p-selectin glycoprotein ligand-1 negatively regulates … · p-selectin glycoprotein ligand-1...
TRANSCRIPT

of July 1, 2018.This information is current as
Regulates T-Cell Immune ResponsesP-Selectin Glycoprotein Ligand-1 Negatively
HirataMasanori Matsumoto, Masayuki Miyasaka and Takako
http://www.jimmunol.org/content/183/11/7204doi: 10.4049/jimmunol.0902173November 2009;
2009; 183:7204-7211; Prepublished online 4J Immunol
MaterialSupplementary
3.DC1http://www.jimmunol.org/content/suppl/2009/11/04/jimmunol.090217
Referenceshttp://www.jimmunol.org/content/183/11/7204.full#ref-list-1
, 19 of which you can access for free at: cites 38 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 1, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

P-Selectin Glycoprotein Ligand-1 Negatively Regulates T-CellImmune Responses1
Masanori Matsumoto,* Masayuki Miyasaka,* and Takako Hirata2*†
Cell surface sialomucins often act as antiadhesive molecules by virtue of their extended structure and negative charge. CD43 is onesuch sialomucin, expressed on most leukocytes. P-selectin glycoprotein ligand-1 (PSGL-1) is another sialomucin expressed byleukocytes. It serves as a major selectin ligand, but no antiadhesive role for it has been described. In this study, we showed thatPSGL-1-deficient T cells, like CD43-deficient T cells, exhibited increased adhesion and proliferation compared with wild-type cells.The loss of both PSGL-1 and CD43 led to a further increase in T cell adhesion and proliferation. The reexpression of full-lengthPSGL-1 or CD43 in double-deficient CD4� T cells reversed their increased adhesion and proliferation phenotype. Using chimericconstructs of human CD8 and either PSGL-1 or CD43, we demonstrated that the intracellular domain of PSGL-1 or CD43 isrequired for suppressing proliferation but not adhesion. Furthermore, in a mouse model of inflammatory bowel disease inducedby the adoptive transfer of naive T cells into RAG-deficient hosts, a PSGL-1 deficiency exacerbated the development of inflam-mation. These results reveal a novel regulatory role for PSGL-1 in T cell adhesion and proliferation and suggest that PSGL-1negatively regulates T cell immune responses in vivo. The Journal of Immunology, 2009, 183: 7204–7211.
C ell surface sialomucins represent a family of glycopro-teins that are highly glycosylated with sialylated O-linkedglycans. Because of their extended structure and the neg-
ative charge imparted by their sialic acid residues, some sialomu-cins can provide a repulsive barrier around a cell that inhibits cell-cell interactions. CD43 is one such molecule (1). It is expressed bymost hematopoietic cells, including T cells. The extracellular do-main of both human and mouse CD43 contains more than 80serine or threonine residues, most of which are glycosylated byO-linked glycans that are heavily sialylated; this domain has arod-like structure that is predicted to extend 45 nm from the lipidbilayer (2). Supporting evidence for its antiadhesive role is ob-served in vitro by the enhanced homotypic adhesion and prolifer-ation of CD43-deficient (CD43�/�) T cells (3), and in vivo by theirincreased migration into secondary lymphoid organs (4). CD43�/�
neutrophils also show enhanced rolling and adhesion both in vitroand in vivo (5, 6). These antiadhesive functions, however, are notevident when CD43 interacts with its specific ligands (7). Severalpotential ligands for CD43 have been reported, including ICAM-1(8), galectin-1 (9), and sialoadhesin (10). In particular, we andothers reported that CD43 on mouse Th1 cells and human T lym-phoblasts is modified with sialylated and fucosylated carbohydratestructures to interact with E-selectin (11, 12). This interaction
plays an important role in mediating the migration of activated Tcells to inflamed skin (13, 14).
P-selectin glycoprotein ligand-1 (PSGL-1;3 CD162) (3) is an-other sialomucin expressed by most leukocytes (15, 16). The ex-tracellular domain of both human and mouse PSGL-1 contains�40 potential O-glycosylation sites. Like CD43, PSGL-1 is ahighly extended molecule: its ectodomain is estimated to be 50-nmlong (17). PSGL-1 was originally identified as the major ligand forP-selectin, and subsequent studies showed that it also binds theother two members of the selectin family, E- and L-selectin. Stud-ies using PSGL-1-deficient (PSGL-1�/�) mice showed that, onactivated T cells, PSGL-1 serves as the major ligand for P-selectinas well as being an E-selectin ligand (18, 19). Similarly, PSGL-1on neutrophils binds P- and E-selectin to mediate their rolling onendothelial cells and migration into inflamed tissues (20, 21).These well-recognized functions of PSGL-1 are critically depen-dent on its glycosylation, which endows it with the ability to bindselectins. Thus, although PSGL-1 is abundantly expressed on mosthematopoietic cells, its function in cells that are not equipped withthe machinery to modify it to bind selectins, such as naive T cells,remains unknown.
To explore the function of PSGL-1 independent of its role as aselectin ligand, we exploited our previous findings that the relatedsialomucin CD43 has dual functions: it is usually antiadhesive, butwhen appropriately glycosylated in certain cell types, it can be-come a proadhesive selectin ligand (5, 6). We investigated whetherthe PSGL-1 expressed on T cells has antiadhesive or regulatoryroles, using T cells isolated from wild-type (WT), PSGL-1�/�,CD43�/�, and PSGL-1/CD43 double-knockout (DKO) mice. Weshow that PSGL-1�/� T cells exhibited enhanced adhesion andproliferation. Loss of both PSGL-1 and CD43 resulted in a furtherincrease in T cell adhesion and proliferation. The increased adhe-sion and proliferation of the DKO T cells were reversed by thereintroduction of PSGL-1 or CD43. The intracellular domain of
*Laboratory of Immunodynamics, Department of Microbiology and Immunology,Graduate School of Medicine and World Premier International Immunology FrontierResearch Center, Osaka University, Suita, Osaka, Japan; and †Center for Innovationin Immunoregulative Technology and Therapeutics, Graduate School of Medicine,Kyoto University, Kyoto, Japan
Received for publication July 6, 2009. Accepted for publication September 29, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by a Grant-in-Aid from the Ministry of Education, Culture,Sports, Science and Technology, Japan; a Grant-in-Aid for Scientific Research fromthe Japan Society for the Promotion of Science; and a Grant of Long-Range ResearchInitiative (LRI) by the Japan Chemical Industry Association (JCIA).2 Address correspondence and reprint requests to Dr. Takako Hirata, Center for In-novation in Immunoregulative Technology and Therapeutics, Graduate School ofMedicine, Kyoto University, Yoshida-Konoe-cho, Sakyo-ku, Kyoto, Japan. E-mailaddress: [email protected]
3 Abbreviations used in this paper: PSGL-1, P-selectin glycoprotein ligand-1; WT,wild type; DKO, PSGL-1 and CD43 double-knockout; FL, full length.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0902173
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

PSGL-1 or CD43 was required for the reversal of the increasedproliferation, but not for reducing the adhesion. Moreover, in vivo,the PSGL-1 deficiency exacerbated the development of T cell-mediated colitis. Thus, in addition to its well-characterized role asa selectin ligand, these results point to a novel role for PSGL-1 innegatively regulating T cell adhesion and proliferation and in con-trolling T cell-mediated immune responses in vivo.
Materials and MethodsMice
C57BL/6 (B6) mice were purchased from CLEA Japan. PSGL-1�/� miceon a B6 background were provided by B. Furie (Harvard Medical School,Boston, MA). RAG-1-deficient (RAG-1�/�) mice on a B6 background andCD43�/� mice on a B6 � 129S4/SvJae background were purchased fromThe Jackson Laboratory. CD43�/� mice were back-crossed more than sixgenerations onto the B6 genetic background. DKO mice were generated bybreeding PSGL-1�/� mice with CD43�/� mice. All mice were used when6 to 10 wk old. The mice were housed at the Institute of ExperimentalAnimal Sciences at Osaka University Medical School. All studies and pro-cedures were approved by the Ethics Review Committee for Animal Ex-perimentation of the Osaka University Graduate School of Medicine.
Isolation of T cells
Total T cells and CD4� T cells were isolated from the spleen and theperipheral and mesenteric lymph nodes using a BD IMag Mouse T Lym-phocyte Enrichment Set and CD4 T Lymphocyte Enrichment Set (bothfrom BD Biosciences), respectively. Isolated CD4� T cells were stainedwith anti-CD45RB-FITC (C363-16A; BioLegend), anti-CD25-PE (PC61;BD Biosciences), and anti-CD4-allophycocyanin (RM4-5; BD Biosciences),and naive CD4� T cells (CD4�CD25�CD45RBhigh) were sorted byFACSAria (BD Biosciences).
Construction of retroviral vectors
Full-length mouse PSGL-1 (PSGL-1FL) cDNA was inserted into thepMXs-IG vector (22), provided by T. Kitamura (University of Tokyo, To-kyo, Japan), at the BamHI and XhoI sites. The viral long terminal repeatsof this vector drive the expression of PSGL-1 and enhanced GFP via abicistronic mRNA containing an internal ribosomal entry site element.cDNAs for full-length mouse CD43 (CD43FL) and full-length human CD8(CD8FL) were amplified from mouse Th1-cell cDNA and human thymuscDNA, respectively. To produce chimeric constructs in which the extra-cellular and transmembrane domain of mouse PSGL-1 or CD43 was fusedto the intracellular domain of human CD8 (PSGL-1/CD8 or CD43/CD8),or the extracellular and transmembrane domain of human CD8 was fusedto the intracellular domain of mouse PSGL-1 or CD43 (CD8/PSGL-1 orCD8/CD43), the corresponding regions were amplified and ligated into thepMXs-IG vector.
Retroviral infections
The improved retrovirus packaging cells, PLAT-E (23), were transfectedby calcium phosphate precipitation with 20 �g of the pMXs-IG vectorcontaining one of the constructs described above. At 12 h post transfection,the calcium phosphate-containing medium was replaced, and the cells werecultured for an additional 48 h. The virus-containing supernatant from thePLAT-E cells was harvested, added to a six-well plate coated with 20�g/ml recombinant fibronectin fragment CH-296 (RetroNectin; Takara),and then centrifuged at 1,500 � g for 2 h. To infect T cells, purified CD4�
T cells from WT or DKO mice were stimulated with plate-immobilizedanti-CD3� (10 �g/ml; 145-2C11; BD Biosciences) plus anti-CD28 (10�g/ml; 37.51; BD Biosciences) for 48 h and added to the retrovirus-coatedplate in the presence of 10 ng/ml IL-2 (R&D Systems). After 48 h ofinfection, the cells were washed, transferred to uncoated dishes, and cul-tured for an additional 3 days.
Homotypic aggregation assays
Isolated T cells (1 � 105 cells), naive CD4� T cells (1 � 105 cells), orretrovirus-transduced CD4� T cells (1 � 104 cells) were added to 96-wellculture plates and incubated with PMA (100 ng/ml) for 5 h in the presenceof calcium or EDTA. In some experiments, T cells from DKO mice wereadded to the wells and incubated with PMA in the presence of 10 �g/mlanti-CD11a (M17/4; BD Biosciences), anti-ICAM-1 (KAT-1; provided byK. Kato, Juntendo University, Tokyo, Japan), anti-CD18 (GAME-46; BDBiosciences), anti-CD11b (M1/70; BD Biosciences), or isotype controls.
To examine the effect of neuraminidase treatment, the isolated T cellsfrom WT, PSGL-1�/�, CD43�/�, and DKO mice were incubated withor without neuraminidase from Clostridium perfringens (25 mU/ml;Sigma-Aldrich) for 1 h and cultured with PMA for 5 h. The number ofunaggregated cells was determined from the samples before they weredissociated. To determine the total number of cells in each sample, theaggregates were dissociated to the single-cell level by the addition ofEDTA. The percentage of cells that had aggregated (percent aggrega-tion) was calculated as the number of cells in aggregates (total numberof cells minus the number of single cells) over the total number of cells.
Cell adhesion assays
Isolated T cells or naive CD4� T cells (1 � 105 cells) were stimulated withPMA (100 ng/ml), and then added to 96-well culture plates coated with 10�g/ml fibronectin (Sigma-Aldrich) in the presence of calcium or EDTA. Inblocking experiments, PMA-stimulated T cells from DKO mice wereadded to the wells in the presence of anti-CD49d (R1-2; BD Biosciences),anti-CD29 (Ha2/5; BD Biosciences), anti-�7 integrin (FIB27; BD Bio-sciences), or isotype controls. In some experiments, the isolated T cellswere incubated with or without neuraminidase, stimulated with PMA, andadded to 96-well plates coated with fibronectin. The plates were incubatedfor 20 min, unbound cells were removed, and the number of bound cellswas determined by photographing the cells and counting them.
Proliferation assays
Isolated T cells or naive CD4� T cells (1 � 105 cells) were labeled with3.3 �M CFSE for 10 min at 37°C and stimulated with plate-immobilizedanti-CD3� (1 �g/ml) and anti-CD28 (1 �g/ml) for 48 h. The cells werestained with anti-CD4-PE (RM4-5; BD Biosciences) and anti-CD8-APC(53.6.7; BD Biosciences) and then analyzed on a FACSCalibur. Daughtergenerations of CD4� and CD8� T cells were determined by measuring theCFSE dilution. In some experiments, retrovirus-transduced CD4� T cells(1 � 104 cells) were stimulated with plate-immobilized anti-CD3� (1 �g/ml) and anti-CD28 (1 �g/ml) in the presence of 1 �Ci/well of [3H]thymi-dine (GE Healthcare) for 24 h. Cells were harvested onto a UniFilter-96GF/C (PerkinElmer) using a UniFilter-96 Harvester (PerkinElmer), and theincorporated radioactivity was measured using a TopCount NXT scintil-lation counter (PerkinElmer).
Mouse colitis model
We adapted a mouse model for colitis in which colitis is induced in im-munodeficient mice by the transfer of naive T cells (24, 25). Naive CD4�
T cells (5 � 105) sorted from WT, PSGL-1�/�, CD43�/�, and DKO micewere injected via the tail vein into RAG-1�/� mice. The body weight ofthe recipient mice was monitored throughout the experiment. The micewere analyzed 6 wk after the T cell transfer. They were assigned a diseaseactivity score that was the sum of four parameters: hunching and wastingwere scored 0 or 1, colon thickening 0–3, and stool consistency 0–3, asdescribed (26). Tissue samples from the middle colon of untreated or T cellrecipient RAG-1�/� mice were prepared for histological staining withH&E. Individual sections were scored blind, according to the followingindependent criteria: cell infiltration 0–3, crypt elongation 0–3, and num-ber of crypt abscesses 0–3 (26).
Isolation of cells infiltrating the large intestine lamina propria
Cells in the large intestine lamina propria were prepared by digesting theintestinal tissue fragments depleted of intraepithelial lymphocytes with 400U/ml collagenase D (Roche) and 10 �g/ml DNase I (Roche) for 30 min at37°C. Lymphocytes were further enriched by centrifugation on 40/75%Percoll (GE Healthcare). Total cell numbers were determined with a he-mocytometer. The percentage of CD4� T cells was determined by stainingthe cells with anti-CD4-APC and analyzing them on a FACSCalibur (BDBiosciences).
Statistical analysis
Data are presented as the mean � SEM. Statistical analyses were per-formed using the 2-tailed unpaired Student’s t test.
ResultsT cells deficient in PSGL-1 and/or CD43 show an increase inhomotypic aggregation and adhesion
To examine the role of PSGL-1 as an antiadhesive molecule on Tcells, we first tested T cells isolated from WT, PSGL-1�/�,CD43�/�, and DKO mice for homotypic aggregation upon PMA
7205The Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

stimulation. In accordance with a previous report (3), CD43�/� Tcells showed an increase in homotypic aggregation compared withWT cells (Fig. 1A). The percentage of aggregated PSGL-1�/� Tcells was also increased, and the loss of both PSGL-1 and CD43led to a further increase in aggregation (Fig. 1A). The addition ofEDTA completely abolished the aggregation of cells of all geno-types, confirming that the aggregation was calcium dependent(data not shown). The percentage of CD4� and CD8� T cells orthose with a naive or memory phenotype was not significantlydifferent among the four genotypes (data not shown). Similar re-sults were obtained when naive CD4� T cells from the four ge-notypes were tested for homotypic aggregation (supplementaryFig. S1A).4 To determine which adhesion molecules mediate theaggregation, we examined the effect of Ab treatment on the ho-motypic aggregation of PMA-stimulated DKO T cells. The percentaggregation was significantly reduced in the presence of an anti-�L
(CD11a), anti-ICAM-1, or anti-�2 (CD18) mAb (Fig. 1B), but notin the presence of an anti-�M (CD11b) mAb (data not shown),indicating that the aggregation was mediated by the interactionbetween LFA-1 and ICAM-1. Thus, PSGL-1 and CD43 both pre-vented the homotypic aggregation of PMA-stimulated T cells,which was mediated by LFA-1 and ICAM-1.
To determine whether PSGL-1 and CD43 regulate integrin-me-diated adhesive processes in general, we tested the adhesion ofPMA-stimulated T cells to fibronectin immobilized on plasticplates. Consistent with the results of the homotypic aggregation
assays (Fig. 1A), the adhesion of PMA-stimulated PSGL-1�/� Tcells to fibronectin was greater than that of WT cells (Fig. 1C). Theadhesion of DKO T cells was further increased compared withcells deficient in either PSGL-1 or CD43 (Fig. 1C). We confirmedthat the deficiency of PSGL-1 and/or CD43 also increased thebinding of PMA-stimulated naive CD4� T cells to fibronectin(supplementary Fig. S1B). The adhesion was largely mediated by�4�1 integrin, because it was blocked by treatment of the cells withan anti-�4 (CD49d) or anti-�1 (CD29) mAb (Fig. 1D).
To examine whether the deficiency of PSGL-1 and/or CD43affects the expression of other adhesion molecules, the cell surfaceexpression of PSGL-1, CD43, and integrin chains on T cells wasmeasured by flow cytometry. The expression level of PSGL-1 onCD43�/� cells and of CD43 on PSGL-1�/� cells was comparableto that on WT cells (supplementary Fig. S2). The expression ofintegrin chains �L, �4, �1, �2, and �7, as well as ICAM-1, was alsocomparable among the four genotypes (supplementary Fig. S2 anddata not shown). These results suggest that the increased aggrega-tion and adhesion of T cells deficient in PSGL-1 and/or CD43 isnot due to alterations in the expression of other adhesion mole-cules, but rather is a direct effect of their deficiency.
T cells deficient in PSGL-1 and/or CD43 show enhancedproliferative responses
We next investigated the proliferative responses of T cells from WT,PSGL-1�/�, CD43�/�, and DKO mice. The isolated T cells werelabeled with CFSE and stimulated with anti-CD3� plus anti-CD28 for2 days. The proliferative responses of both the CD4� and CD8� Tcells were determined by measuring the dilution of the CFSE. Inagreement with published reports (3, 27), both the CD4� (Fig. 2A)and the CD8� (Fig. 2B) T cells from CD43�/� mice exhibited in-creased proliferative responses as did the CD4� and CD8� T cellsfrom the PSGL-1�/� mice (Fig. 2). The DKO T cells showed anadditional enhancement in proliferation (Fig. 2). Similar results wereobtained with naive CD4� T cells (supplementary Fig. S3). Thesedata suggest that both PSGL-1 and CD43 function to regulate theproliferative responses of T cells.
The extracellular/transmembrane domain of PSGL-1 or CD43 isresponsible for the reversal of increased T cell aggregation
To verify that the deficiency of PSGL-1 and/or CD43 was respon-sible for the increased adhesion and proliferation, we reintroducedeither PSGL-1 or CD43 into DKO T cells using a retroviral genetransfer system. DKO CD4� T cells were infected with a GFP-containing control retrovirus vector (pMXs-IG) or with vectorsencoding full-length PSGL-1 (PSGL-1FL) or CD43 (CD43FL).Transduced cells were sorted based on their GFP expression level,4 The online version of this article contains supplementary material.
FIGURE 1. Loss of PSGL-1 and CD43 increases T cell adhesion. A,Homotypic aggregation of PMA-stimulated T cells. T cells isolated fromWT, PSGL-1�/�, CD43�/�, and DKO mice were incubated with PMA(100 ng/ml) for 5 h. The percentage of cells within aggregates was deter-mined. B, Effects of Ab treatment on the homotypic aggregation of PMA-stimulated T cells. T cells from DKO mice were incubated with PMA for5 h in the presence of anti-CD11a, anti-ICAM-1, anti-CD18, or isotypecontrols. C, Adhesion of PMA-stimulated T cells to fibronectin. The iso-lated T cells were stimulated with PMA (100 ng/ml), added to 96-wellplates coated with fibronectin, and incubated for 20 min. Unbound cellswere removed, and the number of bound cells was determined. D, Effectsof Ab treatment on T cell adhesion to fibronectin. PMA-stimulated T cellsfrom DKO mice were added to fibronectin-coated plates and incubated inthe presence of anti-CD49d, anti-CD29, anti-�7 integrin, or isotype con-trols. Results represent one of three similar experiments. Data are presentedas means � SEM. �, p � 0.05; ��, p � 0.01; ���, p � 0.001.
FIGURE 2. Loss of PSGL-1 and CD43 increases T cell proliferation.The isolated T cells from WT, PSGL-1�/�, CD43�/�, and DKO mice werelabeled with CFSE and stimulated with plate-immobilized anti-CD3� andanti-CD28 for 48 h. Daughter generations of CD4� (A) and CD8� (B) Tcells were determined by measuring the dilution of CFSE. Results repre-sent one of three similar experiments. Data are presented as means � SEM.��, p � 0.01 vs WT.
7206 ROLE OF PSGL-1 IN T-CELL IMMUNE RESPONSES
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
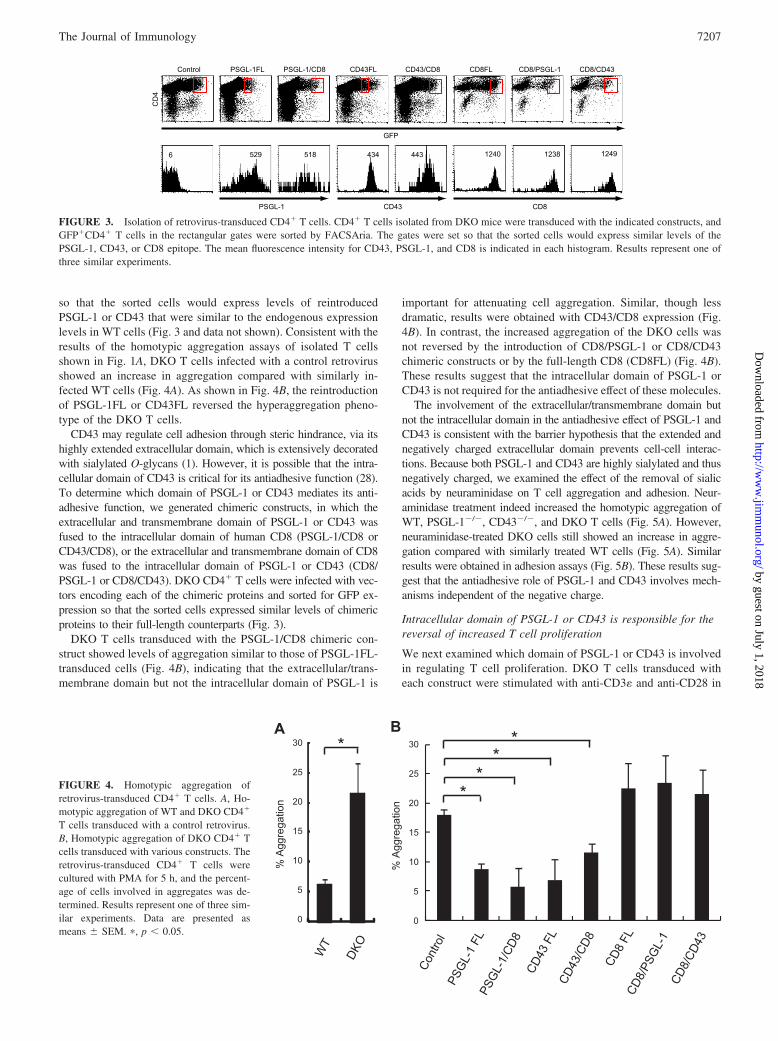
so that the sorted cells would express levels of reintroducedPSGL-1 or CD43 that were similar to the endogenous expressionlevels in WT cells (Fig. 3 and data not shown). Consistent with theresults of the homotypic aggregation assays of isolated T cellsshown in Fig. 1A, DKO T cells infected with a control retrovirusshowed an increase in aggregation compared with similarly in-fected WT cells (Fig. 4A). As shown in Fig. 4B, the reintroductionof PSGL-1FL or CD43FL reversed the hyperaggregation pheno-type of the DKO T cells.
CD43 may regulate cell adhesion through steric hindrance, via itshighly extended extracellular domain, which is extensively decoratedwith sialylated O-glycans (1). However, it is possible that the intra-cellular domain of CD43 is critical for its antiadhesive function (28).To determine which domain of PSGL-1 or CD43 mediates its anti-adhesive function, we generated chimeric constructs, in which theextracellular and transmembrane domain of PSGL-1 or CD43 wasfused to the intracellular domain of human CD8 (PSGL-1/CD8 orCD43/CD8), or the extracellular and transmembrane domain of CD8was fused to the intracellular domain of PSGL-1 or CD43 (CD8/PSGL-1 or CD8/CD43). DKO CD4� T cells were infected with vec-tors encoding each of the chimeric proteins and sorted for GFP ex-pression so that the sorted cells expressed similar levels of chimericproteins to their full-length counterparts (Fig. 3).
DKO T cells transduced with the PSGL-1/CD8 chimeric con-struct showed levels of aggregation similar to those of PSGL-1FL-transduced cells (Fig. 4B), indicating that the extracellular/trans-membrane domain but not the intracellular domain of PSGL-1 is
important for attenuating cell aggregation. Similar, though lessdramatic, results were obtained with CD43/CD8 expression (Fig.4B). In contrast, the increased aggregation of the DKO cells wasnot reversed by the introduction of CD8/PSGL-1 or CD8/CD43chimeric constructs or by the full-length CD8 (CD8FL) (Fig. 4B).These results suggest that the intracellular domain of PSGL-1 orCD43 is not required for the antiadhesive effect of these molecules.
The involvement of the extracellular/transmembrane domain butnot the intracellular domain in the antiadhesive effect of PSGL-1 andCD43 is consistent with the barrier hypothesis that the extended andnegatively charged extracellular domain prevents cell-cell interac-tions. Because both PSGL-1 and CD43 are highly sialylated and thusnegatively charged, we examined the effect of the removal of sialicacids by neuraminidase on T cell aggregation and adhesion. Neur-aminidase treatment indeed increased the homotypic aggregation ofWT, PSGL-1�/�, CD43�/�, and DKO T cells (Fig. 5A). However,neuraminidase-treated DKO cells still showed an increase in aggre-gation compared with similarly treated WT cells (Fig. 5A). Similarresults were obtained in adhesion assays (Fig. 5B). These results sug-gest that the antiadhesive role of PSGL-1 and CD43 involves mech-anisms independent of the negative charge.
Intracellular domain of PSGL-1 or CD43 is responsible for thereversal of increased T cell proliferation
We next examined which domain of PSGL-1 or CD43 is involvedin regulating T cell proliferation. DKO T cells transduced witheach construct were stimulated with anti-CD3� and anti-CD28 in
FIGURE 3. Isolation of retrovirus-transduced CD4� T cells. CD4� T cells isolated from DKO mice were transduced with the indicated constructs, andGFP�CD4� T cells in the rectangular gates were sorted by FACSAria. The gates were set so that the sorted cells would express similar levels of thePSGL-1, CD43, or CD8 epitope. The mean fluorescence intensity for CD43, PSGL-1, and CD8 is indicated in each histogram. Results represent one ofthree similar experiments.
FIGURE 4. Homotypic aggregation ofretrovirus-transduced CD4� T cells. A, Ho-motypic aggregation of WT and DKO CD4�
T cells transduced with a control retrovirus.B, Homotypic aggregation of DKO CD4� Tcells transduced with various constructs. Theretrovirus-transduced CD4� T cells werecultured with PMA for 5 h, and the percent-age of cells involved in aggregates was de-termined. Results represent one of three sim-ilar experiments. Data are presented asmeans � SEM. �, p � 0.05.
7207The Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

the presence of [3H]thymidine for 24 h, and proliferative responseswere determined by [3H]thymidine incorporation. The increasedproliferation of DKO T cells was reversed by the reexpression of
either PSGL-1FL or CD43FL (Fig. 6), indicating that this hyper-proliferative phenotype was indeed due to the loss of PSGL-1and/or CD43. In contrast, expression of PSGL-1/CD8 or CD43/CD8 had little effect (Fig. 6). These results suggest that the extra-cellular/transmembrane domain of PSGL-1 or CD43 does not playa major role in regulating T cell proliferation, and suggest that theintracellular domain of PSGL-1 or CD43 is involved in this pro-cess. Indeed, the introduction of CD8/PSGL-1 or CD8/CD43 de-creased the proliferative response compared with CD8FL (Fig. 6).These results suggest that the intracellular domain of PSGL-1 andCD43 is required for the negative regulation of T cell proliferation.
PSGL-1 and CD43 regulate the development of CD4�
T cell-mediated colitis in vivo
The above results suggested that PSGL-1 and CD43 both regulateT cell adhesion and proliferation. To determine whether they reg-ulate T cell-mediated immune responses, which require optimal Tcell adhesion and proliferation in vivo, we used a mouse model ofinflammatory bowel disease, which is caused by adoptively trans-ferring naive T cells into SCID or RAG-1�/� recipient mice (24,25). For our experiments, we used CD4�CD25�CD45RBhigh Tcells from WT, PSGL-1�/�, CD43�/�, and DKO mice for theadoptive transfer into RAG-1�/� recipient mice. The recipientmice were analyzed 6 wk after cell transfer. RAG-1�/� mice thatreceived WT cells developed colitis, with clinical signs such asdiarrhea and wasting. RAG-1�/� mice that received PSGL-1�/�,CD43�/�, or DKO cells also developed colitis. The body weightof the RAG-1�/� mice with colitis at 6 wk after cell transfer wasreduced compared with untreated RAG-1�/� mice, but was com-parable regardless of the T cell genotype that was transferred (datanot shown). To quantify the severity of the colitis, we determinedthe disease activity as the sum of four parameters: hunching, wast-ing, colon thickening, and stool consistency. The disease activityscore of the mice that received WT cells was 3.6 (n � 8), whereasthat of the mice receiving PSGL-1�/�, CD43�/�, or DKO cellswas 5.6 (n � 7), 5.0 (n � 10), or 5.7 (n � 10), respectively (Fig.7A). Thus, the single and double knockout phenotypes all exacer-bated the induced colitis.
We also performed histological analyses of colon cross-sec-tions from untreated or RAG-1�/� mice or RAG-1�/� mice that
FIGURE 5. Effect of neuraminidase treatment on T cell aggregation andadhesion. A, Homotypic aggregation of PMA-stimulated T cells treatedwith neuraminidase. The isolated T cells from WT, PSGL-1�/�, CD43�/�,and DKO mice were incubated with or without neuraminidase for 1 h andcultured with PMA for 5 h. The percentage of cells involved in aggregateswas determined. B, Adhesion of PMA-stimulated T cells treated with neur-aminidase to fibronectin. The isolated T cells were incubated with or with-out neuraminidase, stimulated with PMA, added to 96-well plates coatedwith fibronectin, and incubated for 20 min. Unbound cells were removed,and the number of bound cells was determined. �, p � 0.05.
FIGURE 6. Proliferative responses of ret-rovirus-transduced DKO CD4� T cells. Theretrovirus-transduced DKO CD4� T cellswere stimulated with plate-immobilized anti-CD3� and anti-CD28 in the presence of[3H]thymidine for 24 h, and proliferative re-sponses were determined by [3H]thymidineincorporation. Results represent one of threesimilar experiments. Data are presented asmeans � SEM. �, p � 0.05; ��, p � 0.01.
7208 ROLE OF PSGL-1 IN T-CELL IMMUNE RESPONSES
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

received the transferred naive CD4� T cells as described above.Six weeks after the transfer, typical pathologic changes of colitis,such as cell infiltration, crypt elongation, and crypt abscesses, wereobserved in mice that received the naive CD4� T cells (Fig. 7B).For the histological colitis score, individual sections were scoredfor cell infiltration, crypt elongation, and the number of crypt ab-scesses. The mice that received PSGL-1�/�, CD43�/�, or DKOcells showed a higher colitis score than those that received WTcells (Fig. 7C).
Because marked cell infiltration was observed in the colon of theT cell recipient mice, we examined the number of CD4� T cells
in the colon of the mice that received the WT, PSGL-1�/�,CD43�/�, or DKO cells. In the colon of the mice that receivedDKO cells, the number of infiltrated CD4� T cells was �2-foldgreater than in the mice that received WT cells (Fig. 7D). Theseresults suggest that PSGL-1 and CD43 both negatively regulate thedevelopment of CD4� T cell-mediated colitis in vivo.
DiscussionAlthough it is well established that PSGL-1 serves as a selectinligand to mediate the migration of T cells and neutrophils intoinflamed sites, none of its other roles are well understood. In this
FIGURE 7. Loss of PSGL-1 exacer-bates T cell-mediated colitis. NaiveCD4� T cells were isolated from WT,PSGL-1�/�, CD43�/�, and DKO miceby FACSAria, and transferred intoRAG-1�/� mice. The recipient mice de-veloped colitis and were analyzed at 6wk after T cell transfer. A, Disease ac-tivity score. The mice were scored fordisease activity as described in Materi-als and Methods. B, Histological analy-sis of colon cross-sections. Tissue sam-ples from the middle colon wereprepared for histological staining withH&E. Crypt abscesses are indicated byarrowheads. C, Histological colitisscore. Individual sections were scoredblind as described in Materials andMethods. D, The number of CD4� Tcells in the large intestine lamina pro-pria. Cells were isolated from the largeintestine lamina propria, enriched forleukocytes by percoll, and stained withanti-CD4-PE. The numbers of CD4� Tcells were determined. Values aremeans � SEM for seven to ten mice ineach experimental group. �, p � 0.05;��, p � 0.01.
7209The Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

study, we provide evidence that PSGL-1 is a negative regulator ofT cell adhesion and proliferation in vitro and of T cell immuneresponses in vivo.
Sialomucins often act as antiadhesive molecules and the sialo-mucin CD43 has both antiadhesive and proadhesive functions (7).Previous studies demonstrated that the targeted disruption of CD43in mice leads to enhanced T cell adhesion (3). We speculated thatPSGL-1, a sialomucin that shares many features with CD43, suchas an extended extracellular structure, abundant expression on Tcells, and microvillar localization, might also function as an anti-adhesive molecule. Our data showed that a deficiency of PSGL-1enhanced T cell adhesion and that the loss of both PSGL-1 andCD43 had an additive effect on T cell adhesion. Retroviral genetransfer experiments suggested that the antiadhesive function ofPSGL-1 and CD43 was mediated by their extracellular/transmem-brane domain. A similar requirement for the extracellular/trans-membrane domain was reported for the antiadhesive function ofCD34 on mast cells (29). These results are consistent with thebarrier hypothesis that the highly extended and negatively chargedextracellular domain of sialomucins provides a physical and elec-trostatic barrier around a cell. The observation that a PSGL-1and/or CD43 deficiency enhanced T cell adhesion even after re-moval of sialic acids suggests that the antiadhesive role of theirextracellular/transmembrane domain involves mechanisms inde-pendent of the electrostatic barrier. In contrast, Walker et al. (28)previously reported that the expression of a mutant CD43 thatlacks the intracellular domain in a CD43�/� T cell line did notreverse its hyperaggregation phenotype. The reason for this incon-sistency is unknown, although it is possible that the lack of theintracellular domain may have affected the configuration of theextracellular domain in the cell line used in their study.
Our study also demonstrated that a deficiency of PSGL-1 enhancesthe proliferative responses of T cells. Consistent with previous studies(3, 27), we also demonstrated that CD43�/� T cells show an increasein proliferative responses, and the loss of both PSGL-1 and CD43additively increased proliferation. These results are in contrast to thereport by Carlow et al. (30), which showed that a CD43 deficiencyhad no effect on T cell responsiveness. In fact, we did not observe theenhanced proliferation of CD43�/� T cells when the cells were stim-ulated with higher or lower concentrations of anti-CD3� plus anti-CD28 (data not shown). In addition, when the proliferative responsewas assessed by [3H]thymidine incorporation, the role of PSGL-1 andCD43 as antiproliferative molecules was most prominent within 1 dayafter stimulation (data not shown), suggesting that PSGL-1 and CD43regulate T cell proliferation during the early phase.
We also showed that the intracellular domain of PSGL-1 andCD43 is required for their antiproliferative effect. In support of thisfinding, two groups have published data demonstrating that aCD43 mutant lacking the intracellular domain is ineffective in re-verting the hyperproliferative phenotype of CD43�/� T cells (28,31). Although the intracellular domains of PSGL-1 and CD43 haveno obvious homology, this domain is known to be involved insignal transduction. In addition, the intracellular domain interactswith the adapter proteins ezrin and moesin (32, 33), which functiontogether to regulate T cell activation (34). The cytoplasmic domainof CD43 is also phosphorylated upon PMA stimulation (35). How-ever, it remains unknown whether PSGL-1 and CD43 under phys-iological conditions bind to yet-undefined receptors to deliver aninhibitory signal for proliferation. Additional experiments will berequired to clarify the mechanism of the antiproliferative effectmediated by the intracellular domains of PSGL-1 and CD43.
Besides PSGL-1 and CD43, several sialomucins, includingCD34, podocalyxin, and endomucin, which can serve as L-selectinligands when modified with selectin-binding glycans, are reported
to have similar dual functions (36–38). We propose that thesesialomucins generally function as antiadhesive molecules to pre-vent inappropriate cell adhesion and activation, but when they aremodified to bind selectins in specific cells or tissues, they switchroles and become proadhesive, promoting cell-cell interactions.
We found that, in vivo, a deficiency of PSGL-1 as well as CD43exacerbated the development of inflammation in a mouse colitismodel induced by the adoptive transfer of naive CD4� T cells intoRAG�/� hosts. This colitis model is reported to be mediated byTh1 cells that migrate into the colon in a selectin-independentmanner (24). Importantly, we showed that the numbers of CD4� Tcells in the colon of DKO cell-recipient mice were significantlyhigher than in mice that received WT or singly deficient cells.These results are consistent with a role for PSGL-1 and CD43 innegatively regulating T cell adhesion and proliferation.
In conclusion, our study shows for the first time that PSGL-1functions as a negative regulator of T cell adhesion and prolifer-ation, adding this molecule to the expanding category of dual-functioning sialomucins. Together with CD43, PSGL-1 constitutesan additional layer of regulation for T cell adhesion and prolifer-ation, and possibly for T cell-mediated immune responses.
AcknowledgmentsWe thank Dr. Bruce Furie for the PSGL-1-deficient mice, Dr. KazunoriKato for the anti-ICAM-1 mAb, Dr. Toshio Kitamura for the pMXs-IGvector and PLAT-E cells, and Yuko Furukawa and Norie Yoshizumi fortechnical assistance. We also thank Dr. Takeshi Watanabe for valuablecomments.
DisclosuresThe authors have no financial conflict of interest.
References1. Fukuda, M., and S. Tsuboi. 1999. Mucin-type O-glycans and leukosialin. Bio-
chim. Biophys. Acta 1455: 205–217.2. Cyster, J. G., D. M. Shotton, and A. F. Williams. 1991. The dimensions of the T
lymphocyte glycoprotein leukosialin and identification of linear protein epitopesthat can be modified by glycosylation. EMBO J. 10: 893–902.
3. Manjunath, N., M. Correa, M. Ardman, and B. Ardman. 1995. Negative regula-tion of T-cell adhesion and activation by CD43. Nature 377: 535–538.
4. Stockton, B. M., G. Cheng, N. Manjunath, B. Ardman, and U. H. von Andrian.1998. Negative regulation of T cell homing by CD43. Immunity 8: 373–381.
5. Woodman, R. C., B. Johnston, M. J. Hickey, D. Teoh, P. Reinhardt, B. Y. Poon,and P. Kubes. 1998. The functional paradox of CD43 in leukocyte recruitment:a study using CD43-deficient mice. J. Exp. Med. 188: 2181–2186.
6. Matsumoto, M., A. Shigeta, M. Miyasaka, and T. Hirata. 2008. CD43 plays bothantiadhesive and proadhesive roles in neutrophil rolling in a context-dependentmanner. J. Immunol. 181: 3628–3635.
7. Ostberg, J. R., R. K. Barth, and J. G. Frelinger. 1998. The Roman god Janus: aparadigm for the function of CD43. Immunol. Today 19: 546–550.
8. Rosenstein, Y., J. K. Park, W. C. Hahn, F. S. Rosen, B. E. Bierer, andS. J. Burakoff. 1991. CD43, a molecule defective in Wiskott-Aldrich syndrome,binds ICAM-1. Nature 354: 233–235.
9. Baum, L. G., M. Pang, N. L. Perillo, T. Wu, A. Delegeane, C. H. Uittenbogaart,M. Fukuda, and J. J. Seilhamer. 1995. Human thymic epithelial cells express anendogenous lectin, galectin-1, which binds to core 2 O-glycans on thymocytesand T lymphoblastoid cells. J. Exp. Med. 181: 877–887.
10. van den Berg, T. K., D. Nath, H. J. Ziltener, D. Vestweber, M. Fukuda, I. van Die,and P. R. Crocker. 2001. Cutting edge: CD43 functions as a T cell counterre-ceptor for the macrophage adhesion receptor sialoadhesin (Siglec-1). J. Immunol.166: 3637–3640.
11. Matsumoto, M., K. Atarashi, E. Umemoto, Y. Furukawa, A. Shigeta,M. Miyasaka, and T. Hirata. 2005. CD43 functions as a ligand for E-selectin onactivated T cells. J. Immunol. 175: 8042–8050.
12. Fuhlbrigge, R. C., S. L. King, R. Sackstein, and T. S. Kupper. 2006. CD43 is aligand for E-selectin on CLA� human T cells. Blood 107: 1421–1426.
13. Matsumoto, M., A. Shigeta, Y. Furukawa, T. Tanaka, M. Miyasaka, andT. Hirata. 2007. CD43 collaborates with P-selectin glycoprotein ligand-1 to me-diate E-selectin-dependent T cell migration into inflamed skin. J. Immunol. 178:2499–2506.
14. Alcaide, P., S. L. King, C. J. Dimitroff, Y. C. Lim, R. C. Fuhlbrigge, andF. W. Luscinskas. 2007. The 130-kDa glycoform of CD43 functions as an E-selectin ligand for activated Th1 cells in vitro and in delayed-type hypersensi-tivity reactions in vivo. J. Invest. Dermatol. 127: 1964–1972.
15. McEver, R. P., and R. D. Cummings. 1997. Role of PSGL-1 binding to selectinsin leukocyte recruitment. J. Clin. Invest. 100: S97–103.
7210 ROLE OF PSGL-1 IN T-CELL IMMUNE RESPONSES
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

16. Vestweber, D., and J. E. Blanks. 1999. Mechanisms that regulate the function ofthe selectins and their ligands. Physiol. Rev. 79: 181–213.
17. Patel, K. D., M. U. Nollert, and R. P. McEver. 1995. P-selectin must extend asufficient length from the plasma membrane to mediate rolling of neutrophils.J. Cell Biol. 131: 1893–1902.
18. Hirata, T., G. Merrill-Skoloff, M. Aab, J. Yang, B. C. Furie, and B. Furie. 2000.P-Selectin glycoprotein ligand 1 (PSGL-1) is a physiological ligand for E-selectinin mediating T helper 1 lymphocyte migration. J. Exp. Med. 192: 1669–1676.
19. Hirata, T., B. C. Furie, and B. Furie. 2002. P-, E-, and L-selectin mediate mi-gration of activated CD8� T lymphocytes into inflamed skin. J. Immunol. 169:4307–4313.
20. Yang, J., T. Hirata, K. Croce, G. Merrill-Skoloff, B. Tchernychev, E. Williams,R. Flaumenhaft, B. C. Furie, and B. Furie. 1999. Targeted gene disruption dem-onstrates that P-selectin glycoprotein ligand 1 (PSGL-1) is required for P-selec-tin-mediated but not E-selectin-mediated neutrophil rolling and migration. J. Exp.Med. 190: 1769–1782.
21. Xia, L., M. Sperandio, T. Yago, J. M. McDaniel, R. D. Cummings,S. Pearson-White, K. Ley, and R. P. McEver. 2002. P-selectin glycoprotein li-gand-1-deficient mice have impaired leukocyte tethering to E-selectin under flow.J. Clin. Invest. 109: 939–950.
22. Kitamura, T., Y. Koshino, F. Shibata, T. Oki, H. Nakajima, T. Nosaka, andH. Kumagai. 2003. Retrovirus-mediated gene transfer and expression cloning:powerful tools in functional genomics. Exp. Hematol. 31: 1007–1014.
23. Morita, S., T. Kojima, and T. Kitamura. 2000. Plat-E: an efficient and stablesystem for transient packaging of retroviruses. Gene Ther. 7: 1063–1066.
24. Powrie, F., M. W. Leach, S. Mauze, S. Menon, L. B. Caddle, and R. L. Coffman.1994. Inhibition of Th1 responses prevents inflammatory bowel disease in scidmice reconstituted with CD45RBhi CD4� T cells. Immunity 1: 553–562.
25. Powrie, F. 1995. T cells in inflammatory bowel disease: protective and patho-genic roles. Immunity 3: 171–174.
26. de Jong, Y. P., A. C. Abadia-Molina, A. R. Satoskar, K. Clarke, S. T. Rietdijk,W. A. Faubion, E. Mizoguchi, C. N. Metz, M. Alsahli, T. ten Hove, et al. 2001.Development of chronic colitis is dependent on the cytokine MIF. Nat. Immunol.2: 1061–1066.
27. Thurman, E. C., J. Walker, S. Jayaraman, N. Manjunath, B. Ardman, andJ. M. Green. 1998. Regulation of in vitro and in vivo T cell activation by CD43.Int. Immunol. 10: 691–701.
28. Walker, J., and J. M. Green. 1999. Structural requirements for CD43 function.J. Immunol. 162: 4109–4114.
29. Drew, E., J. S. Merzaban, W. Seo, H. J. Ziltener, and K. M. McNagny. 2005.CD34 and CD43 inhibit mast cell adhesion and are required for optimal mast cellreconstitution. Immunity 22: 43–57.
30. Carlow, D. A., S. Y. Corbel, and H. J. Ziltener. 2001. Absence of CD43 fails toalter T cell development and responsiveness. J. Immunol. 166: 256–261.
31. Tong, J., E. J. Allenspach, S. M. Takahashi, P. D. Mody, C. Park, J. K. Burkhardt,and A. I. Sperling. 2004. CD43 regulation of T cell activation is not through stericinhibition of T cell-APC interactions but through an intracellular mechanism.J. Exp. Med. 199: 1277–1283.
32. Serrador, J. M., M. Nieto, J. L. Alonso-Lebrero, M. A. del Pozo, J. Calvo,H. Furthmayr, R. Schwartz-Albiez, F. Lozano, R. Gonzalez-Amaro,P. Sanchez-Mateos, and F. Sanchez-Madrid. 1998. CD43 interacts with moesinand ezrin and regulates its redistribution to the uropods of T lymphocytes at thecell-cell contacts. Blood 91: 4632–4644.
33. Serrador, J. M., A. Urzainqui, J. L. Alonso-Lebrero, J. R. Cabrero, M. C. Montoya,M. Vicente-Manzanares, M. Yanez-Mo, and F. Sanchez-Madrid. 2002. A juxta-membrane amino acid sequence of P-selectin glycoprotein ligand-1 is involved inmoesin binding and ezrin/radixin/moesin-directed targeting at the trailing edge ofmigrating lymphocytes. Eur. J. Immunol. 32: 1560–1566.
34. Shaffer, M. H., R. S. Dupree, P. Zhu, I. Saotome, R. F. Schmidt,A. I. McClatchey, B. D. Freedman, and J. K. Burkhardt. 2009. Ezrin and moesinfunction together to promote T cell activation. J. Immunol. 182: 1021–1032.
35. Mody, P. D., J. L. Cannon, H. S. Bandukwala, K. M. Blaine, A. B. Schilling,K. Swier, and A. I. Sperling. 2007. Signaling through CD43 regulates CD4 T-celltrafficking. Blood 110: 2974–2982.
36. Baumheter, S., M. S. Singer, W. Henzel, S. Hemmerich, M. Renz, S. D. Rosen,and L. A. Lasky. 1993. Binding of L-selectin to the vascular sialomucin CD34.Science 262: 436–438.
37. Sassetti, C., K. Tangemann, M. S. Singer, D. B. Kershaw, and S. D. Rosen. 1998.Identification of podocalyxin-like protein as a high endothelial venule ligand forL-selectin: parallels to CD34. J. Exp. Med. 187: 1965–1975.
38. Kanda, H., T. Tanaka, M. Matsumoto, E. Umemoto, Y. Ebisuno, M. Kinoshita,M. Noda, R. Kannagi, T. Hirata, T. Murai, M. Fukuda, and M. Miyasaka. 2004.Endomucin, a sialomucin expressed in high endothelial venules, supports L-se-lectin-mediated rolling. Int. Immunol. 16: 1265–1274.
7211The Journal of Immunology
by guest on July 1, 2018http://w
ww
.jimm
unol.org/D
ownloaded from