information to users broken or indistinct print,...
TRANSCRIPT

The effects of water stress on gas-exchange in Pinus brutia, var eldarica
Item Type text; Thesis-Reproduction (electronic)
Authors Soumana, Diallo Amadou, 1957-
Publisher The University of Arizona.
Rights Copyright © is held by the author. Digital access to this materialis made possible by the University Libraries, University of Arizona.Further transmission, reproduction or presentation (such aspublic display or performance) of protected items is prohibitedexcept with permission of the author.
Download date 06/07/2018 12:56:21
Link to Item http://hdl.handle.net/10150/277326

INFORMATION TO USERS
The most advanced technology has been used to photograph and
reproduce this manuscript from the microfilm master. UMI films the
text directly from the original or copy submitted. Thus, some thesis and
dissertation copies are in typewriter face, while others may be from any
type of computer printer.
The quality of this reproduction is dependent upon the quality of the copy submitted. Broken or indistinct print, colored or poor quality
illustrations and photographs, print bleedthrough, substandard margins,
and improper alignment can adversely affect reproduction.
In the unlikely event that the author did not send UMI a complete
manuscript and there are missing pages, these will be noted. Also, if
unauthorized copyright material had to be removed, a note will indicate
the deletion.
Oversize materials (e.g., maps, drawings, charts) are reproduced by
sectioning the original, beginning at the upper left-hand corner and
continuing from left to right in equal sections with small overlaps. Each
original is also photographed in one exposure and is included in
reduced form at the back of the book.
Photographs included in the original manuscript have been reproduced
xerographically in this copy. Higher quality 6" x 9" black and white
photographic prints are available for any photographs or illustrations
appearing in this copy for an additional charge. Contact UMI directly to order.
University Microfilms International A Bell & Howell Information Company
300 North Zeeb Road, Ann Arbor, Ml 48106-1346 USA 313/761-4700 800/521-0600


Order Number 1341237
The effects of water stress on gas-exchange in Pinus brutia, var. eldarica
Soumana, Diallo Amadou, M.S.
The University of Arizona, 1990
U MI 300 N. Zeeb R& Ann Arbor, MI 48106


THE EFFECTS OF WATER STRESS ON GAS-EXCHANGE
IN PINUS BRUTIA. VAR. ELDARICA
by
Diallo Amadou Soumana
A Thesis Submitted to the Faculty of the
THE DEPARTMENT OF SOIL AND WATER SCIENCE
In Partial Fulfillment of the Requirements For the Degree of
MASTER OF SCIENCE
In the Graduate College
THE UNIVERSITY OF ARIZONA
19 9 0

STATEMENT BY AUTHOR
2
This thesis has been submitted in partial fulfillment of requirements for an advanced degree at the University of Arizona and is deposited in the University Library to be made available to borrowers under rules of the Library.
Brief quotations from this thesis are allowable without special permission, provided that accurate acknowledgement of source is made. Requests for permission for extended quotation from or reproduction of this manuscript in whole or in part may be granted by the head of the major department or the Dean of the Graduate College when in his or her judgement the proposed use of the material is in the interests of scholarship. In all other instances, however, permission must be obtained from the author.
SIGNED: SoonftMlV
APPROVAL BY THESIS DIRECTOR
This thesis has been approved on the date shown below:
Qu*~ t \ . -71/ln-nMAa f o i l , g . mo Allan D. Matthias (Pat© Associate Professor of Soil and Water Science

3
ACKNOWLEDGEMENTS
The author wishes to specially thank Dr. Allan Matthias,
his thesis Advisor and Major Professor, for his continuous
advice, helpful suggestions and patience throughout the entire
study. I am very grateful to Dr. J. W. Moon and Dr. J. R.
Simpson for their technical contribution and assistance in the
study. I also thank Dr. Richard Jensen for the use of his
laboratory and instrumentation for the gas-exchange
measurements.
Finally, special gratitude is extended to my parents in
Niger for their patience, to my sponsor in Washington D.C. and
Carol Davis at the International Agricultural Program,
University of Arizona, for their encouragement and support,
and to all Americans.

4
TABLE OF CONTENTS
Page
LIST OF ILLUSTRATIONS 5
LIST OF TABLES 6
ABSTRACT 7
CHAPTER 1, INTRODUCTION 8
CHAPTER 2, LITERATURE REVIEW 11
CHAPTER 3, MATERIALS AND METHODS 13
Experimental Units, Treatments, and Design 13
Environmental Monitoring within Shade House .... 16
Gas-Exchange Measurement Procedures 17
Computational and Data Analysis Procedures 20
CHAPTER 4, RESULTS AND DISCUSSION 22
CHAPTER 5, SUMMARY AND CONCLUSION 29
APPENDIX A: Air Temperature within Shade House .... 31
APPENDIX B: Solar Radiation within Shade House .... 32
APPENDIX C: Photosynthetic Parameters for Four Conifer Species 33
REFERENCES 34

5
LIST OF ILLUSTRATIONS
Figure Page
1 Schematic diagram of open-chamber system used for gas-exchange measurements 18
2 C02 assimilation rate (A) versus intercellular C02 mole fraction (C,) for non-stressed (control) plants, stressed plants (medium ^»L < -2.0 MPa), and plants irrigated 7 days following 2 to 3 days medium stress 23

6
LIST OF TABLES
Table Page
1 Description of control and treatment designations 14
2 Gas-exchange parameter mean values measured for potted Pinus brutia plants under well watered (control), medium water stress, and water stress + 7 days recovery conditions .... 24

7
ABSTRACT
Pinus brutia var. eldarica is often considered to possess
photosynthetic characteristics that make it highly tolerant to
drought conditions. However, very little is known about it»s
photosynthetic response to water stress under either
laboratory or field imposed drought conditions. The purpose
of this study was to utilize laboratory gas-exchange
measurements to determine the effects of water stress and
recovery from stress on photosynthetic capacity of potted
Pinus brutia var. eldarica plants. Analysis of the rate
assimilation (A) versus intercellular C02 (C^) data indicates
that recovery of carboxylation efficiency (g'J (i.e. Rubisco
activity and amount) does occur after daily re-watering of
moderately stressed (^2 - -2.0 MPa) Pinus brutia. However,
these same data indicate that irreversible damage to cell
membranes results in an inability to re-generate RubP.
Although inferences on the effect of stress on A in field
grown Pinus brutia is tenuous, these laboratory results
indicate that Pinus brutia may not be as drought tolerant as
commonly believed.

8
CHAPTER 1
INTRODUCTION
Water stress due to drought is often the single most
important environmental factor affecting the survival and
growth of a plant species (Kramer,1983). Newly planted tree
seedlings are particularly sensitive to water stress because
their poorly developed root systems (Brix, 1979) are often
inadequate for preventing severe plant water deficits (Davies
et al.,1973; Kozlowski,1975) during drought. Tolerance of
seedlings to water stress is, therefore, an important factor
affecting their survival and growth.
Plant water stress can cause large reductions in
photosynthetic activity. This response is mainly the result
of both increasing resistance to C02 diffusion and a reduction
in the efficiency of the photosynthetic mechanism (Boyer,
1976; Hinckley et al., 1981). Plants previously exposed to
water stress continue photosynthesis at lower water potentials
than do plants which have never experienced any water stress
(Ashton, 1956; Ackerson and Hebert, 1981; Matthew and Boyer,
1984) .
Attempts at introducing new tree species, such as Pinus
brutia, to an arid or semi-arid region often meet with
failure, because planted tree seedlings are frequently unable
to survive the stress imposed by drought conditions within the

9
region. The effects on photosynthesis due to prolonged periods
of water stress through lack of rain and/or irrigation is a
primary cause of failure to establish tree seedlings.
Pinus brutia is a species of pine commonly believed to
have characteristics favorable for growth in arid and semi-
arid regions, such as the southwestern United States. It is
relatively unique among species of pine in that it is readily
adaptable to growth in soils with high pH (Abido, 1986). It
is also commonly believed to be tolerant to drought
conditions, however, the response of the photosynthetic
capacity of Pinus brutia to water stress has not been studied
either in the laboratory (potted plants) or under field
conditions.
Techniques of gas-exchange have made possible the
detailed investigation of the effects of water stress on
photosynthesis in Pinus brutia. For almost all other plant
species previously studied, it is well documented that severe
water stress causes a decrease in net photosynthesis (Brix,
1969). Furthermore, there is evidence that the capacity of
the mesophyll to assimilate carbon is sometimes affected by
water stress, as is also stomatal conductance (Boyer, 1976).
Gas-exchange measurements have also shown a reduction in
quantum yield and in light-saturated rates of C02 assimilation
(Boyer, 1971; Mooney et al., 1977).

10
The effects of water stress on the so-called "mesophyll
conductance" (e.g. initial slopes ofA/C,. curve) of carbon have
been examined by several authors, but the results remain
equivocal (Troughton and Slatyer, 1969; Laisk and Oya, 1971;
Jones, 1973; Moldau, 1973; Coolatz, 1977; O'Toole et al.,
1976; Mooney et al.,1977; Radin and Ackerson, 1981). The
results are equivocal in respect to many different methods of
imposing water stress that have been used, thus, making
interspecies comparisons difficult.
Measurements of the rate of C02 assimilation (A) to
intercellular C02 mole fraction (C{) response functions for
several pine tree species (e.g. Pinus silvestris. Pinus
contorta. and Pinus radiatal have shown that rapidly induced,
short-term water stress leads to reductions in assimilation in
plants grown under high C)2 (C, 660 /xmol mol"1) but not in
plants grown under ambient C02 (C{ - 330 /mol mol'1) (Havrane
and Brenecke, 1978).
Although studies of photosynthesis have been made for
many species of pine (see Brix, 1969), there is, however,
little information available at present on photosynthesis in
Pinus brutia (Havrane and Benecke, 1978). The overall
objective of this study was, therefore, to determine the
effects of water stress, and recovery from water stress on
photosynthetic capacity in potted Pinus brutia var. eldarica
under laboratory conditions.

11
CHAPTER 2
LITERATURE REVIEW
The basic physiological parameters describing
photosynthesis are obtained from the response of the rate of
C02 assimilation (A) to the intercellular C02 concentration
(C{). Seedling physiological response to water stress has
been measured by leaf moisture content, transpiration,
diffusive resistance, photosynthesis and respiration. The
effect of moisture stress on the physiology of the needle has
generally been observed to decrease transpiration, close
stomates, and reduce photosynthesis. In addition, Abido
(1986) used electron microscopy to determine that chloroplasts
of mesophyll cells Pinus brutia were markedly disrupted when
relative water content decreased to 62 percent during water
stress conditions.
Second year loblolly pine seedlings were found to
decrease carbon dioxide uptake with increasing diffusion
pressure deficit (DPD) of the needles and photosynthesis was
found to be highly positively correlated with transpiration
rate (Brix, 1962). Bilan et al. (1977) and Millan and Allen
(1971) showed that stomatal aperture and mesophyll resistance
could both play a part in photosynthesis reductions. Most
frequently cited as the cause of the decline in photosynthesis
as a result of water stress, several authors have observed a

12
major effect of non stomatal factors on photosynthetic
inhibition (Boyer and Bowen, 1970; Cornic et al. 1983;
Hutmatcher and Krieg, 1983).
A photosynthesis response to lower water potentials would
convey an advantage to plants during periods of drought.
Photosynthesis and growth in such plants could continue longer
during the period of water deficit. This response, however,
has been examined in only a limited number of species and has
not been investigated in Pinus brutia var. eldarica.

13
CHAPTER 3
MATERIALS AND METHODS
Experimental Units. Treatments, and Design
Ninety potted Pinus brutia var. Eldarica seedlings formed
the experimental units of this study. The one-year old trees
were approximately 0.3 to 0.5 m tall at the time of the study,
which was done during April, May, and June 1989. Prior to the
study, the trees were transplanted to 4-L plastics pots in a
1 soil: 2 sphagnum peat: 2 perlite (by volume) medium with pH
of 6.2. The medium was amended with 0.744 kg of treble
superphosphate, 0.946 kg of potassium nitrate, 0.496 kg
magnesium sulfate, 4 kg ground calcite limestone, and 0.062 kg
Peters Frit Industries Trace Elements No. 555 (Peters
Fertilizers Products, Fogesville, Pa.) per cubic meter. During
plant establishment, the potted trees were fertilized at each
watering with 200 mg L"1 each of N and K supplied from 517 mg
L"1 KN03 and 367 mg L'1 NH4N03. The fertilizer solution was
maintained at pH 6.0 by injecting 75% (w/w) technical grade
phosphoric acid into the irrigation system.
On 1 April 1989 the 90 trees were placed at random within
an array of 18 rows (5 plants per row with 0.25 m spacing) on
a bench in a 50% shade house at the University of Arizona
Campus Agricultural Center in Tucson. Note that the trees
were acclimated to the reduced light intensity within the

14
Table 1. Description of control and treatment designations
Designation Description
Control
Low i|rL
Medium i|tL
Plants irrigated daily in shade house. VL did not become less than about - 0.5 MPa as determined at time of gas-exchange measurements.
Gas-exchange measurements made at end of a 3 to 5 day interval during which plants did not receive any irrigation.
- - 4.0 MPa at time of measurements.
Gas-exchange measurements made at end of a 2 to 3 day interval during which plants did not receive any irrigation. i|»L - - 1.0 to -1.5 MPa at time of measurements.
Medium \|»L + Recovery Gas-exchange measurements made at end of 7 day recovery period with daily irrigations following a 2 to 3 day interval with no irrigations. -- 2.0 MPa at time of measurements.
Low i(fL + Recovery Gas-exchange measurements made at end of 7 day recovery period with daily irrigations following a 3 to 5 day interval with no irrigations. i|»L -- 3.0 MPa at time of measurements.

15
shade house for a period of 6 weeks prior to the establishment
of water stress treatments in April 1989. Establishment of
treatments commenced on 2 April. The treatments were
designated (see Table 1) as high stress, medium stress, and
control. Also, recovery of the plants over about a seven day
period from the stress conditions was studied. Both the high
and medium stress treatments involved withholding irrigation
for several days. The high stress plants did not receive
irrigation water for intervals of 3 to 5 days, resulting in
needle water potentials that decreased to about -4.0 MPa. The
medium stress plants did not receive irrigation water for
intervals of 2 to 3 days, resulting in needle water potentials
of about -1.5 MPa. The control treatment involved daily
irrigation with tap water which maintained needle water
potentials at about -0.5 MPa. During selected periods of the
study, several of the high stress and medium stress plants
were irrigated daily for about 7 days following a stress cycle
in order to permit investigation of the recovery of the
photosynthetic process following stress conditions. Needle
water potentials (i|rL) at the end of the 7 day recovery period
were about -2.0 MPa and -3.0 MPa for the medium and high
stress plants, respectively. Needle water potentials were
measured with a Soil-Moisture Equipment Model 1000 pressure
chamber immediately before and after the gas-exchange
measurements. The needle water potential measurements were

16
facilitated by removal of three to four 0.05 to 0.1 m long
branches from a plant under study. Water potential values
reported here are averages of readings taken on all plants
within a treatment.
Environmental Monitoring within Shade House
Air temperature (at 1.5 m height above ground) and solar
radiation conditions within the shade house were continuously
monitored between 3 April 1989 and 13 June 1989 with a
radiation-shielded thermistor (Campell Scientific Inc.(CSI;
Logan, Utah) model 107) and a Licor (Lincoln, Nebraska) LI
200S pyranometer, respectively. Output signals from both
sensors were sampled at 10 second intervals with a CSI 2IX
Micrologger and averaged over a 60 minute period to yield mean
air temperature (° C) and solar radiation flux density (MJ m'2
h"1). All data were processed by the Micrologger and were
stored on cassette tape for later analysis. As illustrated in
Appendix A, air temperatures were relatively invariant from
day-to-day throughout the study, with the exception of a brief
cooling trend during mid-April. Typically, daily maximum air
temperatures within the shade house were about 35 to 40 ° C,
and minimum temperatures were about 10 to 15 ° C. Incoming
solar radiation within the shade house was also relatively
invariant from day-to-day during the study, with total flux
densities typically in the range of 6 to 9 MJ m"2 d'1. Data

I
17
presented in Appendix B shows that the diurnal variations in
solar radiation flux density were similar from day-to-day.
The data presented in Appendices A and B, therefore, show
that the environmental conditions within the shade house were
relatively constant on a day-to-day basis. Thus, there
should not have been any marked confounding effect of changing
environmental conditions on the water stress treatments
imposed during the study. The data also show that the
relatively warm and sunny conditions were favorable for rapid
imposition of water stress in the potted plants studied.
Gas-Exchanae Measurement Procedures
Gas-exchange response to increasing C02 was measured in
a laboratory on the University of Arizona campus on well
watered (control) plants, water stressed plants, and water
stressed + recovery plants at intervals throughout the study.
At the completion of each stress cycle and each recovery
cycle, gas-exchange responses of approximately 4 stressed
plants and 4 control plants were measured.
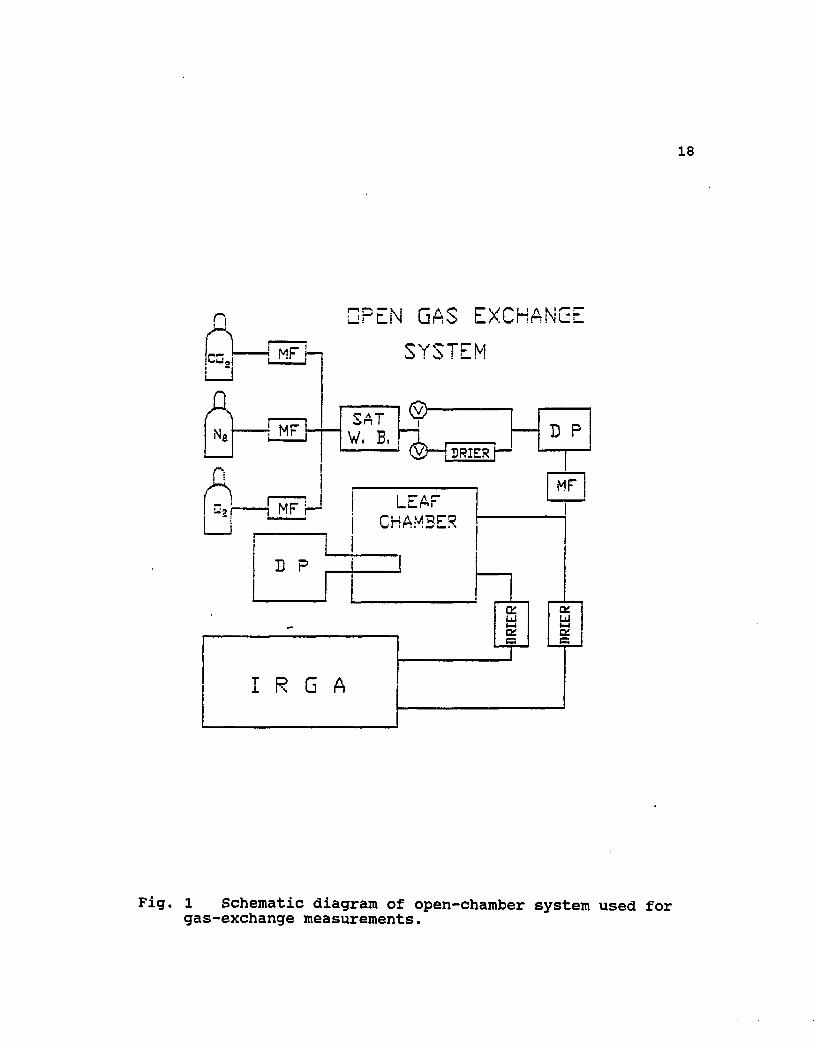
18
A raj [mf>
-TMF
SPEN GAS EXCHANG!
SYSTEM
SAT V, B,
0"
(v)—! 3RIIR
LEAF CHAMBER
I R G A
D P
MF
zc LLI
0 £ zc LLI U I—i t-l Q£ ae. CS c
Fig. 1 Schematic diagram of open-chamber system used for gas-exchange measurements.

19
Gas-exchange measurements were made using an open system
and temperature controlled glass cuvette (see Fig. 1, leaf
chamber) as previously described by Perchorowicz et al.
(1981). Oxygen, nitrogen, and C02 were mixed in a manifold
using a multiple valve flow control system. The flow rates
into the mixing chamber were measured with mass flow meters
(Hastings Model ST 2263 and ST 2664). Response of C02
assimilation (A) to increasing C02 was monitored by measuring
C02 gas-exchange in step increments from low (<100 /xmol mol"1)
to high C02 (1000 nmol mol"1) mole fractions. The reference,
sample and the C02 differential concentrations between the
inlet and the outlet gas from the cuvette were measured with
an infrared gas analyzer (IRGA in Fig.l) (ADC Model LCA-2).
Flow rate into the glass cuvette was controlled at 3 L min"1
and the air temperature inside the water-cooled cuvette was
controlled throughout at about 25 ± 0.5 t. Air and needle
temperatures were monitored using thin-wire thermocouples.
Vapor pressure deficit (VPD) was maintained at 1.5 to 2.0 kPa
by saturating the gas stream in a temperature controlled water
bath, and then drying part of the air stream with silica gel.
Vapor pressure of the inlet and outlet gas was measured with
dew-point hygrometers (EG&G Model 911). Light supplied by a
1000 W metal halide lamp was filtered through 0.08 m of water
and neutral density screens. The irradiance level was measured
with a quantum sensor (Licor Model LI-180) positioned between

20
the screen and cuvette, and was maintained at a PPF between
1400 and 1600 jumol s"1 m'2. Assimilation rate (A) was
expressed on a per unit needle area basis. Total needle area
within the cuvette was determined by removing all needles from
the plant under study upon completion of gas-exchange
measurements. The needles were then dried at about 60°C
overnight. A regression equation developed for area versus
needle dry weight was then used to calculate total needle area
for each plant studied.
Computational and Data Analysis Procedures
Gas-exchange parameter values were calculated using
standard equations given by Moon and Flore (1986). Non-linear
regression equations were fitted to the calculated C02
assimilation (A) versus intercellular C02 concentration (cf)
data for use in determining A at light and C02 saturated
conditions (A^) and carboxylation efficiency (g'm). A^ for
each fitted curve was estimated as the value of A
corresponding to the "plateau" region of the curve where the
slope (dA/dCj) approaches zero. Carboxylation efficiency
(g'm) was determined from the slope of the linear portion of
the A/Cj curve.
Statistical analyses were made using Dunnett's one-sided
t-test (Steele and Torrie, 1960) to determine whether or not
photosynthetic parameter means for the stressed plants were

21
significantly different from the mean values determined for
the non-stressed control plants.
The relative stomatal limitation (I, expressed in units
of percent) to photosynthesis is the proportionate decrease in
C02 assimilation (A) that may be attributed to the stomata and
other gas phase limitations. This parameter is, thus, useful
for separating stomatal from mesophyll limitations to
photosynthesis (Long, 1985). Relative stomatal limitation is
calculated from Eq. 1 (Farquhar and Sharkey, 1982) given as:
1= A" A°ioo (1) A
where the rate of C02 assimilation (A) is calculated at the
normal atmospheric C02 mole fraction of about 350 /xmol mol"1,
and A0 is the rate which would occur if there was no stomatal
limitation. A0 is the value of A interpolated from the A
versus C, curve at C{ - 350 jtmol mol'1.

22
CHAPTER 4
RESULTS AND DISCUSSION
The characteristic changes in photosynthetic rate, (A)
with intercellular C02 concentration (Cf), are shown in Fig.
2 for the control plants, the medium stress plants, and the
medium stress + recovery plants.
Figure 2 shows that for the control plants the increase
in A with C. was linear over the low C02 mole fraction range.
At about 400 nmol mol"1 C02, the slope of the curve gradually
decreased. At this C02 mole fraction, referred to by Farquhar
and Sharkey (1982) as the inflection point, the rate of
assimilation (A) increases more slowly with increased c{. The
photosynthetic rate for the control plants reached a maximum
value (A^) of 27.7 fimol (C02) m"2 s"1 at a mole fraction of
about 650 jumol mol*1. Figure 2 also shows that medium water
stress (i.e. i|fL <-2.0 MPa) in Pinus brutia markedly decreased
C02 assimilation rates. In addition, the slope (g*m) in the
linear portion of the A/C. curve for the medium stressed
treatment is considerably lower than the slope for the control
plants. In contrast to the low g'm for the medium stressed
plants, the g'm for the recovery plants was nearly identical
to that for the control plants. However, the Amax values for
both stressed treatments shown in Fig. 1 were considerably
below the A(I)0X for the control plants.

23
</> 30-N I E
(N ° 20-^ O ~
O E 3. 1 0 -
• CONTROL ^ < -0.5 MPa
© STRESS + 7 DAYS RECOVERY
• STRESS < -2.0 MPa
C. /zmol mol"1
800
Fig. 2. CO, assimilation rate (A) versus intercellular C02 mole fraction (CJ for non-stressed (control) plants, stressed plants (medium ijr^ < -2.0 MPa), and plants irrigated 7 days following 2 to 3 days medium stress.
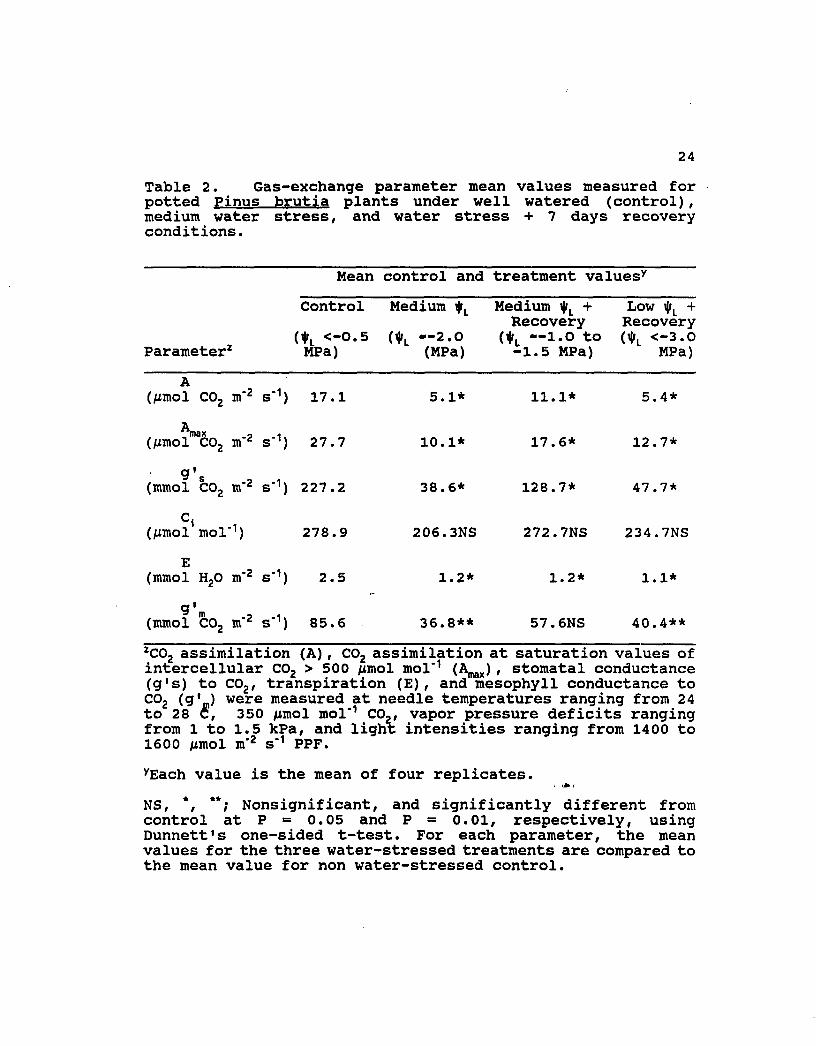
24
Table 2. Gas-exchange parameter mean values measured for potted Pinus brutia plants under well watered (control), medium water stress, and water stress + 7 days recovery conditions.
Mean control and treatment valuesy
Parameter z
Control
(*L <-0.5 MPa)
Medium
(1»L —2.0 (MPa)
Medium i|rL + Recovery
(^r, —1.0 to -1.5 MPa)
LOW i|»L + Recovery (l|lL <-3.0
MPa)
A (Mmol C02 m"2 s" 1) 17.1 5.1* 11.1* 5.4*
Anax (/mol co2 m"2 s' 1) 27.7 10.1* 17.6* 12.7*
g's (mmol C02 m"2 s" 1) 227.2 38.6* 128.7* 47.7*
ci (jumol mol • 1 ) 278.9 206.3NS 272.7NS 234.7NS
E (mmol H20 m"2 s" 1) 2.5 1.2* 1.2* 1.1*
g'm (mmol C02 m'2 s" 1) 85.6 36.8** 57.6NS 40.4**
ZC02 assimilation (A), C02 assimilation at saturation values of intercellular C02 > 500 /xmol mol"1 (Amax), stomatal conductance (g's) to C02, transpiration (E), and mesophyll conductance to C02 (g'_) were measured at needle temperatures ranging from 24 to 28 C, 350 /umol mol*1 CO,, vapor pressure deficits ranging from 1 to 1.5 kPa, and light intensities ranging from 1400 to 1600 /mol m"2 s"1 PPF.
yEach value is the mean of four replicates.
NS, *, **; Nonsignificant, and significantly different from control at P = 0.05 and P = 0.01, respectively, using Dunnett's one-sided t-test. For each parameter, the mean values for the three water-stressed treatments are compared to the mean value for non water-stressed control.

I
25
Photosynthetic and transpiratory parameter values
determined from gas-exchange measurements and from the A
versus C{ curves (Fig. 2) are summarized in Table 2. Note
that Table 2 does include values for the low i|rL + recovery
treatment. It does not, however, contain values for the low
t|il (see Table 1 for description of this treatment) because the
A response to C, for this treatment was virtually nonexistent
due to the severity of the 3- to 5-day water stress condition.
As shown in Table 2, there was measurable recovery in the A
versus C{ response in the highly stressed plants following 7
days watering.
Results summarized in Fig. 2 and in Table 2 give an
indication that Pinus brutia var. eldarica may not be as
drought tolerant as commonly believed. Analysis of the rate
of assimilation (A) versus intercellular C02 mole fraction (C.)
data indicates that recovery of the efficiency of
carboxylation (g'm) (i»e., ribulose l,5-biphosphate
carboxylase/oxygenase (Rubisco) activity and amount) does
occur after daily re-watering of moderately (but not highly)
stressed Pinus brutia plants. This is brought out clearly in
Table 2 which shows that the g• m value (57.6) for the medium
stress + recovery plants is not significantly different from
the g'm value (85.6) for the control plants. This indicates
that the Rubisco proteins in the medium stressed plants did
regenerate during the re-watering phase. However, these same

26
data (in Table 2) indicate that irreversible damage to cell
membranes may have resulted in an inability to re-generate
ribulose 1,5-biphosphate (RubP) by photosynthetic electron
transport. This is seen by the fact that the A^ value (17.6)
for the medium stress + recovery treatment was significantly
different from the A^ value (27.7) for the control plants.
Table 2 shows that all stress levels did significantly
reduce the transpiratory parameters (i.e. transpiration, E;
and stomatal conductance to water vapor, g's) relative to the
control. It is noteworthy that reduction in g*s due to water
stress did not, on the basis of C,- data, impose a severe
stomatal limitation to C02 diffusion to the intercellular
spaces. This is seen by the fact that the water stress
treatment C{ values in Table 2 were slightly lower but not
significantly different from the mean value (278.9) for the
non-stressed control. The measured slight decrease in C{ with
decreasing \|iL in Pinus brutia is, interestingly, opposite to
the increase in C. with decreased i|»L measured by Farquhar and
Sharkey (1982) in Encilia farinosa.
In contrast to the C,. data, however, it is interesting
that evaluation of the relative stomatal limitation (I, Eq. 1)
for each of the A/C{ curves given in Fig. 2 indicates that
stomatal limitation in the C02 pathway did reduce A in the
stressed plants. Calculations with Eq. 1 show that I values
were about 22, 37, and 49 % in the control, medium stress +

27
recovery, and medium stress (no recovery) plants,
respectively. The change from 22 to 49 % represents a
proportionate increase in stomatal limitation of 123 % when i|rL
decreased from about -0.5 MPa to -2.0 MPa. In contrast, Long
(1985) reported that for Helianthus annuus a lowering of i|iL
from -0.33 MPa to -1.63 MPa increased I from 0.27 to 0.31,
which was a proportionate increase of only 13 %. Stomatal
limitation data analysis for other species of pine have not,
to my knowledge, been reported in the literature.
Appendix C presents a tabulation of A, g's, and Cj values
taken from the literature (Wong et al., 1979, Brix, 1979) for
Pinus species: radiata, contorta, and silvestris. It also
presents the values (taken from Table 2) for non-water-
stressed brutia determined from this study. The data were
collected under somewhat different environmental and
silvicultural conditions, thus, making direct comparisons
difficult. The data presented, however, qualitatively
indicate that A values were similar among the four species
with the exception of contorta, which reportedly has a very
low carbon dioxide assimilation rate of 6.4 /zmol C02 m"2 s"1.
Interestingly, brutia has the largest stomatal conductance
(227.2 mmol m"2 s"1) of the four species considered, which may
be an advantage for carbon assimilation under non-water-
stressed conditions, but a disadvantage for conserving water.
The stomatal conductance for brutia is also markedly higher

28
than the value of 1.5 mmol m"2 s"1 reported for one-year old
needles of Pinus resinosa (see Long, 1985) Intercellular C02
concentrations (C,) among the species listed in Appendix C are
similar. Of Pinus contorta Doual.. Pinus radiata. and Pinus
svlvestris. Pinus contorta was determined by Brix (1969) to be
the most sensitive to water stress. However, results from
this study indicate that brutia may be equally sensitive to
water stress.

CHAPTER 5
29
SUMMARY AND CONCLUSIONS
This study was done to determine the effects of water
stress and recovery from water stress on the photosynthetic
capacity of Pinus brutia var. eldarica. Measurements of gas-
exchange were made using an open chamber system in a
laboratory. Water stress imposition was done on potted, one-
year old, eldarica plants that were grown under shade house
environmental conditions. Stress treatments included
withholding irrigation of plants to allow \|rL to decrease to
values ranging from about -2 MPa (medium stress) and to about
-4 MPa (high stress). Recovery from stress conditions was
facilitated by daily irrigation of plants for a period of
about 7 days. Control plants were irrigated daily throughout
the study.
Results showed that:
1) Assimilation (A) decreased with decreasing tJil .
2) Water stress decreased transpiration (E) and stomatal
conductance (g*8).
3) Intercellular C02 mole fraction (C{) decreased
slightly, but not significantly, with increased stress.
4) Relative stomatal limitation to gas phase C02
diffusion to the site of carboxylation apparently increased
with increasing stress.

30
5) Full recovery of Rubisco enzyme activity and amount
from medium stress did occur following 7 days irrigation.
Full recovery did not occur from high stress.
6) Full regeneration of RubP did not occur following 7
days irrigation of stressed plants, thus indicating
irreversible damage to cell membranes and photosynthetic
capacity.
7) Although inferences on the effects of water stress on
photosynthesis in field grown Pinus brutia is tenuous, these
laboratory results indicate that Pinus brutia may not be as
drought tolerant as commonly believed. And that other
physiological characteristics, such as its adaptability to
growth in high pH soil, make it a suitable plant for wood and
biomass production in arid and semi-arid regions.

APPENDIX A: Air temperature within shade house
31
0.2' 0.4 0.6 o.e
3 April to 13 June (hours x 1000)
APPENDIX B: Solar radiation within shade house
32
1.2
1.1
I 0.9
0.8
0.7
0.6
0.5
0.4
0.3
0.2
0.1
0 0.2 0.4 0.6 0.8
3 April to 13 June (Hours z 1000)

33
APPENDIX C: Photosynthetic parameters for four conifer species
- Species Parameter/units radiata contorta silvestris brutia
A (/xmol C02 m"z s"1) 20.6 £74 17.0 17.1 g»s (mmol C02 m"2 s"1) 133.0 100.0 96.0 227.2
(Mmol mol'1) 291.0 284.9 289.7 278.9

34
REFERENCES
Abido, M.S. 1986. Morpho-physiological evaluations of Aleppo and Brutia pine seedlings under two different moisture regimes. Ph.D. dissertation. University of Arizona Library. 116 pages.
Ackerson, R.C. and R.R. Hebert. 1981. Osmoregulation in cotton in response to water stress. 1. alterations in photosynthesis, leaf conductance, translocation and ultrastructure. Plant Physiol. 67:484-488.
Ashton, F. M. 1956. Effects of a series of cycles of alternating low and high soil water contents on the rate of apparent photosynthesis in sugar cane. Plant Physiol. 31:266-274.
Bilan, M.V., C.T. Hogan, and H.B. Carter. 1977. Stomatal opening, transpiration,.and needle moisture in Loblolly pine seedlings from Texas seed sources. For. Sci. 23:457-462.
Boyer, J.S. 1976. Water deficits and photosynthesis. In Water Deficits and Plant Growth, vol. IV (T.T. Kozlowski, ed.) pp. 153-190. Academic Press, New York. 383 pages.
Boyer, J.S. 1971. Non stomatal inhibition of photosynthesis in sunflower at low leaf water potentials and high light intensities. Plant Physiol. 48:532-536.
Boyer, J.S., and B.L. Bowen. 1970. Inhibition of oxygen evolution in choloroplasts isolated from leaves with low water potentials. Plant Physiology 45, 612-615.
Bravdo, B. 1968. Decrease in net photosynthesis caused by respiration. Plant Physiol. 43:479-483.
Brix, H. 1962. The effect of water stress on the rates of photos^pthesis and respiration in tomato plants and loblolly pine seedlings. Physiologia 22:10.
Brix, H. 1969. Effects of plant water stress on photosynthesis and survival of four conifers. Can. J. Forest. Res. 9:160-165.
Collatz, G.J. 1977. Influence of certain environmental factors on photosynthesis and photorespiration in Simondsia chinensis. Planta 134:127-132.

35
Cornic, G.Y., L. Prioul, and G. Lovason. 1983. Stomatal and non-stomatal contribution in the decline in leaf net C02 uptake during rapid water stress. Physiol. Plant. 58:295-302.
Davies, W.J. 1973. Effects of transplanting on physiological responses and growth of shade trees, pages 22-30. Jn The 48th International Shade Tree Conf. Proc. Urbana, ill.
Farquhar, G.D., and T.D. Sharkey. 1982. Stomatal conductance and photosynthesis. Ann. Rev. Plant Physiol. 33:341-345.
Havrane, W.M., and U. Benecke. 1978. The influence of soil moisture on water potential, transpiration, and potosynthesis of conifer seedlings. Plant and Soil 49:91-103.
Hutchmatcher, R.B., and D.R. Krieg. 1983. Photosynthetic rate control in cotton. Plant Physiol. 73:658-661.
Hinckley, T.M., Duhne, A.R. Hinckley, and H. Richter. 1981. Temperate hardwood forests. In Water Deficits and Plant Growth, vol.. VI (T.T. Kozlowski, ed.) pp. 153-208. Academic Press, New York, 582 pages.
Jones, H.G. 1973. Limiting factors in photosynthesis. New Physiologist. 72:1089-1094.
Jones, H.G. 1985. Partitioning stomatal and non stomatal limitation to photosynthesis. Plant Cell Environ. 8:95-104.
Kozlowski, T.T. 1975. Effects of transplanting and site on water relations in trees. Am. Nurseryman. 141:84-94.
Kramer, P.J. 1983. Elevated atmospheric partial pressure of co2 and plant growth. I. Interaction of nitrogen nutrition and photosynthetic capacity in C3 and C4 plants. Oecologia 44, 68-74.
Laisk, A. 1977. Kinetics of photosynthesis and photorespiration of C3 plants. Namka, Moscow.
Laisk, A., and V. Oya. 1971. Change in resistance of Aspen mesophyll as a response to rapid leaf drying. Physiol. Res. 18:553-562.
Long, S.P. 1985. Leaf gas exchange. In Photosynthetic Mechanisms and the Environment. (J. Barber and N.R. Baker, ed.) pp. 453-499. Elsevier Science Publishers.

36
Matthews, M.A., and J.S. Boyer. 1984. Acclimation of photosynthesis to low leaf water potentials. Plant Physiol. 74:161-166.
Milan, A.B., and R. Allen. 1971. Changes in radio sensitivity of pine seedlings subjected to water stress during chronic gamma irradiation. Health Physics 15, 1652-1662.
Moldau, H. 1973. Effects of various water regimes on stomatal and mesophyll conductances of bean leaves. Photosynthetica. 7:1-7.
Moon, J.W., Jr., and J.A. Flore 1986. A basic computer program for calculation of photosynthesis, stomatal conductance, and related parameters in an open gas-exchange system. Photosyn. Res. 7:269-279.
Mooney, H.A., 0. Bjorkman, and G.J. Collatz. 1977. Photosythetic acclimation to temperature and water stress in the desert shrub Larrea divaricata. Year Book, Carnegie Inst., Washington, D.C. 76:328-335.
O'Toole, J.C., R.K. Crookton, K.J. Treharne, and J.L. Ozbun. 1976. Mesophyll resistance and carboxylase activity. Plant Physiol. 57:465-468.
Perchorowics, J.T., D.A. Raynes, and R.G. Jensen. 1981. Light limitation of photosynthesis and activation of ribulose bisphosphate carboxylase in wheat seedling. Proc. Natl. Acad. Sci. USA 78:2985-2989.
Plant, Z., and B. Bravdo. 1973. Response of carbon dioxide fixation to water stress. Plant Physiol. 52:28-32.
Radin, J.W., and G.C. Ackerson. 1981. Water relations of cotton plants under nitrogen deficiency. III. Stomatal conductance, photosynthesis, and abscisic acid accumulation during drought. Plant Physiol. 67:115-119.
Sharkey, T.D. 1985. Photosynthesis in intact leaves in C3 plants: physics, physiology, and rate limitations. The Botanical Rev. 51:53-105.
Steele, R.G., and J.H. Torrie. 1960. Principles and Procedures of Statistics. New-York. 480 pages.
Troughton, J.H., and R.O. Slatyer. 1969. Plant water status and carbon dioxide exchange in cotton leaves. Aust. J. Biol. Sci. 22:289-302.

37
Wong, S.C. 1979. Elevated atmospheric partial pressure of CO2 and plant growth. I. nteraction of nitrogen nutrition and photosynthetic capacity in C3 and C4 plants. Oecologia 44, 68-74.
Wong, S.C., and I.R. Cowan. 1979. Stomatal conductance correlates with photosynthetic capacity. Nature 282:424-426.