integral lipid in human hair follicle - … · integral lipid in human hair follicle won-soo lee,...
TRANSCRIPT

Integral Lipid in Human Hair Follicle
Won-Soo Lee,� Tak Heon Oh,� Seung Hyun Chun,� Soo Young Jeon,� Eun Young Lee,� Sanghoon Lee,�
Won-Seok Park,w and Sungjoo Hwangz�Department of Dermatology and Institute of Hair & Cosmetic Medicine, Wonju College of Medicine, Yonsei University, Wonju, Korea; wAmorepacific R&D Center,Yongin, Korea; zDr Hwang’s Hair Clinic, Seoul, Korea
On the hair surface, cells are covered with a thin lipid layer (LL) covalently bonded to hair proteins. This integral
hair lipid is different from sebaceus lipid. We conducted this study to examine the lipid distribution in human hair
follicle. Transmission electron microscopy was performed to observe the ultrastructure of the LL. Hair follicles
were cut and observed longitudinally along the hair axis. For transmission electron microscopy, new fixative (Lee’s
fixative: composed of OsO4 and RuO4) was designed as the conventional fixatives such as OsO4 or RuO4 alone
were not appropriate for staining for hair follicle lipid. In addition, we measured the chemical composition of
integral hair lipid by high-performance thin-layer chromatography. From the above experimental procedure, it was
discovered that the lipid in the hair follicle was mainly distributed in hair cuticle and keratinized inner root sheath. A
multitude of lamellar granule is observed in the vicinity of the above LL. The chemical composition of integral hair
lipid was different from those of epidermal or sebaceous lipids. It is assumed that the LL in the hair follicle is similar
to the epidermal LL playing an important role as a skin barrier in the stratum corneum. We proposed here the new
terminology ‘‘hair barrier’’ from this point of view.
Key words: hair barrier/hair follicle/integral hair lipid/transmission electron microscopyJ Investig Dermatol Symp Proc 10:234 –237, 2005
In mammals, the lipid layer (LL) has been known to preventpercutaneous water loss and regulate percutaneous ab-sorption in the stratum corneum (Lampe et al, 1983). And,the amount of lipid content is a more important factor forpercutaneous absorption of certain substance than thethickness and number of LL in the stratum corneum (Eliaset al, 1981). The LL in the stratum corneum has been knownto function as a physiologic barrier, but there have been fewstudies on LL and its function in hair. Furthermore, therehave been no reports on lipid distribution in hair folliclewithin scalp tissue. The lipid distribution on the hair surfacehas been reported in recent years. The lipid content of hu-man hair, takes up to 0.7%–1.3% in total components ofhair and is different from the composition of the lipid in theepidermis. This lipid content in hair shows no significantdifference in each mammal unlike in sebaceous gland (Ni-cloaides et al, 1968; Lee et al, 1996; Jolles et al, 1997; Be-rtrand et al, 2003). The LL in hair is the only continuousstructure that plays a key role in maintenance of hair integ-rity including hydrophobicity and stiffness. On the hair sur-face, cells are covered with a thin LL covalently bonded tohair proteins (Allen, 1994). Surface cells in hair are abundantin fatty acids unlike the keratinized area of epidermis orsebaceous gland in hair, and about 40% of such fatty acidswere composed of 18-methyl-eicosanoic acid (18-MEA)and known to be bound to proteins by ester or thioesterbond in living or keratinized cells (Wertz and Downing, 1988,
1989). The synthesis of 18-MEA requires the oxidative dec-arboxylation of isoleucine. Maple syrup urine disease(MSUD) is caused by an inherited deficiency in the enzymebranched chain 2-oxo acid dehydrogenase, which leads tothe accumulation of branched chain a-keto-acids derivedfrom the amino acids, leucine, isoleucine, and valine. Trans-mission electron microscopy studies of developing hair fib-ers show a structural defect in the fiber shaft in hair frompatients with MSUD. This defect is confined to the cuticle ofthe fiber, where the cuticle membrane directly apposes theintercellular material. Thus, the defect indicates that 18-MEA is located exclusively on the upper surface of fibercuticle cells (Jones et al, 1996).
Results
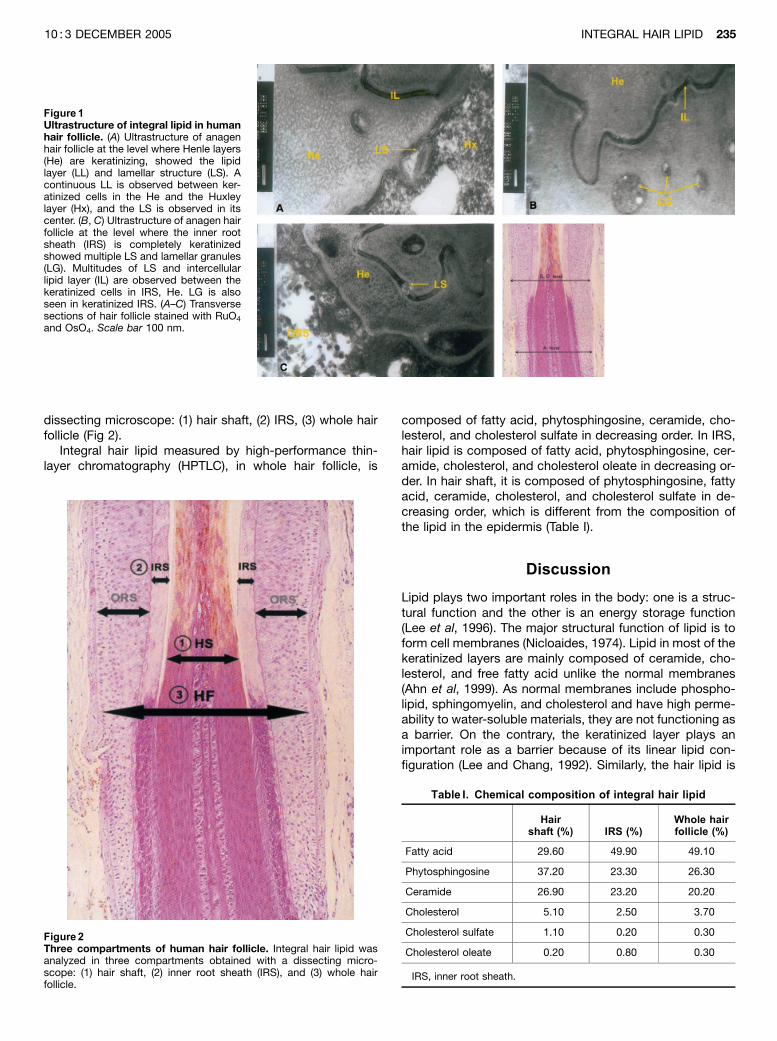
From electron microscopy of anagen hair follicle, it wasdiscovered that the lipid in hair follicle was mainly dis-tributed in hair cuticle and keratinized inner root sheath(IRS) Electron microscopy showed the presence of a lipidmultilayer, similar to the keratinized epidermis in the marginof keratinized IRS. A continuous LL is observed in IRS andthe lamellar structure (LS) is observed in its center (Fig 1A).A multitude of lamellar granules (LG) and LS are observed inthe vicinity of the above lipid multilayer (Figs 1B,C). Similarto LG in the keratinized epidermis, some of the LG in ker-atinized IRS were connected with a lipid multilayer.
Quantitative analysis of intrinsic hair lipid Integral hairlipid was analyzed in three compartments obtained with a
Abbreviations: HPTLC, high-performance thin-layer chromatogra-phy; IL, intercellular lipid; IRS, inner root sheath; LG, lamellar gran-ule; LL, lipid layer; LS, lamellar structure; ORS, outer root sheath
Copyright r 2005 by The Society for Investigative Dermatology, Inc.
234
dissecting microscope: (1) hair shaft, (2) IRS, (3) whole hairfollicle (Fig 2).
Integral hair lipid measured by high-performance thin-layer chromatography (HPTLC), in whole hair follicle, is
composed of fatty acid, phytosphingosine, ceramide, cho-lesterol, and cholesterol sulfate in decreasing order. In IRS,hair lipid is composed of fatty acid, phytosphingosine, cer-amide, cholesterol, and cholesterol oleate in decreasing or-der. In hair shaft, it is composed of phytosphingosine, fattyacid, ceramide, cholesterol, and cholesterol sulfate in de-creasing order, which is different from the composition ofthe lipid in the epidermis (Table I).
Discussion
Lipid plays two important roles in the body: one is a struc-tural function and the other is an energy storage function(Lee et al, 1996). The major structural function of lipid is toform cell membranes (Nicloaides, 1974). Lipid in most of thekeratinized layers are mainly composed of ceramide, cho-lesterol, and free fatty acid unlike the normal membranes(Ahn et al, 1999). As normal membranes include phospho-lipid, sphingomyelin, and cholesterol and have high perme-ability to water-soluble materials, they are not functioning asa barrier. On the contrary, the keratinized layer plays animportant role as a barrier because of its linear lipid con-figuration (Lee and Chang, 1992). Similarly, the hair lipid is
Figure 1Ultrastructure of integral lipid in humanhair follicle. (A) Ultrastructure of anagenhair follicle at the level where Henle layers(He) are keratinizing, showed the lipidlayer (LL) and lamellar structure (LS). Acontinuous LL is observed between ker-atinized cells in the He and the Huxleylayer (Hx), and the LS is observed in itscenter. (B, C) Ultrastructure of anagen hairfollicle at the level where the inner rootsheath (IRS) is completely keratinizedshowed multiple LS and lamellar granules(LG). Multitudes of LS and intercellularlipid layer (IL) are observed between thekeratinized cells in IRS, He. LG is alsoseen in keratinized IRS. (A–C) Transversesections of hair follicle stained with RuO4
and OsO4. Scale bar 100 nm.
Figure 2Three compartments of human hair follicle. Integral hair lipid wasanalyzed in three compartments obtained with a dissecting micro-scope: (1) hair shaft, (2) inner root sheath (IRS), and (3) whole hairfollicle.
Table I. Chemical composition of integral hair lipid
Hairshaft (%) IRS (%)
Whole hairfollicle (%)
Fatty acid 29.60 49.90 49.10
Phytosphingosine 37.20 23.30 26.30
Ceramide 26.90 23.20 20.20
Cholesterol 5.10 2.50 3.70
Cholesterol sulfate 1.10 0.20 0.30
Cholesterol oleate 0.20 0.80 0.30
IRS, inner root sheath.
INTEGRAL HAIR LIPID 23510 : 3 DECEMBER 2005

mainly composed of fatty acid, cholesterol sulfate, cer-amide, and cholestrol. It is the only continuous structureamong the total components of hair (Wertz and Downing,1989). So, we termed the above LL as the so-called hairbarrier, in that it may be the counterpart for skin barrier. TheLL in hair plays a key role in the maintenance of hair integ-rity, including hydrophobicity and stiffness (Jones and Ri-vett, 1997). Orwin (1979) reported that a globular particleexists in intercellular space during the development of haircuticles and moves to an external area, forming the con-tinuation of the LS. The LS outside the hair cuticle is formedwith the rupture of membrane outside the hair cuticle, andpaired new LS, assumed to be a LL, is formed to fill thespace outside the hair cuticle and between IRS cuticles(Jones et al, 1994). When IRS cuticle and hair cuticle sep-arate from each other, the LS on one side covers the outersurface of the hair cuticle and that on the other side is po-sitioned on the surface of IRS cuticle. There have beenstudies on the LL between hair cuticles in hair; it has beenknown to be present as a cell membrane complex com-posed of a d layer, core structure relatively light stained,covered with two b layers in sandwich form (Jones and Ri-vett, 1997). In addition, lateral force microscope and forcemodulation atomic force microscope (AFM) have revealed auniform layer on the surface of the newly formed cuticlesurface that is not readily apparent from topographic AFMimaging. This layer is of low friction and is almost certainlythat of the upper b layer, a covalently linked layer of 18-MEAto a proteinacous support, most probably that of the ep-icuticle (Smith and Swift, 2002). But there have been noreports on lipid distribution in hair follicle. We conductedelectron microscopic observation on hair follicle. It is diffi-cult to observe the LL with the conventional staining methodusing OsO4 or RuO4. OsO4 cannot reveal lipid component intissue. On the other hand, RuO4, which is routinely used intorevealing epidermal lipids cause severe hair damage. So,we prepared a new fixative (Lee’s fixative) to minimize hairinjury and observe the LL in the hair follicle. We observedthe lipid multilayer, and structures similar to LG in the vi-cinity of the lipid multilayer in keratinized cells. We found outthat the LL was mainly positioned in the hair cuticle andkeratinized IRS. The LL in the hair follicle is assumed to besimilar to the epidermal LL functioning as a skin barrier. Inquantitative analysis of intrinsic hair lipid, the report of rel-atively high amounts of phytosphingosine was remarkable,although we do not know its significance. The othersphingosines were negligible. This study is considered toprovide baseline data for the studies on lipid distribution inthe hair follicle, and more studies will be required to exam-ine the chemical component and the function of the LL inthe hair follicle.
Materials and Methods
Electron microscopy of normal scalp hair follicle Small piecesof a total of five normal adults’ scalp tissue from occipital regionwere used in this study after receiving informed consent. Sixty hairfollicles were obtained under the surgical microscope. The cross-section of the hair follicle was fixated in Karnovsky solution (2%glutaraldehydeþ 2% paraform) and rinsed in 0.1 M sodium ca-codylate, and post-fixated with Lee’s fixative (0.5% RuO4:2%
OsO4:0.2 M cacodylate buffer¼ 1:1:1), which was designed forminimizing hair injury and for better viewing the LL in the hair fol-licle, at room temperature for one and half hour. After that, tissuesection was dehydrated in alcohol solutions and substituted withpropylene oxide, and embedded in Epon mixture solution in thetransverse position. Embedded tissue section was prepared withultramicrotome with a thickness of 70–80 nm, and double stainedwith uranyl acetate and lead citrate. Then, the tissue section wasobserved in a transmission electron microscope (JEM-1200EDXII,80 kV: JEOL, Tokyo, Japan).
Quantitative analysis of intrinsic hair lipid The extraction andseparation of hair lipid were performed as described previously byWertz and Downing (1988). Free lipid was discarded, and co-valently bound integral hair lipids were analyzed. The extractedlipids were separated by one-dimensional HPTLC on 20 � 10 cmglass plates coated with silica gel (Silicagel 60, Merck, Darmstadt,Germany) using the ‘‘total development system’’ (Choi et al, 2002).In the ‘‘Total’’ development system, the lipids were separated bysequential development in (1) methanol:chloroform:water (19:79:2),(2) methanol:chloroform:ethylether:hexane (20:25:40:15), (3) chlo-roform:ethylether:hexane (25:50:25), (4) ethylether:hexane (50:50),and (5) hexane (100). Serial dilutions of appropriate lipid standardswere used for quantification. The quantification was performed af-ter staining (copper acetate and copper sulfate in phosphoric acid)and charring, using a photodensitometer (Videoscan: CCIR Color3CCD camera, CAMAG, Muttenz, Switzerland) with VideoscanHPTLC Evaluation Software Ver. 1.01, CAMAG). The results wereexpressed as the means of three different samples � SD (MicroCalOrigin 3.0: MicroCal Software Inc., Northampton, MA).
This study was supported by Hair Research Grant from Pacific R&DCenter, and a research fund from Drs Woo and Hann’s Skin Clinic,Seoul, Korea.
DOI: 10.1111/j.1087-0024.2005.10113.x
Manuscript received September 20, 2004; revised March 10, 2005;accepted for publication May 10, 2005
Address correspondence to: Won-Soo Lee, MD, PhD, Department ofDermatology, Wonju College of Medicine, Yonsei University, 162 Ilsan-Dong, Wonju, Kangwon-Do 220-701, Korea. Email: [email protected]
References
Ahn SK, Hwang SM, Choi EH, et al: Morphologic characteristics of skin barrier
and stratum corneum. J Skin Barrier Res 1:22–28, 1999
Allen DE: Lipids. In: Woods AE, Ellis RC (eds). Laboratory Histopathology: A
Complete Reference Section 6. 1994; p 3.1–3.31
Bertrand L, Doucet J, Simionovici A, et al: Lead-revealed lipid organization in
human hair. Biochim Biophys Acta 1620:218–224, 2003
Choi EH, Park W-S, Son E-D, et al: The effect of change in epidermal calcium
gradient on stratum corneum lipid and epidermal differentiation. In: Marks
R, Leveque J-L, Voegeli R (eds). The Essential Stratum Corneum. Lon-
don: Martin Dunitz, 2002; p 77–80
Elias PM, Cooper ER, Korc A, et al: Percutaneous transport in rotation to stratum
corneum structure and lipid composition. J Invest Dermatol 76:297–301,
1981
Jones LN, Horr TJ, Kaplin IJ: Formation of surface membranes in developing
mammalian hair fibres. Micron 25:589–595, 1994
Jones LN, Peet DJ, Danks DM, et al: Hairs from patients with maple syrup urine
disease show a structural defect in the fiber cuticle. J Invest Dermatol
106:461–464, 1996
Jones LN, Rivett DE: The role of 18-methyleicosanoic acid in the structure and
formation of mammalian hair fibers. Micron 28:469–485, 1997
Lampe MA, Burlingame AL, Whitney JA, et al: Human stratum corneum lipids:
Characterization and regional variations. J Lipid Res 24:120–130, 1983
Lee SH, Chang SN: Epidermal lipid and skin barrier. J Aerospace Environ Med
2:15–24, 1992
236 LEE ET AL JID SYMPOSIUM PROCEEDINGS

Lee SH, Koo SW, Hwang MS, et al: The rate of recovery after disruption of the
skin barrier and the composition of human surface lipids at different an-
atomic sites of skin. Korean J Dermatol 34:38–45, 1996
Nicloaides N: Skin lipids: Their biochemical uniqueness. Science 186:19–26,
1974
Nicloaides N, Fu HC, Rice GR: The skin surface lipids of man compared with
those of eighteen species of animals. J Invest Dermatol 51:83–89, 1968
Orwin DF: The cytology and cytochemistry of the wool follicle. Int Rev Cytol
60:331–374, 1979
Smith JR, Swift JA: Lamellar subcomponents of the cuticular cell membrane
complex of mammalian keratin fibres show friction and hardness contrast
by AFM. J Microscopy 206:182–193, 2002
Wertz PW: Integral lipids of hair and stratum corneum. In: Formation and
structure of human hair. Basel: Brikhauser, 1997; p 227–237
Wertz PW, Downing DT: Integral lipids of human hair. Lipids 23:878–881, 1988
Wertz PW, Downing DT: Integral lipids of mammalian hair. Comp Biochem Physiol
92:759–761, 1989
INTEGRAL HAIR LIPID 23710 : 3 DECEMBER 2005