mutant alcohol dehydrogenase (adh iii) presequences that affect
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, June 1990, p. 2801-28080270-7306/90/062801-08$02.00/0Copyright C 1990, American Society for Microbiology
Mutant Alcohol Dehydrogenase (ADH III) Presequences That AffectBoth In Vitro Mitochondrial Import and In Vitro Processing by the
Matrix ProteaseDUANE T. MOONEY, DAVID B. PILGRIM,t AND ELTON T. YOUNG*
Department ofBiochemistry, University of Washington, Seattle, Washington 98195
Received 26 July 1989/Accepted 2 March 1990
Point mutations in the presequence of the mitochondrial alcohol dehydrogerase isoenzyme (ADH III) havebeen shown to affect either the import of the precursor protein into yeast mitochondria in vivo or its processingwithin the organelle. In the present work, the behavior of these mutants during in vitro import into isolatedmitochondria was investigated. All point mutants tested were imported with a slower initial rate than that ofthe wild-type precursor. This defect was corrected when the precursors were treated with urea prior to import.Once imported, the extent of processing to the mature form of mutant precursors varied greatly and correlatedwell with the defects observed in vivo. This result was not affected by prior urea treatment. When matrixextracts enriched for the processing protease were used, this defect was shown to be due to failure of theprotease to efficiently recognize or cleave the presequence, rather than to a lack of access to the precursor. Therate of import of two ADHM precursors bearing internal deletions in the leader sequence was similar to thoseof the point mutants, whereas a deletion leading to the removal of the 15 amino-terminal amino acids waspoorly imported. The mature amino terminus of wild-type ADH In was determined to be Gln-25. Mutant mOl(Ser-26 to Phe), which reduced the efficiency of cleavage in vitro by 80%, was cleaved at the correct site.
Most proteins imported into the mitochondrion are syn-thesized in the cytoplasm as a precursor containing a tran-sient presequence at their amino terminus. (For reviews, seereferences 11, 17, and 20). From experiments with hybridproteins, the presequence (also called an import or leadersequence) is sufficient for mitochondrial targeting and import(13, 15, 27). Unfolding of the precursors during or beforebinding to mitochondria requires the action of the hsp70class of proteins (10) and can be mimicked during in vitroimport by incubation of precursors in urea prior to import(12). Once imported, the hsp60 protein facilitates refolding,oligomerization, and perhaps processing by the matrix pro-tease (7). The chelator-sensitive, matrix-associated proteasespecifically removes the amino-terminal presequence (15 to80 amino acids) (5) and has no demonstrated activity againstnonmitochondrial proteins. A majority of all precursorsimported to the matrix, even those destined for the inter-membrane space, are processed by this protease (21). Thesites of matrix protease action on several imported yeastproteins have been determined (1, 8, 20, 22, 24, 25); how-ever, no common sequence or motif near the cleavage site isevident.We have chosen the mitochondrial alcohol dehydrogenase
isoenzyme (ADH III) of the yeast Saccharomyces cerevisiaefor the study of targeting and import of mitochondrialproteins. The nuclear gene (ADH3) that endcodes this en-zyme has been cloned, and its DNA sequence has beendetermined (31). When the ADH3 sequence was comparedwith those of the closely related ADHI and ADH2 genes, an
amino-terminal extension was inferred in the ADH III pro-tein. This peptide extension was shown to be necessary andsufficient to allow correct binding and import into the mito-chondrial matrix (27, 28). Deletions and amino acid substi-
* Corresponding author.t Present address: MRC Laboratory of Molecular Biology, Cam-
bridge CB2 2QH, United Kingdom.
tutions in the leader peptide have been shown to affect thekinetics with which the mature form ofADH III is processedin vivo (23). In the present work, import of mutant precursorproteins into isolated mitochondria and processing by par-tially purified matrix protease were studied to determinewhether this defect was at the level of import or processing,or both. In addition, the amino terminus of mature ADH IIIwas determined for the wild-type protein and one mutant.
MATERIALS AND METHODS
Yeast strains and growth. Saccharomyces cerevisiae D273-10B (ATCC 24657) and 900-17-la were grown and used as
previously described (23).Plasmid construction, DNA manipulation, and yeast trans-
formation. Restriction nuclease digestion, cloning proce-dures, and yeast transformation were described previously(31). DNA fragments were isolated from low-melting-pointagarose as suggested by the supplier (Bethesda ResearchLaboratories, Inc., Gaithersburg, Md.).Mutant m80, whose primary ADH III translation product
lacks amino acids 1 through 28, was constructed as follows.Plasmid YEp-ADH3-m77 (23) (10 ,ug) was digested withXbaI. To remove the overhang while retaining the SphIrecognition site, the mixture was treated at room tempera-ture for 2 min with mung bean nuclease (New EnglandBioLabs, Inc., Beverly, Mass.) (see the details of the con-struction of m75 and m76 [23]). This fragment was restrictedwith EcoRI. The EcoRI-XbaI fragment that contained thepromoter and initiation codon of ADH3 was isolated andligated, along with a 1.3-kilobase-pair PvuII-SalI fragment ofADH3, into EcoRI-SalI-cut pMW5 vector (23). The resultingADH3 gene has been deleted from the HpaI to the PvuII sitein the wild-type sequence, but retains the initiator ATG. Thepredicted amino-terminal sequence of the m80 ADH IIIprotein is Met-Pr-Ser-Ala-Ile-Pro, where the underlinedamino acids are novel. The construction was verified byrestriction enzyme analysis. When expressed in S. cerevi-
2801
Vol. 10, No. 6
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 02
Dec
embe
r 20
21 b
y 31
.170
.60.
85.

2802 MOONEY ET AL.
TABLE 1. Alterations in ADH III presequence and their effects on mitochondrial import
% of imported % of proteinProteina ADH III amino acid sequenceb protein remaining bound but not
uncleavedc imported'
10 20 30WT M L R T S T L F T R R V Q P S L F S R N I L R L Q S T A A I P <1 3m76 M L R T S T L F T R R V Q P - A I P >99 8m77 - M P S R N I L R L Q S T A A I P NI 4m80 M P S A I P NI 1m81 M L R T S R V QP S L F S R N I L R L Q S T A A I P <5 1
mO4 C 16m73 I 13m78 I 56m69 I 94mOl F 80m79 F I 98mO6 I F I >99COX IV <1 1ADH I NI <0.5
a On the basis of the rate of appearance of mature ADH III in the pulse-labeling experiments of Pilgrim and Young (23), m73 and mO4 were imported and cleavedas rapidly as the wild-type, i.e., class I. Class II mutants mOl, m69, and m79 produced the mature form less rapidly than the wild type. Class III mutants mO6,m76, and m77 were not processed. Import of mutants m78, m80, and m81 was not previously examined.
b Mutations leading to the indicated amino acid alterations have been reported previously (23). Dashes indicate deleted amino acids. The arrow ( ) indicatesthe in vitro processing site.
' The total ADH III imported in Fig. 1 was measured by scanning densitometry. From these values the percentage of precursor present was determined. Thedata for the COX IV precursor were taken from Fig. 2A. NI, Not imported.
d The percentage of precursor bound but not imported was determined by first measuring the total amount of precursor bound plus precursor imported byomitting the proteinase K treatment prior to sedimentation, and then subtracting from that value the amount imported after proteinase K treatment andsedimentation.
siae, the plasmid led to the production of an active enzymeof the predicted size.An ADH3 gene carrying a DNA deletion which leads to
the removal of amino acids 6 through 10 in the ADH IIIprecursor, m81, has been generated as follows. The yeastexpression plasmid bearing the wild-type ADH3 gene,YEpADH3-9 (31), was subjected to three sets of doublerestriction digestions. The plasmid was restricted withEcoRI and Sall, and the 6.8-kb vector fragment was isolated.Digestion with EcoRI and HpaI allowed the purification ofthe 305-base-pair fragment 5' to the translational start site.Finally, the translated sequence (1,336 base pairs) wasisolated after XbaI-SaiI digestion. These three fragmentswere combined with a double-stranded oligonucleotide ofsequence
5'-AACACCATCATGTTGAGAACGTCACGTGTCCAACCAAGCCTATTTTTTGTGGTAGTACAACTCTTGCAGTGCACAGGTTGGTTCGGATAAAAGATC-5'
and ligated for 16 h at 12°C. DNA sequence analysis fromthe ADH3 AvaI site of recombinants that demonstrated ashort deletion by restriction analysis confirmed this se-quence. The inferred amino acid sequence is presented inTable 1. For in vitro expression of this pentapeptide deletionof the ADH III leader sequence, the HpaI-SaIl fragmentfrom YEpADH3-m81, containing the ADH3-m81 structuralgene, was cloned into SmaI-SalI digested pSP65.SDS-PAGE. 35S-labeled proteins were analyzed by sodium
dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) (18). The gels were treated as described previously(23).
In vitro protein synthesis. The wild-type and mutant ADH3genes were placed under the control of the SP6 promoter inthe plasmids pSP64 and pSP65 (Promega Biotec, Madison,Wis.). The SalI-BamHI fragment ofADH1, from the plasmidpADC1(-150) (3), was cloned into plasmid pSP64 to yield anADH1 gene under the control of the SP6 promoter. A similar
construction, including the yeast cytochrome oxidase sub-unit IV (COX4) gene, was kindly provided by D. Allison.Before transcription, these SP6 plasmids were restricted at asite 3' to the translational stop codon. Runoff transcriptionthen resulted in RNA bands of discrete sizes. In vitrotranscription was performed as described by Pilgrim andYoung (23), except that each nucleoside triphosphate waspresent at 500 p.M and the m7GpppG was omitted. Theprotein precursors were synthesized in vitro at 25°C for 2 hunless otherwise indicated (23). When urea was used topartially denature the precursor proteins, 20 ,1 of translationmix was added to 10.8 mg of urea or to 40 pul of 9 M urea.Once dissolved at 20°C, the resulting solution containing 6 Murea was incubated for 15 min at 20°C.
In vitro protein import into mitochondria. Mitochondriawere prepared from S. cerevisiae (23) and suspended in 0.5ml of 0.6 M sorbitol-20 mM N-2-hydroxyethylpiperazine-N'-2-etharesulfonic acid (HEPES)-KOH (pH 7.4). The yieldwas determined by diluting 10 pul of the mitochondrialsuspension in 1 ml 0.6% SDS. Following incubation at 100°Cfor 4 min, the A280 was measured. The approximate proteinconcentration was calculated as 1 A280 unit = 0.5 mg ofprotein per ml. Energy import mix (160 p.l) (0.6 M mannitol,50 mM HEPES-KOH [pH 7.4], 6 mM GTP, 1 mM ATP, 5mM phosphoenolpyruvate, 1 mM MgCl2, 1 mM dithiothrei-tol, 2 mM L-methionine, 40 mM KCI, 2.4 U of pyruvatekinase per ml) was warmed to 20°C, and 10 pul of fresh invitro-translated protein was added. The import reaction wasinitiated by addition of 100 pug of fresh mitochondria, incu-bated at 20°C for the times indicated, and then stopped byincubation on ice in the presence of 10 p.l of freshly preparedproteinase K (5 mg/ml; Sigma Chemical Co., St. Louis, Mo.)for 30 min. Mitochondria were reisolated from all mixturesby a 10-min centrifugation at 4°C through a 0.6-ml sucrosecushion (20% [wtlvol] sucrose, 100 mM KCl, 2 mM MgCl2,20 mM HEPES-KOH [pH 7.4]) in a microcentrifuge. The
MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 02
Dec
embe
r 20
21 b
y 31
.170
.60.
85.

IMPORT AND PROCESSING OF ADH III 2803
pellet was suspended in 25 ,ul of sorbitol-HEPES buffer,made 1 x in SDS sample buffer, and boiled for 2 min beforesamples were loaded for SDS-PAGE. Electrophoresis andfluorography were performed as described previously (23).
Preparation and assay of mitochondrial matrix protease.The mitochondrial matrix protease was partially purified byadaptation of the methods of Bohni et al. (4). Yeast cellswere grown to late logarithmic phase in 8 liters of YEPmedium containing 3% glycerol, and mitochondria wereprepared from 52 g (wet weight) of cells. The final mitochon-drial suspension in 0.6 M sorbitol-20 mM HEPES-KOH (pH7.4) was diluted 10-fold in 10 mM Tris (pH 7.4)-30 mMNaCl-1 mM phenylemthylsulfonyl fluoride and held on icefor 30 min. The mitoplasts were lysed at 0°C by 30 passes ina tight-fitting Dounce homogenizer, and the particulate frac-tion was removed by sedimentation for 10 min at 17,000 x g.The supernatant was applied at 4°C to a prewashed DE-52column (1.5 by 6.5 cm; Whatman International, Ltd., Maid-stone, England) equilibrated with 10 mM Tris (pH 7.4)-30mM NaCl. After a 10-ml wash with this buffer, the columnwas resolved with a linear 60-ml 30 to 600 mM NaClgradient. Column fractions of 1 ml were obtained and held at-70°C until assayed (see below). Fractions containing ma-trix protease activity were pooled and concentrated approx-imately 10-fold at 0°C by using an ultrafiltration apparatusfitted with a PM10 filter (Amicon Corp., Danvers, Mass.).The concentrate was diluted 10-fold in 10 mM Tris (pH 7.4)at 0°C, and the volume was again reduced 10-fold. The finalsolution, containing 5.2 mg of protein in 2 ml, was made 50%in glycerol and stored at -70°C.
In vitro assay of the matrix protease was carried out at30°C in 20 to 50 ,ul of 10 mM Tris (pH 7.5)-50 uM CoCl2-50p.M ZnCl2-1 mM phenylmethylsulfonyl fluoride. Freshlyprepared in vitro-synthesized 35S-labeled precursor proteins(4 to 8 ,ul) were added, and the reaction was started by two-to fourfold dilution of the protease preparation into this mix.When the kinetics of the cleavage reaction were analyzed,samples were removed at the times indicated, made 1 x inSDS sample buffer, and stored at -20°C until SDS-PAGEanalysis. The cleavage was maximal after 30 to 60 min andwas typically 15 to 60% of the input precursor. This resultdoes not represent a loss in enzyme activity over time, asCOX IV precursor was cleaved by the protease when addedafter 1 h of reaction (data not shown).
Determination of the amino terminus of mature ADH III.An in vitro translation mix was prepared, lacking alanine,isoleucine, and [35S]methionine. It contained 15 ,uCi ofL-[U-14C]alanine (174 mCi/mmol) and 20 ,uCi of L-[4,5-3H]isoleucine (95 Ci/mmol) (Amersham Corp., ArlingtonHeights, Ill.) in a total volume of 100 ,ul. Translation wascarried out for 1.5 h at 30°C. In vitro cleavage by the matrixprotease was performed under standard conditions, in a300-,ul total volume containing 100 p.l of protease. After 1 hat 30°C, the reaction mix was chilled to 4°C and diluted to660 p.l. The final solution contained 1% Triton X-100, 0.3 MNaCl, 0.1 M Tris (pH 7.4), 10 mM EDTA, and 300 ,ud ofanti-ADH antiserum. The immune reaction was incubated at4°C for 2 h with gentle mixing, 435 p.l of crude Staphylococ-cus A cell suspension (Sigma) was added, and gentle mixingwas continued for 2 h at 4°C. The particulate fraction wascollected by centrifugation, and the pellet was washed threetimes with 1-ml portions of a buffer containing 0.1% SDS, 10mM NaPO4 (pH 8), and 2 mM EDTA. The final pellet wassuspended in 300 pul of 10 mM NaPO4 (pH 8)- 2% SDS-2.5%2-mercaptoethanol and boiled for 7 min. Insoluble materialwas removed by centrifugation, and the clear supernatant
was dialyzed overnight at 4°C in 1 liter of 1 mM NaPO4 (pH7.4)- 0.1% SDS. The final solution contained 1.2 x 105 cpmof 3H and 4.8 x 103 cpm of 14C.The 3H- and 14C-labeled ADH III was sequenced on an
amino acid sequencer (model 890C; Beckman Instruments,Inc., Fullerton, Calif.) by using a program (6) with 0.1 MQuadrol buffer and polybrene (26). Ten Edman degradationcycles were carried out, and the anilinethiazolinone deriva-tives were concentrated by evaporation of the chlorobutanesolvent. The derivatives were dissolved in 10 ml of Aquasolscintillation cocktail (Du Pont, NEN Research Products,Boston, Mass.) and counted in a scintillation counter (modelLS-230; Beckman).
RESULTS
Mutants with deletions and point mutations in the prese-quence of ADH III have previously been assayed in vivo forkinetics of accumulation of mature ADH III in the mitochon-drial matrix (23). To examine separately mitochondrial im-port and processing of the mutant ADH III precursors, therate of import into purified, energized mitochondria wasmeasured in vitro (Fig. 1A). Bound, but not imported,precursors were distinguished from imported precursor mol-ecules by their sensitivity to proteinase K digestion. Bothprecursor and mature forms of imported ADH III wereidentified after proteinase K-treated mitochondria were pel-leted through a sucrose pad. SDS-PAGE followed by auto-radiography allowed the rate of import to be quantitated byscanning densitometry. Several control experiments wereperformed to demonstrate that import of both mutant andwild-type precursors was normal. When the energy gradientacross the inner membrane was disrupted with the ionophorevalinomycin, import was blocked and no ADH III wasprotected (data not shown). As a further control, there wasno protection when the mitochondria were lysed with deter-gent after import but prior to proteinase K treatment (datanot shown).
In vitro import of point mutant ADH III precursors. Twobands present in import lanes of m69 and other mutants wereshown to have the apparent molecular weights expected forprecursor and mature ADH III (Fig. 1A). Although little ifany wild-type ADH III precursor was proteinase K pro-tected during the import reactions shown in Fig. 1A, theprecursors of all mutants tested were a significant fraction ofthe imported protein. The extent of processing to the matureform varied greatly among the mutants, ranging from lessthan 1% to more than 95% (Table 1). This defect in process-ing was characteristic for each mutant and corroborated theresults of in vivo imports (23). No processing during in vitroimport of mO6 was detected. The total percent import at eachtime point was estimated by scanning densitometry forwild-type and mutant ADH III proteins (Fig. 1B). A two- tothreefold decrease in the rate of import relative to the wildtype was observed for all mutants tested.Urea treatment stimulates import of the ADH HI precursor.
During the development of the foregoing experiments, therate of import of the ADH III precursor was compared withthat of COX IV, since only low levels of added ADH IIIprecursor were typically imported in 20 min at 20°C. Theinitial import rate of COX IV precursor was four- to fivefoldgreater than that of the ADH III precursor (Fig. 2A) andsuggests that the extent of import of COX IV may besignificantly greater as well. Other lines of evidence havealso suggested that in vitro synthesis of the ADH III precur-sor can lead to import-incompetent conformation(s) which
VOL. 10, 1990
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 02
Dec
embe
r 20
21 b
y 31
.170
.60.
85.

2804 MOONEY ET AL.
AC 2 WT 7
m
c
m
mO42 7
20
-tw
20
c
c
40
m692 7
mOl2 7
B
*I////
a142 1!
-i
20
20
.WT
,m 73
.m79m06
-mO.m044
mint e s _minutes
FIG. 1. In vitro mitochondrial import of ADH III wild-type andmutant precursors. After the energy import mix was brought to20°C, fresh, in vitro-translated precursors were added and theimport reaction was initiated by the addition of mitochondria.Aliquots were removed and treated with proteinase K, and themitochondria were sedimented through sucrose. (A) Autoradio-graphs of SDS-PAGE analysis. Lane C contains 7.5% of theprecursor removed at each time point. (B) The percentage of totalADH III imported was determined by scanning densitometry.Equivalent amounts of radioactive mutant precursors, synthesizedin samples of the same reticulocyte lysate, were added in eachimport reaction.
may represent a significant fraction of the total in vitro-translated precursor. Gel filtration chromatography of invitro-synthesized ADH III precursor demonstrated thatsome material eluted at a position consistent with the knowntetrameric structure of the mature, matrix-associated en-zyme. However, a significant fraction of the added precursoreluted in the void volume, suggesting that the precursor hadaggregated (data not shown).
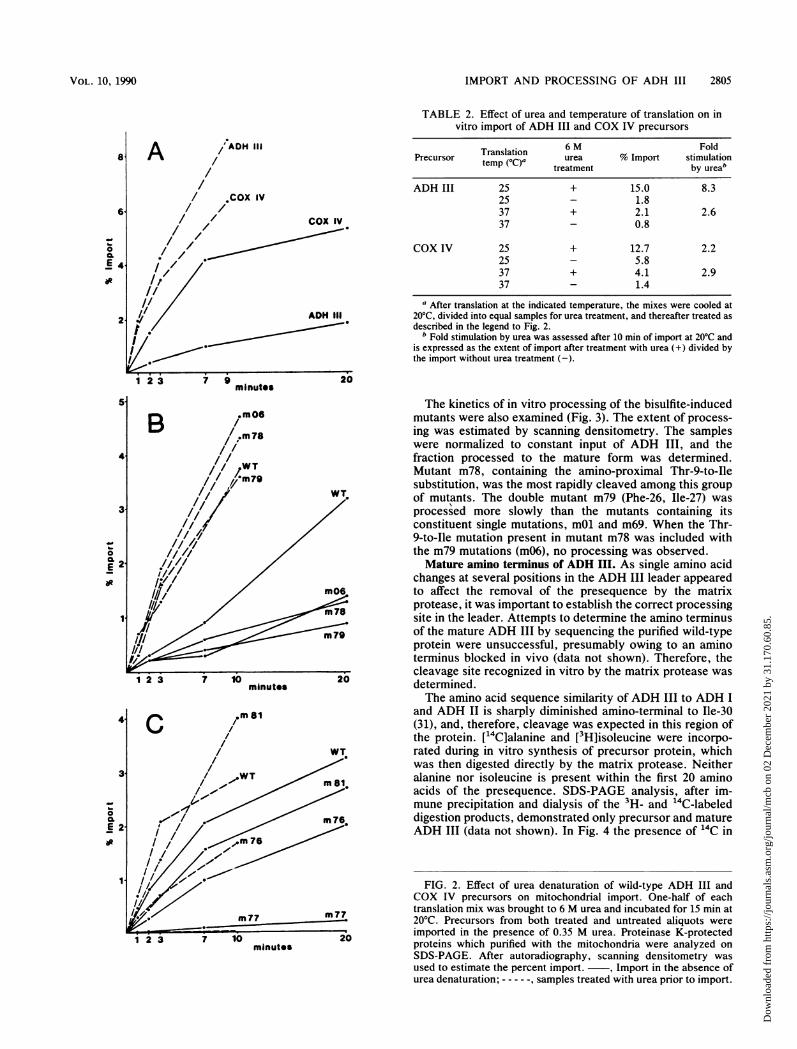
Pretreatment of precursors with urea has been shown tostimulate mitochondrial import, presumably by unfolding theprecursors (12, 30). Incubation of COX IV and ADH IIIprecursors, synthesized at 25°C, in 6 M urea for 15 min at20°C significantly stimulated their rates of import, makingtheir rates more similar in the presence of urea than in itsabsence (Fig. 2A; Table 2). The rate of import of ADH III
precursor was stimulated 5- to 10-fold. The final concentra-tion of urea in all import reactions, 0.35 M, was shown tohave no effect on these rates (12; data not shown). Alsosuggestive of the presence of import-incompetent structureswas the effect of the temperature of translation upon theurea-dependent stimulation of import. Import of COX IVprecursors synthesized at 25 and 37°C was stimulated to asimilar degree by urea treatment (Table 2). However, thestimulation of import by urea treatment of the ADH IIIprecursor synthesized at 37°C was significantly reduced fromthe stimulation seen after synthesis at 25°C, suggesting thatan import-incompetent structure(s) was formed during syn-thesis at 37°C which cannot be corrected by the ureatreatment used.The effect of urea treatment on the import of the mutants
with point mutations in ADH III was examined (Fig. 2B).The import rates of all mutants tested were stimulated byprior urea treatment to rates indistinguishable from that ofthe wild type when translated at 25°C, even though theywere all slower than the wild type in the absence of urea.When wild-type and mutant precursors were synthesized at37°C, the urea-mediated stimulations of their import rateswere similar.
Deletion mutants of the ADH III presequence. Deletionmutants m76, m77, m80, and m81 displayed various degreesof import deficiency. The pentapeptide deletion in m81 led toa reduction in import rate which was similar to those for thepoint mutants (Fig. 2C), and the precursor was processed toa form which comigrated with the mature wild-type protein(data not shown). Consistent with these results, expressionof the m81 precursor in S. cerevisiae showed that it wasenzymatically active and properly localized to the mitochon-drial matrix (data not shown). The rate of import of thelarger, internal deletion mutant m76 was also reduced rela-tive to the wild type. No processing of m76 was demon-strated. Import of both m81 and m76 was stimulated, al-though to differing degrees, by treatment with urea.
Import of the m77 precursor revealed a more severedefect. Although similar amounts of m77 and wild-typeprecursors were bound to mitochondria at 30°C (Table 1),less than 0.3% of m77 was protected from protease after a20-min import, whereas more than 3% of the wild-typeprecursor was protected (Fig. 2C). Urea was unable tostimulate the import of m77. Although m81 was processedafter import, no action of the matrix protease in vitro couldbe demonstrated on the m77 precursor, even though thewild-type processing site was present (data not shown). Themutant lacking the entire leader sequence, m80, bound at alow but significant level (Table 1); however, no import wasobserved (data not shown).
In vitro proteolytic processing of the ADH III precursor.The altered rates of processing observed for the pointmutants during in vitro import could be due to defects atseveral points in the import process. The last step, process-ing by the matrix protease, was examined by first partiallypurifying the protease (see Materials and Methods) and thenperforming the processing reaction in vitro by using ADH IIIprecursors synthesized in vitro. The matrix protease specif-ically cleaves native precursor proteins (4, 9). The rate ofcleavage of the wild-type ADH III precursor was examinedin vitro by using a partially purified preparation of the matrixprotease (Fig. 3). In general, 15 to 60% of the input precursorwas cleaved. This cleavage was sensitive to chelating agents(data not shown) and produced an ADH III protein whichmigrated on SDS-PAGE at the same rate as did ADH IIIpurified from S. cerevisiae (data not shown).
MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 02
Dec
embe
r 20
21 b
y 31
.170
.60.
85.
IMPORT AND PROCESSING OF ADH III 2805
IIADH III
/
i/// / .cox iv ox I
I/// /
/// ADH"".I-~/I,
I.~~~~~~~~~~~~~
TABLE 2. Effect of urea and temperature of translation on invitro import of ADH III and COX IV precursors
Translation 6 M FoldPrecursor temn(Ca urea % Import stimulationtemp (°C)a treatment by ureab
ADH III 25 + 15.0 8.325 - 1.837 + 2.1 2.637 - 0.8
COX IV 25 + 12.7 2.225 - 5.837 + 4.1 2.937 - 1.4
a After translation at the indicated temperature, the mixes were cooled at20°C, divided into equal samples for urea treatment, and thereafter treated asdescribed in the legend to Fig. 2.
b Fold stimulation by urea was assessed after 10 min of import at 20°C andis expressed as the extent of import after treatment with urea (+) divided bythe import without urea treatment (-).
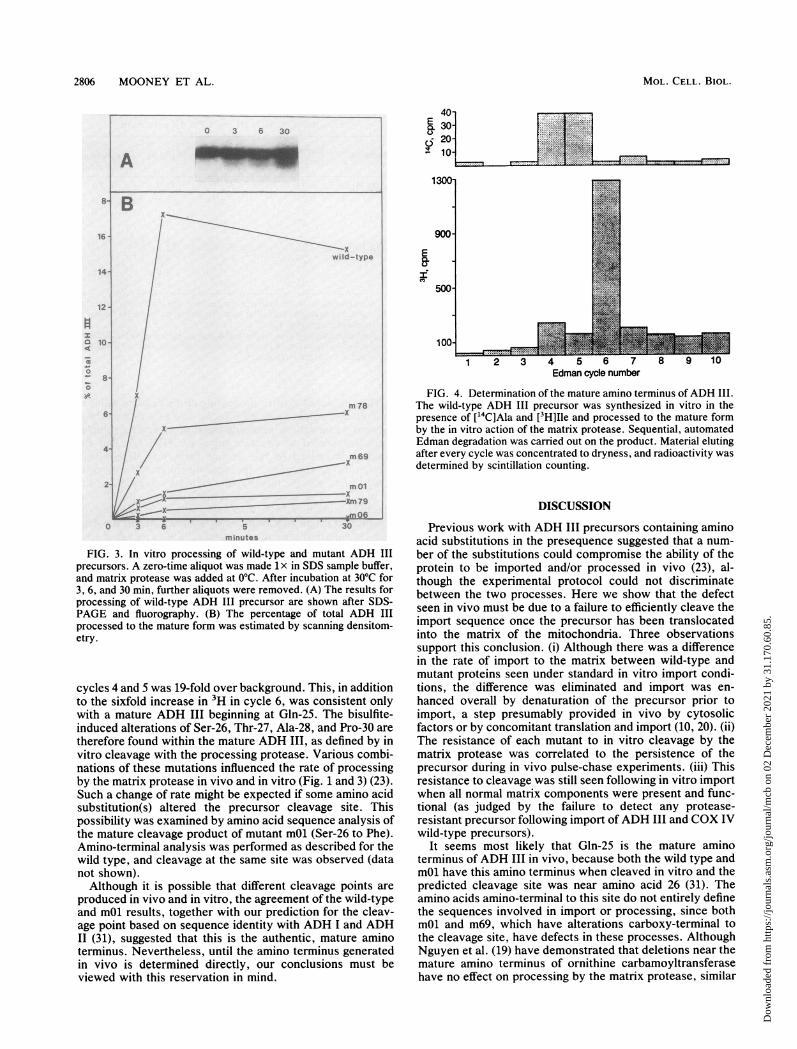
The kinetics of in vitro processing of the bisulfite-inducedmutants were also examined (Fig. 3). The extent of process-ing was estimated by scanning densitometry. The sampleswere normalized to constant input of ADH III, and thefraction processed to the mature form was determined.Mutant m78, containing the amino-proximal Thr-9-to-Ilesubstitution, was the most rapidly cleaved among this groupof mutants. The double mutant m79 (Phe-26, Ile-27) wasprocessed more slowly than the mutants containing itsconstituent single mutations, mOl and m69. When the Thr-9-to-Ile mutation present in mutant m78 was included withthe m79 mutations (mO6), no processing was observed.Mature amino terminus of ADH III. As single amino acid
changes at several positions in the ADH III leader appearedto affect the removal of the presequence by the matrixprotease, it was important to establish the correct processingsite in the leader. Attempts to determine the amino terminusof the mature ADH III by sequencing the purified wild-typeprotein were unsuccessful, presumably owing to an aminoterminus blocked in vivo (data not shown). Therefore, thecleavage site recognized in vitro by the matrix protease wasdetermined.The amino acid sequence similarity of ADH III to ADH I
and ADH II is sharply diminished amino-terminal to Ile-30(31), and, therefore, cleavage was expected in this region ofthe protein. [14C]alanine and [3H]isoleucine were incorpo-rated during in vitro synthesis of precursor protein, whichwas then digested directly by the matrix protease. Neitheralanine nor isoleucine is present within the first 20 aminoacids of the presequence. SDS-PAGE analysis, after im-mune precipitation and dialysis of the 3H- and "4C-labeleddigestion products, demonstrated only precursor and matureADH III (data not shown). In Fig. 4 the presence of 14C in
FIG. 2. Effect of urea denaturation of wild-type ADH III andCOX IV precursors on mitochondrial import. One-half of eachtranslation mix was brought to 6 M urea and incubated for 15 min at20°C. Precursors from both treated and untreated aliquots wereimported in the presence of 0.35 M urea. Proteinase K-protectedproteins which purified with the mitochondria were analyzed onSDS-PAGE. After autoradiography, scanning densitometry wasused to estimate the percent import. , Import in the absence ofurea denaturation; ---, samples treated with urea prior to import.
8
6
E 4
2
1 2 3 7 minutes 20
-ft
0
E
-
0
E
minut*s
VOL. 10, 1990
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 02
Dec
embe
r 20
21 b
y 31
.170
.60.
85.
2806 MOONEY ET AL.
0 3 6 30
_U_~I
13001
BX
16-i II16 1,
i X| wild-type
141 i
121 /
10
<
8
61 1 m78
:/X
41/ / tmOl61~~~~~~~~~~6
21/ 08m69
X R~~~~~~~~~060 3 6 5 30
minutes
FIG. 3. In vitro processing of wild-type and mutant ADH IIIprecursors. A zero-time aliquot was made 1lx in SDS sample buffer,and matrix protease was added at 0°C. After incubation at 30°C for3, 6, and 30 mm, further aliquots were removed. (A) The results forprocessing of wild-type ADH III precursor are shown after SDS-PAGE and fluorography. (B) The percentage of total ADH IIIprocessed to the mature form was estimated by scanning densitom-etry.
cycles 4 and 5 was 19-fold over background. This, in additionto the sixfold increase in 3H in cycle 6, was consistent onlywith a mature ADH III beginning at Gln-25. The bisulfite-induced alterations of Ser-26, Thr-27, Ala-28, and Pro-30 aretherefore found within the mature ADH III, as defined by invitro cleavage with the processing protease. Various combi-nations of these mutations influenced the rate of processingby the matrix protease in vivo and in vitro (Fig. 1 and 3) (23).Such a change of rate might be expected if some amino acidsubstitution(s) altered the precursor cleavage site. Thispossibility was examined by amino acid sequence analysis ofthe mature cleavage product of mutant mOl (Ser-26 to Phe).Amino-terminal analysis was performed as described for thewild type, and cleavage at the same site was observed (datanot shown).Although it is possible that different cleavage points are
produced in vivo and in vitro, the agreement of the wild-typeand mOl results, together with our prediction for the cleav-age point based on sequence identity with ADH I and ADHII (31), suggested that this is the authentic, mature aminoterminus. Nevertheless, until the amino terminus generatedin vivo is determined directly, our conclusions must beviewed with this reservation in mind.
900q
Ea.a)I-
500-
100-1=1 2
Edman cycle number
FIG. 4. Determination of the mature amino terminus ofADH III.The wild-type ADH III precursor was synthesized in vitro in thepresence of [14C]Ala and [3H]Ile and processed to the mature formby the in vitro action of the matrix protease. Sequential, automatedEdman degradation was carried out on the product. Material elutingafter every cycle was concentrated to dryness, and radioactivity wasdetermined by scintillation counting.
DISCUSSION
Previous work with ADH III precursors containing aminoacid substitutions in the presequence suggested that a num-ber of the substitutions could compromise the ability of theprotein to be imported and/or processed in vivo (23), al-though the experimental protocol could not discriminatebetween the two processes. Here we show that the defectseen in vivo must be due to a failure to efficiently cleave theimport sequence once the precursor has been translocatedinto the matrix of the mitochondria. Three observationssupport this conclusion. (i) Although there was a differencein the rate of import to the matrix between wild-type andmutant proteins seen under standard in vitro import condi-tions, the difference was eliminated and import was en-hanced overall by denaturation of the precursor prior toimport, a step presumably provided in vivo by cytosolicfactors or by concomitant translation and import (10, 20). (ii)The resistance of each mutant to in vitro cleavage by thematrix protease was correlated to the persistence of theprecursor during in vivo pulse-chase experiments. (iii) Thisresistance to cleavage was still seen following in vitro importwhen all normal matrix components were present and func-tional (as judged by the failure to detect any protease-resistant precursor following import ofADH III and COX IVwild-type precursors).
It seems most likely that Gln-25 is the mature aminoterminus ofADH III in vivo, because both the wild type andmOl have this amino terminus when cleaved in vitro and thepredicted cleavage site was near amino acid 26 (31). Theamino acids amino-terminal to this site do not entirely definethe sequences involved in import or processing, since bothmOl and m69, which have alterations carboxy-terminal tothe cleavage site, have defects in these processes. AlthoughNguyen et al. (19) have demonstrated that deletions near themature amino terminus of ornithine carbamoyltransferasehave no effect on processing by the matrix protease, similar
40-E 30-& 20-- 10
-
MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 02
Dec
embe
r 20
21 b
y 31
.170
.60.
85.

IMPORT AND PROCESSING OF ADH III 2807
deletions carboxy-terminal to the processing site of theATP2 gene product do affect processing (2, 29). Such con-tributions from the mature protein may vary, depending onthe secondary structures of the regions analyzed. The pri-mary sequence at the mature amino terminus of ADH IIImay affect processing by influencing a local secondarystructure which affects the action of the matrix protease.Mutations nearer the amino terminus of the leader had less
severe effects on processing than those nearer the cleavagesite when analyzed as single mutations. Their effects can begreat, however, when tested in combination with othermutations. The effects of the Thr-9-to-Ile alteration (m78)were similar to those of two other single mutations, mO4 andm73 when analyzed individually. When tested in combina-tion with the m79 double mutant no processing was observedin either in vitro assay. Consistent with other studies (16, 19,29), import of ADH III does not require cleavage of thepresequence.The analysis of deletion mutants in the presequence sug-
gested functional redundancy of the import information inthe leader. The pentapeptide TLFTR (positions 6 through10) is similar to that beginning at residue 15, SLFSR.Deletion mutant m76 retained only the amino-terminal 14amino acids of the presequence, and mutant m81 containeda deletion of amino acids 6 to 10. These mutant precursorswere imported with similar kinetics, but m76, no longercontaining the wild-type processing site, was not detectablyprocessed. The amino-terminal deletion in m77 eliminatedamino acids 1 to 17 and disrupted both the import andprocessing signals present in the wild-type precursor.We previously showed (23) that certain mutants were
imported and processed with rates indistinguishable fromthat of the wild type during in vivo import experiments.However, in vitro import analysis demonstrated that nomutant precursors were imported as rapidly as the wild type.Similar differences between in vitro and in vivo importexperiments have also been reported for mutations of COX4(14) and F1lo ATPase (29). Although the cause of thisdifference has not been specifically determined, a largefraction of the intact precursor used in in vitro experimentsis proposed to have folded into a structure which is resistantto processing. When synthesis and import occur concomi-tantly in vivo, this aberrant structure may not exist insignificant amounts owing to the proposed chaperoningfunction of hsp70 (10). The presence of such structures afterin vitro synthesis is inferred from the effects of urea upon theinitial rate of import in vitro, since the mutant-dependentdecrease in rate relative to the wild type, as well as the rateof the wild type itself, is influenced positively by urea. Aproblem in protein folding may also be reflected in the invitro processing data, since a large fraction of the inputwild-type precursor is never processed, even though thematrix protease remains active. The rapid, and linear, initialrate of processing may reflect enzyme action on a smallfraction of the precursor population which is in a processing-competent conformation. The decrease in rate and finalextent of processing observed for the mutants tested mayreveal that the processing-competent conformation is evenless stable for mutant precursors.
This mutational analysis of the ADH III precursor hasshown that a broad spectrum of alterations, both pointmutations and deletions, affect both mitochondrial importand precursor processing. No entirely redundant elementshave been observed, since all mutations have led to analtered phenotype at one or more levels. It is proposed thatthe amino acid sequence and secondary structure(s) of the
leader sequence, as well as the adjacent sequences in themature amino terminus of ADH III, all contribute informa-tion for normal mitochondrial binding, import, and process-ing.
ACKNOWLEDGMENTS
This study was supported by Public Health Service grantGM33779 from the National Institutes of Health. D.P. was sup-ported by a postgraduate scholarship from the Natural Sciences andEngineering Research Council, Ottawa, Ontario, Canada.We thank Roger Wade, Harry Charbonneau, and Ken Walsh for
their advice in designing the amino-terminal sequencing experimentsand for their excellent technical expertise in performing the auto-mated Edman degradations. We also thank D. Allison and G.Parraga for critical reading of this manuscript.
LITERATURE CITED1. Beckman, J. D., P. 0. Ljungdahl, J. L. Lopez, and B. L.
Trumpower. 1987. Isolation and characterization of the nucleargene encoding the Rieske iron-sulfur protein (RIP1) from Sac-charomyces cerevisiae. J. Biol. Chem. 262:8901-8909.
2. Bedwell, D. M., D. J. Klionsky, and S. C. Emr. 1987. The yeastFl-ATPase 1B subunit precursor contains functionally redundantmitochondrial protein import information. Mol. Cell. Biol. 7:4038-4047.
3. Beier, D. R., and E. T. Young. 1982. Characterization of aregulatory region upstream of the ADR 2 locus in S. cerevisiae.Nature (London) 300:724-728.
4. Bohni, P. C., G. Daum, and G. Schatz. 1983. Import of proteinsinto mitochondria: partial purification of a matrix-located pro-tease involved in cleavage of mitochondrial precursor polypep-tides. J. Biol. Chem. 258:4937-4943.
5. Bohni, P., S. Gasser, C. Leaver, and G. Schatz. 1980. In A. M.Kroon and C. Saccone (ed.), The organization and expression ofthe mitochondrial genome, p. 423-433. Elsevier/North-HollandPublishing Co., Amsterdam.
6. Brauer, A. W., M. N. Margolies, and E. Haber. 1975. Theapplication of 0.1 M quadrol to the microsequence of proteinsand the sequence of tryptic peptides. Biochemistry 14:3029-3035.
7. Cheng, M., F.-U. Harti, J. Martin, R. Poliock, F. Kalousek, W.Neupert, E. Hallberg, R. Hallberg, and A. Horwich. 1989.Mitochondrial heat-shock protein hsp60 is essential for assem-bly of proteins imported into yeast mitochondria. Nature (Lon-don) 337:620-625.
8. Cumsky, M. G., C. E. Trueblood, C. Ko, and R. 0. Poyton.1987. Structural analysis of two genes encoding divergent formsof yeast cytochrome c oxidase subunit V. Mol. Cell. Biol.7:3511-3519.
9. Daum, G., P. C. Bohni, and G. Schatz. 1982. Import of proteinsinto mitochondria. Cytochrome b2 and cytochrome c peroxidaseare located in the intermembrane space of yeast mitochondria.J. Biol. Chem. 257:13028-13033.
10. Deshaies, R., B. Koch, M. Werner-Washburne, E. Craig, and R.Schekman. 1988. A subfamily of stress proteins facilitates trans-location of secretory and mitochondrial precursor polypeptides.Nature (London) 332:800-805.
11. Douglas, M. G., M. T. McCammon, and A. Vassarotti. 1986.Targeting proteins into mitochondria. Microbiol. Rev. 50:166-178.
12. Eilers, M., S. Hwang, and G. Schatz. 1988. Unfolding andrefolding of a purified precursor protein during import intoisolated mitochondria. EMBO J. 7:1139-1145.
13. Horwich, A. L., F. Kalousek, and L. E. Rosenberg. 1985.Arginine in the leader peptide is required for both import andproteolytic cleavage of a mitochondrial precursor. Proc. Natl.Acad. Proc. USA 82:4930-4933.
14. Hurt, E. C., D. S. Allison, U. Miller, and G. Schatz. 1987.Amino-terminal deletions in the presequence of an importedmitochondrial protein block the targeting function and proteo-lytic cleavage of the presequence at the carboxy terminus. J.Biol. Chem. 262:1420-1424.
VOL. 10, 1990
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 02
Dec
embe
r 20
21 b
y 31
.170
.60.
85.

2808 MOONEY ET AL.
15. Hurt, E. C., B. Pesold-Hurt, and G. Schatz. 1984. The amino-terminal region of an imported mitochondrial precursor poly-peptide can direct cytoplasmic dihydrofolate reductase into themitochondrial matrix. EMBO J. 3:3149-3156.
16. Hurt, E. C., B. Pesold-Hurt, K. Suda, W. Oppliger, and G.Schatz. 1985. The first twelve amino acids (less than half thepresequence) of an imported mitochondrial protein can directmouse cytosolic dihydrofolate reductase into the yeast mito-chondrial matrix. EMBO J. 4:2061-2068.
17. Hurt, E. C., and A. P. G. M. van Loon. 1986. How proteins findmitochondria and intramitochondrial compartments. TrendsBiochem. Sci. 11:204-207.
18. Laemmeli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
19. Nguyen, M., C. Argan, W. P. Sheffield, A. W. Bell, D. Shields,and G. C. Shore. 1987. A signal sequence domain essential forprocessing, but not import, of mitochondrial pre-ornithine car-bamyl transferase. J. Cell Biol. 104:1193-1198.
20. Nicholson, D. W., and W. Neupert. 1988. Synthesis and assem-bly of mitochondrial proteins, p. 677-746. In R. C. Das andP. W. Robbins (ed.), Protein transfer and organelle biogenesis.Academic Press, Inc., San Diego, Calif.
21. Ohashi, A., J. Gibson, I. Gregor, and G. Schatz. 1982. Import ofproteins into mitochondria. The precursor of cytochrome ci isprocessed in two steps, one of them heme-dependent. J. Biol.Chem. 257:13042-13047.
22. Patterson, T. E., and R. 0. Poyton. 1986. COX8, the structuralgene for yeast cytochrome c oxidase subunit VIII. J. Biol.Chem. 261:17192-17197.
23. Pilgrim, D. B., and E. T. Young. 1987. Primary structurerequirements for correct sorting of the yeast mitochondrial
protein ADH III to the yeast mitochondrial matrix space. Mol.Cell. Biol. 7:294-304.
24. Pratje, E., G. Mannhaupt, G. Michaelis, and K. Beyreuther.1983. A nuclear mutation prevents processing of a mitochondri-ally encoded membrane protein in Saccharomyces cerevisiae.EMBO J. 2:1049-1054.
25. Takeda, M., W.-J. Chen, J. Saltzgaber, and M. G. Douglas.1986. Nuclear genes encoding the yeast mitochondrial ATPasecomplex. J. Biol. Chem. 261:15126-15133.
26. Tarr, G. E., J. F. Beecher, M. Bell, and D. J. McKean. 1978.Polyquarternary amines prevent peptide loss from sequenators.Anal. Biochem. 84:622-627.
27. van Loon, A. P. G. M., A. W. Brandli, and G. Schatz. 1986. Thepresequences of two imported mitochondrial proteins containinformation for intracellular and intramitochondrial sorting. Cell44:801-812.
28. van Loon, A. P. G. M., and E. T. Young. 1986. Intracellularsorting of alcohol dehydrogenase isoenzymes in yeast: a cyto-solic location reflects absence of an amino-terminal targetingsequence for the mitochondrion. EMBO J. 5:161-165.
29. Vassarotti, A., W.-J. Chen, C. Smagula, and M. G. Douglas.1987. Sequences distal to the mitochondrial targeting sequencesare necessary for the maturation of the F1-ATPase P-subunitprecursor in mitochondria. J. Biol. Chem. 262:411-418.
30. Verner, K., and B. Lemire. 1989. Tight folding of a passengerprotein can interfere with the targeting function of a mitochon-drial presequence. EMBO J. 8:1491-1495.
31. Young, E. T., and D. B. Pilgrim. 1985. Isolation and DNAsequence of ADH3, a nuclear gene encoding the mitochondrialisozyme of alcohol dehydrogenase in Saccharomyces cerevi-siae. Mol. Cell. Biol. 5:3024-3034.
MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 02
Dec
embe
r 20
21 b
y 31
.170
.60.
85.