investigations of the chymotrypsin-catalyzed hydrolysis of ... · hypothesis that the ph dependence...
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEM~~Y Vol. 242, No. 5, Issue of March 10, PP. 919-929, 1967
P&ted in U.S.A.
Investigations of the Chymotrypsin-catalyzed Hydrolysis of Specific Substrates
I. THE pH DEPENDENCE OF THE CATALYTIC HYDROLYSIS OF N-ACETYL-L-TRYPTOPHANAMIDE BY THREE FORMS OF THE ENZYME AT ALKALINE pH*
(Received for publication, July 7, 1966)
ALBERT HIMOE,~ PAUL C. PARKS,~ AND GEORGE P. HESS~
From the Xection of Biochemistry and Molecular Biology, Division of Biological Sciences, Cornell University, Ithaca, New York 14850
SUMMARY
The pH dependence of the steady state kinetic parameters K,(app) and $,t of the a-, acetylated 6-, and ar-chymotrypsin- catalyzed hydrolysis of the specific amide substrate N-acetyl- r.-tryptophanamide, has been investigated in the neutral and alkaline pH regions. A completely automatic technique for measuring the hydrolysis of amides was used. New results and important aspects of the chymotrypsin-catalyzed hy- drolysis of a specific amide substrate which have emerged from this investigation are the following.
The pH dependence of the catalytic reaction at alkaline pH is adequately accounted for by the pH dependence of K,(app). In earlier experiments we have shown that in amide hydrolysis the steady state kinetic parameter K,(app) is a measure of an over-all enzyme-substrate dissociation constant (K’,). (a) K,(app) was observed to be pH-de- pendent in the hydrolysis of N-acetyl-r&yptophanamide as catalyzed by all three forms of the enzyme. Analysis of this pH dependence shows that an ionizing group with pK(app) - 8.5 is involved; an ionizing group of the enzyme with this pK(app) has been implicated in all chymotrypsin-catalyzed reactions that have been studied. (b) The catalytic rate constant kat was found to be pa-independent in the pH region 8 to 10 for the &chymotrypsin-catalyzed reaction and in the pH region 8 to 9.2 for the a-chymotrypsin-catalyzed reaction. (c) The pH dependence of the steady state kinetic parameters kcat and K,(app) in the chymotrypsin-catalyzed hydrolysis of iV-acetyl-L-tryptophanamide is consistent with the findings of our earlier studies of the individual steps in the reaction between chymotrypsin and diisopropyl fluoro- phosphate and in the chymotrypsin-catalyzed hydrolysis of N-acetyl-L-phenylalaninamide: studies which indicated that the pH dependence of chymotrypsin-catalyzed hydrolysis above pH 8 is due to the effect of pH on the formation of
* We are grateful to the National Institutes of Health and the National Science Foundation for financial support.
1 United States Public Health Service Postdoctoral Trainee. 5 National Institutes of Health Postdoctoral Fellow. f[ To whom reprint requests should be addressed at 210 Savage
Hall.
chymotrypsin complexes and not on the bond-breaking step. (d) The experiments reported here are not in agreement with earlier studies of Bender and his co-workers, which indicated that the pH dependence of chymotrypsin-catalyzed reactions above pH 8 is due to the effect of pH on the bond- breaking step, the formation of chymotrypsin-substrate complexes being pa-independent.
The 6 form of chymotrypsin is a more efficient catalyst than the (Y form in the hydrolysis of N-acetyl-L-tryptophanamide, as indicated by both the observed maximum rate of reaction and the binding of substrate to enzyme. Both the values and the pH dependencies of the steady state kinetic parameters are different for these two forms of the enzyme, but are the same for 6- and acetylated b-chymotrypsin. Only limited and qualified information regarding differences in the catalytic properties of a- and d-chymotrypsin has been previously available.
These and previous experiments are consistent with the hypothesis that the pH dependence of chymotrypsin-cata- lyzed reactions at alkaline pH is due to a pH-dependent equilibrium between two major conformations of the enzyme, with the same ionizing group of pK (app) - 8.5 controlling both the equilibrium between enzyme conformations and the decrease in catalytic properties of the enzyme. This hypothe- sis leads to several predictions, discussed in this paper, which can be tested experimentally.
Only limited and qualified information regarding chymo- trypsin-catalyzed reactions at alkaline pH is available (1). It is known that the catalytic properties of the enzyme toward both specific and model substrates decrease above pH 8 (2-4). Steady state kinetic experiments with cY-chymotrypsin and specific substrate esters and amides have indicated that an ionizing group of the enzyme with pK(app) - 8.5 is implicated in this decrease of enzymic activity (5, 6). Recent steady state kinetic experiments have led to the conclusion (5-9) that the pH de- pendence of chymotrypsin-catalyzed reactions at alkaline pH values is due to the effect of hydrogen ion concentration on the
919
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

920 Chymotrypsin-catalyzed Hydrolysis of Specific Substrates. 1 Vol. 242, No. 5
bond-breaking step and that the formation of enzyme-substrate complexes is pH-independent. However, this conclusion ap- peared questionable in view of investigations (10-22) of the individual steps in the stoichiometric reaction of chymotrypsin with its specific inhibitor DFPl and in the chymotrypsin-cata- Iyzed hydrolysis of N-acetyl-L-phenylalaninamide. It has been shown that the chymotrypsin-DFP reaction is similar to chymo- trypsin-catalyzed hydrolysis of specific substrates in that its reaction rate also decreases above pH 8 (10, 20, 23), with implica- tion in this decrease of an ionizing group of the enzyme of
ok - 9 (20). Kinetic studies of individual steps in the DFP reaction (19, 20), made through measurements of phos- phorylation rate and changes in the spectral and optical rotatory properties of the enzyme (10-19) suggested, however, a different mechanism for the pH dependence above pH 8 than the one proposed (5-9) for substrate hydrolysis. The DFP studies indicated that the observed decrease in phosphorylation rate above pH 8 is due to the effect of pH on a chymotrypsin-inhibitor complex which is formed by a reversible process which precedes the phosphorylation step. A similar effect of pH on enzyme- substrate complex formation was deduced from studies of in- dividual steps in the chymotrypsin-catalyzed hydrolysis of N-acetyl-n-phenylalaninamide (22). The nature of this pH effect has been suggested by experiments (17, 21) with acetylated &chymotrypsin which suggest that the level of pH affects an equilibrium between active and inactive conformations of the enzyme, and thereby the formation of enzyme complexes and the catalytic properties of the enzyme.
Because of these experimental findings, we are now conducting an investigation of chymotrypsin-catalyzed reactions of specific substrates at alkaline pH values. As a first step in these studies, we have measured the pH dependence of the steady state kinetic parameters, K,(app) and kcat, of the 6- and acetylated d-chymo- trypsin-catalyzed hydrolysis of the specific amide substrate ATA,’ and have reinvestigated the previously studied (6) (Y- chymotrypsin-catalyzed hydrolysis of the same substrate. Acetylated &chymotrypsin was used in this work because of previous evidence (21) that this enzyme exhibits, above pH 8, a pH-dependent equilibrium between two major conformations; the c+ and &chymotrypsin were used for purposes of comparison. An amide was chosen as specific substrate because recent experi- ments (24) indicated that in the chymotrypsin-catalyzed hy- drolysis of amides, but not esters, K,(app) is a direct measure of the enzyme-substrate equilibrium constants. Observation of a pH dependence of K,(app) in the hydrolysis of ATA would therefore indicate a pH dependence in the formation of enzyme- substrate complexes.
The measurements reported in this study were greatly facili- tated by the development of a completely automatic technique for measuring the hydrolysis of amides (25), a technique which as far as enzyme concentration, precision of measurement, and time involved are concerned, compares favorably with the pa-stat method used for measuring the kinetics of ester hydrolysis. Steady state kinetic parameters of 6- or acetylated b-chymo- trypsin-catalyzed hydrolysis of a specific amide substrate have not been reported prior to this study. A preliminary report of a part of this investigation has appeared (26).
1 The abbreviations used are: DFP, diisopropyl fluorophos- phate; ATA, N-acetyl-n-tryptophanamide.
EXPERIMENTAL PROCEDURE
Materials
Three times crystallized chymotrypsinogen (Lots 703, 781, and 8936) and three times crystallized, salt-free cr-chymotrypsin were obtained from Worthington. The &chymotrypsin was prepared just prior to each kinetic run by activating chymo- trypsinogen under conditions that are known to lead essentially to the 6 form of the enzyme (27). The ar-chymotrypsin and some of the &chymotrypsin preparations were passed through Sephadex columns before use, as suggested by Yapel et al. (28). The procedure was to dissolve 100 mg of enzyme in approximately 10 ml of cold 1 mM HCl solution, pass the solution through a Sephadex (G-25 coarse) column (90 X 2.5 cm) which had been equilibrated with 1 mu HCl, elute the sample with 1 mM HCl in a cold room (-4”), and collect the first fraction emerging from the column for use in the experiments. No difference in enzyme properties was noted between b-chymotrypsin samples which had been passed through Sephadex columns and those which had not.
Acetylated &chymotrypsin was prepared by acetylating chymotrypsinogen and then activating it, as described pre- viously (21). The product was examined by quantitative end group determination, Van Slyke amino nitrogen analysis, and spectrophotometric titration of free tyrosyl residues, which indicated that the material was &chymotrypsin with all e-amino groups of lysine and an average of 1.5 of the 2 titratable tyrosine residues acetylated. Deacetylation of tyrosyl residues was ac- complished by bringing a cold acetylated &chymotrypsin prep- aration (5 mg per ml) to pH 11.5 by addition of 0.2 M KOH in a pH-stat, and allowing the solution to stand at room temperature for 3 hour. The solution was then adjusted to pH 5.3 in an ice bath, and diluted for subsequent kinetic experiments. Several of these preparations were checked both spectrophotometrically and by Van Slyke amino nitrogen analysis to make certain that all tyrosyl residues but none of the amino groups had been un- blocked. All the analytical procedures used for checking the acetylated d-chymotrypsin preparations have been described in detail (21).
Protein concentrations were determined spectrophotometri- tally at 280 rnp; molar extinction coefficients used were 5 x lo4 M-I cm-r for unacetylated enzyme (29), and 4.7 X lo4 M-I cm-l for the acetylated derivatives (21). A molecular weight of 25,000 was assumed (30). Concentration of active enzyme was determined by the N-trans-cinnamoyl imidazole method (31). These determinations were made with aliquots of the enzyme stock solutions used for the kinetic experiments.
N-Acetyl-L-tryptophanamide, chromatographically pure, with melting point of 192-193” and [o(]t3 = +19.1 + 1.2”, was ob- tained as Lot 4739 from Cycle. In some instances, the material was recrystallized from methanol-ether-petroleum ether. N- trans-Cinnamoyl imidazole, with melting point of 133-134”, was prepared from cinnamoyl chloride and imidazole (reagent grade, Eastman) by the method of Schonbaum, Zerner, and Bender (31). Ninhydrin was obtained from Nutritional Biochemicals, and ninhydrin stock solutions were prepared according to a modification of the method of Moore and Stein (32). The preparation of the ninhydrin solutions used in the kinetic experi- ments has been described (25). Sephadex G-25 (coarse) was obtained from Pharmacia. All other chemicals were reagent grade and obtained from Mallinckrodt.
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Issue of March 10, 1967 Himoe, Parks, and Hess 921
Bu$er Compositims-(a) In all experiments of Fig. 2, and in all experiments of Figs. 3 and 4 except those characterized by IJ, the final buffer compositions were as follows: pH 5.7 to 8, 0.02 M
potassium phosphate buffer; pH 8 to 9.2, 0.02 M Tris-HCl buffer; pH 9.2 to 10.5, 0.02 M potassium carbonate buffer. Sufficient KC1 was added to all these buffers to give an ionic strength of 0.16 M. (b) In the experiments of Figs. 3 and 4 characterized by q , and in all experiments of Fig. 5, the final buffer compositions
were as follows: pH 5.7 to 8.3, 0.1 M potassium phosphate-O.1 M
KCl; pH 9 to 10.5, 0.1 M potassium carbonate-O.1 M KCl. ilbove pH 10.5, mixtures of 0.1 M potassium phosphate and
0.1 M KOH were used as buffers.
Methods and Apparatus
JReasurement of pH--A Radiometer pH meter, type TTTI, with type B electrode, was used for all the pH determinations. Measurements were made with reference to Beckman standard buffers of pH values within 1 pH unit of the sample solutions.
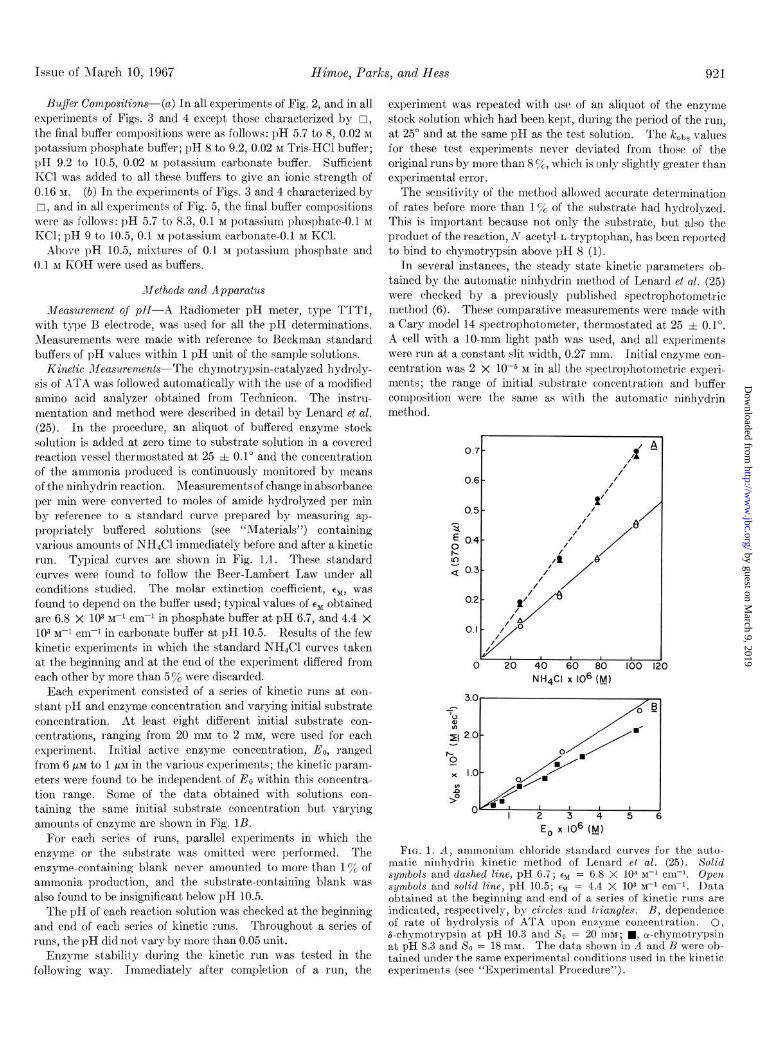
Kinetic JIeasurements-The chymotrypsin-catalyzed hydroly- sis of ATA was followed automatically with the use of a modified amino acid analyzer obtained from Technicon. The instru- mentation and method were described in detail by Lenard et al. (25). In the procedure, an aliquot of buffered enzyme stock solution is added at zero time to substrate solution in a covered reaction vessel thermostated at 25 f 0.1” and the concentration of the ammonia produced is continuously monitored by means of the ninhydrin reaction. Measurements of change in absorbance per min were converted to moles of amide hydrolyzed per min by reference to a standard curve prepared by measuring ap- propriately buffered solutions (see “Materials”) containing various amounts of NH&l immediately before and after a kinetic run. Typical curves are shown in Fig. IA. These standard curves were found to follow the Beer-Lambert Law under all conditions studied. The molar extinction coefficient, enb, was found to depend on the buffer used; typical values of ehl obtained are 6.8 x 103 M-’ cm-r in phosphate buffer at pH 6.7, and 4.4 x 103 M-I cm+ in carbonate buffer at pH 10.5. Results of the few kinetic experiments in which the standard NH&l curves taken at the beginning and at the end of the experiment differed from each other by more than 5% were discarded.
Each experiment consisted of a series of kinetic runs at con- stant pH and enzyme concentration and varying initial substrate concentration. At least eight different initial substrate con- centrations, ranging from 20 mM to 2 mM, were used for each experiment. Initial active enzyme concentration, Eo, ranged from 6 PM to 1 PM in the various experiments; the kinetic param- eters were found to be independent of EO within this concentra- tion range. Some of the data obtained with solutions con- taining the same initial substrate concentration but varying amounts of enzyme are shown in Fig. 1B.
For each series of runs, parallel experiments in which the enzyme or the substrate was omitted were performed. The enzyme-containing blank never amounted to more than 1 y0 of ammonia production, and the substrate-containing blank was also found to be insignificant below pH 10.5.
The pH of each reaction solution was checked at the beginning and end of each series of kinetic runs. Throughout a series of runs, the pH did not vary by more than 0.05 unit.
Enzyme stability during the kinetic run was tested in the following way. Immediately after completion of a run, the
experiment was repeated with use of an aliquot of the enzyme stock solution which had been kept, during the period of the run, at 25” and at the same pH as the test solution. The kobs values for these test experiments never deviated from those of the original runs by more than 8 ojg, which is only slightly greater than experimental error.
The sensitivity of the method allowed accurate determination of rates before more than 1% of the substrate had hydrolyzed. This is important because not only the substrate, but also the product of the reaction, N-acetyl-L-tryptophan, has been reported to bind to chymotrypsin above pH 8 (1).
Tn several instances, the steady state kinetic parameters ob- tained by the automatic ninhydrin method of Lenard et al. (25) were checked by a previously published spectrophotometric method (6). These comparative measurements were made with a Cary model 14 spectrophotometer, thermostated at 25 5 0.1”. A cell with a IO-mm light path was used, and all experiments were run at a constant slit width, 0.27 mm. Initial enzyme con- centration was 2 X low5 M in all the spectrophotometric experi- ments; the range of initial substrate concentration and buffer composition were the same as with the automatic ninhydrin method.
NH4CI x IO6 (11)
J 6 E, x IO6 (Ml
FIG. 1. A, ammonium chloride standard curves for the auto- matic ninhydrin kinetic method of Lenard et al. (25). Solid symbols and dashed line, pH 6.7; tM = 6.8 X 103 M-’ cm+. Open symbols and solid line, pH 10.5; Q, = 4.4 X 103 M-I cm-‘. Data obtained at the beginning and end of a series of kinetic runs are indicated, respectively, by circles and triangles. B, dependence of rate of hydrolysis of ATA upon enzyme concentration. 0, &chymotrypsin at pH 10.3 and So = 20 mM; n , cu-chymotrypsin at pH 8.3 and 5’0 = 18 mM. The data shown in A and B were ob- tained under the same experimental conditions used in the kinetic experiments (see “Experimental Procedure”).
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
922 Chymotrypsin-catalyyxed Hydrolysis of Specific Substrates. I Vol. 242, No. 5
The steady state kinetic constants were calculated from the rate data by means of a digital computer program written for the Lineweaver-Burk form of the Michaelis-Menten rate equation (33). Data weighting and the calculation of the standard errors of the kinetic constants were performed as discussed by Wilkinson (34). The kobs values were obtained by dividing u,bs by Eo. The E. was determined by the N-trans-cinnamoyl imidazole method (31). For illustrative purposes, some of the data were plotted according to the method of Eadie (35).
Deacylation of trans-Cinnamoyl Chymotrypsin-The rate of deacylation of trans-cinnamoyl o(- or &chymotrypsin at 25” was carried out as described by Bender, Schonbaum, and Zerner (36). The rate of reaction was followed by observing both the forma- tion of trans.cinnamic acid at 260 rnp and the disappearance of trans-cinnamoyl chymotrypsin at 310 mp. The trans-cinnamoyl chymotrypsin was generated in situ by addition of a small amount, 50 ~1, of trans-cinnamoyl imidazole in acetonitrile to 2.7 ml of a chymotrypsin solution in 1 mM acetate buffer of pH 4.3, to give a final concentration of 1.1 X 10e5 M trans-cinnamoyl imidazole and 1.2 X low5 M enzyme. Deacylation was started by addition of 0.3 ml of concentrated buffer to the reaction mixture. First order rate constants were calculated from the recorder traces by the method of Guggenheim (37).
RESULTS
A comparison of the rate data obtained by the automatic ninhydrin method of Lenard et al. (25) and the spectrophoto-
-I
k .,bs/so (!f’sec“)
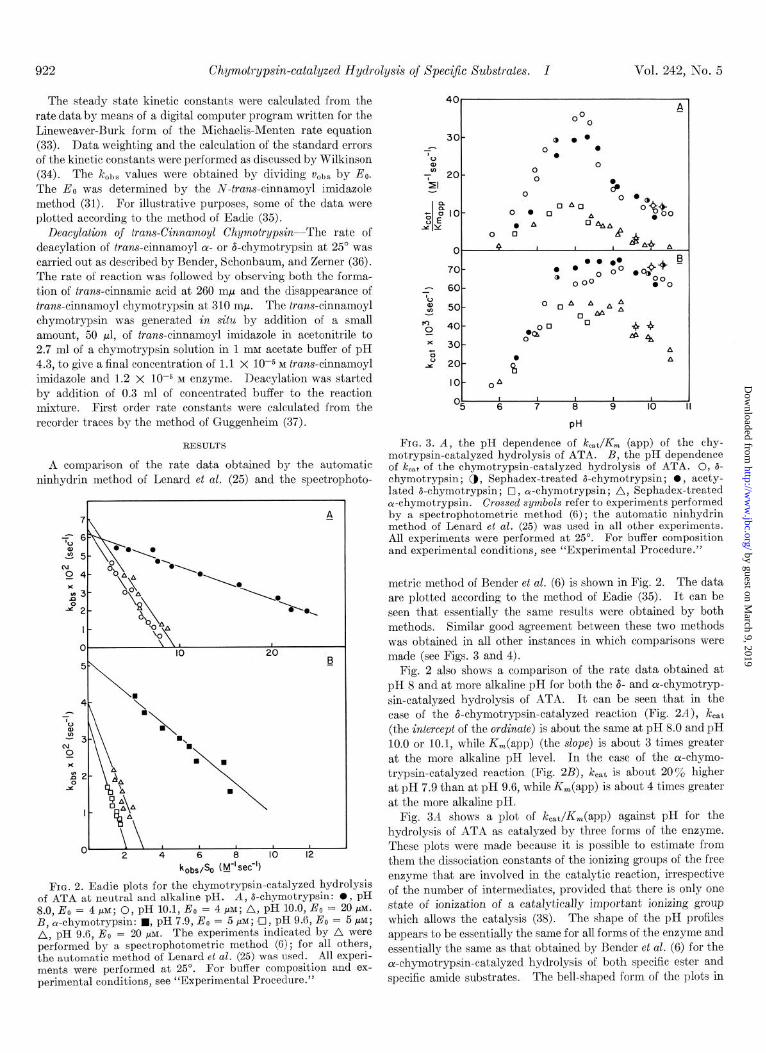
FIG. 2. Eadie plots for the chymotrypsin-catalyzed hydrolysis of ATA at neutral and alkaline pH. A, ii-chymotrypsin: 0, pH 8.0, Eo = 4 PM; 0, pH 10.1, Eo = 4 PM; A, pH 10.0, Eo = 20 PM. B, a-chymotrypsin: n , pH 7.9, Eo = 5 pM; .O, pH 9.6, Eo = 5 pM; A, pH 9.6, E. = 20 PM. The experiments indicated by A were performed by a spectrophotometric method (6); for all others, the automatic method of Lenard et al. (25) was used. All experi- ments were performed at 25”. For buffer composition and ex- perimental conditions, see “Experimental Procedure.”
= ‘0 5 50
v-l 0 40
- x 30 'r 2 20
I OnA A AA AA A
Cl 00 0
Oeg Ob &B
A A
IO OA
O5 I I I , I 6 7 8 9 IO
PH
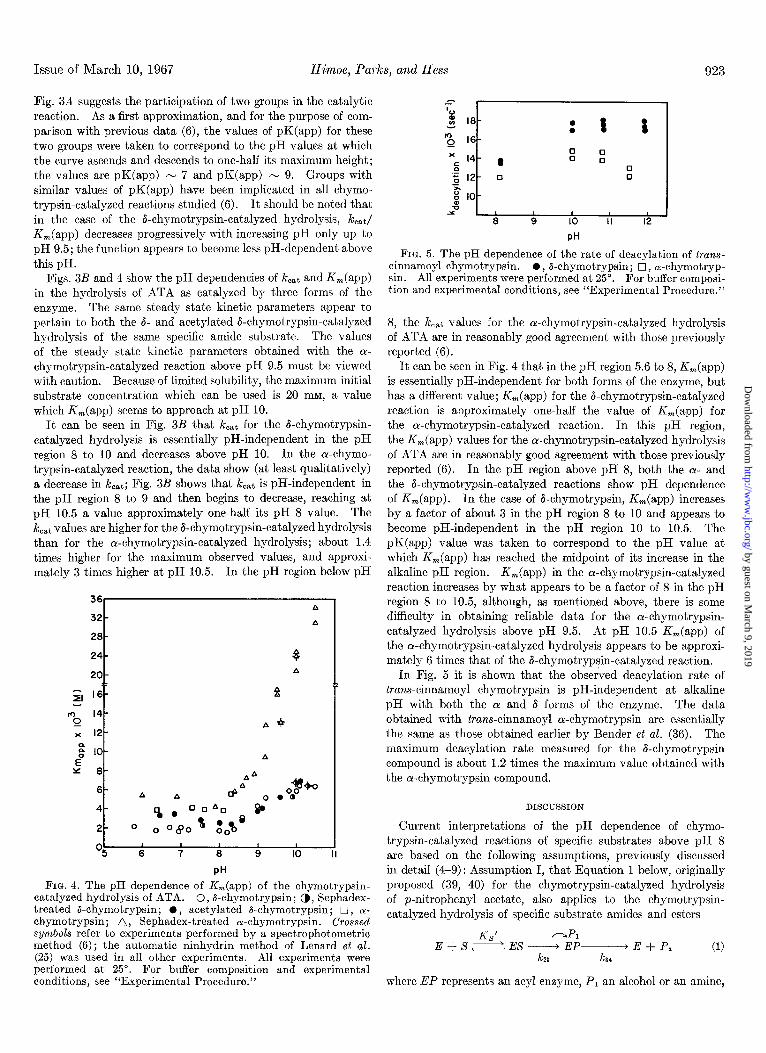
FIG. 3. A, the pH dependence of kJK, (app) of the chy- motrypsin-catalyzed hydrolysis of ATA. B, the pH dependence of k,,t of the chymotrypsin-catalyzed hydrolysis of ATA. 0, 6- chymotrypsin; 0, Sephadex-treated &chymotrypsin; 0, acety- lated &chymotrypsin; 17, cu-chymotrypsin; A, Sephadex-treated a-chymotrypsin. Crossed symbols refer to experiments performed by a spectrophotometric method (6); the automatic ninhydrin method of Lenard et al. (25) was used in all other experiments. All experiments were performed at 25”. For buffer composition and experimental conditions, see “Experimental Procedure.”
metric method of Bender et al. (6) is shown in Fig. 2. The data are plotted according to the method of Eadie (35). It can be seen that essentially the same results were obtained by both methods. Similar good agreement between these two methods was obtained in all other instances in which comparisons were made (see Figs. 3 and 4).
Fig. 2 also shows a comparison of the rate data obtained at pH 8 and at more alkaline pH for both the 6- and cr-chymotryp- sin-catalyzed hydrolysis of ATA. It can be seen that in the case of the &chymotrypsin-catalyzed reaction (Fig. 2A), kcat (the intercept of the ordinate) is about the same at pH 8.0 and pH 10.0 or 10.1, while K,(app) (the slope) is about 3 times greater at the more alkaline pH level. In the case of the a-chymo- trypsin-catalyzed reaction (Fig. 2B), keat is about 20% higher at pH 7.9 than at pH 9.6, while K,(app) is about 4 times greater at the more alkaline pH.
Fig. 38 shows a plot of k,,t/Ki,(app) against pH for the hydrolysis of ATA as catalyzed by three forms of the enzyme. These plots were made because it is possible to estimate from them the dissociation constants of the ionizing groups of the free enzyme that are involved in the catalytic reaction, irrespective of the number of intermediates, provided that there is only one state of ionization of a catalytically important ionizing group which allows the catalysis (38). The shape of the pH profiles appears to be essentially the same for all forms of the enzyme and essentially the same as that obtained by Bender et al. (6) for the oc-chymotrypsin-catalyzed hydrolysis of both specific ester and specific amide substrates. The bell-shaped form of the plots in
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Issue of March 10, 1967 Himoe, Parks, and Hess 923
Fig. 3A suggests the participation of two groups in the catalytic reaction. As a first approximation, and for the purpose of com- parison with previous data (6), the values of pK(app) for these two groups were taken to correspond to the pH values at which the curve ascends and descends to one-half its maximum height; the values are pK(app) - 7 and pK(app) N 9. Groups with similar values of pK(app) have been implicated in all chymo- trypsin-catalyzed reactions studied (6). It should be noted that in the case of the &chymotrypsin-catalyzed hydrolysis, lc& K,(app) decreases progressively with increasing pH only up to pH 9.5; the function appears to become less pH-dependent above this pH.
Figs. 3B and 4 show the pH dependencies of k,,$ and K,(app) in the hydrolysis of ATA as catalyzed by three forms of the enzyme. The same steady state kinetic parameters appear to pertain to both the 6- and acetylated &chymotrypsin-catalyzed hydrolysis of the same specific amide substrate. The values of the steady state kinetic parameters obtained with the (Y- chymotrypsin-catalyzed reaction above pH 9.5 must be viewed with caution. Because of limited solubility, the maximum initial substrate concentration which can be used is 20 mM, a value which K,(app) seems to approach at pH 10.
It can be seen in Fig. 3B that kcat for the &chymotrypsin- catalyzed hydrolysis is essentially pH-independent in the pH region 8 to 10 and decreases above pH 10. In the ar-chymo- trypsin-catalyzed reaction, the data show (at least qualitatively) a decrease in k,,t; Fig. 3B shows that kcat is pH-independent in the pH region 8 to 9 and then begins to decrease, reaching at pH 10.5 a value approximately one-half its pH 8 value. The kcat values are higher for the &chymotrypsin-catalyzed hydrolysis than for the oL-chymotrypsin-catalyzed hydrolysis; about 1.4 times higher for the maximum observed values, and approxi- mately 3 times higher at pH 10.5. In the pH region below pH
0 ;; 14- - AO x 12-
$ IO- E
A
= 8- AA
6- &A mo
A A 0 0 oo” 4-
%. 0 0 A0 8
2- 0 0 080 8 .g 00
O5 1 I
6 7 8 9 IO II
PH
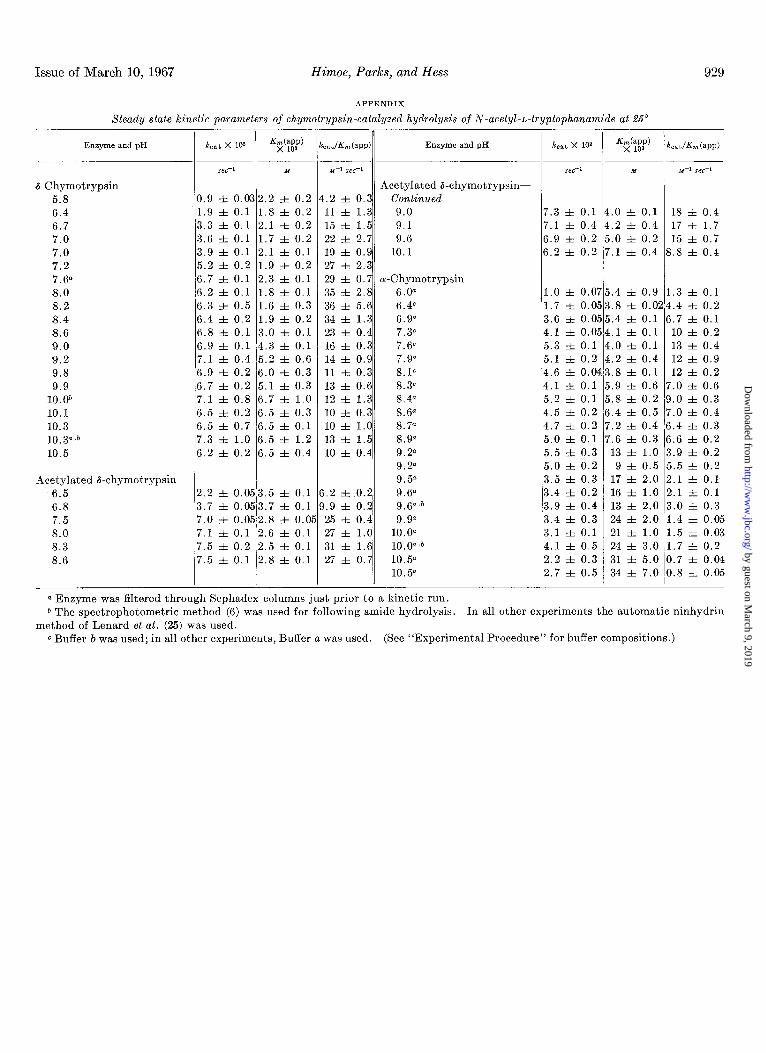
FIG. 4. The pH dependence of &(app) of the chymotrypsin- catalyzed hydrolysis of ATA. 0, khymotrypsin; a, Sephadex-
treated 8-chymotrypsin; l , acetylated d-chymotrypsin; 0, o1- chymotrypsin; a, Sephadex-treated cr.chymotrypsin. Crossed symbols refer to experiments performed by a spectrophotometric method (6); the automatic ninhydrin method of Lenard ef, al. (25) was used in all other experiments. All experiments were performed at 25”. For buffer composition and experimental conditions, see “Experimental Procedure.”
PH
FIG. 5. The pH dependence of the rate of deacylation of trans- cinnamoyl chymotrypsin. l , &chymotrypsin; q , a-chymotryp- sin. All experiments were performed at 25”. For buffer composi- tion and experimental conditions, see “Experimental Procedure.”
8, the k,,t values for the cr-chymotrypsin-catalyzed hydrolysis of ATA are in reasonably good agreement with those previously reported (6).
It can be seen in Fig. 4 that in the pH region 5.6 to 8, K,(app) is essentially pa-independent for both forms of the enzyme, but has a different value; K,(app) for the &chymotrypsin-catalyzed reaction is approximately one-half the value of K,(app) for the cr-chymotrypsin-catalyzed reaction. In this pH region, the K,(app) values for the cr-chymotrypsin-catalyzed hydrolysis of ATA are in reasonably good agreement with those previously reported (6). In the pH region above pH 8, both the (Y- and the &chymotrypsin-catalyzed reactions show pH dependence of K,(app). In the case of B-chymotrypsin, K,(app) increases by a factor of about 3 in the pH region 8 to 10 and appears to become pH-independent in the pH region 10 to 10.5. The pK(app) value was taken to correspond to the pH value at which K,(app) has reached the midpoint of its increase in the alkaline pH region. K,(app) in the cr-chymotrypsin-catalyzed reaction increases by what appears to be a factor of 8 in the pH region 8 to 10.5, although, as mentioned above, there is some difficulty in obtaining reliable data for the cr-chymotrypsin- catalyzed hydrolysis above pH 9.5. At pH 10.5 K,(app) of the a-chymotrypsin-catalyzed hydrolysis appears to be approxi- mately 6 times that of the &chymotrypsin-catalyzed reaction.
In Fig. 5 it is shown that the observed deacylation rate of trans-cinnamoyl chymotrypsin is pH-independent at alkaline pH with both the o( and 6 forms of the enzyme. The data obtained with trans-cinnamoyl Lu-chymotrypsin are essentially the same as those obtained earlier by Bender et al. (36). The maximum deacylation rate measured for the d-chymotrypsin compound is about 1.2 times the maximum value obtained with the oc-chymotrypsin compound.
DISCUSSION
Current interpretations of the pH dependence of chymo- trypsin-catalyzed reactions of specific substrates above pH 8 are based on t.he following assumptions, previously discussed in detail (4-9): Assumption I, that Equation 1 below, originally proposed (39, 40) for the chymotrypsin-catalyzed hydrolysis of p-nitrophenyl acetate, also applies to the chymotrypsin- catalyzed hydrolysis of specific substrate amides and esters
KS’ -PI E+SeES----+EP-E+Pz
k23 ka 0)
where EP represents an acyl enzyme, PI an alcohol or an amine,
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

924 Chymotrypsin-catalyzed Hydrolysis of Speci$c Substrates. I Vol. 242, No. 5
PZ a free acid, and S a specific chymotrypsin substrate, i.e. a derivative of an aromatic amino acid. Assumption II, that deacylation (&) is rate-limiting in the chymotrypsin-catalyzed hydrolysis of ester substrates except at very alkaline pH (see Assumption IV below), and that acylation (k23) is rate-limiting in the corresponding amide substrate hydrolysis. Therefore, in amide hydrolysis kcat is a measure of k23 and K,(app) is a measure of K’,. Assumption III, that K’, is pa-independent, that lcs4 is pa-independent above pH 8, and that kZ3 exhibits a bell-shaped pa-rate profile, the descending limb at alkaline pH implicating an ionizing group with pK(app) - 8.5. And Assumption IV, that as a consequence of Assumptions II and III, a pa-dependent change in the rate-limiting step occurs in the catalyzed hydrolysis of esters. Below pH 8, the deacylation step (with rate constant ks4) is rate-limiting; at some pH above pH 8, however, the bell-shaped pH profile of the acylation step (with rate constant k23) crosses below the pH-independent pH profile of the deacylation step, and the acylation step becomes rate-limiting.
These assumptions, and the experimental data on which they are based, must be re-examined in terms of the present work on ATA. The following considerations are pertinent.
1. The assumption that Kts, as measured by K,(app) in amide hydrolysis, is pa-independent, is inconsistent with the data (Fig. 4) showing that K,(app) of the a- and d-chymo- trypsin-catalyzed hydrolysis of ATA increases progressively with pH in the pH region 8 to 10.5. The pH dependence of K, (app) implicates (5,6) an ionizing group with pK(app) - 8.5, such as has been implicated in the known decrease (2-4) of the cata- lytic properties of chymotrypsin above pH 8. The experiments shown in Fig. 4 included the catalyzed hydrolysis of ATA by three forms of chymotrypsin, and involved about 30 determina- tions of the steady state kinetic parameters, with each determina- tion based on at least eight different kinetic runs.
The assumption of the pH independence of K’s and the pH dependence of kost above pH 8 was based on the following experi- ments: (a) investigations of the oc-chymotrypsin-catalyzed hydrolysis of ATA (4, 6), in which the reaction was measured by recording the difference spectrum at 306 rnp between ATA and the product N-acetyl-L-tryptophan. A difficulty with these investigations is that the published procedure requires that small absorbance differences be measured against a rela- tively high background, especially at high pH values where high enzyme concentrations are required, for the enzyme as well as substrate absorbs light at 306 mp. Another source of error in these previous experiments was revealed by the present work with the chymotrypsin-catalyzed hydrolysis of ATA. When a Gary model 14 spectrophotometer was used on the automatic slit width program (41), the results obtained were in reasonable agreement with published data (4, 6), but did not agree with results obtained by the automatic ninhydrin method of Lenard et al. (25). It was found that the reason for the disagreement between the two methods was the use of variable instrumental slit width in the Cary spectrophotometric measurements. With the automatic slit width program, the slit width is changed as initial substrate concentration is varied, and a result is a change in Atm the molar absorption difference coefficient, which is used as a measure of the hydrolysis of the amide (6). When the hydrolysis of ATA was followed in the Cary spectrophotom- eter with use of the same slit width regardless of initial substrate
concentration, as proper use of the instrument requires (41), excellent agreement between the spectrophotometric method and the automatic ninhydrin method was obtained (Figs. 2 to 4). (b) Investigations of the cr-chymotrypsin-catalyzed hy- drolysis of N-acetyl-n-phenylalaninamide and N-acetyl-nn- phenylalaninamide (6), in which the ammonia liberated was measured with use of Nessler’s reagent. A difficulty with these experiments is that the data at pH 8 and above are few and show considerable scatter; the K,(app) values vary by a factor of over 3 in the pH region 7.6 to 9.95, but in an irregular fashion, making any conclusion about the pH dependence of K,(app) proble- matic. Although the solid line drawn through the points (Fig. 5 in Reference 6) indicates that koat decreases above pH 8, the actual data look rather similar to those obtained in the cu-chymo- trypsin-catalyzed hydrolysis of ATA (Fig. 3B). Direct deter- mination (22) of the over-all enzyme-substrate dissociation constants of o(- or b-chymotrypsin and N-acetyl-n-phenylalanin- amide, with spectral changes of the enzyme near 290 rnp used as a measure of the enzyme-substrate complex, led to the conclu- sion that in this reaction also, pH dependence above pH 8 is due to the pH-dependent formation of an enzyme-substrate complex and not due to the effect of pH on the bond-breaking step per se. (c) Investigations of the acylation of a-chymo- trypsin byp-nitrophenyl acetate (6,42). Conclusions drawn from these experiments, namely that this reaction follows Michaelis- Menten kinetics and that I&, t’he acylation rate constant, and K’s can be evaluated separately, are questionable in view of a recent study of this reaction by Faller and Sturtevant (43). In the earlier work of Bender et al. (6, 42), the evaluation of data suffers from several difficulties. In those experiments in which Eo >> So, the trouble is that chymotrypsin concentrations suffi- ciently greater than the reported K’, values of approximately 1 x 1OP M cannot be obtained. In those experiments in which So >> EO, evaluation of the constants k23 and K’, from the equa- tion
k ohs = kzsSo(So + K’s)-’ + k34 (2)
suffers from the fact that the authors ignore the kZ4 term. Faller and Sturtevant, in their recent studies (43), found that the kZ4 term in Equation 2 is not insignificant compared to kobs at pH 7.0, and must be considered. In a thorough investigation of the acylation reaction at pH 7.0 in isopropanol-water and di- oxane-water mixtures containing various concentrations of organic solvent and buffer, the reaction was shown to follow second order kinetics under all conditions. When comparable, the second order rate constants are in excellent agreement with those of Bender et al., i.e. 2200 M-l se@ at pH 7.0 and 25” in 1% (v/v) isopropanol-water (43), and 2490 M-I see-1 at pH 7.1 and 25” in 1.6% (v/v) acetonitrile-water (6). This agree- ment between the data is not surprising, since inclusion of the lcs4 term has only a slight effect on the second order term, &a/K’, in Equation 2. The inclusion of the ks4 term does, however, have a large effect in the determination of whether the data follow second order or Michaelis-Menten kinetics. This is illustrated by recalculations (43) of earlier data of Gutfreund and Sturtevant (40) for the acylation of chymotrypsin by p- nitrophenyl acetate at pH 6.5 and pH 7.8. The authors (43) showed that inclusion of the ks4 term in the calculations gave second order kinetics rather than Michaelis-Menten kinetics as first reported (40). Inclusion of the ks4 term in Equation 2
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of March 10, 1967 Himoe, Parks, and Hess
becomes even more important for evaluation of data obtained above pH 8, since k&s decreases above pH 8 (6) while kz4 is presumably pH-independent.
The data of Bender et al. (6) can therefore be interpreted to mean that the decrease in k&s for the acylation of chymotrypsin by p-nitrophenyl acetate above pH 8 is comparable to a similar decrease observed in other chymotrypsin-catalyzed reactions; in particular the reaction of chymotrypsin with DFP, where the second order term k~s/K’, was observed to decrease above pH 8. The conclusion of the authors that the acylation rate constant (k23) itself is pH-dependent is not warranted in view of the observations of Failer and Sturtevant.
2. Also called into question by the present ATA study is the suggestion (5, 6) that the observed pH dependence of K,(app) in the chymotrypsin-catalyzed hydrolysis of N-acetyl-n-trypto- phan ethyl ester is due to the pH dependence of k23. This inter- pretation is based on Assumption III above, that K’s is pH- independent and lc23 is pH-dependent. However, there exists no evidence which indicates that K’s for the ester is pH-inde- pendent. On the contrary, pH dependence of Kls is suggested by the pH dependence of K,(app) in the chymotrypsin-catalyzed hydrolysis of the corresponding amide substrate (Fig. 4).
3. The observation (5, 6) that the decrease in kCat of the ol-chymotrypsin-catalyzed hydrolysis of N-acetyl-L-tryptophan ethyl ester occurs above pH 10 has been considered as evidence for Assumption IV that the rate-limiting step of the reaction changes from ks4 to k23 at high pH. The experiments on the a-chymotrypsin-catalyzed hydrolysis of ATA, the corresponding amide, certainly do not support this assumption. In amide hydrolysis, k23 is supposedly rate-limiting in the entire pH range, yet the experimental results (Fig. 3) show only a moderate pH dependence of kcat at pH 10. The assumption of pH inde- pendence of ka4 in all chymotrypsin-catalyzed reactions at alkaline pH values is based on the observation that the rate of deacylation of the model compound N-trans-cinnamoyl cu-chymo- trypsin is pH-independent in the pH region 8 to 12 (36). This one example hardly precludes the pH dependence at high pH values of the deacylation of N-acetyl-L-tryptophyl chymo- trypsin (EP in Equation I), assuming that such a compound exists.
4. The nonidentity of the pH dependence of kcat and k,,J K,(app) in the chymotrypsin-cat.alyzed hydrolysis of ATA (Fig. 3) contradicts Assumptions II and III which require that in amide hydrolysis. kcat and k,JK,(app) have the same pH dependence. The data presented indicate that the pH de- pendence of the catalytic reaction above pH 8 is complex (38).
In view of previous experiments (24) which indicate that K,(app) in the chymotrypsin-catalyzed hydrolysis of specific amide substrates is a direct measure of the over-all enzyme- substrate dissociation constant, it can be deduced that the alkaline pH dependence of K,(app) in the ATA reaction (Fig. 4) is mainly due to the effect of pH on the formation of chymo- trypsin complexes. Since this pH dependence of K,(app) adequately accounts for the decrease in catalytic properties of the enzyme at alkaline pH, it can be concluded that the ATA data are consistent with the hypothesis that the origin of the alkaline pH dependence of reaction is the same in chymotrypsin- catalyzed hydrolysis of specific substrates as it is in the DFP- chymotrypsin reaction (see introductory section). This possi- bility of a relationship between pH-dependent conformational
C-NH
;
(Chymotrypsinogen)
FIG. 6. Scheme for the formation of chymotrypsin-substrate complexes, showing ionization equilibria and equilibria between two main conformations of the enzyme, A and I, in the free state and in complexes with substrate. The equilibrium constants kL and k‘L apply to the pH 7 form of the enzyme (conformation A) and kH and k’n apply to the pH 10.5 form of the enzyme (conforma- tion I). Although not explicitly shown, free substrate concen- tration %So is involved in the equilibria shown.
changes of the enzyme above pH 8 and the pH dependence of the chymotrypsin-catalyzed hydrolysis of specific substrates at alkaline pH can be considered, for purposes for discussion, in terms of the scheme shown in Fig. 6. The formation of enzyme-substrate complexes involving two conformations of chymotrypsin, A and I, is presented graphically in this figure. The main conformation is considered to be A at neutral pH, and I at alkaline pH. The ionizations of AH and IH, with ionization constants KA and KI, respectively, are considered to be due to one and the same ionizing group in both conforma- tions.
The equations pertaining to the scheme in Fig. 6 above pH 8 appear in Table I. They are based on the following assump- tions: (a) that there exists a pH-dependent equilibrium between two major conformations of the enzyme in the alkaline pH region, This assumption is based on experiments (44-47) which indicate that the conformations of chymotrypsinogen and chymotrypsin are different at neutral pH; on several experiments (17, 21, 48) which suggest that the conformations of the zymogen and the enzyme are similar above pH 10; and on data (44) which indicate that the specific rotation values for chymotrypsinogen and chymotrypsin are different at neutral pH and the same above pH 10. (b) That the pa-dependent equilibrium between two major conformations of the enzyme is characterized by a single ionization constant. This assumption is based on experiments with acetylated chymotrypsinogen and acetylated d-chymo- trypsin (17, 21) which show that when the specific rotation of
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

926 Chymotrypsin-catalyzed Hydrolysis of XpeciJic Substrates. I Vol. 242, No. 5
TABLE I Dejhitions and equations pertaining to Fig. 6
I. Definitions pertaining to Fig. 6
kL = FH < 1
K =(A)(H) A
K = U)(H)
(AH) I----
ul3
The assumption of a pH-dependent equilibrium between two ma- jor conformations of the enzyme, with conformation Ipredominat- ing at high pH and conformation A predominating at neutral pH, requires k~ < 1 and k,~ > 1. The constants k’.~,, k’~, K’A, and K’I are defined in the same way as kL, kH, KA, and KI, but in- volve the enzyme-substrate complexes SAH, SA, NH, and 81, rather than the corresponding enzyme forms.
K (SO)(AH) (So) (A)
SAH = (SAH) KSA - --
GSA)
K (So) UH) c30~ (I)
SIH = (SIH) Ksr = __ @I)
kAn, kA, krE, and kl are defined as the first order rate constants for the conversion to free enzyme and product of SAH, SA, XIH, and SI, respectively. II. Equations pertaining to free enzyme in absence of substrate
Eo=EH+E=AH+TI+A+I
KIkr,=KAkx
K*s = (I + A)(H) = K
U-H + AH)
0 + kx) = _ KAkx
A(l+
III. Equations pertaining to Fig. 6
Eo=EH+E+SEH+SE=AH+IH+A
+I+SAH+SZH+SA+SI
KSAHKSA K K’A k’H = K’I k’L = KA kx __ __
SAEKSIH
&A Ksr =KrkL----
KSIH KSI
K* (SI + SA)(H) (1 + k’d
ES = tsm + XAHj = K'A ~ = m K’A k’z (1 + k’d
where
SAH=H+ SA = K’A +
SIH = H k’L c$ SI = K’A k’x 4
Eo So ‘= S&H+ K*zs) f KSAHCH + K*E)
2)obs = kaH[8AHl + km[SIHl + ka[SAl + kr[SIl
= ,#@A& + km H k’L + kAKl.4 + krK’ak’H1
= +kl[H + K’A kzl
where
kt = kAH $ klH kII, and kp = &A + kI k’d
(km + km k’d
(H + K*s) K&w) = KSAH cH + K*zsj
k cat = kl (H + K’A kz)
(H + K*E&
h (H +K’A kz)
k,,tlK&pp) = - KSAH (H + K*E)
(1) (2)
(3)
(4)
(5)
(6)
(7)
(53)
(9)
(10)
(11)
the enzyme at neutral pH is considered characteristic of one conformation of the enzyme (conformation A in Fig. 6), and at pH 10.5 characteristic of another conformation (conformation I in Fig. 6), the pH-dependent change in specific rotation follows the ionization of a single group with pK(app) - 8.3. (c) That the pH-dependent cyclic equilibrium between two major con- formations of the enzyme accounts for the pH dependence of the chymotrypsin-catalyzed hydrolysis above pH 8. (d) That an ionizing group with pK(app) - 7, a group implicated in all chymotrypsin-catalyzed reactions (for a review of this subject, see Reference 9), is completely ionized above pH 8 and need not be considered in the alkaline pH region. (e) That the pertinent enzyme-substrate complexes (81, XIH, XA, and &lH in Fig. 6) are in equilibrium with enzyme and substrate under the condi- tions used in the experiment. This assumption is based on stopped flow experiments with chymotrypsin and the substrates ATA (24) and N-acetyl-L-phenylalaninamide (22). The only intermediates which could be observed in these experiments were formed in less than 3 msec, while kobs for the catalytic reaction was less than 0.06 see-r. Furthermore, the concentra- tion dependence of the detected enzyme-substrate complexes yielded enzyme-substrate dissociation constants which are in excellent agreement with K,(app), a constant obtained from steady state kinetic experiments.
Some properties of the scheme in Fig. 6 which are pertinent to the chymotrypsin-catalyzed hydrolysis of ATA are as follows.
Equation 9 (Table I) gives the relationship between K,(app) and pH above pH 8, regardless of the number of ES complexes that are converted to free enzyme and product. It shows that K,(app) will bepH-dependent whenKeE # K*xs. Theobserved pH dependence of K,(app) in the catalytic hydrolysis of ATA above pH 8 requires that pK*,rs > PK*~. This leads to the prediction, which can be tested, that the binding of specific amide substrates by chymotrypsin is accompanied by proton uptake by the enzyme at alkaline pH. It also leads to the prediction that the conversion of A conformation to I conforma- tion must occur at higher pH in presence of substrate than in absence of substrate, since pK*zs controls the pH-dependent cyclic equilibrium of the ES complexes; this can be tested by measuring the change in specific rotation of the enzyme when enzyme and substrate are mixed at alkaline pH. A proton uptake by the enzyme and a change in the pH dependence of the specific rotation of the enzyme have already been observed in the DFP-chymotrypsin reaction above pH 7 (17, 20, 21).
It should be noted (see Equations 3 and 6, Table I) that K*, and K*Es are complex constants related to the ionization constants of conformations A and I in the free enzyme and the enzyme-substrate complexes, respectively. The values of K*z and KEEP can be estimated from data on the pH dependencies of steady state kinetic parameters, provided that the limiting high pH values of the pertinent parameters can be measured.
K*B can be calculated from Equation 11 (Table I), which gives the relationship between k&K&app) and pH above pH 8. The midpoint of the pH-dependent decrease of Ic,,J K,(app), which is defined as 3 [(k,,tlK,(app)) maximum va~ue -l- &dL(app)) limiting high pHvaluelisgivenby[kl(K*~ -I- K'~kz)l
[2K*EKs,&1. Substitution of this expression for k&Kn(app) in Equation 11 gives [HI+ = K*z, or pH = pK*B. The experi- ments represented in Fig. 3A indicate that in the 6- or acetylated &chymotrypsin-catalyzed hydrolysis of ATA, k,,JK,(app) reaches its limiting high pH value at about pH 10, and that the
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of March 10, 1967 Himoe, Parks, and Hess 927
midpoint of the decrease in k&Ki-,(app) is at about pH 8.5; thus, PK*~ is about 8.5. An ionizing group of about this pK(app) value has also been implicated in the control of the pH dependence of the specific rotation of the same enzyme (21). The specific rotation measurements indicate that this ionizing group controls the equilibrium between two major conformations of the enzyme; the measurements reported in Fig. 34 indicate that this ionizing group is important in the catalytic reaction and is responsible for the decrease in catalytic properties of the enzyme above pH 8.
K*E8 can be estimated in a manner similar to the evaluation of K*E from data on the pH dependence of either K,(app) or k cat.
Equation 10 (Table I), which gives the relationship between kcat and pH, shows that the limiting cases of the pH dependence of koat yield a bell-shaped pH profile when K’,k2 < K*ES, and a pH-independent profile when K’,kz = KEEP. Therefore, the proposed relationship between the pH-dependent cyclic equilib- rium of two major conformations of the enzyme and the pH dependence of the steady state kinetic parameters accommodates the pH dependence of kcat in both c+ and &chymotrypsin- catalyzed hydrolysis of ATA above pH 8 (Fig. 3B).
In previous treatments (6, 38) of the pH dependence of en- zyme-catalyzed reactions, the pH dependence of k&K,(app) has been considered to be a property of the free enzyme only, and to be the same for all neutral substrates. For the example treated here, the pH dependence of k&K,(app) depends not only on K*,, but also on K’,kg and as can be seen from Equation 6, K’, reflects not only the properties of the free enzyme, but also the various enzyme-substrate dissociation constants and the properties of the cyclic equilibrium of these complexes. Differences in substrate are therefore expected to result in differences in K’, and thereby affect the pH dependence of LdLdapp).
It is to be noted from the scheme in Fig. 6 that an equilibrium between two main conformations of the enzyme exists at all pH values. This suggests the possibility that the conformational changes of the enzyme and the spectral changes of the enzyme at 290 rnp, which have been deduced to accompany the chymo- trypsin-DFP reaction at all pH values, are due to the perturba- tion of the equilibrium between these two main conformations of the enzyme.
It should be noticed that the scheme in Fig. 6 is an extension of an earlier model, (20, 21) which was based on studies of in- dividual steps of the reaction of chymotrypsin with DFP (10-21). In this earlier model, the formation of a reversible chymotrypsin complex was considered to involve only the A conformations of the enzyme. In the scheme in Fig. 6, the substrate is con- sidered to bind to both the I and A conformations of the enzyme. This assumption is justified by the experiments depicted in Fig. 4, which indicate that K,(app) for the &chymotrypsin-catalyzed hydrolysis of ATA becomes pH-independent above pH 10, and by reports of Vaslow and Doherty (49) that chymotrypsinogen, which is considered to resemble the conformation of the enzyme at high pH (conformation 1 in Fig. 6) binds virtual substrates. While the earlier model (20, 21) predicted the pH dependence above pH 8 of the formation of chymotrypsin complexes, the pH independence of the bond-breaking step, the hydrogen ion uptake by the enzyme, and the change in specific rotation of the enzyme upon binding by substrate, the present model also takes
into account the differences in the catalytic properties of various forms of chymotrypsin in the hydrolysis of ATA.
It is important to emphasize that although both chemical and physical studies are consistent with the existence of a pH- dependent cyclic equilibrium between two major conformations of the enzyme, the suggested effect of such an equilibrium on chymotrypsin-catalyzed reactions of specific substrates is a hypothesis. Analysis of the pH dependence of chymotrypsin- catalyzed hydrolysis of specific amides according to this hy- pothesis is valuable, however, because it suggests measurable changes of the enzyme which can be used to investigate individual steps in the catalytic reaction. According to the scheme of Fig. 6, interaction of the enzyme with specific substrates may be accompanied by hydrogen ion uptake, and by changes in optical rotatory and spectral properties of the enzyme. The determina- tion of the steady state kinetic parameters is also a part of the study of individual reaction steps, since these parameters give the over-all equilibrium and rate constants with which the individual steps of the reactions must be reconciled.
An additional new and important aspect of chymotrypsin- catalyzed hydrolysis indicated by the data presented in this paper is the comparison of catalytic activities of the three forms of the enzyme that were studied. There is experimental evidence that there actually were three different enzyme forms. End group determinations of several batches of three times crystal- lized Worthington or-chymotrypsin preparations have led to the conclusion that the preparation is essentially the LY form (21). Although the &chymotrypsin preparation was not well charac- terized, the acetylated &chymotrypsin preparation was; end group analysis (21) was used in characterizing this enzyme also. The steady state kinetic parameters of the 6- and acetylated &chymotrypsin-catalyzed hydrolysis of ATA appear to be identical (7; Figs. 3 and 4). However, the data on the hydrolysis of ATA presented in Figs. 2 to 4 indicate that d-chymotrypsin is a more efficient catalyst than a-chymotrypsin as far as the observed maximum rate of hydrolysis and the binding of the substrate are concerned. Similarly, the data in Fig. 5 indicate that trans.cinnamoyl d-chymotrypsin is deacylated more effi- ciently than the corresponding ar-chymotrypsin compound. The only previous indications of differences in catalytic prop- erties between the two forms of the enzyme have been the qualita- tive observations (27, 50, 51) that the specific activity of 6- chymotrypsin is greater than that of the a: form near pH 8.2
REFERENCES
1. NIEMANN, C.,Science, 143, 1287 (1964). 2. NORTHROP, J. H., KUNITZ, M., AND HERRIOT, R. M., Crystal-
2 Note Added in Proof-In a short paper (52), which appeared after this manuscript had been submitted for publication, Bender et al. have retracted their previous findings (4, 6) on the variation with pH of kcrat and of Km (app) for the chymotrypsin-catalyzed hydrolysis of ATA at alkaline pH values and have also corrected their previous data (5, 6) on the pH dependence of kCRt for the hydrolysis of N-acetyl-L-tryptophan ethyl ester. The results presented by Bender, Gibian, and Whelan for the hydrolysis of ATA are in qualitative agreement with those presented in this paper and those published previously (26,53). Bender et al. have also proposed a model for the pH dependence of chymotrypsin- catalyzed hydrolyses at alkaline pH values which becomes iden- tical with a model proposed earlier (21) when the notations used by Bender et al. (E*H+, E*, EH+, E, K*H, Klp, K~H, and KT) are converted to the earlier notation (IH, I, AH, A, KI, KA, KL, and KH, respectively).
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

928 Chymotrypsin-catalyzed Hydrolysis of Specific Substrates. I Vol. 242, No. 5
3.
4.
5.
G.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22. 23.
24.
25.
line enzymes, Ed. 2, Columbia University Press, New York, 26. HIMOE, A., PARKS, P. C., AND HESS, G. P., Federation Proc., 1948. 24, 473 (1965).
HUANG, H.T., AND NIEMANN, C.,J. Am. Chem. Sot., 73, 1541 27. JACOBSEN, C. F., Compt. Rend. Trav. Lab. Carlsberg, Ser. (1951). Chim., 25, 325 (1947).
BENDER, M. L., CLEMENT, G. E., K~ZDY, F. J., AND ZERNER, 28. YAPEL, A., HAN, M., LUMRY, R., ROSENBERG, A., AND SHIAO, 1). F., J. Am. Chem. Sot., 88, 2573 (1966).
29. DIXON, G. H., AND NEURATH, H., J. Biol. Chem., 225, 1049 (1957).
B., J. Am. Chem. Sot., 86, 358.(1965). BENDER. M. L.. AND CLEMENT. G. E.. Biochem. Biowhus. Res.
Commnn., 12; 339 (1963). ’ ’ 1 ”
BENDER, l% L., CLEMENT, G. E., KBZDY, F. J., AND HECK, 30. WILCOX, P. E., KRAUT, J., WADE, R. D., AND NEURATH, H., H.. J. Am. Chem. Sot.. 86, 3680 (1964). Biochim. Biowhus. Acta. 24. 72 (1957).
ZER~ER, B., AND BENDER, G. L., J: Am: Chem. Sot., 86, 3669
BENDER, M. L., AND K~ZDY, F. J., J. Am. Chem. Sot., 86, 3704 (1964).
(1964). BENDER, M. L., AND K~ZDY, F. J., Ann. Rev. Biochem., 34,
49 (1965). WOOTTON, J. F., AND HESS, G. P., J. Am. Chem. Sot., 84, 440
(1962). HAVSTEEN, B. H., AND HESS, G. P., J. Am. Chem. Sot., 84,448
(19G2). HAVSTEEN, B. H., AND HESS, G. P., J. Am. Chem. Sot., 84,491
(1962). LABOUESSE, B., HAVSTEEN, B. H., AND HESS, G. P., Proc.
Natl. Acad. Sci. U. S., 48, 2137 (1962). OPPENHEIMER, H. L., MERCOUROFF, J., AND HESS, G. P.,
Biochim. Biophys. Acta, 71, 78 (1963). HAVSTEEN, B. H., AND HESS, G. P., J. Am. Chem. Sot., 85,791
(1963). HAVSTEEN, B. H., LABOUESSE, B., AND HESS, G. P., J. Am.
Chem. Sot., 86, 796 (1963). LABOUESSE, B., OPPENHEIMER, H., AND HESS, G. P., in T. W.
GOODWIN, J. I. HARRIS, AND B. S. HARTLEY (Editors), Structure and activity of enzymes, Academic Press, Inc., New York, 19G4, p. 134.
MOON, A-Y., MERCOUROFF, J., AND HESS, G. P., J. Biol. Chem., 240, 717 (1965).
HAVSTEEN, B. H., AND HESS, G. P., Biochem. Biophys. Res. Commun., 14, 313 (1964).
MOON, A-Y., STURTEVANT, J. M., AND HESS, G. P., J. Biol. Chem.. 240. 4204 (1965).
OPPENH&&, H. i., ~ABOUESSE, B., AND HESS, G. P., J.
Biol. Chem.. 241, 2720 (1966). HIMOE, A., AND HI&, G. k., Federation Proc., 26,650 (1966). MOUNTER, L. A., ALEXANDER, H. C., TUCK, K. D., AND DIEN,
L. T. H., J. Biol. Chem., 226, 867 (1959). BRANDT, K. G., AND HESS, G. P., Biochem. Biophys. Res.
Commun., 22, 447 (1966). LENARD, J., JOHNSON, S. L., HYMAN, R. W., AND HESS, G. P.,
Anal. Biochem., 11, 30 (1965).
31. SCHONBAUM, G: 6., ZERN~R, B., AND BENDER, M. L., J. Biol.
32. MOORE, S., AND STEIN, W. H., J. BioZ. Chem., 211, 908 (1954). 33. LINEWEAVER, H., AND BURK, D., J. Am. Chem. SOC., 66, 658
Chem., 236, 2930 (1961).
(1934). 34. WILKINSON, G. N., Biochem. J., 80, 324 (1961). 35. EADIE, G. S., J. Biol. Chem., 146, 85 (1942). 36. BENDER, M. L., SCHONBAUM, G. R., AND ZERNER, B., J. Am.
Chem. Sot., 84, 2562 (1962). 37. GUGGENHEIM, E. A., Phil. Mag., Ser. 7, 2, 538 (1926). 38. PELLER, L., AND ALBERTY, R. A., J. Am. Chem. Sot., 81, 5907
(1959). 39. HARTLEY, B. S., AND KILBY, B. A., Biochem. J., 66, 288 (1954). 40. GUTFREUND, H., AND STURTEVANT, J. M., Biochem. J., 63,
658 (1956). 41. Optimum Spectrophotometer Parameters, Application Report
AR-14-2, Cary Instruments, Applied Physics Corporation, Pasadena, California, September, 1964.
42. K~ZDY, F. J., AND BENDER, M. L., Biochemistry, 1,1097 (1962). 43. FALLER, L., AND STURTEVANT, J. M., J. Biol. Chem., 241, 4825
(1966). 44. NEURATH, H., RUPLEY, J. A., AND DREYER, W. J., Arch. Bio-
them. Biophys., 66, 243 (195G). 45. IMAHORI, K., YOSHIDA, A., AND HASHIZUME, H., Biochim.
Biophys. Acta, 45, 380 (1960). 4G. FASMAN, G. D., AND FOSTER, R. J., Federation Proc., 24, 472
(1965). 47. RAVAL, D. N., AND SCHELLMAN, J. A., Biochim. Biophys. Acta,
107, 463 (1965). 48. BILTONEN, R., LUMRY, R., MADISON, V., AND PARKER, H.,
Proc. iVat1. Acad. Sci. U. S., 64, 1412 (1965). 49. VASLOW, F., AND DOHERTY, D. G., J. Am. Chem. Sot., 76, 928
(1953). 50. BETTELHEIM, F. R., ANU NEURATH. H.. J. Biol. Chem.. 212.
241 (1955): ’ , ,
51. ROVERY, M., POILROUX, M., YOSHIDA, A., AND DESNUELLE, P., Biochim. Biophys. Acta, 23, GO8 (1957).
52. BENDER, M. L., GIBIAN, M. J., AND WHELAN, D. J., Proc. Natl. Acad. Sci. U.S.. 66.833 (19%).
53. HIMOE, A., AND HESS, G. ‘P., lkochem. Biophys. Res. Commun., 23, 234 (19G6).
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Issue of March 10, 1967 Himoe, Parks, and Hess
APPENDIX
Steady state kinetic parameters of chymotrypsin-catalyzed hydrolysis of N-acetyl-L-tryptophanamide at 25’
929
Enzyme and pH k,,t X 102 &&Ps”
XC’ M
&Chymotrypsin 5.8 0.9 f 0.032.2 f 0.2 6.4 1.9 f 0.1 1.8 + 0.2 6.7 3.3 f 0.1 2.1 * 0.2 7.0 3.6 f 0.1 1.7 A 0.2 7.0 3.9 f 0.1 2.1 f 0.1 7.2 5.2 f 0.2 1.9 f 0.2 7.6” 6.7 f 0.1 2.3 rk 0.1 8.0 6.2 f 0.1 1.8 + 0.1 8.2 6.3 f 0.5 1.6 f 0.3 8.4 6.4 f 0.2 1.9 f 0.2 8.6 6.8 f 0.1 3.0 f 0.1 9.0 6.9 f 0.1 4.3
0.4 15.2 f 0.1
9.2 7.1 f f 0.6 9.8 6.9 f 0.2 ‘6.0 f 0.3 9.9 6.7 f 0.2 5.1 f 0.3
10.0” 7.1 f 0.8 6.7 f 1.0 10.1 6.5 f 0.2 6.5 zk 0.3 10.3 6.5 f 0.7 6.5 31 0.1 10.3”JJ 7.3 f 1.0 6.5 f 1.2 10.5 6.2 f 0.2 6.5 f 0.4
Acetylated 8-chymotrypsin 6.5 2.2 f 0.05,3.5 f 0.1 6.8 3.7 f 0.0513.7 f 0.1 7.5 7.0 zk 0.05,2.8 xk 0.01 8.0 7.1 rk 0.1 2.6 + 0.1 8.3 7.5 f 0.2 2.5 f 0.1 8.6 7.5 f 0.1 2.8 f 0.1
M
.o f 0.1
.2 f 0.4
.o f 0.2
.I f 0.4
:at/&(ap~) Enzyme and pH k,,t X 102 oat/Knn(app)
M-1 s&r’ see-’ M-1 seco
Acetylated &chymotrypsin- .2 f 0.3 Continued 11 f 1.3 9.0 7.3 f 0.1 18 + 0.4 15 zt 1.5 9.1 7.1 f 0.4 17 f 1.7 22 f 2.7 9.6 6.9 f 0.2 15 f 0.7 19 f 0.9 10.1 6.2 f 0.2 .8 f 0.4 27 f 2.3 29 f 0.7 ol-Chymotrypsin 35 f 2.8 6.0Q 1.0 f o.oi .3 + 0.1 36 f 5.6 6.4” 1.7 f 0.0: .4 f 0.2 34 f 1.3 6.ga 3.6 f 0.0: .7 f 0.1 23 f 0.4 7.3c 4.1 f O.OE 10 f 0.2 16 f 0.3 7.6” 5.3 f 0.1 13 f 0.4 14 f 0.9 7.9” 5.1 f 0.2 12 f 0.9 11 f 0.3 8.1c 4.6 f 0.04 12 f 0.2 13 f 0.6 8.3” 4.1 f 0.1 .O f 0.6 12 f 1.3 8.4” 5.2 f 0.1 .o f 0.3 10 f 0.3 8.6” 4.5 f 0.2 .o f 0.4 10 f 1.0 8.7,= 4.7 f 0.2 .4 f 0.3 13 f 1.5 8.9” 5.0 f 0.1 .6 f 0.2 10 f 0.4 9.20 5.5 f 0.3 .9 f 0.2
9.2a 5.0 f 0.2 .5 f 0.2 9.5” 3.5 f 0.3 .l Z!Z 0.1
.2 f 0.2 9.6” 3.4 f 0.2 .l f 0.1
.9 f 0.2 9.6”,* 3.9 f 0.4 .o f 0.3 25 rt 0.4 9.9” 3.4 f 0.3 .4 f 0.05 27 f 1.0 10.04 3.1 f 0.1 .5 f 0.03 31 f 1.6 10.Oa-* 4.1 f 0.5 .7 f 0.2 27 f 0.7 10.5” 2.2 f 0.3 .7 f 0.04
10.50 2.7 f 0.5 .8 f 0.05
a Enzyme was filtered through Sephadex columns just prior to a kinetic run. * The spectrophotometric method (6) was used for following amide hydrolysis. In all other experiments the automatic ninhydrin
method of Lenard et al. (25) was used. c Buffer b was used; in all other experiments, Buffer a was used. (See “Experimental Procedure” for buffer compositions.)
.4 zt 0.9
.8 f 0.0:
.4 f 0.1
.l f 0.1
.o f 0.1
.2 f 0.4
.8 f 0.1
.9 f 0.6
.8 f 0.2
.4 f 0.5
.2 f 0.4
.6 f 0.3 13 f 1.0
9 f 0.5 17 f 2.0 16 f 1.0 13 f 2.0 24 f 2.0 21 f 1.0 24 f 3.0 31 f 5.0 34 f 7.0
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Albert Himoe, Paul C. Parks and George P. HessENZYME AT ALKALINE pH
N-ACETYL-l-TRYPTOPHANAMIDE BY THREE FORMS OF THEI. THE pH DEPENDENCE OF THE CATALYTIC HYDROLYSIS OF
Investigations of the Chymotrypsin-catalyzed Hydrolysis of Specific Substrates:
1967, 242:919-929.J. Biol. Chem.
http://www.jbc.org/content/242/5/919Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/242/5/919.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on March 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from