islet amyloid polypeptide in human insulinomas - diabetes
TRANSCRIPT

Islet Amyloid Polypeptide in HumanInsulinomasEvidence for Intracellular AmyloidogenesisTIMOTHY D. O'BRIEN, ALEXANDRA E. BUTLER, PATRICK C. ROCHE, KENNETH H. JOHNSON,AND PETER C. BUTLER
Amyloid deposits that characteristically form in thepancreatic islets of patients with non-insulin-dependent diabetes mellitus (NIDDM) and ininsulinomas are both derived from islet amyloidpolypeptide (IAPP). Evidence from previous studieshas suggested that deposition of IAPP-derivedamyloid is related to inherent amyloidogenicsequences present within normal human IAPP,together with an increased production and localconcentration of IAPP. However, whether theaggregation of IAPP to form amyloid fibrils is primarilyan intra- or extracellular event is not clear. To addressthis question, we studied 20 human insulinomas bylight and electron microscopy. By light microscopy,amyloid deposits were demonstrated in 13 of 20 (65%)human insulinomas. Furthermore, evaluation of Congored-stained tumor sections showed small, globular orirregular, congophilic amyloid deposits within thecytoplasm of many tumor cells in 10 of 13 (77%)amyloid-containing insulinomas. Dense, punctateareas of IAPP immunoreactivity within tumor cellscorresponded with the congophilic intracellulardeposits. Ubiquitin immunoreactivity also wasobserved as punctate intracellular labeling and withinlarge extracellular amyloid deposits. Among the 10insulinomas available for electron microscopicevaluation, pathological lAPP-immunoreactive(immunogold) deposits were found in 3 of 5insulinomas in which amyloid was demonstrated bylight microscopy and in none of 5 tumors foundnegative for amyloid by light microscopy. Morphologyof lAPP-immunoreactive deposits varied from thosewith the classical distinct 7- to 10-nm diameternonbranching fibrils to those with distinct but faint
From the Endocrine Research Unit, Mayo Clinic, Rochester; and the Depart-ment of Veterinary Pathobiology, University of Minnesota, St. Paul, Minnesota.
Address correspondence and reprint requests to Dr. Timothy D. O'Brien,Endocrine Research Unit, Mayo Clinic, Rochester, MN 55905.
Received for publication 10 February 1993 and accepted in revised form23 September 1993.
NIDDM, non-insulin-dependent diabetes mellitus; IAPP, islet amyloidpolypeptide; IA, islet amyloid; TBS-T, tris-buffered saline with 0.05% tween.
fibrillarity to those without discernable fibrils. In eachof the 3 tumors with ultrastructurally demonstrableamyloid deposits, intracellular aggregates oflAPP-immunoreactive amyloid fibrils were observedeither free in the cytoplasmic matrix or inmembrane-bound structures. IntracellularlAPP-immunoreactive deposits were up to 10 urn inthe greatest dimension. Free intracellular aggregatesof amyloid fibrils sometimes contained degeneratingsecretory vesicles and fragments of membranousorganelles. These observations suggest thatlAPP-derived amyloid deposits are initially formedintracellularly and are subsequently released to theextracellular space by exocytosis of themembrane-bound structures and/or after necrosis ofthe tumor cells. Our observations raise the possibilitythat lAPP-derived islet amyloid deposits(characteristically present in NIODM patients) also maybe initially formed within the cytoplasm of p-cells, thusleading to p-cell necrosis and a reduction of p-cellmass as seen in NIDDM. Diabetes 43:329-36,1994
Islet amyloid polypeptide (IAPP) is a 37 amino acidpolypeptide that is produced primarily by the pancre-atic p-cells (1,2,3) and is copackaged (4,5) andcosecreted with insulin (6-10). IAPP is the protein
precursor of the amyloid deposits that are the character-istic lesions of the pancreatic islets in non-insulin-depen-dent diabetes mellitus (NIDDM) (11). IAPP also formsamyloid deposits in human (15-17) and canine (18)pancreatic endocrine tumors, and in fact, a humaninsulinoma provided one source of amyloid from whichIAPP was first identified (1). Because the occurence ofislet amyloid (IA) is closely associated with NIDDM, anunderstanding of the pathogenesis of IA may contributeto the elucidation of the underlying cause and pathogen-esis of NIDDM.
Because IA and the amyloid in insulinomas are bothderived from IAPP, similar factors may possibly play a
DIABETES, VOL. 43, FEBRUARY 1994 329
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/43/2/329/360700/43-2-329.pdf by guest on 14 February 2022

IAPP IN INSULINOMAS
role in amyloidogenesis in both conditions. In this regard,the site (i.e., intra- versus extracellular) of initial IAPP-derived amyloid formation or deposition has not beenclearly established in either pancreatic islets or in pan-creatic endocrine tumors. However, we have demon-strated previously intracellular IA in nondiabetic cats withminimal extracellular IA (12) and also have demonstratedlAPP-immunoreactive amyloid within islet cells of catswith diabetes (4). Others also have observed both intra-cellular and extracellular amyloid in islets of macaqueswith NIDDM (13). It is therefore possible that IA mayinitially form intracellularly in (3-cells and then may bereleased to the extracellular space.
Although it is difficult to obtain fresh human pancreatictissue suitable for electron microscopic studies in NIDDMpatients, surgically excised insulinoma specimens canbe fixed immediately for optimal electron microscopicand immunohistochemical study. Therefore, in this study,we have examined 20 human insulinomas by immuno-histochemistry and electron microscopy to determineboth the frequency of IAPP-derived amyloid deposits inhuman insulinomas and the potential importance of intra-cellular IAPP aggregation and fibrillogenesis in the de-generation and destruction of p-cells.
RESEARCH DESIGN AND METHODSTwenty human insulinomas were obtained from the ar-chives of the Department of Pathology, Mayo Clinic,Rochester, Minnesota. Of the 20 cases, 13 comprised allinsulinoma accessions over the 1 -year period before thebeginning of the study. Samples for light microscopywere available for all 13 cases, whereas samples forelectron microscopy were available in only 3 of thesecases. An additional 7 cases were selected solely on thebasis of having samples available for both light andelectron microscopic evaluation. Age, sex, and tumorlocation for each patient is given in Table 1.
After surgical removal, each tumor was fixed in 10%neutral buffered formalin solution and routinely pro-cessed for paraffin embedding. For each tumor, 4-Limthick serial sections were mounted on chromalum-coatedglass slides, deparaffinized, and stained with hematoxy-lin and eosin, Congo red, and the peroxidase-anti-peroxidase method for insulin (guinea pig anti-insulin1/400, Dako, Santa Barbara, CA), IAPP (anti-human-amylin 1/1000, Peninsula, Belmont, CA), insulin C-pep-tide (1/100, Dako), and ubiquitin (1/500, Dako) asdescribed previously (3). As reported previously (8), theanti-IAPP serum had 100% cross-reactivity with humanIAPP, 0.001% cross-reactivity with human calcitoningene-related peptide, and no measurable cross-reac-tivity with human calcitonin, human insulin, human gluca-gon, human pancreatic polypeptide, porcine pancreast-atin, porcine neuropeptide Y, vasoactive intestinalpeptide, or somatostatin. Negative immunocytochemicalcontrol studies were performed in which the primaryantiserum was replaced with nonimmune serum.
For electron microscopic evaluation, tumor samplesfrom 10 of 20 cases (1,4,10,14-20) were immediatelyminced (1-mm3 pieces), fixed in Trump's fluid, and
Table 1Insulinoma patient data
Case(number)
123456789
1011121314151617181920
Sex
FMMFMFMMFFMFMFFFFFFM
Age(years)
7035687047474354667264244375609
27297444
Tumor location
TailTail and bodyBodyTailTailBodyUncinate processTailHeadTailBodyHeadTailHeadHeadHeadUncinate processHeadBodyTail
Duration(months)
24722
2436
1362448362
152430302424122448
Duration in months of clinical signs related to insulinoma.
postfixed in 1% osmium tetroxide. Samples were thendehydrated in a graded series of ethanol solutions,embedded in Spur's resin, thin sectioned, placed oncopper grids, and stained with lead citrate and uranylacetate. For immunogold labeling, thin sections wereplaced on celloidin membranes, etched with saturatedsodium metaperiodate for 16 min, washed with water,and then incubated in tris-buffered saline with 0.05%tween (TBS-T) and 1% bovine serum albumin. Sectionswere incubated with the primary antiserum (1:1000 rabbitanti-human lAPP^y [Peninsula] or 1:400 guinea piganti-insulin [Dako]) for 2 h at 37°C and were rinsed withTBS-T. Sections were then incubated with goat anti-rabbit-immunoglobulin (IAPP) or protein-A (insulin) con-jugated to colloidal gold for 2-3 h at room temperatureand were rinsed in TBS-T. Sections were counterstainedwith uranyl acetate. Immunocytochemical negative con-trol samples consisted of omission of the primary anti-serum and application of the gold-conjugated secondaryantibody or protein-A gold solution alone.
Each case was independently evaluated by two pa-thologists (A.E.B., T.D.O.) to confirm histopathologicaldiagnosis, to assess the presence of amyloid deposits,and to evaluate immunoreactivity to IAPP and insulin inamyloid and/or tumor cells.
RESULTSClinical characteristics of cases included are summa-rized in Table 1. Statistical comparison (analysis ofvariance) of duration of clinical illness related to insuli-noma between patients with IAPP-derived amyloid intheir insulinoma versus those without amyloid revealedno significant difference (P> 0.05). The histological ap-pearance of all tumors was consistent with previousdescriptions of human pancreatic endocrine tumors (14).Briefly, tumors were composed of solid cords or palisad-
330 DIABETES, VOL. 43, FEBRUARY 1994
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/43/2/329/360700/43-2-329.pdf by guest on 14 February 2022
T.D. O'BRIEN AND ASSOCIATES
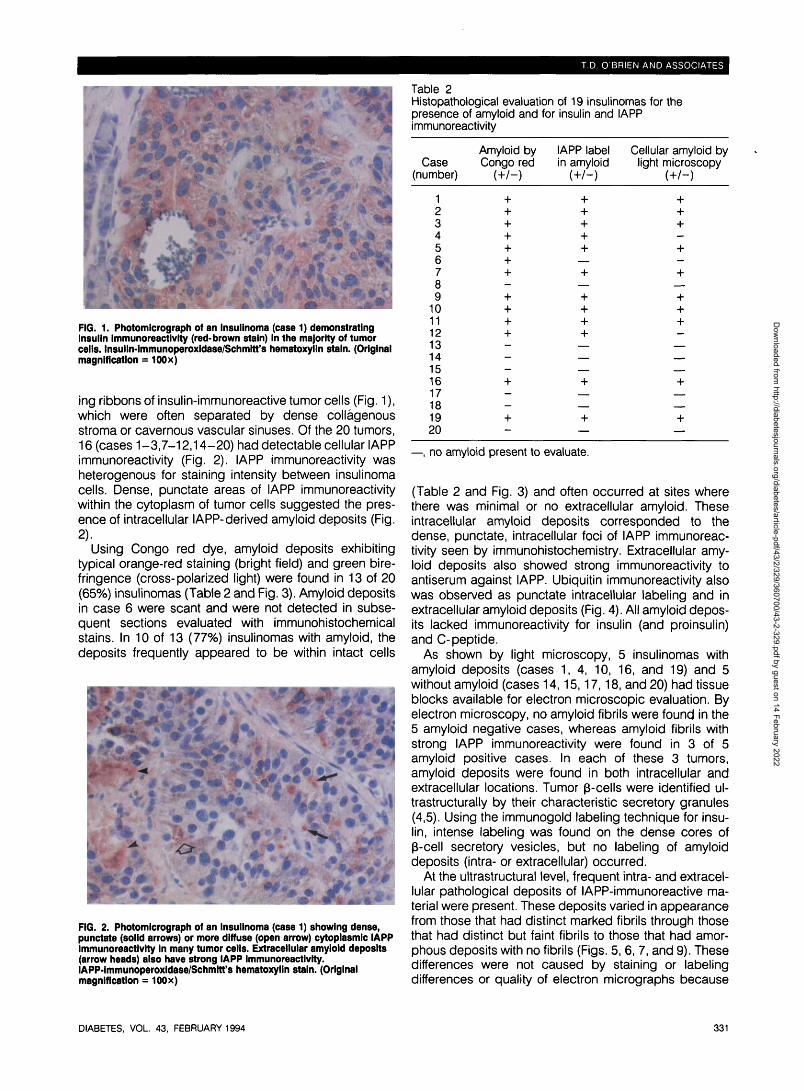
FIG. 1. Photomicrograph of an Insullnoma (case 1) demonstratingInsulin immunoreactivity (red-brown stain) In the majority of tumorcells. Insulln-immunoperoxidase/Schmitt's hematoxylln stain. (Originalmagnification = 100x)
ing ribbons of insulin-immunoreactive tumor cells (Fig. 1),which were often separated by dense collagenousstroma or cavernous vascular sinuses. Of the 20 tumors,16 (cases 1-3,7-12,14-20) had detectable cellular IAPPimmunoreactivity (Fig. 2). IAPP immunoreactivity washeterogenous for staining intensity between insulinomacells. Dense, punctate areas of IAPP immunoreactivitywithin the cytoplasm of tumor cells suggested the pres-ence of intracellular lAPP-derived amyloid deposits (Fig.2).
Using Congo red dye, amyloid deposits exhibitingtypical orange-red staining (bright field) and green bire-fringence (cross-polarized light) were found in 13 of 20(65%) insulinomas (Table 2 and Fig. 3). Amyloid depositsin case 6 were scant and were not detected in subse-quent sections evaluated with immunohistochemicalstains. In 10 of 13 (77%) insulinomas with amyloid, thedeposits frequently appeared to be within intact cells
FIG. 2. Photomicrograph of an Insulinoma (case 1) showing dense,punctate (solid arrows) or more diffuse (open arrow) cytoplasmlc IAPPImmunoreactivity In many tumor cells. Extracellular amyloid deposits(arrow heads) also have strong IAPP immunoreactivity.lAPP-lmmunoperoxidase/Schmltt's hematoxylin stain. (Originalmagnification = 100x)
Table 2Histopathological evaluation of 19 insulinomas for thepresence of amyloid and for insulin and IAPPimmunoreactivity
Case(number)
123456789
1011121314151617181920
Amyloid byCongo red
+++++—++++———+_—+-
IAPP labelin amyloid
+ ++++
+
++++
+
+—
Cellular amyloid bylight microscopy
+—+_+
+++_
+—
—, no amyloid present to evaluate.
(Table 2 and Fig. 3) and often occurred at sites wherethere was minimal or no extracellular amyloid. Theseintracellular amyloid deposits corresponded to thedense, punctate, intracellular foci of IAPP immunoreac-tivity seen by immunohistochemistry. Extracellular amy-loid deposits also showed strong immunoreactivity toantiserum against IAPP. Ubiquitin immunoreactivity alsowas observed as punctate intracellular labeling and inextracellular amyloid deposits (Fig. 4). All amyloid depos-its lacked immunoreactivity for insulin (and proinsulin)and C-peptide.
As shown by light microscopy, 5 insulinomas withamyloid deposits (cases 1, 4, 10, 16, and 19) and 5without amyloid (cases 14,15,17,18, and 20) had tissueblocks available for electron microscopic evaluation. Byelectron microscopy, no amyloid fibrils were found in the5 amyloid negative cases, whereas amyloid fibrils withstrong IAPP immunoreactivity were found in 3 of 5amyloid positive cases. In each of these 3 tumors,amyloid deposits were found in both intracellular andextracellular locations. Tumor p-cells were identified ul-trastructurally by their characteristic secretory granules(4,5). Using the immunogold labeling technique for insu-lin, intense labeling was found on the dense cores ofp-cell secretory vesicles, but no labeling of amyloiddeposits (intra- or extracellular) occurred.
At the ultrastructural level, frequent intra- and extracel-lular pathological deposits of lAPP-immunoreactive ma-terial were present. These deposits varied in appearancefrom those that had distinct marked fibrils through thosethat had distinct but faint fibrils to those that had amor-phous deposits with no fibrils (Figs. 5, 6, 7, and 9). Thesedifferences were not caused by staining or labelingdifferences or quality of electron micrographs because
DIABETES, VOL. 43, FEBRUARY 1994 331
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/43/2/329/360700/43-2-329.pdf by guest on 14 February 2022

IAPP IN INSULINOMAS
FIG. 3. Photomicrograph of a Congo red-stained Insullnoma (case 1)Illuminated by cross-polarized light showing congophilic extracellular(solid arrow) amyloid deposits and small congophilic foci (arrowheads) that appear to be Intracellular. Note the typical greenbirefringence of amyloid. Congo red stain. (Originalmagnification = 100x)
these different appearances can be observed in thesame section. For the purposes of this study, we consid-ered both the marked fibrillar and the faint but distinctlyfibrillar IAPP aggregates as amyloid. The justification ofthis was that the faintly fibrillar material showed fibrils thatwere consistent with amyloid fibrils (i.e., 7- to 10-nmdiameter nonbranching fibrils). Furthermore, the size andlocation of these deposits correlated with the congophilicand lAPP-immunoreactive intracellular amyloid depositsidentified in tumor cells by light microscopy.
At the ultrastructural level, IAPP-derived amyloid waspresent in tumor p-cells 1) in intracellular deposits, 2) inextracellular deposits, and 3) in deposits that wereapparently being extruded from the cell to the extracel-lular space. Intracellular lAPP-immunoreactive depositsin tumor cells were found in several ultrastructural loca-tions and in sizes ranging from 250 nm up to 10 (xm. Thelarger amyloid deposits observed by electron micros-
FIG. 4. Photomicrograph of an insullnoma (case 2) demonstratingdense, punctate intracellular staining (arrows) In tumor cells andintense labeling of extracellular amyloid deposits (arrow heads).Ubiquitin-immunoperoxidase/Schmitt's hematoxylin stain. (Originalmagnification = 100x)
FIG. 5. Electron micrograph of an insulinoma p-cell (case 1) showingintracellular amyloid with faintly but distinctly fibrillar morphology(arrows) that is free within the cytoplasm adjacent to the nucleus ofthe cell. Note the extensive IAPP immunoreactivity (15-nmimmunogold label) of this deposit. (Bar = 750 nm)
copy corresponded in size and location with amyloiddeposits seen by light microscopy. The smallest intracel-lular amyloid deposits were too small to be reliablydetected by light microscopy. Intracellular amyloid de-posits often were free within the tumor cell cytoplasm andwere not surrounded by a membrane (Figs. 5 and 6).Such deposits were contiguous with the adjacent cyto-plasm of the affected cell. This type of deposit was seenat various locations including sites adjacent to mitochon-dria and next to the nucleus deep within the cell (Fig. 5).The larger intracellular amyloid deposits (up to 10 u.m ingreatest dimension) displaced normal cellular organelles(Fig. 6). However, even these large amyloid depositswere completely surrounded by cytoplasm. Within someof these large deposits, profiles of remaining cellularorganelles were found, but no evidence was observed ofextracellular components such as collagen fibrils (Fig. 6).The lAPP-immunoreactive material in these large depos-its consisted of the faintly but distinctly fibrillar amyloidand amorphous deposits without apparent fibrillarity.
Other intracellular lAPP-immunoreactive deposits werefound within round to elongated, membrane-boundstructures (Fig. 7). The membrane-bound structureswere of variable size (-250-1,000 nm in greatest dimen-sion) and were sometimes found adjacent to golgi (Fig.7). Tightly packed but distinct lAPP-immunoreactivefibrils were usually more apparent in the larger mem-brane-bound structures, whereas the smallest of thesemembrane-bound structures contained lAPP-immunore-active material that, like portions of the larger intracellularamyloid deposits, often had a granular or amorphousappearance with little apparent fibrillar structure. Thelack of fibrillar appearance of some lAPP-immunoreac-tive deposits may reflect an early phase in amyloidogen-esis or in some regions may have been partly caused bythe plane of section being perpendicular to the long axisof the amyloid fibrils.
IAPP immunoreactivity also was noted frequently on
332 DIABETES, VOL. 43, FEBRUARY 1994
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/43/2/329/360700/43-2-329.pdf by guest on 14 February 2022

T.D. O'BRIEN AND ASSOCIATES
.**• e W - . "4 ' . ** - * * • *;
FIG. 6. 4 : Electron micrograph of an insullnoma p-cell (case 1) that IsIsolated from adjacent cells and amyloid deposits and Is completelysurrounded by erythrocytes. Note the large lAPP-lmmunolabeled (15nm gold) deposit central In the cell that Is displacing normal cellularorganelles. (Bar = 2.0 |im). B: Higher magnification of a region fromcell in Fig. 6 showing the large Intracellular amyloid deposit. Note thatwhereas the amyloid deposit is fairly sharply delimited, it Is notbounded by membrane. The amyloid is composed of faint but distinct,7- to 10-nm diameter fibrils (arrows) that are strongly Immunolabeledfor IAPP. This deposit also contains several profiles of membranousorganelles (arrow heads). (Bar = 500 nm)
FIG. 7. Electron micrograph of a tumor p-cell (case 1) containingmultiple well-demarcated lAPP-lmmunolabeled (15 nm gold) amyloiddeposits comprised of compact arrays of 7- to 10-nm diameter fibrils.Some deposits are bounded by distinct membranes (arrow). Note thelocalization of small membrane-bound lAPP-lmmunolabeled material(arrow heads) adjacent to golgi apparatus. (Bar = 750 nm)
loid fibrils appeared to project from multiple locations inthe p-cell cytoplasm into the extracellular amyloid depos-its (Fig. 9). Within these p-cells, the lAPP-amyloid fibrilswere often found in close proximity to intracellular or-ganelles (e.g., mitochondria) with no intervening cellmembrane (i.e., are clearly in contact with cytoplasm).Amyloid bundles originating from these sites were con-tinuous with dense bundles of amyloid fibrils within theextracellular deposits. Tumor cell basement membranesand extracellular spaces were obliterated at these sitesof tumor cell/amyloid interface.
DISCUSSIONIn this study, we sought to determine the frequency andultrastructural location of IAPP-derived amyloid formationin human insulinoma tissue. In three previous studies(15-17), the observation of lAPP-immunoreactive amy-loid within tumor p-cells was mentioned but gave littledetail concerning the intracellular locations and did notprovide any estimate of how frequently this occurred inIAPP-producing tumors. We report the presence of IAPP-
the dense cores of tumor p-cell secretory vesicles (Fig.8). IAPP immunoreactivity was seen less often in second-ary lysosomes (Fig. 8), however, no amyloid fibrils wereobserved in these sites. Insulin immunoreactivity wasfound only on the dense core of the tumor p-cell secre-tory vesicles.
IAPP-derived extracellular amyloid deposits occurredvariously as small accumulations between tumor p-cells,perivascularly between tumor p-cells and basementmembranes of adjacent capillaries, or as large masseswithout discemable vascular orientation. Perivascularamyloid often was intermixed with cellular debris, gran-ular material (without IAPP immunoreactivity), and a fewcollagen fibrils. Large masses of amyloid sometimesappeared to be compressing adjacent tumor cells andgenerally contained few or no resident cells.
The relationship between p-cells and amyloid depositsat some sites was suggestive of IAPP amyloid beingextruded from cells. In these locations, bundles of amy-
FIG. 8. Electron micrograph of an insulinoma p-cell (case 1)demonstrating secondary lysosomes (arrows) with IAPPImmunolabeling but without detectable formation of amyloid fibrils.(Bar = 200 nm)
DIABETES, VOL. 43, FEBRUARY 1994 333
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/43/2/329/360700/43-2-329.pdf by guest on 14 February 2022

IAPP IN INSULINOMAS
FIG. 9. Electron micrograph of a tumor p-cell (case 16) with distinctbundles of lAPP-lmmunolabeled (15 nm gold) amyloid fibrils extendingfrom the cytoplasm Into a large extracellular amyloid deposit. Notethat amyloid fibrils occur in close proximity to a mitochondrion withno intervening cell membrane (arrows). (Bar = 400 nm)
immunoreactive amyloid deposits in 13 of 20 (65%)insulinomas studied by light microscopy. Furthermore,light microscopy showed that 77% of the amyloid-con-taining tumors had amyloid deposits within tumor cells.lAPP-immunoreactive intracellular amyloid fibrils weredemonstrated in all three of the tumors in which amyloidwas identified ultrastructurally.
A point of particular interest in this study is the spec-trum of appearances of pathological IAPP aggregates inthese tumors from those with marked and distinct fibrils,through those with less distinct but faint fibrils, to thosewith no fibrils. We emphasize that each of these types ofaggregates are highly immunoreactive for IAPP and thatnone of these aggregates occur in healthy normal p-cells(i.e., all are pathological). These are unlikely to be threedistinct unrelated forms of IAPP aggregate but morelikely represent part of a spectrum of evolving amyloid.Thus, it is well established that mature amyloid has adistinct fibrillar appearance and that this material ischemically stable and strongly resistant to enzymatic orchemical degradation (i.e., biologically inert). Therefore,it is highly improbable that the loosely aggregated, faintlyfibrillar, lAPP-immunoreactive material is derived fromthe above but quite likely that the reverse is true. Amyloidfibrils are formed by a process of aggregation of the unitpeptides (i.e., IAPP) in repetitive antiparallel p-pleatedsheet conformation that results in the progressive elon-gation of the amyloid fibrils. Furthermore, in vitro experi-ments have shown that amyloid fibrils undergoelongation from an initial nidus of fibril formation. There-fore, it is not surprising that regions of early amyloidformation lack detectable fibrils because at this point inamyloidogenesis the fibrils are not sufficiently large toobserve at the limits of resolution of biological electronmicroscopy. Taken together, we therefore propose thatthe nonfibrillar but markedly abnormal lAPP-immunore-active deposits likely mature through the faintly fibrillardeposits and finally to the mature distinct fibrils that have
been the classic definition of amyloid at the ultrastructurallevel.
Theoretically, it might be argued that the apparentintracellular deposits represent invaginations of extracel-lular amyloid. However, several factors indicate thatintracellular formation of IAPP-derived amyloid depositsoccurs. First, lAPP-immunoreactive amyloid depositswere found deep within the cytoplasm adjacent to thenucleus and mitochondria, and furthermore, no mem-brane delimiting such deposits from the surroundingp-cell cytoplasm was found (Fig. 5). Second, intracellularamyloid deposits contained no extracellular componentssuch as collagen fibrils that are often admixed withextracellular amyloid deposits. Third, p-cells that weresurrounded by hemorrhage and isolated from surround-ing tumor cells and connective tissues were found tocontain large (up to 10 u,m in greatest dimension) intra-cellular lAPP-immunoreactive deposits (Fig. 6). Thesedeposits were completely surrounded by p-cell cyto-plasm and often occupied much of the cytoplasm anddisplaced normal organelles. Furthermore, no extracellu-lar amyloid was adjacent to these cells. Material withinthese deposits showed extensive immunogold labelingfor IAPP and consisted of loosely aggregated 7- to10-nm diameter, distinct but faint fibrils (consistent withamyloid fibrils), and granular or amorphous material,which did not have classical amyloid morphology. Fourth,because ubiquitination is an energy-dependent intracel-lular process, the colocalization of ubiquitin with IAPP inall amyloid deposits further argues for an intracellularorigin for these deposits.
The presence of intracellular amyloid could eitherreflect intracellular formation of amyloid fibrils or cellularuptake (i.e., phagocytosis) of extracellularly formed amy-loid fibrils. The latter seems unlikely because p-cellshave not been shown to have phagocytic properties and,in this study, amyloid was not associated with secondarylysosomes as would be expected following phagocyto-sis. Also, note that intracellular formation of amyloid fibrilshas been proposed recently in several other examples oflocal amyloid deposition involving other precursor pro-teins. Examples include growth hormone and prolactin-derived amyloid in anterior pituitary adenomas (19),p-protein in inclusion body myositis (20) and Alzheimer'sdisease (21), and immunoglobulin light chains in myelomacells (22). If IAPP-derived amyloid is indeed initiallyformed intracellularly in insulinomas, several questionsarise. First, what is the mechanism of intracellular IAPP-derived amyloid formation in insulinomas, and what is theprogression of events leading to formation of the coex-istent extracellular lAPP-derived amyloid? Second, what,if any, is the relationship between the pathogenesis oflAPP-derived amyloid in insulinomas and the same formof amyloid present in the islets of NIDDM patients?
Regarding the mechanism of lAPP-derived amyloidformation, data obtained from molecular biological andbiochemical studies indicate that the amino acid se-quence of human IAPP obtained from insulinoma and IAdeposits (1,2) is identical to the normal DNA-predictedsequence (23). Therefore, the mechanism of IAPP amy-loidogenesis does not appear to involve an abnormal
334 DIABETES, VOL. 43, FEBRUARY 1994
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/43/2/329/360700/43-2-329.pdf by guest on 14 February 2022

T.D. O'BRIEN AND ASSOCIATES
form of IAPP occurring because of a gene defect orabnormal proteolytic cleavage of IAPP or pro-IAPP toproduce an amyloidogenic fragment. These observa-tions are supported further by a study showing that thecDNA-predicted IAPP sequences from 25 NIDDM pa-tients were completely normal (24).
It is also known that only a select few species (e.g.,humans, macaques, and cats) develop IAPP-derived IAdeposits (11). This species specificity of IA formation isdue to the peculiar amino acid sequence of the IAPP ofthese species-spanning residues 20-29 (25-27). Thus,although IAPP structure shows close sequence homol-ogy between all species in both the amino-terminal(residues 1-19) and carboxy-terminal (residues 30-37),regions with less homology are present in residues20-29, especially between rodents and other mamma-lian species. Secondary structural predictions of thisregion of the human IAPP molecule indicate a propensityto form the p-pleated sheet secondary structure neces-sary to form amyloid fibrils (23). These predictions havebeen confirmed in vitro with synthetic peptides corre-sponding to the amino acid sequence of human and catIAPP in the 20-29 region that, under conditions of highconcentration, forms amyloid fibrils (27). In contrast,under similar circumstances the corresponding syntheticpeptides in the rat, guinea pig, and mouse do not formamyloid fibrils (27). Thus, the species specificity ofIAPP-derived amyloid formation appears to be causedby an inherent amyloidogenic midsequence in the IAPPof these species that promotes fibrillogenesis if a suffi-ciently high concentration of the protein is achieved.'However, additional (and as yet unknown) factors mustalso be involved in the amyloidogenesis of IAPP becauseIAPP is normally produced and secreted by all individu-als, yet only a minority develop IAPP-derived amyloiddeposits and NIDDM. This is further illustrated by the factthat canine IAPP contains the putative amyloidogenicsequence (28) and forms amyloid in canine pancreaticendocrine tumors (18), but dogs do not develop IA orNIDDM.
Although the IAPP-derived amyloid fibrils were com-monly found intracellularly in insulinoma cells in thisstudy, the majority of the amyloid was deposited extra-cellularly. Extracellular amyloid deposits might arise viaseveral different mechanisms, either alone or in combi-nation. Potential mechanisms include the secretion orextrusion of amyloid fibrils from the tumor cells as hasbeen suggested in pituitary adenomas (19). This mech-anism is supported by our observation that the amyloid-containing, membrane-bound structures within tumorp-cells were sometimes seen near the cell surface andby the observation of deep cytoplasmic invaginations ofamyloid that were seen in some tumor p-cells adjacent toextracellular amyloid deposits. This extrusion of pre-formed IAPP-derived amyloid from p-cells, which areoriented around capillaries, may explain the observationthat early IA deposits are often located perivascularly(29). Another potential route for movement of amyloid tothe extracellular space might involve formation of mas-sive accumulations of intracytoplasmic (either free ormembrane-bound) amyloid fibrils followed by p-cell
death and lysis with release of amyloid to the extracellularspace. This mechanism is supported by our observationthat cellular debris was commonly found within andadjacent to the amyloid deposits. We also cannot ex-clude the possibility that IAPP-derived amyloid simulta-neously forms extracellularly along with intracellularformation.
Finally, the question arises as to the potential relation-ship between the pathogenesis of IAPP-derived IA foundin NIDDM and the IAPP-derived amyloid found in insuli-noma tissue. If IAPP-derived amyloid in NIDDM alsoinitially forms intracellularly, as suggested by the currentultrastructural studies in insulinomas, it is feasible thatthis could lead to p-cell death and subsequent release ofthe amyloid to the extracellular space. Several lines ofevidence from previous studies support this scenario.First, the mean volume of islets in NIDDM patientsremains unaltered from those in health even when >50%of the islet is occupied by amyloid (30). This suggeststhat as the p-cells are lost a comparable volume ofamyloid occupies the former cellular volume. Second,intracellular IAPP-derived amyloid deposits have beendescribed in prediabetic and diabetic cats (4,12). Thatthis has not as yet been described in human NIDDM maybe due to several factors. First, studies thus far haveused tissues obtained at autopsy, which, because ofautolysis, adversely affects histological and ultrastruc-tural details. Autolysis also reduces the ability to detectIAPP immunoreactivity, which might interfere with detec-tion of small intracellular deposits that were seen in thisstudy. Last, because most cases evaluated at autopsyare in the advanced stages of the disease, early lesionssuch as intracellular amyloidogenesis will not be evident,and remaining amyloid will be principally extracellular.
The development of IAPP-derived amyloid deposits(either in pancreatic islets or in insulinomas) intracellu-larly implies that cellular mechanisms, which normallyprevent IAPP from aggregating into fibrils, are over-whelmed. This may be due to either an increased rate ofsynthesis of IAPP beyond levels that p-cells can processor an intrinsic defect in this protective mechanism suchthat even normally expressed amounts of IAPP cannot beaccommodated. Further studies are needed to explorethe possible sites of abnormalities in IAPP processing,trafficking, or degradation that would account for theformation of amyloid fibrils from this normally expressedpeptide in patients with NIDDM.
ACKNOWLEDGMENTSThis study was supported by Public Health ServiceGrants RO1-DK-44341, RR-585A, and DK-36734; a grantfrom Pfizer Pharmaceuticals; and the Mayo Foundation.
The authors also acknowledge the excellent technicalassistance of Margaret J. Hukee for preparation of theimmunogold-labeled specimens for electron micros-copy.
REFERENCES1. Westermark P, Wernstedt C, Wilander E, Hayden DW, O'Brien TD,
Johnson KH: Amyloid fibrils in human insulinoma and islets ofLangerhans of the diabetic cat are derived from a neuropeptide-like
DIABETES, VOL 43, FEBRUARY 1994 335
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/43/2/329/360700/43-2-329.pdf by guest on 14 February 2022

IAPP IN INSULINOMAS
protein also present in normal islet cells. Proc Natl Acad Sci USA84:3881-85, 1987
2. Westermark P, Wernstedt C, O'Brien TD, Hayden DW, Johnson KH:Islet amyloid in type II human diabetes mellitus and adult diabeticcats is composed of a novel putative polypeptide hormone. Am JPathol 127:414-17, 1987
3. Cooper GJS, Willis AC, Clark A, Turner RC, Sim RB, Reid KBM:Purification and characterization of a peptide from amyloid-richpancreases of type II diabetic patients. Proc Natl Acad Sci USA84:8628-32, 1987
4. Johnson KH, O'Brien TD, Hayden DW, Jordan K, Ghobrial HKG,Mahoney WC, Westermark P: Immunolocalization of islet amyloidpolypeptide (IAPP) in pancreatic p-cells by means of peroxidase-antiperoxidase (PAP) and protein A-techniques. Am J Pathol 130:1-8, 1988
5. Lukinius A, Wilander E, Westermark GT, Engstrom U, Westermark P:Colocalization of islet amyloid polypeptide and insulin in the p-cellsecretory granules of the human pancreatic islets. Diabetologia32:240-44, 1989
6. Fehmann HC, Weber V, Goke R, Goke B, Arnold R: Cosecretion ofamylin and insulin from isolated rat pancreas. FEBS Lett 262:279-81,1990
7. Inoue K, Hisatomi A, Umeda F, Nawata H: Release of amylin fromperfused rat pancreas in response to glucose, arginine, p-hydroxy-butyrate, and gliclazide. Diabetes 40:1005-1009, 1991
8. Butler PC, Chou J, Carter WB, Wang Y, Bu B, Chang D, Chang J,Rizza RA: Effects of meal ingestion on plasma amylin concentrationin NIDDM and nondiabetic humans. Diabetes 39:752-56, 1990
9. O'Brien TD, Westermark P, Johnson KH: Islet amyloid polypeptide(IAPP) and insulin secretion from isolated perfused pancreas of fed,fasted, glucose-treated,and dexamethasone-treated rats. Diabetes40:1701-706, 1991
10. Ogawa A, Harris V, McCorkle SK, Unger RH, Luskey KL: Amylinsecretion from the rat pancreas and its selective loss after strepto-zotocin treatment. J Clin Invest 85:973-76, 1990
11. Johnson KH, O'Brien TD, Betsholtz C, Westermark P: Islet amyloid,islet-amyloid polypeptide, and diabetes mellitus. N Engl J Med321:513-18, 1989
12. Yano BL, Hayden DW, Johnson KH: Feline insular amyloid: ultra-structural evidence for intracellular formation by nonendocrine cells.Lab Invest 45:149-56, 1981
13. Clark A, DeKoning E, Hansen B, Bodkin N, Morris JF: Islet amyloidin glucose intolerant and spontaneous diabetic Macaca mulattamonkeys. In Frontiers in Diabetes Research, Lessons From AnimalDiabetes, ///Shafrir E, Ed. Jersalem, Smith-Gordan, 1990, p. 502-506
14. Mukai K, Greider MH, Grotting JC, Rosai J: Retrospective study of77 pancreatic endocrine tumors using the immunoperoxidasemethod. Am J Surg Pathol 6:378-99, 1982
15. Lukinius A, Wilander E, Stridsberg M, Eriksson B, Oberg K: Electron-microscopical immunocytochemical study of a pancreatic isletamyloid polypeptidoma. Endocr Pathol 2:169-75, 1991
16. Toshimori H, Narita R, Nakazato M, Asai J, Mitsukawa T, KangawaK, Matsuo H, Takahashi K, Matsukura S: Islet amyloid polypeptide in
insulinoma and in the islet of the pancreas of nondiabetic anddiabetic subjects. Virchows Archiv Pathol Anat 418:411-17, 1991
17. Rindi G, Terenghi G, Westermark G, Westermark P, Moscoso G,Polak JM: Islet amyloid polypeptide in proliferating pancreaticp-cells during development, hyperplasia, and neoplasia in humansand mice. Am J Pathol 138:1321-34, 1991
18. O'Brien TD, Westermark P, Johnson KH: Canine pancreatic endo-crine tumors: islet amyloid polypeptide and calcitonin gene-relatedpeptide immunoreactivity in amyloid and tumor cells. Vet Pathol27:200-204, 1990
19. Saitoh Y, More H, Matsumoto K, Ushio Y, Hayakawa T, Mori S, AritaN, Mogami H: Accumulation of amyloid in pituitary adenomas. ActaNeuropathol 68:87-92, 1985
20. Askanas V, Engel WK, Alvarez RB: Light and electron microscopiclocalization of p-amyloid protein in muscle biopsies of patients withinclusion-body myositis. Am J Pathol 141:31-36, 1992
21. Wisniewski HM, Wegiel KC, Wang M, Kujawa M, Lach B: Ultrastruc-tural studies of the cells forming amyloid fibers in classical plaques.Can J Neurol Sci 16:535-42, 1989
22. Drurie BGM, Perskey B, Soehnlen BJ, Grogan TM, Salmon SE:Amyloid production in human myeloma stem-cell culture, withmorphological evidence of amyloid secretion by associated macro-phages. N EnglJ Med 307:1689-92, 1982
23. Betsholtz C, Svensson V, Rorsman F, Engstrom U, Westermark GT,Wilander E, Johnson KH, Westermark P: Islet amyloid polypeptide(IAPP): cDNA cloning and identification of an amyloidogenic regionassociated with the species-specific occurrence of age-relateddiabetes mellitus. Exp Cell Res 183:484-93, 1989
24. Nishi M, Bell Gl, Steiner DF: Islet amyloid polypeptide (amylin): noevidence of an abnormal precursor sequence in 25 type II (non-insulin-dependent) diabetic patients. Diabetologia 33:628-30,1990
25. Betsholtz C, Christmansson L, Engstrom U, Rorsman F, Jordan K,O'Brien TD, Murtaugh M, Johnson KH, Westermark P: Structure ofcat islet amyloid polypeptide and identification of amino acidresidues of potential signficance for islet amyloid formation. Diabe-tes 39:118-22, 1990
26. Betsholtz C, Christmansson L, Engstrom U, Rorsman F, Svensson V,Johnson KH, Westermark P: Sequence divergence in a specificregion of islet amyloid polypeptide (IAPP) explains differences inislet amyloid formation between species. FEBS Lett 251:261-64,1989
27. Westermark P, Engstrom U, Johnson KH, Westermark GT, BetsholtzC: Islet amyloid polypeptide: pinpointing amino acid residues linkedto amyloid fibril formation. Proc Natl Acad Sci USA 87:5036-40,1990
28. Jordan K, Murtaugh MP, O'Brien TD, Westermark P, Betsholtz C,Johnson KH: Canine IAPP cDNA sequence provides importantclues regarding diabetogenesis and amyloidogenesis in type IIdiabetes. Biochem Biophys Res Commun 169:502-508, 1990
29. Westermark P: Fine structure of islets of Langerhans in insularamyloidosis. Virchows Archiv Pathol Anat 359:1-18, 1973
30. Westermark P: Quantitative studies of amyloid in the islets ofLangerhans. UpsalaJ Med Sci 77:91-94, 1972
336 DIABETES, VOL. 43, FEBRUARY 1994
Dow
nloaded from http://diabetesjournals.org/diabetes/article-pdf/43/2/329/360700/43-2-329.pdf by guest on 14 February 2022