improved translation efficiency of injected mrna during early embryonic development
TRANSCRIPT

TECHNIQUES
Improved Translation Efficiency of InjectedmRNA During Early Embryonic DevelopmentMaria Fink, Gabriele Flekna,† Alfred Ludwig, Thomas Heimbucher, and Thomas Czerny*
Injection techniques are a powerful approach to study gene function in fish and frog model systems. Inparticular, in vitro transcribed mRNA is broadly used for such misexpression experiments. Sequenceelements flanking the coding region, such as untranslated repeats and polyadenylation sequences, areknown to affect the stability and the translation efficiency of mRNA. Here we show that in early embryos,poly(A) signals strongly contribute to the activity of the injected mRNA. Of interest, they only marginallyaffect mRNA stability, whereas the translation efficiency is dramatically enhanced. Combination of apoly(A) tail and an SV40 late poly(A) signal leads to highly synergistic effects of the two elements forinjected mRNA. Compared with established vector systems, we detected a 20-fold improvement for mRNAderived from the novel transcription vector pMC. Developmental Dynamics 235:3370–3378, 2006.© 2006 Wiley-Liss, Inc.
Key words: mRNA injection; UTR; �-globin; SV40 late poly(A); T7TS; pCS2; pMC
Accepted 18 September 2006
INTRODUCTION
Ectopic gene expression is an impor-tant method to study gene function,complementing the analysis of mu-tants. It allows investigators to testfor sufficiency of gene action and,through the application of antimor-phic molecules, also for loss-of-func-tion experiments. In fish and frogmodel systems, transient misexpres-sion techniques based on microinjec-tion have been developed. mRNA in-jected at the one or two-cell stage isreadily distributed and translatedwithin the embryo and allows the ex-pression of products even before mid-blastula transition (Vize et al., 1991;Hyatt and Ekker, 1999). This tech-nique is, therefore, ideal to analyze
effects of misexpression on early de-velopment of the embryos (Harlandand Misher, 1988).
The stability and the translationefficiency of mRNA represents animportant control of gene expression(reviewed in Wilusz and Wilusz,2004). During processing, a 7-methylguanosine cap-structure and a poly(A)tail are added to the transcript in thenucleus. After nuclear export, themRNA molecule is present in the cy-toplasm as a circular complex, due tointeractions between poly(A) bindingproteins and the elF4F-cap complex(Sachs et al., 1997; Mazumder et al.,2003). Bridging the mRNA termini en-hances the recruitment of new ribo-somal subunits by elF4F, resulting in
enhanced translation. In addition,this complex inhibits the action of thedecapping enzyme, which destabilizesmRNAs by removing the cap (Dehlinet al., 2000; Pickering and Willis,2005). Furthermore, poly(A) se-quences block mRNA exonucleases(Ford et al., 1997). The poly(A) tail,therefore, represents a key factor forboth mRNA stabilization as well astranslation efficiency (de Moor et al.,2005). Cytoplasmic activities thatmodify the length of this tail, there-fore, critically influence gene expres-sion. Both cytoplasmic deadenylationas well as cytoplasmic adenylation actupon mRNA molecules and are knownto be regulated during embryonic de-velopment (Huarte et al., 1992; Rich-
The Supplementary Material referred to in this article is available at http://www.interscience.wiley.com/jpages/1058-8388/suppmatInstitute of Animal Breeding and Genetics, University of Veterinary Medicine, Vienna, AustriaGrant sponsor: Austrian Science Fund (FWF); Grant number: P13138; Grant number: P15185.†Dr. Flekna’s present address is Institute of Meat Hygiene, Meat Technology and Food Science, University of Veterinary Medicine,Veterinarplatz 1, A-1210 Vienna, Austria.*Correspondence to: Thomas Czerny, Institute of Animal Breeding and Genetics, University of Veterinary Medicine, Veteri-narplatz 1, A-1210 Vienna, Austria. E-mail: [email protected]
DOI 10.1002/dvdy.20995Published online 26 October 2006 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 235:3370–3378, 2006
© 2006 Wiley-Liss, Inc.

ter, 1999; de Moor et al., 2005). Dead-enylation is the default pathway formRNA molecules in the cytoplasm.Stepwise reduction of the poly(A) tailup to a certain limit eventually endsin a rapid degradation by exonucle-ases (Copeland and Wormington,2001; Wilusz and Wilusz, 2004). Thedecay rate of mRNA is further regu-lated by AU-rich sequence elementsmainly located in the 3� untranslatedregion (UTR; de Moor et al., 2005).
The UTRs can influence both thestability and the translatability of themRNA. Untranslated AUGs and sta-ble secondary structures in the 5�UTR affect translation initiation(Pickering and Willis, 2005), and se-quence-specific binding of proteins toelements in both 5� and 3� UTRs canaffect degradation, polyadenylation,and mRNA localization (Wickens etal., 1997; Kloc et al., 2002). Especiallyduring embryonic development, thereis a wealth of examples of 3� UTRs andpolyadenylation events influencingtranslation efficiency (Seydoux, 1996;Kuersten and Goodwin, 2003).
During early development, mater-nal mRNAs have to direct develop-ment before the onset of zygotic tran-
scription. A mechanism has, therefore,evolved that translationally masks apool of maternal mRNAs duringoozyte maturation, which are then ac-tivated during early development(Paillard and Osborne, 2003). Inacti-vation involves deadenylation of themRNAs, whereas cytoplasmic adeny-lation reactivates the dormant tran-scripts (Richter, 1999). Two discretesignals in the 3� UTR are necessaryfor this process, the nuclear polyade-nylation site AAUAAA and a U-richcytoplasmic polyadenylation element(CPE) with a consensus sequence ofUUUUUAU (Wormington, 1993). Thesequence of the CPE and its positionrelative to AAUAAA may control thetiming and extent of polyadenylation(Richter, 1999).
We tested different UTRs andpoly(A) sequences for their efficiencyin mRNA injection experiments. Sur-prisingly variation of the UTRs hadlittle effect, contrary to the poly(A) se-quences. Both artificial poly(A)stretches as well as the SV40 latepoly(A) signal strongly improvedtranslation activity, but not the stabil-ity of the RNA. Combined applicationof these elements in a new transcrip-
tion vector resulted in highly syner-gistic activity of the elements for in-jected mRNA in early embryos.
RESULTS
Pax2 UTRs Reduce theActivity of Injected mRNA
The Pax2 gene is a key player in mid/hindbrain development and conse-quently leads to strong phenotypeswhen misexpressed in early embryos(Kelly and Moon, 1995; Okafuji et al.,1999; Ye et al., 2001). We tried to re-peat these experiments in medakafish by injecting in vitro transcribedcapped mRNA of zebrafish Pax2 intoone-cell stage embryos. Contrary toour expectations, we did not observeany phenotypes in the mid/hindbrainregion (data not shown). When weraised the amount of injected mRNA,increasing numbers of embryos werearrested during gastrulation, but thesurviving embryos still appeared un-affected during somitogenesis. We as-sumed, that rapid degradation of theinjected mRNA in combination withthe slow development of medaka couldcause these unexpected results. Be-cause mRNA stability is mainly regu-lated by untranslated regions (UTR),we tested the effects of the Pax2 UTRsin a construct containing the fireflyluciferase reporter gene flanked bythese UTRs (Fig. 1; Pax2 luc). Wemaintained the endogenous transla-tion start signals of Pax2 by fusing theluciferase coding region directly to theATG of the Pax2 cDNA. In vitro tran-scribed mRNA of this construct wasthen injected into medaka embryos.Luciferase activity measurements af-ter 24 hr revealed sevenfold reducedvalues when compared with a refer-ence group of embryos injected withmRNA containing luciferase togetherwith artificial UTRs derived from thecell culture expression vector pKW10(construct “KW luc”, Table 1; Fig. 1).At 48 hr post-fertilization (hpf), thedifference in luciferase activity be-tween the two constructs was evenmore pronounced (38-fold, Table 1).Because the context of the start codonfor both constructs was similar(CCCATGG for Pax2 luc and AC-CATGG for KW luc), these data sug-gest a destabilizing effect of the Pax2UTR sequences. These results were
Fig. 1. Schematic presentation of the injected mRNAs. All mRNAs contain the firefly luciferasecDNA. KW luc, CS luc, and CSas luc mRNA contain artificial untranslated regions (UTRs) derivedfrom pKW10 (KWluc) and pCS2 (CS luc and CSas luc). The 5� and the 3� UTRs of the zebrafish Pax2gene are part of Pax2 luc mRNA and the Xenopus �-globin UTRs of T7TS luc, T7TS luc pA, and MCluc. Except for T7TS luc, all mRNAs contain either the early (e-pA) or the late (l-pA) SV40polyadenylation signal. In addition, T7TS luc, T7TS luc pA, and MC luc mRNAs contain an artificialA30 poly(A) sequence (AAAA).
IMPROVED TRANSLATION EFFICIENCY OF mRNA 3371

further supported by cell culture ex-periments. The two constructs, bothcontaining a CMV promoter, wereused for transient DNA transfection ofNIH 3T3, HeLa, and U2-OS cells. Theexperiments revealed a 10- to 25-foldreduced luciferase activity for thePax2 UTR-containing construct, de-pending on the cell line used (Table 1).Therefore, Pax2 UTR sequences nega-tively influence mRNA activity, possi-bly by enhancing the degradation rateboth in embryonic and in cell culturecells.
Contrary to Pax2, UTRs of othergenes have been shown to increase theactivity of injected mRNA. The �-glo-bin UTRs present in the pT7TS con-struct are widely used for misexpres-sion experiments (Hyatt and Ekker,1999). Indeed when we applied thesesequences we could improve markergene expression compared with thepKW10-derived mRNA (Table 1; T7TSluc pA), but the effects were only two-to fourfold. In cell culture experimentsaddition of the �-globin sequences af-fected the luciferase activity twofoldat most, both positively and nega-tively, depending on the cell line (Ta-ble 1).
SV40 PolyadenylationSignals Strongly AffectmRNA Activity in EarlyEmbryos
The pCS2 vector (Rupp et al., 1994;Turner and Weintraub, 1994) repre-sents another backbone frequentlyused for misexpression experiments.Of interest, we observed an almost 20-fold increase in luciferase activity formRNA derived from this constructcompared with the KW luc reference(Table 2). The pCS2 vector containsartificial UTRs and an SV40 poly(A)signal. Such a signal is also present inthe T7TS luc pA construct and could,therefore, not explain the dramatic in-crease of reporter gene activity forpCS2. However, we realized that thetwo constructs differ in the orienta-tion of the inserted SV40 fragment,which contains poly(A) signals in bothorientations. Thus in pCS2, the latepoly(A) signal is used, whereas inT7TS luc pA, the oppositely orientedearly poly(A) signal of the SV40 virusis used. To test whether one of the twopoly(A) signals preferentially affects
mRNA activity, we generated a pCSconstruct with a reversed poly(A) se-quence. mRNA derived from this con-struct (CSas luc, Table 2) indeed re-sulted in strongly reduced luciferaseactivities, comparable to that forT7TS luc pA. Of interest, these manip-ulations of the expression vectors didnot affect luciferase activity in cell cul-ture cells (Table 2), indicating that theeffects of the poly(A) signals are re-stricted to embryonic development.Therefore, viral SV40 poly(A) se-quences specifically affect the activityof injected mRNA in early embryos.
Combined SequenceElements SynergisticallyAffect mRNA Activity inEarly Embryos
Because both pT7TS, as well as pCS2vectors, positively affected the activityof injected mRNA, we tested a combi-nation of their elements. The result-
ing vector pMC contains the backboneof pCS2, including the CMV promoterand the SV40 poly(A) signal and the 5�and 3� UTRs of the �-globin gene to-gether with a poly(A) tail of the pT7TSconstruct. When we tested mRNA de-rived from this construct, we sawstrong synergistic effects of the com-bined sequences. Compared with theisolated elements in pT7TS or pCS2,pMC-derived mRNA resulted in al-most 20-fold higher luciferase activityin the embryos (Table 3). Again thecorresponding cell culture experi-ments did not reveal similar effects.
The results for all constructs arecompared in Figure 2. In cell culture(mean values for all three cell lineswere calculated), �-globin UTRs andthe orientation of the SV40 poly(A)sequences showed almost no effects,only the Pax2 UTRs significantly re-duced luciferase activity. Contrary tocell culture cells, the different ele-ments strongly affected the results for
TABLE 1. Pax2 UTRs Reduce the Activity of Injected mRNAa
Construct
Embryo Cell culture
24 hpf 48 hpf NIH HeLa U2-OS
KW luc 100 (9) 205 (3) 100 100 100100 (13)
Pax2 luc 8.8 (9) 5.4 (4) 10.0 4.0 4.616 (16)
T7TS luc pA 378 (7) 298 (7) 202 58 101201 (9)
aThe data represent relative luciferase activities of the indicated constructs obtainedafter mRNA injection into medaka embryos or DNA transfection into the indicated celllines. Values for the “KW luc” construct were set at 100. The numbers of the injectedembryos are given in brackets. Two series of injections were performed, each includingall three mRNAs. The results of the two series are presented in the first and secondrows, respectively. The cell culture results are based on four independent transfectionsexperiments. UTRs, untranslated regions; hpf, hours postfertilization
TABLE 2. Orientation of the SV40 Sequences Affects the Activity ofInjected mRNAa
Construct
Embryo Cell culture
24 hpf NIH HeLa U2-OS
KW luc 100 (11) 100 100 100100 (17)
CSas luc 233 (10) 198 54 98149 (14)
CS luc 1944 (10) 110 46 1021859 (17)
aFor details, see Table 1.
3372 FINK ET AL.

mRNA injections in embryos. ThuspMC-derived mRNA was 385-foldmore active than KW luc mRNA, andcompared with the established tran-scription vector pCS2 and pT7TS, ledto a 20-fold improvement in earlymedaka embryos.
The cell culture experiments sug-gested that the activating potential ofpMC might be restricted to embryonicdevelopment. To compare differentconstructs during later stages of de-velopment, we performed in ovo elec-troporation of chick embryos. With
this technique, we transferred DNAinto neural tube cells during somito-genesis (Hamburger and Hamiltonstage [HH] 12–14, 16–20 somites). Wetested KW luc, CS luc, and MC lucconstructs, all containing a CMV pro-moter. Experimental variations werecorrected by coelectroporation of a Re-nilla luciferase reference construct.The electroporated embryos were cul-tured for 24 hr (HH18–20), dissected,and lysed to measure luciferase activ-ity. The corrected values did not showany beneficial effects for �-globin or
SV40 late poly(A) sequences. MC lucvalues were slightly below that for CSluc and for KW luc (1.6-fold and 1.5-fold,respectively). The improved expressionof pMC-derived mRNA, therefore,seems to be restricted to the earlieststages of embryonic development.
�-globin and SV40 SequencesOnly Marginally AffectmRNA Stability
The simplest way to mechanisticallyexplain the enhancing effects of the�-globin and poly(A) sequences wouldbe a prolonged stability of the injectedmRNA. To test this hypothesis, weperformed a kinetic study, where wemeasured the relative amount ofmRNA in the embryos directly afterinjection (reference value at 0 hpf) andat 6, 24, and 48 hpf. As endogenousreference, we selected the house-keep-ing gene �-actin (Takagi et al., 1994;Nakaniwa et al., 2005). To correct forinjection inconsistencies, we lysed 10medaka embryos for each time pointand isolated the total RNA. After re-verse transcription, real-time poly-merase chain reaction (PCR) was car-ried out by performing two differentassays, one for quantifying the lucif-erase and another one for quantifyingendogenous �-actin mRNA. Quantify-ing all RNA samples (n � 21), we de-termined constant levels of maternal�-actin mRNA (0 and 6 hpf) and 19.3-and 39.7-fold higher amounts in theembryos at 24 and 48 hpf, respec-tively. After having defined the abso-lute levels for this reference gene ateach stage, we used it to determinethe relative luc mRNA concentrationwithin each individual sample. Thecorrected values for the injected MCluc, CS luc, and KW luc mRNAs weresubsequently used to calculate themRNA decay rates (Fig. 3). Assuminga constant decay rate, the half-life forKW luc mRNA was 2.9 hr, that forpCS2 was 3.4 hr, and that for pMCwas 3.5 hr. These calculations give av-erage values covering the first 48 hr ofdevelopment. Close inspection of thedata points, however, indicates somedeviations from a linear relationship(almost all data points at 6 hpf areabove, those for 24 hpf are below thelines). During the earliest stages untilmid-blastula transition (at 8 hpf inmedaka; Aizawa et al., 2003) mRNA
Fig. 2. Summary of mRNA injection and DNA transfection experiments. Relative luciferase activ-ities for mRNA injection experiments into early medaka embryos (embryo) were calculated fromTables 1 to 3, relative to MC luc mRNA, which was set at 100. Likewise the relative luciferaseactivities of the cell culture experiments (cell culture) were determined; here mean values for thethree cell lines were calculated. The error bars shown indicate the standard deviation.
TABLE 3. Combined Effects of T7TS- and pCS2-DerivedSequence Elements
Construct
Embryo Cell culture
24 hpf NIH HeLa U2-OS
MC luc 100 (13) 100 100 100100 (12)
T7TS luc 4.5 (5) - - -7.0 (10)
CS luc 5.7 (9) 96 50 1004.4 (6)
aThe data represent relative luciferase activities of the indicated constructs obtainedafter mRNA injection into medaka embryos or DNA transfection into the indicated celllines. Values for the “MCluc” construct were set at 100. The numbers of the injectedembryos are given in brackets. Two series of injections were performed, each includingall three mRNAs. The results of the two series are presented in the first and secondrows, respectively. The cell culture results are based on four independent transfectionsexperiments. The T7TS construct lacks a promoter and was therefore excluded fromthe cell culture experiments. hpf, hours past fertilization.
IMPROVED TRANSLATION EFFICIENCY OF mRNA 3373

indeed has been reported to be highlystable in Xenopus (Vize et al., 1991).The half-life measured for injectedmRNA after this stage was 2.5 hr forfrog embryos (Harland and Misher,1988), which is similar to the valueswe determined for medaka. The deg-radation rates for the three mRNAsfollowed the same order as their lucif-erase activities after injection, withpMC being most active. However, thevariations are marginal and cannotexplain the dramatic differences inmarker gene activity (385-fold differ-ence in luciferase activity between MCluc and KW luc mRNAs). Therefore,mRNA stability can be excluded as amolecular mechanism to explain thebeneficial effects of the transcriptionvectors.
Injected mRNA Is DegradedWith Similar Rates asEndogenous mRNA
Injected RNA differs in several as-pects from natural mRNA. At its 5�end, an artificial cap has to be inte-grated, which in vivo is added duringnuclear processing of the mRNA. Inaddition, it lacks a terminal poly(A)sequence. We, therefore, asked thequestion whether this artificial RNAdiffers in its half-life from in vivo pro-cessed mRNA. For this purpose, weused a heat shock transgenic line ex-pressing the luciferase coding region,which is flanked by exactly the samesequences present in the pMC vector(Bajoghli et al., 2004). This artificialHSE promoter produces a burst ofmRNA during heat shock treatment,which then diminishes according to itsnatural degradation rate. Quantifica-tion of the mRNA levels was per-formed using the same real-time re-verse transcriptase-PCR (RT-PCR)strategy as before (in this case individ-ual embryos were quantified). The re-sulting decay rate (3.5 hr half-life)was identical to that calculated formRNA injection (Fig. 3). The timewindow used for determination oftheir half-life differed for the two mR-NAs, nevertheless our data indicatethat natural and artificial mRNAs inthe early embryo are degraded atroughly the same rate.
DISCUSSION
Stability of the InjectedmRNA
During embryonic development, regu-latory genes typically are active onlyduring small time windows of compe-tence. This transient function ex-plains the reduced stability of regula-tory proteins, which has to becomplemented by a short half-life ofthe corresponding mRNAs. We thusdetected a reduced activity of the Pax2mRNA in our experiments, mediatedby its 5� and 3� UTRs. The critical roleof the UTRs in the stability of injectedmRNA first became clear when a longhalf-life for �-globin mRNA was de-tected in Xenopus embryos (Gurdon etal., 1974). The application of heterolo-gous UTRs was, therefore, selected asa general strategy for injection exper-iments. Two vector systems were de-veloped for this purpose, which arebroadly used, pT7TS and pCS2. ThepT7TS vector (A. Johnson and P.Krieg, unpublished) is based on �-glo-bin sequences inserted into pGEM4Z
and an artificial poly(A) sequence,consisting of 30 A residues. The pCS2vector (Rupp et al., 1994; Turner andWeintraub, 1994) contains a CMVpromoter, artificial UTRs, and theSV40 late poly(A) signal. It can, there-fore, also be used for DNA injectionexperiments. The artificial UTRs lackdestabilizing elements and conse-quently behave neutral in the embryo(van der Velden et al., 2001). We es-tablished an assay system based onthe firefly luciferase gene, to quantita-tively determine protein activities ininjected embryos. Using this systemfor medaka fish, we indeed observedbeneficial effects for both transcrip-tion vector systems. pT7TS and pCS2derived mRNA resulted in more than100-fold higher luciferase activitycompared with mRNA containing theendogenous UTRs of the Pax2 gene(Tables 1–3; Fig. 2).
The mRNA stabilizing functions ofthe heterologous UTRs were the basicidea for construction of these two vec-tor systems (Vize et al., 1991; Hyattand Ekker, 1999). Attempts to com-
Fig. 3. Decay rates of luciferase mRNAs in early embryos. The indicated mRNAs (MC luc, CS luc,and KW luc) were injected at the one-cell stage into medaka embryos and then quantified by realtime reverse transcriptase-polymerase chain reaction (RT-PCR). mRNA levels at 0 hpf (directly afterinjection) were set at 100, all other values were calculated relative to this level. The decay rates weredetermined for each mRNA [0.237 h�1 for KW luc mRNA (y � 100e�0.237x), 0.205 h�1 for CS luc,and 0.198 h�1 for MC luc] by assuming a linear relationship between 0 and 48 hours postfertilization(hpf). Similarly luciferase mRNA levels were quantified after heat shock activation of a stablyintegrated transgene in medaka embryos (HSE luc). The RNA level directly after heat shock was setat 100 (the time of the heat shock is indicated by an arrow). The decay rate for this endogenouslyprocessed mRNA was calculated using 0.199 h�1, which is almost identical to that of the injectedpMC.
3374 FINK ET AL.

bine the positive effects of the two vec-tor systems, therefore, concentratedon the UTRs. A combination of thepT7TS-derived �-globin UTRs withthe pCS2 backbone nevertheless re-sulted in decreased activity of the in-jected mRNA (van der Velden et al.,2001), thus indicating that the effectsof the UTRs are not additive. We com-pared mRNA derived from pT7TS andpCS2 to that of the typical cell cultureexpression vector pKW10 (Adams etal., 1992), which has an architecturesimilar to pCS2 [CMV promoter, arti-ficial UTRs, and SV40 early poly(A)signal]. Both transcription vectorspositively affected the activity of in-jected mRNA in early embryos (Fig. 2;24-fold and 19-fold, respectively), butwhen we provided the three con-structs with identical poly(A) signals,the mRNA of the two specialized tran-scription vectors was only threefold(pT7TS luc pA) or twofold (pCSas luc)more active compared with thepKW10 vector. The improvementsseen for the �-globin UTRs, therefore,were weak compared with the strongeffects published both for cell culturecells (Malone et al., 1989) as well asfor early embryos (Vize et al., 1991).However, in these experiments, the�-globin UTRs were compared withmRNAs lacking UTRs at all. ArtificialUTRs, as for example used in thepKW10 vector, therefore, provide al-most the same efficiency as the �-glo-bin sequences. The improved lucif-erase activity we observed for thetranscription vectors has to be attrib-uted mainly to elements outside theUTRs.
When we quantified the injectedmRNA in the embryos over the timeby real-time PCR, we observed onlyminor differences between the vectors.After 24 hr, pCS2-derived mRNA waspresent in the embryos at a 2.2-foldhigher concentration compared withthat for pKW10 mRNA (calculatedfrom the decay rates presented in Fig.3), which, however, cannot explain a19-fold higher luciferase activity forthis mRNA measured at the sametime (Table 2). For the chimeric pMCvector, a 2.6-fold higher mRNA levelwas determined, contrasted by a 385-fold higher luciferase activity (Fig. 3;Tables 2, 3). Constructs that lackUTRs and poly(A) sequences com-pletely, produce unstable mRNA,
which is rapidly degraded in the em-bryos starting from mid-blastula tran-sition (Richter, 1999). However, com-pared with mRNA containingartificial UTRs, pT7TS and pCS2 donot substantially improve the stabilityof injected mRNA in medaka embryos.
Poly(A) Signals StronglyAffect the Activity ofInjected mRNA
Efficient polyadenylation at the 3� endof the mRNA is considered to be im-portant for both mRNA stability andtranslation efficiency. Initial polyade-nylation is part of the mRNA process-ing in the nucleus (Zhao et al., 1999).Injected mRNA is not processed, andconsequently, artificial A tails havebeen added to the in vitro transcribedmRNA (Vize et al., 1991). One strat-egy is to include this tail into the tran-scription vector, which has been donefor the pT7TS by adding 30 A resi-dues. Such elements indeed stronglyaffect the stability of injected mRNAin the embryo (Vize et al., 1991). Fur-ther elongation of the tail to 75 A res-idues does not enhance this effect (vander Velden et al., 2001). pCS2 lackssuch an artificial poly(A) tail, never-theless mRNA derived from this vec-tor is not degraded in the embryo. Thedetermined half-life was 3.4 hr, whichwas similar to that for pMC derivedmRNA, containing the A30 sequence ofpT7TS (3.5 hr). Furthermore, the sta-bility of pCS2 mRNA was in the samerange as for mRNA produced from anintegrated transgene, where a poly(A)tail is added during processing withinthe nucleus (3.5 hr; Fig. 3).
One possible explanation for the ac-tivity of pCS2-derived mRNA wouldbe cytoplasmic polyadenylation. Thisprocess is active in early embryos andis known to be regulated by U-richsequence elements cooperating with apoly(A) signal (Wormington, 1993;Richter, 1999). Our poly(A) replace-ment experiments (CS luc vs. CSasluc) indicate, that a fragment of 198nucleotides of the SV40 genome (2534-2731), including the late poly(A) sig-nal, mediates the positive effects onmRNA activity. Contrary to this find-ing, the SV40 early poly(A) signal(present on the same fragment in theopposite orientation) lacks positive ef-fects, but instead reduced the activity
of pT7TS-derived mRNA (compareT7TS luc and T7TS luc pA in Fig. 2).Processing of SV40 virus transcriptsis regulated by several different se-quence elements. Among them, two el-ements, 13 and 51 nucleotides up-stream of the AAUAAA sequence havebeen shown to cooperate with this sig-nal for efficient cleavage and polyade-nylation of late viral transcripts(Schek et al., 1992). It is, however, notclear whether the function of these el-ements is restricted to nuclear pro-cessing. Although the sequences areU-rich, we could not detect a CPE (re-viewed in Richter, 1999). Neverthe-less, such effects have been observedalso for mRNAs lacking defined CPEs(Verrotti et al., 1996).
Poly(A) sequences improved the ac-tivity of the mRNA in early embryosafter injection at the one-cell stage. Nosignificant advantages were observedwhen the expression was initiatedduring neurulation in chick embryosor in cell culture cells (summarized inFig. 4). The positive effects are, there-fore, either restricted to the earlieststages of embryonic development, orhave to do with specific properties ofartificially synthesized and injectedmRNA. Interestingly these stages cor-relate with the early embryonic activ-ity of cytoplasmic polyadenylation(Sachs, 1990; Richter, 1999), suggest-ing important contributions of thismechanism to the activity of injectedmRNA.
Highly Efficient mRNAProduced From theChimeric pMC Vector
Both the A30 sequence of pT7TS andthe SV40 late poly(A) signal present inpCS2 positively affect the mRNA ac-tivity in early embryos (van der Vel-den et al., 2001; and our SV40 poly(A)swapping experiments). When wecombined these two elements, we ob-tained the chimeric pMC vector (Sup-plementary Figure S1, which can beviewed at http://www.interscience.wiley.com/jpages/1058-8388/suppmat),including also the �-globin UTRs.This combination resulted in a highlysynergistic activity in medaka em-bryos, suggesting independent effectsof the two elements. At 24 hpf, wemeasured its luciferase activity at 20-fold higher levels compared with
IMPROVED TRANSLATION EFFICIENCY OF mRNA 3375
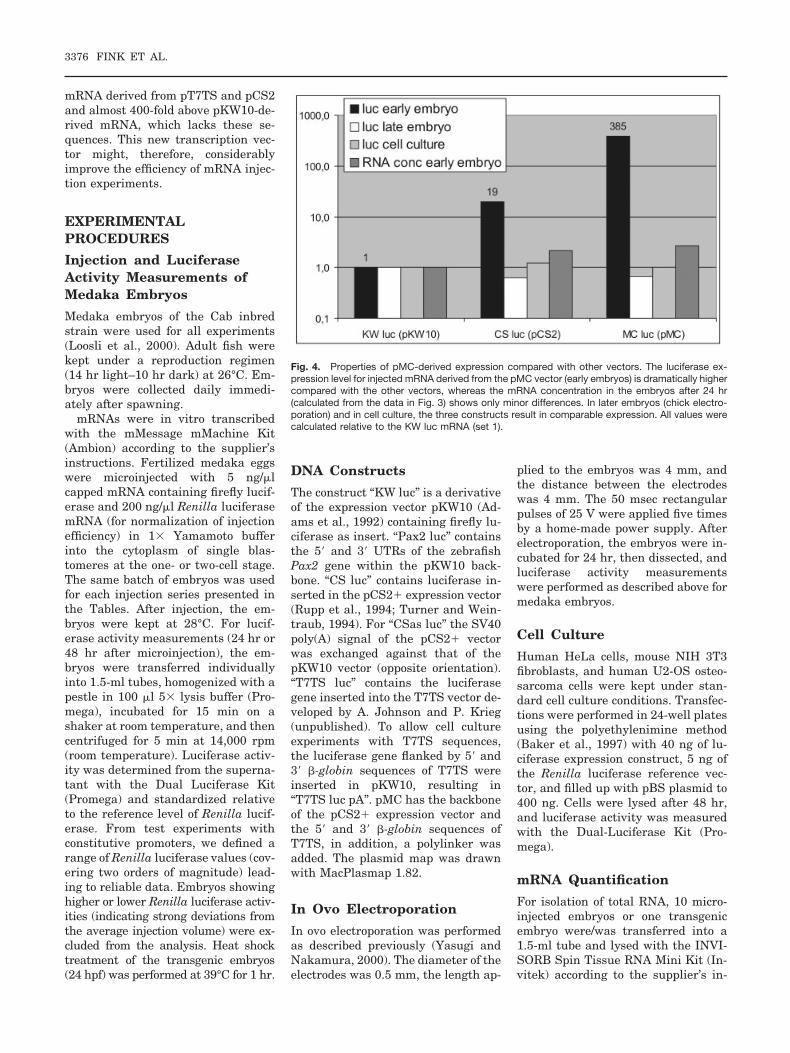
mRNA derived from pT7TS and pCS2and almost 400-fold above pKW10-de-rived mRNA, which lacks these se-quences. This new transcription vec-tor might, therefore, considerablyimprove the efficiency of mRNA injec-tion experiments.
EXPERIMENTALPROCEDURES
Injection and LuciferaseActivity Measurements ofMedaka Embryos
Medaka embryos of the Cab inbredstrain were used for all experiments(Loosli et al., 2000). Adult fish werekept under a reproduction regimen(14 hr light–10 hr dark) at 26°C. Em-bryos were collected daily immedi-ately after spawning.
mRNAs were in vitro transcribedwith the mMessage mMachine Kit(Ambion) according to the supplier’sinstructions. Fertilized medaka eggswere microinjected with 5 ng/�lcapped mRNA containing firefly lucif-erase and 200 ng/�l Renilla luciferasemRNA (for normalization of injectionefficiency) in 1� Yamamoto bufferinto the cytoplasm of single blas-tomeres at the one- or two-cell stage.The same batch of embryos was usedfor each injection series presented inthe Tables. After injection, the em-bryos were kept at 28°C. For lucif-erase activity measurements (24 hr or48 hr after microinjection), the em-bryos were transferred individuallyinto 1.5-ml tubes, homogenized with apestle in 100 �l 5� lysis buffer (Pro-mega), incubated for 15 min on ashaker at room temperature, and thencentrifuged for 5 min at 14,000 rpm(room temperature). Luciferase activ-ity was determined from the superna-tant with the Dual Luciferase Kit(Promega) and standardized relativeto the reference level of Renilla lucif-erase. From test experiments withconstitutive promoters, we defined arange of Renilla luciferase values (cov-ering two orders of magnitude) lead-ing to reliable data. Embryos showinghigher or lower Renilla luciferase activ-ities (indicating strong deviations fromthe average injection volume) were ex-cluded from the analysis. Heat shocktreatment of the transgenic embryos(24 hpf) was performed at 39°C for 1 hr.
DNA Constructs
The construct “KW luc” is a derivativeof the expression vector pKW10 (Ad-ams et al., 1992) containing firefly lu-ciferase as insert. “Pax2 luc” containsthe 5� and 3� UTRs of the zebrafishPax2 gene within the pKW10 back-bone. “CS luc” contains luciferase in-serted in the pCS2� expression vector(Rupp et al., 1994; Turner and Wein-traub, 1994). For “CSas luc” the SV40poly(A) signal of the pCS2� vectorwas exchanged against that of thepKW10 vector (opposite orientation).“T7TS luc” contains the luciferasegene inserted into the T7TS vector de-veloped by A. Johnson and P. Krieg(unpublished). To allow cell cultureexperiments with T7TS sequences,the luciferase gene flanked by 5� and3� �-globin sequences of T7TS wereinserted in pKW10, resulting in“T7TS luc pA”. pMC has the backboneof the pCS2� expression vector andthe 5� and 3� �-globin sequences ofT7TS, in addition, a polylinker wasadded. The plasmid map was drawnwith MacPlasmap 1.82.
In Ovo Electroporation
In ovo electroporation was performedas described previously (Yasugi andNakamura, 2000). The diameter of theelectrodes was 0.5 mm, the length ap-
plied to the embryos was 4 mm, andthe distance between the electrodeswas 4 mm. The 50 msec rectangularpulses of 25 V were applied five timesby a home-made power supply. Afterelectroporation, the embryos were in-cubated for 24 hr, then dissected, andluciferase activity measurementswere performed as described above formedaka embryos.
Cell Culture
Human HeLa cells, mouse NIH 3T3fibroblasts, and human U2-OS osteo-sarcoma cells were kept under stan-dard cell culture conditions. Transfec-tions were performed in 24-well platesusing the polyethylenimine method(Baker et al., 1997) with 40 ng of lu-ciferase expression construct, 5 ng ofthe Renilla luciferase reference vec-tor, and filled up with pBS plasmid to400 ng. Cells were lysed after 48 hr,and luciferase activity was measuredwith the Dual-Luciferase Kit (Pro-mega).
mRNA Quantification
For isolation of total RNA, 10 micro-injected embryos or one transgenicembryo were/was transferred into a1.5-ml tube and lysed with the INVI-SORB Spin Tissue RNA Mini Kit (In-vitek) according to the supplier’s in-
Fig. 4. Properties of pMC-derived expression compared with other vectors. The luciferase ex-pression level for injected mRNA derived from the pMC vector (early embryos) is dramatically highercompared with the other vectors, whereas the mRNA concentration in the embryos after 24 hr(calculated from the data in Fig. 3) shows only minor differences. In later embryos (chick electro-poration) and in cell culture, the three constructs result in comparable expression. All values werecalculated relative to the KW luc mRNA (set 1).
3376 FINK ET AL.

structions. Reverse transcription (RT)reactions were performed with the Re-vert Aid First Strand cDNA SynthesisKit (Fermentas) with 1/15 of the totalRNA using hexamer primers accord-ing to the supplier’s instructions.
Two-step quantitative real-timePCR was carried out using each 1/50of the obtained cDNA. Controls were“minus RT” reactions. The hot startAmpliTaq Gold DNA polymerase wasactivated by incubation at 95°C for 10min followed by amplification of 1 �l ofRT reaction product for 45 cycles at95°C for 15 sec and 60�C for 1 min.Fluorescence “real-time” measure-ments recorded by either the ABIPRISM 7700 or 7900HT Sequence De-tection Systems (Applied Biosystems)were transformed into CT values us-ing the SDS software version 1.9.1and 2.1, respectively. For generationof standard curves, dilution series ofplasmid DNA were made. All reac-tions were performed at least in dupli-cates. Real-time PCR yielded optimalamplification efficiency (E � 0.9).
Primers and Probes
Primers were designed using thePrimer Express TM 1.5 software andare given below as 5�–3� sequence.Standard TaqMan probes were la-beled with the fluorescent quencherdye 6-carboxytetramethylrhodamine(TAMRA) and with the reporter dye6-carboxyfluorescein (6FAM). Primersand probe for the medaka �-actin genewere TCCACCATGAAGATCAAGAT-CATT, TGTTTGGCTGGGGAGGAGand 6FAM-TCCATCGTCCACCGCA-AATGCTTCT-TAMRA. Oligonucleo-tides for quantitation of luciferasemRNA were TGGATTACGTCGCCA-GTCAAG, TTCGGTACTTCGTCCA-CAAACA and 6FAM-AAAGGCCAA-GAAGGGCGGAAAGTCC-TAMRA.
ACKNOWLEDGMENTSWe thank Anders Fjose and OlivierDestree for providing DNA constructs.
REFERENCES
Adams B, Dorfler P, Aguzzi A, Kozmik Z,Urbanek P, Maurer Fogy I, BusslingerM. 1992. Pax-5 encodes the transcriptionfactor BSAP and is expressed in B lym-
phocytes, the developing CNS, and adulttestis. Genes Dev 6:1589–1607.
Aizawa K, Shimada A, Naruse K, MitaniH, Shima A. 2003. The medaka midblas-tula transition as revealed by the expres-sion of the paternal genome. Gene ExprPatterns 3:43–47.
Bajoghli B, Aghaallaei N, Heimbucher T,Czerny T. 2004. An artificial promoterconstruct for heat-inducible misexpres-sion during fish embryogenesis. Dev Biol271:416–430.
Baker A, Saltik M, Lehrmann H, Killisch I,Mautner V, Lamm G, Christofori G, Cot-ten M. 1997. Polyethylenimine (PEI) is asimple, inexpensive and effective re-agent for condensing and linking plas-mid DNA to adenovirus for gene deliv-ery. Gene Ther 4:773–782.
Copeland PR, Wormington M. 2001. Themechanism and regulation of deadeny-lation: identification and characteriza-tion of Xenopus PARN. RNA 7:875–886.
de Moor CH, Meijer H, Lissenden S.2005. Mechanisms of translational con-trol by the 3� UTR in development anddifferentiation. Semin Cell Dev Biol 16:49 –58.
Dehlin E, Wormington M, Korner CG,Wahle E. 2000. Cap-dependent dead-enylation of mRNA. EMBO J 19:1079 –1086.
Ford LP, Bagga PS, Wilusz J. 1997. Thepoly(A) tail inhibits the assembly of a3�-to-5� exonuclease in an in vitro RNAstability system. Mol Cell Biol 17:398–406.
Gurdon JB, Woodland HR, Lingrel JB.1974. The translation of mammalianglobin mRNA injected into fertilizedeggs of Xenopus laevis I. Message sta-bility in development. Dev Biol 39:125–133.
Harland R, Misher L. 1988. Stability ofRNA in developing Xenopus embryosand identification of a destabilizing se-quence in TFIIIA messenger RNA. De-velopment 102:837–852.
Huarte J, Stutz A, O’Connell ML, GublerP, Belin D, Darrow AL, Strickland S,Vassalli JD. 1992. Transient transla-tional silencing by reversible mRNAdeadenylation. Cell 69:1021–1030.
Hyatt TM, Ekker SC. 1999. Vectors andtechniques for ectopic gene expressionin zebrafish. Methods Cell Biol 59:117–126.
Kelly GM, Moon RT. 1995. Involvementof wnt1 and pax2 in the formation ofthe midbrain-hindbrain boundary inthe zebrafish gastrula. Dev Genet 17:129 –140.
Kloc M, Zearfoss NR, Etkin LD. 2002.Mechanisms of subcellular mRNA local-ization. Cell 108:533–544.
Kuersten S, Goodwin EB. 2003. Thepower of the 3� UTR: translational con-trol and development. Nat Rev Genet4:626 –637.
Loosli F, Koster RW, Carl M, Kuhnlein R,Henrich T, Mucke M, Krone A, Wittbrodt
J. 2000. A genetic screen for mutationsaffecting embryonic development inmedaka fish (Oryzias latipes). Mech Dev97:133–139.
Malone RW, Felgner PL, Verma IM. 1989.Cationic liposome-mediated RNA trans-fection. Proc Natl Acad Sci U S A 86:6077–6081.
Mazumder B, Seshadri V, Fox PL. 2003.Translational control by the 3�-UTR: theends specify the means. Trends BiochemSci 28:91–98.
Nakaniwa M, Hirayama M, Shimizu A,Sasaki T, Asakawa S, Shimizu N,Watabe S. 2005. Genomic sequences en-coding two types of medaka hemopexin-like protein Wap65, and their gene ex-pression profiles in embryos. J Exp Biol208:1915–1925.
Okafuji T, Funahashi J, Nakamura H.1999. Roles of Pax-2 in initiation of thechick tectal development. Brain Res DevBrain Res 116:41–49.
Paillard L, Osborne HB. 2003. East ofEDEN was a poly(A) tail. Biol Cell 95:211–219.
Pickering BM, Willis AE. 2005. The impli-cations of structured 5� untranslated re-gions on translation and disease. SeminCell Dev Biol 16:39–47.
Richter JD. 1999. Cytoplasmic polyadenyl-ation in development and beyond. Micro-biol Mol Biol Rev 63:446–456.
Rupp RA, Snider L, Weintraub H. 1994.Xenopus embryos regulate the nuclearlocalizationofXMyoD.GenesDev8:1311–1323.
Sachs A. 1990. The role of poly(A) in thetranslation and stability of mRNA. CurrOpin Cell Biol 2:1092–1098.
Sachs AB, Sarnow P, Hentze MW. 1997.Starting at the beginning, middle, andend: translation initiation in eukaryotes.Cell 89:831–838.
Schek N, Cooke C, Alwine JC. 1992. Defi-nition of the upstream efficiency elementof the simian virus 40 late polyadenyla-tion signal by using in vitro analyses.Mol Cell Biol 12:5386–5393.
Seydoux G. 1996. Mechanisms of transla-tional control in early development. CurrOpin Genet Dev 6:555–561.
Takagi S, Sasado T, Tamiya G, Ozato K,Wakamatsu Y, Takeshita A, Kimura M.1994. An efficient expression vector fortransgenic medaka construction. MolMar Biol Biotechnol 3:192–199.
Turner DL, Weintraub H. 1994. Expres-sion of achaete-scute homolog 3 in Xeno-pus embryos converts ectodermal cells toa neural fate. Genes Dev 8:1434–1447.
van der Velden AW, Voorma HO, ThomasAA. 2001. Vector design for optimal pro-tein expression. Biotechniques 31:572,574, 576–580, passim.
Verrotti AC, Thompson SR, Wreden C,Strickland S, Wickens M. 1996. Evolu-tionary conservation of sequence ele-ments controlling cytoplasmic polyade-nylylation. Proc Natl Acad Sci U S A93:9027–9032.
Vize PD, Melton DA, Hemmati-BrivanlouA, Harland RM. 1991. Assays for gene
IMPROVED TRANSLATION EFFICIENCY OF mRNA 3377

function in developing Xenopus embryos.Methods Cell Biol 36:367–387.
Wickens M, Anderson P, Jackson RJ. 1997.Life and death in the cytoplasm: mes-sages from the 3� end. Curr Opin GenetDev 7:220–232.
Wilusz CJ, Wilusz J. 2004. Bringing therole of mRNA decay in the control of geneexpression into focus. Trends Genet 20:491–497.
Wormington M. 1993. Poly(A) and transla-tion: development control. Curr OpinCell Biol 5:950–954.
Yasugi S, Nakamura H. 2000. Genetransfer into chicken embryos as an ef-fective system of analysis in develop-mental biology. Dev Growth Differ 42:195–197.
Ye W, Bouchard M, Stone D, Liu X, Vella F,Lee J, Nakamura H, Ang SL, Busslinger
M, Rosenthal A. 2001. Distinct regula-tors control the expression of the mid-hindbrain organizer signal FGF8. NatNeurosci 4:1175–1181.
Zhao J, Hyman L, Moore C. 1999. Forma-tion of mRNA 3� ends in eukaryotes:mechanism, regulation, and interrela-tionships with other steps in mRNA syn-thesis. Microbiol Mol Biol Rev 63:405–445.
3378 FINK ET AL.