flow and transport in porous tissues part i
TRANSCRIPT

Flow and Transport in Porous TissuesPart I
Alberto Bellin, DICAM
February 18, 2020
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 1 / 37

Contents
1 Overview of the cardiovascular system
2 The Endothelial cell
3 Microcirculation and solute exchange
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 2 / 37

References
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 3 / 37

References (continued)
J. R. Levick, An Introduction to Cardiovascular Physiology, HodderArnold, and Hachette Company, London, UK, 2000
R. L. Fournier, Basic Transport Phenomena in BiomedicalEngineering, CRC Press, Boca raton, FL, 2012
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 4 / 37

Chapter 1 of J. R. Levick, Introduction to CardiovascularPhysiology
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 5 / 37

Transport processes in the body system
The heart and blood vessels evolved to transport oxygen andnutrients rapidly;
material transport from the bloodstream to the cells relies on diffusion(τ ' x2/(2D)), where D is the diffusion coefficient(D ' 0.6× 10−9 ÷ 2× 10−9m2/s for diluite aqueous solutions and10−11 ÷ 10−10m2/s for biological molecules);
diffusion is a passive transport process and it is driven by the rapidrandom thermal motion of molecules;
in the presence of a concentration gradient the random movement ofmolecules results in a net diffusive transport in the direction of theconcentration gradient;
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 6 / 37
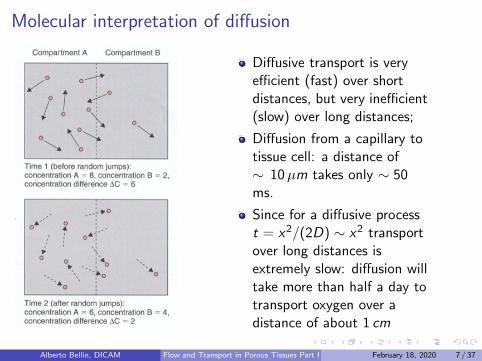
Molecular interpretation of diffusion
Diffusive transport is veryefficient (fast) over shortdistances, but very inefficient(slow) over long distances;
Diffusion from a capillary totissue cell: a distance of∼ 10µm takes only ∼ 50ms.
Since for a diffusive processt = x2/(2D) ∼ x2 transportover long distances isextremely slow: diffusion willtake more than half a day totransport oxygen over adistance of about 1 cm
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 7 / 37
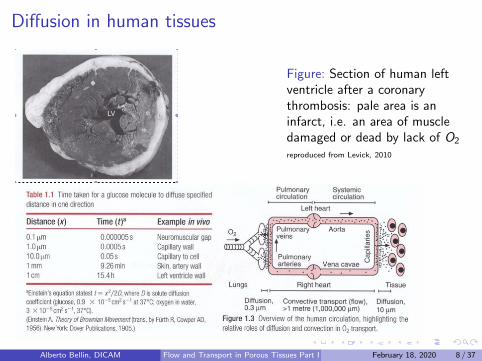
Diffusion in human tissues
Figure: Section of human leftventricle after a coronarythrombosis: pale area is aninfarct, i.e. an area of muscledamaged or dead by lack of O2
reproduced from Levick, 2010
.
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 8 / 37

Functions of the cardiovascular system
Primary function of the cardiovascular system (CVS): rapidconvective transport. O2 in the blood stream takes ∼ 30 s travelfrom the lungs to the smallest blood vessels (about 1m apart),whereas diffusion would take more than five years!
the CVS is also a control system: it distributes hormones to thetissues and secret bioactive agents (nitric oxide, etc.)
the CVS is crucial for body temperature regulation
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 9 / 37
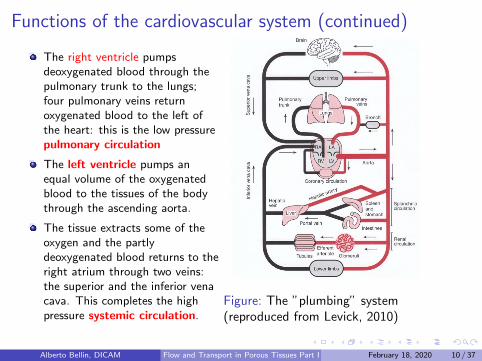
Functions of the cardiovascular system (continued)
The right ventricle pumpsdeoxygenated blood through thepulmonary trunk to the lungs;four pulmonary veins returnoxygenated blood to the left ofthe heart: this is the low pressurepulmonary circulation
The left ventricle pumps anequal volume of the oxygenatedblood to the tissues of the bodythrough the ascending aorta.
The tissue extracts some of theoxygen and the partlydeoxygenated blood returns to theright atrium through two veins:the superior and the inferior venacava. This completes the highpressure systemic circulation.
Figure: The ”plumbing” system(reproduced from Levick, 2010)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 10 / 37
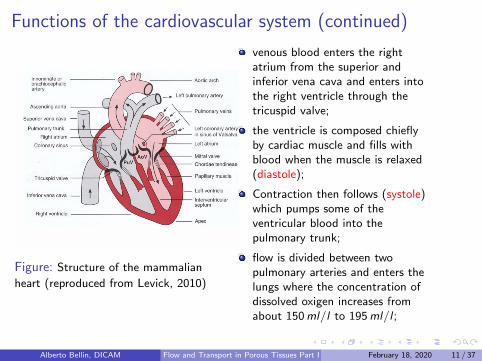
Functions of the cardiovascular system (continued)
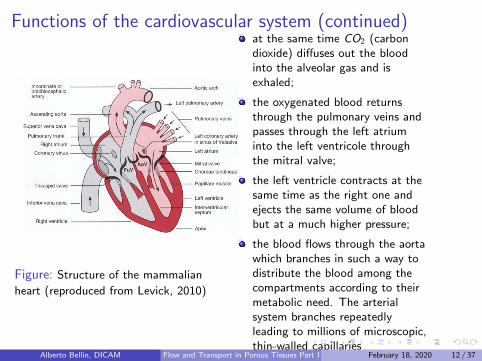
Figure: Structure of the mammalian
heart (reproduced from Levick, 2010)
venous blood enters the rightatrium from the superior andinferior vena cava and enters intothe right ventricle through thetricuspid valve;
the ventricle is composed chieflyby cardiac muscle and fills withblood when the muscle is relaxed(diastole);
Contraction then follows (systole)which pumps some of theventricular blood into thepulmonary trunk;
flow is divided between twopulmonary arteries and enters thelungs where the concentration ofdissolved oxigen increases fromabout 150ml/l to 195ml/l ;
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 11 / 37
Functions of the cardiovascular system (continued)
Figure: Structure of the mammalian
heart (reproduced from Levick, 2010)
at the same time CO2 (carbondioxide) diffuses out the bloodinto the alveolar gas and isexhaled;
the oxygenated blood returnsthrough the pulmonary veins andpasses through the left atriuminto the left ventricole throughthe mitral valve;
the left ventricle contracts at thesame time as the right one andejects the same volume of bloodbut at a much higher pressure;
the blood flows through the aortawhich branches in such a way todistribute the blood among thecompartments according to theirmetabolic need. The arterialsystem branches repeatedlyleading to millions of microscopic,thin-walled capillaries
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 12 / 37
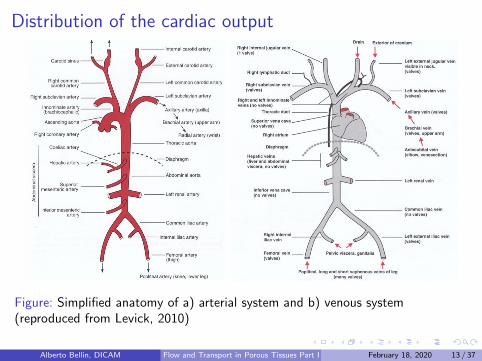
Distribution of the cardiac output
Figure: Simplified anatomy of a) arterial system and b) venous system(reproduced from Levick, 2010)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 13 / 37
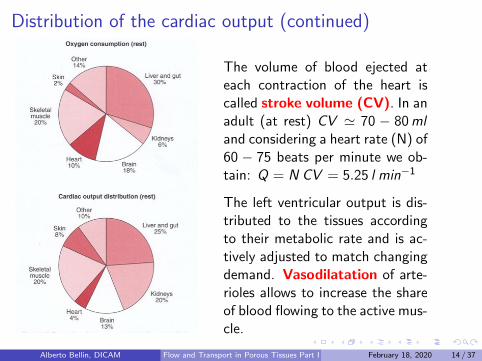
Distribution of the cardiac output (continued)
The volume of blood ejected ateach contraction of the heart iscalled stroke volume (CV). In anadult (at rest) CV ' 70 − 80mland considering a heart rate (N) of60 − 75 beats per minute we ob-tain: Q = N CV = 5.25 l min−1
The left ventricular output is dis-tributed to the tissues accordingto their metabolic rate and is ac-tively adjusted to match changingdemand. Vasodilatation of arte-rioles allows to increase the shareof blood flowing to the active mus-cle.
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 14 / 37

Elementary ”hydraulics” of the CVS
Flow is driven by the pressure induced by the heart: Arterial pressureis pulsatile because the heart eject blood intermittently (systole),with rest in between (diastole).
During diastole the systemic arterial pressure decays from a peak of∼ 120mmHg to ∼ 80mmHg ; In pulmonary artery the pressuredecays from 25mmHg to 10mmHg
flow depends on the diameter of the vessels
flow is typically laminar and in a first approximation blood can beconsidered as a Newtonian fluid, therefore Q = K ∆P, where K is theconductivity and ∆P is the variation of the pressure;
at the capillary level dissolved gasses, glucose and other metabolitesdiffuse between the capillary blood reaching the cells.
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 15 / 37
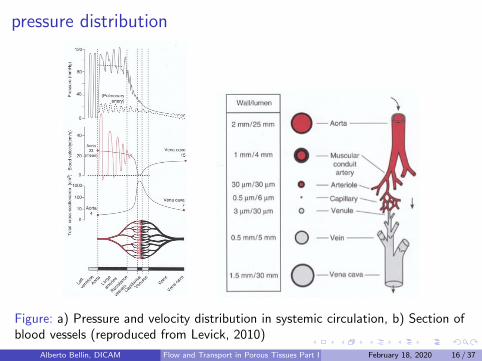
pressure distribution
Figure: a) Pressure and velocity distribution in systemic circulation, b) Section ofblood vessels (reproduced from Levick, 2010)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 16 / 37

Flow of a Newtonian incompressible fluid in a rigid tube
Starting point: Navier Stockes (NS) equation1:
ρ∂u
∂t+ ρu · ∇u = ρ g −∇p + µ∇2u (1)
where: u = (u, v ,w) [L/T ] is the fluid velocity vector, p[Force/L2] = [Mass/(LT 2)] is the pressure, ρ [Mass/L3] is the fluiddensity, µ [Mass/(LT )] is the fluid viscosity and g [L/T 2] is the vector ofthe gravity acceleration.
Modified pressure P is defined as follows: P = p − p0 − ρ g · xTherefore NSE becomes:
ρ∂u
∂t+ ρu · ∇u = −∇P + µ∇2u (2)
1see, Batchelor, G.K, An Introduction to Fluid Dynamics, Cambridge UniversityPress, 1967
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 17 / 37

Steady unidirectional flow
Hyp:(1) the velocity vector has the same direction everywhere and isindependent on the distance in the flow direction x : u · ∇u = 0;(2) the flow is steady: ∂u/∂t = 0Under these conditions the NSE simplifies to:
∂P
∂x= µ
(∂2u
∂y2+∂2u
∂z2
)(3)
with:∂P
∂y= 0,
∂P
∂z= 0, v = w = 0 (4)
Or:
∂2u
∂y2+∂2u
∂z2= −G
µ(5)
−G is a constant pressure gradient.
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 18 / 37

Poiseuille flow
For a long (horizontal) tube of circular section under the action of apressure difference between the two ends Equation (5) reads as:
1
r
d
dr
[rd u
d r
]= −G
µ(6)
which solved yields:
u =G
4µ
(−r2 + A ln r + B
)(7)
The singularity at r = 0 can be eliminated by imposing A = 0 and theconstant B is obtained by imposing u = 0 for r = a, where a is radius ofthe tube:
u =G
4µ
(−r2 + a2
)(8)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 19 / 37

Poiseuille flow (continued)
The flux of volume passing any section of the tube (discharge) is given by:
Q =
∫ a
0u 2π r dr =
π G a4
8µ=π a4(p0 − p1)
8µ l(9)
p0 = p(x = 0), p1 = p(x = l), where l is the length of the tube.The tangential stress at the wall of the tube is:
τ(a, θ) = µ
(d u
d r
)r=a
= −1
2G a
the tangential stress does not change with the angle θ.The total frictional force, in the direction of flow, on a length l of the tubeis:
F = −∫ 2π
0τ(a, θ) a d θ = 2π a l(
1
2G a) = π a2(p0 − p1)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 20 / 37

Poiseuille flow (continued)
As expected the total frictional force on the tube is equal to the differencebetween the total force at the inlet (π a2 p0) and at the outlet (π a2 p1)sections of the tube.The rate at which mechanical energy is dissipated by viscosity, per unitmass of fluid, is:
Φ =µ
ρ
(d u
d r
)2
=G 2 r2
4 ρµ
The total rate of dissipation in the fluid instantaneously filling the length lof the tube is: ∫ a
0Φ 2π r ρ dr =
π l G 2 a4
8µ= l Q G
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 21 / 37

Peripheral Resistance
The volumetric flux in the tube (9) can be written as follows:
Q = K (p0 − p1), where K = π a4
8µ l is the conductance. On the other hand,resistance can be defined as follows: R = 1/K .
Basic rules for resistances in series and
parallel (reproduced from Levick, 2010)
For n resistances in series theequivalent conductance is:
Keq,s =1∑n
i=1 Ri=
1∑ni=1 1/Ki
For n resistances in parallel theequivalent conductance is:
Keq,p =n∑
i=1
Ki
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 22 / 37

Effective conductance for elements in series
n elements connected in series (i.e. vessels with different radius) areinterested by the same flux Q; the pressure drops between the two ends ofthe j−th resistance is pj−1 − pj , j = 1, ..., n. Therefore:
Q = K1(p0 − p1) = K2(p1 − p2) = ....... = Kn(pn−1 − pn)
or equivalently:
p0 − p1 =Q
K1; p1 − p2 =
Q
K2; ......; pn−1 − pn =
Q
Kn
Summing up all the left and right hand terms of these equations leads to:
(p0 − p1) + (p1 − p2) + .......+ (pn−1 − pn) = Q
[1
K1+
1
K2+ ......+
1
Kn
]
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 23 / 37

Effective conductance for elements in series (continued)
Since the left hand term sums up to p0 − pn, i.e. the total pressure drop,the following relationship can be established between conductivities of thevessels and an effective global conductance:
p0 − pn = Qn∑
j=1
1
Kj=
Q
Keff
Therefore:
Keff =1∑n
j=11
Kj
(10)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 24 / 37

Effective conductance for elements in parallel
n elements connected in parallel are subjected to the same pressure drop∆p = p0 − p1 and each of them conducts a fraction of the total discharge:
Q1 = K1 ∆p, Q2 = K2 ∆p, ........, Qn = Kn ∆p
Thus, the total discharge assumes the following expression:
Q =n∑
j=1
Kj ∆ p = Keff ∆ p
Therefore:
Keff =n∑
j=1
Kj
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 25 / 37

Global resistances
The total resistance of the human systemic circulation is∼ 0.02mmHg minml−1 (pressure is measured in Pa = N/m2 in SIunits; 1mmHg = 133.32Pa = 0.0013332 bar = 0.0013 atm
The resistance of the pulmonary circulation is∼ 0.003mmHg minml−1, because pulmonary vessels are shorter andwider than their systemic counterpart.
changes in vascular resistance regulate local blood flow
As blood traverses the arterial tree, the increase of the number n ofvessels through branching outweighs the reduction of the vessel size(cross-sectional area π a2): the total cross-sectional area of thecirculation increases from aorta to capillary bed. The length ofthe capillaries in a adult human sums up to 40 000 km
Capillary blood velocity is ∼ 1/200 th of arterial blood velocity, dueto the vast increase in cross-sectional area. Slow flow gives to the redcells enough time to exchange oxygen and carbon dioxide.
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 26 / 37
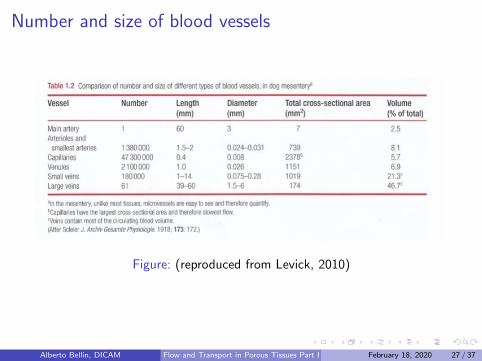
Number and size of blood vessels
Figure: (reproduced from Levick, 2010)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 27 / 37
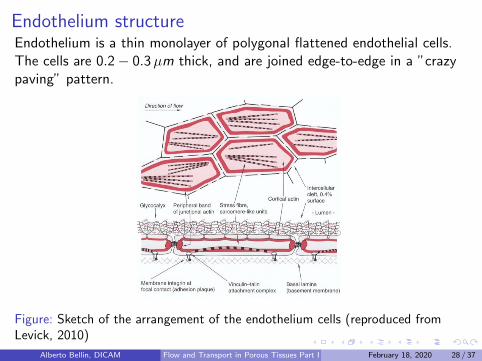
Endothelium structureEndothelium is a thin monolayer of polygonal flattened endothelial cells.The cells are 0.2− 0.3µm thick, and are joined edge-to-edge in a ”crazypaving” pattern.
Figure: Sketch of the arrangement of the endothelium cells (reproduced fromLevick, 2010)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 28 / 37

Functions of the endotheliumEndothelium regulates blood-tissue exchangeEndothelial surface enzymes modify vasoactive peptides in thebloodstreamEndothelium secretes both antithrombotic and pro-clotting factorEndothelium participates in the inflammatory defense againspathogensEndothelium initiates new blood vessel formationEndothelial dysfunction contributes to atheroma (basis for the diseaseentity atherosclerosis)
Figure: Sketch of the reduction of arterial lumen due to atheroma
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 29 / 37
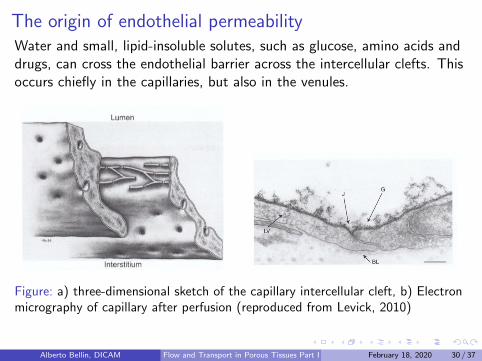
The origin of endothelial permeabilityWater and small, lipid-insoluble solutes, such as glucose, amino acids anddrugs, can cross the endothelial barrier across the intercellular clefts. Thisoccurs chiefly in the capillaries, but also in the venules.
Figure: a) three-dimensional sketch of the capillary intercellular cleft, b) Electronmicrography of capillary after perfusion (reproduced from Levick, 2010)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 30 / 37

The origin of endothelial permeability (continued)
Geometry of the clefts: 20 nm wide and occupy only 0.2− 0.4% ofthe capillary surface.
Junctional strands composed by rows of protein particles emergingfrom the facing cell surfaces: they overlap forming the tight junctions
The endothelium is coated internally by a hydrated gel, with athickness of 60− 570 nm, called glycocalyx.
Glycoalyx is composed by an assemblage of carbohydrate polymericfilaments and form a size-selective and charge-selective molecularsieve that is permeable to water and other small molecules, but not toplasma proteins or at herogenic low-density lipoproteins: glycocalyxmakes the endothelium semipermeable.
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 31 / 37

Glycocalyx functions
Glycocalyx is mechano-sensor, i.e. it senses changes in blood flow: forexample the flow-stimulated secretin of nitric oxide by endothelium isimpaired if glycocalyx is degraded.
The glycocalyx lubrificates the deformed red cells while squeezing in asingle file along capillary
Glycocalyx can degrade and shed into the bloodstream in response to:a) ischaemia-reperfusing injury, b) atrial natriuretic peptide, c)atherogenic oxidized low-density lipoprotein (LDL) and d) atheroma
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 32 / 37
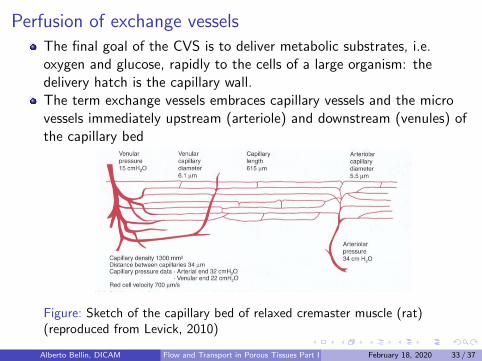
Perfusion of exchange vesselsThe final goal of the CVS is to deliver metabolic substrates, i.e.oxygen and glucose, rapidly to the cells of a large organism: thedelivery hatch is the capillary wall.The term exchange vessels embraces capillary vessels and the microvessels immediately upstream (arteriole) and downstream (venules) ofthe capillary bed
Figure: Sketch of the capillary bed of relaxed cremaster muscle (rat)(reproduced from Levick, 2010)
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 33 / 37

Capillary bedCapillary density is adapted to tissue function: skeletal muscle contains300-1000 capillaries per mm2 cross-section, or 1-3 capillaries per musclefibre. Endurance training stimulates angiogenesis and can rise capillary/fibre ratio to 6-8. In the myocardium and brain, where metabolic rate ishigh, the capillary density is also high ∼ 3000 per mm2.
High capillary density increases the endothelial surface area availablefor gas and nutrient exchange. Capillary surface: from ∼ 100 cm2 pergram in skeletal muscle to ∼ 500 cm2 per gram in myocardium orbrain
high capillary density reduces the distance between the bloodstreamand the cells
capillaries can be well perfused by blood or ”closed” depending onwhether the individual arteriole connected to them is relaxed orcontracted. In a skeletal muscle some arterioles are relaxed and otherare contracted at the same time leading to a heterogeneous tissueperfusion
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 34 / 37

Capillary transit time
Individual arteriole alternates between contracted and relaxed states.For example in skeletal muscle the cycle time is of ∼ 15 s and isregular; in other tissue, such as skin, vasomotor is less regular.
The time a cell takes to pass through (travel) the capillary bed iscalled the transit time. Transit time is the time allowed to blood tounload oxygen, glucose and other substances and upload urea CO2,etc. Transit time changes because of vasomotor. However, a typicaltransit time in a well perfused capillary is 0.5− 2 s, resulting in ablood velocity of 300− 1000µm/s. During exercise transit time mayreduce to 0.25 s, because arterioles dilate and blood velocity increases.
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 35 / 37
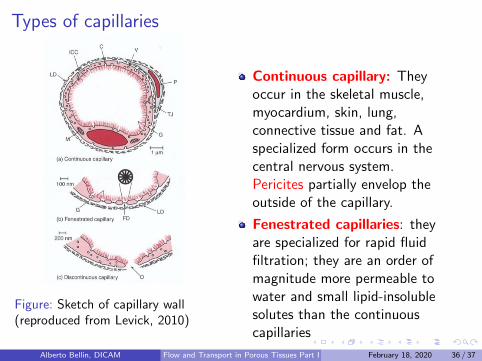
Types of capillaries
Figure: Sketch of capillary wall(reproduced from Levick, 2010)
Continuous capillary: Theyoccur in the skeletal muscle,myocardium, skin, lung,connective tissue and fat. Aspecialized form occurs in thecentral nervous system.Pericites partially envelop theoutside of the capillary.
Fenestrated capillaries: theyare specialized for rapid fluidfiltration; they are an order ofmagnitude more permeable towater and small lipid-insolublesolutes than the continuouscapillaries
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 36 / 37

Types of capillaries (continued)
Continuous capillary exchange processes: The intercellular clefttransmit water and small lipid-insoluble solutes; The luminalGlycocalyx excludes plasma proteins from access to the cleft(however, it is permeable to water and small solute);
Fenestrated capillary: They are found in tissues that specialize influid exchange (kidneys, exocrine glands, intestinal mucosa, synoviallining of joints, choroid plexus, ciliary body of the eye) and inendocrine glands. The endothelium is perforated by clusters of small,circular windows, or fenestrae, of diameter 50-60 nm. Each fenestra isbridged by an extremely thin membrane, the diaphragm, of thickness4-5 nm, except in renal glomerular capillaries, where the diaphragm isabsent.
Discontinuous capillaries allow blood cell turnover. They haveendothelial gaps over 100 nm wide with the corresponding opening inthe basal lamina. They are highly permeable, even to plasma proteins.
Alberto Bellin, DICAM Flow and Transport in Porous Tissues Part I February 18, 2020 37 / 37