mechanism of the fermentation of … of the fermentation of lactose by yeasts by morrison rogosa*...
TRANSCRIPT

MECHANISM OF THE FERMENTATION OF LACTOSE BY YEASTS
BY MORRISON ROGOSA*
(From the Bureau of Dairy Industry, Agricultural Research Administration, United States Department of Agriculture, Washington)
(Received for publication, April 17, 1948)
There are two fundamental theories concerning the fermentation of disaccharides. One presupposes an “indirect” and the other a “direct” fermentation. A theory proposed by Fischer (l-3) was that no compound sugar is fermented directly as such, but that the compound sugar is cleaved by appropriate hydrolytic enzymes into its monose constituents, the lat- ter actually being fermented as such. Briefly stated this is the classical theory of the indirect fermentation of compound sugars.
Leibowitz and Hestrin (4) stated that:
“Fischer’s explanation, owing largely perhaps to the high authority of its author, was widely accepted. Essentially a speculative theory based on an attractive corre- lation, it assumed in the minds of both chemists and biologists the status of a proved fact. Even though Fischer himself had not even attempted to subject his theory to crucial quantitative test, the theory went almost unchallenged for nearly a gener- ation. Moreover, its scope was broadened and the belief became general that the Fischer mechanism described not only alcoholic fermentation of oligosaccharides by yeast, but also the fermentations of oligosaccharides by bacteria, and equally the glycolysis of compound sugars by tissues of the higher forms of life. Laborde’s [(5)] early criticism of the Fischer view in its application to sucrose metabolism in hetero- ferment,ative lactic acid bacteria was simply ignored.”
Willstatter and Oppenheimer (6) observed that lactose-fermenting yeasts may ferment lactose at a faster rate than they ferment the monose constitutents of lactose, glucose, and galactose, either individually or in mixtures in concentrations equivalent to those of lactose. They found that the concentration of lactase in yeast extra& was insufficient to hydrolyze lactose at a rate equivalent to the fermentation of lactose by the intact cells. In some instances the lactase content of yeast extracts was negligible or zero. Because technical difficulties make it impossible to conduct ex- periments on lactase with the intact cell, certain objections can be raised to a positive statement that direct fermentation occurs. Thus, the evi- dence for direct fermentation presented by Willstatter is suggestive and inferential, but not conclusive.
Hestrin (7) demonstrated that lactose can be fermented more rapidly than hexoses by using a yeast strain from Palestine Leben, and Myrback
* Present address, National Institute of Health, Bethesda, Maryland. 413
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from

414 LACTOSE FERMENTATION BY YEAST
and Vasseur (8) obtained a similar result with Saccharomyces fragilis and Torula cremoris.
Kluyver and Custers (9) noted that Blastodendrion intermedium fer- mented lactose, but did not respire in it, whereas the organism both fer- mented and respired in glucose. The lack of respiration in lactose would indicate t.hat glucose, on which the cells do respire, was not formed.
Wright (IO, 11) has suggested a direct fermentation of lactose by Strep- tococcus th.ermophd?.rs. In t,he present author’s laboratory, experiments with Streptococcus thcrmophilus have resulted in more rapid fermentation of lactose than of glucose. Fermentation of glucose was always slow or slight and in one instance a fermentation of lactose was nearly complete before glucose was fermented at all.
Hoff-Jgrgensen et al. (12) have shown that one strain resembling Lucto- bacillus bulgaricus will grow in a medium containing glucose plus lactose but will not grow in the same medium containing glucose only.
The evidence for the direct fermentation of maltose and certain other carbohydrates has been reviewed by Leibowitz and Hestrin (4).
Experiments in this laboratory in which attempts have been made to isolate a lactase from extracts of lactose-fermenting yeasts have yielded disappointing results. ,4t different times either no lactase was found or the quantity present was negligible. In the latter case the presence of lactase was a transitory phenomenon. These experiments induced us to undertake the work reported here.
The evidence for the direct fermentation of lactose, as presented by other workers, is persuasive and is based primarily on the kinetics of the fermenta- tion. However, cert,ain objections are difficult to invalidate.
It is the purpose of this paper to present data showing the relative rates of fermentation by diverse types of lactose-fermenting yeasts, as well as to present new and more definite proof of the direct fermentation of lactose by lactose-fermenting yeasts.
EXPERIMENTAL
Methods
The organisms employed in this work were Torulopsis keJir 149, Sac- charomyces la&is 131, Torula lactosa 168, Zygosaccharomyces lactis 90, Torulopsis sphaerica 13, Xaccharomyces anamensis 145,l type F 93,2 Candida pseudotropicalis 32, Mycotorula la&is 130, Saccharomyces fragilis 15, and Torulopsis cremoris 2.
1 This strain, received from the Lister Institute, is obviousIy incorrectly desig- nated. According to Stelling-Dekker (13) Xaccharomyces anamensis should ferment maltose but not lactose.
2 Type F appears to be a variety of Sacchar~mycesfragilis unable to ferment inulin,
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from

M. ROGOSA 415
According to Diddons and Lodder (14), Mycotorula la& is a representa- tive of Can&da pseudotropicalis and To&a lu~tosa a strain of Can&da pseudotropicalis var. lactosa. These organisms are as representative a group of la.ctose-fermenting yeasts and yeast-like fungi as it, was possible to obtain.
Strains 145, 93, 32, 130, 2, and 15 were incubated at 37” and strains 131, 149, 90, 13, and 168 at 30”. These temperatures are within the optimal range for both growth and fermentation.
The source cultures used for inoculating the experimental fermentations were grown on agar slopes of the following composition: Bacto-tryptone 0.5 per cent, Bacto-yeast extract 0.3 per cent, KH2P04 0.2 per cent, agar 2 per cent, and glucose 2 per cent. Except for one type of experiment, which will be described later, the stock cultures had been grown for many transfers (five times weekly for nearly a year) on the above medium con- taining glucose as the sole carbohydrate. Since many yeasts are capable of adapting themselves to the fermentation of certain carbohydrates, particularly galactose, glucose was chosen in order to avoid the complica- tions of adaptation. The importance and significance of this point will be discussed later.
The inocula for the fermentations were grown on agar slopes for 48 hours. The cells were washed from the surface and suspended in sterile distilled water. The suspension was shaken mechanically and uniform aliquots in any given experiment were inoculated into the fermentation flasks which contained a medium of the following composition: Bacto-tryptone 0.5 per cent, Bacto-yeast extract 0.3 per cent, KHzP04 0.2 per cent, 85 per cent lactic acid 0.04 per cent, and the appropriate carbohydrates or mixtures of carbohydrates at t,he desired concentrations. The pH of this medium was approximately 5.1 and remained constant during autoclaving. The flasks were of 125 ml. capacity and contained ‘78 gm. of the fermentation medium which had been dispensed by an aut,omatic pipet.te. The flasks were plugged with cotton and autoclaved no longer than 5 minutes at 15 pounds steam pressure in order to avoid decomposition of the sugars.
Glucose and galactose were Pfanstiehl products with specific rotations of +52.5” and +80.5”, respectively. The lactose was recrystallized from C.P. lactose and treated with norit A six times. The specific rotation of the lactose was +52.5” at equilibrium. These values were obtained inde- pendently by t\vo analysts and are in agreement with accept’ed values (15).
The progress of the fermentations was followed by a direct, determina- tion of t,he residual suga.r by polarimetric a.nalysis. 3 times the normal weight of the sugar were taken in each case and clarified with lead sub- acetate. The normal weights were accepted as 32.248 gm. for glucose, 32.857 gm. for lactose, and 21.429 gm. for galact,ose. All polarimetric
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from
416 LACTOSE FERMENTATION BY YEAST
analyses were conducted at 20”. Results of many analyses were usually within 0.05 per cent of the known concentrations of the various carbohy- drates. For the analyses of mixtures of glucose and galactose, the primary fermentation with the lactose-fermenting yeast was terminated by mo- mentary autoclaving. After cooling, the flask was reinoculated with Saccharomyces chevalieri, American Type Culture Collection No. 9804. This yeast, ferments glucose but not lactose or galactose, and it does not become adapted to the fermentation of galactose even after long periods of time. Thus the residual galactose was determined directly, and the residual glucose in a mixture of glucose and galactose was determined by
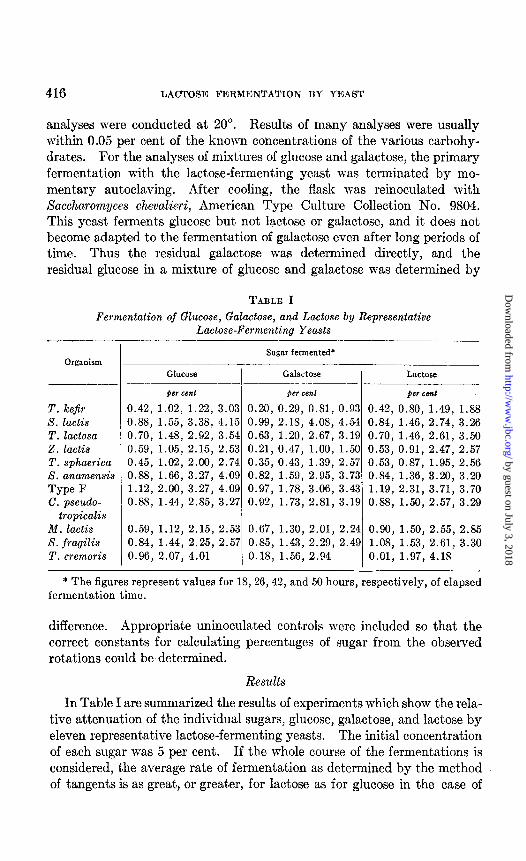
TABLE I
Fermentation of Glucose, Galactose, and Lactose by Representative Lactose-Fermenting Yeasts
Sugar fermented* Organism -- .-
GlUCOSe Gala&.x Lactose --
)CI cent per cent #cr cm1
T. kefir 0.42, 1.02, 1.22, 3.03 0.20, 0.29, 0.81, 0.93 0.42, 0.80, 1.49, 1.88 S. lactis 0.88, 1.55, 3.38, 4.15 0.99, 2.18, 4.08, 4.54 0.84, 1.46, 2.74, 3.26 T. lactosa 0.70, 1.48, 2.92, 3.54 0.63, 1.20, 2.67, 3.19 0.70, 1.46, 2.61, 3.50 2. lactis 0.59, 1.05, 2.15, 2.53 0.21, 0.47, 1.00, 1.50 0.53, 0.91, 2.47, 2.57 T. sphaerica 0.45, 1.02, 2.00, 2.74 0.35, 0.43, 1.39, 2.57 0.53, 0.87, 1.95, 2.56 S. anamensis 0.88, 1.66, 3.27, 4.09 0.82, 1.59, 2.95, 3.73 0.84, 1.36, 3.20, 3.20 Type F 1.12, 2.00, 3.27, 4.09 0.97, 1.78, 3.06, 3.43 1.19, 2.31, 3.71, 3.70 C. pseudo- 0.88, 1.44, 2.85, 3.27 0.92, 1.73, 2.81, 3.19 0.88, 1.50, 2.57, 3.29
tropicalis M. lactis 0.59, 1.12, 2.15, 2.53 0.67, 1.30, 2.01, 2.24 0.90, 1.50, 2.55, 2.85 S. fragilis 0.84, 1.44, 2.25, 2.57 0.85, 1.43, 2.29, 2.49 1.08, 1.53, 2.61, 3.30 T. cremoris 0.96, 2.07, 4.01 0.18, 1.56, 2.94 0.01, 1.97, 4.18
* The figures represent values for 18,26,42, and 50 hours, respectively, of elapsed fermentation time.
difference. Appropriate uninoculated controls were included so that the correct constants for calculating percentages of sugar from the observed rotations could be determined.
Results
In Table I are summarized the results of experiments which show the rela- tive attenuation of the individual sugars, glucose, galactose, and lactose by eleven representative lactose-fermenting yeasts. The initial concentration of each sugar was 5 per cent. If the whole course of the fermentations is considered, the average rate of fermentation as determined by the method of tangents is as great, or greater, for lactose as for glucose in the case of
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from

M. ROGOSA 417
several organisms. The average rates, expressed as the percentage of sugar fermented per hour, in the case of glucose and lactose, respectively, were 0.077 and 0.079 for Torula lactosa, 0.088 and 0.098 for type F, 0.065 and 0.064 for Candida pseudotropicalis, 0.052 and 0.057 for Mycotorula la&s, 0.052 and 0.068 for Saccharomyces jragilis, and 0.10 and 0.174 for Torulopsis cremoris. Moreover the maximal rates of a fermentation were greater for lactose than for glucose with all the above organisms and also with Zygosaccharomyces lactis and Saccharomyces anamensis. This must mean that lactose was being dissimilated more rapidly than glucose, at least during one stage of the fermentations. And this occurred even with organisms that could be considered glucose-adapted strains.
A striking result, as Table I shows, was the relative slowness with which galactose was fermented by most of the strains. Only four of the eleven strains fermented galactose at average rates, throughout the entire fer- mentation, that were equal to or in excess of those for glucose. Of these four strains only two, Saccharomyces la&is and S. jrqilis, consistently fermented galactose as well as they fermented glucose, or better, at nearly all intervals of the fermentations, and only one strain, S. lactis, fermented galactose more readily than either glucose or lactose for the entire duration of the fermentations. With this organism the average rates for glucose, galactose, and lactose, respectively, were 0.086, 0,099, 0.067; the maximal rates were 0.114, 0.149, and 0.080. Thus, in only one of eleven instances can it be said that galactose was fermented fairly consistently at a rate greatly in excess of that of lactose.
One peculiar feature of the galactose fermentation is the occasional in- stance in which galactose is fermented more slowly than either glucose or lactose during the greater part of the fermentation, until suddenly galactose begins to be fermented in an explosive-like manner (see the results with Torulopsis sphaerica, Table I), The rate of fermentation may change abruptly from a value of 0.047 to 0.147 for a relatively short period. The author is not referring to changes from the lag or induction period to the logarithmic phase of fermentation, but rather to abrupt changes of rate within the logarithmic phase itself. Obviously, such an abrupt change would affect the average rate as well as the maximal rate of fermentation and make it appear from the figures that galactose is fermented better than it really is during most of the fermentation period.
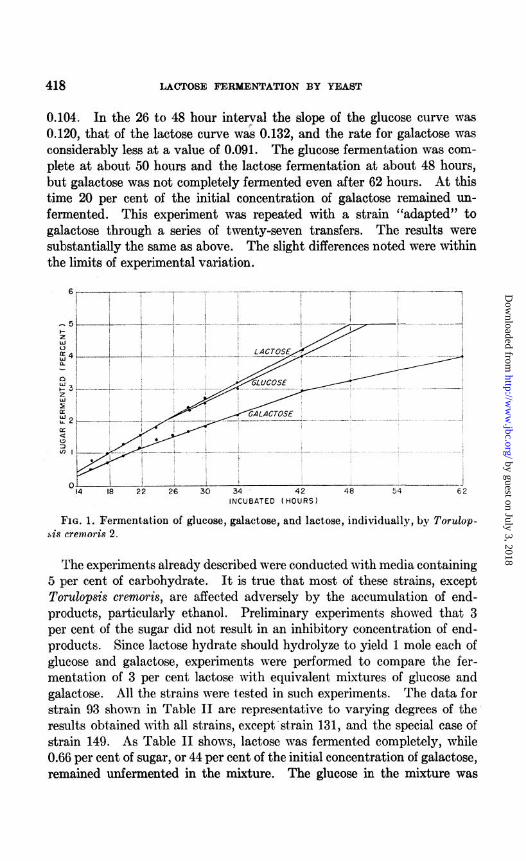
The results shown in Table I do not describe the course of a complete fermentation in which all the init.ial sugar is finally fermented. Fig. 1, however, does depict the entire courses of the fermentations of glucose, galactose, and lactose by Torulopsis cremoris. During the 14 to 26 hour interval, the slopes of the glucose and lactose curves were practically identical at a value of 0.142, and the slope of the galactose curve was only
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from
418 LACTOSE FERMENTATION BY YEAST
0.104. In the 26 to 48 hour interyal the slope of the glucose curve was 0.120, that of the lactose curve was 0.132, and the rate for galactose was considerably less at a value of 0.091. The glucose fermentation was com- plete at about 50 hours and the lactose fermentation at about 48 hours, but galactose was not completely fermented even after 62 hours. At this time 20 per cent of the initial concentration of galactose remained un- fermented. This experiment was repeated with a strain “adapted” to galactose through a series of twenty-seven transfers. The results were substantially the same as above. The slight differences noted were within the limits of experimental variation.
INCUBATED I HOURS)
FIG. 1. Fermentation of glucose, galactose, and lactose, individually, by TOT&~- sis cremoris 2.
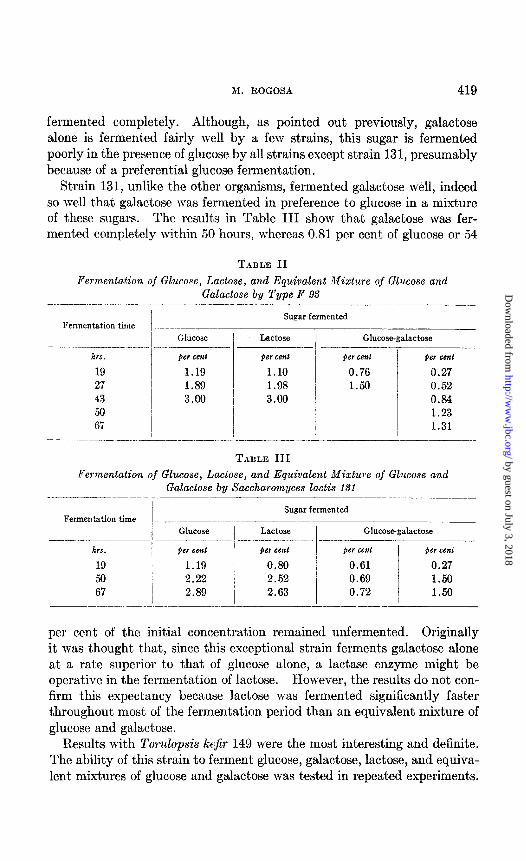
The experiments already described were conducted with media containing 5 per cent of carbohydrate. It is true that most of these strains, except Torulopsis cremoris, are affected adversely by the accumulation of end- products, particularly ethanol. Preliminary experiments showed that 3 per cent of the sugar did not result in an inhibitory concentration of end- products. Since lactose hydrate should hydrolyze to yield 1 mole each of glucose and galactose, experiments were performed to compare the fer- mentation of 3 per cent lactose with equivalent mixtures of glucose and galactose. All the strains were t,est,ed in such experiments. The data for strain 93 shown in Table II are representative to varying degrees of the results obtained with all strains, except’strain 131, and the special case of strain 149. As Table II shows, lactose was fermented completely, while 0.66 per cent of sugar, or 44 per cent of the initial concentration of galactose, remained unfermented in the mixture. The glucose in the mixture was
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from
M. ROGOSA 419
ferment,ed completely. Although, as pointed out previously, galactose alone is fermented fairly well by a few strains, this sugar is fermented poorly in the presence of glucose by all strains except strain 131, presumably because of a preferential glucose fermentation.
Strain 131, unlike the other organisms, fermented galactose well, indeed so well that galactose was fermented in preference to glucose in a mixture of these sugars. The results in Table III show that galactose was fer- mented completely within 50 hours, whereas 0.81 per cent of glucose or 54
TABLE II
Perm,entation of Glucose, Lactose, and Equivalent Mixture of Glucose and Galactose by Type P 93
..-- .-I-... Glucose I Lactose
hrs. per cent per cent
19 1.19 1.10 27 1.89 / 1.98 43 3.00 I 3.00 50 I
67 /
I
I
Glucose-galactose
J+er cemt )er cc?u
0.76 0.27 1.50 0.52
0.84 1.23 1.31
TABLE III
Fermentation of Glucose, Lactose, and Equivalent Mixture of Glucose and Galactose by Saccharomyces la&is 131
Fermentation time I __-.--.--- Glucose
- hrs. fm cent
19 1.19 50 2.22 67 I 2.89
Sugar fermented ..-___-
Lactose Glucose-galactose -..
per cenl per cent per cent
0.80 2.52 j
0.61 0.27 0.69 1.50
2.63 0.72 / 1.50 --____
per cent of the initial concentration remained unfermented. Originally it was thought that, since this exceptional strain ferments galactose alone at a rate superior to that of glucose alone, a lactase enzyme might be operative in the fermentation of lactose. However, the results do not con- firm this expectancy because lactose was fermented significantly faster throughout most of the fermentation period t,han an equivalent mixture of glucose and galactose.
Results with Torulopsis liefir 149 were the most interesting and definite. The ability of this strain to ferment glucose , galactose, lactose, and equiva- lent mixtures of glucose and galact,ose was t*ested in repeated experiments.
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from
420 LACTOSE FERMENTATION BY YEAST
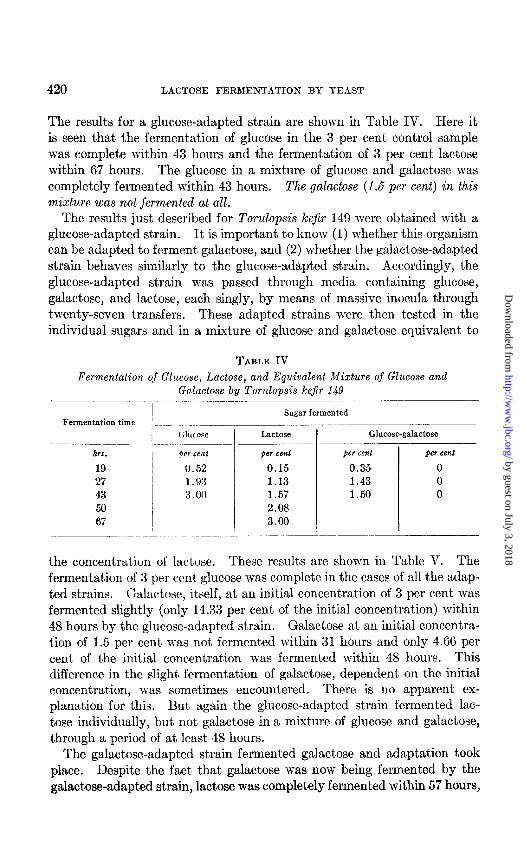
The results for a glucose-adapted strain are shown in Table IV. Here it is seen that the fermentation of glucose in the 3 per cent control sample was complete within 43 hours and the fermentation of 3 per cent lactose within 67 hours. The glucose in a mixture of glucose and galactose was completely fermented within 43 hours. The galactose (1.5 per cent) in this mixture was not fermented at all.
The results just described for Torulopsis kefir 149 were obtained with a glucose-adapted strain. It is important to know (1) whether this organism can be adapted to ferment galactose, and (2) whether the galactose-adapted strain behaves similarly to the glucose-adapted strain. Accordingly, the glucose-adapted strain was passed through media containing glucose, galactose, and lactose, each singly, by means of massive inocula through twenty-seven transfers. These adapted strains were then tested in the individual sugars and in a mixture of glucose and galactose equivalent to
TABLE IV
Fermentation of Glucose, Lactose, and Equivalent Mixture of Glucose and Galactose by Torulopsis kefir 149
- I
Fermentation time
hm.
19 27 43 50 67
GlUt-OSe
fler cent
0.52 1.93 3.00
.I. Sugar fermented
Lactose Glucose-galactose -
per cent per cent per cent
0.15 0.35 0 1.13 1.43 0 1.57 1.50 0 2.08 3.00
- -
the concentration of lactose. These results are shown in Table V. The fermentat,ion of 3 per cent glucose was complete in the cases of all the adap- ted strains. Galactose, itself, at an initial concentration of 3 per cent was fermented slightly (only 14.33 per cent of the initial concentration) within 48 hours by the glucose-adapted strain. Galactose at an initial concentra- tion of 1.5 per cent was not fermented within 31 hours and only 4.66 per cent of the initial concentration was fermented within 48 hours. This difference in the slight fermentation of galactose, dependent, on the initial concentration, was sometimes encountered. There is no apparent ex- planation for this. But again the glucose-adapt.ed strain fermented lac- tose individually, but not galactose in a mixture of glucose and galactose, through a period of at least 48 hours.
The galactose-adapted strain fermented galactose and adaptation took place. Despite the fact that galactose was now being fermented by the galactose-adapted strain, lactose was completely fermented within 57 hours,
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from
M. ROGOSA 421
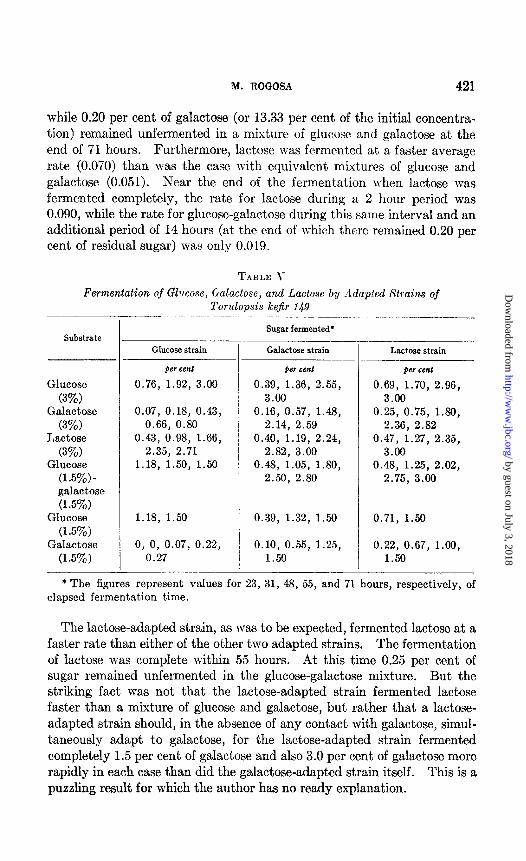
While 0.20 per cent of galactose (or 13.33 per cent of the initial concentra- tion) remained unfermented in a mixture of glucose and gdactose at the end of 71 hours. Furthermore, lactose was fermented at a faster average rate (0.070) than was the case with equivalent mixtures of glucose and galactose (0.051). Near the end of the fermentation when lactose was fermented completely, the rate for lactose during a 2 hour period was 0.090, while the rate for glucose-galactose during this same interval and an additional period of 14 hours (at, the end of which t,here remained 0.20 per cent of residual sugar) was only 0.019.
TABLE \
Fermentation of Glucose, Galactose, and Lactose bg Adapted Strains of
Substrate
Glucose
(3%) Galactose
(3%) Lactose
(3%) Glucose
(1.5%) - galactose
(1.5%) Glucose
(1.5%) Galactose
(1.5%)
I Torulopsis kejir 149 -
Sugar fermented*
Glucose strain
per Cent
0.76, 1.92, 3.00
0.07, 0.18, 0.43, 0.66, 0.80
0.43, 0.98, 1.66, 2.35, 2.71
1.18, 1.50, 1.50
1.18, 1.50 0.39, 1.32, 1.50 0.71, 1.50
0, 0, 0.07, 0.22, 0.10, 0.55, 1.25, 0.22, 0.67, 1.00, 0.27 1.50 1.50
Galactose strain
per cent ger teat
0.39, 1.36, 2.55, 0.69, 1.70, 2.96, 3.00 3.00
0.16, 0.57, 1.48, 0.25, 0.75, 1.80, 2.14, 2.59 2.36, 2.82
0.40, 1.19, 2.24, 0.47, 1.27, 2.35, 2.82, 3.00 3.00
0.48, 1.05, 1.80, 0.48, 1.25, 2.02, 2.50, 2.80 2.75, 3.00
Lactose strain
* The figures represent values for 23, 31, 48, 55, and 71 hours, respectively, of elapsed fermentation time.
The lactose-adapted strain, as was to be expected, fermented lactose at a faster rate than either of the other two adapted strains. The fermentation of lactose was complete within 55 hours. At this time 0.25 per cent of sugar remained unfermented in the glucose-galactose mixture. But the striking fact was not that the lactose-adapted strain fermented lactose faster than a mixture of glucose and galactose, but rather that a lactose- adapted strain should, in the absence of any contact with galactose, simul- taneously adapt to galactose, for the lactose-adapted strain fermented completely 1.5 per cent of galactose and also 3.0 per cent of galactose more rapidly in each case than did the galactose-adapted strain itself. This is a puzzling result for which the author has no ready explanation.
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from

422 LACTOSE FERMENTATIOS BY YEAST
DISCUSSION
All of the organisms studied ferment,ed lactose at a faster rate than that at which they fermented a mixture of glucose and galactose. In addition, however, it must be remembered that one glucose-adapted strain fermented lactose completely but did not ferment galactose in a mixture of glucose and galactose. It is difficult on the basis of this direct result to argue that a preliminary hydrolysis of lactose in a fermentation is necessary.
The reason that lactose is fermented faster than a mixture of glucose and galactose lies in the fact that galactose, itself, normally is fermented with relative difficulty. On the basis of present knowledge it is only possible to speculate as to the reason for the slowness of the galactose fermentation.
The importance of phosphorus in carbohydrate metabolism is now well established. If preliminary hydrolysis of lactose in a fermentation does
LACTOSE + ADENOSINE-TRIPHOSPHATE m LACTOSE-PHOSPHATE tADENOSINE- DIPHOSPHATE
ENZYME
ENZYMES PYRUVIC ACID. . ..(- LACTOSE-PHOSPHATE
ENZYME
GLYCOGEN+H3P04
not occur, then it is necessary to postulate a reaction analogous to that of Price, Cori, and Colowick (16). This is shown in the accompanying scheme.
The reaction scheme may not be exactly as shown, but it is not unreason- able to assume the formation of phosphoric esters with disaccharides. Doudoroff et al. (17) have already demonstrated the phosphorolysis of sucrose and related compounds in ferment,ation by Pseudomonas sac- charophila.
The existence of galactose-l-phosphate has been demonstrated by Kosterlitz (18). The author cannot explain the simultaneous adaptation of a lactose-adapted strain to galactose. The possibility of the existence of a lactose-phosphate ester has not yet been investigated. Until these gaps in our knowledge are bridged, it is not possible to argue with assurance concerning the mechanism of lactose fermentation. Considerable work remains to be done.
SUMMARl-
The lactose-fermenting yeasts adapted to glucose, galactose, or lactose fermented lactose faster than they fermented an equivalent mixture of
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from

M. ROGOSA 423
glucose and galactose. Xormally this is because of a relatively weal; gdactose fermentation.
One lactose-adapted strain, in the absence of any contact with galactose, simultaneously became adapted to the fermentation of galactose.
One glucose-adapted strain fermented lactose completely, while it did not ferment galactose in a mixture of glucose and galactose.
Enzymatic hydrolysis of lactose is unnecessary for the fermentation of lactose. Fermentation may be “direct” by means of phosphorolysis and phosphorylat,ion.
The significance of these results in relation to modern schemes of carbo- hydrate metabolism is discussed.
BIBLIOGRAPHY
1. Fischer; E., Ber. them. Ges., 27, 3479 (1894). 2. Fischer, IL, 2. physio2. Chem., 26, 72 (1898). 3. Fischer, E., and Lindner, P., Ber. them. Ges., 28, 3034 (1895). 4. Leibowitz, J., and Hestrin, S., in Nord, F. F., and We&man, C. I-I., Advances in
enzymology and related subjects of biochemistry, New York, 6, 87 (1945). 5. Laborde, I., Compt. rend. Acad., 138, 228 (1904). 6. Willstatter, R., and Oppenheimer, G., Z. physiol. Chem., 118, 168 (1922). 7. Hestrin S., Dissertation, Hebrew University, Jerusalem (1941) (cf. (4)). 8. Myrbiick, K., and Vasseur, E., 2. physiol. Chem., 277,171 (1943). 9. Kluyver, A., and Custers, M., Antonie van Leeuwenhoek, J. Microbial. and Serol.,
6, 121 (1940) (cf. (4)). 10. Wright, H., J. Path. and Baci., 42, 31 (1936); 43, 487 (1936). 11. Wright, H., J. Path. and Bact., 46, 117 (1937); 46,261 (1938). 12. Hoff-Jergensen, E., Williams, W. L., and Snell, R. E., J. Biol. Chem., 168, 773
(1947). 13. Stelling-Dekker, N. X, Verhandel. Koninkl. Akad., Wetensch. Amsterdam, A.fdeel.
N&Lurk., 28, sect. 2, 1 (1931). l-4. Diddens, Ii. A., and Lodder, J., Die anaskosporogenen Hefen, Amsterdam, pt. 2
(1945). 15. Prowne, C. A., and Zerban, F. W., Physical and chemical methods of sugar analy-
sis, New York, 3rd edition (1941). 16. Price, W. II., Cori, C. F., and Colowick, S. P., J. Biol. C’hem., 160, 633 (1945). 17. Doudoroff, M., Kaplan, N., and Hassid, W. Z., J. Biol. Chem., 148, 67 (1943). 18. Kosterlitz, If., J. Physiol., 93, 34 (1938).
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from

Morrison RogosaOF LACTOSE BY YEASTS
MECHANISM OF THE FERMENTATION
1948, 175:413-423.J. Biol. Chem.
http://www.jbc.org/content/175/1/413.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/175/1/413.citation.full.haccessed free atThis article cites 0 references, 0 of which can be
by guest on July 3, 2018http://w
ww
.jbc.org/D
ownloaded from