nitric oxide negatively regulates abscisic acid … oxide negatively regulates abscisic acid...
TRANSCRIPT

Nitric oxide negatively regulates abscisic acid signalingin guard cells by S-nitrosylation of OST1Pengcheng Wanga,1, Yanyan Dua,b,1, Yueh-Ju Houa, Yang Zhaoa,c, Chuan-Chih Hsud, Feijuan Yuanb, Xiaohong Zhua,W. Andy Taod, Chun-Peng Songb, and Jian-Kang Zhua,c,2
aDepartment of Horticulture and Landscape Architecture and dDepartment of Biochemistry, Purdue University, West Lafayette, IN 47907; bDepartment ofBiology, Institute of Plant Stress Biology, State Key Laboratory of Cotton Biology, Henan University, Kaifeng 475001, China; and cShanghai Center for PlantStress Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai 200032, China
Contributed by Jian-Kang Zhu, December 8, 2014 (sent for review October 23, 2014)
The phytohormone abscisic acid (ABA) plays important roles inplant development and adaptation to environmental stress. ABAinduces the production of nitric oxide (NO) in guard cells, but howNO regulates ABA signaling is not understood. Here, we show thatNO negatively regulates ABA signaling in guard cells by inhibitingopen stomata 1 (OST1)/sucrose nonfermenting 1 (SNF1)-relatedprotein kinase 2.6 (SnRK2.6) through S-nitrosylation. We foundthat SnRK2.6 is S-nitrosylated at cysteine 137, a residue adjacent tothe kinase catalytic site. Dysfunction in the S-nitrosoglutathione(GSNO) reductase (GSNOR) gene in the gsnor1-3mutant causes NOoveraccumulation in guard cells, constitutive S-nitrosylation ofSnRK2.6, and impairment of ABA-induced stomatal closure. Intro-duction of the Cys137 to Ser mutated SnRK2.6 into the gsnor1-3/ost1-3 double-mutant partially suppressed the effect of gsnor1-3on ABA-induced stomatal closure. A cysteine residue correspond-ing to Cys137 of SnRK2.6 is present in several yeast and humanprotein kinases and can be S-nitrosylated, suggesting that theS-nitrosylation may be an evolutionarily conserved mechanismfor protein kinase regulation.
NO | ABA | drought | GSNOR | stomata
Abscisic acid (ABA) plays critical roles in seed dormancy andgermination, plant growth, and adaptation to environmental
challenges (1, 2). Stresses, such as drought and high salt con-ditions, increase ABA concentration in plants as a result of ABAbiosynthesis or ABA release from its inactive, conjugated forms(3). In the presence of ABA, the ABA receptors in the PYR1(Pyrabactin Resistance 1)/PYL (PYR1-Like)/RCAR (Regula-tory Component of ABA receptor) protein family bind to andinhibit the activity of clade A protein phosphatase 2Cs (PP2Cs),which are considered as coreceptors and negative regulators ofABA signaling (4–6). This process then results in the releaseof sucrose nonfermenting 1 (SNF1)-related protein kinase 2s(SnRK2s) from suppression by the PP2Cs. As central componentsof the ABA signaling pathway, the activated SnRK2s phosphory-late dozens of downstream effectors to regulate various physio-logical processes, including stomatal closure, root growth anddevelopment, seed dormancy, seed germination, and flowering (7).As the gateway for photosynthetic CO2 uptake and transpi-
rational water loss, stomata are critical for plant growth andphysiology (8). ABA regulates stomatal movement and muta-tions in ABA biosynthesis genes (9), or in the PYL or SnRK2.6(also known as OST1) genes cause open-stomata phenotypes(10). On the other hand, dysfunction of the PP2Cs or over-expression of RCAR1/PYL9 causes stomatal closure (5). Amongthe three SnRK2s, SnRK2.2, -2.3, and -2.6, which are most im-portant for ABA signaling, SnRK2.6 is preferentially expressedin guard cells and plays a critical role in stomatal regulation,whereas SnRK2.2 and -2.3 are mainly expressed in seeds andyoung seedlings and are thus more important for seed germi-nation and seedling growth (4, 11). SnRK2.6 phosphorylates theslow (S-type) anion channel associated 1 and inward potassium
channel KAT1 (K+ channel in Arabidopsis thaliana 1) to causestomatal closure (12, 13).ABA also triggers the generation of several second-messenger
molecules, such as calcium, inositol phospholipids, and nitricoxide (NO) and reactive oxygen species (ROS) (14–17). Thesesecond messengers are involved in ABA regulation of stomatalclosure and other physiological processes (15, 18–20). Amongthese second messengers, NO has been well documented to haveimportant roles in ABA signal transduction. ABA induces NOgeneration in roots and guard cells (15, 21). Exogenous applicationof NO can trigger stomatal closure, whereas application of theNO scavenger 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (c-PTIO) inhibits stomatal closure (22), sug-gesting a positive role of exogenous NO in stomatal closure.Several studies have suggested that exogenous NO may affectstomatal responses to ABA by regulating an inward K+ channel(20), an anion channel (23), and the generation of nitratedcGMP (24), although the direct target of NO in ABA signalingremains unclear.In plants, NO is produced by the nitrite-dependent nitrate re-
ductase pathway (15, 25) and a pathway dependent on the nitricoxide associated 1 (NOA1) protein (21), although NOA1 is notan NO synthase (26). NO regulates many physiological processesin plants, including responses to phytohormones, such as ABA,cytokinin, auxin, gibberellins, and salicylic acid, immunity againstpathogens, senescence, and flowering (15, 27–31). The deficiencyin NO generation in the nia1nia2noa1 triple mutant results inABA-hypersensitive stomatal closure (21), suggesting a negative
Significance
Drought stress induces the accumulation of the plant stresshormone abscisic acid (ABA). ABA then quickly activates theprotein kinase OST1/SnRK2.6 to phosphorylate a number ofproteins in guard cells, resulting in stomatal closure to reducetranspirational water loss. How SnRK2.6 is deactivated andhow ABA signaling may be desensitized are unclear. This studyfound that nitric oxide (NO) resulting from ABA signalingcauses S-nitrosylation of SnRK2.6 at a cysteine residue closeto the kinase catalytic site, which blocks the kinase activity.Dysfunction of S-nitrosoglutathione (GSNO) reductase causesGSNO overaccumulation in guard cells and ABA insensitivity instomatal regulation. This work thus reveals how ABA-inducedNO functions in guard cells to inactivate SnRK2.6 to negativelyfeedback regulate ABA signaling.
Author contributions: P.W. and J.-K.Z. designed research; P.W., Y.D., Y.-J.H., Y.Z., C.-C.H.,F.Y., and X.Z. performed research; P.W., Y.D., W.A.T., C.-P.S., and J.-K.Z. analyzed data;and P.W., X.Z., and J.-K.Z. wrote the paper.
The authors declare no conflict of interest.1P.W. and Y.D. contributed equally to this work.2To whom correspondence should be addressed. Email: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1423481112/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1423481112 PNAS | January 13, 2015 | vol. 112 | no. 2 | 613–618
PLANTBIOLO
GY

role of endogenous NO in ABA signaling. NO overaccumulatesin Arabidopsis gsnor1(S-nitrosoglutathione reductase 1)/hot5 (sen-sitive to hot temperatures 5)/par2 (paraquat resistant 2) mutantplants that are impaired in the GSNO reductase gene (32–34).The gsnor1-3 mutant is hypersensitive to heat stress (33) andbacterial pathogen (28, 31), but is more resistant to oxidativestress (34). Characterization of gsnor1 mutant plants suggestedthat GSNOR regulates multiple developmental and metabolicprograms in Arabidopsis (35). In cytokinin signaling, NO causesS-nitrosylation and inhibition of the histidine phosphotransferprotein AHP1 (Arabidopsis histidine phosphotransfer protein 1)(27). In salicylic acid signaling, the S-nitrosylation of the salicylicacid receptor NPR1 facilitates its oligomerization (31). In addi-tion, NO regulates cell death in plant immunity by S-nitrosylationof the NADPH oxidase AtRBOHD (28).Here, we show that GSNO and Cys-NO (S-nitrosocysteine)
can inhibit SnRK2.6 by S-nitrosylation. The S-nitrosylation ofSnRK2.6 occurs at Cys137, which is adjacent to the catalytic loopof the kinase. A Cys137 to Ser mutation causes the kinase to beresistant to inhibition by GSNO in vitro, whereas a Cys137 to Trpmutation results in an inactive kinase in vitro and in vivo. In-troduction of the Cys137 to Ser mutated form of SnRK2.6 intogsnor1-3/ost1-3 (open stomata 1–3, a null allele of snrk2.6) par-tially suppresses the defect in stomatal closure caused by over-accumulation of SNOs because of the gsnor1-3 mutation. Ourresults suggest that ABA induced S-nitrosylation of SnRK2.6functions to negatively feedback regulate ABA signaling in plants.Moreover, the S-nitrosylation of cysteine137 is evolutionarilyconserved in some AMPK/SNF1-related kinases and glycogensynthase kinase 3/SHAGGY-like kinases (SKs) in plants, yeast,and mammals, suggesting that S-nitrosylation–mediated inhi-bition may be a general regulatory mechanism for these eukary-otic protein kinases.
ResultsGSNO Inhibits SnRK2.6 by S-Nitrosylation of Cys-137 in Vitro. To testwhether NO may regulate the activity of components in the coreABA signaling pathway, we tested the effects of the NO donors,GSNO, Cys-NO, and SNAP (S-Nitroso-N-acetylpenicillamine),on the activity of SnRK2.6 in vitro because SnRK2.6 is a keyplayer in the ABA regulation of guard cells and its activity can bereadily assayed in vitro and in vivo. As shown in Fig. 1A and Fig.S1, GSNO, Cys-NO, and SNAP inhibited the activity of SnRK2.6in a dose-dependent manner, but such inhibitory effects werereversed by DTT, indicating that the effects of NO donors relyon the thiol-based redox status. To determine whether SnRK2.6is S-nitrosylated in the presence of GSNO, we used a biotin-switch method in which the S-nitrosylated cysteine is labeledwith biotin and subsequently detected by antibiotin immuno-blot analysis (27, 31). As shown in Fig. 1B, GSNO inducedS-nitrosylation of SnRK2.6 in a dose-dependent manner, and ap-plication of DTT abolished the GSNO induced S-nitrosylation.Six cysteine residues in SnRK2.6 are putative target sites ofS-nitrosylation. To determine which cysteine is required forGSNO-mediated inhibition, we mutated each cysteine (C) toserine (S) separately and tested the effect of GSNO on themutated SnRK2.6. Of the six mutations, only the Cys137Ser(C137S) mutation eliminated the inhibitory effect of GNSO (Fig.1C). The C137S mutation also caused a slight reduction inSnRK2.6 kinase activity (Fig. 1 C and D). The SnRK2.6C137W
with a Trp substitution at Cys137 that may mimic S-nitrosylation(27, 36) did not show any detectable kinase activity (Fig. 1D).Based on the crystal structure of SnRK2.6 resolved by a recentstudy (37), the side-chain of C137 is exposed on the surface andhas only 8 Å distance from D140 in the catalytic loop and D160in the Mg-binding loop in the catalytic cleft of SnRK2.6 (Fig. 1E,Left). The C137W mutation likely interferes directly with thekinase catalytic activity.
To further verify the S-nitrosylation of SnRK2.6 at Cys137, wesubjected freshly purified SnRK2.6 to a biotin-switch assay,where free cysteines are blocked with S-methymethane thiosul-fonate (MMTS) and nitrosylated cysteines are labeled withbiotin. Mass spectrometry was performed immediately after thelabeling of biotin. The assay detected the S-nitrosylation of Cys137in SnRK2.6 but none of the other cysteine was S-nitrosylated inthe presence of the blocking reagent MMTS (Fig. S2). In theabsence of MMTS, Cys131, Cys159, Cys203, and Cys250 werealso labeled by biotin (Fig. S3). The peptide containing Cys107was too short to be detected by mass spectrometry after trypsindigestion. These results suggested that S-nitrosylation occurredspecifically at Cys137 after GSNO treatment. We also detectedthe dehydrogenation of Cys137 and Cys131 in the tryptic fragmentsafter the biotin-switch assay (Fig. S4), suggesting that a disulfidebond may form between Cys137 and Cys131 in some SnRK2.6molecules. Interestingly, the C131S mutation in SnRK2.6 (C131S)appeared to increase the level of detectable S-nitrosylation relativeto the wild-type SnRK2.6 (WT) (Fig. S5). When Cys137 was mu-tated, virtually no S-nitrosylated SnRK2.6 was detected in thepresence of GSNO (Fig. S5). Collectively, these results show thatSnRK2.6 is S-nitrosylated at Cys137 in the presence of NO donorsand that the S-nitrosylation of SnRK2.6 blocks its kinase ac-tivity in vitro.
ABA Enhances the S-Nitrosylation of SnRK2.6 in Vivo. To furtherconfirm that the Cys137 is modified by S-nitrosylation in vivo, wesubjected total protein extracts from ost1-3 mutant plants com-plemented with native promoter driven GFP-tagged wild-typeSnRK2.6 (ost1-3/SnRK2.6WT-GFP) to biotin-switch analysis. Biotin-labeled SnRK2.6-GFP was purified with streptavidin beads anddetected by anti-GFP immunoblot analysis. As shown in Fig. 2A,a low level of S-nitrosylation of SnRK2.6 could be detectedunder control conditions and exogenous GSNO treatment on the
5000GSNOGSNO
50100 250 500DTT - - - - +
Autoradiograph
-A
Coomassie blue
C
SNO
Total
E
anti-biotin
anti-MBP
Autoradiograph
Coomassie blue
L72H69E65
D140K142
D160
F161
ATP binding loop catalytic loop
C137
αC
αE L72H69E65
D140K142
D160
F161
ATP binding loopcatalytic loop
W137
αC
αE
SnRK2.6(WT) SnRK2.6(C137W)
0 50100250500DTT - - - - +
500 (μM)-
GSNODTT
- ++ -
- ++ -
- ++ -
- ++ -
- ++ -
- ++ -
- ++ -
GSNODTT
- ++ -
- ++ -
- ++ -
Autoradiograph
Coomassie blue
B
D
Autoradiograph
Coomassie blue
500 (μM)0Cys-NO
50100 250 500- - - - +-
Fig. 1. S-nitrosylation at Cys-137 inhibits the activity of SnRK2.6. (A) Nitricoxide donors GSNO and Cys-NO inhibit the activity of SnRK2.6 in a dose-dependent manner. MBP–SnRK2.6 incubated with indicated concentrationof GSNO (Left) and Cys-NO (Right) for 10 min and then [γ-32P]ATP was addedto determine the autophosphorylation of SnRK2.6. In the rightmost lane(DTT+), 1 mM DTT was added into the reaction before adding [γ-32P]ATP.(B) GSNO causes S-nitrosylation of SnRK2.6 as detected by the biotin-switchassay. (C) Effects of C-to-S site-directed mutation of the six cysteines onSnRK2.6 activity upon GSNO (50 μM) or DTT treatment. (D) Effects of C137Sand C137W mutations on the kinase activity of SnRK2.6. (E) Structure ofSnRK2.6 showing the position of Cys-137 (Left) and Trp-137 (Right). ResiduesE65, H69, L72, K142, D160, F161, and C137 (W137) are shown by sticks.
614 | www.pnas.org/cgi/doi/10.1073/pnas.1423481112 Wang et al.

seedlings increased the level of S-nitrosylated SnRK2.6 in ost1-3/SnRK2.6WT-GFP transgenic plants. In ost1-3/SnRK2.6C137S-GFPplants, in which the Cys137 is mutated to Serine, S-nitrosylatedSnRK2.6 was undetectable. These results suggest that S-nitrosylationof SnRK2.6 at Cys137 occurs in vivo.We compared the water loss and ABA-induced stomatal clo-
sure in Col, ost1-3, and transgenic plants expressing wild-typeand mutated forms of SnRK2.6. Only SnRK2.6WT-GFP com-plemented the water loss (Fig. 2B) and stomatal closure(Fig. 2C) phenotypes of ost1-3. The transgenic plant ost1-3/SnRK2.6C137W-GFP, which contains the S-nitrosylation–mimickingmutation at Cys137, was insensitive to ABA-induced stomatal clo-sure (Fig. 2C) and had a higher rate of water loss than the wild-type(Fig. 2B). SnRK2.6C137S-GFP partially complemented the pheno-types of ost1-3 (Fig. 2 B and C). Consistent with these phenotypes,in-gel kinase assays showed that SnRK2.6C137S-GFP had a reducedkinase activity and that SnRK2.6C137W-GFP is a “dead” kinasein vivo (Fig. 2D). Taken together, our in vitro and in vivo datasuggest that S-nitrosylation at Cys137 of SnRK2.6 abolishes itskinase activity.
Consistent with a previous report (15), we found that ABAincreased NO generation in guard cells (Fig. 2E). To testwhether ABA affects the S-nitrosylation of SnRK2.6, we exam-ined the S-nitrosylation of SnRK2.6-GFP upon ABA treatment.Treatment with 50 μM ABA for 15 min did not increase theS-nitrosylation of SnRK2.6. However, ABA treatment for 30 or60 min increased the level of SnRK2.6 S-nitrosylation (Fig. 2F).As expected, preincubation with the NO-scavenger c-PTIO blockedS-nitrosylation of SnRK2.6 (Fig. 2F). These results indicate thatprolonged treatment with ABA enhances the S-nitrosylationof SnRK2.6.
The NO Overaccumulation Mutant gsnor1-3 Is Insensitive to ABA-Induced Stomatal Closure, a Phenotype That Is Partially Suppressedby the Cys137S Mutation in SnRK2.6. We hypothesized that NOproduction inside guard cells may negatively regulate ABAsignaling through SnRK2.6 S-nitrosylation. To test this hy-pothesis, we investigated ABA responses in gsnor1-3 mutantplants, which overproduce SNOs because of a deficiency of theGSNO reductase in Arabidopsis (32). S-nitrosylation of proteinssuch as AHP1 (27) and SABP3 (salicylic acid-binding protein 3)(38), is increased in gsnor1-3 mutant plants. In gsnor1-3 leaves,NO overaccumulated in guard cells, where the GSNOR1 isexpressed (Fig. S6 A and B). Without ABA treatment, the levelof S-nitrosylation of SnRK2.6 was higher in gsnor1-3/ost1-3/SnRK2.6 than ost1-3/SnRK2.6 plants (Fig. 3A). After ABAtreatment, the S-nitrosylation levels were similar. In the gsnor1-3mutant, ABA was substantially less effective in inducing stomatalclosure than in the Col-0 wild-type plants (Fig. 3B). However,exogenous H2O2, but not SNP or GSNO, could still inducestomatal closure in gsnor1-3 (Fig. S7A). The stomatal in-sensitivity of gnsor1-3 to ABA is similar to that of the ost1-3,abi1-1, and abi2-1 mutants (Fig. 3B) (39, 40). Like these singlemutants, gsnor1-3/abi1-1 and gsnor1-3/ost1-3 double-mutantplants are equally insensitive to ABA in stomatal closure (Fig.3B). Consistent with the impaired ABA response in guard cells,the excised leaves of gsnor1-3 showed a higher water-loss ratethan wild-type leaves (Fig. 3C). Two other T-DNA insertionmutants, gsnor1-4 and hot5-4 in the WS (Wassilewskija) back-ground, also exhibited higher water loss than their wild-typecontrol plants (Fig. 3C). However, hot5-1, which is anothermissense allele of GSNOR1 in the Col-0 background (33),showed a similar water loss rate as the Col-0 wild-type (Fig.3C). Immunoblot and GSNOR activity assays revealed thatgsnor1-3, gsnor1-4, and hot5-4 were virtually null alleles (Fig.S7 D and E) (33), whereas hot5-1 plants contained a similaramount of GSNOR protein as the wild-type and retainedabout half of the GSNOR activity (Fig. S7 D and E) (33).Therefore, the transpirational water-loss phenotypes are con-sistent with the GSNOR protein and activity levels. Consistentwith its higher transpirational water loss, the leaf surfacetemperature of gsnor1-3 was significantly lower than that of theCol-0 wild-type, especially under drought stress (Fig. S8). Ourresults suggest that increased NO accumulation in the gsnor1mutants causes elevated S-nitrosylation of SnRK2.6, resultingin ABA insensitivity in guard cells and higher transpirationalwater loss.We examined the kinase activity of SnRK2.6 in gsnor1 mu-
tant plants using an in-gel kinase assay (11, 41), which involvesseparating plant proteins in SDS-polyacrylamide gels contain-ing ABF2 fragments as a kinase substrate and renaturation ofthe proteins with DTT after removal of SDS from the gels beforedetecting protein bands with kinase activities by incubation with[λ-32P]ATP. The assay revealed higher SnRK2.6 kinase activities inhot5-4 and gsnor1-4 mutant plants (Fig. S7F). Because the DTTused for protein renaturation in the assay would reverseS-nitrosylation and possibly some other modifications on Cysresidues of SnRK2.6, the results suggest that S-nitrosylated
0 20 40 60 80 1000
10203040506070
Wat
er lo
ss (%
)
ost1-3 Col-0
ost1-3/2.6C137W ost1-3/2.6C137S ost1-3/2.6WT
D
in-gel kinase assay
anti-GFP
Time (min)
A
C
E
0 30 60
SNO-SnRK2.6
Total
c-PTIO
Fluo
resc
ence
inte
nsity
(A
rbitr
ary
unit)
0
20
60
80
40
100 control
15 30 60
ABA
0
15
ost1-3/ SnRK2.6
ost1-3/ 2.6C137S
GSNO
SNO-SnRK2.6
Total
+ + - -
B S
tom
atal
ape
rture
(μm
)
1
3
4
2
5
- + - + - + - + - +
anti-GFP
anti-GFP
+ 50 μM ABA (min)
Time (min)
F
ABA * * *
* *
*
* * *
0
control ABA
Fig. 2. ABA up-regulates the S-nitrosylation of Cys-137 of SnRK2.6 in vivo.(A) GSNO-induced S-nitrosylation of wild-type but not Cys173Ser mutatedform of SnRK2.6-GFP from plants. Twelve-day-old seedlings were treatedwith or without 250 μM GSNO for 30 min. The biotin-switch and immunoblotassays were performed as described in SI Materials and Methods. (B and C)Analysis of water loss (B) and stomata responses to 10 μMABA (C) in the wild-type, ost1-3 mutant, and ost1-3 transgenic lines expressing SnRK2.6WT-GFP,SnRK2.6C137S-GFP, and SnRK2.6C137W-GFP. Treatment with water was used ascontrol. Error bars indicate SD. n = 3–5 independent experiments. More than20 stomata were measured for each treatment in each experiment inC. Student’s t test, *P < 0.05 (significantly different from ost1-3). (D) In-gelkinase assay showing SnRK2.6 activity before (−) and after (+) 50 μM ABAtreatment in the wild-type, the ost1-3 mutant, and ost1-3 transgenic linesexpressing SnRK2.6WT-GFP, SnRK2.6C137S-GFP, and SnRK2.6C137W-GFP. Thepositions of SnRK2.6 and GFP-fused SnRK2.6 are indicated by open and closedtriangles, respectively. (E) Average FDA-FM fluorescence intensity indicatingNO levels in guard cells upon ABA (10 μM) treatment in Col wild-type.Treatment with water was used as control. Error bars indicate SD (n > 6 from3 independent experiments). Student’s t test, *P < 0.01. (F) ABA regulation ofS-nitrosylation of SnRK2.6 in ost1-3/SnRK2.6WT-GFP transgenic plants. Twelve-day-old seedlings were treated with or without 50 μM ABA for the indicatedtimes. The seedlings treated with 200 μM c-PTIO were used as a control. Thebiotin-switch and immunoblot assays were performed as described in SIMaterials and Methods.
Wang et al. PNAS | January 13, 2015 | vol. 112 | no. 2 | 615
PLANTBIOLO
GY
and other Cys-modified SnRK2.6 in the gsnor1 mutants arenot further subjected to any irreversible and inhibitory mod-ifications.To confirm that the ABA insensitivity in stomatal closure in
gsnor1 mutant plants is a result of SnRK2.6 S-nitrosylation, we in-troduced native promoter-driven SnRK2.6WT and SnRK2.6C137S intothe gsnor1-3mutant by crossing with ost1-3mutant plants expressingthe SnRK2.6 constructs, and analyzed the water loss and stomatalresponses in the resulting gsnor1-3/ost1-3/SnRK2.6WT and gsnor1-3/ost1-3/SnRK2.6C137S plants. The SnRK2.6C173S mutant but not thewild-type SnRK2.6 partially restored ABA-induced stomatal closurein the gsnor1-3 mutant (Fig. 3D). The water-loss defect of thegsnor1-3/ost1-3 double-mutant was also partially rescued by in-troduction of SnRK2.6C173S but not the wild-type SnRK2.6 (Fig. 3E).
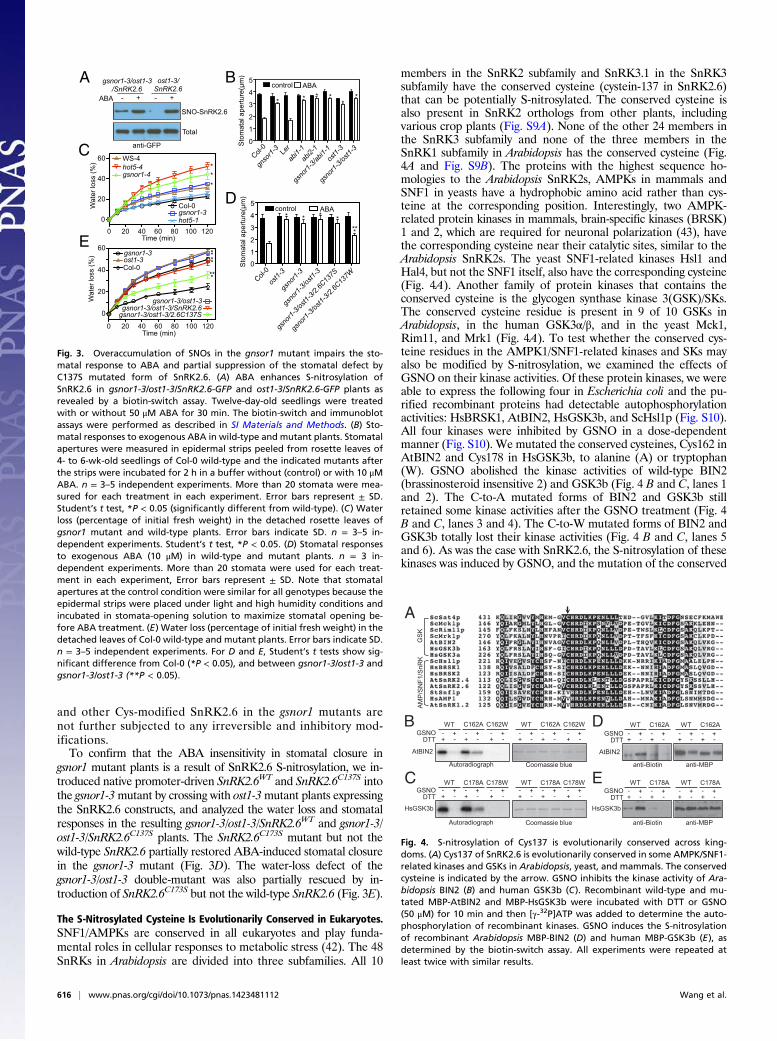
The S-Nitrosylated Cysteine Is Evolutionarily Conserved in Eukaryotes.SNF1/AMPKs are conserved in all eukaryotes and play funda-mental roles in cellular responses to metabolic stress (42). The 48SnRKs in Arabidopsis are divided into three subfamilies. All 10
members in the SnRK2 subfamily and SnRK3.1 in the SnRK3subfamily have the conserved cysteine (cystein-137 in SnRK2.6)that can be potentially S-nitrosylated. The conserved cysteine isalso present in SnRK2 orthologs from other plants, includingvarious crop plants (Fig. S9A). None of the other 24 members inthe SnRK3 subfamily and none of the three members in theSnRK1 subfamily in Arabidopsis has the conserved cysteine (Fig.4A and Fig. S9B). The proteins with the highest sequence ho-mologies to the Arabidopsis SnRK2s, AMPKs in mammals andSNF1 in yeasts have a hydrophobic amino acid rather than cys-teine at the corresponding position. Interestingly, two AMPK-related protein kinases in mammals, brain-specific kinases (BRSK)1 and 2, which are required for neuronal polarization (43), havethe corresponding cysteine near their catalytic sites, similar to theArabidopsis SnRK2s. The yeast SNF1-related kinases Hsl1 andHal4, but not the SNF1 itself, also have the corresponding cysteine(Fig. 4A). Another family of protein kinases that contains theconserved cysteine is the glycogen synthase kinase 3(GSK)/SKs.The conserved cysteine residue is present in 9 of 10 GSKs inArabidopsis, in the human GSK3α/β, and in the yeast Mck1,Rim11, and Mrk1 (Fig. 4A). To test whether the conserved cys-teine residues in the AMPK1/SNF1-related kinases and SKs mayalso be modified by S-nitrosylation, we examined the effects ofGSNO on their kinase activities. Of these protein kinases, we wereable to express the following four in Escherichia coli and the pu-rified recombinant proteins had detectable autophosphorylationactivities: HsBRSK1, AtBIN2, HsGSK3b, and ScHsl1p (Fig. S10).All four kinases were inhibited by GSNO in a dose-dependentmanner (Fig. S10). We mutated the conserved cysteines, Cys162 inAtBIN2 and Cys178 in HsGSK3b, to alanine (A) or tryptophan(W). GSNO abolished the kinase activities of wild-type BIN2(brassinosteroid insensitive 2) and GSK3b (Fig. 4 B and C, lanes 1and 2). The C-to-A mutated forms of BIN2 and GSK3b stillretained some kinase activities after the GSNO treatment (Fig. 4B and C, lanes 3 and 4). The C-to-W mutated forms of BIN2 andGSK3b totally lost their kinase activities (Fig. 4 B and C, lanes 5and 6). As was the case with SnRK2.6, the S-nitrosylation of thesekinases was induced by GSNO, and the mutation of the conserved
0 20 40 60 80 100 120
control ABA
Sto
mat
al a
pertu
re(μ
m)B
3
5
A W
ater
loss
(%)
Time (min)
gsnor1-3
gsnor1-3/ost1-3
ost1-3
gsnor1-3/ost1-3/SnRK2.6gsnor1-3/ost1-3/2.6C137S
Col-0
C
E
0
20
40
60
gsnor1-3/ost1-3 /SnRK2.6
ost1-3/ SnRK2.6
ABA
SNO-SnRK2.6
Total
+ + - -
anti-GFP
Sto
mat
al a
pertu
re(μ
m)
0
1
3
4
2
5 control ABA
D
*
* * * * * *
* *
*
* *
**
* * * * **
0
1
3
4
2
5
0 20 40 60 80 100 120
Wat
er lo
ss (%
)
Col-0 gsnor1-3
WS-4 hot5-4 gsnor1-4
hot5-1 0
20
40
60
Time (min)
* * *
Fig. 3. Overaccumulation of SNOs in the gnsor1 mutant impairs the sto-matal response to ABA and partial suppression of the stomatal defect byC137S mutated form of SnRK2.6. (A) ABA enhances S-nitrosylation ofSnRK2.6 in gsnor1-3/ost1-3/SnRK2.6-GFP and ost1-3/SnRK2.6-GFP plants asrevealed by a biotin-switch assay. Twelve-day-old seedlings were treatedwith or without 50 μM ABA for 30 min. The biotin-switch and immunoblotassays were performed as described in SI Materials and Methods. (B) Sto-matal responses to exogenous ABA in wild-type and mutant plants. Stomatalapertures were measured in epidermal strips peeled from rosette leaves of4- to 6-wk-old seedlings of Col-0 wild-type and the indicated mutants afterthe strips were incubated for 2 h in a buffer without (control) or with 10 μMABA. n = 3–5 independent experiments. More than 20 stomata were mea-sured for each treatment in each experiment. Error bars represent ± SD.Student’s t test, *P < 0.05 (significantly different from wild-type). (C) Waterloss (percentage of initial fresh weight) in the detached rosette leaves ofgsnor1 mutant and wild-type plants. Error bars indicate SD. n = 3–5 in-dependent experiments. Student’s t test, *P < 0.05. (D) Stomatal responsesto exogenous ABA (10 μM) in wild-type and mutant plants. n = 3 in-dependent experiments. More than 20 stomata were used for each treat-ment in each experiment, Error bars represent ± SD. Note that stomatalapertures at the control condition were similar for all genotypes because theepidermal strips were placed under light and high humidity conditions andincubated in stomata-opening solution to maximize stomatal opening be-fore ABA treatment. (E) Water loss (percentage of initial fresh weight) in thedetached leaves of Col-0 wild-type and mutant plants. Error bars indicate SD.n = 3–5 independent experiments. For D and E, Student’s t tests show sig-nificant difference from Col-0 (*P < 0.05), and between gsnor1-3/ost1-3 andgsnor1-3/ost1-3 (**P < 0.05).
GS
KA
MP
/SN
F1/S
nRK
AtBIN2
HsGSK3b
WT C162A C162WGSNO
DTT -++-
Autoradiograph Coomassie blue
A
B
anti-Biotin anti-MBP
EC
D-++-
-++-
WT C162A C162W
-++-
-++-
-++-
WT C162A
-++-
-++-
WT C162A
-++-
-++-GSNO
DTT
AtBIN2
WT C178A C178WGSNO
DTT -++-
-++-
-++-
WT C178A C178W
-++-
-++-
-++-
WT C178A
-++-
-++-
WT C178A
-++-
-++-GSNO
DTTHsGSK3b
Autoradiograph Coomassie blue anti-Biotin anti-MBP
Fig. 4. S-nitrosylation of Cys137 is evolutionarily conserved across king-doms. (A) Cys137 of SnRK2.6 is evolutionarily conserved in some AMPK/SNF1-related kinases and GSKs in Arabidopsis, yeast, and mammals. The conservedcysteine is indicated by the arrow. GSNO inhibits the kinase activity of Ara-bidopsis BIN2 (B) and human GSK3b (C). Recombinant wild-type and mu-tated MBP-AtBIN2 and MBP-HsGSK3b were incubated with DTT or GSNO(50 μM) for 10 min and then [γ-32P]ATP was added to determine the auto-phosphorylation of recombinant kinases. GSNO induces the S-nitrosylationof recombinant Arabidopsis MBP-BIN2 (D) and human MBP-GSK3b (E), asdetermined by the biotin-switch assay. All experiments were repeated atleast twice with similar results.
616 | www.pnas.org/cgi/doi/10.1073/pnas.1423481112 Wang et al.

cysteine blocked the S-nitrosylation of the proteins (Fig. 4 D andE). Our finding of the regulation of BIN2 by S-nitrosylation isconsistent with the observation that brassinosteroid treatmentinduces NO accumulation in maize leaves (44). The conservationof the cysteine and the S-nitrosylation–dependent regulation of theactivities of the kinases from diverse organisms suggest thatS-nitrosylation–mediated inhibition is a general regulatorymechanism for these eukaryotic protein kinases.
DiscussionOur study has revealed a novel mechanism by which NO regu-lates ABA signaling in Arabidopsis. We provided evidence thatABA enhances SnRK2.6 S-nitrosylation and the S-nitrosylationfeedback inhibits ABA signaling. The evidence includes: (i) ex-ogenous NO donors caused S-nitrosylation of SnRK2.6 andinhibited its activity in vitro, and an S-nitrosylation-mimickingmutation abolished its kinase activity; (ii) ABA enhanced thelevel of nitrosylated SnRK2.6 in planta; and (iii) NO-over-producing mutant plants were less sensitive to ABA in stomatalclosure, but the defect was partially suppressed by ectopic ex-pression of C137S mutated form of SnRK2.6. Our study revealedSnRK2.6 as a new S-nitrosylation target that bridges NO-medi-ated redox signaling and ABA signaling.In the absence of ABA, PP2Cs inhibit SnRK2.6 activity by
both dephosphorylation at Ser175 and direct binding (45). In thepresence of ABA, however, the binding of PYLs to PP2Csreleases SnRK2.6 from inhibition. SnRK2.6 is activated veryquickly by ABA because 2 min of ABA treatment is enough tocause a strong activation of SnRK2.6 (41). In contrast to theearly activation of SnRK2.6, NO-mediated inhibition of SnRK2.6is expected to happen later during ABA treatment because thelevel of S-nitrosylated SnRK2.6 is not increased after 15 min ofABA application (Fig. 2F). NO accumulation did not reach peaklevels until 30 min after ABA application (Fig. 2E) (46). Thenotion that NO accumulation and SnRK2.6 S-nitrosylation areslow or late events during ABA treatment is consistent with theobservation that ABA-induced NO generation is dependent onthe quick burst of ROS (47). The accumulation of endogenousNO may function as one of the negative feedback mechanisms toprevent overactivation of ABA signaling in guard cells. Thisfeedback regulation is achieved by S-nitrosylation of SnRK2.6.ABA-activated SnRK2.6 phosphorylates NADPH oxidases tocause ROS production in guard cells (18, 48). The S-nitrosylationof the NADPH oxidase AtRBOHD has been shown to causeinhibition of the NADPH oxidase (28), and thus may alsocontribute to the NO-mediated negative feedback regulation ofABA signaling. Although detailed dynamic changes in the levelsof activated SnRK2.6, NO, and S-nitrosylated SnRK2.6 in guardcells are not yet known, available evidence suggests the followingscenario: ABA treatment causes very fast and strong activationof SnRK2.6 in guard cells. The activated SnRK2.6 phosphor-ylates many downstream effector proteins to cause stomatalclosure, and it also phosphorylates the NADPH oxidasesAtRBOHD and AtRBOHF to cause a burst of ROS, which inturn causes NO accumulation in the guard cells. When NOaccumulates to high levels, it causes S-nitrosylation and inhibitionof SnRK2.6 and the NADPH oxidases. This inhibition serves todesensitize ABA signaling. The phenomenon of desensitization ofABA signaling was observed 20 y ago, when water deficit stresswas shown to reduce the stomatal sensitivity to ABA (49).In gsnor1 mutant plants, overaccumulated SNOs cause con-
stitutive S-nitrosylation of SnRK2.6 such that the kinase is in-activated, and thus the stomata are insensitive to ABA (Fig. 3).In support of our conclusion that endogenous NO has a negativerole in ABA signaling in guard cells, a recent study demonstratedthat the NO-deficient mutant nia1nia2noa1 is hypersensitive toABA in stomatal closure (21). The kinase activity of S-nitro-sylated SnRK2.6 may be resumed quickly, depending on cellular
redox homeostasis. Protein S-nitrosylation depends on theproximity of the protein to NO sources, and on factors fortransnitrosylation and denitrosylation, such as thioredoxin andglutathione levels (50). The reversible inhibition of SnRK2 byS-nitrosylation may fine-tune the strength or duration of SnRK2.6activation in response to ABA. Such dynamic control may beimportant for plants to balance stress resistance with growth thatcannot happen if stomata are fully closed as a result of continuedactivation of SnRK2.6. Because NO originates from nitrogensources, NO-mediated stomatal regulation through SnRK2.6S-nitrosylation may also help coordinate nitrogen supply withphotosynthetic carbon availability.The negative effect of endogenous NO on ABA signaling in
guard cells is supported by genetic analysis using both NO de-ficient and overaccumulation mutants (Fig. 3) (21). On the otherhand, several studies have found a positive role of exogenousapplication of NO in ABA-induced stomatal closure (15, 20,22, 24). Exogenous NO might promote stomatal closure byregulating ion channels (20, 23), or by generating nitrated cGMP(24). Alternatively, the observed positive role of exogenousNO on ABA responses in guard cells might be caused by sec-ondary effects, such as induction of ROS or other second mes-sengers (24, 47).The activities of protein kinases are tightly controlled by multiple
mechanisms. In most cases, dephosphorylation at the phosphory-lated sites in the “activation segment” causes kinase deactivation(51). For SnRK2.6, the phosphorylation of Ser175 is necessary forits full activity (37). The negative regulators of ABA signaling,PP2Cs, inhibit SnRK2.6 partly by dephosphorylation at Ser175.Similarly, in brassinosteroid signaling, AtBIN2 is dephosphorylatedand inactivated by BSU1 (bri1 SUPPRESSOR 1), a Kelch-repeatdomain-containing protein phosphatase (52). In contrast to thephosphorylation-mediated activation, the phosphorylation ofGSK3 at serine 9 by Akt kinase inhibits its activity (53), suggestinga “pseudo-substrate” mechanism to reduce the kinase activity (51).Our finding that GSNO-mediated S-nitrosylation of HsBRSK1,ScHsl1p, GSKs, and AtSnRK2s suggests a third way to deactivatethese protein kinases. The cysteine in the catalytic cleft exists onlyin certain members of the SnRK/GSK family. In most proteinkinases, the conserved cysteine is substituted by a hydrophobicamino acid. Other cysteines not in the position near the catalyticloop could still be modified by S-nitrosylation, leading to the in-hibition of some kinases (54–56). The S-nitrosylation of cysteinein the catalytic cleft reported here represents a unique mechanismin the regulation of some of the AMPK/SNF1-related and GSKfamily of protein kinases.
Materials and MethodsPlant Materials and Growth Condition. The Arabidopsis wild-type Columbia-0(Col-0), Wassilewskija-4 (WS-4), and Landsberg (Ler) plants were used in thisstudy. The gsnor1-3 (GABI_315D11), hot5-1 (CS66011), hot5-4 (FLAG_298F11),gsnor1-4 (FLAG_220G07), and ost1-3 (Salk_008068) mutant seeds were orderedfrom the Arabidopsis Biological Resource Center. Seeds were germinated onhalf-strength MS agar plates containing 1.5% sucrose. For the measurementof water loss or stomatal apertures, 10-d-old seedlings on MS plates weretransferred to soil and grown under short-day conditions (12 h light, 100–120 μmol·m−2·s−1) at 22 °C.
Epidermal Strip Bioassay and Water Loss Measurement. Stomatal bioassayexperiments were performed as described previously (15). Epidermal stripswere peeled from the rosette leaves of 4- to 6-wk-old seedlings that wereplaced under the light and high humidity for 12 h and incubated in stomata-opening solution containing 50 mM KCl, 10 mM MES, pH 6.15 in a growthchamber for 1.5 h to maximize stomatal opening before ABA was added.Stomatal apertures were measured 2 h after 10 μM ABA was added. Theapertures of about 60 stomata were measured in three independentexperiments.
For the measurement of water loss, detached rosette leaves of 4-wk-oldplants were placed in weighing dishes and left on the laboratory benchwith
Wang et al. PNAS | January 13, 2015 | vol. 112 | no. 2 | 617
PLANTBIOLO
GY

light (30–40 μmol·m−2·s−1). Fresh weight was monitored at the indicatedtimes. Water loss was expressed as a percentage of initial fresh weight.
In Vitro Kinase Assay and In-Gel Kinase Assay. The in vitro kinase assay and in-gel kinase assay were performed as described previously (11); details areprovided in SI Materials and Methods.
In Vitro and in Vivo S-Nitrosylation Assays. In vitro and in vivo S-nitrosylationassays were performed as described (28, 31); details are provided inSI Materials and Methods.
Other Methods. Details for other methods are provided in SI Materials andMethods, including recombinant protein expression and site-directed mu-tagenesis, structure information of SnRK2.6, histochemical detection of GUSactivity, infrared thermography imaging, NO detection by confocal micros-copy, and homology search and sequence alignment. The primers used inthis study are listed in Table S1.
ACKNOWLEDGMENTS. This work was supported by NIH Grant R01GM059138(to J.-K.Z.) and National Natural Science Foundation of China Grants 91017001and 31171363 (to P.W.).
1. Cutler SR, Rodriguez PL, Finkelstein RR, Abrams SR (2010) Abscisic acid: Emergence ofa core signaling network. Annu Rev Plant Biol 61:651–679.
2. Klingler JP, Batelli G, Zhu JK (2010) ABA receptors: The START of a new paradigm inphytohormone signalling. J Exp Bot 61(12):3199–3210.
3. Lee KH, et al. (2006) Activation of glucosidase via stress-induced polymerizationrapidly increases active pools of abscisic acid. Cell 126(6):1109–1120.
4. Fujii H, et al. (2009) In vitro reconstitution of an abscisic acid signalling pathway.Nature 462(7273):660–664.
5. Ma Y, et al. (2009) Regulators of PP2C phosphatase activity function as abscisic acidsensors. Science 324(5930):1064–1068.
6. Park S-Y, et al. (2009) Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 324(5930):1068–1071.
7. Wang P, et al. (2013) Quantitative phosphoproteomics identifies SnRK2 protein ki-nase substrates and reveals the effectors of abscisic acid action. Proc Natl Acad Sci USA110(27):11205–11210.
8. Kim TH, Böhmer M, Hu H, Nishimura N, Schroeder JI (2010) Guard cell signal trans-duction network: advances in understanding abscisic acid, CO2, and Ca2+ signaling.Annu Rev Plant Biol 61:561–591.
9. Léon-Kloosterziel KM, et al. (1996) Isolation and characterization of abscisic acid-deficient Arabidopsis mutants at two new loci. Plant J 10(4):655–661.
10. Gonzalez-Guzman M, et al. (2012) Arabidopsis PYR/PYL/RCAR receptors play a majorrole in quantitative regulation of stomatal aperture and transcriptional response toabscisic acid. Plant Cell 24(6):2483–2496.
11. Fujii H, Verslues PE, Zhu J-K (2007) Identification of two protein kinases required forabscisic acid regulation of seed germination, root growth, and gene expression inArabidopsis. Plant Cell 19(2):485–494.
12. Geiger D, et al. (2009) Activity of guard cell anion channel SLAC1 is controlled bydrought-stress signaling kinase-phosphatase pair. Proc Natl Acad Sci USA 106(50):21425–21430.
13. Sato A, et al. (2009) Threonine at position 306 of the KAT1 potassium channel is es-sential for channel activity and is a target site for ABA-activated SnRK2/OST1/SnRK2.6protein kinase. Biochem J 424(3):439–448.
14. Allen GJ, et al. (2001) A defined range of guard cell calcium oscillation parametersencodes stomatal movements. Nature 411(6841):1053–1057.
15. Desikan R, Griffiths R, Hancock J, Neill S (2002) A new role for an old enzyme: Nitratereductase-mediated nitric oxide generation is required for abscisic acid-induced sto-matal closure in Arabidopsis thaliana. Proc Natl Acad Sci USA 99(25):16314–16318.
16. Lee Y, et al. (1996) Abscisic acid-induced phosphoinositide turnover in guard cellprotoplasts of Vicia faba. Plant Physiol 110(3):987–996.
17. McAinsh MR, Brownlee C, Hetherington AM (1990) Abscisic acid-induced elevation ofguard cell cytosolic Ca2+ precedes stomatal closure. Nature 343:186–188.
18. Pei Z-M, et al. (2000) Calcium channels activated by hydrogen peroxide mediate ab-scisic acid signalling in guard cells. Nature 406(6797):731–734.
19. Kwak JM, et al. (2003) NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J 22(11):2623–2633.
20. Garcia-Mata C, et al. (2003) Nitric oxide regulates K+ and Cl− channels in guard cellsthrough a subset of abscisic acid-evoked signaling pathways. Proc Natl Acad Sci USA100(19):11116–11121.
21. Lozano-Juste J, León J (2010) Enhanced abscisic acid-mediated responses in nia1-nia2noa1-2 triple mutant impaired in NIA/NR- and AtNOA1-dependent nitric oxidebiosynthesis in Arabidopsis. Plant Physiol 152(2):891–903.
22. Neill SJ, Desikan R, Clarke A, Hancock JT (2002) Nitric oxide is a novel component ofabscisic acid signaling in stomatal guard cells. Plant Physiol 128(1):13–16.
23. Vahisalu T, et al. (2008) SLAC1 is required for plant guard cell S-type anion channelfunction in stomatal signalling. Nature 452(7186):487–491.
24. Joudoi T, et al. (2013) Nitrated cyclic GMP modulates guard cell signaling in Arabi-dopsis. Plant Cell 25(2):558–571.
25. Wang P, Du Y, Li Y, Ren D, Song C-P (2010) Hydrogen peroxide-mediated activation ofMAP kinase 6 modulates nitric oxide biosynthesis and signal transduction in Arabi-dopsis. Plant Cell 22(9):2981–2998.
26. MoreauM, Lee GI, Wang Y, Crane BR, Klessig DF (2008) AtNOS/AtNOA1 is a functionalArabidopsis thaliana cGTPase and not a nitric-oxide synthase. J Biol Chem 283(47):32957–32967.
27. Feng J, et al. (2013) S-nitrosylation of phosphotransfer proteins represses cytokininsignaling. Nat Commun 4:1529.
28. Yun B-W, et al. (2011) S-nitrosylation of NADPH oxidase regulates cell death in plantimmunity. Nature 478(7368):264–268.
29. Terrile MC, et al. (2012) Nitric oxide influences auxin signaling through S-nitrosylationof the Arabidopsis TRANSPORT INHIBITOR RESPONSE 1 auxin receptor. Plant J 70(3):492–500.
30. He Y, et al. (2004) Nitric oxide represses the Arabidopsis floral transition. Science305(5692):1968–1971.
31. Tada Y, et al. (2008) Plant immunity requires conformational changes [corrected] ofNPR1 via S-nitrosylation and thioredoxins. Science 321(5891):952–956.
32. Feechan A, et al. (2005) A central role for S-nitrosothiols in plant disease resistance.Proc Natl Acad Sci USA 102(22):8054–8059.
33. Lee U, Wie C, Fernandez BO, Feelisch M, Vierling E (2008) Modulation of nitrosativestress by S-nitrosoglutathione reductase is critical for thermotolerance and plantgrowth in Arabidopsis. Plant Cell 20(3):786–802.
34. Chen R, et al. (2009) The Arabidopsis PARAQUAT RESISTANT2 gene encodes anS-nitrosoglutathione reductase that is a key regulator of cell death. Cell Res 19(12):1377–1387.
35. Xu S, Guerra D, Lee U, Vierling E (2013) S-nitrosoglutathione reductases are low-copynumber, cysteine-rich proteins in plants that control multiple developmental anddefense responses in Arabidopsis. Front Plant Sci 4:430.
36. Yonashiro R, et al. (2012) Mitochondrial ubiquitin ligase MITOL blocks S-nitrosylatedMAP1B-light chain 1-mediated mitochondrial dysfunction and neuronal cell death.Proc Natl Acad Sci USA 109(7):2382–2387.
37. Ng LM, et al. (2011) Structural basis for basal activity and autoactivation of abscisicacid (ABA) signaling SnRK2 kinases. Proc Natl Acad Sci USA 108(52):21259–21264.
38. Wang Y-Q, et al. (2009) S-nitrosylation of AtSABP3 antagonizes the expression ofplant immunity. J Biol Chem 284(4):2131–2137.
39. Merlot S, Gosti F, Guerrier D, Vavasseur A, Giraudat J (2001) The ABI1 and ABI2protein phosphatases 2C act in a negative feedback regulatory loop of the abscisicacid signalling pathway. Plant J 25(3):295–303.
40. Murata Y, Pei ZM, Mori IC, Schroeder J (2001) Abscisic acid activation of plasmamembrane Ca2+ channels in guard cells requires cytosolic NAD(P)H and is differen-tially disrupted upstream and downstream of reactive oxygen species production inabi1-1 and abi2-1 protein phosphatase 2C mutants. Plant Cell 13(11):2513–2523.
41. Yoshida R, et al. (2002) ABA-activated SnRK2 protein kinase is required for de-hydration stress signaling in Arabidopsis. Plant Cell Physiol 43(12):1473–1483.
42. Hardie DG (2007) AMP-activated/SNF1 protein kinases: Conserved guardians of cel-lular energy. Nat Rev Mol Cell Biol 8(10):774–785.
43. Kishi M, Pan YA, Crump JG, Sanes JR (2005) Mammalian SAD kinases are required forneuronal polarization. Science 307(5711):929–932.
44. Zhang A, et al. (2011) Nitric oxide mediates brassinosteroid-induced ABA biosynthesisinvolved in oxidative stress tolerance in maize leaves. Plant Cell Physiol 52(1):181–192.
45. Soon F-F, et al. (2012) Molecular mimicry regulates ABA signaling by SnRK2 kinasesand PP2C phosphatases. Science 335(6064):85–88.
46. Zhang A, et al. (2007) Nitric oxide induced by hydrogen peroxide mediates abscisicacid-induced activation of the mitogen-activated protein kinase cascade involved inantioxidant defense in maize leaves. New Phytol 175(1):36–50.
47. Bright J, Desikan R, Hancock JT, Weir IS, Neill SJ (2006) ABA-induced NO generationand stomatal closure in Arabidopsis are dependent on H2O2 synthesis. Plant J 45(1):113–122.
48. Sirichandra C, et al. (2009) Phosphorylation of the Arabidopsis AtrbohF NADPH oxi-dase by OST1 protein kinase. FEBS Lett 583(18):2982–2986.
49. Peng Z-Y, Weyers JDB (1994) Stomatal sensitivity to abscisic acid following waterdeficit stress. J Exp Bot 45(6):835–845.
50. Martínez-Ruiz A, et al. (2013) Specificity in S-nitrosylation: A short-range mechanismfor NO signaling? Antioxid Redox Signal 19(11):1220–1235.
51. Harwood AJ (2001) Regulation of GSK-3: A cellular multiprocessor. Cell 105(7):821–824.
52. Kim T-W, et al. (2009) Brassinosteroid signal transduction from cell-surface receptorkinases to nuclear transcription factors. Nat Cell Biol 11(10):1254–1260.
53. Cross DAE, Alessi DR, Cohen P, Andjelkovich M, Hemmings BA (1995) Inhibition ofglycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature378(6559):785–789.
54. Reynaert NL, et al. (2004) Nitric oxide represses inhibitory kappaB kinase throughS-nitrosylation. Proc Natl Acad Sci USA 101(24):8945–8950.
55. Park H-S, Huh S-H, Kim M-S, Lee SH, Choi E-J (2000) Nitric oxide negatively regulatesc-Jun N-terminal kinase/stress-activated protein kinase by means of S-nitrosylation.Proc Natl Acad Sci USA 97(26):14382–14387.
56. Whalen EJ, et al. (2007) Regulation of β-adrenergic receptor signaling by S-nitrosylationof G-protein-coupled receptor kinase 2. Cell 129(3):511–522.
618 | www.pnas.org/cgi/doi/10.1073/pnas.1423481112 Wang et al.