outer membrane as a diffusion barrier in salmonezza
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 250, No. 18, Issue of September 25, pp. 7359-7365, 1975
Printed in U. S. A.
Outer Membrane as a Diffusion Barrier in SalmoneZZa typhimurium PENETRATION OF OLIGO- AND POLYSACCHARIDES INTO ISOLATED OUTER MEMBRANE VESICLES AND CELLS WITH DEGRADED PEPTIDOGLYCAN LAYER*
(Received for publication, January 13,1975)
TAIJI NAKAE AND HIROSHI NIKAIDO
From the Department of Bacteriology and Immunology, University of California, Berkeley, California 94 720
In Escherichia coli and Salmonella typhimurium, the cell wall that contains both the outer membrane layer and the peptidoglycan layer acts as a barrier of the molecular sieve type for the penetration of uncharged saccharides (G. Decad, T. Nakae, and H. Nikaido (1974) Fed. Proc. 33, 1240). Here we examined which of the layers of the cell wall limited the size of the penetrating molecules, by studying the penetration of saccharides into (a) cells whose peptidoglycan layer had been destroyed by lysozyme treatment or growth in the presence of penicillin and (b) isolated outer membrane vesicles. We found that peptidoglycan-defective cells were similar to intact, plasmolyzed cells in that they allowed a partial penetration of stachyose (molecular weight 666), but essentially excluded saccharides with molecular weights higher than 900 to 1000. We also found that the isolated outer membrane acted as a penetration barrier for saccharides. These observations led us to conclude that the outer membrane, rather than peptidoglycan, sets the size limit for the penetration of uncharged, hydrophilic molecules through the E. coli or S. typhimurium cell wall. The isolated outer membrane, however, had an exclusion limit much higher than that found in intact cells. This “leakiness” could be decreased either by the use of mutants producing extremely deficient lipopolysaccharide, or by trypsin treatment of the isolated membrane followed by heating and slow cooling in the presence of Mg *+ We feel that these observations . are consistent with the hypothesis that the resealing of the ruptured outer membrane during the isolation procedure is often incomplete, and that cracks and holes thus generated are responsible for the “leakiness” of the isolated membrane vesicles.
Gram-negative bacteria such as Escherichia coli or Salmo- nella typhimurium are covered by a double membrane system (3, 4). The inner, or cytoplasmic membrane is surrounded by the “outer membrane,” and the peptidoglycan layer is located between these two membranes. Since the components of known active transport systems are usually found in the cytoplasmic membrane (5, 6), the outer membrane had been thought as a rather inert structure that allows the penetration of all low molecular weight substances.
Increasing numbers of evidence that have accumulated in recent years suggest, however, that the outer membrane indeed constitutes a barrier for the penetration of certain kinds of molecules. The relevant data include the following: (a) E. coli and Salmonella mutants defective in the biosynthesis of lipoplysaccharide, a component of the outer membrane, were found to be more sensitive to various antibiotics, dyes, and bile salts (7-12), and to allow the more rapid penetration of a dye, crystal violet (13). (b) Short exposure of E. coli or Salmonella
* This study was supported, in part, by United States Public Health Service Grant 5 ROl AI09644-05 and Grant BC-20 from the American Cancer Society. This is the fourth paper of the series on bacterial outer membrane; the preceding papers include Refs. 1 and 2.
to EDTA was found to release about one-half of the lipopoly- saccharide from the outer membrane of the cells and at the same time make the cells sensitive to antibiotics to which they are normally resistant (14-17). (c) Kinetics of P-thiogalactoside exit from E. coli suggested the presence of a partial diffusion barrier outside the cytoplasmic membrane (18). (d) E. coli cannot utilize peptides larger than a certain size limit, a finding which is consistent with the existence of a molecular sieving barrier on its surface (19).
In spite of the presence of these many pieces of indirect evidence, no direct study of outer membrane permeability has been reported. Therefore, we have examined the penetration of oligosaccharides across the cell wall (i.e. outer membrane plus peptidoglycan) of plasmolyzed E. coli and S. typhimurium cells, and found that the cell wall constitutes a significant diffusion barrier for a molecule as small as stachyose (666 daltons) (2). This paper extends these oligo- and polysaccha- ride penetration studies to two new systems: (a) isolated outer membrane vesicles, and (b) cells with degraded peptidoglycan layer. The results suggest that the outer membrane layer, rather than peptidoglycan layer, sets the size limit for the penetration of saccharides through cell wall.
7359
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7360
EXPERIMENTAL PROCEDURE
Bacterial Strains-Salmonella typhimurium LT2 and its mutants were used. The latter, HN202 (galE503) (i.e. strain M-l of Ref. 20) and TA2168 (hisC3076 galE506 r/a-1009) (21) produce incomplete lipo- polysaccharides of Rc and Re types (22), respectively.
Isolation of Outer Membrane Vesicles-Bacteria were grown and the outer membrane vesicles were isolated according to the procedure of Osborn and co-workers (23), except that the spheroplasts were lysed by slowly pouring the spheroplast suspension into 5 to 10 volumes of ice-cold distilled water containing pancreatic deoxyribonuclease (Sigma, type I) (10 pglml) and pancreatic ribonuclease (Sigma, type XIIB) (10 pg/ml). With TA2168 the separation of the outer membrane band from the intermediate band (“M band”) was insufficient; we therefore collected carefully only the leading portion of the former band. Such a preparation contained less than 5% of inner membrane, as determined by DPNH oxidase assay (23).
For permeability experiments, the vesicles were used immediately after their preparation. Less than 2% of the total phospholipids in these preparations correspond to lysophosphatidylethanolamine.
Determination of Outer Membrane Permeability-The standard procedure with the isolated outer membrane was as follows. To an appropriate buffer solution containing membrane vesicles (containing 3 to 5 mg of protein), [“Clglycerol (or [“C]sucrose) (0.1 to 0.15 PCi), and SH-oligosaccharide (or (‘Hlinulin or [SH]dextran) (0.3 to 0.45 PCi) were added. For the purpose of minimizing the possible adsorp- tion of radioactive compounds to vesicles, the buffer usually con- tained nonradioactive sucrose and glycerol at the final concentrations of 0.25 and 0.01 M. In some experiments, nonradioactive oligo- saccharide, inulin, or dextran was also added, but this did not alter the results.
The reaction mixture (final volume, 0.50 ml) in 1.5.ml plastic centrifuge tubes (Eppendorf, No. 3810) was mixed immediately for 5 s with a Vortex-type mixer, and then was kept at room temperature for 10 min without further agitation. The tubes were centrifuged at room temperature in a SS-34 rotor (equipped with rubber adapters, Cata- logue No. 317, Ivan Sorvall) of a Sorvall RC-2B centrifuge for 10 min at 19,000 rpm (25,800 x g at the tip of centrifuge tubes). The supernatant was rpmoved and the inside wall of the centrifuge tubes was carefully wiped with a piece of tissue paper. The pellet was resuspended in 1.00 ml of water by mixing vigorously with a Vortex-type mixer, and after 10 min at room temperature the tubes were centrifuged again as before.
The first supernatant (50 ~1) plus 450 ~1 of water, and the second supernatant (500 ~1) were transferred to separate vials, and the radioactivity was determined as described below.
The space (in microliters) permeable to a given substance in the pellet was calculated according to the following formula:
Permeable space (~1) = total radioactivity in second supernatant
radioactivity/F1 of first supernatant
radioactivity in 500 ~1 of second supernatant = x 100
radioactivity in 50 ~1 of first supernatant
The permeable space, however, was affected by the completeness of the removal of the first supernatant; variations of &5% were seen among similar samples. In order to circumvent this difficulty, we took advantage of the double label technique. Usually our “C-labeled compounds could penetrate freely through the outer membrane (2) (see also “Results”). Thus the permeable space for “C compounds would be a sum of (a) intravesicular space, (b) intervesicular space, and (c) volume of the first supernatant that could not be removed. If the 3H-labeled compound used was completely impermeable through the membrane, the permeable space for the 3H compound will be the sum of b and c above. If we take the difference (permeable space for “C compound) (permeable space for 3H compound), the result would be equal to a, or intravesicular space, and this is not affected by the completeness of removal of the first supernatant. We therefore routinely calculated this difference, “3H-impermeable space,” which is equal to intravesicular space for nonpenetrating compounds, close to zero for fully permeable compounds, and in between for partially penetrating compounds.
For reasons described under “Results” we calculate the intravesicu- lar space by using 16,500 daltons dextran, which we assume to be completely impermeable through the outer membrane. The degree of penetration for any other 3H-labeled compound is:
1 _ $H-impermeable space for that compound 1 x 100 (%) intravesicular space
Plasmolyzed Celkl-HN202 cells were grown in L broth (24) (glucose omitted) (1,500 ml in g-liter Erlenmeyer flask) at 37’ with vigorous shaking (200 rpm, New Brunswick gyrotatory shaker model G-52). When the cell density reached about 1 mg (wet weight)/ml, the cells were harvested by centrifugation at 6,000 x g for 5 min. (This as well as all subsequent operations were carried out at room temperature.) The cells were washed once with 0.1 M NaCl, and suspended in 4 to 6 ml of 0.1 M NaCl. The penetration of saccharides was tested in a manner similar to that described above for the outer membrane vesicles, except that (a) cells (50 to 150 mg wet weight) were used instead of membrane vesicles, (b) centrifugation was at 1,780 x g rather than at 25,800 x g, and (c) NaCl was added to the reaction mixture to a final concentra- tion of 0.5 M in order to produce plasmolysis. [“C]Sucrose was used as the compound that penetrates through the cell wall but not through the inner, cytoplasmic membrane. The “SH-impermeable space” in this case thus was related to the size of, and the degree of penetration of aH-saccharides into the periplasmic space.
Lysozyme-treated, Plnsmolyzed Celki-HN202 was grown and har- vested as described for outer membrane preparation (23). Cells (about 1 g wet weight) were suspended in 100 ml of 0.75 M sucrose-O.01 M Tris-HCl (pH 7.5)-100 pg/ml of lysozyme, and the suspension was kept at 0” for 10 min, collected by centrifugation (2700 x g for 10 min), and finally resuspended in 4 to 6 ml of 0.75 M sucrose-O.01 M Tris-HCl (pH 7.5)-100 rg/ml of lysozyme. Although most cells still retained the rodlike shape, slow dilution with 2 volumes of cold water converted about 70% of the cells into spherical forms, an observation indicating that peptidoglycan was digested in at least 70% of the cells (25). The penetration of saccharides into these cells was tested in a manner similar to that described above for plasmolyzed cells, except that NaCl was not added and that the incubation was done in 0.75 M sucrose-O.01 M Tris-HCl (pH 7.5)-100 fig/ml of lysozyme.
Penicillin Spheroplasts-An exponential phase culture of HN202 in 400 ml of medium M-9 (26) (cell density: 0.3 mg dry weight/ml) was diluted 4-fold with Difco antibiotics medium No. 3 containing 20% sucrose, 0.2% MgSO,, and 0.1% penicillin G, prewarmed to 37”. The B-liter Erlenmeyer flask containing the suspension was gently (40 rpm) shaken at 37’ on a water bath shaker (New Brunswick, model G-77) for 2 hours, and the spheroplasts were collected by centrifugation (2000 x g, 10 min) and were suspended very carefully in about 4 ml of antibiotics medium No. 3 (Difco) containing 20% sucrose, 0.2% MgSO,, and 0.1% penicillin G. Examinations with a phase-contrast microscope showed that more than 99% of cells had been converted into spherical forms. For the determination of penetration of sac- charides, the conditions used were similar to those used for plasmolyzed cells, except that the incubation was done in antibiotics medium No. 3 containing 20% sucrose, 0.2% MgSO,, and 0.1% penicillin G.
Chemicals-Chemicals used were of the best grades commercially available. [“C]Sucrose, [“Clglucose, and [“Clglycerol were obtained from Schwarz/Mann. 13HlRaffinose and 13Hlstachvose were the eift from Mr. G. Decad of this department. . _ ”
Fractionation of [3HjDewtran and [3Hynulin-About 750 PCi of [methoxy-3H]dextran (New England Nuclear Corp., NET-427A, Lot 622-252, specific! activity 1 mCi/3 mg) were applied to a column (2.54 x 51 cm) of Bio-Gel P-30, 100 to 200 mesh (Bio-Rad Labora- tories, Richmond, Calif.), which was eluted with 0.1 M NaCl. The highest peak of radioactivity was found in fractions eluted shortly after the void volume. However, significant amounts of radioac- tivity were found in almost all fractions ranging from the void volume to the complete inclusion volume. Neighboring fractions were pooled and purified by repeating gel filtration usually twice on Bio-Gel P-30, or Bio-Gel P-10, or both. Fractions on either side of “peak” fractions were discarded in each gel filtration step, and consequently the puri- fied [$H]dextran preparations apparently had a narrow molecular weight distribution as judged by the sharp elution pattern in analytical gel filtration. In the example shown in Fig. 1, for instance, a monodisperse inclusion volume marker, CoCl, was eluted in a “peak” with a half-height width of 3.8 ml, and the [*H]dextran “peak” had the half-height width only a little wider, 5.1 ml, thus indicating the presence of very little heterogeneity.
[3H]Inulin (New England Nuclear Corp., NET-314) was similarly fractionated first on a column of Bio-Gel P-4 and then on columns of Bio-Gel P-4 or P-2.
The two [3H]dextran fractions with the highest molecular weight were individually co-chromatographed with Dextran TlO (Lot 3205, M, (number average) 9,740, Pharmacia) on a column (1.27 x 85 cm) of Bio-Gel A-0.5m (200 to 400 mesh, Bio-Rad), which was eluted with 0.1 M NaCl-0.02% NaN,. These two [SH]dextran fractions were eluted at
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from
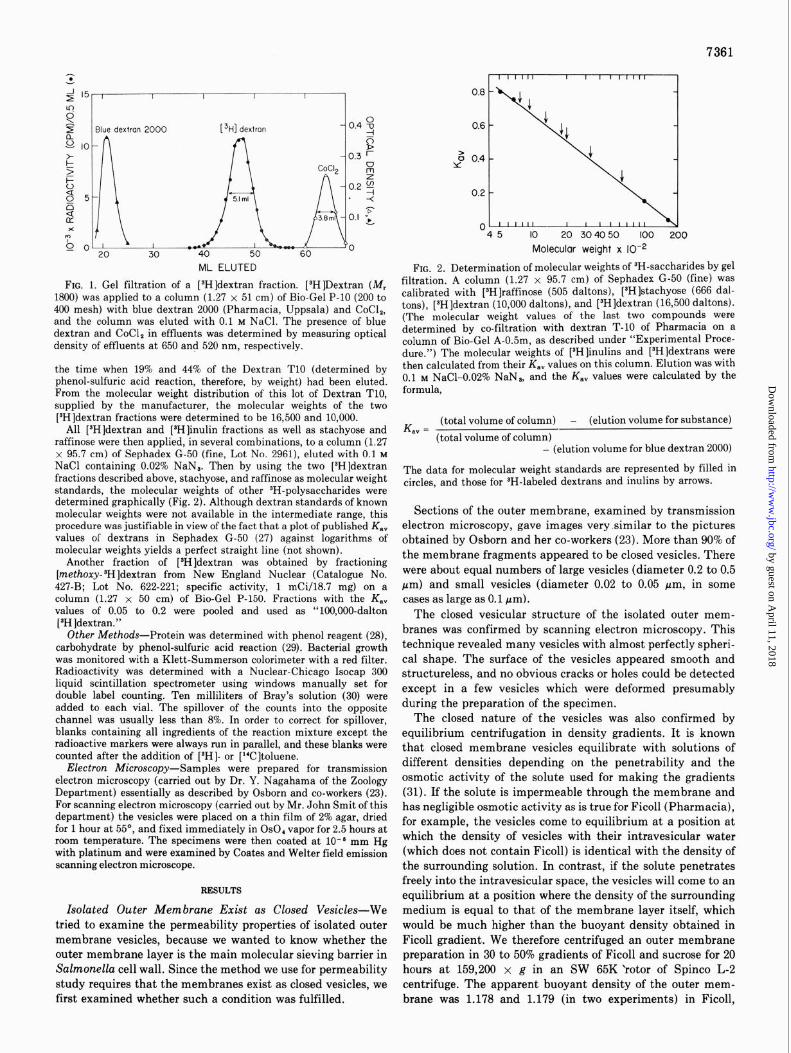
7361
r3tt] dextron
ML ELUTED
FIG. 1. Gel filtration of a [*H]dextran fraction. [sHIDextran (iVf, 1600) was applied to a column (1.27 x 51 cm) of Bio-Gel P-10 (200 to 400 mesh) with blue dextran 2000 (Pharmacia, Uppsala) and CoCl,, and the column was eluted with 0.1 M NaCl. The presence of blue dextran and CoCl, in effluents was determined by measuring optical density of effluents at 650 and 520 nm, respectively.
the time when 19% and 44% of the Dextran TlO (determined by phenol-sulfuric acid reaction, therefore, by weight) had been eluted. From the molecular weight distribution of this lot of Dextran TlO, supplied by the manufacturer, the molecular weights of the two [JH]dextran fractions were determined to be 16,500 and 10,000.
All [3H]dextran and [$H]inulin fractions as well as stachyose and raffinose were then applied, in several combinations, to a column (1.27 x 95.7 cm) of Sephadex G-50 (fine, Lot No. 2961), eluted with 0.1 M NaCl containing 0.02% NaN,. Then by using the two [BH]dextran fractions described above, stachyose, and raffinose as molecular weight standards, the molecular weights of other ‘H-polysaccharides were determined graphically (Fig. 2). Although dextran standards of known molecular weights were not available in the intermediate range, this procedure was justifiable in view of the fact that a plot of published K,, values of dextrans in Sephadex G-50 (27) against logarithms of molecular weights yields a perfect straight line (not shown).
Another fraction of [Wldextran was obtained by fractioning [methoxy-SH]dextran from New England Nuclear (Catalogue No. 427-B; Lot No. 622-221; specific activity, 1 mCi/18.7 mg) on a column (1.27 x 50 cm) of Bio-Gel P-150. Fractions with the K., values of 0.05 to 0.2 were pooled and used as “100,000-dalton [SH]dextran.”
Other Methods-Protein was determined with phenol reagent (28), carbohydrate by phenol-sulfuric acid reaction (29). Bacterial growth was monitored with a Klett-Summerson calorimeter with a red filter. Radioactivity was determined with a Nuclear-Chicago Isocap 300 liquid scintillation spectrometer using windows manually set for double label counting. Ten milliliters of Bray’s solution (30) were added to each vial. The spillover of the counts into the opposite channel was usually less than 8%. In order to correct for spillover, blanks containing all ingredients of the reaction mixture except the radioactive markers were always run in parallel, and these blanks were counted after the addition of [*HI- or [“Cltoluene.
Electron Microscopy-Samples were prepared for transmission electron microscopy (carried out by Dr. Y. Nagahama of the Zoology Department) essentially as described by Osborn and co-workers (23). For scanning electron microscopy (carried out by Mr. John Smit of this department) the vesicles were placed on a thin film of 2% agar, dried for 1 hour at 55’, and fixed immediately in 0~0, vapor for 2.5 hours at room temperature. The specimens were then coated at 1O-s mm Hg with platinum and were examined by Coates and Welter field emission scanning electron microscope.
RESULTS
Isolated Outer Membrane Exist as Closed Vesicles-We tried to examine the permeability properties of isolated outer membrane vesicles, because we wanted to know whether the outer membrane layer is the main molecular sieving barrier in Salmonella cell wall. Since the method we use for permeability study requires that the membranes exist as closed vesicles, we first examined whether such a condition was fulfilled.
Y
45 IO 20 304050 100 200
Molecular weight x IO-2
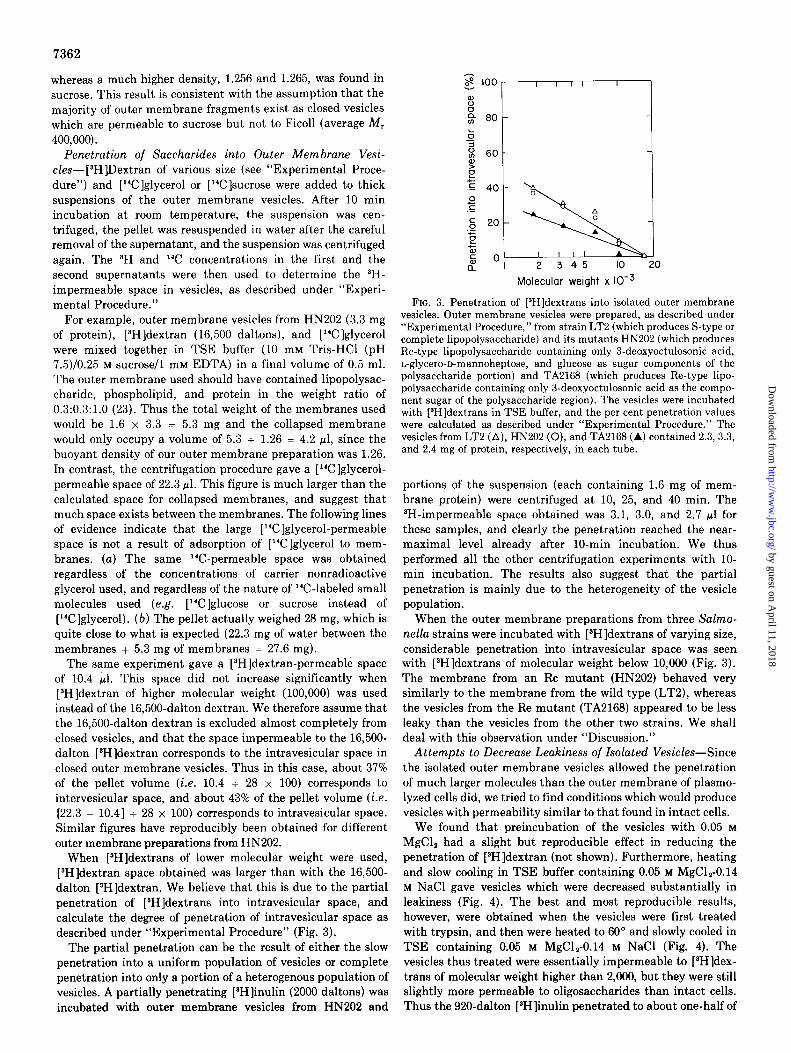
FIG. 2. Determination of molecular weights of 3H-saccharides by gel filtration. A column (1.27 x 95.7 cm) of Sephadex G-50 (fine) was calibrated with [aH]raffinose (505 daltons), [SH]stachyose (666 dal- tons), taH]dextran (10,000 daltons), and tBH]dextran (16,500 daltons). (The molecular weight values of the last two compounds were determined by co-filtration with dextran T-10 of Pharmacia on a column of Bio-Gel A-0.5m, as described under “Experimental Proce- dure.“) The molecular weights of [*H]inulins and [$H]dextrans were then calculated from their K., values on this column. Elution was with 0.1 M NaCl-0.02% NaN,, and the K,, values were calculated by the formula,
K = (totalvolumeof column) - (elution volume for substance)
‘” (total volume of column) - (elution volume for blue dextran 2000)
The data for molecular weight standards are represented by filled in circles, and those for *H-labeled dextrans and inulins by arrows.
Sections of the outer membrane, examined by transmission electron microscopy, gave images very similar to the pictures obtained by Osborn and her co-workers (23). More than 90% of the membrane fragments appeared to be closed vesicles. There were about equal numbers of large vesicles (diameter 0.2 to 0.5 pm) and small vesicles (diameter 0.02 to 0.05 pm, in some cases as large as 0.1 pm).
The closed vesicular structure of the isolated outer mem- branes was confirmed by scanning electron microscopy. This technique revealed many vesicles with almost perfectly spheri- cal shape. The surface of the vesicles appeared smooth and structureless, and no obvious cracks or holes could be detected except in a few vesicles which were deformed presumably during the preparation ,of the specimen.
The closed nature of the vesicles was also confirmed by equilibrium centrifugation in density gradients. It is known that closed membrane vesicles equilibrate with solutions of different densities depending on the penetrability and the osmotic activity of the solute used for making the gradients (31). If the solute is impermeable through the membrane and has negligible osmotic activity as is true for Ficoll (Pharmacia), for example, the vesicles come to equilibrium at a position at which the density of vesicles with their intravesicular water (which does not contain Ficoll) is identical with the density of the surrounding solution. In contrast, if the solute penetrates freely into the intravesicular space, the vesicles will come to an equilibrium at a position where the density of the surrounding medium is equal to that of the membrane layer itself, which would be much higher than the buoyant density obtained in Ficoll gradient. We therefore centrifuged an outer membrane preparation in 30 to 50% gradients of Ficoll and sucrose for 20 hours at 159,200 x g in an SW 65K ‘rotor of Spinco L-2 centrifuge. The apparent buoyant density of the outer mem- brane was 1.178 and 1.179 (in two experiments) in Ficoll,
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from
7362
whereas a much higher density, 1.256 and 1.265, was found in sucrose. This result is consistent with the assumption that the majority of outer membrane fragments exist as closed vesicles which are permeable to sucrose but not to Ficoll (average M, 400,000).
Penetration of Saccharides into Outer Membrane Vesi- cles- [SH]Dextran of various size (see “Experimental Proce- dure”) and [“Clglycerol or [“C]sucrose were added to thick suspensions of the outer membrane vesicles. After 10 min incubation at room temperature, the suspension was cen- trifuged, the pellet was resuspended in water after the careful removal of the supernatant, and the suspension was centrifuged again. The ‘H and “C concentrations in the first and the second supernatants were then used to determine the $H- impermeable space in vesicles, as described under “Experi- mental Procedure.”
For example, outer membrane vesicles from HN202 (3.3 mg of protein), [‘Hldextran (16,500 daltons), and [‘Clglycerol were mixed together. in TSE buffer (10 mM Tris-HCl (pH 7.5)/0.25 M sucrose/l mM EDTA) in a final volume of 0.5 ml. The outer membrane used should have contained lipopolysac- charide, phospholipid, and protein in the weight ratio of 0.3:0.3:1.0 (23). Thus the total weight of the membranes used would be 1.6 x 3.3 = 5.3 mg and the collapsed membrane would only occupy a volume of 5.3 + 1.26 = 4.2 ~1, since the buoyant density of our outer membrane preparation was 1.26. In contrast, the centrifugation procedure gave a [“Clglycerol- permeable space of 22.3 ~1. This figure is much larger than the calculated space for collapsed membranes, and suggest that much space exists between the membranes. The following lines of evidence indicate that the large [“Clglycerol-permeable space is not a result of adsorption of [“C]glycerol to mem- branes. (a) The same “C-permeable space was obtained regardless of the concentrations of carrier nonradioactive glycerol used, and regardless of the nature of “C-labeled small molecules used (e.g. [“Clglucose or sucrose instead of [“Clglycerol). (b) The pellet actually weighed 28 mg, which is quite close to what is expected (22.3 mg of water between the membranes + 5.3 mg of membranes = 27.6 mg).
The same experiment gave a [3H]dextran-permeable space of 10.4 ~1. This space did not increase significantly when [3H]dextran of higher molecular weight (100,000) was used instead of the 16,500-dalton dextran. We therefore assume that the 16,500-dalton dextran is excluded almost completely from closed vesicles, and that the space impermeable to the 16,500- dalton [SH]dextran corresponds to the intravesicular space in closed outer membrane vesicles. Thus in this case, about 37% of the pellet volume (i.e. 10.4 + 28 x 100) corresponds to intervesicular space, and about 43% of the pellet volume (i.e. [22.3 - 10.41 + 28 x 100) corresponds to intravesicular space. Similar figures have reproducibly been obtained for different outer membrane preparations from HN202.
When ]sH]dextrans of lower molecular weight were used, [SH]dextran space obtained was larger than with the 16,500- dalton [SH]dextran. We believe that this is due to the partial penetration of [SH]dextrans into intravesicular space, and calculate the degree of penetration of intravesicular space as described under “Experimental Procedure” (Fig. 3).
The partial penetration can be the result of either the slow penetration into a uniform population of vesicles or complete penetration into only a portion of a heterogenous population of vesicles. A partially penetrating [9H]inulin (2000 daltons) was incubated with outer membrane vesicles from HN202 and
Molecular weight x 10e3
FIG. 3. Penetration of [3H]dextrans into isolated outer membrane vesicles. Outer membrane vesicles were prepared, as described under “Experimental Procedure,” from strain LT2 (which produces S-type or complete lipopolysaccharide) and its mutants HN202 (which produces Rc-type lipopolysaccharide containing only 3-deoxyoctulosonic acid, L-glycero-n-mannoheptose, and glucose as sugar components of the polysaccharide portion) and TA2168 (which produces Retype lipo- polysaccharide containing only 3-deoxyoctulosonic acid as the compo- nent sugar of the polysaccharide region). The vesicles were incubated with [SH]dextrans in TSE buffer, and the per cent penetration values were calculated as described under “Experimental Procedure.” The vesicles from LT2 (A), HN202 (O), and TA2168 (A) contained 2.3,3.3, and 2.4 mg of protein, respectively, in each tube.
portions of the suspension (each containing 1.6 mg of mem- brane protein) were centrifuged at 10, 25, and 40 min. The ‘H-impermeable space obtained was 3.1, 3.0, and 2.7 ~1 for these samples, and clearly the penetration reached the near- maximal level already after IO-min incubation. We thus performed all the other centrifugation experiments with lo- min incubation. The results also suggest that the partial penetration is mainly due to the heterogeneity of the vesicle population.
When the outer membrane preparations from three Salmo- nella strains were incubated with [SH]dextrans of varying size, considerable penetration into intravesicular space was seen with [3H]dextrans of molecular weight below 10,000 (Fig. 3). The membrane from an Rc mutant (HN202) behaved very similarly to the membrane from the wild type (LT2), whereas the vesicles from the Re mutant (TA2168) appeared to be less leaky than the vesicles from the other two strains. We shall deal with this observation under “Discussion.”
Attempts to Decrease Leakiness of Isolated Vesicles-Since the isolated outer membrane vesicles allowed the penetration of much larger molecules than the outer membrane of plasmo- lyzed cells did, we tried to find conditions which would produce vesicles with permeability similar to that found in intact cells.
We found that preincubation of the vesicles with 0.05 M
MgCl, had a slight but reproducible effect in reducing the penetration of [sH]dextran (not shown). Furthermore, heating and slow cooling in TSE buffer containing 0.05 M MgCl,-0.14 M NaCl gave vesicles which were decreased substantially in leakiness (Fig. 4). The best and most reproducible results, however, were obtained when the vesicles were first treated with trypsin, and then were heated to 60” and slowly cooled in TSE containing 0.05 M MgCl,-0.14 M NaCl (Fig. 4). The vesicles thus treated were essentially impermeable to [“Hldex- trans of molecular weight higher than 2,000, but they were still slightly more permeable to oligosaccharides than intact cells. Thus the 920-dalton [*H]inulin penetrated to about one-half of
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7363
Molecular weight x IO-’
water bath. Such “trypsinized, heated in MgCl,” vesicles (triangles) were tested for permeability in TSE buffer-50 mM MgCl,-140 mM
FIG. 4. Penetration of oligo- and polysaccharides into outer mem- brane vesicles and plasmolyzed cells of HN202. Outer membrane preparations (circles, crosses, and triangles) and cells plasmolyzed in
NaCl. Three experiments with different membrane preparations were
0.5 M NaCl (squares) were tested for penetration as described under “Experimental Procedure.”
oerformed with both untreated and treated vesicles. Svmbols not filled
3H-Labeled compounds used and their molecular weights (in parentheses) were: raffinose (504), stachyose (666), inulin (920), inulin (1,380), dextran (1,800), inulin (2,000), dextran (3,300), dextran (6,300), dextran (lO,OOO), and dextran (16,500). Untreated outer membranes (circles) were tested in TSE buffer. Portions of the outer membrane preparation (8 to 10 mg protein/ml) were also treated with trypsin (250 &ml) for 1 hour at 37” in TSE buffer containing 0.1 mg/ml of chloramphenicol, MgCl, and NaCl were then added to the final concentrations of 50 and 140 mM, respectively, and the suspension was heated at 60” for 30 min followed bv the slow cooling (4 to 5 hours) to room temnerature in a turned off
plasmolyzed cells were similar to that of the plasmolyzed cells in that 50% penetration was observed with the saccharides of about 700 daltons (Fig. 5). Thus the molecular sieving proper- ties of cell wall were not affected significantly by the absence of continuous peptidoglycan layers. 1
Very similar results’ were also obtained with penicillin spheroplasts (Fig. 5), which are known to contain extensively damaged peptidoglycan layer (32-34). (Penicillin spheroplasts and lysozyme-treated cells, however, allowed the penetration of small amounts of saccharides of 1000 to 2000 daltons. This finding will be discussed under “Discussion.“) These results indicate that the main diffusion barrier in the cell envelope lies in the outer membrane, not in the peptidoglycan layer.’
In one experiment, we sought to create a situation which was complementary to those described above; we tried to damage the outer membrane while leaving the peptidoglycan layer intact. HN202 cells were washed in a solution containing 0.05 M
EDTA and were plasmolyzed in 0.5 M NaCl containing about 0.04 M EDTA. EDTA is known to take off a significant portion (under our conditions 15%) of lipopolysaccharide molecules from the outer membrane layer (14). The cell wall of these
cells, however, was still essentially impermeable to SH-saccha- rides of molecular weight higher than 1800. Thus the removal of much lipopolysaccharide did not increase the permeability
of cell wall to polysaccharides.
DISCUSSION
in (0, A), half-filled in (0, A), and completely filled in (0, A) correspond to these three experiments. In other experiments, the vesicles were heated and slowly cooled in TSE buffer-50 mM MgCl,-140 mM NaCl without trypsin digestion; data from a typical experiment are shown (crosses). As a control, exponential phase cells of HN202 were plasmolyzed in 0.5 M NaCl and the penetration of 3H-oligosaccharides was determined (squares) as described under “Experimental Proce- dure.”
barrier for the penetration of oligosaccharides (2). In that study, however, the precise location of the barrier has not been established.
By using plasmolyzed cells of Escherichiu coli and Salmo- nella typhimurium, Decad and we have found previously that the cell wall of these organisms acted as a molecular sieve-type
the intravesicular space in the treated vesicles, whereas with plasmolyzed cells similar half-penetration was obtained with [3H]stachyose (666 daltons) (2). It may be argued that trypsin might simply be converting many vesicles into lipopolysac- charide-phospholipid “liposomes” that do not contain pro- teins, thereby decreasing the permeability in general. How- ever, such a situation is unlikely because only 30% of proteins were released from vesicles as a result of trypsin treatment, and because all vesicles were still permeable to sucrose after the treatment, although liposomes are known to be impermeable to sucrose (1).
Procedures which did not significantly decrease the leakiness of vesicles include the following: (a) trypsin treatment alone,
(b) trypsin treatment followed by heating and slow cooling in TSE buffer, and (c) heating and slow cooling in TSE buffer.
Permeability of Cells with Degraded Peptidoglycan Layers -Another system we used for investigating the role of peptido- glycan and outer membrane in saccharide penetration was the cells with damaged peptidoglycan layers.
In a series of experiments we treated plasmolyzed HN202 cells with a high concentration of lysozyme (100 pg/ml). Under these conditions lysozyme can digest the peptidoglycan layer of intact cells (25); the fact that the digestion did occur in our
experiment was ascertained by the occurrence of dilution- induced conversion into spherical forms (see “Experimental Procedure”). The permeability of these lysozyme-treated,
Thus the barrier properties could be due to the presence of (a) peptidoglycan layer, (b) outer membrane, or (c) both of these layers. We have found in this study that the isolated outer membrane, which are largely devoid of peptidoglycan, did act as a diffusion barrier. This observation clearly rules out the Hypothesis a above, that peptidogylcan is entirely responsible for the barrier properties of E. coli and Salmonella cell wall.
When we consider Hypotheses b and c above, we should keep in mind the fact that the peptidoglycan-rich cell wall of Gram-positive bacteria was shown to act as molecular sieve- type barriers (31). It therefore seems likely that Hypothesis c is correct, but the real question is which of the two layers determines the size limit for diffusing molecules. Since the exclusion limit for E. coli or Salmonella cell wall (about 900 daltons or less for oligosaccharides) is much smaller than that found for the cell wall of Gram-positive bacteria (e.g. about 100,000 daltons for Bacillus megaterium (35)*, it seems rather
unlikely that the peptidoglycan acts as a finer sieve in E. coli or Salmonella. This possibility appears even less likely if we consider the fact that the peptidoglycan layer in E. coli or Salmonella is much thinner and less extensively cross-linked
‘Although sucrose, rather than NaCl, was used as the plasmolyzing agent in these experiments, our unpublished oligosaccharide penetra- tion experiments have shown that sucrose-plasmolyzed cells behave in a manner very similar to NaCl-plasmolyzed cells.
‘Although it has been claimed that the “monodisperse exclusion threshold” for this system is 1200 daltons, the data presented are inconsistent internally, since 25 to 65% penetration has been obtained with “Dextran T-40 ” “T-20,” and “T-10” from Pharmacia, which contain few molecules smaller than 1200 daltons (less than 0.1 to 0.5% in weight).
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from
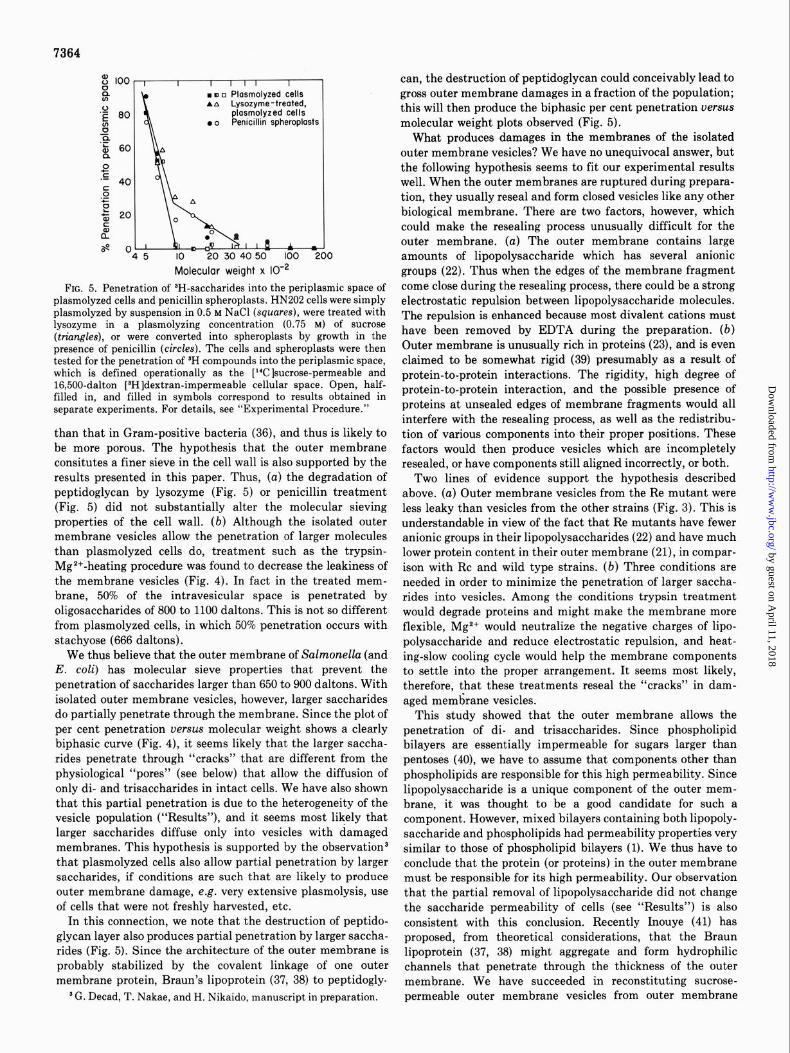
7364
n q q Plasmalyzed cells ra Lvsozvme-treated.
pjoskolyzed cells l o Penicillin spheroplasts
Molecular weight x 10m2
FIG. 5. Penetration of ‘H-saccharides into the periplasmic space of nlasmolvzed cells and nenicillin SDheronlasts. HN202 cells were simply plasmolyxed by suspension in 0.5~ N&l (squares), were treated &h lysozyme in a plasmolyzing concentration (0.75 M) of sucrose (triangles), or were converted into spheroplasts by growth in the presence of penicillin (circles). The cells and spheroplasts were then tested for the penetration of W compounds into the periplasmic space, which is defined operationally as the [Wlsucrose-permeable and 16,500-dalton [JH]dextran-impermeable cellular space. Open, half- filled in, and filled in symbols correspond to results obtained in separate experiments. For details, see “Experimental Procedure.”
than that in Gram-positive bacteria (36), and thus is likely to be more porous. The hypothesis that the outer membrane consitutes a finer sieve in the cell wall is also supported by the results presented in this paper. Thus, (a) the degradation of peptidoglycan by lysozyme (Fig. 5) or penicillin treatment (Fig. 5) did not substantially alter the molecular sieving properties of the cell wall. (b) Although the isolated outer membrane vesicles allow the penetration of larger molecules than plasmolyzed cells do, treatment such as the trypsin- Mg2+-heating procedure was found to decrease the leakiness of the membrane vesicles (Fig. 4). In fact in the treated mem- brane, 50% of the intravesicular space is penetrated by oligosaccharides of 800 to 1100 daltons. This is not so different from plasmolyzed cells, in which 50% penetration occurs with stachyose (666 daltons) .
We thus believe that the outer membrane of Salmonella (and E. coli) has molecular sieve properties that prevent the penetration of saccharides larger than 650 to 900 daltons. With isolated outer membrane vesicles, however, larger saccharides do partially penetrate through the membrane. Since the plot of per cent penetration uerws molecular weight shows a clearly biphasic curve (Fig. 4), it seems likely that the larger saccha- rides penetrate through “cracks” that are different from the physiological “pores” (see below) that allow the diffusion of only di- and trisaccharides in intact cells. We have also shown that this partial penetration is due to the heterogeneity of the vesicle population (“Results”), and it seems most likely that larger saccharides diffuse only into vesicles with damaged membranes. This hypothesis is supported by the observationS that plasmolyzed cells also allow partial penetration by larger saccharides, if conditions are such that are likely to produce outer membrane damage, e.g. very extensive plasmolysis, use of cells that were not freshly harvested, etc.
In this connection, we note that the destruction of peptido- glycan layer also produces partial penetration by larger saccha- rides (Fig. 5). Since the architecture of the outer membrane is probably stabilized by the covalent linkage of one outer membrane protein, Braun’s lipoprotein (37, 38) to peptidogly-
can, the destruction of peptidoglycan could conceivably lead to gross outer membrane damages in a fraction of the population; this will then produce the biphasic per cent penetration versus molecular weight plots observed (Fig. 5).
What produces damages in the membranes of the isolated outer membrane vesicles? We have no unequivocal answer, but the following hypothesis seems to fit our experimental results well. When the outer membranes are ruptured during prepara- tion, they usually reseal and form closed vesicles like any other biological membrane. There are two factors, however, which could make the resealing process unusually difficult for the outer membrane. (a) The outer membrane contains large amounts of lipopolysaccharide which has several anionic groups (22). Thus when the edges of the membrane fragment come close during the resealing process, there could be a strong electrostatic repulsion between lipopolysaccharide molecules. The repulsion is enhanced because most divalent cations must have been removed by EDTA during the preparation. (b) Outer membrane is unusually rich in proteins (23), and is even claimed to be somewhat rigid (39) presumably as a result of protein-to-protein interactions. The rigidity, high degree of protein-to-protein interaction, and the possible presence of proteins at unsealed edges of membrane fragments would all interfere with the resealing process, as well as the redistribu- tion of various components into their proper positions. These factors would then produce vesicles which are incompletely resealed, or have components still aligned incorrectly, or both.
Two lines of evidence support the hypothesis described above. (a) Outer membrane vesicles from the Re mutant were less leaky than vesicles from the other strains (Fig. 3). This is understandable in view of the fact that Re mutants have fewer anionic groups in their lipopolysaccharides (22) and have much lower protein content in their outer membrane (21), in compar- ison with Rc and wild type strains. (b) Three conditions are needed in order to minimize the penetration of larger saccha- rides into vesicles. Among the conditions trypsin treatment would degrade proteins and might make the membrane more flexible, MgZ+ would neutralize the negative charges of lipo- polysaccharide and reduce electrostatic repulsion, and heat- ing-slow cooling cycle would help the membrane components to settle into the proper arrangement. It seems most likely, therefore, that these treatments reseal the “cracks” in dam- aged membrane vesicles.
This study showed that the outer membrane allows the penetration of di- and trisaccharides. Since phospholipid bilayers are essentially impermeable for sugars larger than pentoses (40), we have to assume that components other than phospholipids are responsible for this high permeability. Since lipopolysaccharide is a unique component of the outer mem- brane, it was thought to be a good candidate for such a component. However, mixed bilayers containing both lipopoly- saccharide and phospholipids had permeability properties very similar to those of phospholipid bilayers (1). We thus have to conclude that the protein (or proteins) in the outer membrane must be responsible for its high permeability. Our observation that the partial removal of lipopolysaccharide did not change the saccharide permeability of cells (see “Results”) is also consistent with this conclusion. Recently Inouye (41) has proposed, from theoretical considerations, that the Braun lipoprotein (37, 38) might aggregate and form hydrophilic channels that penetrate through the thickness of the outer membrane. We have succeeded in reconstituting sucrose-
‘G. Decad, T. Nakae, and H. Nikaido, manuscript in preparation. permeable outer membrane vesicles from outer membrane
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

proteins, lipopolysaccharides, and phospholipids (42); thus it may become possible to identify the permeability-conferring protein with this technique.
The data presented here do not distinguish between the penetration through pores and the diffusion mediated by carrier proteins. However, evidence to be presented elsewhere8 appears to support the pore mechanism.
Finally, we must emphasize that the hydrophilic pores are not the only pathway through which small molecules move across the outer membrane. Firstly, there are hydrophilic molecules that are too large to go through the pore, yet must diffuse through the outer membrane to support the growth of cells. It is most interesting that various phage and colicin “receptor proteins” on the surface of E. coli and S. typhimu- rium were found to bind these large molecules specifically. Examples include the binding of vitamin B,, (1357 daltons) by colicin E-receptor (43), that of enterochelin (746 daltons) by colicin B-, I-, V-receptor (44), and that of ferrichrome (740 daltons) by T5-, @O-, colicin M-receptor (45). In these cases the binding by specific proteins (carriers?) is obviously needed for the transmembranous diffusion of these large molecules. Sec- ondly, the penetration of oligosaccharides is essentially unaf- fected by mutations that affect the biosynthesis of lipopolysac- charides (2).5 Yet we know that the alteration of lipopoly- saccharide structure drastically alters the permeability of the outer membrane to certain dyes and antibiotics (7-13), which therefore must diffuse through the outer membrane by a different mechanism. We believe that these substances move across the membrane by first dissolving into the hydrophobic interior of the membrane; the properties of this “hydrophobic pathway” will be discussed in a separate paper.’
1. 2. 3.
4.
5. 6. 7.
a.
9.
REFERENCES
Nikaido, H., and Nakae, T. (1973) J. Infect. Dis. 128, 522-526 Decad, G., Nakae, T., and Nikaido, H. (1974) Fed. Proc. 33,124O Glauert, A. M., and Thornley, M. J. (1969) Annu. Reu. Microbial.
23, 159-198 Freer, J. H., and Salton, M. R. J. (1971) in Microbial Toxins
(Weinbalm, G., Kadis, S., and Ajl, S. J., eds) Vol. IV, pp. 67-126, Academic Press, New York
Sistrom, W. R. (1958) Biochim. Biophys. Acta 29,579-587 Kaback, H. R. (1970) Annu. Reu. Biochem. 39, 561-598 Roantree, R. J., Kuo, T., MacPhee, D. G., and Stocker, B. A. D.
(1969) Clin. Res. 17, 157 Schlecht, S., and Schmidt, G. (1969) Zentrabl. Bakteriol. Parasi-
tenkd. Znfektionskr. Hyg. Abt. ZOrig. 212,505-511 Schmidt, G., Schlecht, S., and Westphal, 0. (1969) Zentrabl.
Bakteriol. Parasitenkd. Znfektionskr. Hyg. Abt. Z Orig. 212, 88-96
10.
11.
12.
13.
14. 15. 16.
17.
18.
19. 20. 21.
22.
23.
24. 25.
26. 27.
28
29
7365
Schlecht, S., and Westphal, 0. (1970) Zentrabl. Bakteriol. Parasi- tenkd. Znfektionskr. Hyg. Abt. Z Orig. 213,356-381
Wilkinson, R. G., Gemski, P., and Stocker, B. A. D. (1972) J. Gen. Microbial. 70, 527-554
Tamaki, S., Sato, T., and Matsuhashi, M. (1971). J. Bacterial. 105, 968-975
Gustafsson, P., Nordstriim, K., and Normark, S. (1973) J. Bacte- riol. 116, 893-900
Leive, L. (1965) Biochem. Biophys. Res. Commun. 21, 290-296 Leive, L. (1968) J. Biol. Chem. 243, 2373-2380 Hamilton-Miller, J. M. T. (1965) Biochem. Biophys. Res. Com-
mun. 20, 688-691 Muschel, L. H., and Gustafson, L. (1968) J. Bacterial. 95,
2010-2013 Robbie, J. P., and Wilson, T. H. (1969) Biochim. Biophys. Acta
173, 234-244 Payne, J. W., and Gilvarg, C. (1968) J. Biol. Chem. 243,6291-6299 Fukasawa, T., and Nikaido, H. (1961) Genetics 46, 1295-1303 Ames, G. F.-L., Spudich, E. N., and Nikaido, H. (1974) J. Bacterial.
117, 406-416 Liideritz, O., Galanos, C., Risse, H., Ruschmann, E., Schlecht, S.,
Schmidt, G., Schulte-Holthausen, H., Wheat, R., Westphal, O., and Schlosshardt, J. (1966) Ann. N. Y. Acad. Sci. 133,349-374
Osborn, M. J., Gander, J. E., Parisi, E., and Carson, J. (1972) J. Biol. Chem. 247, 3962-3972
Bertani, G. (1951) J. Bacterial. 62, 293-300 Birdsell, D. C., and Cota-Robles, E. H. (1967) J. Bacterial. 93,
427-437 Anderson, E. H. (1946) Proc. Natl. Acad. Sci. U. S. A. 32,120-128 Laurent, T. C., and Killander, J. (1964) J. Chromatogr. 14,
317-330 Lowry, 0. H., Rosebrough, N. J., Farr, A. L., and Randall, R. J.
(1951) J. Biol. Chem. 193, 265-275 Dubois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., and
Smith, F. (1956) Anal. Chem. 28, 350-356 30. Bray, G. A. (1960) Anal. Biochem. 1, 279-285 31. Wallach, D. F. H., and Kamat, V. B. (1964) Proc. Natl. Acad. Sci.
U. S. A. 52, 721-728
32. Murray, R. G. E., Steed, P., and Elson, H. E. (1965) Can. J. Microbial. 11, 547-560
33. Martin, H. H. (1964) J. Gen. Microbial. 36, 441-450 34. Wise, E. M., Jr., and Park, J. T. (1965) Proc. Natl. Acad. Sci. U. S.
A. 54, 75-81 35. Scherrer, R., and Gerhardt, P. (1971) J. Bacterial. 107, 718-735 36. Schleifer, K. H., and Kandler, 0. (1972) Bacterial. Reu. 36,
407-477 37. Braun, V., and Sieglin, J. (1970) Eur. J. Biochem. 13,33&-346 38. Braun, V., and Wolff, H. (1970) Eur. J. Biochem. 14,387-391 39. Henning, U., Rehn, K., and Hoehn, B. (1973) Proc. Natl. Acad.
Sci. U. S. A. 70,2033-2036 40. Bangham, A. D., De Gier, J., and Greville, G. D. (1967) Chem.
Phys. Lipids 1,225-246 41. Inouye, M. (1974) Proc. Natl. Acad. Sci. U. S. A. 71,239&2400 42. Nakae, T. (1975) Biochem. Biophys. Res. Commun. 64,1224-1230 43. Di Masi, D. R., White, J. C., Schnaitman, C. A., and Bradbeer. C.
(1973) J. Bacterial. 115,506-513 44. Guterman, S. K. (1973) J. Bacterial. 114,1217-1224
’ H. Nikaido, manuscript in preparation. 45. Wayne, R., and Neilands, J. B. (1975) J. Bacterial. 121,497-503
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from

T Nakae and H Nikaidodegraded peptidoglycan layer.
oligo- and polysaccharides into isolated outer membrane vesicles and cells with Outer membrane as a diffusion barrier in Salmonella typhimurium. Penetration of
1975, 250:7359-7365.J. Biol. Chem.
http://www.jbc.org/content/250/18/7359Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/250/18/7359.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 11, 2018
http://ww
w.jbc.org/
Dow
nloaded from