interaction of central and superficial peripheral thermosensors in control of thermoregulatory...
TRANSCRIPT

Physiology & Behavior, Vol. 23, pp. 503-512. Pergamon Press and Brain Research Publ., 1979. Printed in the U.S.A.
Interaction of Central and Superficial Peripheral Thermosensors in Control of Thermoregulatory
Behaviors of the Rat
W A R R E N W. R O B E R T S A N D A L A N B. F R O L
Depar tment o f Psychology, University o f Minnesota, Minneapolis, M N 55455
R e c e i v e d 10 J a n u a r y 1979
ROBERTS, W. W. AND A. B. FROL. Interaction of central and superficial peripheral therrnosensors in control of thermoregulatory behaviors of the rat. PHYSIOL. BEHAV. 23(3)503-512, 1979.--The interaction of localized central and superficial peripheral thermal stimulation was studied in rats using diathermic warming of different brain areas controlling postural extension, locomotion, and grooming and a thermal floor warmed from thermally neutral 26.5 to 33.5 or 40.5°C, which induced locomotion and grooming. When behaviors were elicited by both central and superficial stimuli, combined stimuli were additive (activity) or partially additive subject to a ceiling (grooming). When behaviors were elicited by only one (or predominantly one) of two stimuli, combinations of the stimuli evidenced inhibition (grooming), multiplicative summation (activity), or mixtures of facilitation at low intensities and inhibition at high intensities (extension). Behaviors not elicited in significant amounts by either stimulus did not increase when both were combined (extension). It was concluded that the varied influences on thermoregulatory behaviors exerted by superficial and central thermosensors acting separately and in combination can largely explain the differences in behavioral response patterns induced by various thermal stress conditions that distribute heat differently between the surface and core of the body.
Thermoregulation Thermoreceptors Grooming Locomotor activity
Septal area Preoptic area Hypothalamus Midbrain
WHEN exposed to warm environments, rats typically in- crease their grooming and locomotor activity and, after some delay, spread out on the floor in an extended posture [11, 30, 35]. Since ambient heat produces a spatially and temporally complex pattern of stimulation of superficial thermosensors and deep thermosensors in the brain and body core, more specific thermal techniques have been used to warm lo- calized peripheral and central sites to determine their re- spective contributions to this behavior pattern. Warming limited to superficial or deep peripheral thermosensors elicits grooming and locomotor activity, but little or no postural extension [27]. Localized warming of a number of brain regions induces extension, grooming, or locomotion, depending on location. Postural extension is produced by warming of the medial preoptic and anterior hypothalamic area (MPO-AH) [29], the spinal cord [21], and the dorsal medulla [22], although the last was not observed by Roberts and Mooney [29]. Grooming is produced by warming of the medial posterior hypothalamus (MPH), and locomotor ac- tivity mainly by warming three zones located in the septal area (SA), medial midbrain (MM), and dorsomedial medulla [29].'This and other evidence that different groups of ther- mosensors have different effects on thermoregulatory re- sponses [1, 8, 10, 22, 26, 31, 32, 33] led to the conclusion that
thermoregulatory responses are not controlled by a unitary integrated signal representing a weighted combination of thermosensory inputs [5, 7, 14], but by separate and rela- tively independent linkages between particular thermosen- sors and particular response mechanisms, with any positive or negative summation between different inputs taking place within or functionally close to specific response mechanisms [29].
One predictable consequence of this fractional mode of organization is variation in the relative amounts of grooming, locomotion, and postural extension as a function of the dis- tribution of hyperthermia between superficial and central thermosensors. Since different thermal conditions, such as acute ambient heat, chronic ambient heat, and exercise, pro- duce different distributions of heat between superficial and central thermosensors, this fractional mode of organization could adjust the relative amounts of the three behaviors to maximize their joint effectiveness and efficiency for particu- lar conditions. Although some effects of different distribu- tions of hyperthermia can be predicted from available knowledge of the effects of localized superficial and central warming, the predicitions are limited by lack of knowledge of possible interaction effects of combinations of localized superficial and central warming, especially in cases where
1This research was supported by Grant MH 06901 from the National Institute of Mental Health to W. W. Roberts. Grateful acknowledge- ment is made to Eunice K. Roberts for assistance with the histology, and to Jennifer A. Nagel for assistance in collecting some of the data.
C o p y r i g h t © 1979 Bra in R e s e a r c h P u b l i c a t i o n s Inc . - -0031-9384/79/090503-10501.50/0

N 5C~ ROBERIS A, '1) t:R{)l.
responses are not elicited by both sets of sensors. The pur- pose of the present study was to examine the interactions of superficial thermosensors with several groups of central thermosensors to determine when summation occurs, and whether it is additive, multiplicative, or possibly even in- hibitory. A thermal floor, whose temperature could be in- creased rapidly for periods too brief to alter deep body tem- perature, was used as the source of superficial thermal stimu- lation. This was chosen partly because of its effectiveness in an earlier study [27] and partly because of its proximity to important peripheral thermosensors in the skin of the scrotum of the rats 115, 16, 17, 18]. Localized central warm- ing was produced by diathermy electrodes in the MPO-AH extension zone, the MPH grooming zone, or the SA and/or MM activity zones.
METHOD
Animals
Animals were Long-Evans pigmented male retired breeder rats aged 6-8 months and weighing 450-650 g ob- tained from Simonsen Laboratories, Gilroy, CA. They were housed at 23-25°C and were tested during the light phase of a 13 hr light, 11 hr dark cycle.
Apparatus
The source of the diathermic current was a 2 MHz Grass LM-2 lesion maker set for maximum output, with a variable resistance in series with the rat to control current level with maximal stability. The voltage drop across a 10 ohm resistor in series with the animal was rectified and monitored on a low frequency oscilloscope as a DC index of relative current level. An inductance was placed in parallel with the animal to shunt any DC current that might result from rectification in the biological or electronic parts of the circuit. The diather- mic current was conducted to the animal through overhead wires connected to a slip ring that allowed free rotation of leads.
The two test cages had copper floors 30.5 cm in diameter and 0.6 cm wire mesh walls 37.5 cm high. Soldered to the undersurface of each floor was a coil of 0.8 cm copper tubing that altered floor temperature when water of different tem- peratures was circulated through it. The undersurfaces of the floors were thermally insulated by 6 cm of urethane foam. The thermally conditioned water was supplied by three insu- lated tanks whose temperature was regulated by thermostat-controlled immersion heaters supplemented by immersion cooling coils. Two high capacity tubing pumps circulated water through the floor coils continuously, and a set of 2- and 4-way solenoid valves was used to switch con- nections between the two cages and the three tanks under the semi-automatic control of a bank of timers. The tempera- tures of the water tanks were set at levels that would produce maximum floor temperatures of 26.5, 33.5, and 40.5°C, Floor sUrface temperatures were measured 4 cm from the wall in the region most frequently occupied by the rat using a minia- ture copper-constantan thermocouple made of 0.15 mm di- ameter wires with a soldered junction about 0.20×0.25 ram, a thermocouple digital thermometer (Bailey BAT-8, Bailey Instruments, Saddle Brook, N J), and a polygraph. The floor temperature reached 60% of maximum 22 sec after the start of the temperature change, and 90% of maximum 43 sec later. The effect of heat exchange between body and floor on the temperature where the two were in contact was tested in
an anesthetized animal at body temperature~ of 34-.35 and 37-38°C. In general, the thermal capacity of the floor was great enough so that the body had relatively little effect on floor temperature in areas of contact. At the highest floor temperature, 40.5°C, the average static change when a forefoot, a hindfoot, or the scrotum was held over the ther- mocouple was only -0.6°C at the lower body temperature and -0.2°C at the higher body temperature. At the lowest floor
temperature, 26.5°C, the changes were 0.7 and 2.0°C, re- spectively, the latter probably due to vasodilation present at the higher body temperatnre.
Surgery
Under 60 mg/kg sodium pentobarbital anesthesia, dia- thermic warming electrodes were implanted stereotaxically centered on the midline in the MPO-AH area, MPH, SA, or MM, where local warming induces postural extension, grooming, or increased locomotion [29]. Most rats received a single electrode array, but a small number received two ar- rays located in the SA and MM areas for locomotor activity. The electrodes were made of four stainless steel insect pins. 0.40 mm in diameter, insulated with Formvar except for 2 or 3 mm at the tips and joined in a rectangular array so that the tips enclosed a 2 mm cube or a rectangle 2×2×3 mm. They were connected to a miniature socket attached to the skull with dental cement and stainless steel screws.
Pro~'edlll'#
Preliminary tests. After a minimum of one week of con- valescence, a series of preliminary diathermic warming ses- sions was given to determine the reliability and purity of elicited activity, grooming, and postural extension. Each session consisted of 10-40 rain adaptation in the test cages followed by 12-15 2 min trials or, later in the preliminary test sequence, 6-8 4 rain trials. Two rats received alternating trials in each session, so that the intertrial intervals for each animal averaged about 160 or 280 sec. The ambient tempera- ture was about 25°C. A water bottle was available on the cage wall, and the cages were separated by an opaque screen. During test trials, measures were taken of locmotor activity (number of times both hind feet crossed a quadrant line) and the duration of grooming, body extension with head up or on the floor, and rearing (forepaws off the floor, but not grooming). The duration measures were mutually exclusive. During intertrial intervals (i.e., while the other rat was tested with diathermy), the only behavior of the resting rat that was measured was the dominant category (activity, groomingi or extension) controlled by the thermosensitive area where the electrodes were implanted. Occasionally (<5% of sessions) animals became visibly drowsy and were gently aroused dur- ing one or two intertrial intervals by an air puff, nudge, or tilt of the cage that was not strong enough to directly elicit grooming or activity. The radio frequency current was started at a low level and increased by small steps after sev- eral trials at each level until a reliable response was obtained, behavioral evidence of a lesion appeared (e.g., hyperactiv- ity), or the current reached levels at which prior experience had shown lesions were likely. The small number of animals that had electrode arrays in both septal area and midbrain received separate tests with each electrode array during the first two sessions, after which both arrays were used to, gether for simultaneous warming of both sites. Since the two arrays of electrodes were connected to the diathermy source

CENTRAL AND S U P E R F I C I A L SENSORS IN T H E R M O R E G U L A T I O N 505
in parallel and the impedance of the septal electrodes (meas- ured by substitution) averaged 27% lower than that of the midbrain electrodes, it was necessary to add carbon resistors to the septai circuit to maintain an approximate balance of power consumption at the two sites. When comparison of response measures during diathermic warming trials and in- tertrial intervals indicated that extension, grooming, or ac- tivity was elicited rel iably by the thermal stimulus, a mod- ified staircase method was used to select a low current value that would elicit a slightly suprathreshold response and a higher current that would induce a moderately strong but submaximal response. About the same time, trials were lengthened to 4 min, the value used in the interaction tests. The average number of preliminary test sessions given all animals was 6.0 (range 2-20).
No attempt was made to measure the local temperature changes produced by the diathermy current. The signifi- cance and usefulness of such measures in the conditions of the present study was considered insufficient to warrant their costs in increased tissue damage caused by the addi- tional bulk of a thermocouple, and the potential distortion of the radio-frequency field by an additional pair of wires. Since the specific locations and density distributions of thermosen- sors are unknown, it is impossible to select appropriate sites for directly sampling receptor temperature [2]. In addition, the spatial pattern of warming produced by the four elec- trode array as visualized in albumin is complex and suscep- tible to alteration by varying brain resistivity. This together with the residual variability of stereotaxic placement creates too much spatial and interanimal variability in the stimulus applied to central receptors and in the location of ther- mocouples or thermistors for any single temperature meas- ure to have more than rough qualitative significance. Lastly, because many thermoregulatory responses are normally con- trolled by summation of thermosensors located in more than one brain region [2, 5, 21, 22, 29, 34], localized thermal stimulation usually requires greater than normal temperature changes [4] that cannot be accepted as quantitative indi- cators of the normal temperature-response function when thermal changes are general throughout the brain and spinal cord. However , since the temperature threshold for tissue damage is 45-49°C [3, 6, 9] and small lesions were evident in about one third of the rats in the interaction tests, it is likely that the average maximum temperature at the higher level of diathermy in the interaction tests was within 43-48°C. Since temperature is proportional to current squared and the ratio of the lower to the higher DC indexes of RF current was 0.67, the average maximum temperature at the lower diath- ermy level was probably within 40.0-M2.5°C.
The strength, purity, and trial-to-trial reliability of re- sponses to diathermy varied appreciably across animals, probably mainly due to stereotaxic variability but also possibly due to local variation in precise location or density of thermosensor cells. Since the relatively complex pattern of results often yielded by interaction studies requires con- siderable precision to establish significant trends and differ- ences in trends, it was necessary to select animals that dis- played maximally strong, pure, and reliable responses to diathermic warming. Of the total of 160 rats operated and given preliminary testing, 37 were selected to commence the interaction study with peripheral stimuli. Almost half of the rejected animals had electrodes in the vicinity of the SA, where the elicited activity was usually weak and variable from trial to trial, although consistent across animals if aver- aged across a large number of trials. Of 60 rats implanted
with SA electrodes alone, 77% were more active on diath- ermic warming trials than on control trials, 15% were more active on control trials, and 8% were equally active (omitting equal cases, sign test z=4.99, p<0.001). When diathermy electrodes were implanted in both septal area and midbrain, the yield of adequately strong and reliable activity responses was much improved. Although most rats were eliminated by quality criteria, 24 were lost due to accidental brain lesions (as inferred from behavioral changes) or detachment of the socket. After initiation of the interaction tests, 8 of the 37 rats that began the test were eliminated because of emergence of a second response (2), accidental creation of a large lesion (2, confirmed in histology in one case, inferred from behavior in other), and loss or weakening of the initial response (4, possibly due to lesioning, though not histologi- cally verified). Consequently, the final experimental data was obtained from 8 rats having MPO-AH electrodes that elicited posturai extension, 11 rats having MPH electrodes that elicited grooming, and 10 rats whose electrodes elicited locomotor activity, of which 4 had electrodes in the MM, 1 in the SA, and 5 in both areas.
Interaction tests. The test conditions consisted of all nine possible combinations of three levels of central warming (zero, slightly suprathreshold, and moderately suprathresh- old) and three levels of peripheral warming via the thermal floors (26.5, 33.5, 40.5°C). All conditions were tested once in each 9-trial daily session. The 29 experimental rats were tested for 12 sessions, except for three who received 6, 8, or 10 sessions because they died or the cranial socket became detached. The order of stimuli was counterbalanced within and across animals, with the requirement that each level of central warming and each level of peripheral warm- ing should occur once in each block of 3 trials. Two rats were tested in each session with each rat ' s test trials occurring during the intertrial intervals of the other rat. Before the start of a session, the animals received 10-30 min adaptation in the test cages. At the beginning of each trial, the thermal floor stimulus was initiated 15 sec before the onset of central diathermic warming because of the slower onset of the form- er. For 240 sec following the onset of central warming, grooming, activity, extension, and rearing were timed or counted as in the preliminary tests. At the end of the trial, the diathermic stimulus was turned off, the floor temperature was returned to 26.5°C, and the data was recorded during a 60 sec period before initiation of the next trial for the other rat. Thus, the intertrial interval for each rat was about 375 sec. Unlike the preliminary testing, behavior measures were not taken during the intertrial interval. The ambient tempera- ture recorded from a thermometer between the cages at the start and end of each session averaged 25.1°C. Water was continually available but was seldom consumed.
If the strength of responses to diathermy on trials when the floor temperature was neutral (26.5°C) appeared to in- crease or decrease in strength in the course of the test series, suggesting a change in threshold, additional diathermy trials were given after the regular test session to verify the threshold change and determine the adjustment in current needed to compensate for it. As during the preliminary tests, when animals occasionally (<5% of trials) became visibly drowsy, they were aroused by gentle tactile stimuli or tilting during the intertrial interval.
Histology
After completion of testing, the 29 rats that completed the interaction tests and a small number that were eliminated in

506 ROBER'IS AND FROI,
preliminary testing were anesthetized and perfused with 10% Formalin, and frozen section histology was performed. Elec- trodes were localized in the atlas of Pellegrino and Cushman [25] by direct projection of tissue sections on atlas diagrams at matched magnification.
Statistical Analysis
Each rat 's scores were averaged across sessions to obtain a single value for each response measure. The data for each group was first subjected to an overall 2-way analysis of variance for repeated measures [361 for each response meas- ure to test main effects of central and peripheral warming and their interaction. Next, one-way anovas were performed at each level of each variable to determine the reliability of effects at each level, which was mainly useful in cases of significant interactions. A few comparisons of pairs of points were made with t tests, or with the Wilcoxon test when zero scores were numerous.
RESULTS AND DISCUSSION
Single Thermal Stimuli
When superficial and central thermal stimuli were pre- sented separately, they elicited behaviors similar to those obtained in earlier studies employing somewhat different apparatus and procedures [27,29].
Superficial warming. Warming of superficial peripheral receptors alone via the thermal floor elicited principally grooming, locomotor activity, and rearing as indicated by the dotted lines in Fig. 2B, -D, -F and 3B, -D, -F. These effects were significant well beyond the 0.001 level when all three groups were combined, F 's(2,56)= 12.27-55.67, and in most cases reached the 0.05 level or better within groups. Rearing, which occurred only 1/5- 1/3 as much as grooming, is not presented in a separate figure because it evidenced trends similar to locomotor activity. Since rearing also resembled locomotion in the frequency with which it was accompanied by sniffing and looking around, rearing and locomotion may represent upward and horizontal components of a general activity syndrome facilitated by thermal stimuli [23]. In ad- dition to the main responses of grooming, locomotion, and rearing, the warm floor also induced very weak extension shown by dotted lines in Fig. 1B, -D, -F, Wilcoxon T(24)=0, p<0.001. In terms of mean duration per min, the extension induced by the warm floor was only 8% of the amount eli- cited by the higher level of MPO-AH warming and only 13% of the maximum amount displayed toward the end of earlier ambient heat tests summarized in Fig. 4. The failure to detect this weak effect in the earlier study of Roberts and Martin [27] was probably due to the smaller number of animals and test trials or other procedural differences. The possibility that the warm floor might have raised MPO-AH or spinal cord temperatures enough to cause the extension was exam- ined in two conscious rats, each of which had a miniature thermocouple chronically implanted in the MPO-AH region and another in a reentrant tube in the dorsal abdominal cav- ity near the aorta. The recording system had a resolution of less than 0.1°C and the schedule of floor temperatures, trial durations, and intertrial intervals was the same as in the interaction tests. The average change in brain and abdominal temperatures from the beginning to the end of trials with the 40.5°C floor temperature was 0.02 and 0.09°C respectively,
thus ruling out the possibility that the thermal floor might have raised brain and/or body core temperatures.
The 33.5°C floor temperature failed to elicit significant increases in locomotion, grooming or rearing, indicating that the threshold for elicitation of these responses by the ventral skin area warmed by the floor lies above 33.5°C. This is in agreement with the report of Hellon, Hensel, and Schafer [ i 5] that the thresholds of warm thermosensors in the skin of the rat scrotum fall within a range of about 33-36°C. Addi- tional thermosensitivity has been found in the rat or other species in a variety of superficial sites that were not exposed to the warm floor, including the face, nasal air passage, ears, and back [19, 20, 28]. Since there is considerable spatial summation of peripheral thermosensor input in the elicita- tion of grooming and locomotion [28], it is likely that the functions relating behavioral responses to surface tempera- tures are steeper during natural ambient heat exposure when all superficial sensors are warmed than during the spatially limited ventral warming in the present study.
Since the highest floor temperature, 40.5°C, was within or c lose to the upper range of environmental t empera tures that feral rats may normal ly exper ience and did not elicit stronger responses than 37-38°C ambient heat, it was prob- ably no more aversive or traumatic than elevated tempera- tures that occur under natural conditions. It was well below the lowest available determination of rats" escape threshold for heat pain, 43.7°C, or the threshold of nociceptive neurons in the cord or thalamus of rats, 42-43°C [241.
Central warming. In the absence of superficial thermal stimulation, central diathermic warming of the MPO-AH area induced postural extension shown by the dotted line in Fig. 1A, warming of the SA/MM areas elicited locomotor activity and rearing shown in Fig. 3E, and warming of the MPH area predominantly induced grooming shown in Fig. 2C, F's(2,14--20)= 13.17-133.92, a l l p ' s <0.001. MPH warm- ing also elicited a small increase in activity (Fig. 3C), about 22% of the amount produced by SA/MM warming, F(2,20)=8.44, p<.003. In addition to these facilitatory e f fects, MPO-AH warming inhibited grooming (Fig. 2A), F(2,14)=4.68, p<0.03. SA/MM warming produced a similar inhibitory trend (Fig. 2E) that fell short of statistical significance in tests of central warming alone, but was sig- nificant in the overall anova combining SA/MM effects at all floor temperatures, F(2,18) =5.03, p <0.02.
Examination of the histology of the 29 rats that provided data in the interaction tests confirmed that the diathermic warming electrodes were located in brain areas where ex- tension, grooming, and locomotion were obtained in an ear- lier study [29]. The average midline coordinates of the cen- ters of the electrode arrays in the atlas of Pellegrino and Cushman [25] were: MPO-AH, A7.7, H-I.0; MPH, A5.4, H-2.5; SA, A8.9, H2.1 : MM, A2.7, H-2.1. These values were very close to the centers of the effective areas for diathermic warming in the study of Roberts and Mooney [29], except for a lmm dorsal shift of the SA electrodes to minimize possible overlap with the MPO-AH zone and a lmm anterior shift of the MPH electrodes to minimize overlap with the MM zone. A small amount of tissue damage ( l -3mm a) was evident in eight cases and a larger amount (3.8, 6.7mm:b in two cases. The ratio of the lesion volume to the 8 or 12 mm :~ zone enclosed by the uninsulated electrode tips averaged 0.21. The animals having lesions did not evidence weaker behav- iors in the interaction tests, but the currents required to elicit the behaviors averaged 35% higher than in the nonlesioned rats.

CENTRAL A N D S U P E R F I C I A L SENSORS IN T H E R M O R E G U L A T I O N 507
PO-AH C=.O01 P..02 CP=.O01
35
5 0
25
20.
Z
(.) b.I t n I0.
Z 5.
Z 0
I-- 0
A p,3&5=.O01 / / " ~'~I ~'6.5=.001
/ / f
,, ,, " ......"" ~,,, , ,~40.5~,00t
/,., / .,.'"
C l:3
Z 0
T40.5 ".14 Z . . . . . ~33 .s t~ .-. 2 . . ' : . - . - .7 . . . . . . . . . . . . . . . .+ize.5 I-- • • "
xSA/MM E
0 ~. :...:_..: . . . . . . . . . . . . . x . . . . . . . . . . . 1 6 L'O vii
, C E N T R A L W A R M I N G
J 71
I
s
B
~ f l l = 002
~ - i LO=.O5
..~ . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . iO
D
Io I . . . . . -.::_: _ , ,t .o - - .L .{jill ~ : : . - - - . ~ : :: -- . . . . .
z&5 3&5 46.s F L O O R T E M P E R A T U R E , °C
FIG. 1. Effects of central and superficial peripheral warming on postural extension during 4 min trials. Data plotted with respect to central warming on left, with respect to peripheral warming on right. MPO-AH=medial preoptic-anterior hypothalamic warming, MPH=medial posterior hypothalamic warming, SA/MM=septal area and/or medial midbrain warming. P values of overall anovas in upper left; C=central, P=peripheral, CP=central-peripheral interaction. P values of 1-way anovas for each stimulus level on right of each curve. Anovas not used
where zero scores were numerous. Bars indicate standard errors.
Combined Thermal Stimuli
Postural extension. Fig. 1A and -B show how the exten- sion response was affected when central warming of the MPO-AH area, which by itself elicits strong extension, was combined with superficial peripheral warming via the ther- mal floor, which induces very weak extension when pre- sented alone. The effect of the thermal floor on centrally- induced extension was biphasic, as indicated by a significant interaction, F(4,28)=10.90, p<0.001. At the high level of MPO-AH warming, the addition of superficial warming at 40.5°C reduced extension by about 33% below the amount at 26.5°C or 33.5°C, t 's(7)=2.72, 5.72, p ' s <0.03, 0.001. At the low level of MPO-AH warming, which by itself elicited only about one third as much extension as the high level, the addition of mild superficial warming at 33.5°C produced a small but s ignif icant inc rease in ex tens ion , t (7)=2.38 , p <0.05.
The reduction in extension at the high level of MPO-AH warming may have resulted from response interference from the increased grooming and locomot ion el ic i ted by the 40.5°C floor and/or from possible mild aversiveness of pro- longed ventral contact with the warm but nonpainful floor
during sustained extension, perhaps related to the unfavor- able heat exchange when floor temperature exceeds body temperature. The mild facilitatory effect of the low level of peripheral warming on the smaller amount of extension el- cited by the low level of MPO-AH warming is consistent with the capacity of the warm floor alone to elicit weak ex- tension, but the absence of significant summation at higher levels of central or peripheral warming and the significant inhibition when both were high indicate that the contribution of peripheral thermal stimuli to the elicitation of extension is small and limited to relatively mild central and superficial heat levels.
Fig. 1C, -D, -E, -F show the effects of combining warming of the MPH and SA/MM areas, which did not induce exten- sion when presented alone, with peripheral warming, which elicited weak extension. Because of numerous zero scores and positive skewing, it was not feasible to apply analysis of variance. However , the generally low scores under all con- ditions indicate that MPH and SA/MM thermosensors were as ineffective in facilitation of extension as in its direct elici- tation.
Grooming. Fig. 2C and -D show the effects on grooming of combined warming of the MPH area and the thermal floor,

508 RO BE RT S ANI) FROI.
MPO-AH C:.02 P:.O01 A CP:.02
15.
I0 1 k ~ ~ \ "-- ] ~ ~ 40.5-%14
I .......................... . . . . . . . .
1 2 6 . 5 = , 0 3 o
MPH Z 2 5
(.) LLI20 (,9
7
15• Z 0
I--
or I0,
£3
( -9 5 Z
o o Q: 0
~sA/MM 15.
IO
C / / ~ 33.5=.001
C='O01 t ~ 40.5=.001 P=O02 C P = . O O ' ~ . . . ' I 26.5=.001
] . ' . . - . " /
...<.~ . f -
.,.)
/ /
/
C=.02 P=,07
............................. . . . . . . . . . . . " " " • 40.5 =.07
-" ~" "~" :.-.I 26.5=-25 l I 33.5=,02
B T OLO03
.... "r LO=,O01
" T / " • ' ~ 1 H 1=.002
- < 7 I /
D
J
. . . . . . . . . . 1'" " - • •~o=.oo,
. . . . . . . . . . . . . . . . . I O <09
_ . . . . . . . . . . . . . . . . . . . . I . . . . . _ . . . . . . . Lo .52
I '
(~ L'O H'I 2~.5 3~).5 46 .5 CENTRAL WARMING FLOOR TEMPERATURE, °C
FIG. 2. Effects of central and superficial peripheral warming on grooming during 4 min trials. Abbreviations as in Fig. 1.
which both elicit grooming when warmed separately. There was a significant interaction of central and peripheral stimuli, F(4,40)=10.54, p<0.001, chiefly due to summation at the lower level of central warming, but not at the higher level, where grooming appeared to be limited by a ceiling in the vicinity of 22-25 sec/min, which was about 25% greater than the maximum amount evidenced during exposure to ambient temperatures of 37-38°C in previous studies summarized in Fig. 4.
Fig. 2E and -F show the effect of combining SA/MM warming, which inhibits grooming, with peripheral warming, which elicits grooming. The resulting negative summation
was near-additive, since the interaction fell far short of s tat i s t ica l s ign i f i cance . H o w e v e r . the interact ion of MPO-AH warming, which also inhibits grooming, with peripheral warming shown in Fig. 2A and -B was significant. F(4,28)=3.83, p<0 .02 , mainly due to the greater inhibitory effect of the MPO-AH warming on grooming elicited by the lower level of floor warming. Whether the inhibitory effect of SA/MM and MPO-AH warming on grooming was due to specific inhibition of grooming or to general response inter- ference from the strong locomotor activity or extension that they principally induced cannot be determined from the available data. The finding that MPO-AH warming did not
CENTRAL AND S U P E R F I C I A L SENSORS IN T H E R M O R E G U L A T I O N 509
M P O - A H A 40.5=.001
2.0 c =,oo= / I P=.ool / I
1.5 = " ±
1.0. ~
_ _[ . . . . . . . . . . . . I33s=2~ I - . . . . 1 - = ..... ~ . ' : . 7 . . ~ .
t . . . . . . . . 1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I z 6 ' ~ ' ~ 0
Mq Z 1.5.
if) ~.0 1.0. Z 03 o n.- .5. o
C . / . . . . . . . . . . . / ~ 4 0 5 001 • =.
C=.O01 P=.O01 C P = . O 0 7 ~
I I 265-003 ~- ~ .,;. :. ~ "~" ~-,~ 33.5=.00
>_- ~ ....... _= =.._. . . . . . . . . . . . . -.~ . . . . . . . . . . . . . .
t - 0
r--- S A / M M (..) < 3.0. E
C= 002 ~ 40.5-.03 n.- P=~OO~ / _1 o ,_ _ / II 0 2.5.
~ ....... ~ 26.s..oo,
/ , .....
I. ' 1.5. •
I.o. ..: -::-' :~2"2" I "
.5. I"
0
B HI,.O01
/ / / ~ [L.O=.O01
, , , , / "~,,,,, . . . . . I o'.oo=
T / . - " i
D
~ H I = .001
t~ lO=.OOj
_._I ~ . ~ . . . - ....... t o oo,
I . . . . . . . . . . . . . / ' ~ . ........
F ~ . ~ ~ HI =.001
. ~ ~ ~ ~ .." .. "" "'..L O='O01T
T . . . . . . . . . . . . . 1"
1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . I
6 L'O I'll 26.5 3'5.5 4().5 C E N T R A L WARMING FLOOR TEMPERATURE, *C
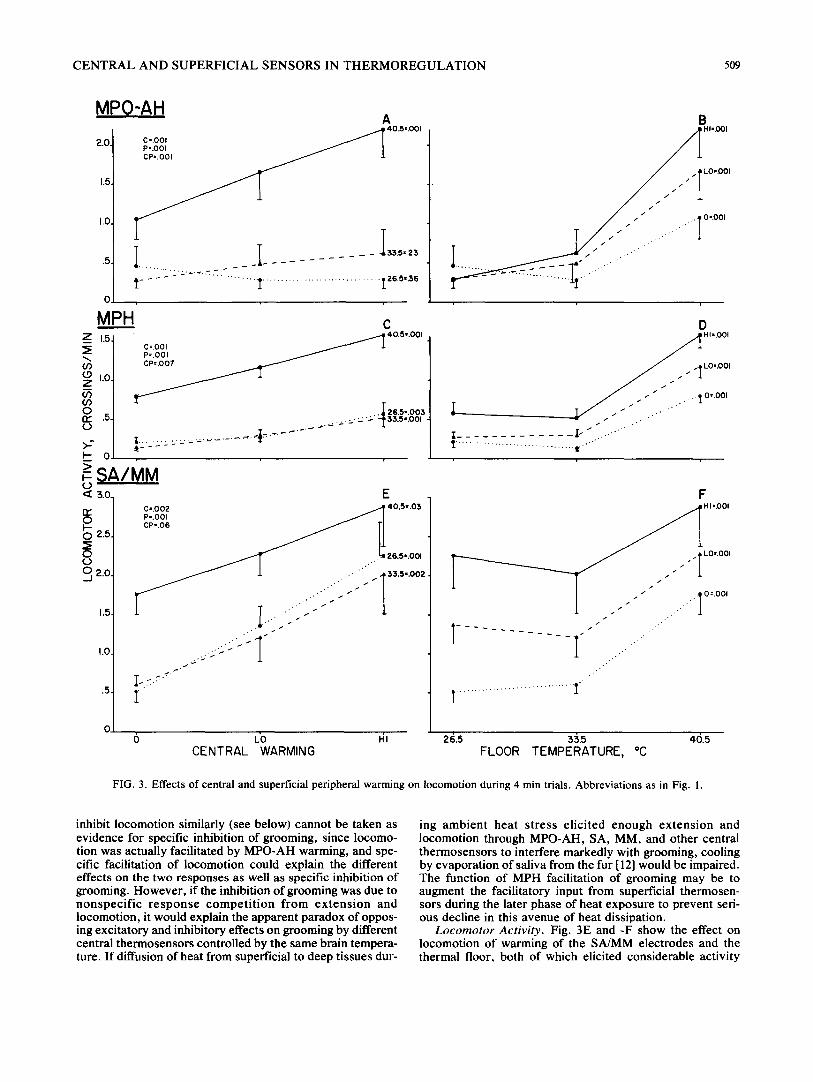
FIG. 3. Effects of central and superficial peripheral warming on locomotion during 4 min trials. Abbreviations as in Fig. 1.
inhibit locomotion similarly (see below) cannot be taken as evidence for specific inhibition of grooming, since locomo- tion was actually facilitated by MPO-AH warming, and spe- cific facilitation of locomotion could explain the different effects on the two responses as well as specific inhibition of grooming. However , if the inhibition of grooming was due to nonspec i f i c r e s p o n s e c o m p e t i t i o n f rom ex t ens ion and locomotion, it would explain the apparent paradox of oppos- ing excitatory and inhibitory effects on grooming by different central thermosensors controlled by the same brain tempera- ture. If diffusion of heat from superficial to deep tissues dur-
ing ambien t hea t s t ress e l ic i ted enough ex tens ion and locomotion through MPO-AH, SA, MM, and other central thermosensors to interfere markedly with grooming, cooling by evaporation of saliva from the fur [121 would be impaired. The function of MPH facilitation of grooming may be to augment the facilitatory input from superficial thermosen- sors during the later phase of heat exposure to prevent seri- ous decline in this avenue of heat dissipation.
Locomotor Activity. Fig. 3E and -F show the effect on locomotion of warming of the SA/MM electrodes and the thermal floor, both of which elicited considerable activity

510 ROBERTS AND FRO].
2O Z
"~ f5
z JO
7
b--
":~ 5, c~
c3
O.
GROOMING
e, • > -
>
F-
LOCOMOTION
4 .
3;
o I o 2 0 30 4 0 ' I ( ) ' 2 '0 ' 3'0 ' 4'O TIME IN MIN TIME IN MIN
Z
20
~5 Z
I0.
p - <~
0 0
B O D Y E X T E N S I O N
f ~
/ // /
I0 20 30 4 0 TIME IN MIN
FIG. 4. Different temporal patterns of grooming, locomotion, and extension during exposure to neutral and elevated ambient temperatures. Mean of results from 245 rats in published [28,30] and unpublished studies. Dotted lines represent tests at 25-27°C and solid lines tests at 37-38°C. Largest standard error for grooming was 0.76 and for locomotion was 0.32. Activity measures obtained in stabilimeters; absolute values not directly comparable with Fig. 3. Rectal temperatures averaged 38.0°C at the end of
25--27°C tests and 39.9~C at end of 37-38°C tests.
when presented alone. In combination, the summation of the two thermal stimuli did not depart significantly from additivi- ty, interaction F(4,36)=2.49, p<0.06. In Fig. 3C and -D, the combination of MPH warming, which elicited a small amount of locomotion, with the thermal floor evidenced a small but significant "mult ipl icat ive" interaction in which the slopes of curves relating locomotion to warming of one are increased when the other area was warmed, F(4,40)=4.13, p<0.007. In Fig. 3A and -B, the combination of MPO-AH warming, which had no significant effect on locomotion when presented alone, with peripheral warming produced a similar though stronger multiplicative interac- tion, F(4,28)=7.33, p<0.001. The capacity of MPH and MPO-AH warming to elicit and/or facilitate locomotion to a small to moderate degree is consistent with an earlier finding that weak or high threshold activity was produced by some warming electrodes in these areas [29].
Rearing, which followed similar trends to locomotor ac- tivity under most conditions, failed to evidence facilitation during combined MPO-AH and peripheral warming, prob- ably because the elicited extension interfered more with ver- tical rearing than horizontal locomotion.
Relation to Ambient Heat Behavior
A major aim in the analysis of the contributions of differ- ent groups of localized thermosensors in the control of ther- moregulatory responses is an eventual synthesis of the ther- moregulatory response pattern under different conditions of thermal stress. In the case of the rat (and probably some other species), where different responses or combinations of responses are controlled by different groups of central and peripheral thermosensors, fully quantitative synthesis pre- sents major difficulties and it may be necessary to settle for qualitative or semiquantitative formulations. Of especial im- portance is the changing distribution of thermosensor input as the rat encounters, endures, and under free ranging con- ditions, escapes from ambient heat. At first, only superficial peripheral thermosensors are warmed by elevated air and ground temperatures. With continued exposure, deep body temperatures of rats slowly rise by 2-3°C over a period of 30-60 min [13], causing a gradual warming of central and deep peripheral thermosensors. During the initial period of superficial thermosensor dominance and the subsequent
shift to combined superficial and deep thermosensor control, rats ' behavioral responses to heat follow disparate time courses. Fig. 4 shows the different temporal patterns of grooming, locomotion, and postural extension during 45 min exposure to moderate ambient heat at 37/38°C for a large number of rats in previous published [28,30] and unpublished studies.
Most of the response differences apparent in Fig. 4 can be explained by the effects of separate and combined superficial and central thermal stimulation studied in the present and preceding papers [27,29]. Thermoregulatory grooming ap- pears in the first 5 min of exposure to ambient heat, and increased locomotion is evident in the second 5 min after the decline of exploratory activity evoked at both neutral and warm temperatures by the shift of cages into the thermal chamber. Both of these strong early responses and the ab- sence or near absence of postural extension may be attrib- uted to the capacity of superficial peripheral thermosensors to elicit grooming and locomotion, and their near-complete inability to induce extension [27]. As exposure to ambient heat continues, and slowly rising deep temperatures gradu- ally warm central and deep peripheral thermosensors [13], grooming and locomotion follow separate time courses, and postural extension appears and steadily increases. During the first 15 min, grooming rises moderately, F(2,488)=4.16. p <0.01, perhaps because of the changed environment, since a similar increase in grooming is evident at the neutral tem- perature, or because of the gradual warming of the cage floor and microclimate just above it, which required 10-15 min to reach 90% of maximum. After the first 15 rnin of ambient heat, grooming declines steadily to about 50% of the maximum, F(6,1464)=36.77, p<0.001, while locomotion rises steadily to about 50% above the amount during the second 5 rain, F(7,1708)=6.61, p<0.001. The decline in grooming may be attributed to the inhibition or interfering responses produced by central thermosensors in the MPO- AH, SA, and MM areas in the present study, perhaps sup- plemented by other sensors, while the increase m locomo- tion may be attributed to the uniformly facilitory influence of MPO-AH, MPH, SA, MM and possibly other thermosen- sors. The slow and steady rise of postural extension during ambient heat exposure follows the same time course as the gradual rise in deep body temperature under similar condi- tions [13] and may be attributed to the near-complete de-

C E N T R A L AND S U P E R F I C I A L SENSORS IN T H E R M O R E G U L A T I O N 511
pendence of extension on central thermosensors in the MPO-AH, spinal cord, and possibly the medulla [21, 22, 29].
The inhibition of high levels of extension exerted by the 40.5°C floor (Fig. 1B) suggests that extension may be some- what damped by superficial thermosensors at high ambient temperatures. If so, escape from ambient heat followed by rapid reduction of superficial sensor input with maintained central sensor activity would be expected to produce a tem- porary increase in extension. At the same time, activity and grooming would be expected to fall markedly, partly because of withdrawal of the potent influence of superficial ther- mosensors over these responses, partly because MPO-AH and MPH facilitation of locomotion is largely multiplicative and therefore dependent on simultaneous input from super- ficial thermosensors, and partly because MPO-AH and SA/MM inhibition of grooming would oppose MPH facilita- tion of grooming. Such a behavioral shift after heat escape would reduce heat production by muscle activity, diminish the likelihood of further locomotion back to a warm en- vironment, and conserve body water. A similar outcome
would be predicted in cases of endogenous hyperthermia produced by exertion and would have similar benefits. While the effect of heat escape or exercise hyperthermia on behav- ioral responses has not been tested directly, a recent study in our laboratory invest igated the effects on extension grooming, and activity in a thermally neutral environment of 1.5-2.0°C increases in deep body temperature produced in one experiment by microwaves and in the other by sub- cutaneous injection of isoproterenoi in rats previously adapted to cold. In both cases, deep hyperthermia in a neu- tral environment elicited exceptionally large increases in postural extension that were 60-100% greater than the maximum induced by ambient heat in Fig. 4. Grooming and activity, on the other hand, failed to increase significantly and in most cases declined below control levels. These find- ings confirm the prediction with respect to the general ther- mal conditions present after escape from ambient heat or exercise hyperthermia and extend the generality of the frac- tional input-output analysis in this and preceding papers to a different pattern of superficial and deep hyperthermia.
REFERENCES
1. Adair, E. R. Hypothalamic control of thermoregulatory behav- ior. In: Recent Studies o f Hypothalamic Function, edited by K. Lederis and K. E. Cooper. Basel: Karger, 1974, pp. 341-358.
2. Adair, E. R. and R. O. Rawson. Autonomic and behavioral temperature regulation: Unilateral vs bilateral preoptic thermal stimulation. Pflugers Arch. ges. Physiol. 352: 91-103, 1974.
3. Alberts, W. W., E. W, Wright, B. Feinstein and G. von Bonin. Experimental radiofrequency brain lesion size as a function of physical parameters. J. Neurosurg. 25: 421-423, 1966.
4. Baldwin, B. A. and J. O. Yates. The effects of hypothalamic temperature variation and intracarotid cooling on behavioral thermoregulation in sheep. J. Physiol. 265: 705-720, 1977.
5. Bligh, J. Temperature Regulation in Mammals and other Ver- tebrates. Amsterdam: North Holland, 1973.
6. Brodkey, J. S., Y. Miyazaki, F. R. Ervin and V. H. Mark. Reversible heat lesions with radiofrequency current: A method of stereotactic localization. J. Neurosurg. 21: 4%53, 1964.
7. Cabanac, M. Temperature regulation. Ann. Rev. Physiol. 37: 415--439, 1974.
8. Carlisle, H.J. and D. L. Ingram. The effects of heating and cooling the spinal cord and hypothalamus on thermoregulatory behaviour in the pig. J. Physiol., Lond. 231: 353-364, 1973.
9. Dusser de Barenne, J. G. and H. M. Zimmerman. Changes in the cerebral cortex produced by thermocoagulation: A sugges- tion to neurosurgery. Archs. Neurol. Psychiat. 33: 123-131, 1935.
10. Fuller, C. A., B. A. Horwitz and J. M. Horowitz. Shivering and nonshivering thermogenic responses of cold-exposed rats to hypothalamic warming. Am. J. Physiol. 228: 151%1524, 1975.
11. Hainsworth, F. R. Saliva spreading, activity, and body tempera- ture regulation in the rat. Am. J. Physiol. 212: 1288--1292, 1967.
12. Hainsworth, F. R. and E. M. Stricker. Salivary cooling by rats in the heat. In: Physiological and Behavioral Temperature Regulation, edited by J. D. Hardy, A. P. Gagge and J. A. J. Stolwijk. Springfield, II1.: Thomas, 1970, pp. 611-626.
13. Hainsworth, F. R. and E. M. Stricker. Evaporative cooling in the rat: Differences between salivary glands as thermoregula- tory effectors. Can. J. Physiol. Pharmac. 49: 573-580, 1971.
14. Hammel, H. T. Regulation of internal body temperature. Ann. Rev. Physiol. 30: 641-710, 1968.
15. Hellon, R. F., H. Hensel and K. Schafer. Thermal receptors in the scrotum of the rat. J. Physiol. 248: 34%357, 1975.
16. Hellon, R. F. and N. K. Misra. Neurones in the dorsal horn of the rat responding to scrotal skin temperature changes. J. Physiol. 232: 375-388, 1973.
17. Hellon, R. F. and N. K. Misra. Neurones in the ventrobasal complex of the rat thalamus responding to scrotal skin tempera- ture changes. J. Physiol. 232: 38%399, 1973.
18. Hellon, R. F., N. K. Misra and K. A. Provins. Neurons in the somatosensory cortex of the rat responding to scrotal skin tem- perature changes. J. Physiol. 232: 401-411, 1973.
19. Kenshalo, D. R., D. G. Duncan and C. Weymark. Thresholds for thermal stimulation of the inner thigh, footpad, and face of cats. J. comp, physiol. Psychol. 63: 133-138, 1967.
20. Kluger, M. J., R. R. Gonzalez and J. D. Hardy. Peripheral thermal sensitivity in the rabbit. Am. J. Physiol. 222: 1031-1034, 1972.
21. Lin, M. T., T. H. Yin and C. Y. Chai. Effects of heating and cooling of spinal cord on CV and respiratory responses and food and water intake. Am. J. Physiol. 223: 626-631, 1972.
22. Lipton, J. M. Thermosensitivity of medulla oblongata in control of body temperature. Am. J. Physiol. 224: 890-897, 1973.
23. Mackintosh, J. H., M. R. A. Chance and A. P. Silverman. The contribution of ethological techniques to the study of drug ef- fects. In: Handbook of Psychopharmacology. Vol. 7. Principles of Behavioral Pharmacology., edited by L. L. Iversen, S. D. Iversen and S. H. Snyder. New York: Plenum Press, 1977, pp. 3-35.
24. Mitchell, D. and R. F. Hellon. Neuronal and behavioral re- sponses in rats during noxious stimulation of the tail. Proe. R. Soc. Lond. B. 197: 16%194, 1977.
25. Pellegrino, L. J. and A. J. Cushman. A Stereotaxic Atlas of the Rat Brain. New York: Appleton-Century-Crofts, 1967.
26. Roberts, W. W., E. H. Bergquist and T. C. L. Robinson. Ther- moregulatory grooming and sleep-like relaxation induced by local warming of preoptic area and anterior hypothalamus in opossum. J. comp. physiol. Psychol. 67: 182-188, 1969.
27. Roberts, W. W. and J. R. Martin. Peripheral thermoreceptor control of thermoregulatory responses of the rat. J. comp. physiol. Psychol. 87: 111)9-1118, 1974.
28. Roberts, W. W. and J. R. Martin. Effects of lesions in central thermosensitive areas on thermoregulatory responses in rat. Physiol. Behav. 19: 503-511, 1977.
29. Roberts, W. W. and R. D. Mooney. Brain areas controlling thermoregulatory grooming, prone extension, locomotion, and tail vasodilation in rat. J. comp. physiol. Psychol. 86: 470--480, 1974.
30. Roberts, W. W., R. D. Mooney and J. R. Martin. Ther- moregulatory behaviors of laboratory rodents. J. comp. physiol. Psychol. 86: 693-699, 1974.

512 R O B E R T S AN 13 FR()[ .
31. Satinoff, E. Neural integration of thermoregulatory responses. In: Limbic and Autonomic Nervous Systems Research. edited by L. V. DiCara. New York: Plenum Press, 1974, pp. 41-83.
32. Satinoff, E. Neural organization and evolution of thermal regu- lation in mammals. Science 201: 16-22, 1978.
33. Schmidt, I. Effect of central thermal stimulation on the ther- moregulatory behavior of the pigeon, t~fl,gers Arch. ,ges. Physiol. 363: 271-272, 1976,
34. Simon, E. Temperature regulation: The spinal cord as a s~e of extrahypothalamic thermoregulatory functions. /x'cv~ P/;3si,/ Biochem. PharmacoL 71: 1-76, 1974.
35. Swift, R. W. and R. M. Forbes. The heat production of the fasting rat in relation to the environmental temperature. J. N,tr. 18: 307-318, 1939.
36. Winer, B. J. Statistical Principles in Experimental l)esi~,n. New York: McGraw-Hill, 1971, p. 575.