is the effect of trees on soil properties mediated by soil fauna? a case study from post-mining...
TRANSCRIPT

Forest Ecology and Management xxx (2013) xxx–xxx
Contents lists available at SciVerse ScienceDirect
Forest Ecology and Management
journal homepage: www.elsevier .com/locate / foreco
Is the effect of trees on soil properties mediated by soil fauna? A case studyfrom post-mining sites
Jan Frouz a,⇑, Miluše Livecková b, Jana Albrechtová c, Alica Chronáková d, Tomáš Cajthaml a,e, Václav Pizl d,Ladislav Hánel d, Josef Stary d, Petr Baldrian e, Zuzana Lhotáková c, Hana Šimácková a, Šárka Cepáková b,d
a Institute of Environmental Studies, Charles University in Prague, Benátská 2, CZ 128 00 Prague, Czech Republicb Department of Ecosystem Biology, University of South Bohemia in Ceské Budejovice, CZ 370 05 Ceské Budejovice, Czech Republicc Department of Experimental Plant Biology, Charles University in Prague, Vinicná 5, CZ 128 44 Prague, Czech Republicd Institute of Soil Biology, Biology Centre AS CR, Na Sádkách 7, CZ 370 05 Ceské Budejovice, Czech Republice Institute of Microbiology AS CR, Vídenská 1083, CZ 142 20 Prague, Czech Republic
a r t i c l e i n f o
Article history:Available online xxxx
Keywords:BioturbationSoil faunaMicroorganismsReclamationEarthwormsFoliage chemistry
0378-1127/$ - see front matter � 2013 Elsevier B.V. Ahttp://dx.doi.org/10.1016/j.foreco.2013.02.013
⇑ Corresponding author. Tel.: +420 221951902.E-mail address: [email protected] (J. Frouz).
Please cite this article in press as: Frouz, J., et al.Ecol. Manage. (2013), http://dx.doi.org/10.1016
a b s t r a c t
The effects of tree species differing in foliage and litter chemistry on the chemical, micro-morphological,and biological properties of soil were studied on post-mining sites afforested with one of six tree species(spruce, pine, larch, oak, lime, and alder) and also on sites left to natural succession (dominated by wil-low). The sites were located on a large colliery spoil heap that had been produced by the mining of coal inalkaline tertiary clays near the city of Sokolov, Czech Republic. Because no topsoil had been applied to thesites, soil development resulted from in situ interactions among the deposited overburden (spoil), trees,and soil biota. Soil formation differed markedly among sites afforested with different tree species. Onsites with trees producing litter with a low C/N ratio (the deciduous species), the organic Oe layer wasnarrow or absent and a thick organomineral A layer was evident. On sites with trees producing litter witha high C/N ratio (the evergreen species), in contrast, a thick Oe layer and a thin A layer were evident.Besides C/N ratio, earthworm abundance and earthworm bioturbation activity (measured as the amountof earthworm casts in the topsoil) were the strongest predictors of A layer thickness and C accumulationin the mineral topsoil. Sites with higher C accumulation in mineral soil had higher microbial biomass andlower microbial respiration, which may have contributed to the higher C storage. The gradient of biotur-bation was correlated with changes in the composition of the bacterial community and other soil biota,but partial correlation showed that the effects of litter quality and bioturbation were largely independent.Overall, the results indicate that the effect of tree species on soil development is substantially mediatedby soil fauna activity and especially by earthworm bioturbation.
� 2013 Elsevier B.V. All rights reserved.
1. Introduction
Trees are a dominant factor in forest ecosystems and affectmany ecosystem properties including soil. The effect of trees onsoil is important because soil is a fundamental component of eco-systems, and changes in soil properties are likely to change futurevegetation development on the site (Odum, 1969; Frouz et al.,2008a; Jangida et al., 2011). Soil development also generates astrong feedback on existing vegetation and may affect understoryvegetation and tree growth (Mudrák et al., 2010). Because soil isan important pool of many biogenic elements such as C, N, andP, and because soil retains and releases large volumes of water,understanding how soil is affected by trees is necessary forunderstanding local and global biogeochemical cycles and for
ll rights reserved.
Is the effect of trees on soil pro/j.foreco.2013.02.013
understanding water storage and the water cycle (Brady and Weil,2008). Understanding the role of trees in soil carbon storage and incarbon cycling is particularly important because of the potential tomitigate the effects of global climate change (Lal, 2004). Finally,understanding how trees affect soil may help us to understandthe effects of species traits and consequently the importance ofbiodiversity for ecosystem functioning (Schimel et al., 1994).
Trees have both direct and indirect effects on soil. Direct effectsresult from root growth, which supports the formation of soil poresand soil aggregates (Angers and Caron, 1998), and from the pro-duction of litter and root exudates, which greatly affects soil organ-ic matter content, chemistry, and soil structure (Brady and Weil,2008). Indirect effects include those on soil biota. Some plants sup-port decomposer communities (fungal- or bacterial-based detritiv-orous food webs) that are beneficial to them (Wardle, 2006) eitherbecause of the litter that they produce or because of the long-termeffects of the particular tree species on soil properties (Vivanco and
perties mediated by soil fauna? A case study from post-mining sites. Forest

2 J. Frouz et al. / Forest Ecology and Management xxx (2013) xxx–xxx
Austin, 2008). The effect of trees on the physical and chemicalproperties of soil can be mediated by soil fauna, which may sub-stantially affect topsoil properties as a consequence of bioturbation(Lavelle et al., 1997; Scullion and Malik, 2000; Ponge, 2003; Frouzet al., 2007a).
Research in forest ecosystems has usually focused on the effectsof soil fauna on litter decomposition and litter removal from thesoil surface (Setälä et al., 1996; Aubert et al., 2010). Much less re-search has been conducted on the effect of fauna on the physicaland chemical properties of topsoil formation as a consequence ofbioturbation and aggregate formation (Frouz et al., 2007a).Although these processes have been the subject of various manip-ulation experiments in laboratory or field microcosms (Frouz et al.,2007a, 2008b), how bioturbation and aggregate formation differbeneath different tree species in the field has not been sufficientlyelucidated.
The effect of trees on soil properties has usually been studied inafforested agricultural land or other non-forest land where thesoils are already well developed (De Schrijver et al., 2011) andhas seldom been studied during the early development of soil fromparent material (Frouz et al., 2008a, 2009). Understanding the ef-fect of trees in such situation is important for several reasons. Asubstantial amount of land with poorly developed soil has beenformed by human activities such as mining and the building ofhighways and dams. Bare substrates in these areas often differmarkedly from natural soils in having extreme texture consistingfrom narrow range either too fine (mostly clay) or too coarse par-ticles (sand and gravel) structure resulting from weathering of par-ent material without soil aggregates and by lacking recent inputsof organic matter (Bradshaw, 1997). These areas have a severely re-duced ability to perform ecological functions, and although at-tempts at remediating them often include afforestation, soilrecovery is a basic precondition in this effort. A better understand-ing of the mechanisms of soil formation and the role of trees in thisprocess would increase our ability to restore these disturbed sites.In addition to providing this practical benefit, studying these sitescan also help us understand how new soils form from parent mate-rial and can allow us to observe processes that are difficult to ob-serve in mature soils. The latter applies to the effect of soil faunaon soil formation. Soil fauna form structures that are quite persis-tent in soil (Lavelle et al., 1997), and the effect of soil fauna in new
Fig. 1. Map of individual study sites, dashed line represent heap margin, shaded area reT = lime, A = alter, Q = oak, L = larch, Pn = pine, Pc = spruce, and S = unreclaimed or ‘‘succ
Please cite this article in press as: Frouz, J., et al. Is the effect of trees on soil proEcol. Manage. (2013), http://dx.doi.org/10.1016/j.foreco.2013.02.013
soils where no such structures are present should be clearly visible(Frouz et al., 2007b).
The principal objective of this study was to evaluate the effect oftree species on soil faunal communities and on the contribution ofsuch fauna to changes in soil properties during early pedogenesis.
2. Materials and methods
2.1. Study sites
The study was carried out at a large colliery spoil heap (2.5 kmwide and 10 km long) near the town of Sokolov, Czech Republic(50�1404400N, 12�410400E, 450–520 m a.s.l.). The heap wasformed during the last 40 years by the dumping of overburdengenerated by brown coal mining. The overburden consisted of ter-tiary clay material, dominated by illite and kaolinite, with a pHnear 8 (Šourková et al., 2005a; Helingerová et al., 2010). The meanannual precipitation in this area is 650 mm, and the mean annualtemperature is 6.8 �C.
The heap was covered by a mosaic of patches (hereafter referredto as sites) that were overgrown with different types of vegetationof varying age. Each site covered an area of several hectares. Sitesthat had been reclaimed (afforested) by the planting of one of sixtree species were chosen for the study, and these site types werelime (Tilia cordata), alder (Alnus glutinosa and Alnus incana), oak(Quercus robur), larch (Larix decidua), pine (Pinus concorta and Pinusnigra), and spruce (Picea omorica and Picea pungens). Unreclaimedsites, which were dominated by willow (Salix caprea) and whichare referred to as ‘‘succession sites’’, were also included. Each ofthe seven types was represented by four replicate sites, whichwere randomly located on the heap (Fig. 1). In order to select thesites all patches covered by selected species falling to given agerange (22–32 years) was mapped, numbered, and four of themwas randomly selected, inside each patch the sampling site coordi-nates were randomly generated at least 10 m for the patch edge.Vegetation and soil properties of the sites are summarized in Ta-ble 1. No topsoil had been spread on the surface of either the affor-ested or unreclaimed sites. The surface of the afforested sites hadbeen leveled before the trees were planted. No leveling had beenconducted at the unreclaimed sites, where the surface consistedof longitudinally oriented, 1-m-high ridges and depressions
present area of heap covered by woody vegetation, dots represent sampling points,ession’’ sites in which willow was dominant. Numbers 1–4 represent replication.
perties mediated by soil fauna? A case study from post-mining sites. Forest
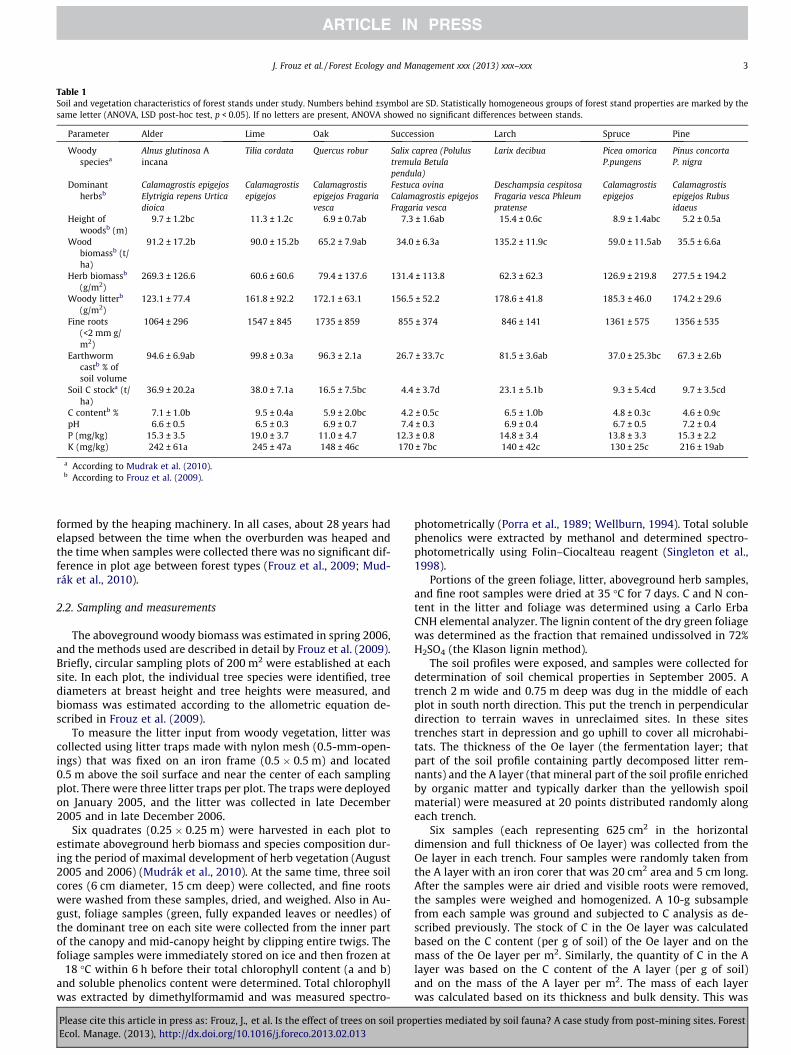
Table 1Soil and vegetation characteristics of forest stands under study. Numbers behind ±symbol are SD. Statistically homogeneous groups of forest stand properties are marked by thesame letter (ANOVA, LSD post-hoc test, p < 0.05). If no letters are present, ANOVA showed no significant differences between stands.
Parameter Alder Lime Oak Succession Larch Spruce Pine
Woodyspeciesa
Alnus glutinosa Aincana
Tilia cordata Quercus robur Salix caprea (Polulustremula Betulapendula)
Larix decibua Picea omoricaP.pungens
Pinus concortaP. nigra
Dominantherbsb
Calamagrostis epigejosElytrigia repens Urticadioica
Calamagrostisepigejos
Calamagrostisepigejos Fragariavesca
Festuca ovinaCalamagrostis epigejosFragaria vesca
Deschampsia cespitosaFragaria vesca Phleumpratense
Calamagrostisepigejos
Calamagrostisepigejos Rubusidaeus
Height ofwoodsb (m)
9.7 ± 1.2bc 11.3 ± 1.2c 6.9 ± 0.7ab 7.3 ± 1.6ab 15.4 ± 0.6c 8.9 ± 1.4abc 5.2 ± 0.5a
Woodbiomassb (t/ha)
91.2 ± 17.2b 90.0 ± 15.2b 65.2 ± 7.9ab 34.0 ± 6.3a 135.2 ± 11.9c 59.0 ± 11.5ab 35.5 ± 6.6a
Herb biomassb
(g/m2)269.3 ± 126.6 60.6 ± 60.6 79.4 ± 137.6 131.4 ± 113.8 62.3 ± 62.3 126.9 ± 219.8 277.5 ± 194.2
Woody litterb
(g/m2)123.1 ± 77.4 161.8 ± 92.2 172.1 ± 63.1 156.5 ± 52.2 178.6 ± 41.8 185.3 ± 46.0 174.2 ± 29.6
Fine roots(<2 mm g/m2)
1064 ± 296 1547 ± 845 1735 ± 859 855 ± 374 846 ± 141 1361 ± 575 1356 ± 535
Earthwormcastb % ofsoil volume
94.6 ± 6.9ab 99.8 ± 0.3a 96.3 ± 2.1a 26.7 ± 33.7c 81.5 ± 3.6ab 37.0 ± 25.3bc 67.3 ± 2.6b
Soil C stocka (t/ha)
36.9 ± 20.2a 38.0 ± 7.1a 16.5 ± 7.5bc 4.4 ± 3.7d 23.1 ± 5.1b 9.3 ± 5.4cd 9.7 ± 3.5cd
C contentb % 7.1 ± 1.0b 9.5 ± 0.4a 5.9 ± 2.0bc 4.2 ± 0.5c 6.5 ± 1.0b 4.8 ± 0.3c 4.6 ± 0.9cpH 6.6 ± 0.5 6.5 ± 0.3 6.9 ± 0.7 7.4 ± 0.3 6.9 ± 0.4 6.7 ± 0.5 7.2 ± 0.4P (mg/kg) 15.3 ± 3.5 19.0 ± 3.7 11.0 ± 4.7 12.3 ± 0.8 14.8 ± 3.4 13.8 ± 3.3 15.3 ± 2.2K (mg/kg) 242 ± 61a 245 ± 47a 148 ± 46c 170 ± 7bc 140 ± 42c 130 ± 25c 216 ± 19ab
a According to Mudrak et al. (2010).b According to Frouz et al. (2009).
J. Frouz et al. / Forest Ecology and Management xxx (2013) xxx–xxx 3
formed by the heaping machinery. In all cases, about 28 years hadelapsed between the time when the overburden was heaped andthe time when samples were collected there was no significant dif-ference in plot age between forest types (Frouz et al., 2009; Mud-rák et al., 2010).
2.2. Sampling and measurements
The aboveground woody biomass was estimated in spring 2006,and the methods used are described in detail by Frouz et al. (2009).Briefly, circular sampling plots of 200 m2 were established at eachsite. In each plot, the individual tree species were identified, treediameters at breast height and tree heights were measured, andbiomass was estimated according to the allometric equation de-scribed in Frouz et al. (2009).
To measure the litter input from woody vegetation, litter wascollected using litter traps made with nylon mesh (0.5-mm-open-ings) that was fixed on an iron frame (0.5 � 0.5 m) and located0.5 m above the soil surface and near the center of each samplingplot. There were three litter traps per plot. The traps were deployedon January 2005, and the litter was collected in late December2005 and in late December 2006.
Six quadrates (0.25 � 0.25 m) were harvested in each plot toestimate aboveground herb biomass and species composition dur-ing the period of maximal development of herb vegetation (August2005 and 2006) (Mudrák et al., 2010). At the same time, three soilcores (6 cm diameter, 15 cm deep) were collected, and fine rootswere washed from these samples, dried, and weighed. Also in Au-gust, foliage samples (green, fully expanded leaves or needles) ofthe dominant tree on each site were collected from the inner partof the canopy and mid-canopy height by clipping entire twigs. Thefoliage samples were immediately stored on ice and then frozen at�18 �C within 6 h before their total chlorophyll content (a and b)and soluble phenolics content were determined. Total chlorophyllwas extracted by dimethylformamid and was measured spectro-
Please cite this article in press as: Frouz, J., et al. Is the effect of trees on soil proEcol. Manage. (2013), http://dx.doi.org/10.1016/j.foreco.2013.02.013
photometrically (Porra et al., 1989; Wellburn, 1994). Total solublephenolics were extracted by methanol and determined spectro-photometrically using Folin–Ciocalteau reagent (Singleton et al.,1998).
Portions of the green foliage, litter, aboveground herb samples,and fine root samples were dried at 35 �C for 7 days. C and N con-tent in the litter and foliage was determined using a Carlo ErbaCNH elemental analyzer. The lignin content of the dry green foliagewas determined as the fraction that remained undissolved in 72%H2SO4 (the Klason lignin method).
The soil profiles were exposed, and samples were collected fordetermination of soil chemical properties in September 2005. Atrench 2 m wide and 0.75 m deep was dug in the middle of eachplot in south north direction. This put the trench in perpendiculardirection to terrain waves in unreclaimed sites. In these sitestrenches start in depression and go uphill to cover all microhabi-tats. The thickness of the Oe layer (the fermentation layer; thatpart of the soil profile containing partly decomposed litter rem-nants) and the A layer (that mineral part of the soil profile enrichedby organic matter and typically darker than the yellowish spoilmaterial) were measured at 20 points distributed randomly alongeach trench.
Six samples (each representing 625 cm2 in the horizontaldimension and full thickness of Oe layer) was collected from theOe layer in each trench. Four samples were randomly taken fromthe A layer with an iron corer that was 20 cm2 area and 5 cm long.After the samples were air dried and visible roots were removed,the samples were weighed and homogenized. A 10-g subsamplefrom each sample was ground and subjected to C analysis as de-scribed previously. The stock of C in the Oe layer was calculatedbased on the C content (per g of soil) of the Oe layer and on themass of the Oe layer per m2. Similarly, the quantity of C in the Alayer was based on the C content of the A layer (per g of soil)and on the mass of the A layer per m2. The mass of each layerwas calculated based on its thickness and bulk density. This was
perties mediated by soil fauna? A case study from post-mining sites. Forest

4 J. Frouz et al. / Forest Ecology and Management xxx (2013) xxx–xxx
based on previous chronosequence studies indicating no C accu-mulation below A layer (Šourková et al., 2005a; Frouz and Kalcík,2006), To correct for fossil C content in 0.75 m was subtracted fromC content in A layer see Frouz et al. (2009) for more details. Brieflymajor source of fossil C in the overburden is type II kerogen, orig-inating from algae that once grow in Miocene lake. Kerogen ishomogeneously distributed in clay overburden (Kríbek et al.,1998). Overburden is dumped in layers which has been homoge-nized by transportation and heaping (Frouz et al., 2011). Samplesfrom 75 cm depth bring us approximately to middle of such layerand was used here as an estimate of fossil C contribution.
In spring 2005 and 2006 (30.3.2005 and 5.4.2006) a corer with10 cm2 area was used to collect 5 cores (0–5 cm depth) of the min-eral soil from each plot. The litter layer was removed before thesesamples were collected an all cores from one site was mixed beforeanalysis. Half of these samples were air dried and sieved using2 mm screen and then used for various chemical analyses the sec-ond half was refrigerated and used for soil microbiology. In 2006samples of Oe layer was sampled as well if Oe layer was presentrefrigerated and used for PLFA.
The pH was measured in a water solution (1:5). The contents ofavailable Ca and K were determined using ion-sensitive electrodesin a 1% citric acid solution extract (1:5, sample:liquid, shaken for1 h). Water-soluble P was determined in a water solution (1:5,sample:liquid, shaken for 1 h) (Murphy and Riley, 1962).
Basal respiration (BR) was estimated by the incubation methodin which 10 g of soil per sample was incubated for 2 days at 20 �Cin a 200-mL vial; the carbon dioxide that evolved from the soil wastrapped in 3 mL of 0.5 M NaOH and then quantified by titrationwith 0.05 M HCl after addition of BaCl2. Bottles without soil wereused to assess CO2 trapped during incubation (from air closed invials) and during handling. Microbial biomass was determined bythe chloroform fumigation–extraction method (Vance et al.,1987). Metabolic quotient known also as specific respiration wascalculated as microbial respiration divided by microbial biomass.
The samples for PLFA analysis were extracted with a mixture ofchloroform–methanol–phosphate buffer (1:2:0.8). Phospholipidswere separated using solid-phase extraction cartridges (LiChrolutSi 60, Merck), and the samples were subjected to mild alkalinemethanolysis as described previously (Šnajdr et al., 2008). The freemethyl esters of PFLAs were analyzed by gas chromatography–mass spectrometry (450-GC, 240-MS ion trap detector, Varian,Walnut Creek, CA, USA). The GC instrument was equipped with asplit/splitless injector, and a DB-5MS column was used for separa-tion (60 m, 0.25 mm i.d., 0.25 mm film thickness). The temperatureprogram began at 60 �C and was held for 1 min in splitless mode.Then the splitter was opened, and the oven was heated to 160 �Cat a rate of 25 �C min�1. The second temperature ramp was up to280 �C at a rate of 2.5 �C min�1, and this temperature was main-tained for 10 min. The solvent delay time was set at 8 min. Thetransfer line temperature was set at 280 �C. Mass spectra were re-corded at 1 scan s�1 under electron impact at 70 eV, mass range50–350 amu. Methylated fatty acids were identified according totheir mass spectra and by using a mixture of chemical standardsfrom Sigma–Aldrich, Prague, Czech Republic and Matreya LLC,Pleasant Gap, PA, USA. Fungal biomass was quantified based on18:2x6,9 content (PLFAfun), and bacterial biomass (PLFAbact)was quantified as a the sum of i14:0, i15:0, a15:0, 16:1x7t,16:1x9, 16:1x7, 10Me-16:0, i17:0, a17:0, cy17:0, 17:0, 10Me-17:0, 10Me-18:0, and cy19:0 (Sampedro et al., 2009). The PLFAfungal/bacterial ratio was calculated as the ratio of PLFAfun/PLFAbact.
Total community DNA was extracted from 2006 samples using0.25 g of soil that had not been dried (Ultra Clean�Soil DNA Isola-tion Kit, Mo-Bio, USA) and was quantitatively characterized spec-trophotometrically (NanoDrop 2000, Thermo Scientific) and
Please cite this article in press as: Frouz, J., et al. Is the effect of trees on soil proEcol. Manage. (2013), http://dx.doi.org/10.1016/j.foreco.2013.02.013
qualitatively characterized by agarose gel electrophoresis. Bacte-rial community composition was studied by 16S rRNA gene frag-ment-PCR-DGGE analysis. DGGE was performed with the IngenyPhorU DGGE system (Ingeny PhorU system, Ingeny, Leiden, NL)and with 100 ng of 473-bp amplicons generated by the primerset GC-968f/1401r on a 6% polyacrylamide at 60 �C for 16 h. DNAfragments were separated on a denaturing gradient of 40–65%;complete details are provided by Chronáková et al. (2012).
Soil fauna was sampled the same term as samples for microbialparameters (30.3.2005 and 5.4.2006). For analysis of soil micro-fauna (nematodes), five samples (area of each 10 cm2, depth5 cm) were collected from each plot and combined to form onecomposite sample per plot. An additional set of samples was col-lected in the same manner and used for analysis of soil mesofauna(oribatid mites and collembolans). For nematode analysis, aftereach composite sample was mixed, 10 g was extracted with a mod-ified Baermann funnel (Hánel, 1995). Oribatid mites and collembo-lans were extracted with a Tullgren apparatus. For analysis ofearthworms, five samples (area of each 125 cm2, depth 5 cm) werecollected from each plot and combined to form one compositesample per plot. Macrofauna were extracted with a Kempson appa-ratus. We do not go into species in this study but there were sev-eral specialized studies dealing with soil fauna composition in thesame post-mining area. As concern earthworms based on previousresearch Lumbricus rubellus and Aporectodea caliginosa belong tomost common species in the area (Frouz et al., 2001, 2008a).
To study soil microstructure, one iron box 5 � 5 � 10 cm wasused to sample soil monolith of 0–5 cm depth from middle of eachtrench. The soil was embedded in epoxide resin and thin soil sec-tions were prepared as described in Frouz et al. (2009). The propor-tion of earthworm cast in soil volume was quantified using thecross intercept method; 1000 crosses were projected on each thinsoil section and intercepts of earthworm casts with the crosseswere recorded. The proportion of intercepts of a cast was used asan estimate for the proportion of volume of a given structure inthe investigated soil (Frouz et al., 2009).
2.3. Statistical analysis
One-way ANOVAs were used to compare individual parametersamong sites with different site types. Two-way ANOVAs were usedto compare the ratios of fungal and bacterial PLFAs in the Oe and Alayers. Linear correlation coefficients of partial correlations wereused to explore the relationships between individual parameters.Bonferonni correction was applied on linear correlation coeffi-cients, however, to indicate potential trends; all correlation coeffi-cients significant at p < 0.05 are shown. If data were collected for2 years, mean values for both years for each plot were used forregression or ANOVA. Statistica 6.0 was used for statistical analy-ses. The effect of tree stand on bacterial community as indicatedby DGGE was studied using principal component analysis (PCA),which was computed by CANOCO 4.0.
3. Results
Although wood biomass differed significantly among sites, andwas highest in larch sites and lowest in succession sites, organicmatter input into soil (litter input, herb biomass, and fine root bio-mass) did not differ among sites (Table 1). Soil C stock significantlydiffered among the sites and was highest in lime and alder sitesand lowest in succession sites (Table 1). The soil C stock in siteswas positively correlated with the C content in the topsoil, thedepth of the A horizon, and proportion of earthworm cast in topsoil(Table 2). Soil pH and water-soluble P did not significantly differamong the sites. Available K content was highest in lime and alder
perties mediated by soil fauna? A case study from post-mining sites. Forest

Tabl
e2
Line
arco
rrel
atio
nco
effi
cien
tbe
twee
nlit
ter
C/N
and
quan
tita
tive
para
met
ers
from
Tabl
es1
and
2.O
nly
sign
ifica
ntco
rrel
atio
nco
effi
cien
tp
<0.
05ar
egi
ven,
corr
elat
ion
coef
fici
ents
that
com
ply
wit
hco
ndit
ions
ofBo
nfer
onny
corr
ecti
onar
ebo
ld.N
umbe
rsin
the
firs
tro
wco
rres
pond
wit
hnu
mbe
red
nam
esof
para
met
ers
liste
din
the
firs
tco
lum
n.
12
34
56
78
910
1112
1314
1516
1-W
ood
biom
ass
2-W
oody
litt
er3-
Eart
hw
orm
cast
4-A
laye
r0.
606
5-O
ela
yer
�0.
411
6-So
ilC
stoc
k0.
650
0.73
27-
Cco
nte
nt
0.61
30.
536
0.78
18-
pH�
0.42
1�
0.63
69-
P�
0.42
20.
461
�0.
450
10-K
0.38
90.
409
0.46
40.
442
11-L
itte
rC
/N�
0.44
0�
0.53
2�
0.49
3�
0.42
812
-Ear
thw
orm
�0.
371
0.38
20.
532
0.51
60.
370
13-M
icro
bial
biom
ass
0.52
80.
376
�0.
472
0.52
30.
713
�0.
477
0.41
10.
424
0.37
614
-Mic
robi
alre
spir
atio
n�
0.45
8�
0.46
6�
0.45
2�
0.56
2�
0.40
40.
405
�0.
420
15-M
etab
olic
quot
ien
t�
0.61
2�
0.44
6�
0.59
6�
0.65
0�
0.44
5�
0.41
6�
0.66
80.
819
16-F
/BPL
FA�
0.44
00.
585
17-N
emat
oda
�0.
550
0.39
4�
0.45
318
-Ori
bati
da0.
671
0.75
0�
0.48
90.
721
0.66
40.
545
�0.
585
0.73
30.
649
�0.
575
�0.
666
19-C
olle
mbo
la�
0.49
10.
585
0.46
90.
595
J. Frouz et al. / Forest Ecology and Management xxx (2013) xxx–xxx 5
Please cite this article in press as: Frouz, J., et al. Is the effect of trees on soil proEcol. Manage. (2013), http://dx.doi.org/10.1016/j.foreco.2013.02.013
and lowest in spruce sites (Table 1). Litter quality differed greatlyamong the sites (Table 3). Although C content in the litter wasquite similar among the sites, N content was greater in the litterproduced by deciduous vs. evergreen trees (Table 3). As a conse-quence, the C/N ratio was significantly lower in deciduous thanin evergreen litter.
The C content in green foliage was somewhat lower than in lit-ter and was significantly higher in the foliage of evergreen thandeciduous trees (Table 3). The N content was significantly lowerand the C/N ratio was significantly higher in the foliage of ever-green than deciduous trees. In general, variation in N and C/Nwas greater in green foliage than in litter (Table 3). In foliage, Ncontent was positively correlated with total chlorophyll content(r = 0.777, p < 0.001). N chlorophyll contents in foliage were corre-lated with the depth of the A layer (r = 0.592 for N and 0.573 forchlorophyll, p < 0.002), and both were also positively correlatedwith the C stock in soil (r = 0.461 for N and 0.501 for chlorophyll,p < 0.05). Lignin and phenolic contents of foliage were not corre-lated with any of the other investigated foliage parameters. Ligninand phenolic contents were highest in the leaves of S. caprea,which was the dominant tree in the succession sites.
Earthworm density significantly differed among the forest sites(Fig. 2) and were highest in the alder sites and lowest in the succes-sion sites. Earthworm density was positively correlated with theproportion of earthworm casts in the soil profile as well as withthe depth of the A horizon and with the soil C stock (Table 2). Nem-atode abundance did not significantly differ among the sites (Ta-ble 4). The abundance of oribatid mites was highest in alder sitesand lowest in spruce sites (Table 4), while the abundance of Col-lembola was highest in spruce and lowest in larch sites. Numbersof nematodes and oribatid mites were negatively correlated withthe C/N ratio while the numbers of Collembola were positively cor-related with the C/N ratio and with fungal and bacterial biomass(Table 2).
Microbial respiration and microbial biomass showed oppositetrends among the sites (Table 4). Microbial biomass was highestin lime sites and lowest in succession sites, while respiration(and metabolic quotient) was highest in succession sites and low-est in lime sites; the latter pattern was also evident for metabolicquotient. Microbial respiration was positively correlated with soilC content while microbial respiration and metabolic quotient werenegatively correlated with C content (Table 2). Microbial biomasswas negatively correlated with the thickness of the Oe layer, whilemicrobial respiration was negatively correlated with the abun-dance of earthworm casts in the topsoil (Table 2). Factors that indi-cate soil bioturbation such as a high C content, a thin Oe layer, orthe abundance of worm casts were associated with increasedmicrobial biomass, and negatively correlated with metabolic quo-tient. Microbial biomass, respiration, and specific respiration werenot correlated with the fungal/bacterial (F/B) ratio based on PLFAs.F/B in the A horizon did not differ significantly among the sites ex-cept that F/B was higher at spruce sites than at other sites. F/B waspositively correlated with the C/N ratio of litter (Table 2). F/B wasgenerally higher in the Oe horizon, if that layer was present, thanin the A horizon (p = 0.0002, two-way ANOVA), but this differencewas significant only for spruce (Table 4).
With regard to the composition of the bacterial community asdetermined by DGGE and PCA, the second ordination axis closelycorresponded with parameters that reflected the presence or ab-sence of bioturbation such as earthworm density, presence ofworm casts in soil, or the thickness of the Oe layer (Fig. 3).
Because the formation of a humus horizon and the accumula-tion of C in the topsoil seem to be important factors that determinemany other soil biological parameters, we explored the relation-ship between A horizon thickness and topsoil C content vs. litterquality (as indicated by the C/N ratio) and bioturbation (as indi-
perties mediated by soil fauna? A case study from post-mining sites. Forest
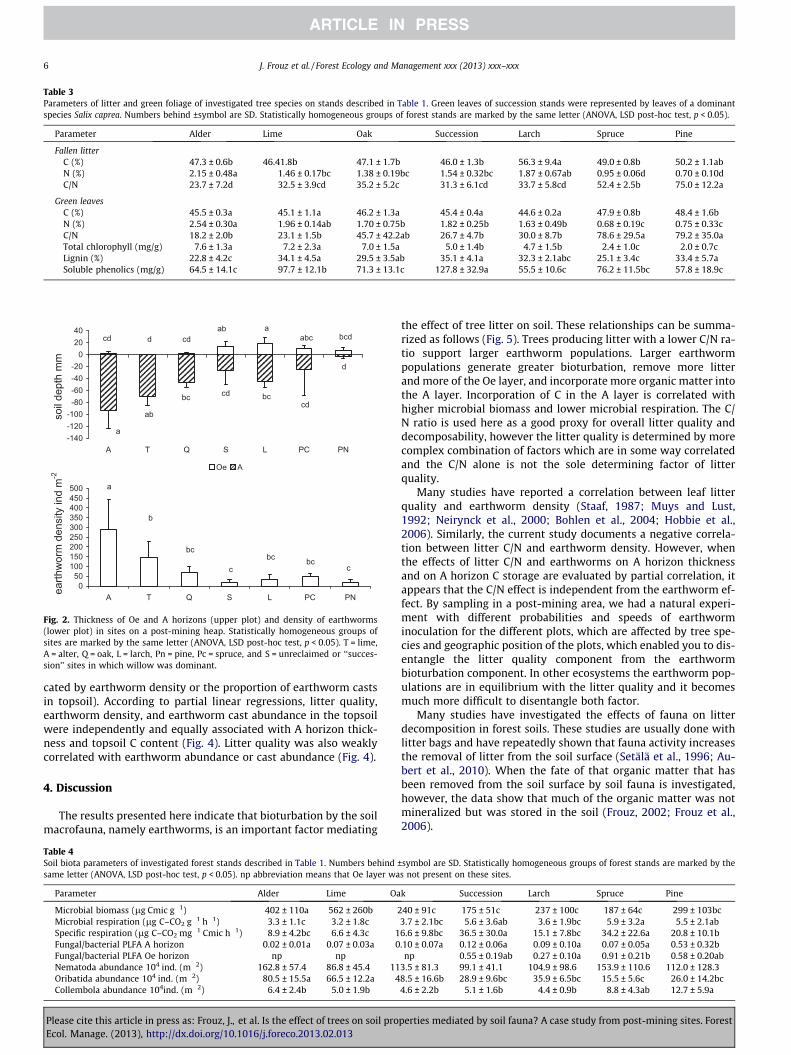
Table 3Parameters of litter and green foliage of investigated tree species on stands described in Table 1. Green leaves of succession stands were represented by leaves of a dominantspecies Salix caprea. Numbers behind ±symbol are SD. Statistically homogeneous groups of forest stands are marked by the same letter (ANOVA, LSD post-hoc test, p < 0.05).
Parameter Alder Lime Oak Succession Larch Spruce Pine
Fallen litterC (%) 47.3 ± 0.6b 46.41.8b 47.1 ± 1.7b 46.0 ± 1.3b 56.3 ± 9.4a 49.0 ± 0.8b 50.2 ± 1.1abN (%) 2.15 ± 0.48a 1.46 ± 0.17bc 1.38 ± 0.19bc 1.54 ± 0.32bc 1.87 ± 0.67ab 0.95 ± 0.06d 0.70 ± 0.10dC/N 23.7 ± 7.2d 32.5 ± 3.9cd 35.2 ± 5.2c 31.3 ± 6.1cd 33.7 ± 5.8cd 52.4 ± 2.5b 75.0 ± 12.2a
Green leavesC (%) 45.5 ± 0.3a 45.1 ± 1.1a 46.2 ± 1.3a 45.4 ± 0.4a 44.6 ± 0.2a 47.9 ± 0.8b 48.4 ± 1.6bN (%) 2.54 ± 0.30a 1.96 ± 0.14ab 1.70 ± 0.75b 1.82 ± 0.25b 1.63 ± 0.49b 0.68 ± 0.19c 0.75 ± 0.33cC/N 18.2 ± 2.0b 23.1 ± 1.5b 45.7 ± 42.2ab 26.7 ± 4.7b 30.0 ± 8.7b 78.6 ± 29.5a 79.2 ± 35.0aTotal chlorophyll (mg/g) 7.6 ± 1.3a 7.2 ± 2.3a 7.0 ± 1.5a 5.0 ± 1.4b 4.7 ± 1.5b 2.4 ± 1.0c 2.0 ± 0.7cLignin (%) 22.8 ± 4.2c 34.1 ± 4.5a 29.5 ± 3.5ab 35.1 ± 4.1a 32.3 ± 2.1abc 25.1 ± 3.4c 33.4 ± 5.7aSoluble phenolics (mg/g) 64.5 ± 14.1c 97.7 ± 12.1b 71.3 ± 13.1c 127.8 ± 32.9a 55.5 ± 10.6c 76.2 ± 11.5bc 57.8 ± 18.9c
-140-120-100
-80-60-40-20
02040
A T Q S L PC PN
soil
dept
h m
m
Oe A
050
100150200250300350400450500
A T Q S L PC PN
earth
wor
m d
ensi
ty in
d m
-2
a
b
bcbc bc
c c
aabc bcd
abcd cdd
a
ab
bc bccd
cd
d
Fig. 2. Thickness of Oe and A horizons (upper plot) and density of earthworms(lower plot) in sites on a post-mining heap. Statistically homogeneous groups ofsites are marked by the same letter (ANOVA, LSD post-hoc test, p < 0.05). T = lime,A = alter, Q = oak, L = larch, Pn = pine, Pc = spruce, and S = unreclaimed or ‘‘succes-sion’’ sites in which willow was dominant.
6 J. Frouz et al. / Forest Ecology and Management xxx (2013) xxx–xxx
cated by earthworm density or the proportion of earthworm castsin topsoil). According to partial linear regressions, litter quality,earthworm density, and earthworm cast abundance in the topsoilwere independently and equally associated with A horizon thick-ness and topsoil C content (Fig. 4). Litter quality was also weaklycorrelated with earthworm abundance or cast abundance (Fig. 4).
4. Discussion
The results presented here indicate that bioturbation by the soilmacrofauna, namely earthworms, is an important factor mediating
Table 4Soil biota parameters of investigated forest stands described in Table 1. Numbers behind ±same letter (ANOVA, LSD post-hoc test, p < 0.05). np abbreviation means that Oe layer wa
Parameter Alder Lime Oa
Microbial biomass (lg Cmic g�1) 402 ± 110a 562 ± 260b 2Microbial respiration (lg C–CO2 g�1 h�1) 3.3 ± 1.1c 3.2 ± 1.8cSpecific respiration (lg C–CO2 mg�1 Cmic h�1) 8.9 ± 4.2bc 6.6 ± 4.3c 1Fungal/bacterial PLFA A horizon 0.02 ± 0.01a 0.07 ± 0.03a 0Fungal/bacterial PLFA Oe horizon np npNematoda abundance 104 ind. (m�2) 162.8 ± 57.4 86.8 ± 45.4 11Oribatida abundance 104 ind. (m�2) 80.5 ± 15.5a 66.5 ± 12.2a 4Collembola abundance 104ind. (m�2) 6.4 ± 2.4b 5.0 ± 1.9b
Please cite this article in press as: Frouz, J., et al. Is the effect of trees on soil proEcol. Manage. (2013), http://dx.doi.org/10.1016/j.foreco.2013.02.013
the effect of tree litter on soil. These relationships can be summa-rized as follows (Fig. 5). Trees producing litter with a lower C/N ra-tio support larger earthworm populations. Larger earthwormpopulations generate greater bioturbation, remove more litterand more of the Oe layer, and incorporate more organic matter intothe A layer. Incorporation of C in the A layer is correlated withhigher microbial biomass and lower microbial respiration. The C/N ratio is used here as a good proxy for overall litter quality anddecomposability, however the litter quality is determined by morecomplex combination of factors which are in some way correlatedand the C/N alone is not the sole determining factor of litterquality.
Many studies have reported a correlation between leaf litterquality and earthworm density (Staaf, 1987; Muys and Lust,1992; Neirynck et al., 2000; Bohlen et al., 2004; Hobbie et al.,2006). Similarly, the current study documents a negative correla-tion between litter C/N and earthworm density. However, whenthe effects of litter C/N and earthworms on A horizon thicknessand on A horizon C storage are evaluated by partial correlation, itappears that the C/N effect is independent from the earthworm ef-fect. By sampling in a post-mining area, we had a natural experi-ment with different probabilities and speeds of earthworminoculation for the different plots, which are affected by tree spe-cies and geographic position of the plots, which enabled you to dis-entangle the litter quality component from the earthwormbioturbation component. In other ecosystems the earthworm pop-ulations are in equilibrium with the litter quality and it becomesmuch more difficult to disentangle both factor.
Many studies have investigated the effects of fauna on litterdecomposition in forest soils. These studies are usually done withlitter bags and have repeatedly shown that fauna activity increasesthe removal of litter from the soil surface (Setälä et al., 1996; Au-bert et al., 2010). When the fate of that organic matter that hasbeen removed from the soil surface by soil fauna is investigated,however, the data show that much of the organic matter was notmineralized but was stored in the soil (Frouz, 2002; Frouz et al.,2006).
symbol are SD. Statistically homogeneous groups of forest stands are marked by thes not present on these sites.
k Succession Larch Spruce Pine
40 ± 91c 175 ± 51c 237 ± 100c 187 ± 64c 299 ± 103bc3.7 ± 2.1bc 5.6 ± 3.6ab 3.6 ± 1.9bc 5.9 ± 3.2a 5.5 ± 2.1ab6.6 ± 9.8bc 36.5 ± 30.0a 15.1 ± 7.8bc 34.2 ± 22.6a 20.8 ± 10.1b.10 ± 0.07a 0.12 ± 0.06a 0.09 ± 0.10a 0.07 ± 0.05a 0.53 ± 0.32bnp 0.55 ± 0.19ab 0.27 ± 0.10a 0.91 ± 0.21b 0.58 ± 0.20ab
3.5 ± 81.3 99.1 ± 41.1 104.9 ± 98.6 153.9 ± 110.6 112.0 ± 128.38.5 ± 16.6b 28.9 ± 9.6bc 35.9 ± 6.5bc 15.5 ± 5.6c 26.0 ± 14.2bc4.6 ± 2.2b 5.1 ± 1.6b 4.4 ± 0.9b 8.8 ± 4.3ab 12.7 ± 5.9a
perties mediated by soil fauna? A case study from post-mining sites. Forest

Fig. 3. Ordination diagram of principal component analysis (PCA) based on DGGE ofthe bacterial communities in sites on a post-mining heap. Numbers by each axisrepresent the percentage of data variability explained by the axis. Arrows representenvironmental variables. Variables that correlate significantly with the first orsecond ordination axis are marked by solid lines, and the correlation coefficientfollowed by the number of the axis is indicated in parentheses. T = lime, A = alter,Q = oak, L = larch, Pn = pine, Pc = spruce, and S = unreclaimed or ‘‘succession’’ sitesin which willow was dominant.
J. Frouz et al. / Forest Ecology and Management xxx (2013) xxx–xxx 7
In the current study, microbial biomass increased and microbialrespiration decreased with increasing C content in the topsoil. Thisis in agreement with a previous study done on reclaimed and unre-claimed chronosequences at the same post-mining location (Helin-gerová et al., 2010). That microbial biomass increases withincreasing soil C has been observed by many other authors(Šantrucková, 1992; Baer et al., 2002; Bekku et al., 2004). Many re-ports also indicated that metabolic quotient decreased withincreasing C content (Insam and Domsch, 1988; Šantrucková,1992; Bekku et al., 2004; Frouz and Nováková, 2005; Šourkováet al., 2005b), or increasing mirobial biomass (Santruckova andStraskraba, 1991) but this decrease in metabolic quotient seldomled to a decrease in microbial respiration per unit soil mass as wasthe case in the current study. We except that, in addition to mecha-nism described above, bioturbation and consequent coating of organ-ic matter by clay particles make even large proportion of soilmicroflora inactive as already shown in other studies (Six et al., 2002).
The fungal/bacterial (F/B) ratio was positively correlated withthe C/N ratio, which was expected (Fierer et al., 2009), but the F/B ratio was not correlated with microbial respiration. The incorpo-ration of C into the A horizon seems to affect microbial biomass
Fig. 4. Partial correlation coefficients between litter C/N, earthworm abundance, or abundsites on a post-mining heap. The upper correlation coefficients are based on the abuabundance. Thick line mark correlation significant for p < 0.05.
Please cite this article in press as: Frouz, J., et al. Is the effect of trees on soil proEcol. Manage. (2013), http://dx.doi.org/10.1016/j.foreco.2013.02.013
and respiration more strongly than the F/B ratio. This can be ex-plained by the fact that the bioturbation creates quite differentconditions for litter decomposition. Also, comparison of the F/B ra-tios between Oe and A horizons and among sites but within indi-vidual horizons (Table 4) showed that bioturbation substantiallyaffected the F/B ratio. Without bioturbation, litter tends to remainon the soil surface, and organic matter enters the soil either as dis-solved organic matter or as small fragments that remain betweensoil aggregates. With bioturbation, the organic matter is removedfrom the soil surface and mixed into soil aggregates (Frouz et al.,2007a). The incorporation of organic matter in soil aggregates pro-tects organic matter and slows decomposition (Guggenbergeret al., 1996; Zhang et al., 2003; Bossuyt et al., 2005), which is con-sistent with the lower respiration and higher C storage at sites withgreater bioturbation (Frouz et al., 2009). Higher C storage in soil asa result of fauna activity was also reported by Wironen and Moore(2006), who observed higher soil C storage as a consequence of bio-turbation in North American forests affected by earthworm inva-sion. Soil invertebrates and earthworms in particular mayincrease the stability of soil organic matter (Wolters, 2000) bychanging the chemistry of the organic matter (Hopkins et al.,1998), by incorporating the organic matter into soil aggregates,and by physically binding the organic matter (Lavelle and Martin1992; Guggenberger et al., 1996; Six et al., 2002; Zhang et al.,2003; Bossuyt et al., 2005). On the other hand, the supply of easilydecomposable litter may support earthworm populations (Lavelleet al., 1997; Ponge, 2003). This may explain why soil C contentand soil C storage were highest and microbial respiration was low-est in sites with litter that had a low C/N ratio, i.e., in sites wherethe litter was presumably easily decomposed.
Besides being related to the C/N ratio of litter, the C storage intopsoil and the overall C stock were related to those characteristicsof living leaves that indicated high decomposability, i.e., a low C/Nratio and a high chlorophyll content. The correlation of overall Cstock with leaf chlorophyll content suggests that future researchshould explore the possibility of estimating soil carbon stock by re-mote sensing of foliage. Similar relationships have already been re-ported between soil water-extractable dissolved organic carbon,needle chlorophyll content, and laboratory leaf reflectance ofNorway spruce needles in the Krusne hory region (Albrechtováet al., 2008). Chlorophyll content is now commonly estimatedbased on leaf optical properties (Ustin et al., 2009), and leaf opticalproperties (airborne hyperspectral data) may be used for mappingthe health of forests (Mišurec et al., 2012).
In summary, the current study indicates that soil fauna on for-ested spoil heaps greatly change the organization of the topsoil.When soil fauna are not abundant, most organic matter residesin litter and in the Oe horizon. When soil fauna are abundant, mostorganic matter is moved into the mostly mineral A horizon. Thisshift substantially changes the conditions for microbial activityand may be an important driver of microbial activity. Similarly,the effect of earthworms on other soil fauna, such as negative
ance of earthworm casts and the thickness of the A horizon or C content in topsoil inndance of earthworm casts, and the lower coefficients are based on earthworm
perties mediated by soil fauna? A case study from post-mining sites. Forest

Fig. 5. Hypothetical conceptual model describing the relationships between thetree effect and fauna effect on soil properties. When the fauna effect is small, littertends to remain on the soil surface, the Oe horizon is thick, the A horizon is thin,microbial respiration is high, and soil C storage is low. When the fauna effect islarge, litter tends to be transported into the soil, the Oe horizon is thin, the Ahorizon is thick, microbial respiration is low, and soil C storage is high.
8 J. Frouz et al. / Forest Ecology and Management xxx (2013) xxx–xxx
correlations with collembolan density, is consistent with labora-tory experiments showing the reduction of collembolans whenearthworms are present (Mudrák et al., 2010).
It follows that the greatest effect of trees on soil may be indirect,i.e., may result from the effect of tree litter on the bioturbationactivity of soil fauna. Bioturbation changes not only the biomassand activity of microorganisms but also the composition of the soilmicrobial community (Fig. 3) and other soil fauna. The effect ofearthworms on the microbial community has been documentedmany times in short-term laboratory experiments (Frouz et al.,2006, 2007b), but less often in the field.
Acknowledgements
This study was supported by the Czech Science Foundation(Grant No. P504/12/1288); the Ministry of Education, Youth andSports of the Czech Republic (No. LC 06066); and by the Sokolovskáuhelná a.s. coal mining company. We thank Mgr. Drahomíra
Please cite this article in press as: Frouz, J., et al. Is the effect of trees on soil proEcol. Manage. (2013), http://dx.doi.org/10.1016/j.foreco.2013.02.013
Bartáková and Mgr. Monika Kovárová for help with chlorophylland phenolic extractions. We wish to thank Dr. B. Jaffee (JaffeeRe-vises, Bellingham, USA) for reading the manuscript and for lan-guage corrections.
References
Albrechtová, J., Seidl, Z., Aitkenhead-Peterson, J., Lhotáková, Z., Rock, B.N.,Alexander, J.E., Malenovsky, Z., McDowell, W.H., 2008. Spectral analysis ofconiferous foliage and possible links to soil chemistry: are spectral chlorophyllindices related to forest floor dissolved organic C and N? Sci. Total. Environ. 404,424–432.
Angers, A.D., Caron, J., 1998. Plant-induced changes in soil structure: processes andfeedbacks. Biogeochemistry 42, 55–72.
Aubert, M., Margerie, P., Trap, J., Bureau, F., 2010. Aboveground–belowgroundrelationships in temperate forests: plant litter composes and microbiotaorchestrates. For. Ecol. Manage. 259, 563–572.
Baer, S.G., Kitchen, D.J., Blair, J.M., Rice, C.W., 2002. Changes in ecosystem structureand function along a chronosequence of restored grasslands. Ecol. Appl. 12,1688–1701.
Bekku, Y.S., Nakatsubo, T., Kume, A., Koizumi, H., 2004. Soil microbial biomass,respiration rate, and temperature dependence on a successional glacier forelandin Ny-Alesund. Svalbard. Arctic Alpine Res. 36, 395–399.
Bohlen, P.J., Pelletier, D.M., Groffman, P.M., Fahey, T.J., Fisk, M.C., 2004. Influence ofearthworm invasion on redistribution and retention of soil carbon and nitrogenin northern temperate forests. Ecosystems 7, 13–27.
Bossuyt, H., Six, J., Hendrix, P.F., 2005. Protection of soil carbon by microaggregateswithin earthworm casts. Soil Biol. Biochem. 37, 251–258.
Bradshaw, A., 1997. Restoration of mined landside using natural processes. Ecol.Eng. 8, 255–269.
Brady, N.C., Weil, R.R., 2008. The Nature and Properties of Soil, 14th ed. PrenticeHall, Upper Saddle River.
Chronáková, A., Ascher, J., Jirout, J., Ceccherini, M.T., Elhottová, D., Pietramellara, G.,Šimek, M., 2012. Cattle impact on composition of archaeal, bacterial, and fungalcommunities by comparative fingerprinting of total and extracellular DNA. Biol.Fertil. Soils. http://dx.doi.org/10.1007/s00374-012-0726-x.
De Schrijver, A., DeFrenne, P., Staelens, J.E., Verstraeten, G., Muys, B., Vesterdal, L.,Wuyts, K., Van Nevel, L., Schelfhout, S.E., De Neves, S., Verheyen, K., 2011. Treespecies traits cause divergence in soil acidification during four decades ofpostagricultural forest development. Global Change Biol. 18, 1127–1140.
Fierer, N., Strickland, M.S., Liptzin, D., Bradford, M.A., Cleveland, C.C., 2009. Globalpatterns in belowground communities. Ecol. Lett. 12, 1–12.
Frouz, J., 2002. The effect of soil macrofauna on litter decomposition and soilorganic matter accumulation during soil formation in spoil heaps after browncoal mining: a preliminary result. Ekologia (Bratisl) 21, 363–369.
Frouz, J., Kalcík, J., 2006. Accumulation of soil organic carbon in relation to other soilcharacteristic during spontaneous succession in non reclaimed colliery spoilheaps after brown coal mining near Sokolov (the Czech Republic) Ekológia 25,pp. 388–397.
Frouz, J., Nováková, A., 2005. Development of soil microbial properties in top soillayer during spontaneous succession in heaps after brown coal mining inrelation to soil microstructure development. Geoderma 129, 54–64.
Frouz, J., Keplin, B., Pizl, V., Tajovsky, K., Stary, J., Lukešová, A., Nováková, A., Balík, V.,Hánel, L., Materna, J., Düker, C., Chalupsky, J., Rusek, J., Heinkele, T., 2001. Soilbiota and upper soil layers development in two contrasting post-miningchronosequences. Ecol. Eng. 17, 275–284.
Frouz, J., Elhottová, D., Kuráz, V., Šourková, M., 2006. Effect of soil macrofauna onother soil biota and soil formation in reclaimed and unreclaimed post-miningsites: result of field microcosm experiment. Appl. Soil Ecol. 33, 308–320.
Frouz, J., Elhottová, D., Pizl, V., Tajovsky, K., Šourková, M., Picek, T., Maly, S., 2007a.The effect of litter quality and soil faunal composition on organic matterdynamics in post-mining soil: a laboratory study. Appl. Soil Ecol. 37, 72–80.
Frouz, J., Pizl, V., Tajovsky, K., 2007b. The effect of earthworms and othersaprophagous macrofauna on soil microstructure in reclaimed and un-reclaimed post-mining sites in Central Europe. Eur. J. Soil. Biol. 43, 184–189.
Frouz, J., Prach, K., Pizl, V., Hánel, L., Stary, J., Tajovsky, K., Materna, J., Balík, V.,Kalcík, J., Rehounková, K., 2008a. Interactions between soil development,vegetation and soil fauna during spontaneous succession in post-mining sites.Eur. J. Soil Biol. 44, 109–121.
Frouz, J., Lobinske, R., Kalcík, J., Ali, A., 2008b. Effects of the exotic crustacean,Armadillidium vulgare (isopoda), and other macrofauna on organic matterdynamics in soil microcosms in a hardwood forest in central Florida. FloridaEntomol. 91 (2), 328–331.
Frouz, J., Cienciala, E., Pizl, V., Kalcík, J., 2009. Carbon storage in post-mining forestsoil, the role of tree biomass and soil bioturbation. Biogeochemistry 94, 111–121.
Frouz, J., Kalcík, J., Velichová, V., 2011. Factors causing spatial heterogeneity in soilproperties, plant cover, and soil fauna in a non-reclaimed post-mining site. Ecol.Eng. 11, 1910–1913.
Guggenberger, G., Thomas, R.J., Zech, W., 1996. Soil organic matter withinearthworm casts of an anecic–endogeic tropical pasture community.Colombia Appl. Soil Ecol. 3, 263–274.
Hánel, L., 1995. Secondary successional stages of soil nematodes in cambisols ofSouth Bohemia. Nematologica 41, 197–218.
perties mediated by soil fauna? A case study from post-mining sites. Forest

J. Frouz et al. / Forest Ecology and Management xxx (2013) xxx–xxx 9
Helingerová, M., Frouz, J., Šantrucková, H., 2010. Microbial activity in reclaimed andunreclaimed post-mining sites near Sokolov (Czech Republic). Ecol. Eng. 36 (6),768–776.
Hobbie, S.E., Reich, P.B., Oleksyn, J., Ogdahl, M., Zytkowiak, R., Hale, C., Karolewski,P., 2006. Tree species effects on decomposition and forest floor dynamics in acommon garden. Ecology 87, 2288–2297.
Hopkins, D.W., Chudek, J.A., Bignell, D.E., Frouz, J., Webster, E.A., Lawson, T., 1998.Application of 13C NMR to investigate the transformations and biodegradationof organic materials by some soil and litter-dwelling insects. Biodegradation 9,423–431.
Insam, H., Domsch, K.H., 1988. Relationship between soil organic carbon andmicrobial biomass on chronosequences of reclamation sites. Microb. Ecol. 15,177–188.
Jangida, K., Williams, M.A., Franzluebbers, A.F., Schmidt, T.M., Coleman, D.C.,Whitman, W.B., 2011. Land-use history has a stronger impact on soil microbialcommunity composition than aboveground vegetation and soil properties. SoilBiol. Biochem. 43, 2184–2193.
Kríbek, B., Strnad, M., Bohácek, Z., Sykorová, I., Cejka, J., Sobalík, Z., 1998.Geochemistry of Miocene lacustrine sediments from the Sokolov Coal Basin(Czech Republic). Int. J. Coal Geol. 37, 207–233.
Lal, R., 2004. Soil carbon sequestration to mitigate climate change. Geoderma 123,1–22.
Lavelle, P., Martin, A., 1992. Small-scale and large-scale effect of endogeicearthworms on soil organic matter dynamics in soil of humid tropics. SoilBiol. Biochem. 24, 1491–1498.
Lavelle, P., Bignell, D., Lepage, M., Wolters, V., Rogers, P., Ineson, P., Heal, O.W.,Dhillion, S., 1997. Soil function in changing world: the role of invertebrateecosystem engineers. Eur. J. Soil Biol. 33, 159–193.
Mišurec, J., Kopácková, V., Lhotaková, Z., Hanuš, J., Weyermann, J., Entcheva-Campbell, P., Albrechtová, J., 2012. Utilization of hyperspectral image opticalindices to assess the Norway spruce forest health status. J. Appl. Remote Sens. 6,063545. http://dx.doi.org/10.1117/1.JRS.6.063545.
Mudrák, O., Frouz, J., Velichová, V., 2010. Understory vegetation in reclaimed andunreclaimed post-mining forest stands. Ecol. Eng. 36 (6), 783–790.
Murphy, J., Riley, J.P., 1962. A modified single solution method for thedetermination phosphate in natural waters. Anal. Chim. Acta 27, 31–36.
Muys, B., Lust, N., 1992. Inventory of the earthworm communities and the state oflitter decomposition in the forests of Flanders, Belgium, and its implications forforest management. Soil Biol. Biochem. 24, 1677–1681.
Neirynck, J., Mirtcheva, S., Sioen, G., Lust, N., 2000. Impact of Tilia platyphyllos Scop,Fraxinus excelsior L., Acer pseudoplatanus L., Quercus robur L. and Fagus sylvaticaL. on earthworm biomass and physical–chemical properties of a loamy topsoil.For. Ecol. Manage. 133, 275–286.
Odum, E.P., 1969. The strategy of ecosystem development. Science 164, 262–270.Ponge, J.F., 2003. Humus form in terrestrial ecosystem: a framework to biodiversity.
Soil Biol. Biochem. 35, 935–945.Porra, R.J., Thompson, W.A., Kriedemann, P.E., 1989. Determination of accurate
extinction coefficients and simultaneous equations for assaying chlorophylls aand b extracted with four different solvents: verification of the concentration ofchlorophyll standarts by atomic absorption spectroscopy. Biochem. Biophys.Acta 975, 384–394.
Sampedro, I., Giubilei, M., Cajthaml, T., Federici, E., Federici, F., Petruccioli, M.,D’annibale, A., 2009. Short-term impact of dry olive mill residue addition to soilon the resident microbiota. Bioresour. Technol. 100, 6098–6106.
Please cite this article in press as: Frouz, J., et al. Is the effect of trees on soil proEcol. Manage. (2013), http://dx.doi.org/10.1016/j.foreco.2013.02.013
Šantrucková, H., 1992. Microbial biomass, activity and soil respiration in relation tosecondary succession. Pedobiologia 36, 341–350.
Santruckova, H., Straskraba, M., 1991. On the relationship between specificrespiration activity and microbial biomass in soils. Soil Biol. Biochem. 23,525–532.
Schimel, D.S., Braswell, B.H., Holland, E.A., McKeown, R., Ojima, D.S., Painter, T.H.,Parton, W.J., Townsend, A.R., 1994. Climatic, edaphic, and biotic controls overstorage and turnover of carbon in soils. Global Biogeochem. Cycles 8, 279–293.
Scullion, J., Malik, A., 2000. Earthworm activity affecting organic matter, aggregationand microbial activity in soils restored after opencast mining for coal. Soil Biol.Biochem. 32, 119–126.
Setälä, H., Marshall, V.G., Trofymow, J.A., 1996. Influence of body size of soil faunaon litter decomposition and 15N uptake by Poplar in a pot trial. Soil Biol.Biochem. 28, 1661–1675.
Singleton, V.L., Orthofer, R., Lamuela-Raventós, R.M., 1998. Analysis of total phenolsand other oxidation substrates and antioxidants by means of folin–ciocalteureagent. Methods Enzymol 299, 152–178.
Six, J., Conanat, R.T., Paul, E.P., Paustian, K., 2002. Stabilisation mechanism of soilorganic mater: implication for C saturation of soil. Plant Soil 241, 155–176.
Šnajdr, J., Valášková, V., Merhautová, V., Herinková, J., Cajthaml, T., Baldrian, P.,2008. Spatial variability of enzyme activities and microbial biomass in theupper layers of Quercus petraea forest soil. Soil Biol. Biochem. 40, 2068–2075.
Šourková, M., Frouz, J., Šantrucková, H., 2005a. Accumulation of carbon, nitrogenand phosphorus during soil formation on alder spoil heaps after brown-coalmining, near Sokolov (Czech Republic). Geoderma 124, 203–214.
Šourková, M., Frouz, J., Fettweis, U., Bens, O., Hüttl, R.F., Šantrucková, H., 2005b. Soildevelopment and properties of microbial biomass succession in reclaimed post-mining sites near Sokolov (Czech Republic) and near Cottbus (Germany).Geoderma 129, 73–80.
Staaf, H., 1987. Foliage litter turnover and earthworm populations in three beechforests. Oecologia 72, 58–64.
Ustin, S.L., Gitelson, A.A., Jacquemoud, S., Schaepman, M., Asner, G.P., Gamon, J.A.,Zarco-Tejada, P., 2009. Retrieval of foliar information about plant pigmentsystems from high resolution spectroscopy. Remote Sens. Environ. 113, S67–S77.
Vance, E.D., Brookes, P.C., Jenkinson, D.S., 1987. An extraction method for measuringsoil microbial biomass C. Soil Biol. Biochem. 19, 703–707.
Vivanco, L., Austin, A.T., 2008. Tree species identity alters forest litter decompositionthrough long-term plant and soil interactions in Patagonia, Argentina. J. Ecol.96, 727–736.
Wardle, D.A., 2006. The influence of biotic interactions on soil biodiversity. Ecol.Lett. 9, 870–886.
Wellburn, A.R., 1994. The spectral determination of chlorophylls a and b, as well astotal carotenoids, using various solvents with spectrophotometers of differentresolution. J. Plant Physiol. 44, 307–313.
Wironen, M., Moore, T.R., 2006. Exotic earthworm invasion increases soil carbonand nitrogen in an old-growth forest in southern Quebec. Can. J. For. Res. 36,845–854.
Wolters, V., 2000. Invertebrate control of soil organic matter stability. Biol. Fertil.Soils 31, 1–19.
Zhang, X., Wang, J., Xie, H., Wang, J., Zech, W., 2003. Comparison of organiccompounds in the particle-size fractions of earthworm casts and surroundingsoil in humid Laos. Appl. Soil Ecol. 23, 147–153.
perties mediated by soil fauna? A case study from post-mining sites. Forest