influence of tea extract supplementation on bifidobacteria during soymilk fermentation
TRANSCRIPT

International Journal of Food Microbiology 188 (2014) 36–44
Contents lists available at ScienceDirect
International Journal of Food Microbiology
j ourna l homepage: www.e lsev ie r .com/ locate / i j foodmicro
Influence of tea extract supplementation on bifidobacteria duringsoymilk fermentation
Danyue Zhao, Nagendra P. Shah ⁎Food and Nutritional Science — School of Biological Sciences, the University of Hong Kong, Pokfulam Road, Hong Kong
⁎ Corresponding author. Tel.: +852 2299 0836; fax: +E-mail address: [email protected] (N.P. Shah).
http://dx.doi.org/10.1016/j.ijfoodmicro.2014.07.0100168-1605/© 2014 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 21 April 2014Received in revised form 9 June 2014Accepted 13 July 2014Available online 18 July 2014
Keywords:BifidobacteriaTea extractAntibacterial effectCell integrityMembrane lipidsSoymilk fermentation
In this study, the influence of tea extract (TE) supplementation on the viability andmembrane lipid compositionsof Bifidobacterium was investigated. Fermented soymilk-tea (SMT) was produced by culturing selectedbifidobacteria in soymilk supplemented with green or black TE. Culturability of four bacteria in the presence ofvarious concentrations of TE was examined by plate count method. Bifidobacterium longum CSCC 5089(BL5089) and B. longum CSCC 5022 (BL5022) were selected for further study based on their sensitivity to TE.The effect of TE supplementation on bacterial cell viability and integrity was assessed by flow cytometry incombination with fluorescence probes. Total lipids of bacterial cell were extracted using an enzyme-assistantextraction method. Fatty acids (FAs) were determined and quantified by GC–MS. Phospholipids (PLs) were sep-arated by high performance thin-layer chromatography (HPTLC) and their relative abundanceswere determinedby densitometry. Total tea phenolic content (TTP) in SMTs with varying concentrations of TE was quantified byHPLC. Among the four Bifidobacterium monitored, TE only significantly inhibited BL5089 (p b 0.01) in a dose-dependent manner, with minimum inhibition concentrations (MICs) determined to be 15.45 mg/mL TTP forgreen TE and 7.34 mg/mL TTP for black TE. Flow cytometric analysis revealed different staining patterns of cellpopulations and compromise in cell integrity upon exposure to high concentrations of TE. Results from GC–MSshowed that unsaturated to saturated FA ratios significantly decreased (p b 0.01) in the membrane of BL5089cells upon TE exposure. Separation of PLs by HPTLC showed dramatic alterations in phosphatidylcholine andphosphatidylglycerol contents due to TE treatment.
852 2299 9914.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
Bifidobacterium is one generally-recognized genre of probiotics fortheir capacity to restore the balance of gut microbiota and to deliverother health benefits to consumers (Picard et al., 2005). According toFAO/WHO (2002), probiotics are “live microorganisms which, whenconsumed in adequate amounts, confer a health effect on the host”. In ad-dition to the “adequate amounts” of intake, probiotic viability at the pointof consumption is also essential for efficacy consideration (Sanders,2000).
Nowadays, mounting evidence of the health-protective effect ofdietary consumption of phenolic compounds attracts more consumers'attention. However, when probiotics interact with dietary phenoliccompounds, their viability and physiological characteristics may beaffected (Parkar et al., 2008). Tea is known for its high amount of poly-phenols, with catechins and theaflavins being the major phenoliccomponents in green tea (GT) and black tea (BT), respectively (Li et al.,2013). Besides its special taste and aroma, the potential of tea as antiox-idant, anti-carcinogenic and anti-inflammatory agents (Khan and
Mukhtar, 2007) makes it one of the world's most popular beverages. Inaddition, it has been found that tea polyphenols (TPs) are able topromote the growth of commensal bacteria, such as bifidobacteria andyogurt microflora (Streptococcus thermophilus and Lactobacillusdelbrueckii ssp. bulgaricus) (Ankolekar et al., 2011; Jaziri et al., 2009;Najgebauer-Lejko et al., 2011), function as prebiotics (Tzonuis et al.,2008) or redox potential reducing agents (Gaudreau et al., 2013). TPshave also been reported to act as inhibitors to food-borne pathogensand viruses (Lee et al., 2006; Perumalla and Hettiarachchy, 2011;Yi et al., 2010). Tea phenolics exert bactericidal effects mainlythrough damaging cytoplasmic membrane (Shimamura et al., 2007;Sivarooban et al., 2008), binding to or altering cell wall-anchoredproteins (Nakayama et al., 2012) and lipid bilayer (Hashimoto et al.,1999; Ikigai et al., 1993), and alteringmembrane fluidity and permeabil-ity (Yi et al., 2010).
Soymilk has received intense attention and popularityworldwide asa healthy dairy substitute and it is rich in isoflavone. As we reportedrecently (Zhao and Shah, 2014), S. thermophilus ASCC 1275,L. delbrueckii ssp. bulgaricus ASCC 859 and Bifidobacterium longumCSCC 5089 (BL5089) were able to grow well in soymilk. The additionof TE significantly inhibited (p b 0.05) the growth of BL5089 comparedwith the control while it had no harmful effect on yogurt starters.

37D. Zhao, N.P. Shah / International Journal of Food Microbiology 188 (2014) 36–44
Inhibition of bifidobacteria by polyphenols was also reported byTabasco et al. (2011). Hence, it is worth examining whether someother bifidobacteria will be affected by tea components and thebactericidal mechanism. According to Puupponen-Pimiä et al.(2001) and Tabasco et al. (2011), higher resistance to phenolic com-pounds in some lactic acid bacteria (LAB) than other Gram-positivespecies, such as Bifidobacterium, is attributed to metabolism ofpolyphenols into monomeric phenolics. However, to our knowledge,no data is available on such metabolic activity in Bifidobacterium.Moreover, there is limited information on the mechanism of howtea phenolic compounds influence the growth and its effect on cellstructures of bifidobacteria. In this study, four Bifidobacterium strainswere exposed to varying concentrations of tea extract (TE) insoymilk. The influence of TE supplementation on the viability ofbacteria was examined by both the conventional plate count methodand flow cytometric analysis. Compositions of cell membrane fattyacids and phospholipids were also monitored. As far as we know, thisis the first mechanistic study on the inhibitory effect of Bifidobacteriumduring fermentation of a soymilk-tea (SMT) beverage.
2. Materials and methods
2.1. Microorganisms and culture conditions
B. longum CSCC 5022 (BL5022), B. longum CSCC 5089 (BL5089),Bifidobacterium bifidum CSCC 5286 (BB5286) and B. bifidum BB-12(BB12) were used for the production of fermented SMT. Each organismwas previously stored at−80 °C. Multiple transfers were performed toproduce active cultures since storage at−80 °Cwould result in bacteriawith longer lag phase. For activation, 10 mL aliquots of sterile MRScontaining 0.05% (w/v) L-cysteine hydrochloride (cys-MRS) (Sigma,St. Louis, MO, U.S.A.) were inoculated with 2% (v/v) of each organismand incubated at 37 °C for 18 h. After the second transfer in MRSbroth, the activated organisms were transferred into sterile soymilkfor another two transfers at 10% (v/v). Activated cultures were usedfor fermentation.
2.2. Preparation of soymilk
Soymilkwas prepared as per themethod of Donkor and Shah (2007)with somemodifications. In brief, soymilk containing lactose (SML)wasmade by dissolving 4% (w/v) soy protein isolate (DuPont, Shanghai,China), 1% (w/v) α-lactose (Sigma Chemical Co., St. Louis, MO) and0.05% (w/v) L-cysteine hydrochloride in double deionized waterpreheated to 50 °C. Upon reconstitution, SML was autoclaved at120 °C for 15 min. Unlike commercial pasteurization, the advantage ofautoclaving at high temperatures is to kill all existing bacteria or fungi,
Table 1Concentrations of caffeine and tea phenolic compounds in soymilk supplemented with varying
CF GA EGC C EGCG EC
GT1X 13.39 0.92 1.48 0.22 1.07 1.8GT2X 25.53 1.82 2.57 0.44 2.11 4.1GT3X 37.86 2.30 4.80 0.69 3.69 5.8GT4X 51.16 3.07 6.16 0.97 4.67 9.2BT1X 12.42 0.50 0.52 0.11 0.04 0.1BT2X 23.11 1.20 1.02 0.22 0.14 0.2BT3X 34.17 1.70 1.38 0.37 0.27 0.4BT4X 47.09 2.23 1.89 0.53 0.35 0.7BT5X 59.44 2.80 2.37 0.69 0.44 0.9
Data is presented as mean (mg/mL) ± SEM of three independent fermentation products.Abbreviations of phenolic compound: GA: gallic acid; EGC: (−)-epigallocatechin; C: catechTF-1: theaflavins; TF-2: theaflavin-3-gallate; TF-3: theaflavin-3,3′-digallate.1/2/3/4/5× indicates normal/double/triple/quadruple/quintuple concentration of the tea extrextract prepared from tea leaf powder (2%, w/v) brewed in boiling water.N.D. = not detected.
so the result of experiments should be the response of the testingbacteria only.
2.3. Preparation of tea extract (TE)
Green tea (Zhuyeqing tea, Sichuan Province, China) and black tea(Dianhong, Yunnan Province, China) were purchased from local tearetailers. TE was prepared as per the method of Zhao and Shah (2014)with minor modifications. In brief, tea leaf powder (2%, w/v, corre-sponding to the strength of “a normal cup of tea” according to Yam etal. (1997)) was infused in boiling deionized water for 10 min. Greenor black TE were produced by first suction filtered through triple-layered Whatman #1 filter paper twice and then filter-sterilized with0.22-μm membrane (Millipore, Bedford, MA, U.S.A.). The sterilized fil-trates were collected in 50 mL sterile tubes and frozen at −80 °Cbefore freeze-drying using a Virtis freeze mobile (Virtis Co., Gardiner,U.S.A.). The freeze-dried TE powder was stored at −20 °C for furtheruse. TE power was dissolved in the same volume of SML, as that beforefreeze-drying to produce SMT containing “normal concentration (1×)”of TE. Double (2×), triple (3×), quadruple (4×) and quintuple (5×)concentrations of TE supplemented to SMLwere prepared by dissolvingTE in 1/2, 1/3, 1/4 and 1/5 SML volume thatwasused for normal concen-tration, respectively.
2.4. Preparation of fermented soy-milk tea and enumeration of viable cellby plate count method
The activated cultures (3%, v/v) were transferred into 10 mL ofSML or SMT with green TE and black TEs both at normal and doubleconcentrations. Samples were incubated at 37 °C for 24 h andfermented SML or SMT were referred to as FS or FST, respectively.Upon completion of fermentation, viable cells were enumeratedusing cys-MRS agar by pour plate method. Plates were incubated at37 °С for 24 h in an anaerobic jar (BD GasPak™, Sparks, MD, U.S.A.).
2.5. Extraction and HPLC quantification of tea polyphenols
A 2 mL aliquot of FS or FST with varying concentrations of TE wasmixed with 4 mL 80% methanol, vortexed for 1 min and centrifuged at5000 × g for 10min at 20 °С. The non-inoculated SMT or SML incubatedat 37 °С for 24 hwas used as a control and 2mL of a control sample wasmixed with 4 mL of water–methanol–acetic acid (20:78:2, v/v). Theprecipitate was further extracted twice with 2 mL of 80% methanoland the supernatants were combined. All solvents were of HPLC-grade(Fisher Scientific, Pittsburgh, PA, U.S.A.) and pooled supernatants werefiltered through 0.45-μmhydrophilic filter (Corning, N.Y., U.S.A.) beforeloaded onto a HPLC column.
amounts of green tea (GT) or black tea (BT) extract.
ECG TF1 TF2 TF3 Total tea phenolics
2 0.47 N.D. N.D. N.D. 5.982 0.99 N.D. N.D. N.D. 12.056 1.50 N.D. N.D. N.D. 18.854 2.03 N.D. N.D. N.D. 26.153 0.08 0.48 0.20 0.36 2.426 0.17 0.91 0.41 0.67 5.019 0.25 1.31 0.65 0.92 7.348 0.37 1.75 0.81 1.16 9.879 0.48 2.21 1.07 1.49 12.54
in; EGCG: (−)-epigallocatechin gallate; EC: (−)-epicatechin; ECG: (−)-epicatechin gallate;
act supplemented to soymilk to produce soymilk-tea. Normal concentration refers to tea

38 D. Zhao, N.P. Shah / International Journal of Food Microbiology 188 (2014) 36–44
HPLC separation of nine phenolic compounds including gallic acid(GA), catechin (C), (−)-epigallocatechin gallate (EGCG), theaflavin(TF-1), theaflavin-3-gallate (TF-2) and theaflavin-3,3′-digallate(TF-3) (Sigma Chemical Co., St. Louis, MO, U.S.A.); (−)-epigallocatechin(EGC), (−)-epicatechin (EC), (−)-epicatechin gallate (ECG) andcaffeine (CF) (Wako, Osaka, Japan) was achieved as previouslydescribed (Zhao and Shah, 2014). Quantification result is shown inTable 1.
2.6. Determination of minimum inhibitory concentrations (MIC) andminimum bactericidal concentration (MBC) of TE against BL5089 in SMT
Cell growth of BL5089 at 37 °С for 24 h in soymilk-tea (SMT) atvarying concentrations of TE was recorded by plate count method.BL5022 was used as a phenolic-resistant strain for comparison. MICis defined as the lowest total tea phenolic content (TTP) of green orblack TE that completely inhibited the bacterial growth as comparedwith the initial cell population. MBC is defined as the lowest TTP in TEthat killed N99.9% of the initial inoculum (ca. log CFU/mL b 4) (Donget al., 2012). For determining MIC and MBC, cells were plated on cys-MRS agar plate and incubated aerobically at 37 °C for 28 h. TTP weredetermined by adding up all major phenolic compounds in TE asquantified by the HPLC method described above.
2.7. Fluorescent staining of bacterial cells and flow cytometry (FCM)
The impact of TE supplementation on cell viability and cell integrityof BL5089 and BL5022 was investigated by fluorimetric FCM. Cells werecultured in 10 mL cys-MRS containing varying concentrations of TE at37 °С for 24 h. Cells grown in MRS without TE was used as a control.After incubation, cells were harvested by centrifugation (4000 ×g,8 min, 4 °С) and washed twice with filter-sterilized PBS buffer(pH 7.4, 4 °С). Cell pellets were resuspended in 10 mL of buffer andfiltered through 1.0-μm hydrophilic membrane (Costar, Nuclepore,Charlotte, NC, U.S.A.). Cell density of filtrates was adjusted to OD600 =0.5–0.6 before staining. Fluorescence staining with carboxylfluoresceindiacetate (cFDA) and propidium iodide (PI) (Invitrogen, Karlsruhe,Germany) was performed as per the method of Amor et al. (2002).
Samples were analyzed in a BD FACSAria III flow cytometer (BectonDickinson, San Jose, CA, U.S.A.) equipped with an air-cooled laseremitting blue light at 488 nm. Four parameters were recorded usingBD FACSDiva software: forward scatter (FSC), side scatter (SSC), greenfluorescence (cFDA) and red fluorescence (PI). All registered signalswere logarithmically amplified and 10,000 events were recorded persample. Different cell populations were separated and gated accordingto their fluorescence staining pattern and intensity of the premixedsample of 50% freshly harvested and 50% heat-killed (90 °С for30 min) bacterial cells in both single and double-staining assays. Datawas analyzed in FlowJo alias software (version 7.6, TreeStar, Ashland,OR, U.S.A.).
2.8. Extraction and profiling of membrane fatty acids (FAs)
2.8.1. Isolation of bacterial cells and enzymatic digestion of protein matrixEnzymatic digestion of protein was performed in order to free
cells from the FS or FST matrices and to “shave off” bacterial surfaceproteins for more efficient extraction of membrane lipids. Fivemilliliters of FS or FST was homogenized with 40 mL of PBS buffer(pH 7.4, 4 °С) in a 50-mL centrifugal tube. The mixture was centrifugedat 20 ×g for 10min, followed by 150 ×g for 5 min at 4 °С. The superna-tant was transferred to another tube and centrifuged at 4000 ×g for10min at 4 °С. The supernatantwas discarded and the pelletwaswashedwith 40 mL of chilled PBS for twice and re-collected by centrifugation at4000×g for 10min at 4 °С. The cell pelletwas resuspended in 0.5mL (for5 mL culture) of the same buffer. Proteinase K (25 μg/mL, Sigma) wasadded to the cell suspension in the presence of 5 mM dithiothreitol
(DTT) and incubated for 30 min at 37 °С. An aliquot (100 μL) of proteaseinhibitor cocktail solution was then added to stop enzymatic reaction.The cocktail solution was prepared by dissolving one tablet of EDTA-free Protease Inhibitor Cocktail (cOmplete, Roche Applied Science,Penzberg, Germany) in 10 volumes (w/v) of PBS (pH 7.4). The digestedmixture before and after incubation was serially diluted with PBS toassess bacterial viability (data not shown). The subsequent mixturewas centrifuged at 4000 ×g for 10 min at 4 °С and cell pellets werecollected.
2.8.2. Extraction of cellular fatty acidsMembrane FAs were extracted from ca. 0.5 g cell pellets (wet
weight) and then converted to fatty acidmethyl ester (FAME) accordingto the Microbial Identification System (MIS) protocol (1990). Decanoicacid (C10:0) was used as the internal standard and was added tocell isolates before extraction and methylation. The extracts wereevaporated under nitrogen flow and re-dissolved in GC-grade hexane(Fisher Scientific, Pittsburgh, PA, U.S.A.) for injection.
2.8.3. Separation and identification of fatty acids by gas chromatography–mass spectroscopy (GC–MS)
Profiles of bacterial FAME were obtained using an Agilent 6590N-5973N GC–MS system (Agilent, Atlanta, GA, U.S.A.) equipped withan Agilent 7694E auto-sampler and a capillary DB-wax column(30 m × 0.25 mm id, 0.25 μm film thickness; J&W Scientific, Folsom,CA, U.S.A.), with an injection volume of 1 μL (splitless mode). The injec-tor and the detector were both held at 250 °C. The temperature wasprogrammed from 100 °C (held for 1 min) to 190 °C at a rate of 4 °C/min, elevated to 235 °C at a rate of 10 °C/min, and finally to 250 °C ata rate of 4 °C/min (held for 4 min). The carrier gas was helium at arate of 1 mL/min. Results were expressed as relative molar percentages(mol%) for each FA. The ratios of unsaturated to saturated FA (U/S)werecalculated accordingly.
2.9. Extraction and profiling of membrane phospholipids (PLs)
2.9.1. Enzymatic digestion of cell wall and cell surface proteinsApproximately 1 g of cell pellets (wet weight) was suspended in
1mL PBS (pH 6.2, the optimal pHwith ionic strength at ca. 0.18 M forlysozyme according to Dickman and Proctor, 1952) and incubatedwith 1 mg/mL lysozyme (ca. 50,000 units/mg, Sigma–Aldrich,St. Quentin-Fallavier, France) at 37 °C for 1 h, washed once withPBS (pH 7.4), followed by digestion with proteinase K as describedin Section 2.8.1. After digestion, cell pellets were collected and theextraction process was performed under N2 to minimize oxidation.
2.9.2. Neutral and acidic extraction of membrane phospholipids (PLs)For efficient extraction of major PLs from bacterial cells, a two-step
extraction method was employed (Özbalci et al., 2013). For neutral ex-traction, cell pellets were resuspended in 1800 μL chloroform/methanol(1:2, v/v) and rotatively agitated for 60 min at room temperature (RT),while vortexing for 1 min every 10 min. Samples were centrifuged at10,000 ×g for 5 min at 4 °С. The supernatant was transferred to a 5-mLcentrifugal tube, followed by adding 600 μL chloroform and 1 mL 0.8%NaCl solution. The supernatant was vortexed for 1 min and centrifugedat 5500 ×g for 5 min at 4 °С. The lower organic phase was collected inanother glass tube. After neutral extraction, cell pellets were processedwith acidic extraction by resuspending in 1 mL chloroform/methanol/37% HCl (40:80:1, v/v) and rotatively agitated for 30 min at RT, whilevortexing for 30 s every 5 min. The tube was transferred to ice, followedby adding 250 μL cold chloroform and 450 μL cold 0.1 NHCl. Themixturewas vortexed for 1 min before centrifugation (5500 ×g, 5 min at 4 °С).The chloroform-rich phase from both neutral and acidic extraction wascollected in a glass tube and dried under N2 stream. The weight ofdried lipids was determined and stored in chloroform/methanol(90:10, v/v, 50 mg/mL) at −20 °С until analysis.

3.2
4.2
5.2
6.2
7.2
8.2
9.2
SML 1X 2X 3X 4X 5X
log
(CFU
/mL)
B. longum CSCC 5022
GT
BT
**
**** **
****
3.2
4.2
5.2
6.2
7.2
8.2
9.2
SML 1X 2X 3X 4X 5X
log
(CFU
/mL)
B. longum CSCC 5089
GT
BT****
**
**
**
**
****
**
Fig. 1. Viable cell count of B. longum CSCC 5089, B. longum CSCC 5022 in soymilksupplemented with varying concentrations of green tea (GT) or black tea (BT) extract.Data is expressed as the mean log CFU/mL ± SEM of three independent fermentationproducts. 1/2/3/4/5× indicates normal/double/triple/quadruple/quintuple concentrationof the tea extract added to soymilk (Table 1). Asterisks (**) above a data point indicatessignificant difference at a level of p b 0.01 compared with the soymilk containing lactose(SML) control.
39D. Zhao, N.P. Shah / International Journal of Food Microbiology 188 (2014) 36–44
2.9.3. Separation of phospholipids by high performance thin layerchromatography (HPTLC)
Silica gel 60 thin-layer chromatographic plates (Merck, Darmstadt,Germany) were developed with solvent prior to spotting in order toremove impurities from the adsorbent layer. The plates were air-driedfor 10 min and oven activated for 20 min at 100 °С before use. A filterpaper was placed in the developing chamber and the developingsolvent added. The chamber was allowed to saturate for 10 min beforechromatography. For identification, six standard PLs (Sigma), cardiolipin(CL), phosphatidylethanolamine (PE), phosphatidylglycerol (PG),phosphatidylinositol (PI), phosphatidylcholine (PC), and lyso-phosphatidylcholine (lyso-PC) were dissolved in chloroform:methanol(90:10, v/v). The same volume of PL extract and PL standards wasspotted on the plates using spotting capillaries.
Phospholipids were separated on TLC plate using chloroform/methanol/acetic acid/water (71:20:6:2.5, v/v) as mobile phase. Forvisualization of lipid bands, plates were sprayed with the followingreagents: a) 0.5% (w/v) vanillin in ethanol/concentrated sulphuricacid (97:3, v/v) solution, for all polar lipids (Rakhuba et al., 2009);b) ammonium molybdate/perchloric acid reagent, for phospholipids(Nzai and Proctor, 1998). Since the bands detected with vanillinsolution were much sharper in color, bands on plates treated withvanillin solution were scanned in a Molecular Imager XR+ System(Bio-Rad, Hercules, CA, U.S.A.) and analyzed in Image Lab (version4.0, Bio-Rad). Identification of major PLs was achieved by comparingrelative mobility (Rf) values of PL standards and the lipid bands.Relative quantity of individual PLs was determined according to thedensity of detected bands.
2.10. Statistical analysis
For each assay, three or more independently incubated sampleswere prepared for conducting plate count, FCM assay, bacterial FA andPL profiling. The data was subjected to one-way analysis of variance(ANOVA) (Turkey andGames-Howell tests) by SPSS 20.0 (IBMSPSS Sta-tistics, IBM Corp, Somers, NY). Results with p b 0.01 or p b 0.05 wereconsidered statistically significant.
3. Results and discussion
3.1. Impact of tea extract supplementation on culturability and cellularproperties of bifidobacteria
3.1.1. Cell culturabilityCell culturability of four bifidobacteria was assessed by viable cell
count on agar plates and results are displayed in Table 2. All strainsshowed ca. 2 log CFU/mL increase during fermentation of SML, with afold change at 165 for BL5022, followed by BL5089 (44 fold), BB12 (42fold) and BB5286 (41 fold). When cells were exposed to TE at normal
Table 2Cell culturability (cell forming unit/mL) of Bifidobacterium longum CSCC 5022 (BL5022),B. longum CSCC 5089 (BL5089), B. bifidum CSCC 5286 (BB5286) and B. bifidum BB-12(BB12) in soymilk supplementedwith normal (1×) or double (2×) concentration of greentea (GT) or black tea (BT) extract in soymilk.
BL5022 BL5089 BB5286 BB12
Initial 6.99 ± 0.02⁎ 6.57 ± 0.02⁎ 6.61 ± 0.01⁎ 6.87 ± 0.02⁎
SML 9.21 ± 0.01 8.20 ± 0.02 8.22 ± 0.01 8.75 ± 0.01GT1X 9.45 ± 0.02⁎ 7.84 ± 0.02⁎ 8.42 ± 0.01⁎ 8.88 ± 0.02⁎
GT2X 9.27 ± 0.02 7.42 ± 0.02⁎ 8.29 ± 0.03 8.75 ± 0.03BT1X 9.38 ± 0.03 7.87 ± 0.02⁎ 8.58 ± 0.02⁎ 8.79 ± 0.02BT2X 9.28 ± 0.01 7.56 ± 0.03⁎ 8.43 ± 0.02⁎ 8.74 ± 0.02
Data is presented as the mean log CFU/mL ± SEM of three independent fermentationproducts. Asterisks in the same column indicate significant difference at a level of p b 0.05compared with SML. 1/2/3/4/5× indicates normal/double/triple/quadruple/quintupleconcentration of the tea extract added to soymilk (refer to Table 1).
and double concentrations, TE promoted the growth of BL5022,BB5286 and BB12. In particular, GT1X (5.98 mg/mL TTP) favored thegrowth of BL5022 and BT1X (2.42 mg/mL TTP) promoted BB5286significantly (p b 0.05) by increasing 2.6% and 4.3% of their cell popula-tion in SML, respectively. Studies have revealed that the reducing natureof tea polyphenols in TE might protect cells from oxidative stress andcreate a more anaerobic environment (Gaudreau et al., 2013), andthus promoted the growth of bifidobacteria. Similar to our previousstudy (Zhao and Shah, 2014), BL5089 was the only strain whoseproliferation was significantly limited in SMT compared with that inSML, without significant difference among tea types (p b 0.05). Itappears that TEs, mainly tea polyphenols, acted as antibacterial agentsagainst BL5089.
To confirm the deleterious effect of TE on the growth of BL5089, bac-teria were treated with higher TE concentrations in soymilk and thephenolic-resistant strain, BL5022, was used as a positive control. Fig. 1depicts the alteration in viable cell count of BL5089 and BL5022 andthere is a pronounced trend of inhibition against BL5089 by TE in adose-dependent manner. GT exhibited higher inhibitory effects againstBL5089 than BT, which was significant at 3× and 4× TE concentrations(p b 0.01). The MIC was determined to be 2.5× (15.45 mg/mL TTP) forGT and 3× (7.34 mg/mL TTP) for BT, while MBC was 4× (26.15 mg/mLTTP) and 4.5× (11.21 mg/mL TTP) for GT and BT, respectively. Ourresult is in agreement with that reported by Chan et al. (2011), whichindicated that BT had higher MIC values than GT against the pathogenstested. TE is a complex system containing a variety of components andconstituent analysis of green and black TE by HPLC helped to explain
40 D. Zhao, N.P. Shah / International Journal of Food Microbiology 188 (2014) 36–44
the discrepancy (Table 1). As detected by HPLC, the major componentsin GT extract are catechins, gallic acid and caffeine, while for BT,theaflavins, gallic acid and caffeine contribute the majority. Thedifference in phenolic profiles can be the main reason and this wasalso suggested by Chan et al. (2011). Regarding BL5022, although aslight decrease in CFU was found at higher TE concentrations, itobtained a log CFU/mL value of 8.67 at 26.15 mg/mL TTP (GT5X)and 8.31 at 12.54 mg/mL TTP (BT5X), which demonstrated its highphenolic tolerance.
Although cell viability determination by counting bacterial colonieson agar plates has been employed as the standard practice in microbio-logical research and also in food processing industry, it actually deter-mines the culturability of cells adapted to a certain environment onagar plates (Ananta et al., 2004). Mounting evidence demonstratesthat cells grown under stress may stay in a non-culturable but viableor sub-lethally injured state (Mizunoe et al., 2000). On the contrary,some injured cells may resuscitate and recover their ability to replicateon agar plates (Wu, 2008). For more accurate detection of cell popula-tions in different physiological status, flow cytometric analysis wasperformed.
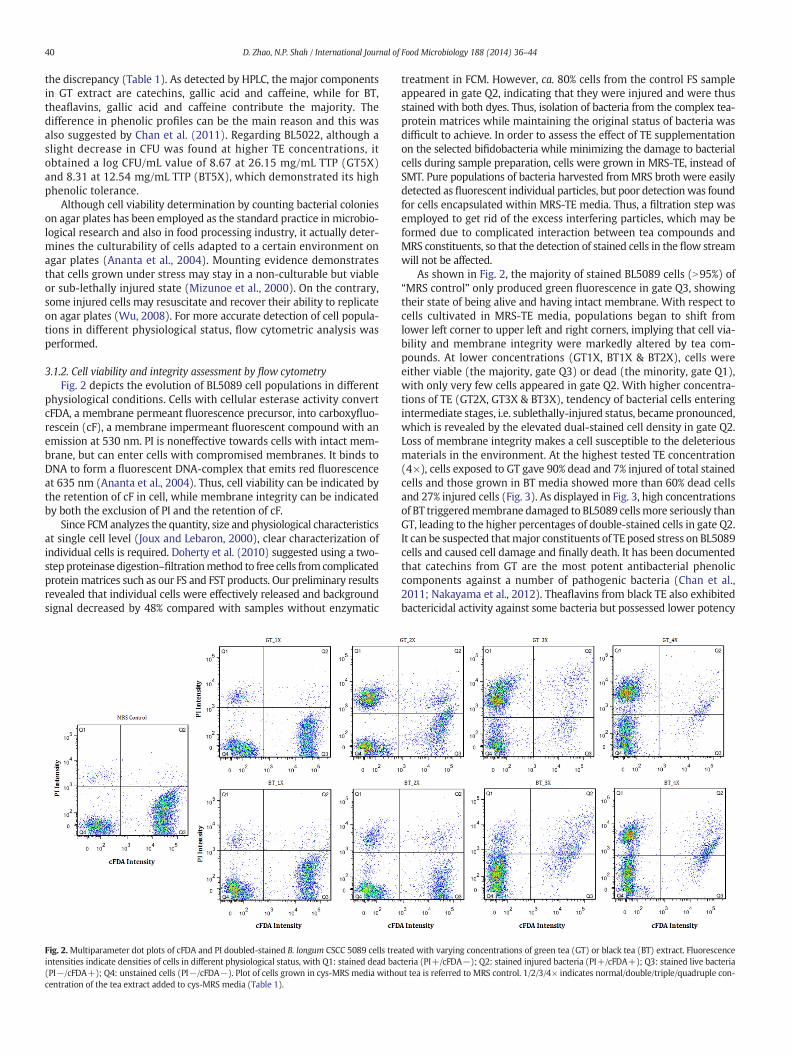
3.1.2. Cell viability and integrity assessment by flow cytometryFig. 2 depicts the evolution of BL5089 cell populations in different
physiological conditions. Cells with cellular esterase activity convertcFDA, a membrane permeant fluorescence precursor, into carboxyfluo-rescein (cF), a membrane impermeant fluorescent compound with anemission at 530 nm. PI is noneffective towards cells with intact mem-brane, but can enter cells with compromised membranes. It binds toDNA to form a fluorescent DNA-complex that emits red fluorescenceat 635 nm (Ananta et al., 2004). Thus, cell viability can be indicated bythe retention of cF in cell, while membrane integrity can be indicatedby both the exclusion of PI and the retention of cF.
Since FCM analyzes the quantity, size and physiological characteristicsat single cell level (Joux and Lebaron, 2000), clear characterization ofindividual cells is required. Doherty et al. (2010) suggested using a two-step proteinase digestion–filtrationmethod to free cells from complicatedproteinmatrices such as our FS and FST products. Our preliminary resultsrevealed that individual cells were effectively released and backgroundsignal decreased by 48% compared with samples without enzymatic
Fig. 2.Multiparameter dot plots of cFDA and PI doubled-stained B. longum CSCC 5089 cells treintensities indicate densities of cells in different physiological status, with Q1: stained dead bac(PI−/cFDA+); Q4: unstained cells (PI−/cFDA−). Plot of cells grown in cys-MRS media withocentration of the tea extract added to cys-MRS media (Table 1).
treatment in FCM. However, ca. 80% cells from the control FS sampleappeared in gate Q2, indicating that they were injured and were thusstained with both dyes. Thus, isolation of bacteria from the complex tea-protein matrices while maintaining the original status of bacteria wasdifficult to achieve. In order to assess the effect of TE supplementationon the selected bifidobacteria while minimizing the damage to bacterialcells during sample preparation, cells were grown in MRS-TE, instead ofSMT. Pure populations of bacteria harvested fromMRS broth were easilydetected as fluorescent individual particles, but poor detectionwas foundfor cells encapsulated within MRS-TE media. Thus, a filtration step wasemployed to get rid of the excess interfering particles, which may beformed due to complicated interaction between tea compounds andMRS constituents, so that the detection of stained cells in the flow streamwill not be affected.
As shown in Fig. 2, the majority of stained BL5089 cells (N95%) of“MRS control” only produced green fluorescence in gate Q3, showingtheir state of being alive and having intact membrane. With respect tocells cultivated in MRS-TE media, populations began to shift fromlower left corner to upper left and right corners, implying that cell via-bility and membrane integrity were markedly altered by tea com-pounds. At lower concentrations (GT1X, BT1X & BT2X), cells wereeither viable (the majority, gate Q3) or dead (the minority, gate Q1),with only very few cells appeared in gate Q2. With higher concentra-tions of TE (GT2X, GT3X & BT3X), tendency of bacterial cells enteringintermediate stages, i.e. sublethally-injured status, became pronounced,which is revealed by the elevated dual-stained cell density in gate Q2.Loss of membrane integrity makes a cell susceptible to the deleteriousmaterials in the environment. At the highest tested TE concentration(4×), cells exposed to GT gave 90% dead and 7% injured of total stainedcells and those grown in BT media showed more than 60% dead cellsand 27% injured cells (Fig. 3). As displayed in Fig. 3, high concentrationsof BT triggeredmembrane damaged to BL5089 cellsmore seriously thanGT, leading to the higher percentages of double-stained cells in gate Q2.It can be suspected thatmajor constituents of TE posed stress on BL5089cells and caused cell damage and finally death. It has been documentedthat catechins from GT are the most potent antibacterial phenoliccomponents against a number of pathogenic bacteria (Chan et al.,2011; Nakayama et al., 2012). Theaflavins from black TE also exhibitedbactericidal activity against some bacteria but possessed lower potency
ated with varying concentrations of green tea (GT) or black tea (BT) extract. Fluorescenceteria (PI+/cFDA−); Q2: stained injured bacteria (PI+/cFDA+); Q3: stained live bacteriaut tea is referred to MRS control. 1/2/3/4× indicates normal/double/triple/quadruple con-

0
10
20
30
40
50
60
70
80
90
100
MRS
Control
GT1X GT2X GT3X GT4X BT1X BT2X BT3X BT4X
Cell
popu
latio
n di
strib
utio
n (%
of s
tain
ed c
ells
) B. longum CSCC 5089
**
**
**
**
**
**
**
**
**
**
**
**
**
**
**
**
**
**
**
**
**
**
**
0
10
20
30
40
50
60
70
80
90
100
MRS
Control
GT1X GT2X GT3X GT4X BT1X BT2X BT3X BT4X
Cell
popu
latio
n di
strib
utio
n (%
of s
tain
ed c
ells
)
B. longum CSCC 5022
Q1: cFDA-/PI+ Q2: cFDA+/PI+ Q3: cFDA+/PI-
**
**
**
** **
**
**
** ** **
**
****
Fig. 3. Distribution of three subpopulations (gate Q1, Q2 and Q3) of Bifidobacterium cellsgrown in varying concentrations of green tea (GT) or black tea (BT) extract. Data is presentedas mean ± SEM of at least three independent experiments. Asterisks (**) above a columnindicates significant difference at a level of p b 0.01 compared with the MRS Control inQ1/Q2/Q3. 1/2/3/4× indicates normal/double/triple/quadruple concentration of the teaextract added to cys-MRS media (Table 1).
41D. Zhao, N.P. Shah / International Journal of Food Microbiology 188 (2014) 36–44
(Jaziri et al., 2009). Viable cell population of BL5022 was slightly de-creased with the increasing TE concentrations, accompanied by an in-crease in injured and dead cells. It appeared that results obtained byplate count method and FCM contradicted since at the low TE concen-trations (GT1X, GT2X, BT1X and BT2X), more loss of cell viability and in-tegrity were observed as compared with the control. However, whencombining populations in Q2 and Q3, the twomethods produced similartrends, i.e. growth of BL5022was promoted at lowTE concentrations. Ac-cording to a review by Wu (2008), under conditions favorable to bacte-ria, injured cells may recover to form colonies on agar plates. Thus, theadditional number of cells observed by plate count may be attributedto the recovery of sub-lethally injured cell on agar plates after incubatingfor 28 h. Nonetheless, the difference in the distribution of stained cellpopulations between BL5089 and BL5022 cell exposed to TE still demon-strated the harmful effects towards BL5089.
3.2. Impact of tea extract supplementation on cell membrane lipids
Bacterial lipid metabolism is closely related to stress responses.Environmental fluctuations may induce alteration in cell membrane
lipid compositions, mostly FAs and polar lipid species to maintain anideal physiological state of cell membrane (Guerzoni et al., 2001;Steponkus et al., 1995). Tea polyphenols and other phenolic acidshave been documented for their high affinity to bind to lipid bilayerand the ability to damage membrane structures (Ikigai et al., 1993;Kumazawa et al., 2004). In order to explain bacterial cell responsesto phenolic stress induced by TE, membrane FAs and PLs of twobifidobacteria were extracted and quantified.
3.2.1. Membrane fatty acid compositionThe relative percentages of major FAs (contributing a mol% higher
than 1) and the unsaturated to saturated FA ratios (U/S) are presentedin Fig. 4, with results for BL5089 in the upper panel and BL5022 in thelower panel. A total of 6 fatty acids were observed in BL5089 mem-branes, and an additional saturated FA, dodecanoic acid (C12:0), wasdetected in BL5022 membranes, regardless of the culture media. ForBL5089, TE substantially altered relative percentages of individual FAsin a dose-dependent manner. Particularly, relative abundances ofC14:0, C18:1, C18:2 and C18:3 were significantly reduced (p b 0.01),while those of C16:0 and C18:0, both saturated FAs, were increased.This lead to gradual decrease in U/S ratios and finally obtained valuesas low as 0.32 (GT4X) and 0.45 (BT4X). It is noteworthy that green TEcaused a sharp decrease in the proportion of C18:2 by 2.1 folds and anincrease in C16:0 by 1.6 folds when its concentration was increasedfrom 2× to 3×. Black tea exposure decreased the U/S ratio steadily,from 0.88 for cells grown in control SML to 0.45 (BT4X). In addition,fluctuations in FA composition of cells grown in GT media, particularlyat high TE concentrations (3× and 4×), were more significant as com-pared with their BT counterparts (p b 0.01). Regarding the phenolic-resistant strain, BL5022, some decrease in unsaturated FA abundancewas also detected but it was much less than that for BL5089. Thevariations of individual FAs in cells exposed to TE were not significant(p b 0.05), except for those of C16:0 and C18:2, compared with that ofthe control.
The distribution of saturated and unsaturated FAs and the fatty acylchain conformation are among the major factors that affect cell mem-brane fluidity, and it is necessary to maintain a suitable fluidity formembrane to function properly (Denich et al., 2003; Murga et al.,1999). In many studies related to the successful adaptation of LAB orbifidobacteria cells to various environmental stresses, such as coldshock (Wang et al., 2005), bile stress (Kimoto-Nira et al., 2009) andacid stress (Wu et al., 2012), increases in U/S ratio were observed. Inour study, however, relative unsaturated FA contents in bothbifidobacteria were reduced, implying that TE, or more specific, TPs,impeded the synthesis of unsaturated FAs in cell membrane andincreased cell rigidity, as pointed out by Tsuchiya (1999). As determinedby Murga et al. (1999), L. acidophilus cells grown at 25 °C showed con-siderably more C18:2 and C16:0 than cells grown at 37 °C, and such al-teration in lipid content enhanced the lipid membrane stability. Similarresultswere also obtained in a study by Kimoto-Nira et al. (2009). It wasalso found that increase in the degree of FA unsaturation or decrease inthe average chain length was a common adaptation response in manystress-adapted bacterial strains (Aricha et al., 2004; Guerzoni et al.,2001). However, these were not found in the phenolic-sensitiveBL5089, which may partly explain the loss of cell viability and integrityfor BL5089 (Figs. 2 and 3), particularly at high TE concentrations.
3.2.2. Membrane phospholipid compositionThe PLs in the cell membrane of BL5089 and BL5022 were also
characterized to investigate the influence of TE supplementation onbacterial growth. As shown in Fig. 5, CL, PE, PG, PI, PC, lyso-PC andan unknown polar lipid (mobility Rf value: 0.790–0.823), werefound to be the major lipid components in the membrane ofBL5089 and BL5022 cells. Although the Folch et al. method (1957),involving the use of a single-phase mixture of chloroform/methanolto extract lipids fromhomogenizedwhole cells, has beenwidely applied

0
5
10
15
20
25
30
35
40
45
50
14:0 16:0 18:0 18:1 18:2 18:3
Dis
trib
utio
n (m
ol%
)
SML
GT1X
GT2X
GT3X
GT4X
BT1X
BT2X
BT3X
BT4X0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
SML 1X 2X 3X 4X
U/S
B. longum CSCC 5089
GT
BT
**
****
**
0
5
10
15
20
25
30
35
40
45
50
12:0 14:0 16:0 18:0 18:1 18:2 18:3
Dis
trib
utio
n (m
ol%
)
Major fatty acids
GT GT GT GT GT GTBT BT BT BT BT BT GT BT
0.30
0.40
0.50
0.60
0.70
0.80
0.90
1.00
SML 1X 2X 3X 4X
U/S
B. longum CSCC 5022
** **** **
GT GT
C
GT GT GT GTBT BT BT BT BT BT
Fig. 4. Variation in membrane fatty acid composition and unsaturated:saturated fatty acid ratio (U/S) of bacterial cells treated with varying concentrations of green tea (GT) or black tea(BT) extract. 1/2/3/4× indicates normal/double/triple/quadruple concentration of the tea extract added to soymilk (Table 1). Data is presented as mean ± SEM of three independentexperiments. Asterisks (**) above a column indicates significant difference at a level of p b 0.01 compared with the soymilk containing lactose (SML) control.
42 D. Zhao, N.P. Shah / International Journal of Food Microbiology 188 (2014) 36–44
for total lipid extraction, the recovery of some charged lipids andminorlipid components was low (Özbalci et al., 2013). In addition, as sug-gested by Filgueiras and den Kamp (1980), CL was poorly extractedfrom Gram-positive bacteria due to the presence of cell wall, possiblydue to the strong interaction between CL and some intracellular constit-uents. Thus, our study first utilized an enzymatic digestion processwithlysozyme and protease K, followed by a two-step extraction using botha neutral chloroform/methanol solution and a HCl-acidified mixture(Limonet et al., 2007). Novik et al. (2006) isolated and analyzed themajor PLs in the representative species of Bifidobacterium and theirfindings substantially agreed with our results. However, the relativeamount of individual PLs varies from strain to strain, and also fromme-dium tomedium for the same strain, as demonstrated by the profiles ofPL extracted from BL5089 and BL5022 cultured in various media.
For BL5089, the majority of PLs was contributed by PE (24.2%),followed by PI (23.0%), PC (21.3%) and PG (16.4%) in cells grown inSML. When TE was added, the abundance of PC was considerablyreduced regardless of TE type, and PG was significantly increased(p b 0.01) and relative percentages of the rest PLs were littlechanged. Relative percentage of PG increased by nearly 50% in cellsgrown in GT4X and BT4X while that of PC dropped to ca. 3% of totaldetected PLs. In contrast, for the phenolic-resistant strain BL5022,relative percentages of PL of cells grown in different media did notvary significantly (p b 0.05). This divergence between the twoB. longum strains may also explain their difference in growth andsurvival in phenolic-rich media. As one primary barrier to the mate-rials exchanging between the external environment and cytoplasm,it can be expected that the structure and the electrostatic propertiesof membrane lipid bilayer should be stable in order to optimize cellgrowth under stress. Our data showed that PL profiles of BL5089
strain were substantially altered upon exposure to TE, rather thanthe phenolic-resistant strain. The net charge of the lipid bilayer wasmore negative due to the significantly higher (p b 0.01) amount ofPG (anionic) but lower PC (zwitterionic). This indicated a shift inthe membrane lipid metabolism towards the synthesis of anionicPLs, which may result in greater electrostatic repulsion betweenthe lipid bilayer and TPs (Lewis and McElhaney, 2000). As found byIkigai et al. (1993), tea catechins damaged membrane integrity andhad high affinity to neutral or positively charged components inthe lipid bilayer, rather than negatively charged components. Thus,increase in synthesis of anionic PLs can be a self-protective responseto phenolic stress. However, additional negative charge on themembrane alters the balance of electrostatic and hydrophobicinteractions with external environment. As a result, the bindingof cationic compounds and elevated level of hydration may leadto conformational changes in the structure of transmembrane α-helicesof membrane proteins and produce the subsequent changes in helicepackings (Lopez et al., 2006). Since there is strict cross-regulationbetween the different membrane components, dramatic alteration inindividual constituents may cause malfunction of cell membrane andfinally lead to death.
Referring to our findings on membrane lipid compositions, themembrane of BL5089 cells was found to be disturbed in an unusualand extreme manner due to TE supplementation. TE, particularlyTPs has been reported to induce irreversible changes to cell mem-brane and cause substantial damage to bacteria (Ikigai et al., 1993).Severe damage in membrane is also related to compromise in cellintegrity, which is demonstrated in the FCM assays (Figs. 2 and 3).As in a micro-biosystem, any dramatic variation in individualcomponents may lead to lethal damage to the whole organism,

0
10
20
30
40
50
60
70
80
90
100
Mem
bran
e ph
osph
olip
id c
ompo
sitio
n (%
)B. longum CSCC 5089
CL
PE
PC
PG
PI
Unknown 1
Lyso-PC
0
10
20
30
40
50
60
70
80
90
100
SML GT2X GT3X GT4X BT2X BT3X BT4X
Mem
bran
e ph
osph
olip
id c
ompo
sitio
n (%
)
B. longum CSCC 5022
CL
PE
PC
PG
PI
Unknown 2
Lyso-PC
Fig. 5. Distribution of individual phospholipids extracted from bacterial cells treated withvarying concentrations of tea extract. Phospholipids: CL, cardiolipin; PE, phosphatidyleth-anolamine; PG, phosphatidylglycerol; PI, phosphatidylinositol; PC, phosphatidylcholine;lyso-PC, lyso-phosphatidylcholine. Unknown, unknown polar lipids detected. 1/2/3/4×indicates normal/double/triple/quadruple concentration of the tea extract added tosoymilk (Table 1). PLs extracted from cells grown in soymilk containing lactose (SML)was used as a control.
43D. Zhao, N.P. Shah / International Journal of Food Microbiology 188 (2014) 36–44
significant modification of cell membrane lipid constituents due toTE supplementation may account for the intolerance of BL5089cells in high-phenolic media.
4. Conclusion
In this study, a novel soymilk-tea beverage was developed forcultivation of selected bifidobacteria. Bifidobacteria exhibited differ-ent responses to tea phenolics and different strains of the samespecies showed distinct sensitivity to tea phenolic compounds. Cellmembrane was one of the targets of tea components as the integrityof BL5089 cell and the composition of its cell membrane lipids weregreatly altered upon 24 h exposure to varying concentrations of TE.This phenolic concentration-dependent alteration in membranelipid compositions and compromise in cell integrity were the possi-ble mechanism for the bactericidal effect of TE on BL5089. The com-bination of dietary phenolic concentrate and probiotics may open anew field of functional food development, while the influence ofphenolic compounds at certain concentrations on those bacteriashould be evaluated in order to prevent the undesired bactericidaleffects.
Acknowledgments
The authors are thankful to Mr. Victor Yeung and Mr. An Ke in theSchool of Biological Sciences, the University of Hong Kong, for theirtechnical assistance and support. The kind supply of soy protein isolateby Solae DuPont China Holding Co., Ltd. (Shanghai, China) is alsoacknowledged.
References
Amor, K.B.,Breeuwer, P.,Verbaarschot, P.,Rombouts, F.M.,Akkermans, A.D.L.,De Vos, W.M.,Abee, T., 2002. Multiparametric flow cytometry and cell sorting for the assessment ofviable, injured, and dead Bifidobacterium cells during bile salt stress. Appl. Environ.Microbiol. 68, 5209–5216.
Ananta, E., Heinz, V., Knorr, D., 2004. Assessment of high pressure induced damage onLactobacillus rhamnosus GG by flow cytometry. Food Microbiol. 21, 567–577.
Ankolekar, C., Johnson, D., Pinto, M.S., Johnson, K., Labbe, R., Shetty, K., 2011. Inhibitorypotential of tea polyphenolics and influence of extraction time against helicobacterpylori and lack of inhibition of beneficial lactic acid bacteria. J. Med. Food 14,1321–1329.
Aricha, B.,Fishov, I.,Cohen, Z.,Sikron, N.,Pesakhov, S.,Khozin-Goldberg, I.,Dagan, R.,Porat,N., 2004. Differences in membrane fluidity and fatty acid composition betweenphenotypic variants of Streptococcus pneumonia. J. Bacteriol. 186, 4638–4644.
Chan, E.W.C.,Soh, E.Y.,Tie, P.P.,Law, Y.P., 2011. Antioxidant and antibacterial properties ofgreen, black, and herbal teas of Camellia sinensis. Pharmacogn. Res. 3, 266–272.
Denich, T.J.,Beaudette, L.A.,Lee, H.,Trevors, J.T., 2003. Effect of selected environmental andphysico-chemical factors on bacterial cytoplasmic membranes. J. Microbiol. Methods52, 149–182.
Dickman, S.R., Proctor, C.M., 1952. Factors affecting the activity of egg white lysozyme.Arch. Biochem. Biophys. 40, 364–372.
Doherty, S.B.,Wang, L.,Ross, R.P.,Stanton, C.,Fitzgerald, G.F.,Brodkorb, A., 2010. Use of vi-ability staining in combination with flow cytometry for rapid viability assessmentofLactobacillus rhamnosusGG in complex protein matrices. Journal of MicrobiologyMethods 82, 301–310.
Dong, L., Tong, Z., Linghu, D., Lin, Y., Tao, R., Liu, J., Tian, Y., Ni, L., 2012. Effects of sub-minimum inhibitory concentrations of antimicrobial agents on Streptococcus mutansbiofilm formation. Int. J. Antimicrob. Agents 39, 390–395.
Donkor, O.N.,Shah, N.P., 2007. Production of β-glucosidase and hydrolysis of isoflavonephytoestrogens by Lactobacillus acidophilus, Bifidobacterium lactis, and Lactobacilluscasei in soymilk. J. Food Sci. 73, M15–M20.
Filgueiras, M.H.,den Kamp, J.A.F., 1980. Cardiolipin, a major phospholipid of gram-positivebacteria that is not readily extractable. Biochim. Biophys. Acta 620, 332–337.
Folch, J., Lees, M., Sloane-Stanley, G.H., 1957. A simple method for the isolation andpurification of total lipids from animal tissues. J. Biol. Chem. 226, 497–509.
Gaudreau, H., Champagne, C.P.,Remondetto, G.E.,Bazinet, L., Subirade, M., 2013. Effect ofcatechins on the growth of oxygen-sensitive probiotic bacteria. Food Res. Int. 53,751–757.
Guerzoni, M.E., Lanciotti, R., Cocconcelli, P.S., 2001. Alteration in cellular fatty acidcomposition as a response to salt, acid, oxidative and thermal stresses in Lactobacillushelveticus. Microbiology 147, 2255–2264.
Hashimoto, T.,Kumazawa, S.,Nanjo, F.,Hara, Y.,Nakayama, T., 1999. Interaction of tea cat-echins with lipid bilayers investigated with liposome systems. Biosci. Biotechnol.Biochem. 63, 2252–2255.
Ikigai, H.,Nakae, T.,Hara, Y., Shimamura, T., 1993. Bactericidal catechins damage the lipidbilayer. Biochim. Biophys. Acta 1147, 132–136.
Jaziri, I.,Ben Slama,M.,Mhadhbi, H.,Urdaci, M.C.,Hamdi, M., 2009. Effect of green and blackteas (Camellia sinensis L.) on the characteristic microflora of yogurt during fermenta-tion and refrigerated storage. Food Chem 112, 614–620.
Joint FAO/WHOWorking Group, 2002. Guidelines for the Evaluation of Probiotics in Food.Food and Agriculture Organization of the United Nations and World HealthOrganization Working Group Report, London, Ontario (ftp://ftp.fao.org/es/esn/food/wgreport2.pdf).
Joux, F., Lebaron, P., 2000. Use of fluorescent probes to assess physiological functions ofbacteria at single-cell level. Microbes Infect. 2, 1523–1535.
Khan, N.,Mukhtar, H., 2007. Tea polyphenols for health promotion. Life Sci. 81, 519–533.Kimoto-Nira, H.,Kobayashi, M.,Nomura, M., Sasaki, K., Suzuki, C., 2009. Bile resistance in
Lactococcus lactis strains varies with cellular fatty acid composition: analysis byusing different growth media. Int. J. Food Microbiol. 143, 226–229.
Kumazawa, S.,Kajiya, K.,Naito, A.,Saito, H.,Tuzi, S.,Tanio, M.,Suzuki, M.,Nanjo, F.,Suzuki, E.,Nakayama, T., 2004. Direct evidence of interaction of a green tea polyphenol, epi-gallocatechin gallate, with lipid bilayers by solid-state nuclear magnetic resonance.Biosci. Biotechnol. Biochem. 68, 1743–1747.
Lee, H.C.,Jenner, A.M.,Lowa, C.S.,Lee, Y.K., 2006. Effect of tea phenolics and their aromaticfecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 157, 876–884.
Lewis, R.N.,McElhaney, R.N., 2000. Surface charge markedly attenuates the nonlamellarphase-forming propensities of lipid bilayer membranes: calorimetric and (31)P-nuclearmagnetic resonance studies ofmixtures of cationic, anionic, and zwitterioniclipids. Biophys. J. 79, 1455–1464.
Li, S., Lo, C.Y., Pan, M.H., Lai, C.S.,Ho, C.T., 2013. Black tea: chemical analysis and stability.Food Funct. 4, 10–18.
Limonet, M., Saffroy, S.,Maujean, F., Linder, M.,Delaunay, S., 2007. A comparison ofdisruption procedures for the analysis of phospholipids from Streptomycespristinaespiralis. Process Biochem. 42, 700–703.

44 D. Zhao, N.P. Shah / International Journal of Food Microbiology 188 (2014) 36–44
Lopez, C.S., Alice, A.F., Heras, H., Rivas, E.A., Sanchez-Rivas, C., 2006. Role of anionicphospholipids in the adaptation of Bacillus subtilis to high salinity. Microbiology152, 605–616.
Microbial ID, 1990. Identification of bacteria by gas chromatography of cellular fatty acids.Technical Note, 101. Midi, Inc., Newark, DE, U.S.A.
Mizunoe, Y.,Wai, S.N.,Ishikawa, T.,Takade, A.,Yoshida, S., 2000. Resuscitation of viable butnonculturable cells of Vibrio parahaemolyticus induced at low temperature understarvation. FEMS Microb. Lett. 186, 115–120.
Murga, M.L.F., Bernik, D., de Valdez, G.F., Disalvo, A.E., 1999. Permeability and stabilityproperties of membranes formed by lipids extracted from Lactobacillus acidophilusgrown at different temperatures. Arch. Biochem. Biophys. 364, 115–121.
Najgebauer-Lejko, D., Sady, M., Grega, T., Walczycka, M., 2011. The impact of teasupplementation on microflora, pH and antioxidant capacity of yoghurt. Int. Dairy J.21, 568–574.
Nakayama, M.,Shigemune, N.,Tsugukuni, T., Jun, H.,Matsushita, T.,Mekada, Y.,Kurahachi,M.,Miyamoto, T., 2012. Mechanism of the combined anti-bacterial effect of green teaextract and NaCl against Staphylococcus aureus and Escherichia coli O157:H7. FoodControl 25, 225–232.
Novik, G.I.,Astapovich, N.I.,Grzegorzewicz, A.,Gamian, A., 2006. Analysis of phospholipidsfrom bifidobacteria. Microbiology 75, 40–45.
Nzai, J.M., Proctor, A., 1998. Phospholipids determination in vegetable oil by thin-layerchromatography and imaging densitometry. Food Chem. 63, 571–576.
Özbalci, C., Sachsenheimer, T.,Brügger, B., 2013. Quantitative analysis of cellular lipids bynano-electrospray ionization mass spectrometry. In: Rapaport, D., Herrmann, J.M.(Eds.), Membrane Biogenesis: Methods and Protocols. Methods in Molecular Biology,1033, pp. 3–20.
Parkar, S.G., Stevenson, D.E., Skinner, M.A., 2008. The potential influence of fruitpolyphenols on colonic microflora and human gut health. Int. J. Food Microbiol.124, 295–298.
Perumalla, A.V.S.,Hettiarachchy, N.S., 2011. Green tea and grape seed extracts—potentialapplications in food safety and quality. Food Res. Int. 44, 827–839.
Picard, C.,Fioramonti, J.,Francois, A.,Robinson, T.,Neant, F.,Matuchansky, C., 2005. Reviewarticle: bifidobacteria as probiotic agents—physiological effects and clinical benefits.Aliment. Pharmacol. Ther. 22, 495–512.
Puupponen-Pimiä, R., Nohynek, L., Meier, C., Kähkonen, M., Heinonen, M., Hopia, A.,Oksman-Caldentey, K.M., 2001. Antimicrobial properties of phenolic compoundsfrom berries. J. Appl. Microbiol. (90), 494–507.
Rakhuba, D.,Novik, G.,Dey, E.S., 2009. Application of supercritical carbon dioxide (scCO2)for the extraction of glycolipids from Lactobacillus plantarum B-01. J. Supercrit. Fluids49, 45–51.
Sanders, M.E., 2000. Considerations for use of probiotic bacteria to modulate humanhealth. J. Nutr. 130, 384S–390S.
Shimamura, T.,Zhao, W.H.,Hu, Z., 2007. Mechanism of action and potential for use of teacatechin as an anti-infective agent. Anti-Infect. Agents Med. Chem. 6, 57–62.
Sivarooban, T.,Hettiarachchy, N.S.,Johnson, M.G., 2008. Transmission electron microscopystudy of Listeria monocytogenes treated with nisin in combination with either grapeseed or green tea extract. J. Food Prot. 71, 2105–2109.
Steponkus, P.L., Uemura, M.,Webb, M.S., 1995. In: Disalvo, E.A., Simon, A.S. (Eds.),Permeability and Stability of Lipid Bilayers. CRC Press, Boca Raton, FL, pp. 77–104.
Tabasco, R., Sánchez-Patán, F.,Monagas, M., Bartolomé, B.,Moreno-Arribas, M.V., 2011.Effect of grape polyphenols on lactic acid bacteria and bifidobacteria growth:resistance and metabolism. Food Microbiol. 28, 1345–1352.
Tsuchiya, H., 1999. Effects of green tea catechins onmembrane fluidity. Pharmacology 59,34–44.
Tzonuis, X., Vulevic, J., Kuhnle, G.G., George, T., Leonczak, J., Gibson, G.R., et al., 2008.Flavanol monomer-induced changes to the human fecal microflora. Br. J. Nutr. 99,782–792.
Wang, Y.,Delettre, J.,Guillot, A.,Corrieu, G.,Béal, C., 2005. Influence of cooling temperatureand duration on cold adaptation of Lactobacillus acidophilus RD758. Cryobiology 50,294–307.
Wu, V.C.H., 2008. A review of microbial injury and recovery methods in food. FoodMicrobiol. 25, 735–744.
Wu, C.,Zhang, J.,Wang, M.,Du, G.,Chen, J., 2012. Lactobacillus casei combats acid stress bymaintaining cell membrane functionality. J. Ind. Microbiol. Biotechnol. 39,1031–1039.
Yam, T.S.,Shah, S.,Hamilton-Miller, J.M.T., 1997.Microbiological activity of whole and frac-tionated crude extracts of tea (Camellia sinensis), and of tea components. FEMS Mi-crobiology Letters 152, 169–174.
Yi, S., Zhu, J., Fu, L., Li, J., 2010. Tea polyphenols inhibit Pseudomonas aeruginosa throughdamage to the cell membrane. Int. J. Food Microbiol. 144, 111–117.
Zhao, D., Shah, N.P., 2014. Antiradical and tea polyphenol-stabilizing ability of functionalfermented soymilk-tea beverage. Food Chem. 158, 262–269.