embryonic birthdate of hypothalamic leptin- activated...
TRANSCRIPT

Embryonic Birthdate of Hypothalamic Leptin-Activated Neurons in Mice
Yuko Ishii and Sebastien G. Bouret
The Saban Research Institute (Y.I., S.G.B.), Neuroscience Program, Children’s Hospital Los Angeles,University of Southern California, Los Angeles, California 90027; and Institut National de la Santé et dela Recherche Médicale (Y.I., S.G.B.), Jean-Pierre Aubert Research Center, Unité 837, NeurobeseLabaratory, University Lille 2, Lille 59045, France
The hypothalamus plays a critical role in the regulation of energy balance. Neuroanatomical andmouse genetic data have defined a core circuitry in the hypothalamus that mediates many of theeffects of leptin on feeding and energy balance regulation. The present study used 5-bromo-2�-deoxyuridine (a marker of dividing cells) and a neuronal marker to systematically examine neu-rogenesis in the mouse embryonic hypothalamus, particularly the birth of neurons that relay leptinsignaling. The vast majority of neurons in hypothalamic nuclei known to control energy balanceis generated between embryonic days (E) 12 and E16, with a sharp peak of neurogenesis occurringon E12. Neurons in the dorsomedial and paraventricular nuclei and the lateral hypothalamic areaare born between E12 and E14. The arcuate and ventromedial nuclei exhibit a relatively longerneurogenic period. Many neurons in these nuclei are born on E12, but some neurons are generatedas late as E16. We also examined the birth of leptin-activated cells by coupling the 5-bromo-2�-deoxyuridine staining with cFos immunohistochemistry. Remarkably, the majority of leptin-acti-vated cells in the adult hypothalamus were also born during a discrete developmental window onE12. These results provide new insight into the development of hypothalamic neurons that controlfeeding and identify important developmental periods when alterations in the intrauterine en-vironment may affect hypothalamic neurogenesis and produce long-term consequences on hy-pothalamic cell numbers. (Endocrinology 153: 3657–3667, 2012)
The hypothalamus has been the traditional focus offeeding regulation because this brain region contains
sets of neurons that are devoted to metabolic regulationand respond directly to peripheral hormonal and nutri-tional signals. Empirical experiments using physical le-sions of specific hypothalamic structures, and, more re-cently, studies using sophisticated transgenic approacheshave specifically revealed the importance of neuronswithin the arcuate nucleus (ARH), ventromedial nucleus(VMH), dorsomedial nucleus (DMH), paraventricularnucleus (PVH), and lateral hypothalamic area (LHA) inthe regulation of body weight and glucose levels (1–7) (andsee Refs. 8 and 9 for reviews). The ARH is the predominantsite for the integration of peripheral blood-borne signals,
including endocrine and metabolic factors. ARH neuronsthat coproduce neuropeptide Y (NPY) and agouti-relatedpeptide (AgRP) or that contain proopiomelanocortin(POMC)-derived peptides directly respond to peripheralhormonal signals, such as the adipocyte-derived hormoneleptin. Both NPY/AgRP- and POMC-containing neuronsproject extensively to other key hypothalamic nuclei, in-cluding the PVH, DMH, and LHA. These target nucleicontain sets of neurons that play a crucial role in energybalance regulation and include neurons that produce an-orexigenic peptides, such as TRH, CRH, and oxytocin inthe PVH, and express orexigenic neuropeptides, such asorexins and melanin-concentrating hormone (MCH) inthe LHA.
ISSN Print 0013-7227 ISSN Online 1945-7170Printed in U.S.A.Copyright © 2012 by The Endocrine Societydoi: 10.1210/en.2012-1328 Received March 23, 2012. Accepted May 4, 2012.First Published Online May 23, 2012
Abbreviations: AgRP, Agouti-related peptide; ARH, arcuate nucleus; BrdU, 5-bromo-2�-deoxyuridine; CNTF, ciliary neurotrophic factor; DMH, dorsomedial nucleus; E, embryonicday; HuC/D, human neuronal protein HuC/HuD; KPBS, potassium PBS; LHA, lateral hypo-thalamic area; MCH, melanin-concentrating hormone; NPY, neuropeptide Y; P, postnatalday; POMC, proopiomelanocortin; PVH, paraventricular nucleus; VMH, ventromedialnucleus.
E N E R G Y B A L A N C E - O B E S I T Y
Endocrinology, August 2012, 153(8):3657–3667 endo.endojournals.org 3657

Impairments of hypothalamic development duringperinatal life may result in lifelong metabolic dysregula-tions due to the importance of the hypothalamus in thecontrol of eating and energy balance (10–12 and see Refs.13–16 for reviews). However, a good understanding of thetiming of normal hypothalamic development in speciesthat are used for the study of metabolic programming,such as the mouse, is particularly critical to adequatelyassess the influence of perinatal environmental factors onhypothalamic development. The formation of the hypo-thalamus is characterized by various developmental pro-cesses that fall into three major categories: 1) the birth ofnew neurons (i.e. neurogenesis); 2) the migration of thesecells to their final destination; and 3) the formation offunctional circuits, which includes axon growth and syn-aptogenesis (see Refs. 17 and 18 for reviews). Axonal tracttracing experiments in mice have revealed that ARH pro-jections develop postnatally and reach their target nucleiwithin distinct temporal domains. Innervation of theDMH occurs on postnatal day (P) 6 followed by innerva-tion of the PVH on P8–10. Projections to the LHA areestablished on P12, and the pattern of ARH projectionsdoes not resemble that of an adult mouse until P18 (19).
Surprisingly little is known regarding the embryonicdevelopment of hypothalamic neurons despite a tremen-dous increase in the interest in the influence of prenatalfactors on lifelong metabolic regulation and hypothalamicdevelopment. Even less information on the precise birth-date of the hypothalamic neurons that mediate leptin’saction is available. Much of the current knowledge on thegeneration of hypothalamic nuclei has been inferred from[3H]thymidine studies, which cannot be used to determinethe chemical phenotype of the generated cells (20–22).The present study used the analog of thymidine 5-bromo-2�-deoxyuridine (BrdU; a marker of dividing cells), withvarious cellular markers to systematically examine thebirthdates of hypothalamic neurons in key hypothalamicnuclei and other areas that are implicated in energy bal-ance regulation, with particular attention on the birthdateof cells that relay leptin signaling.
Materials and Methods
AnimalsTime-pregnant C57BL6/J wild-type mice (Jackson Labora-
tories, Sacamento, CA) were produced in our breeding colony.The mice were mated around midnight and checked for a vaginalplug the next day. The day of conception (sperm positive vaginalsmear) was designated as embryonic day (E) zero (E0). Animalswere provided food and water ad libitum and housed in a 12-hlight, 12-h dark cycle (lights on at 0700 h), temperature-con-
trolled (21–22 C) environment. Animal use was in compliancewith and approved by the Institutional Animal Care and UseCommittee of the Saban Research Institute of Childrens Hospitalof Los Angeles. The day of birth was designated as P0. Litter sizewas culled to six to eight pups (average litter size after culling wasseven pups per litter) at P1 to assure adequate and standardizednutrition until P22 (weaning). Animals were provided ad libitumaccess to standard laboratory chow [Research Diet (New Bruns-wick, NJ)] after weaning. Only male offspring were studied, andeach experimental group in all experiments included offspringfrom four litters.
5-Bromo-2-deoxyuridine injectionsPregnant mice received a single ip injection of BrdU (Sigma,
St. Louis, MO; 160 mg/kg body weight, dissolved in 0.007 N
NaOH) (23) on E10, E12, E14, E16, or E18. For the experimentscombining BrdU and cFos stainings, pregnant mice received ipinjections of BrdU (50 m/kg) on E12, E14, or E16 three times aday (at 0800, 1000, and 1200 h).
Leptin injectionsAdult (P60) offspring of BrdU-injected dams received ip in-
jections of either recombinant murine leptin (3 mg/kg bodyweight; Prepro Tech, Rocky Hill, NJ) or pathogen-free 5 mM Nacitrate buffer (pH 4.0; n � 4 per group). These offspring wereperfused 2 h later as described below.
Tissue preparationMale offspring of the BrdU-injected dams were examined on
P10 in most experiments. The measurement of BrdU and cFosdouble labeling was examined on P60. Mice (n � 4–6 per group)were deeply anesthetized with tribromoethanol and perfusedtranscardially with 0.9% saline, followed by an ice-cold 4%paraformaldehyde solution made in 0.1 M phosphate buffer (pH7.4). The brains were quickly removed from each perfused an-imals, postfixed in the same fixative containing 20% sucrose for2 h at 4 C, and immersed in 20% sucrose in a 0.02 M potassiumPBS (KPBS) solution at 4 C overnight. Frozen coronal sections(25 �m thick, typically at a frequency of 1 in 3) from each brainwere then cut using a cryostat, and every section was mountedonto gelatin-subbed, poly-L-lysine-coated microscope slides andstored in antifreeze at �20 C until use.
ImmunohistochemistryThe sections were processed for double immunofluorescence
after washing in KPBS as described previously (19, 24). A specialprocedure was used to visualize BrdU. Antigen retrieval was per-formed using a microwave. Briefly, the slides were slow boiledfor 10 min, incubated in 0.01 M citrate buffer (pH 6.0) for 30 min,and rinsed in 0.02 M KPBS for 15 min. The sections were placedin 2% normal serum � 0.3% Triton X-100 overnight beforeincubation with either rat anti-BrdU (1:100; Abcam, Inc., Cam-bridge, MA) and mouse anti-human neuronal protein HuC/HuD(HuC/D) antibodies (neuronal marker; 1:250; Invitrogen, Carls-bad, CA) or rat anti-BrdU (1:100; Abcam) and rabbit anti-cFosantibodies (1:1000; Calbiochem, La Jolla, CA) for 48 h at 4 C in2% normal serum � 0.3% Triton X-100. A goat antirat IgGconjugated to Alexa Fluor 488 (1:200; Invitrogen, Eugene, OR)was used to visualize the anti-BrdU. A goat antimouse or goatantirabbit IgG conjugated to Alexa Fluor 568 antibody (1:200;
3658 Ishii and Bouret Embryonic Hypothalamic Neurogenesis Endocrinology, August 2012, 153(8):3657–3667
Invitrogen) was used to visualize the anti-HuC/D and anti-cFosantibodies, respectively. The sections were counterstained withbis-benzamide (1:3000; Invitrogen) to visualize cell nuclei andthe morphological limits of each nucleus. The slides were cov-erslipped with buffered glycerol (pH 8.5) and observed under aZeiss Imager Z1 microscope with a �20 objective (Carl Zeiss,New York, NY).
Data analysis and quantificationTwo independent observers analyzed the sections using a
Zeiss Axio Imager Z1 microscope. Each region of interest wasidentified with the aid of a standard brain atlas, an in-houselibrary of age-matched Nissl-stained sections, and a bis-benz-amide counterstain (Figs. 1-5A). The number of immunoreactivecells for each animal was quantified manually using Image J(National Institutes of Health, Bethesda, MD) in three sectionsthrough the ARH, VMH, DMH, PVH, and LHA. Only brightlyor heavily labeled neurons were considered to ensure the accu-rate determination of neuronal birthdate and proliferation. OnlyBrdU-positive cells with a corresponding bis-benzamide-stainednucleus were included in our quantification. Two categories oflabeled cells were quantified: 1) relative numbers of BrdU-la-beled cells and 2) relative numbers of double-labeled BrdU/HuC/D neurons. The relative numbers of cFos and cFos�BrdUwere assessed in the ARH, VMH, DMH, LHA, and PVH of eachgroup. The results are expressed as the percentage of BrdU/cFosnuclei among the cFos-immunopositive cells. Our measurementswere not intended to provide absolute cell counts but rather toassess relative changes in cell numbers between experimentalgroups. Percentages of cFos-IR cells containing BrdU were com-pared between groups by two-way ANOVA with embryonic dayand treatment group as factors. Fisher’s least significant differ-ence test was used for post hoc comparisons. A P � 0.05 wasdefined as significant.
Results
Pregnant dams were injected with BrdU at 10, 12, 14, 16,or 18 d after conception to assess the birthdate of cells inkey hypothalamic nuclei that are involved in energy bal-ance regulation. The offspring were killed on either P10(for BrdU and HuC/D labelings) or P60 (for leptin inducedcFos), which is when nearly all cells have migrated to theirfinal destinations and differentiated, as well as when themarkers that we studied are expressed at adult levels. BrdUproduced a stereotypical labeling of cell nuclei throughoutthe hypothalamus at all of the ages studied. However, thedegree and timing of neurogenesis varied among the var-ious hypothalamic nuclei.
Birthdate of neurons in the arcuate nucleus of thehypothalamus
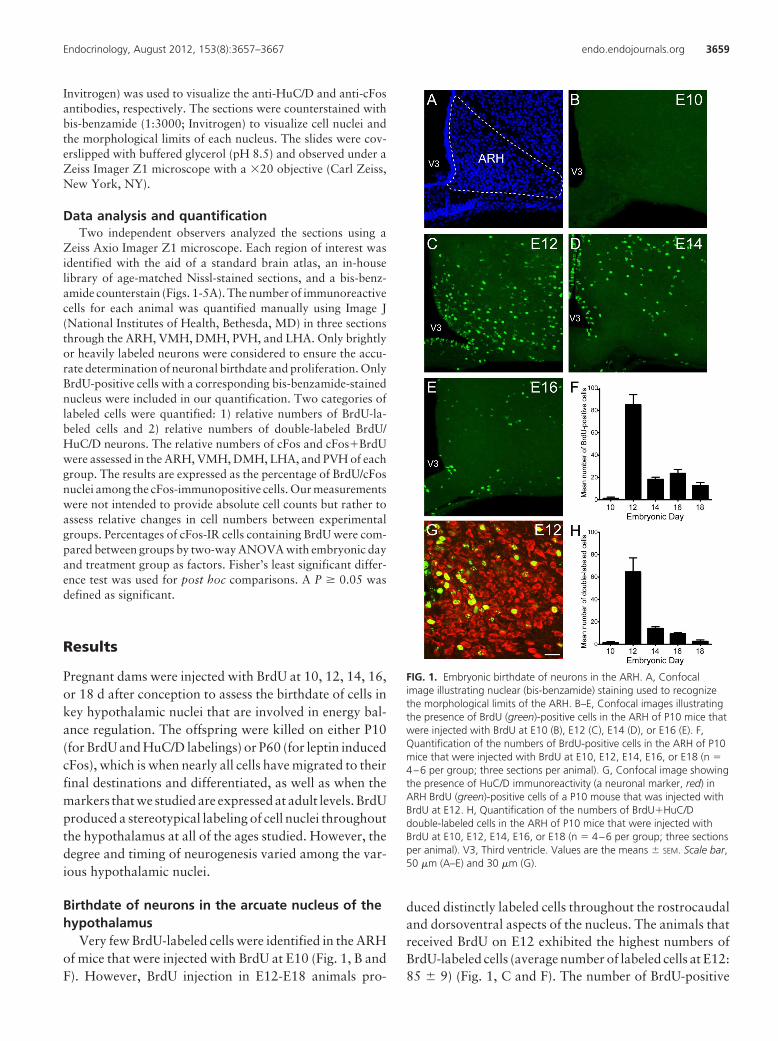
Very few BrdU-labeled cells were identified in the ARHof mice that were injected with BrdU at E10 (Fig. 1, B andF). However, BrdU injection in E12-E18 animals pro-
duced distinctly labeled cells throughout the rostrocaudaland dorsoventral aspects of the nucleus. The animals thatreceived BrdU on E12 exhibited the highest numbers ofBrdU-labeled cells (average number of labeled cells at E12:85 � 9) (Fig. 1, C and F). The number of BrdU-positive
FIG. 1. Embryonic birthdate of neurons in the ARH. A, Confocalimage illustrating nuclear (bis-benzamide) staining used to recognizethe morphological limits of the ARH. B–E, Confocal images illustratingthe presence of BrdU (green)-positive cells in the ARH of P10 mice thatwere injected with BrdU at E10 (B), E12 (C), E14 (D), or E16 (E). F,Quantification of the numbers of BrdU-positive cells in the ARH of P10mice that were injected with BrdU at E10, E12, E14, E16, or E18 (n �4–6 per group; three sections per animal). G, Confocal image showingthe presence of HuC/D immunoreactivity (a neuronal marker, red) inARH BrdU (green)-positive cells of a P10 mouse that was injected withBrdU at E12. H, Quantification of the numbers of BrdU�HuC/Ddouble-labeled cells in the ARH of P10 mice that were injected withBrdU at E10, E12, E14, E16, or E18 (n � 4–6 per group; three sectionsper animal). V3, Third ventricle. Values are the means � SEM. Scale bar,50 �m (A–E) and 30 �m (G).
Endocrinology, August 2012, 153(8):3657–3667 endo.endojournals.org 3659
cells in the ARH decreased by approximately 4- to 5-foldbetween E12 and E14 mice (average number of labeledcells at E14: 18 � 2), and cell numbers remained relativelylow in animals that received BrdU at older ages (averagenumber of cells at E16: 23 � 4; E18: 13 � 3) (Fig. 1, D–F).
We next performed double-labeling experiments using an-tibodies against HuC/D, a marker for differentiated neu-rons, to investigate the chemical phenotype of arcuateBrdU-positive cells. Approximately 60–70% of E12 andE14-labeled BrdU cells also contained HuC/D-immuno-
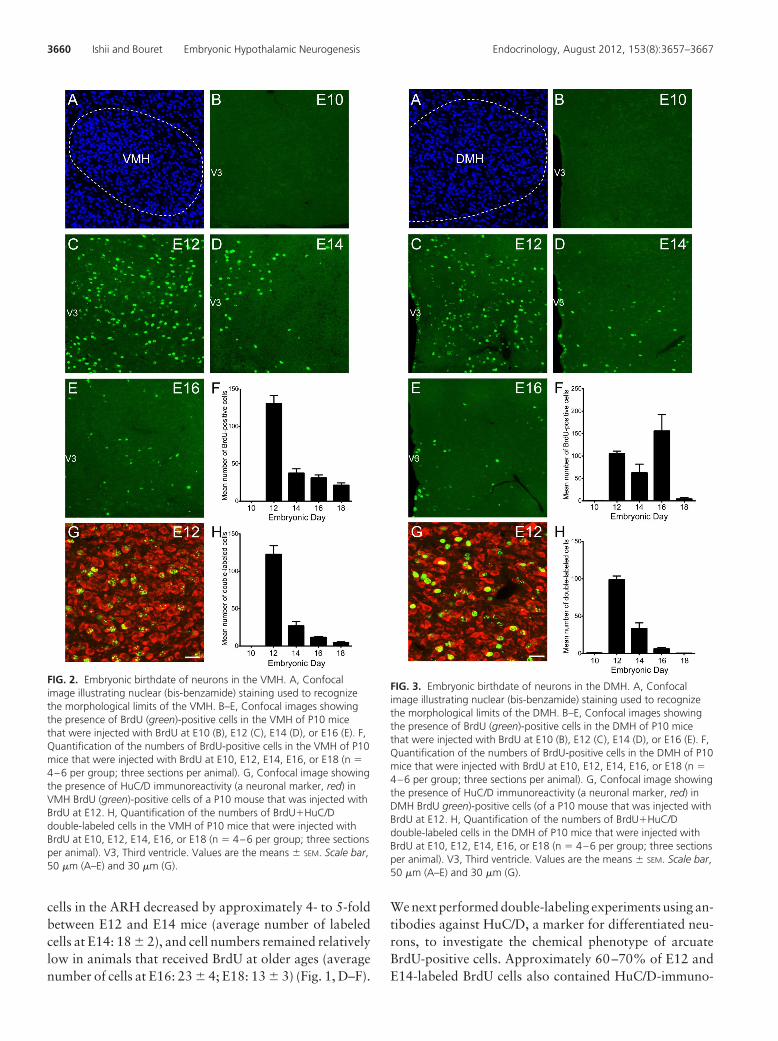
FIG. 2. Embryonic birthdate of neurons in the VMH. A, Confocalimage illustrating nuclear (bis-benzamide) staining used to recognizethe morphological limits of the VMH. B–E, Confocal images showingthe presence of BrdU (green)-positive cells in the VMH of P10 micethat were injected with BrdU at E10 (B), E12 (C), E14 (D), or E16 (E). F,Quantification of the numbers of BrdU-positive cells in the VMH of P10mice that were injected with BrdU at E10, E12, E14, E16, or E18 (n �4–6 per group; three sections per animal). G, Confocal image showingthe presence of HuC/D immunoreactivity (a neuronal marker, red) inVMH BrdU (green)-positive cells of a P10 mouse that was injected withBrdU at E12. H, Quantification of the numbers of BrdU�HuC/Ddouble-labeled cells in the VMH of P10 mice that were injected withBrdU at E10, E12, E14, E16, or E18 (n � 4–6 per group; three sectionsper animal). V3, Third ventricle. Values are the means � SEM. Scale bar,50 �m (A–E) and 30 �m (G).
FIG. 3. Embryonic birthdate of neurons in the DMH. A, Confocalimage illustrating nuclear (bis-benzamide) staining used to recognizethe morphological limits of the DMH. B–E, Confocal images showingthe presence of BrdU (green)-positive cells in the DMH of P10 micethat were injected with BrdU at E10 (B), E12 (C), E14 (D), or E16 (E). F,Quantification of the numbers of BrdU-positive cells in the DMH of P10mice that were injected with BrdU at E10, E12, E14, E16, or E18 (n �4–6 per group; three sections per animal). G, Confocal image showingthe presence of HuC/D immunoreactivity (a neuronal marker, red) inDMH BrdU green)-positive cells (of a P10 mouse that was injected withBrdU at E12. H, Quantification of the numbers of BrdU�HuC/Ddouble-labeled cells in the DMH of P10 mice that were injected withBrdU at E10, E12, E14, E16, or E18 (n � 4–6 per group; three sectionsper animal). V3, Third ventricle. Values are the means � SEM. Scale bar,50 �m (A–E) and 30 �m (G).
3660 Ishii and Bouret Embryonic Hypothalamic Neurogenesis Endocrinology, August 2012, 153(8):3657–3667
reactivity (Fig. 1, G and H). However, only 40 and 20%of E16- and E18-labeled BrdU cells were HuC/D-immu-nopositive, respectively (Fig. 1H). These results indicatethat neurons in the ARH were generated between E12 andE18, with a peak of neurogenesis occurring at E12.
Birthdate of neurons in the ventromedial nucleusof the hypothalamus
No BrdU-labeled cells were observed in the VMH ofmice that received BrdU on E10 (Fig. 2, B and F). In con-trast, animals that received BrdU on E12-E18 exhibited
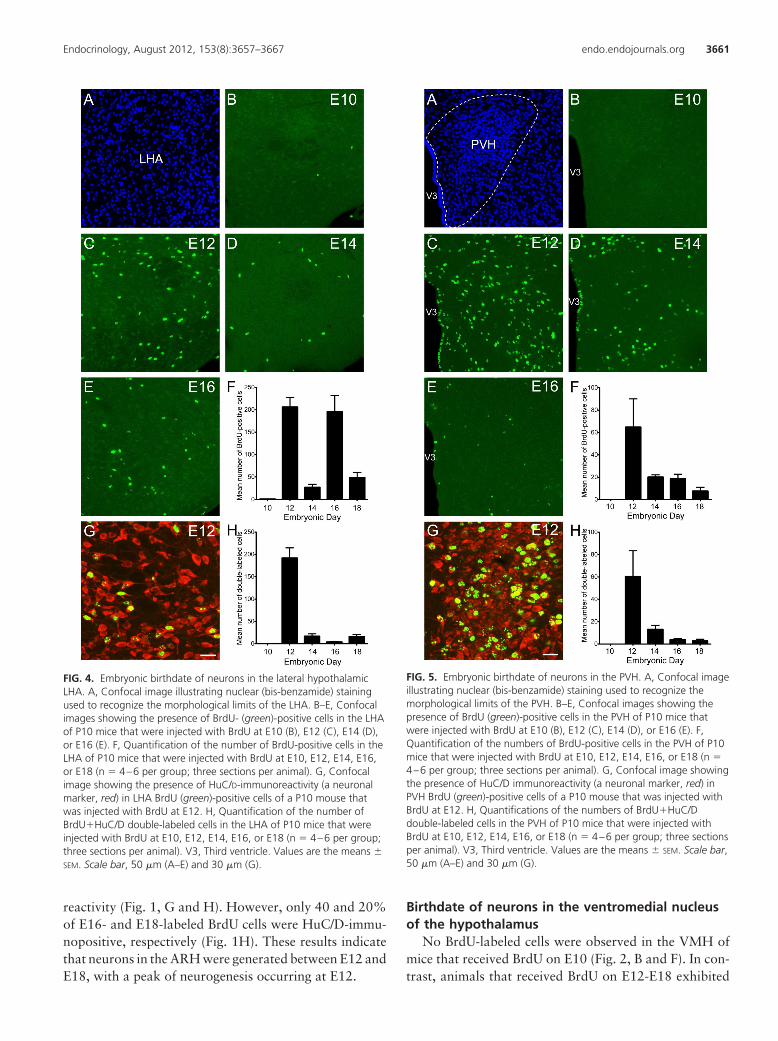
FIG. 4. Embryonic birthdate of neurons in the lateral hypothalamicLHA. A, Confocal image illustrating nuclear (bis-benzamide) stainingused to recognize the morphological limits of the LHA. B–E, Confocalimages showing the presence of BrdU- (green)-positive cells in the LHAof P10 mice that were injected with BrdU at E10 (B), E12 (C), E14 (D),or E16 (E). F, Quantification of the number of BrdU-positive cells in theLHA of P10 mice that were injected with BrdU at E10, E12, E14, E16,or E18 (n � 4–6 per group; three sections per animal). G, Confocalimage showing the presence of HuC/D-immunoreactivity (a neuronalmarker, red) in LHA BrdU (green)-positive cells of a P10 mouse thatwas injected with BrdU at E12. H, Quantification of the number ofBrdU�HuC/D double-labeled cells in the LHA of P10 mice that wereinjected with BrdU at E10, E12, E14, E16, or E18 (n � 4–6 per group;three sections per animal). V3, Third ventricle. Values are the means �SEM. Scale bar, 50 �m (A–E) and 30 �m (G).
FIG. 5. Embryonic birthdate of neurons in the PVH. A, Confocal imageillustrating nuclear (bis-benzamide) staining used to recognize themorphological limits of the PVH. B–E, Confocal images showing thepresence of BrdU (green)-positive cells in the PVH of P10 mice thatwere injected with BrdU at E10 (B), E12 (C), E14 (D), or E16 (E). F,Quantification of the numbers of BrdU-positive cells in the PVH of P10mice that were injected with BrdU at E10, E12, E14, E16, or E18 (n �4–6 per group; three sections per animal). G, Confocal image showingthe presence of HuC/D immunoreactivity (a neuronal marker, red) inPVH BrdU (green)-positive cells of a P10 mouse that was injected withBrdU at E12. H, Quantifications of the numbers of BrdU�HuC/Ddouble-labeled cells in the PVH of P10 mice that were injected withBrdU at E10, E12, E14, E16, or E18 (n � 4–6 per group; three sectionsper animal). V3, Third ventricle. Values are the means � SEM. Scale bar,50 �m (A–E) and 30 �m (G).
Endocrinology, August 2012, 153(8):3657–3667 endo.endojournals.org 3661

labeled cells throughout the VMH. The animals that re-ceived BrdU on E12 exhibited the highest numbers ofBrdU-labeled cells (average number of labeled cells at E12:130 � 11) (Fig. 2, C and F), which is similar to the ARHresults. The numbers of BrdU-labeled cells on E14-E18were moderate and 3–6 times lower than those in the E12animals (average number of cells at E14: 37 � 6; E16: 30 �
4; and E18: 21 � 3) (Fig. 2, D–F). Double-labeling exper-iments using the neuronal marker HuC/D revealed that 90and 70% of BrdU cells that were labeled on E12 and E14,respectively, also contained HuC/D immunoreactivity(Fig. 2, G and H). However, only 40 and 20% of BrdU cellslabeled on E16 and E18, respectively, were HuC/D im-munopositive (Fig. 2H). These data show that VMH neu-rons were generated between E12 and E18 with a peak onE12. Notably, each subdivision of the VMH followed thesame temporal pattern of neurogenesis with a peak of neu-ronal formation occurring on E12 (Table 1).
Birthdate of neurons in the dorsomedial nucleusof the hypothalamus
Virtually no BrdU-labeled cells were identified in theDMH of mice that received BrdU on E10 (Fig. 3, B and F).The injection of BrdU on E12-E16 produced a high num-ber of labeled cells in the DMH (average number of cellsat E12: 105 � 6; E14: 62 � 20; and E16: 155 � 37) (Fig.3, C–F). However, few BrdU-labeled cells were observedin the DMH of animals that received BrdU on E18 (aver-age number of cells at E18: 4 � 3) (Fig. 3F). Double-labeling experiments show that 90 and 60% of cells thatwere labeled with BrdU on E12 and E14 displayed HuC/D-IR, respectively (Fig. 3, G and H). However, only afraction (�5%) of the DMH cells that were generated onE16 acquired a neuronal phenotype. Similarly, less than2% of DMH BrdU cells that were labeled with BrdU onE18 displayed HuC/D immunoreactivity (Fig. 3H). Thesedata show that the majority of neurons in the DMH wereborn between E12 and E14.
Birthdate of neurons located in the LHANo BrdU-labeled cells were observed in the LHA of
mice that received BrdU on E10 (Fig. 4, B and F). However,clear BrdU incorporation was identified in the LHA ofanimals that received BrdU on E12-E18. The highest num-bers of BrdU-labeled cells were identified in animals thatreceived BrdU on E12 (average number of labeled cells atE12: 206 � 21) (Fig. 4, C and F). The number of BrdU-positive cells in the LHA decreased by approximately8-fold between E12 and E14 mice (average number oflabeled cells at E14: 26 � 7). Animals that received BrdUon E16 displayed high numbers of positive cells in the LHA(average number of cells at E16: 195 � 36) (Fig. 4, D–F),and the number of BrdU-immunoreactive cells was dra-matically reduced on E18 (average number of cells at E18:48 � 11) (Fig. 4F). Double-labeling experiments using theHuC/D and BrdU antibodies demonstrated that approx-imately 90 and 60% of the cells that were generated onE12 and E14 acquired a neuronal phenotype, respectively(Fig. 4, G and H). However, less than 3% of the BrdU-positive cells that were generated on E16 were HuC/Dimmunopositive (Fig. 4H). Approximately 30% of thecells in the LHA that were generated on E18 displayed aneuronal phenotype (Fig. 4H). These results indicated thatcells in the LHA were generated primarily during twosharp peaks on E12 and E16 but that neurogenesis in theLHA was largely restricted on E12.
Birthdate of neurons in the paraventricularnucleus of the hypothalamus
No BrdU-labeled cells were observed in the PVH ofmice that received BrdU on E10 (Fig. 5, B and F). In con-trast, numerous BrdU-labeled cells were identified in theVMH of animals that received BrdU on E12-E18. Animalsthat received BrdU on E12 exhibited the highest numbersof BrdU-labeled cells in the PVH (average number of la-beled cells at E12: 64 � 25) (Fig. 5, C and F). The numbersof BrdU-labeled cells in animals that received BrdU onE14-E16 were moderate and approximately three times
TABLE 1. Mean number of BrdU�HuC/D double-labeled cells in distinct anatomical components of the VMH andPVH
E10 E12 E14 E16 E18
VMHDorsomedial part 0 � 0 59.8 � 11.5 22.5 � 5.7 4.0 � 1.8 2.3 � 1.3Central part 0 � 0 37.5 � 5.0 3.0 � 0.7 3.8 � 1.5 1.5 � 0.9Ventrolateral part 0 � 0 25.2 � 3.0 1.2 � 0.9 3.7 � 0.9 0.75 � 0.75
PVNPosterior magnocellular part, lateral zone 0 � 0 14.7 � 1.9 0.2 � 0.2 0.5 � 0.5 0.7 � 0.3Medial parvicellular part, ventral zone 0 � 0 14.0 � 3.8 0.8 � 0.2 1.2 � 0.5 1.0 � 0.0Medial parvicellular part, dorsal zone 0 � 0 35.0 � 6.1 5.2 � 2.7 1.5 � 0.6 2.0 � 1.5Dorsal parvicellular part 0 � 0 16.7 � 5.7 6.8 � 1.9 0.5 � 0.5 0 � 0
3662 Ishii and Bouret Embryonic Hypothalamic Neurogenesis Endocrinology, August 2012, 153(8):3657–3667

lower than those in E12 animals (average number of cellsat E14: 20 � 2; E16: 18 � 4) (Fig. 5, D–F). The numbersof labeled cells in the PVH further decreased in the animalsthat received BrdU on E18 (average number of cells at E18:7 � 3) (Fig. 5F). Double-labeling experiments using theneuronal marker HuC/D demonstrated that 70 and 60%of BrdU-positive cells that were labeled on E12 and E14also contained HuC/D immunoreactivity, respectively(Fig. 5, G and H). However, only 20 and 30% of BrdU-positive cells that were labeled on E16 and E18 wereHuC/D immunopositive, respectively (Fig. 5H). Thesedata show that PVH neurons were generated between E12and E18, with a sharp peak on E12. Notably, each sub-division of the PVH followed the same temporal pattern ofneurogenesis with a peak of neuronal formation occurringon E12 (Table 1).
Birthdate of leptin-activated neurons in thehypothalamus
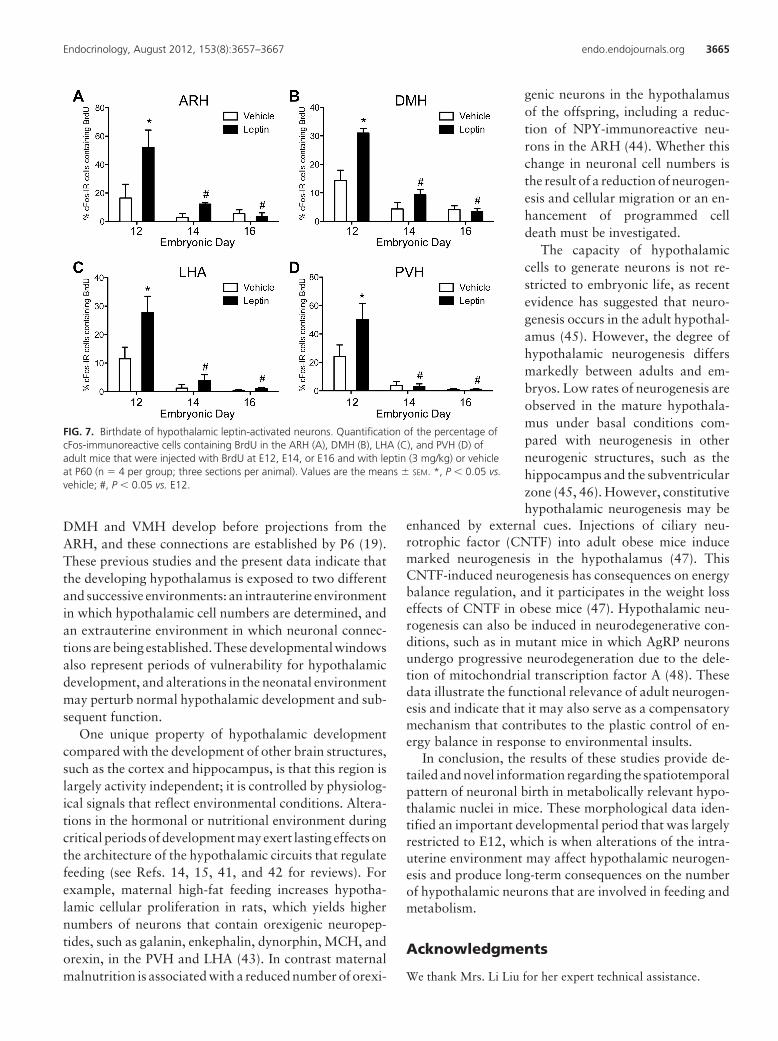
Leptin primarily acts within the hypothalamus to me-diate its actions on feeding and body weight regulation (2,25, 26). We repeated the BrdU injections described aboveand also injected the adult animals with either leptin (3mg/kg, ip) or vehicle alone to determine when hypotha-lamic neurons that convey leptin signaling during adult lifeare born. The brains were stained for cFos becausechanges in cFos staining are generally taken to representan increase in neuronal activity that can be conveyed eitherdirectly by leptin or through transynaptic activation (27).Leptin treatment produced a marked induction of cFos-immunoreactivity in the ARH, DMH, LHA, and PVH ofadult mice (Supplemental Figs. 1 and 2, published on TheEndocrine Society’s Journals Online web site at http://en-do.endojournals.org) as reported previously (19, 28).Double-staining experiments with BrdU indicated that30–50% of leptin-sensitive neurons in the adult hypothal-amus, including in the ARH, DMH, LHA, and PVH, wereborn during a short developmental period that is restrictedon E12 (Figs. 6 and 7). Only a small proportion of hypo-thalamic leptin-activated cells (1–10%) were born duringlater embryonic stages (Figs. 6 and 7).
Discussion
The developmental process of a functional hypothalamicnucleus begins with the birth or terminal mitosis of theneurons within that nucleus. In their landmark analyses ofthe fetal rodent hypothalamus, Altman and Bayer (20) andShimada and Nakamura (21) reported using [3H]thymi-dine, which indicated that cells in the hypothalamic nucleiare primarily derived from precursors that originate in the
proliferative zone surrounding the lower portion of thethird ventricle. This proliferative zone is also known as theneuroepithelium of the third ventricle. These authors fur-ther demonstrated that numerous cells in the hypothala-mus were born embryonically in rats (20) and mice (21).These studies have provided valuable information on thenumbers and timing of hypothalamic cell production, butthey have not investigated the chemical identity of the cellsthat are born at a given time. The present study used BrdUto examine the birthdate of hypothalamic neurons in nu-clei that play a key role in energy balance. Moreover, weperformed a detailed analysis of embryonic hypothalamicneurogenesis in the mouse, which is a widely used modelin molecular genetic studies of feeding physiology. Theadvantages of using BrdU, instead of [3H]thymidine, in-clude a lower background, intranuclear labeling, and thevisualization of the uridine analog with immunofluores-cence, which provides a convenient, simultaneous visual-ization of multiple markers. It is worth noting that highdoses of BrdU can lead to teratogenic malformations andbehavioral changes. However, we did not observe any ma-jor physiological or behavioral alterations nor any grossmorphological changes in our BrdU-injected animals.These observations suggest that it was unlikely that theregime of BrdU administration used in these studies causedsignificant neurotoxicity.
Our present findings show that the vast majority ofneurons in hypothalamic nuclei that control energy ho-meostasis in mice are born between E12 and E14 withpeak birthdates on E12. These findings suggest that theproliferation rate in the neuroepithelium of the third ven-tricle is high at E12 and decreases after E14. They alsocontrast with those reported in rats, which exhibit a rel-atively long neurogenic period that is initiated on E11 andcontinues until E17, with peak birthdates on E13-E15 (17,20). Reports on human fetal chemoarchitecture and cy-toarchitecture have also suggested that early hypotha-lamic neurogenesis is limited to the ninth and 10th weeksof gestation (29–33).
The chemical phenotype of the nonneuronal HuC/D-BrdU� cells remains to be determined. Only a minority ofglial fibrillary acidic protein-positive astrocytes was bornembryonically (data not shown). However, based on theobservation that the hypothalamus is one of the most vas-cularized brain region, it is likely that some of these non-neuronal cells born embryonically become vascular endo-thelial cells. Remarkably, the majority of leptin-activatedcells in the adult hypothalamus (as revealed by cFos-IRafter leptin administration) were born during a develop-mental period that was largely restricted to E12. We choseto use cFos staining to detect leptin-activated cells, insteadof more direct leptin receptor signaling markers such as
Endocrinology, August 2012, 153(8):3657–3667 endo.endojournals.org 3663

phosphorylated signal transducer and activator of tran-scription-3 or phosphorylated ERK1/2 because this pro-tooncogene labels neurons that are both directly and tran-synaptically activated by leptin and identifies both first-and second-order leptin-activated neurons. Brischoux etal. (34) previously reported that MCH neurons in the ratLHA were born between E12 and E13 and that MCHmRNA was detected in the LHA as early as E13. More-over, recent birthdating studies indicate that POMC neu-rons in the ARH are born primarily on E12, and in situhybridization studies have demonstrated that neurons ex-press POMC mRNA on E12 (35, 36). These anatomicalobservations are consistent with an early determination ofcell fate. In addition, specific sets of hypothalamic neuronsmay share a common ontological lineage. For example,Padilla et al. (36) showed that a subpopulation of embry-onic Pomc-expressing precursors subsequently adopts aNPY phenotype in adult mice. These studies also identified
an important developmental period when alterations inthe intrauterine environment may affect hypothalamicneurogenesis and produce enduring structural conse-quences in the hypothalamus.
A second important hypothalamic developmental pe-riod in rodents occurs during the first few weeks of post-natal life when neurons send their axonal projections totheir target sites. Projections from the ARH in mice andrats are immature at birth and primarily develop postna-tally. The development of axonal projections from theARH to each of their target nuclei is not initiated until P6,and these projections are not fully mature, i.e. they do notresemble an adult, until P18 (19). Similar results have beenobserved for the development of AgRP-containing fibersin mice and rats (37, 38), which originate exclusively fromNPY-containing neurons in the ARH (39, 40). Variousparts of the hypothalamus extend axonal projections atdifferent times. For example, efferent projections from the
FIG. 6. Embryonic birthdate of leptin-activated neurons. Confocal images showing leptin-induced cFos immunoreactivity (red), BrdUimmunoreactivity (green), and cFos�BrdU double staining (yellow) in the ARH, DMH, LHA, and PVH of P60 mice that were injected with BrdU atE12 or E14. V3, Third ventricle. Scale bar, 50 �m.
3664 Ishii and Bouret Embryonic Hypothalamic Neurogenesis Endocrinology, August 2012, 153(8):3657–3667
DMH and VMH develop before projections from theARH, and these connections are established by P6 (19).These previous studies and the present data indicate thatthe developing hypothalamus is exposed to two differentand successive environments: an intrauterine environmentin which hypothalamic cell numbers are determined, andan extrauterine environment in which neuronal connec-tions are being established. These developmental windowsalso represent periods of vulnerability for hypothalamicdevelopment, and alterations in the neonatal environmentmay perturb normal hypothalamic development and sub-sequent function.
One unique property of hypothalamic developmentcompared with the development of other brain structures,such as the cortex and hippocampus, is that this region islargely activity independent; it is controlled by physiolog-ical signals that reflect environmental conditions. Altera-tions in the hormonal or nutritional environment duringcritical periods of development may exert lasting effects onthe architecture of the hypothalamic circuits that regulatefeeding (see Refs. 14, 15, 41, and 42 for reviews). Forexample, maternal high-fat feeding increases hypotha-lamic cellular proliferation in rats, which yields highernumbers of neurons that contain orexigenic neuropep-tides, such as galanin, enkephalin, dynorphin, MCH, andorexin, in the PVH and LHA (43). In contrast maternalmalnutrition is associated with a reduced number of orexi-
genic neurons in the hypothalamusof the offspring, including a reduc-tion of NPY-immunoreactive neu-rons in the ARH (44). Whether thischange in neuronal cell numbers isthe result of a reduction of neurogen-esis and cellular migration or an en-hancement of programmed celldeath must be investigated.
The capacity of hypothalamiccells to generate neurons is not re-stricted to embryonic life, as recentevidence has suggested that neuro-genesis occurs in the adult hypothal-amus (45). However, the degree ofhypothalamic neurogenesis differsmarkedly between adults and em-bryos. Low rates of neurogenesis areobserved in the mature hypothala-mus under basal conditions com-pared with neurogenesis in otherneurogenic structures, such as thehippocampus and the subventricularzone (45, 46). However, constitutivehypothalamic neurogenesis may be
enhanced by external cues. Injections of ciliary neu-rotrophic factor (CNTF) into adult obese mice inducemarked neurogenesis in the hypothalamus (47). ThisCNTF-induced neurogenesis has consequences on energybalance regulation, and it participates in the weight losseffects of CNTF in obese mice (47). Hypothalamic neu-rogenesis can also be induced in neurodegenerative con-ditions, such as in mutant mice in which AgRP neuronsundergo progressive neurodegeneration due to the dele-tion of mitochondrial transcription factor A (48). Thesedata illustrate the functional relevance of adult neurogen-esis and indicate that it may also serve as a compensatorymechanism that contributes to the plastic control of en-ergy balance in response to environmental insults.
In conclusion, the results of these studies provide de-tailed and novel information regarding the spatiotemporalpattern of neuronal birth in metabolically relevant hypo-thalamic nuclei in mice. These morphological data iden-tified an important developmental period that was largelyrestricted to E12, which is when alterations of the intra-uterine environment may affect hypothalamic neurogen-esis and produce long-term consequences on the numberof hypothalamic neurons that are involved in feeding andmetabolism.
Acknowledgments
We thank Mrs. Li Liu for her expert technical assistance.
FIG. 7. Birthdate of hypothalamic leptin-activated neurons. Quantification of the percentage ofcFos-immunoreactive cells containing BrdU in the ARH (A), DMH (B), LHA (C), and PVH (D) ofadult mice that were injected with BrdU at E12, E14, or E16 and with leptin (3 mg/kg) or vehicleat P60 (n � 4 per group; three sections per animal). Values are the means � SEM. *, P � 0.05 vs.vehicle; #, P � 0.05 vs. E12.
Endocrinology, August 2012, 153(8):3657–3667 endo.endojournals.org 3665

Address all correspondence and requests for reprints to: Se-bastien G. Bouret, Ph.D., The Saban Research Institute, Neuro-science Program, Children’s Hospital Los Angeles, University ofSouthern California, 4650 Sunset Boulevard, MS no. 135, LosAngeles, California 90027. E-mail: [email protected].
This work was supported by National Institute of HealthGrant DK84142 (to S.G.B.), the Fondation pour la RechercheMédicale (to S.G.B.), the European Union FP7 integrated project(Grant Agreement 266408, “Full4Health,” to S.G.B.), and theAgence Nationale de la Recherche Grant ANR-08-JCJC-0055-01 (to S.G.B.).
Disclosure Summary: The authors (Y.I. and S.G.B.) havenothing to disclose.
References
1. Hetherington AW, Ranson SW 1940 Hypothalamic lesions and ad-iposity in the rat. Anat Rec 78 149–172
2. Elmquist JK, Coppari R, Balthasar N, Ichinose M, Lowell BB 2005Identifying hypothalamic pathways controlling food intake, bodyweight, and glucose homeostasis. J Comp Neurol 493:63–71
3. Dhillon H, Zigman JM, Ye C, Lee CE, McGovern RA, Tang V,Kenny CD, Christiansen LM, White RD, Edelstein EA, Coppari R,Balthasar N, Cowley MA, Chua S Jr, Elmquist JK, Lowell BB 2006Leptin directly activates SF1 neurons in the VMH, and this action byleptin is required for normal body-weight homeostasis. Neuron 49:191–203
4. Ring LE, Zeltser LM 2010 Disruption of hypothalamic leptin sig-naling in mice leads to early-onset obesity, but physiological adap-tations in mature animals stabilize adiposity levels. J Clin Invest120:2931–2941
5. Hill JW, Elias CF, Fukuda M, Williams KW, Berglund ED, HollandWL, Cho YR, Chuang JC, Xu Y, Choi M, Lauzon D, Lee CE, Cop-pari R, Richardson JA, Zigman JM, Chua S, Scherer PE, Lowell BB,Brüning JC, Elmquist JK 2010 Direct insulin and leptin action onpro-opiomelanocortin neurons is required for normal glucose ho-meostasis and fertility. Cell Metabolism 11:286–297
6. Balthasar N, Dalgaard LT, Lee CE, Yu J, Funahashi H, Williams T,Ferreira M, Tang V, McGovern RA, Kenny CD, Christiansen LM,Edelstein E, Choi B, Boss O, Aschkenasi C, Zhang CY, Mountjoy K,Kishi T, Elmquist JK, Lowell BB 2005 Divergence of melanocortinpathways in the control of food intake and energy expenditure. Cell123:493–505
7. Segal-Lieberman G, Bradley RL, Kokkotou E, Carlson M, TromblyDJ, Wang X, Bates S, Myers Jr MG, Flier JS, Maratos-Flier E 2003Melanin-concentrating hormone is a critical mediator of the leptin-deficient phenotype. Proc Natl Acad Sci USA 100:10085–10090
8. Sawchenko PE 1998 Toward a new neurobiology of energy balance,appetite, and obesity: the anatomists weigh in. J Comp Neurol 402:435–441
9. Berthoud HR 2006 Homeostatic and non-homeostatic pathwaysinvolved in the control of food intake and energy balance. Obesity14:197S–200S
10. Bouret SG, Gorski JN, Patterson CM, Chen S, Levin BE, Simerly RB2008 Hypothalamic neural projections are permanently disrupted indiet-induced obese rats. Cell Metab 7:179–185
11. Coupé B, Amarger V, Grit I, Benani A, Parnet P 2010 Nutritionalprogramming affects hypothalamic organization and early responseto leptin. Endocrinology 151:702–713
12. Kirk SL, Samuelsson AM, Argenton M, Dhonye H, Kalamatianos T,Poston L, Taylor PD, Coen CW 2009 Maternal obesity induced by
diet in rats permanently influences central processes regulating foodintake in offspring. PLoS ONE 4:e5870
13. Horvath TL, Bruning JC 2006 Developmental programming of thehypothalamus: a matter of fat. Nat Med 12:52–53
14. Levin B 2006 Metabolic imprinting: critical impact of the perinatalenvironment on the regulation of energy homeostasis. Philos TransR Soc Lond B Biol Sci361:1107–1121
15. Plagemann A 2006 Perinatal nutrition and hormone-dependent pro-gramming of food intake. Horm Res 65:83–89
16. Martin-Gronert MS, Ozanne SE 2005 Programming of appetite andtype 2 diabetes. Early Hum Dev 81:981–988
17. Markakis EA 2002 Development of the neuroendocrine hypothal-amus. Front Neuroendocrinol 23:257–291
18. Bouret SG 2010 Development of hypothalamic neural networkscontrolling appetite. Forum Nutr 63:84–93
19. Bouret SG, Draper SJ, Simerly RB 2004 Formation of projectionpathways from the arcuate nucleus of the hypothalamus to hypo-thalamic regions implicated in the neural control of feeding behaviorin Mice. J Neurosci 24:2797–2805
20. Altman J, Bayer SA 1986 The development of the rat hypothalamus.Adv Anat Embryol Cell Biol 100:1–178
21. Shimada M, Nakamura T 1973 Time of neuron origin in mousehypothalamic nuclei. Exp Neurol 41:163–173
22. Ifft JD 1972 An autoradiographic study of the time of final divisionof neurons in rat hypothalamic nuclei. J Comp Neurol 144:193–204
23. Markakis EA, Swanson LW 1997 Spatiotemporal patterns of secre-tomotor neuron generation in the parvicellular neuroendocrine sys-tem. Brain Res Rev 24:255–291
24. Caron E, Sachot C, Prevot V, Bouret SG 2010 Distribution of leptin-sensitive cells in the postnatal and adult mouse brain. J Comp Neurol518:459–476
25. Myers Jr MG, Münzberg H, Leinninger GM, Leshan RL 2009 Thegeometry of leptin action in the brain: more complicated than asimple ARC. Cell Metab 9:117–123
26. Ahima RS, Saper CB, Flier JS, Elmquist JK 2000 Leptin regulationof neuroendocrine systems. Front Neuroendocrinol 21:263–307
27. Armstrong RC, Montminy MR 1993 Transsynaptic control of geneexpression. Annu Rev Neurosci 16:17–29
28. Elmquist JK, Ahima RS, Maratos-Flier E, Flier JS, Saper CB 1997Leptin activates neurons in ventrobasal hypothalamus and brains-tem. Endocrinology 138:839–842
29. Bugnon C, Fellmann D, Bresson JL, Clavequin MC 1982 Immuno-cytochemical study of the ontogenesis of the CRH-containing neu-roglandular system in the human hypothalamus. C R Seances AcadSci III 294:599–604
30. Burford GD, Robinson IC 1982 Oxytocin, vasopressin and neuro-physins in the hypothalamo-neurohypophysial system of the humanfetus. J Endocrinol 95:403–408
31. Ackland J, Ratter S, Bourne GL, Rees LH 1983 Characterization ofimmunoreactive somatostatin in human fetal hypothalamic tissue.Regul Pept 5:95–101
32. Mai JK, Lensing-Höhn S, Ende AA, Safroniew MV 1997 Develop-mental organization of neurophysin neurons in the human brain.J Comp Neurol 385:477–489
33. Koutcherov Y, Mai JK, Ashwell KW, Paxinos G 2002 Organizationof human hypothalamus in fetal development. J Comp Neurol 446:301–324
34. Brischoux F, Fellman D, Risold PY 2001 Ontogenetic developmentof the diencephalic MCH neurons: a hypothalamic ’MCH area’hypothesis. Eur J Neurosci 13:1733–1744
35. Khachaturian H, Alessi NE, Lewis ME, Munfakh N, FitzsimmonsMD, Watson SJ 1985 Development of hypothalamic opioid neu-rons: a combined immunocytochemical and [3H]thymidine auto-radiographic study. Neuropeptides 5:477–480
36. Padilla SL, Carmody JS, Zeltser LM 2010 Pomc-expressing progen-itors give rise to antagonistic neuronal populations in hypothalamicfeeding circuits. Nat Med 16:403–405
3666 Ishii and Bouret Embryonic Hypothalamic Neurogenesis Endocrinology, August 2012, 153(8):3657–3667

37. Grove KL, Allen S, Grayson BE, Smith MS 2003 Postnatal devel-opment of the hypothalamic neuropeptide Y system. Neuroscience116:393–406
38. Nilsson I, Johansen JE, Schalling M, Hökfelt T, Fetissov SO 2005Maturation of the hypothalamic arcuate agouti-related protein sys-tem during postnatal development in the mouse. Dev Brain Res155:147–154
39. Broberger C, Johansen J, Johansson C, Schalling M, Hökfelt T 1998The neuropeptide Y/agouti gene-related protein (AGRP) brain cir-cuitry in normal, anorectic, and monosodium glutamate-treatedmice. Proc Natl Acad Sci USA 95:15043–15048
40. Bagnol D, Lu XY, Kaelin CB, Day HE, Ollmann M, Gantz I, AkilH, Barsh GS, Watson SJ 1999 Anatomy of an endogenous antago-nist: relationship between Agouti-related protein and proopiomel-anocortin in brain. J Neurosci 19:RC26
41. Bouret SG 2010 Role of early hormonal and nutritional experiencesin shaping feeding behavior and hypothalamic development. J Nutr140:653–657
42. Sullivan EL, Grove KL 2010 Metabolic imprinting in obesity. ForumNutr 63:186–194
43. Chang GQ, Gaysinskaya V, Karatayev O, Leibowitz SF 2008 Ma-ternal high-fat diet and fetal programming: increased proliferationof hypothalamic peptide-producing neurons that increase risk forovereating and obesity. J Neurosci 28:12107–12119
44. García AP, Palou M, Priego T, Sánchez J, Palou A, Picó C 2010Moderate caloric restriction during gestation results in lower arcu-ate nucleus NPY- and alpha MSH-neurons and impairs hypotha-lamic response to fed/fasting conditions in weaned rats. DiabetesObes Metab 12:403–413
45. Kokoeva MV, Yin H, Flier JS 2007 Evidence for constitutive neuralcell proliferation in the adult murine hypothalamus. J Comp Neurol505:209–220
46. Zhao C, Deng W, Gage FH 2008 Mechanisms and functional im-plications of adult neurogenesis. Cell 132:645–660
47. Kokoeva MV, Yin H, Flier JS 2005 Neurogenesis in the hypothal-amus of adult mice: potential role in energy balance. Science 310:679–683
48. Pierce AA, Xu AW 2010 De novo neurogenesis in adult hypothal-amus as a compensatory mechanism to regulate energy balance.J Neurosci 30:723–730
Get ready for the 2012 ABIM board exam in endocrinology, diabetes, and metabolism with Endocrine Board Review, 3rd edition.
http://www.endo-society.org/brdrvw
Endocrinology, August 2012, 153(8):3657–3667 endo.endojournals.org 3667