isolation biochemical characterization s-layer …jb.asm.org/content/170/6/2631.full.pdfisolation...
TRANSCRIPT

Vol. 170, No. 6JOURNAL OF BACTERIOLOGY, June 1988, p. 2631-26380021-9193/88/062631-08$02.00/0Copyright © 1988, American Society for Microbiology
Isolation and Biochemical Characterization of the S-Layer Proteinfrom a Pathogenic Aeromonas hydrophila Strain
JAMES S. G. DOOLEY,' WILLIAM D. McCUBBIN,2 CYRIL M. KAY,2, AND TREVOR J. TRUST'*Department ofBiochemistry and Microbiology, University of Victoria, Victoria, British Columbia, Canada V8W 2Y2,1and Medical Research Council of Canada Group on Protein Structure and Function, Department of Biochemistry,
University ofAlberta, Edmonton, Alberta, Canada T6G 2H7 2
Received 23 October 1987/Accepted 14 March 1988
The regular surface protein array (S layer) present on Aeromonas hydrophila TF7 is composed of a singlespecies of protein of apparent molecular weight 52,000. This protein was extracted from whole cells bytreatment with 0.2 M glycine hydrochloride (pH 3.0). The protein was purified to homogeneity by ion-exchangechromatography and reverse-phase high-performance liquid chromatography. Amino acid compositionanalysis showed that the protein contained 520 residues per molecule, 41% of which were hydrophobic.Cysteine was absent. A pI of 4.6 was determined for the protein, and only a single isoelectric form was detected.The purified protein displayed the hydrophobic characteristic of binding to octyl-Sepharose gels, but the saltaggregation test showed that it did not confer hydrophobicity to the cell surface when present as an intact Slayer. The molecule aggregated strongly in aqueous solution as determined by sedimentation equilibriumstudies. Circular dichroism spectra showed that the S-layer protein was composed of a large amount of D-sheet(approximately 44%), a limited amount of ca-helix (19%), and 12% j-turn, with the remainder of the moleculebeing aperiodic. No significant difference in secondary structure content was measured in the presence of themetal chelator EDTA. The N-terminal amino acid sequence was determined for the first 30 residues. Nosequence homology with other S-layer proteins was found.
Paracrystalline surface protein arrays, or S layers, havebeen observed on a wide variety of archaebacteria andeubacteria (44). These structures normally constitute theoutermost layer of the cell envelope, where they are ideallysited to profoundly influence the interaction between a celland its environment. Indeed, a major function of S layers isthought to be that of providing physical protection from lyticagents including serum proteins (35) and bacteriophages(19). However, there is increasing evidence that proteins canbe multifunctional rather than merely inert barriers. Themost direct evidence for multifunctionality is seen with theS-layer protein of the fish pathogen Aeromonas salmonicida,in which the protein subunits have specific binding sites forporphyrins (25) and immunoglobulins (B. M. Phipps andW. W. Kay, Abstr. Annu. Meet. Am. Soc. Microbiol. 1986,K225, p. 231).
Electron-microscopic studies of S layers have provideddetailed morphological descriptions of subunit arrangementwithin layers, and the recent application of computer-as-sisted image enhancement techniques has provided two- andthree-dimensional models of subunit organization (43). Un-fortunately, the biochemistry of S-layer subunits has re-ceived much less attention than the morphology, in partbecause of the widely held notion that S layers are nothingmore than crude sieves or barriers, and also because ofproblems associated with protein purification and stabilityafter removal of the layer from the cell (26). Recently,however, there has been an increase in interest in thebiochemistry of S-layer subunit proteins which should allowa much better understanding of the structure and biologicalroles of these proteins.S layers are normally composed of a single species of
protein or glycoprotein subunit arranged with hexagonal,tetragonal, or even linear symmetry (44). The subunits range
* Corresponding author.
in molecular weight (Mr) from 30,000 to 200,000 and appearto contain information for layer assembly. Amino acidcomposition data are available for many of the S-layerproteins, but N-terminal amino acid sequence data areavailable only for the S-layer proteins from three strains ofA. salmonicida. These share extensive homology up toresidue 25 (24), but it is not known whether S layers fromdifferent species share any primary sequence homology.We have been studying a group of A. hydrophila strains
that demonstrate enhanced virulence for salmonid fish (34).The members of this group are characterized by unique cellsurface properties including autoaggregation in static brothculture and resistance to the bactericidal activity of normalserum, and they all belong to a single, heat-stable serogroup(30, 34). Recently this phenotype has also been correlatedwith an increased incidence of invasive disease amonghuman A. hydrophila isolates (20). Our previous studieshave demonstrated the presence of a tetragonal S layer on allmembers of the high-virulence group (36). We have identi-fied the S-layer subunit protein from one particular strain ofA. hydrophila (6), and in this paper we report on thepurification and biochemical properties of that protein.
MATERIALS AND METHODSBacterial strains and growth conditions. A. hydrophila
TF7, a member of the previously described A. hydrophilahigh-virulence group (34), was supplied by R. Lallier, Uni-versity of Montreal, Saint Hyacinthe, Quebec, Canada. A.salmonicida A449 was supplied by C. Michel, Institut Na-tional de la Recherche Agronomique, Thiveral-Grignon,France, and the isolation of the S-layer-deficient mutant wasreported earlier (19). Escherichia coli E1049-a-13 (CFA/I+)was a gift from T. Wadstrom, University of Lund, Lund,Sweden. Stock cultures were maintained at -70°C in 15%(vol/vol) glycerol Trypticase soy broth (BBL MicrobiologySystems, Cockeysville, Md.). Cultures were streaked at
2631
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

2632 DOOLEY ET AL.
-70°C onto tryptic soy agar (Difco Laboratories, Detroit,Mich.). A. hydrophila TF7 was grown for 18 h at 37°C, andA. salmonicida strains were grown for 36 h at 20°C. E. coliwas grown for 24 h at 37°C on CFA medium (11).
Isolation of outer membranes. A. hydrophila cells weregrown for 18 h on tryptone soy agar. After being harvested,the cells were washed once in cold 20 mM Tris buffer (pH8.0). One gram (wet weight) of washed cells was resus-pended in 10 ml of 20 mM Tris buffer (pH 8.0). DNase andRNase A (Sigma Chemical Co., St Louis, Mo.) were added,and the cell suspension was then passed through a Frenchpressure cell three times at 16,000 lb/in2. Intact cells wereremoved by two centrifugations at 6,000 x g at 4°C for 30min. Envelopes were collected by centrifugation at 40,000 xg at 4°C for 30 min. The envelopes were suspended in the 20mM Tris buffer (pH 8.0) to give a protein concentration of 1mglml, and outer membranes were prepared by differentialsolubilization of the inner membrane with the detergentsodium N-lauroyl sarcosinate (SLS; W. R. Grace & Co.,Nashua, N.H.) as described by Filip et al. (14).
Electrophoresis. Sodium dodecyl sulfate-polyacrylamidegel electrophoresis (SDS-PAGE) separation of proteins wasperformed by using the discontinuous buffer system ofLaemmli (29) as modified by Ames (1). Samples werestacked in 4.5% (wt/vol) acrylamide and separated in 12.5%(wt/vol) acrylamide. Separating gels of 7.5% (wt/vol) and15% (wt/vol) were also used. Electrophoresis was performedwith a mini-slab gel apparatus (Bio-Rad Laboratories, Rich-mond, Calif.) at 100 V (constant voltage) initially and 200 Vwhen the tracking dye entered the separating gel. Outermembrane protein was applied at S to 10 ,ug per lane.Molecular weights were determined from a plot of thelogarithm of the molecular weight by using SDS-PAGEmolecular weight standards (Bio-Rad). Nondenaturing iso-electric focusing tube gels were run as specified in theBio-Rad manual. Denaturing isoelectric focusing gels wererun in the presence of 8 M urea and 2% Nonidet P-40 in thesystem of O'Farrell (37).
Staining of gels. Proteins and peptides were visualized bystaining with Coomassie brilliant blue R250 (Sigma). Carbo-hydrate was stained by the periodic acid-Schiff procedure(12), and lipopolysaccharide was detected by the method ofTsai and Frasch (49).
Salt aggregation test. The salt aggregation test was per-formed by the method of Lindahl et al. (31). Bacteria weresuspended in 2 mM phosphate buffer (pH 6.8) to a concen-tration of approximately 5 x 109 cells per ml. A 4 M(NH4)2SO4 solution was made up in the phosphate buffer andserially diluted. Equal volumes of bacterial cell suspensionand salt solution were mixed on a glass slide, and thepresence or absence of aggregates was determined after 2min at room temperature. The lowest (NH4)2S04 concentra-tion causing bacterial aggregation was recorded.Chemical determinations. Protein was determined by the
modified Lowry procedure of Markwell et al. (32). Totalcarbohydrate (neutral sugars) was determined by the phenol-sulfuric acid procedure of Dubois et al. (8). 2-Keto-3-deoxy-octulosonic acid was assayed by the thiobarbituric acidmethod of Osborn et al. (39).
Glycine extraction. Glycine extraction was performed by amodification of the method of McCoy et al. (33). After 18 hof incubation, cultures were harvested into 20 mM Trishydrochloride (pH 8.0), washed three times, and then sus-pended in 0.2 M glycine hydrochloride (pH 3.0) (3 g of cellsper 100 ml). The suspension was stirred for 15 min at 4°C,and whole cells were removed by centrifugation at 12,000 x
g for 15 min. The supernatant was neutralized with NaOH,dialyzed against distilled water, and lyophilized.Hydrophobic interaction chromatography. Purified S-layer
protein was made up to a concentration of 1 mg/ml in 0.02M sodium phosphate buffer (pH 7.0) containing 35%(NH42SO4, and 5 ml of this solution was applied to a column(40 by 1 cm) of octyl-Sepharose (Pharmacia, Uppsala, Swe-den) that had been equilibrated with the same buffer. Thecolumn was washed with 2 bed volumes of buffer, and theprotein was eluted at room temperature with a 300-mlgradient of 35 to 0% (NH4)2SO4 in phosphate buffer at a flowrate of 20 ml/h. Fractions (5 ml) were collected, and proteinwas detected by monitoring the effluent at 280 nm. The52,000-molecular-weight S-layer protein (the 52K protein)was identified by SDS-PAGE.
Purification of the S-layer protein. Five grams (wet weight)of cells grown on tryptone soy agar for 18 h at 37°C wereused for each batch of purified protein. The cells wereharvested, washed three times in 20 mM Tris hydrochloride(pH 8.0), and collected by centrifugation at 12,000 x g for 20min at 4°C. The cells were suspended in 0.2 M glycine buffer(pH 3.0) at 3 g of cells per 100 ml of buffer. The suspensionwas stirred at 4°C for 15 min. The cells were removed bycentrifugation at 12,000 x g for 20 min. The supernatant,containing the extracted S-layer protein, was passed througha nitrocellulose filter (pore size, 0.45 pim) to remove anyremaining cells. It was then neutralized and concentrated toapproximately 30 ml by ultrafiltration through a YM 30membrane (Amicon Corp., Lexington, Mass.) operating at25 lb/in2. The extract was desalted on a Sephadex G-25column equilibrated with 20 mM Tris hydrochloride (pH8.0). All the protein eluted just after the void volume of thecolumn. The protein-containing fractions were pooled andloaded onto a DEAE-Sepharose CL-6B column (bed vol-ume, 40 ml) equilibrated with 20 mM Tris hydrochloride (pH8.0). The column was washed with 3 bed volumes of equil-ibration buffer and eluted with a linear gradient of 500 ml of0 to 0.4 M NaCl in the same buffer at a flow rate of 20 ml/h.The major peak, which eluted around 0.1 M NaCl, containedthe 52K S-layer protein. The fractions constituting this peakwere pooled and concentrated to 1 mg/ml by ultrafiltration.All these chromatography and concentration procedureswere carried out at 4°C. Concentrated protein samples weremade up to 0.1 mM phenylmethylsulfonyl fluoride. Oneother purification step was found to be necessary for N-terminal amino acid sequence analysis. The protein from theion-exchange column was applied to an RP-8 Spheri-10reverse-phase high-performance liquid chromatography col-umn (Brownlee Laboratories Inc., Santa Clara, Calif.) andeluted with a solvent gradient system of 0 to 60% acetonitrile(in 0.1% trifluoroacetic acid).Amino acid composition analysis. The purified protein was
dialyzed against distilled water and lyophilized. It was thenhydrolyzed under vacuum in 6 N HCl at 100°C for 18 h. Thehydrolysates were analyzed on a 11BBL amino acid analyzer(Beckman Instruments, Inc., Palo Alto, Calif.) by standardprocedures. Cysteine was analyzed as cysteic acid, andmethionine was analyzed as methionine sulfone followingperformic acid oxidation as outlined by Hirs (16). Trypto-phan was determined by the spectrophotometric procedureof Edelhoch (9, 15) and also by the method of Penke et al.(40),whichinvolvedhydrolysisoftheproteinwithmercaptoeth-anesulfonic acid.
N-terminal sequence analysis. Protein (5 nmol) was appliedto a model 470A gas phase sequencer (Applied Biosystems,Inc., Foster City, Calif.) running a standard operating pro-
J. BACTERIOL.
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
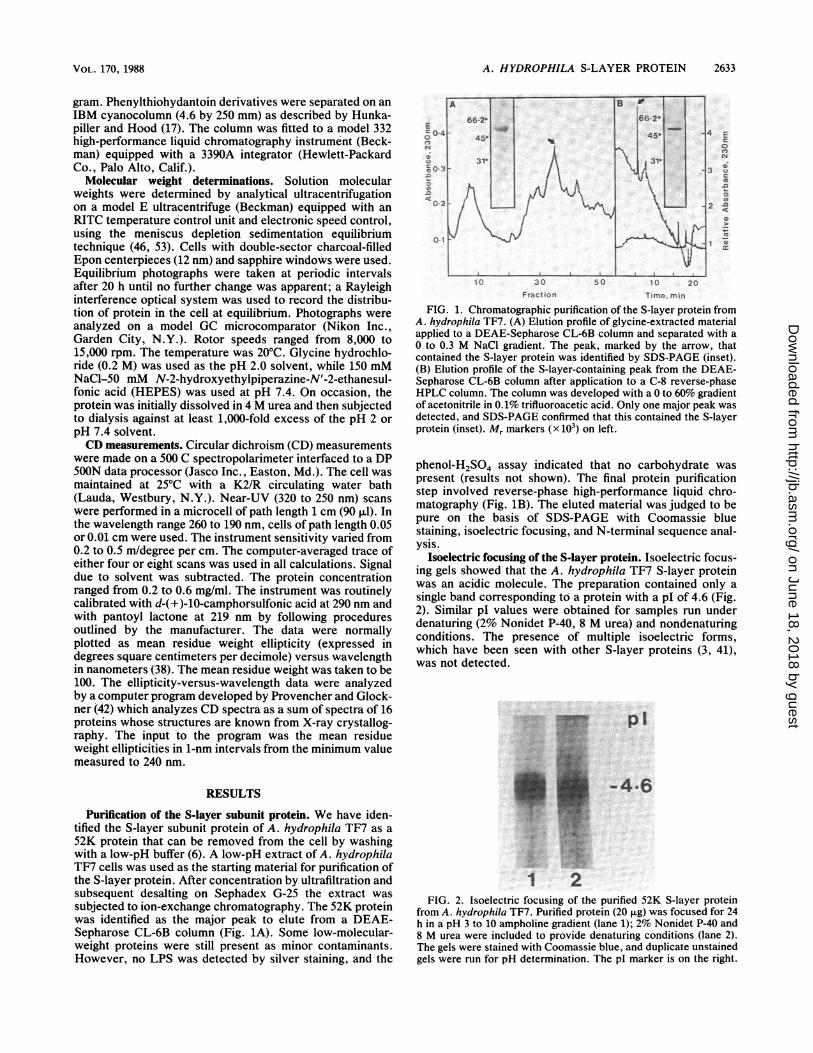
A. HYDROPHILA S-LAYER PROTEIN 2633
gram. Phenylthiohydantoin derivatives were separated on anIBM cyanocolumn (4.6 by 250 mm) as described by Hunka-piller and Hood (17). The column was fitted to a model 332high-performance liquid chromatography instrument (Beck-man) equipped with a 3390A integrator (Hewlett-PackardCo., Palo Alto, Calif.).
Molecular weight determinations. Solution molecularweights were determined by analytical ultracentrifugationon a model E ultracentrifuge (Beckman) equipped with anRITC temperature control unit and electronic speed control,using the meniscus depletion sedimentation equilibriumtechnique (46, 53). Cells with double-sector charcoal-filledEpon centerpieces (12 nm) and sapphire windows were used.Equilibrium photographs were taken at periodic intervalsafter 20 h until no further change was apparent; a Rayleighinterference optical system was used to record the distribu-tion of protein in the cell at equilibrium. Photographs wereanalyzed on a model GC microcomparator (Nikon Inc.,Garden City, N.Y.). Rotor speeds ranged from 8,000 to15,000 rpm. The temperature was 20°C. Glycine hydrochlo-ride (0.2 M) was used as the pH 2.0 solvent, while 150 mMNaCl-50 mM N-2-hydroxyethylpiperazine-N'-2-ethanesul-fonic acid (HEPES) was used at pH 7.4. On occasion, theprotein was initially dissolved in 4 M urea and then subjectedto dialysis against at least 1,000-fold excess of the pH 2 orpH 7.4 solvent.CD measurements. Circular dichroism (CD) measurements
were made on a 500 C spectropolarimeter interfaced to a DP500N data processor (Jasco Inc., Easton, Md.). The cell wasmaintained at 25°C with a K2/R circulating water bath(Lauda, Westbury, N.Y.). Near-UV (320 to 250 nm) scanswere performed in a microcell of path length 1 cm (90 ,ul). Inthe wavelength range 260 to 190 nm, cells of path length 0.05or 0.01 cm were used. The instrument sensitivity varied from0.2 to 0.5 m/degree per cm. The computer-averaged trace ofeither four or eight scans was used in all calculations. Signaldue to solvent was subtracted. The protein concentrationranged from 0.2 to 0.6 mg/ml. The instrument was routinelycalibrated with d-(+)-10-camphorsulfonic acid at 290 nm andwith pantoyl lactone at 219 nm by following proceduresoutlined by the manufacturer. The data were normallyplotted as mean residue weight ellipticity (expressed indegrees square centimeters per decimole) versus wavelengthin nanometers (38). The mean residue weight was taken to be100. The ellipticity-versus-wavelength data were analyzedby a computer program developed by Provencher and Glock-ner (42) which analyzes CD spectra as a sum of spectra of 16proteins whose structures are known from X-ray crystallog-raphy. The input to the program was the mean residueweight ellipticities in 1-nm intervals from the minimum valuemeasured to 240 nm.
RESULTS
Purffication of the S-layer subunit protein. We have iden-tified the S-layer subunit protein of A. hydrophila TF7 as a52K protein that can be removed from the cell by washingwith a low-pH buffer (6). A low-pH extract of A. hydrophilaTF7 cells was used as the starting material for purification ofthe S-layer protein. After concentration by ultrafiltration andsubsequent desalting on Sephadex G-25 the extract wassubjected to ion-exchange chromatography. The 52K proteinwas identified as the major peak to elute from a DEAE-Sepharose CL-6B column (Fig. 1A). Some low-molecular-weight proteins were still present as minor contaminants.However, no LPS was detected by silver staining, and the
.0~~~~~~~~~~~~
10 30 50 1 20Fraction Time. man
FIG. 1. Chromatographic purification of the S-layer protein fromA. hydrophila TF7. (A) Elution profile of glycine-extracted materialapplied to a DEAE-Sepharose CL-6B column and separated with a0 to 0.3 M NaCi gradient. The peak, marked by the arrow, thatcontained the S-layer protein was identified by SDS-PAGE (inset).(B) Elution profile of the S-layer-containing peak from the DEAE-Sepharose CL-6B column after application to a C-8 reverse-phaseHPLC column. The column was developed with a 0 to 60%o gradientof acetonitrile in 0.1% trifluoroacetic acid. Only one major peak wasdetected, and SDS-PAGE confirmed that this contained the S-layerprotein (inset). Mr markers (x 103) on left.
phenol-H2504 assay indicated that no carbohydrate waspresent (results not shown). The final protein purificationstep involved reverse-phase high-performance liquid chro-matography (Fig. iB). The eluted material was judged to bepure on the basis of SDS-PAGE with Coomassie bluestaining, isoelectric focusing, and N-terminal sequence anal-ySiS.
Isoelectric focusing of the S-layer protein. Isoelectric focus-ing gels showed that the A. hydrophila TF7 S-layer proteinwas an acidic molecule. The preparation contained only asingle band corresponding to a protein with a pI of 4.6 (Fig.2). Similar pI values were obtained for samples run underdenaturing (2% Nonidet P-40, 8 M urea) and nondenaturingconditions. The presence of multiple isoelectric forms,which have been seen with other S-layer proteins (3, 41),was not detected.
0 0 -4Z6
0-2122
FIG. 2. Isoelectric focusing of the purified 52K S-layer proteinfrom A. hydrophila TF7. Purified protein (20geg)was focused for 24h in a pH 3 to 10 ampholine gradient (lane 1);2%k Nonidet P-40 and8 M urea were included to provide denaturing conditions (lane 2).The gels were stained with Coomassie blue, and duplicate unstainedgels were run for pH determination. The pI marker is on the right.
VOL. 170, 1988
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

2634 DOOLEY ET AL.
TABLE 1. Amino acid composition of tetragonalS-layer proteins
Mol % in:
Amino acid A. hydrophila A. salmonicidab AzotobacterTF7 A450 vinelandii'
Asx 15.4 14.9 10.9Thr 7.1 8.8 9.5Ser 6.8 3.9 9.5Glx 8.9 9.5 8.0Pro 2.4 2.3 2.9Gly 8.3 4.7 8.0Ala 12.7 8.0 13.3Val 7.5 8.4 8.3Met 1.5 1.0 0.7Ile 2.8 5.1 5.0Leu 8.8 10.1 8.5Tyr 3.2 1.9 2.4Phe 4.4 7.3 5.4His 0.6 1.1 0.0Lys 8.5 6.1 4.9Arg 2.1 4.3 2.1Trp 0.7 1.8 0.5Cys 0.0 0.0 0.0
Total no. of residues 520 473 577per mol
Apparent Mr 52,500 49,200 60,200Hydrophobic residuesa 41% 45% 44.6%
a V, M, I, L, A, F, W, and P.b Kay et al. (24).1 Bingle et al. (3).
Amino acid composition and NH2-terminal sequence analy-sis. Amino acid composition analysis of the purified A.hydrophila TF7 S-layer subunit protein showed that it con-
tained 520 residues per molecuie (Table 1), 41% of whichwere hydrophobic amino acid residues. No cysteic acid wasdetected following performic acid oxidation. The amino acidcomposition and subunit molecular weight of the A. hydro-phila TF7 protein were similar to that of the layer protein ofthe related species A. salmonicida (23, 47) and the tetragonalarray-forming protein ofAzotobacter vinelandii (3) (Table 1).However, when the N-ternminal amino acid sequences of theAeromonas S-layer proteins were compared, no homologywas detected over the first 30 residue positions, even thoughsimilar numbers of hydrophobic and charged residues were
seen in this region (Table 2).S-iayer hydrophobicity. The A. hydrophila TF7 S-layer
protein displayed hydrophobic properties upon extractionfrom the cell. The protein bound to octyl-Sepharose underhigh-ionic-strength conditions and could be eluted only withlow-salt buffers (Fig. 3). However, when present as an intactnative S layer, the S-layer protein did not confer significanthydrophobicity to the cell surface. This was shown by thesalt aggregation test, in which A. hydrophila TF7 cells salted
6621_
451
-'1- S
t ... vS /l
\f \
r j X_; l'
tw - - -_FIG. 3. Analysis of the hydrophobicity of the purified A. hydro-
phila TF7 S-layer protein by hydrophobic interaction chromatogra-phy. The S-layer protein was eluted from an octyl-Sepharosecolumn under conditions of low ionic strength and in the absense of(NH4)2SO4. The protein was identified by SDS-PAGE stained withCoomassie blue (inset). Mr markers (x 103) on left.
out under the same high ionic strength conditions as strainsthat do not possess S layers [>1 M (NH4)2SO4]. In contrast,cells of A. salmonicida that possessed an S layer were
significantly more hydrophobic [0.032 M (NH4)2SO4] thanisogenic mutants that were deficient in the layer [>1 M(NH4)2SO4]. The control CFA/I-producing strain of E. colialso aggregated under low-ionic-strength conditions [0.016M (NH4)2SO4] in keeping with the increased surface hydro-phobicity conferred on the cell by the presence of the CFA/Ifimbriae (13).
Secondary structure of the S-layer protein. CD measure-ments were made to estimate the amount of secondarystructure in the purified S-layer subunit under various exper-imental conditions (Fig. 4). These data are summarized inTable 3. The A. hydrophila TF7 S-layer protein had a limitedamount of a-helix (approximately 19%) at either pH 7.4 or
pH 2 (Fig. 4A and C). The protein had a large amount ofp-pleated sheet (approximately 44%), again constant at bothpH values. The large amount of P-pleated sheet was indi-cated by the single trough at 215 nm. This indicated that no
major conformational changes took place at low pH, imply-ing that the protein was not drastically altered by the low-pHprocedure used to remove it from the cell surface. Asignificant number of p-turns were also present in thesecondary structure.
It has been found that acidic solutions of SDS can be muchmore potent than neutral ones in ordering structure in certainpolypeptides and proteins (52). This is because anionicglutamic and aspartic acid residues can destabilize the in-duced conformation if they are located in the structure-forming segments of the polypeptide chain. Lowering the pHprotonates these residues and removes this disruptive effect.This situation was obtained with the A. hydrophila TF7S-layer protein, since at pH 2 in 0.1% SDS (Fig. 4C) theo-helix content increased from 18% to 42%, with a concom-
itant large decrease in the p-sheet content, as revealed by the
TABLE 2. N-terminal amino acid sequences of S-layer proteins
Species Sequence
A. hydrophila TF7 VNLDTGAGVS FKASGIKVDG AAGTTLGGXA GGA. salmonicida A450a DVVIGPNDNT FVTNSLASVT EQPVLB. brevis 47b APKDGIYIGG NIKKYYSYDV FEMTPQAKAT YA
a Phipps et al. (41).b Tsuboi et al. (50).
J. BACTERIOL.
:4 0 e.:,lkii
..1
iI
Tz
tt
, :1
."; t-
Z. r: e.-
.7-
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

A. HYDROPHILA S-LAYER PROTEIN 2635
200 220 240
I
255 295
O.E
0 -
-30 t
I
-60
zuu 220 24C
WAVELENGTH (nmn)
260 300
200 220 240
WAVELENGTH (nm)
FIG. 4. CD spectra of the S-layer protein from A. hydrophila TF7. S-layer protein dissolved in 50 mM HEPES-150 mM NaCl (pH 7.4) or0.2 M glycine hydrochloride (pH 2.0). (A) Native protein at pH 7.4; (B) native protein plus 0.1% (wt/vol) SDS (pH 7.4); C, native protein (----)and native protein plus 0.1% SDS (-) (pH 2.0); D, native protein (-) and native protein plus 0.1% SDS (----) (pH 7.4); E, native protein(-) and native protein plus 0.1% SDS (----) (pH 2.0).
increase in negative ellipticity and the shift of the singletrough to the bimodal appearance characteristic of a-helicalstructure. On the other hand, at pH 7.4 in the presence of0.1% SDS (Fig. 4B), there was a reduction from 19% to 11%in the amount of ot-helix, with an extensive increase in theamount of p-sheet.The lack of dependence of the A. hydrophila TF7 S layer
on the presence of divalent cations for maintenance of layerintegrity was also confirmed by CD measurements of thelayer in the presence and absence of EDTA. Since EDTAhas no metal-chelating activity at pH 2, the measurementswere taken at pH 7.4. No difference in a-helical content wasnoted either with or without the chelator, but there was anincrease in the amount of 3-sheet in the presence of EDTA,at the expense of less periodic structure (results not shown).Near-UV CD were also measured, since this is the spec-
tral region where contributions are found from asymmetri-
TABLE 3. Far-UV CD data for the A. hydrophilaTF7 S-layer protein
Secondary structure content (%)Solvent
t P Reanr
Helix Sheet Turn Remainder
0.5 M HEPES-.15 M NaCl(pH 7.4)
Native (196-240 nm) 19 43 12 27Native (after solubilization 20 41 9 30
in 4 M urea)Native + SDS (0.1%) 11 68 0 22Native (240-207 nm) 13 55 0 32Native + EDTA (240- 11 79 3 8
207 nm)
0.2 M Glycine-HCl (pH 2)Native (196-240 nm) 16 44 9 30Native (after solubilization 18 44 18 28
in 4 M urea)Native + SDS (0.1%) 42 24 0 34
cally situated aromatic amino acids. The spectra at pH 7.4(Fig. 4D) showed that the TF7 S-layer protein has somedefinite fine structure in this region, with a small peak near292 nm probably due to a tryptophan contribution, the broadtrough near 282 nm representing a tyrosine contribution, andthe sawtooth structure from 270 to 255 nm presumablyarising from phenylalanine residues. The spectrum wassensitive to the presence of 0.1% SDS, but the detergent didnot abolish the fine structure. At pH 2 (Fig. 4E) the signalwas very much reduced, and 0.1% SDS again had essentiallyno effect (Table 3).
Sedimentation analysis was then attempted to estimate themolecular weight of the S-layer protein in solution, at bothpH 2 and pH 7.4. No value for the subunit Mr could beobtained owing to aggregation of the protein under all theconditions tested. At pH 2.0 the protein behaved as a veryhigh-Mr complex with an average Mr of 1.19 x 106. How-ever, less aggregation was evident when the sample wasinitially solubilized in 4 M urea, and an Mr of 408,000 wasobtained. Aggregation was also evident at pH 7.4, with theobservation of an Mr 142,700 species. Under the conditionsof sedimentation (48 h at room temperature), some break-down of the 52K parent was noted. At pH 7.4, lower-molecular-weight fragments could be distinguished, espe-cially in the sample exposed to 4 M urea. A species of Mr29,700 was the most common protein observed. When thismaterial was analyzed by SDS-PAGE, it was found thatproteins of apparent Mr 52,000 and 30,000 were present.SDS-PAGE analysis of the pH 2.0 material gave only the Mr52,000 band (data not shown).
DISCUSSION
Most attempts at purification of S-layer proteins haveinvolved the disruption of the layer by treatment with highconcentrations of chaotropic agents (26, 44). In many casesthe release of S-layer protein in relatively high concentra-tions, and substantially free from other cell components, hasalso required the initial isolation of an envelope fraction. In
A
0 -
-8.
U e)0
j E
0X
_vf Ev:
z 1-P-
0.O
-i E
E
z
4
0
-8
-30
60
D
4Ir1I
VOL. 170, 1988
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

2636 DOOLEY ET AL.
this study we circumvented this step by exploiting the abilityof low pH to disrupt the A. hydrophila TF7 S-layer andrelease it from whole cells with a minimum of contaminants.The glycine extraction is a modification of the procedure ofMcCoy et al. (33), who used it to remove the antiphagocyticmicrocapsular protein from the surface of the animal patho-gen Campylobacter fetus. Importantly, no difference in thesecondary structure content of the purified A. hydrophilaTF7 S-layer protein was found at low or neutral pH, indi-cating that the low-pH treatment did not produce anymeasurable conformational change in the S-layer proteinduring its removal from the cell surface. Indeed, we havefound that low-pH extraction of whole cells is a very usefulmethod of removing S layers in relatively pure form from anumber of gram-negative species, including Aeromonas,Campylobacter, and Pseudomonas species.The A. hydrophila TF7 S layer is morphologically very
similar to the S layer of Azotobacter vinelandii and is alsosimilar to it in overall amino acid composition and subunitmolecular weight (3). Unlike the A. hydrophila layer, how-ever, the Azotobacter vinelandii S layer easily washes off thecell with distilled water, being released in the form oftetramers. The tetramers are capable of self-assembly toform an intact layer in the presence of Ca2" (7). The A.hydrophila TF7 layer differs from that of Azotobacter vine-landii in that it shows no apparent divalent cation involve-ment in assembly, and, indeed, the protein subunits aggre-gate very strongly in aqueous solution in the absence ofdivalent cations. Inclusion of 4 M urea reduced but did noteliminate all the aggregation of the protein subunits. Unfor-tunately, this aggregation in solution made an accuratedetermination of subunit Mr impossible. The aggregation isin accord with the CD finding that the protein has a great dealof 1-structure in its secondary folding, since such systemsoften show a high propensity for aggregation (18, 45). Thesubunit Mr 50,000 tetragonal array-forming protein of A.salmonicida also exhibited some aggregation during sedi-mentation analysis, with 30 to 40% of the material beingpresent as aggregates of Mr > 110,000 (41). The greaterdegree of aggregation exhibited by the A. hydrophila S layerprobably reflects the higher 13-sheet content of this molecule,which is 43% as compared with 28% for the A. salmonicidamolecule. The ,3-sheet content of the A. hydrophila protein isalso significantly higher than the 35% ,B-sheet content of theAzotobacter vinelandii S-layer protein and the 35% 13-sheetcontent (based on infrared measurements) of the S-layerprotein of Deinococcus radiodurans (2). Not one of thesestudies substantiates the prediction of Sletyr and Messner(44), based on amino acid composition data (27), that Slayers would have about 20% 1-sheet structure, and a high(up to 40%) content of a-helix.The a-helix content of the A. hydrophila protein was
measured at approximately 19%, which is much lower thanthe Sleytr and Messner (44) prediction. In the presence ofthe detergent SDS at low pH, a significant enhancement ofa-helix content was noted, from 18% to 42%, which is muchcloser to their prediction. This had also been seen for the A.salmonicida S-layer protein. Given that the hydrophobicenvironment provided by the detergent can alter the second-ary structure content of the protein so dramatically, it ispossible that the molecule has a different conformation whenit is within the native S layer, where hydrophobic interac-tions are thought to be the major forces stabilizing the layer.The overall amino acid composition of the A. hydrophila
TF7 S-layer protein is consistent with that of other S-layerproteins examined to date (10, 44). For example, the absence
of cysteine is characteristic of S-layer proteins, just as it isfor a large number of other bacterial cell surface andexported proteins (44). The hydrophobic amino acid contentof the A. hydrophila protein was lower than those of the A.salmonicida and Azotobacter vinelandii proteins. The puri-fied A. hydrophila S-layer protein did bind strongly tooctyl-Sepharose under conditions of high ionic strength.However, when present as an intact native S layer on thesurface of the cell, the molecule did not appear to contributeto overall cell surface hydrophobicity, as determined by thesalt aggregation test. This contrasts with A. salmonicida, onwhich the assembled S layer does confer significant cellsurface hydrophobicity (48, 51), and this increased hydro-phobicity is thought to enhance the ability of A. salmonicidato associate with macrophages (48). In this regard it isinteresting that S-layer-producing strains ofA. hydrophila donot show any enhanced ability to associate with macro-phages (D. H. Speert and T. J. Trust, unpublished data). Thedifferences in surface hydrophobicity between A. hydrophilaand A. salmonicida appear to be due to differences in theS-layer proteins, since both species have chemically similarlipopolysaccharide O-polysaccharide chains protrudingthrough the S layer (4, 5). This points to differences in thesurface-exposed domains of the S-layer proteins of A. hy-drophila and A. salmonicida. Further evidence for differ-ences in surface structure is the inability of the A. hydrophilaS layer to bind hemin, Congo red, or immunoglobulins, aswell as a lack of immunological cross-reactivity with the A.salmonicida protein by immunofluorescence assay (J. S. G.Dooley and T. J. Trust, unpublished data). The differencesprobably reflect significant differences in the in vivo func-tioning of the respective S layers.The N-terminal amino acid sequence of the A. hydrophila
S-layer protein also displayed virtually no identity with theN-terminal sequence of the A. salmonicida S-layer proteinor with the outer wall protein of Bacillus brevis 47 or theN-terminal sequence of the S layer of C. fetus (D. Dubreuilet al., unpublished data). The lack of homology with the A.salmonicida protein is especially surprising given the geneticrelatness of the two species, the similarity in Mr of thesubunit proteins, and the similar morphology of the assem-bled arrays. Certainly, the subunit proteins of flagella andthe fimbriae produced by members of the family Enterobac-teriaceae and Neisseria, Pseudomonas, and Moraxella spp.which, like S layers, represent exported surface-assembledprotein structures, exhibit considerable N-terminal sequencehomology (21, 22). In these cases, this sequence homology isinterpreted to imply the existence of a single ancestral gene.The limited data currently available suggest that this is notthe case with S-layer proteins.
ACKNOWLEDGMENTSThis study was supported in part by a grant to T.J.T. from the
British Columbia Health Care Foundation and by grants to C.M.K.from the Medical Research Council of Canada and the AlbertaHeritage Foundation for Medical Research. J.S.G.D. was the recip-ient of a University of Victoria graduate student award.The excellent technical assistance of Kimio Oikawa in connection
with the CD measurements is acknowledged.
LITERATURE CITED1. Amies, G. F. 1974. Resolution of bacterial proteins by polyacryl-
amide gel electrophoresis on slabs. Membrane soluble andperiplasmic fractions. J. Biol. Chem. 249:634 644.
2. Baumeister, W., F. Karrenberg, R. Rachel, A. Engel, B. TenHeggler, and W. 0. Saxton. 1982. The major cell envelopeprotein of Micrococcus radiodiurans (R1): structural and chem-
J. BACTERIOL.
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

A. HYDROPHILA S-LAYER PROTEIN 2637
ical characterization. Eur. J. Biochem. 125:535-544.3. Bingle, W. H., J. L. Doran, and W. J. Page. 1986. Characteri-
zation of the surface layer protein from Azotobacter vinelandii.Can. J. Microbiol. 32:112-120.
4. Chart, H., D. H. Shaw, E. E. Ishiguro, and T. J. Trust. 1984.Structural and immunochemical homogeneity of Aeromonassalmonicida lipopolysaccharide. J. Bacteriol. 158:16-22.
5. Dooley, J. S. G., R. Lallier, D. H. Shaw, and T. J. Trust. 1985.Electrophoretic and immunochemical analyses of the lipopoly-saccharides from various strains of Aeromonas hydrophila. J.Bacteriol. 164:263-269.
6. Dooley, J. S. G., and T. J. Trust. 1988. Surface protein compo-sition of Aeromonas hydrophila strains virulent for fish: identi-fication of a surface array protein. J. Bacteriol. 170:499-506.
7. Doran, J. L., W. H. Bingle, and W. J. Page. 1987. Role ofcalcium in assembly of the Azotobacter vinelandii surface array.J. Gen. Microbiol. 133:399-413.
8. Dubois, M., K. A. Gillies, J. K. Hamilton, P. A. Rebers, and F.Smith. 1956. Colorimetric method for determination of sugarsand related substances. Anal. Chem. 28:350-356.
9. Edelhoch, H. 1967. Spectroscopic determination of tryptophanand tyrosine in proteins. Biochemistry 6:1948-1954.
10. Engelhardt, H., W. 0. Saxton, and W. Baumeister. 1986. Three-dimensional structure of the tetragonal surface layer of Sporo-sarcina ureae. J. Bacteriol. 168:309-317.
11. Evans, D. G., D. J. J. R. Evans, and W. Toja. 1977. Hemagglu-tination of human group A erythrocytes of enterotoxigenicEscherichia coli isolated from adults with diarrhea: correlationwith colonization factor. Infect. Immun. 18:330-337.
12. Fairbanks, G., T. L. Steck, and D. F. H. Wallach. 1971.Electrophoretic analysis of the major polypeptides of the humanerythrocyte membrane. Biochemistry 10:2606-2617.
13. Faris, A., T. Wadstrom, and J. H. Freer. 1981. Hydrophobicadsorptive and hemagglutinating properties of Escherichia colipossessing colonization factor antigens (CFA/I or CFA/II), type1 pili, or other pili. Curr. Microbiol. 5:67-72.
14. Filip, C., G. Fletcher, J. L. Wulff, and C. F. Earhart. 1973.Solubilization of the cytoplasmic membrane of Escherichia coliby the ionic detergent sodium-lauryl sarcosinate. J. Bacteriol.115:717-722.
15. Goodwin, T. W., and R. A. Morton. 1946. The spectrophoto-metric determination of tyrosine and tryptophan in proteins.Biochem. J. 40:628-632.
16. Hirs, C. H. W. 1965. The oxidation of ribonuclease withperformic acid. J. Biol. Chem. 219:611-621.
17. Hunkapiller, M. W., and L. E. Hood. 1983. Analysis of phenyl-thiohydantoins by ultrasensitive gradient high-performance liq-uid chromatography. Methods Enzymol. 91:486-493.
18. Ikigai, H., and T. Nakae. 1985. Conformational alteration inalpha-toxin from Staphylococcus aureus concomitant with thetransformation of the water-soluble monomer to the membraneoligomer. Biochem. Biophys. Res. Commun. 130:175-181.
19. Ishiguro, E. E., W. W. Kay, T. Ainsworth, J. B. Chamberlain,R. A. Austen, J. T. Buckley, and T. J. Trust. 1981. Loss ofvirulence during culture of Aeromonas salmonicida at hightemperature. J. Bacteriol. 148:333-340.
20. Janda, J. M., L. S. Oshiro, S. L. Abbott, and P. S. Duffy. 1987.Virulence markers of mesophilic aeromonads: association of theautoagglutination phenomenon with mouse pathogenicity andthe presence of a peripheral cell-associated layer. Infect. Im-mun. 55:3070-3077.
21. Jones, G. W., and R. E. Isaacson. 1984. Proteinaceous adhesinsand their receptors. Crit. Rev. Microbiol. 10:229-260.
22. Joys, T. M. 1986. The covalent structure of the phase-1 flagellarfilament of Salmonella typhimurium and its comparison withother flagellins. J. Biol. Chem. 260:15758-15761.
23. Kay, W. W., J. T. Buckley, E. E. Ishiguro, B. M. Phipps,J. P. L. Monette, and T. J. Trust. 1981. Purification and dispo-sition of a surface protein associated with virulence of Aeromo-nas salmonicida. J. Bacteriol. 147:1077-1084.
24. Kay, W. W., B. M. Phipps, E. E. Ishiguro, R. W. Olafson, andT. J. Trust. 1984. Surface layer virulence A-proteins fromAeromonas salmonicida strains. Can. J. Biochem. Cell Biol.
62:1064-1071.25. Kay, W. W., B. M. Phipps, E. E. Ishiguro, and T. J. Trust. 1985.
Porphyrin binding by the surface array virulence protein ofAeromonas salmonicida. J. Bacteriol. 164:1332-1336.
26. Koval, S. F., and R. G. E. Murray. 1984. The isolation of surfacearray proteins from bacteria. Can. J. Biochem. Cell Biol. 62:1181-1189.
27. Krigbaum, W. R., and S. P. Knutton. 1973. Prediction of theamount of secondary structure in a globular protein from itsamino acid composition. Proc. Natl. Acad. Sci. USA 70:2809-2813.
28. Kurosaka, R., and S. Horiuchi. 1974. Serological properties andamino acid composition of the fimbriae of Aeromonas hydro-phila. Bull. Tokyo Med. Dent. Univ. 21:7-23.
29. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
30. Lallier, R., and P. Daigneault. 1984. Antigenic differentiation ofpili from non-virulent and fish-pathogenic strains of Aeromonashydrophila. J. Fish Dis. 7:509-512.
31. Lindahl, M., A. Faris, T. Wadstrom, and S. Hjerten. 1981. Anew test based on 'salting out' to measure relative surfacehydrophobicity of bacterial cells. Biochim. Biophys. Acta 677:471-476.
32. Markwell, M. A. K., S. M. Haas, L. L. Biaber, and N. E.Tolbert. 1978. A modification of the Lowry procedure to sim-plify protein determination in membrane and lipoprotein sam-ples. Anal. Biochem. 87:206-210.
33. McCoy, E. C., D. Coyle, K. Burda, L. B. Corbeil, and A. J.Winter. 1975. Superficial antigens of Campylobacter (Vibrio)fetus: characterization of the antiphagocytic component. Infect.Immun. 11:517-525.
34. Mittal, K. R., G. Lalonde, D. Leblanc, G. Olivier, and R. Lallier.1980. Aeromonas hydrophila in rainbow trout: relation betweenvirulence and surface characteristics. Can. J. Microbiol. 26:1501-1503.
35. Munn, C. B., E. E. Ishiguro, W. W. Kay, and T. J. Trust. 1982.Role of surface components in serum resistance of virulentAeromonas salmonicida. Infect. Immun. 36:1069-1075.
36. Murray, R. G. E., J. S. G. Dooley, P. W. Whippey, and T. J.Trust. 1988. Structure of an S layer on a pathogenic strain ofAeromonas salmonicida. J. Bacteriol. 170:2625-2630.
37. O'Farrell, P. H. 1975. High resolution two-dimensional electro-phoresis of proteins. J. Biol. Chem. 250:4007-4021.
38. Oikawa, C., C. M. Kay, and W. D. McCubbin. 1968. Theultraviolet circular dichroism of muscle proteins. Biochem.Biophys. Acta 168:164-167.
39. Osborn, M. J., J. E. Gander, E. Parisi, and J. Carson. 1972.Mechanism of assembly of the outer membrane of Salmonellatyphimurimum. J. Biol. Chem. 247:3962-3972.
40. Penke, B., R. Ternezi, and K. Kovass. 1974. A new acidhydrolysis method for determining tryptophan in peptides andproteins. Anal. Biochem. 60:45-50.
41. Phipps, B. M., T. J. Trust, E. E. Ishiguro, and W. W. Kay. 1983.Purification and characterization of the cell surface virulenceA-protein from Aeromonas salmonicida. Biochemistry 22:2934-2939.
42. Provencher, S. W., and J. Glockner. 1981. Estimation of globu-lar protein secondary structure from circular dichroism. Bio-chemistry 20:33-37.
43. Saxton, W. O., and W. Baumeister. 1986. Principles of organi-sation in S-layers. J. Mol. Biol. 187:251-253.
44. Sletyr, U. B., and P. Messner. 1983. Crystalline surface layers onbacteria. Annu. Rev. Microbiol. 37:311-319.
45. Smith, R. 1985. The encephalitogenic protein of myelin formshexamers in which the polypeptides have a pleated-sheet struc-ture. FEBS Lett. 183:331-334.
46. Teller, D. C. 1973. Characterization of proteins by sedimenta-tion equilibrium in the analytical ultracentrifuge. Methods En-zymol. 27:346-411.
47. Trust, T. J., P. S. Howard, J. B. Chamberlain, E. E. Ishiguro,and J. T. Buckley. 1980. Additional surface protein in autoag-gregating strains of atypical Aeromonas salmonicida. FEMS
VOL. 170, 1988
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

2638 DOOLEY ET AL.
Microbiol. Lett. 9:35-38.48. Trust, T. J., W. W. Kay, and E. E. Ishiguro. 1983. Cell surface
hydrophobicity and macrophage association of Aeromonas sal-monicida. Curr. Microbiol. 9:315-318.
49. Tsai, C. M., and C. E. Frasch. 1982. A sensitive silver stain fordetecting lipopolysaccharide in polyacrylamide gels. Anal. Bio-chem. 119:115-119.
50. Tsuboi, A., R. Uchihi, R. Tabata, Y. Takahashi, H. Hashiba, T.Sasaki, H. Yamagata, N. Tsukagoshi, and S. Udaka. 1986.Characterization of the genes coding for two major cell wallproteins from protein-producing Bacillus brevis 47: completenucleotide sequence of the outer wall protein gene. J. Bacteriol.
168:365-373.51. Van Alstine, J. M., T. J. Trust, and D. E. Brooks. 1986.
Differential partition of virulent Aeromonas salmonicida andattenuated derivatives possessing specific cell surface alter-ations in polymer aqueous-phase systems. Appl. Environ. Mi-crobiol. 51:1309-1313.
52. Wu, C.-S., K. Ikeda, and J. T. Yang. 1981. Ordered conforma-tions of polypeptides and proteins in acidic dodecyl sulfatesolution. Biochemistry 20:566-570.
53. Yahantis, D. A. 1964. Equilibrium ultracentrifugation of dilutesolutions. Biochemistry 3:297-317.
J. BACTERIOL.
on June 18, 2018 by guesthttp://jb.asm
.org/D
ownloaded from