exploration of phage-host interactions in the fish pathogen vibrio
TRANSCRIPT

U N I V E R S I T Y O F C O P E N H A G E N
F A C U L T Y O F S C I E N C E
D E P A R T M E N T O F B I O L O G Y
M A R I N E B I O L O G I C A L S E C T I O N
Exploration of phage-host interactions in the fish pathogen Vibrio anguillarum and anti-phage defense strategies
PhD thesis
Demeng Tan
Academic advisor: Mathias Middelboe
Submitted: 16/03/2015

Academic Supervisor:
Professor Mathias Middelboe
Marine Biological Section
Department of Biology
University of Copenhagen
Denmark
Assessment committee:
Reader Martha Clokie
Department of Infection, Immunity & Inflammation
University of Leicester
United Kingdom
Associate Professor Niels-Ulrik Frigaard (Chair)
Marine Biological Section
Department of Biology
University of Copenhagen
Denmark
Associate Professor Lars Jelsbak
Infection Microbiology
Department of Systems Biology
Technical University of Denmark
Denmark
Submitted:
March 16, 2015

1
Contents
Contents ............................................................................................................................................................ 1
Preface ............................................................................................................................................................... 3
Acknowledgements ........................................................................................................................................... 4
Abstract ............................................................................................................................................................. 5
Resumé .............................................................................................................................................................. 7
List of abbreviations .......................................................................................................................................... 9
Introduction ..................................................................................................................................................... 10
1. Aquaculture ............................................................................................................................................. 10
1.1. Current status of aquaculture .......................................................................................................... 10
1.2. Bacterial diseases in aquaculture ..................................................................................................... 10
1.3. Vibriosis and Vibrio anguillarum ...................................................................................................... 11
2. Bacterial communication ......................................................................................................................... 13
2.1. Quorum sensing in Gram-negative bacteria..................................................................................... 13
2.2. Quorum sensing in V. anguillarum ................................................................................................... 14
3. Bacterial biofilm formation and its role in infections .............................................................................. 17
3.1. Biofilm structure ............................................................................................................................... 17
3.2. Biofilm development and variation in biofilm formation ................................................................. 18
3.3. Heterogeneity in biofilms ................................................................................................................. 20
3.4. Diagnosis of biofilms ......................................................................................................................... 21
3.5. Treatment of vibriosis in aquaculture .............................................................................................. 22
4. Alternative disease control in aquaculture ............................................................................................. 24
4.1. Bacteriophages ................................................................................................................................. 24
4.2. Phage classification ........................................................................................................................... 24
4.3. Bacteriophage life cycle .................................................................................................................... 25
4.4. Vibriophages ..................................................................................................................................... 26
4.5. Application of phages to control fish pathogens: Phage therapy .................................................... 28
5. Bacteriophage resistance mechanisms ................................................................................................... 30
5.1. Phage receptor modification ............................................................................................................ 30
5.2. Cutting phage nucleic acids .............................................................................................................. 32

2
5.3. CRISPR/Cas bacterial immune system cleaves bacteriophage DNA ................................................. 32
5.4. Abortive infection (ABi) system ........................................................................................................ 32
5.5. Regulation of phage-host interactions by extracellular signaling molecules ................................... 33
5.6. Implications of phage protection mechanisms: Phage-host coexistence and co-evolution ............ 33
Aims of this Ph.D. project ................................................................................................................................ 35
A summary of this Ph.D. project ...................................................................................................................... 36
Paper I .......................................................................................................................................................... 36
Paper II ......................................................................................................................................................... 37
Paper III ........................................................................................................................................................ 38
Future perspectives ......................................................................................................................................... 39
Paper I .......................................................................................................................................................... 39
Paper II ......................................................................................................................................................... 41
Paper III ........................................................................................................................................................ 43
References ....................................................................................................................................................... 44
Accompanying papers ..................................................................................................................................... 54

3
Preface The project presented in this PhD was carried out under the supervision of Prof. Mathias Middelboe
primarily at the Marine Biological Section, Department of Biology, University of Copenhagen, Denmark.
The work was done in collaboration with Prof. Lone Gram (Department of Systems Biology, Technical
University of Denmark) and Dr. Sine Lo Svenningsen (Department of Biology, University of Copenhagen).
List of publication and manuscripts
1. Demeng Tan, Lone Gram, and Mathias Middelboe. "Vibriophages and their interactions with the
fish pathogen Vibrio anguillarum." Applied and Environmental Microbiology 80.10 (2014): 3128-
3140.
2. Demeng Tan, Amalie Dahl, and Mathias Middelboe. "Vibriophages differentially influence biofilm
formation by Vibrio anguillarum strains." Applied and Environmental Microbiology. (accept,
AEM00518-15)
3. Demeng Tan, Sine Lo Svenningsen, and Mathias Middelboe. "Quorum sensing determines the
choice of anti-phage defense strategy in Vibrio anguillarum." mBio. (in revision, mBio00627-15)

4
Acknowledgements
Studying abroad is probably one of the best things that I have ever done in my life so far. This experience
has not just involved learning scientific skills, but it has also taught me a great deal about independence,
taking initiative, and self-motivation, and has helped me develop a wide range of social skills. I am indebted
to many people for supporting me through the numerous stages of the past three years. Unfortunately, I
will not mention everybody’s name, but there are a few people who deserve a special mention here. To
those who deserve but did not receive mention here, and realized my omission, I thoroughly apologize.
First of all, I want to thank my supervisor, Prof. Mathias Middelboe, for offering me such a valuable
opportunity to let me do my Ph.D. studies with you. You have taught me so many invaluable things, and I
have learned so many important lessons.
I would also like to thank Dr. Sine Lo Svenningsen, for letting me stay in your lab. It has been a
tremendously wonderful time, and I have learned a lot about Molecular Biology and Quorum Sensing.
Thanks again for your patience and enthusiasm. Hopefully, we will have more collaboration in the future
about Quorum Sensing studies.
I am also grateful to Prof. Lone Gram for making our first paper possible.
Hannah Eva Blossom, thank you for proofreading my Introduction Chapter.
I would not have finished my Ph.D. without the love and support of my family, friends and relatives in China.
I wish I could have spent more time with you during the past three years.
And last but not least, my host family in Snekkersten. I cannot express to you how much I appreciated you
over the past three years, with so many nice memories, and your effort to make Sprydet 83 feel like home.

5
Abstract
The disease vibriosis is caused by the bacterial pathogen Vibrio anguillarum and results in large losses in
aquaculture both in Denmark and around the world. Antibiotics have been widely used in antimicrobial
prophylaxis and treatment of vibriosis. Recently, numerous multidrug-resistant strains of V. anguillarum
have been isolated, indicating that antibiotic use has to be restricted and alternatives have to be developed.
Lytic phages have been demonstrated to play an essential role in preventing bacterial infection. However,
phages are also known to play a critical role in the evolution of bacterial pathogenicity development.
Therefore, successful application of phage therapy in the treatment of vibriosis requires a detailed
understanding of phage-host interactions, especially with regards to anti-phage defense mechanisms in the
host.
Part I. As a first approach, 24 V. anguillarum and 13 Vibrio sp. strains, representing considerable temporal
(20 years) and geographic (9 countries) variation in regards to their origins of isolation, and 11 vibriophages
representing three different families (Myoviridae, Siphoviridae, and Podoviridae), were characterized with
respect host range, morphology, genome size and lytic properties. Together the host range of the 11
vibriophages covered all the 37 Vibrio strains in the collection. In addition, the occurrence of unique
susceptibility patterns of the individual host isolates, as well as key phenotypic properties related to phage
susceptibility that were common to the isolated strains were studied.
Part II. In vitro phage-host interactions in two V. anguillarum strains (BA35 and PF430-3) and their
corresponding phages (ΦH20 (Siphoviridae) and KVP40 (Myoviridae)), during growth in the settings of
micro-colonies, biofilms, and the free-living phase were investigated, in order to explore the
resistance/tolerance mechanisms responsible for the different outcomes of these two phage-host
interactions. The study demonstrated large intraspecific differences in phage-protection mechanisms, as
strain BA35 obtained genetic resistance through mutational changes, whereas strain PF430-3 was
protected from phage infection by formation of a biofilm. These different protection mechanisms have
important implications for the phage-host dynamics and efficiency of phage infection.
Part III. To gain an insight into whether bacteria of the Vibrio genus use quorum sensing (QS) to control
their phage-susceptibility, a study of the QS-mediated interaction between phage KVP40 and V.
anguillarum PF430-3 was carried out. The data show that QS does indeed regulate phage-host interactions
in V. anguillarum PF430-3, potentially by using QS transcription factor VanT to repress ompK expression. It
was demonstrated that QS controls the choice of anti-phage defense strategies in the V. anguillarum strain

6
PF430-3, suggesting the presence of dynamic, temporary adaptations to phage infection pressure, while
still securing the ability to produce a functional OmpK receptor.
In conclusion, this thesis provides a first insight into the dynamic vibriophage-host interactions, indicating
the complexity of phage therapy in the treatment of vibriosis, regarding the evolution of anti-phage
defense mechanisms, gene regulation, quorum sensing, biofilm formation, as well as pathogenesis.
Together, these discoveries will enable us to evaluate these potential factors which regulate phage-host
interactions, and on that background, to optimize phage therapeutic strategies against vibriosis with
regards to the composition of applied phage cocktails, the timing of treatment, and the likely outcome of
phage therapy and QS inhibiting molecules.

7
Resumé
Vibrio anguillarum er en vigtig patogen bakterie i akvakultur, hvor den forårsager sygdommen vibriose, som
resulterer i fiskedød og store økonomiske tab i akvakulturindustrien i både Danmark og hele resten af
verden. Typisk bruges antibiotika i behandlingen af vibriose, men forøget forekomst af antibiotika-
resistente V. anguillarum har medført øget fokus på behovet for alternative behandlingsmetoder.
Anvendelse af lytiske bakteriofager har de senere år vist sig som et potentielt alternativ til antibiotika ifm
forebyggelse og behandling af fiskeinfektioner. Denne tilgang er dog også forbundet med udfordringer og
begræsninger, dels fordi fagerne kan spille en rolle i bakteriers patogene egenskaber og dels fordi bakterie
også udvikler resistens overfor fager. Succesful anvendelse af bakteriofager i sygdomsbehandling kræver
derfor et grundigt kendskab til fag-vært interaktioner og specielt en forståelse af anti-fag
forsvarsmekanismer i værtsbakterien.
Del 1. Som en første tilgang dette undersøgtes 24 V. anguillarum og 13 Vibrio sp., stammer som
repræsenterede en betydelige tidsmæssige (20 år) og geografiske (9 forskellige lande) forskellige med
hensyn til stedet for deres oprindelige isolation, samt 11 vibriofager, der repræsenterede 3 forskellige
moroflogiske familier (Myoviridae, Siphoviridae, and Podoviridae). Bakteriofagerne blev karakteriseret med
hensyn til "host range", morfologi, genomstørrelse og lytiske egenskaber, og tilsammen kunne de 11 fag-
isolater inficere alle 37 V. anguillarum stammer i stammesamlingen. Derudover blev mønstre i
inficerbarhed overfore fagerne samt egenskaber associeret med resistens overfor disse undersøgt hos
værtsstammerne.
Del 2. In vitro studier af fag-værtinteraktioner hos to V. anguillarum stammer (BA35 and PF430-3) og deres
associerede fager (ΦH20 (Siphoviridae) og KVP40 (Myoviridae)) blev undersøgt under forskellige vækstfaser
(mikrokolonier, biofilm og fritlevende) med det formål at kortlægge resistens/tolerance mekanismer i de to
stammer. Studiet dokumenterede store intraspecifikke forskelle i de underliggende mekanismer for
beskyttelse mod fager i de to stammer. Stamme BA35 udviklede genetisk resistens gennem mutationer
mens stamme PF430-3 beskyttedes mod fag-infektioner ved dannelse af biofilm. Disse forskelle i
beskyttelsesmekanismer havde store implikationer for fag-vært dynamik og infektions-effektivitet af de to
fager.
Del 3. I dette studie undersøgtes i hvilket omfang Vibrio anguillarum stamme PF430-3 anvender "quorum
sensing" (QS) i reguleringen af tolerance overfor faginfektion. Data påviser at QS regulerer interaktionerne
mellem fag KVP40 og V. anguillarum PF430-3, idet QS medierer en aktivering af transkriptionsfaktor VanT
som leder til en nedjustering af ekspressionen af ompK, som er receptor for KVP40. Arbejdet viser at QS kan

8
kontrollerer skift i anti-fag strategi hos V. anguillarum strain PF430-3, som afhænger af de givne
vækstbetingelser. Disse skift i forsvarsmekanisme indikerer meget dynamiske tilpasninger til fag-
infektionstryk I V. anguillarum PF430-3, som samtidigt sikrer at stammen kan opretholde en funktionel
ompK receptor.
Denne afhandling bidrager således med ny viden om de dynamiske og komplekse interaktioner mellem
vibriofager og deres værtsceller og understreger betydningen af en bedre forståelse af disse i relation til
den løbende udforskning af nye fag-baserede behandlingsmetoder. Disse opdagelser kan således anvendes
i vurderingen af de faktorer der regulerer fag-resistens under forskellige forhold og kan dermed bidrage til
en optimering af fag-anvendelsen i behandling af vibriose, specielt i forhold til sammensætning af fag
cocktails, timing af fag-behandling og potentielt anvendelse af QS hæmmende substrater.

9
List of abbreviations
Abi Abortion infection system
AHLs N-acyl-L-homoserine lactone
AI Autoinducer
CAI-1 cholerae autoinducer-1
cas CRISPR-associated
CRISPR Clustered regularly interspaced short palindromic repeats
CT Cholera toxin
CVEC Conditionally viable environmental cells
CWD Cold water disease
eDNA Extracellular DNA
EMSA Electrophoretic mobility shift assay
EPS Extracellular Polymeric Substances
ERIC Enterobacterial repetitiveelement intergenic consensus sequence
FISH Fluorescence in situ hybridization
ICTV International Committee on Taxonomy of Viruses
LPS Lipopolysaccharide
MLST Multi-locus sequence typing
MRSA Methicillin-resistant S. aureus
OA Oxolinic acid
Omp Outer membrane protein
PFGE Pulsed-field gel electrophoresis
qrr Quorum sensing regulatory RNA
QS Quorum sensing
R-M Restriction-Modification
VBNC Viable but non-culturable
vps Vibrio polysaccharide

10
Introduction
1. Aquaculture
1.1. Current status of aquaculture
A daunting challenge of our time is to ensure adequate nutritional food to the expanding human population
(1). Over the past 50 years, the aquaculture industry has become one of the fastest growing industries
globally, with an annual growth rate of 8.3%. The aquaculture industry not only provides healthy food for
human consumption, but it is extremely important for local economies which rely heavily on the sales of
high valued marine fish and crustaceans (1). According to Figure 1, at the end of 2012, aquaculture
production has already reached 66.6 million tons per year (1). With declining natural fish stocks and global
fisheries, aquaculture plays a vital role in feeding the planet, today and in the future.
Figure 1. World capture fisheries and aquaculture production from 1950 to 2012 (1).
Aquaculture is defined as farming aquatic organisms including fish, molluscs, crustaceans and aquatic
plants. In addition, aquaculture farming not only involves interventions in the rearing process to enhance
production, such as stock, feeding, protection from predators, but also implies stocks cultivation (1).
Despite the fact that aquaculture is a well-controlled and organized industry, problems such as disease
outbreaks can still occur. Disease outbreaks are considered as one of the most significant constraints to
sustainable aquaculture growth (2).
1.2. Bacterial diseases in aquaculture
Fish are susceptible to a wide range of bacterial pathogens. Some of the most important opportunistic
bacterial diseases are vibriosis, salmon rickettsial syndrome (SRS), furunculosis, and cold water disease
(CWD), which is caused by Vibrio anguillarum, Piscirickettsia salmonis, Aeromonas salmonicida, and

11
Flavobacterium psychrophilum, respectively (3-5). These bacterial cells are naturally occurring in the
environment but become serious pathogens when fish hosts are physiologically unbalanced,
immunocompromised or exposed to other stressors, i.e., poor water quality, overstocking, and overfeeding,
etc., which then allows opportunistic bacterial infections to progress (6). Once the abundance of these
opportunistic pathogens reaches a threshold level, they can cause disease. Hence, farmed fish in many
cases are affected by these bacterial diseases, resulting in dynamic unpredictable mortality rates, and
subsequently, severe economic losses. Thus, disease management in aquaculture is becoming more
complex and challenging.
1.3. Vibriosis and Vibrio anguillarum
Vibriosis, a highly fatal haemorrhagic septicaemia, is one of the most prevalent fish diseases and has been
found in more than 50 fresh and salt-water fish species including various species of economic importance
to the larviculture and aquaculture industry, especially during the larval and juvenile stages. These species
include salmon (Salmo salar), rainbow trout (Onchorhynchus mykiss), turbot (Psetta maxima), European sea
bass (Dicentrarchus labrax), gilthead sea bream (Sparus aurata) and ayu (Plecoglossus altivelis) (7). The
pathogen can invade the host either via contaminated food or through the skin epithelial cells, while the
infection is then triggered due to environmental factors commonly found in fish farms such as high
population density or poor water quality (8). Typical external clinical symptoms of the disease include
weight loss, lethargy, and development of red spots on the ventral and lateral areas of the fish, with
swollen, dark skin lesions causing ulceration and bleeding (Figure 2) (9). Nevertheless, in acute pandemic
infections the infection spreads so rapidly that most of the infected fish die even without evidence of any
clinical symptoms (10). Previous studies have reported that V. anguillarum, as one of the most prominent
fish and shellfish pathogens, is associated with vibriosis infection in aquaculture (11-13).
Figure 2. Clinical symptoms of vibriosis infection a sea bass causing haemorrhagic septicaemia on the fin and around the operculum,
figure adapted from Austin (10).

12
V. anguillarum is a Gram-negative, halophilic and facultative anaerobic bacterium, which grows rapidly in
the range of temperatures between 15 and 30 °C on rich media containing 1.5-2% NaCl. According to
Harrell’s study, V. anguillarum was initially divided into two distinct biotypes (1 and 2) (14). However,
through advances in DNA sequence technology, V. anguillarum biotype 1 was reclassified as Listonella
anguillarum, as a result of 5S ribosomal RNA gene sequence analysis (15). Nevertheless, because of its
strong similarities with other Vibrio species, it is generally still identified as V. anguillarum (16). Biotype 2
was reclassified as a new species, V. ordalii (11).
V. anguillarum is composed of 2 chromosomes (3.0 and 1.2 Mbp) with a whole genome size of about 4.2
Mbp and a G+C content of 43-46%. Most of the O1 serotype strains isolated harbor a plasmid which carries
many virulence factors, such as genes affecting chemotaxis and motility, an iron uptake system,
lipopolysaccharides (LPSs) and extracellular products with proteolytic or haemolytic activity (11). Recently,
the genome of the O1 serotype strain V. anguillarum 775 was sequenced. The genome annotation shows
that the majority of the genes for essential cell functions and pathogenicity are located in chromosome 1.
This chromosome also has 8 genomic islands, while chromosome 2 has only 2. It was found that some of
these islands carried potential virulence genes (17).
V. anguillarum strains show close similarities according to 16S rRNA sequences irrespective of time or place
of isolation, suggesting 16S rRNA is highly uniform and stable (18), indicating further genetic fingerprinting
tools such as enterobacterial repetitive element intergenic consensus (ERIC) PCR (19), multi-locus sequence
typing (MLST) (20), and pulsed-field gel electrophoresis (PFGE) (21) are needed for surveillance purposes,
monitoring outbreaks and tracking pathogen spreading.
Currently, 23 different O serotypes (O1-O23) within V. anguillarum are discriminated, each single O
serotype with its own different pathogenicity and host specificity. Among these, only serotypes O1, O2, and
O3, have been reported to associate with vibriosis infection in fish (22). Within serotype O2, three host-
specific sub-serotypes, O2a, O2b and O2c, have been distinguished by Rasmussen (23).

13
2. Bacterial communication
Bacteria have adapted to thrive in diverse ecological niches. Over the long-term evolution, they have
developed sophisticated mechanisms, allowing them to perceive and respond to dynamic environmental
conditions (24).
Quorum sensing (QS) is a ubiquitous cell-cell communication found in bacteria, thus far (25). By using this
QS system, bacteria coordinate their cell density-dependent gene expression to act in unison, via the
production, secretion and subsequent detection of extracellular signaling molecules, namely, autoinducers
(25). Detection of autoinducers allows bacterial cells to distinguish different behavioral modes, i.e. from
low cell densities to high cell densities (25).
2.1. Quorum sensing in Gram-negative bacteria
Most Gram-negative proteobacteria use acyl homoserine lactones (AHLs) as major auotoinducers for
interspecies communication, with homoserine lactone (HSL) rings carrying acyl chains of a range from C4 to
C18 in length (26). The first AHL was identified in bioluminescent bacteria (V. fischeri), which have a
symbiotic relationship with bobtail squid (Euprymna scolopes) (27). By using QS to activate expression of
the luciferase operon to counter-illuminate itself, the squid can easily escape from predators as a
cooperative symbiotic interaction between the squid and V. fischeri (25, 28, 29). The mechanism underling
this phenomenon has been fully characterized. Briefly, in the V. fischeri model, LuxI synthesizes QS
autoinducers, and LuxR acts as a cytoplasmic receptor for AHLs as well as transcription factor of the
luciferase gene (28, 30, 31). However, without AHLs ligand, LuxR is unstable and can be rapidly degraded.
Since AHLs can freely diffuse across cell membranes, their concentration increases correspond to the cell
density increases. By binding to the transcriptional factor LuxR, the AHLs-LuxR complexes turn on the
luxICDABE expression (Figure 3) (28, 32).
Figure 3. A canonical Gram-negative LuxIR-type QS circuits. Red pentagons denote AHL autoinducers. Figure adapted from Ng (28).

14
2.2. Quorum sensing in V. anguillarum
N-acyl homoserine lactone-mediated QS circuits have been identified in V. anguillarum, showing similarities
to many other Vibrio species in previous studies (28, 33-35). These circuits contain components for multiple
QS phosphoreplay system, and control QS-regulated genes via the transcription factor VanT, which is
activated in response to extracellular signaling molecules (35).
In general, at low cell densities, these auto-inducers are also at low concentrations, and the membrane-
binding sensors are kinases, VanN, VanQ and CqsS, which start autophosphorylation, and initiate a cascade,
transferring phosphate onto the phosphotransferase VanU, which phosphorylates the σ54-dependent
response regulator VanO (33, 36). Upon phosphorylation, VanO is activated together with the alternative
sigma factor RpoN, together with the RNA chaperone Hfp, which finally destabilizes vanT mRNA, repressing
expression of the QS transcriptional regulator VanT (33, 36).
At high cell densities, auto-inducer concentrations reach certain thresholds with vanT expression induced,
while VanO is inactivated (35). Specifically, binding of QS-signal molecules to the kinase sensor inhibits
kinase activity, which changes the receptor function from kinase to phosphatase activity, leading to the
dephosphorylation reactions and finally to inactivate VanO. Thus, QS regulatory RNA (qrr) is not expressed
while stabilizing the expression of VanT, leading to the QS-mediated gene regulation (33, 36). Figure 4 is a
model of V. anguillarum QS circuits.

15
Figure 4. V. anguillarum QS-circuits. Solid lines with arrows and bars represent activation and repression of gene expression,
respectively. Solid lines with double arrowheads represent the transfer of phosphoryl groups to each other. At low cell densities,
auto-inducers are at low concentrations. Upon phosphorylation, VanO is activated together with the alternative sigma factor RpoN,
which finally destabilizes vanT mRNA, repressing expression of the QS transcriptional regulator VanT. At high cell densities, auto-
inducers reach a certain threshold with vanT expression induced, while VanO is inactivated. OM and IM indicate outer membrane
and inner membrane, respectively. The P in the circle indicates that the protein is in the phosphorylated state (35).

16
In addition, several N-Acyl homoserine lactone auto-inducers (N-(3-Oxodecanoyl)-L-homoserine lactone, N-
(3-hydroxyhexanoyl)homoserine lactone, and N-hexanoylhomoserine lactone) (Figure 5) have been
identified in stationary-phase V. anguillarum cell-free supernatant at concentrations of approximately 8.5,
9.5, and 0.3 nM, respectively (37).
Figure 5. Structures of three different V. anguillarum autoinducers present in stationary phase cell-free culture supernatant,
namely, N-(3-Oxodecanoyl)-L-homoserine lactone, N-(3-hydroxyhexanoyl)homoserine lactone, and N-hexanoylhomoserine lactone.

17
3. Bacterial biofilm formation and its role in infections
Until recently, a bacterium cell has been considered as an individual cell without interactions with
neighboring cells. This dogma has been challenged by the discovery of the cell-density dependent
bioluminescence of the marine bacterium V. fischeri, and bacterial biofilm formation (27, 38, 39). These
types of gene regulatory phenomena are generally recognized as QS (25). As discussed above QS regulates
multiple bacterial social behaviors, such as bioluminescence (40), sporulation (41), competence (42),
virulence (43), motility (44), and phage-host interactions (45, 46), as well as bacterial biofilm formation (47).
The development of biofilm and QS are meticulously interconnected, as cooperative bacterial group
behaviors in a self-produced extracellular matrix are essential for biofilm formation and development (48).
It is, therefore, necessary to better understand the mechanisms of V. anguillarum evolution and
pathogenicity to develop better animal models and design new strategies and therapeutics for the control
of vibriosis.
During vibriosis infection, bacterial growth in planktonic form is relatively rare. In natural environments and
aquaculture, bacterial infection involves adhesion to the host organism in the form of a biofilm, sustaining
in the persistence of the infection during the vibriosis outbreak (49, 50). Studies have shown, after the first
stages of infection, V. anguillarum formed biofilm-like micro-colonies within the skin mucosal tissues of
rainbow trout, causing chronic infection and contributing to disease persistence in the face of antibiotic
treatment regimens, due to poor antibiotic penetration, nutrient limitation and slow growth, as well as
adaptive stress responses (51, 52). Thus, understanding of the role of bacterial biofilm during infection
should help the management of antibacterial therapy of relevant infections.
3.1. Biofilm structure
Biofilms are matrix enclosed microbial communities that can be established on a range of biological or non-
biological surfaces (53). The complex mixture of hydrated polymers, known as extracellular polymeric
substances (EPS), plays an important role in the function of biofilms, including retention of nutrients and
water as well as protection from antimicrobial agents such as antibiotics, protozoan grazing, host immune
systems and bacteriophages (53). Although polysaccharides are predominant components, EPS also contain
a variety of other substances, including protein, extracellular DNA (eDNA), membrane vesicles, and other
polymers. For instance, in the model organisms of strain Enterococcus faecalis, Staphylococcus aureus, P.
aeruginosa and V. cholerae, eDNA has been shown essential for maintaining the stability of saturated
biofilms (54-57). The structural role of eDNA in promoting biofilm formation is highly variable and depends
on the bacterial strains and growth conditions as well as the stages of the biofilm. The observation of
prophage-stimulated biofilm formation (58, 59), and demonstration that addition of DNase I at the

18
initiation stage of biofilm growth inhibited biofilm formation suggested that prophage mediated lysis
results in the release of eDNA enhancing biofilm formation. However, DNase I-treated experiments did not
affect phage KVP40-meduated V. anguillarum strain PF430-3’s biofilm formation, suggesting that eDNA
released from phage KVP40-mediated lysis did not serve as a critical structural component in the initial
stages of biofilm formation (Tan et al., unpublished). However, the results should be interpreted carefully,
since further work with more controls, such as DNase I inactivation and biofilm penetration, are needed.
3.2. Biofilm development and variation in biofilm formation
Biofilm formation is a multistep process (Figure 6), first involving surface-associated bacterial attachment
followed by immobilization, then combined unidirectional movement along the surface, and finally the
formation of three-dimensional micro-colonies (60). Meanwhile, these sessile biofilm communities can also
rapidly multiply and disperse to planktonic bacterial cells (61).
Figure 6. Five complex development processes involving biofilm maturation. Stage 1-5, initial attachment; irreversible attachment;
maturation I; maturation II; and dissemination, respectively. Figure adapted from Monroe’s study (62).
It turns out that there is a whole range of different types of biofilm development that bacteria use for a
variety of purposes. For instance, V. cholerae bacterium stops biofilm formation when a certain density of
bacterial cells gather together; while other bacterium such as P. aeruginosa bacterium makes biofilms
when the bacterial number reaches a certain density threshold.
P. aeruginosa biofilms, which is contrasted with cholera acute infection case, cause devastating chronic
lung infections in the immunocompromised hosts (63). QS signaling molecules, on the other hand, activate
gene transcriptions which facilitate biofilm formation at high cell densities (64). Another well-known
example of biofilm formation at high cell densities is that of E. coli, in which the physiological and
morphological development of its biofilms have been well studied. In the post-exponential growth phase of
E. coli, while nutrients are no longer optimal yet still available, E. coli cells stop producing flagella, and enter
into stationary phase, in a process of cellular aggregation (65). This aggregation imposes diffusion

19
constraints on material exchanges, preventing both entering or leaving the biofilm system, which impedes,
for example, antimicrobial substances and the interactions between phage receptor and phage binding
proteins (66). However, biofilm sessile cells can also be detached by seeding dispersal, and these detached
cells also involve the establishment of new biofilms, which is of fundamental importance to the
dissemination of pathogens in the infection model (67).
In V. cholerae cases, biofilm formation is activated at low cell densities (47), in contrast to other bacterial
pathogens that induce biofilm formation at high cell densities in the presence of QS signaling molecules.
The ability to produce biofilm at low cell densities not only protects the V. cholerae cells against the host
immune system and gastric acid, but it also defends against host internal bacteria, such as human gut
microbial communities (68). Once the cell densities reach high numbers, bacterial cells stop generating
biofilm, since biofilm production takes up valuable resources, which could otherwise be used in growth and
cell division (47). At high cell densities, QS-mediated expression of the hap protease gene, was shown to
promote release from the epithelium and escape into the environment for successful infection of new
hosts (69). The same phenomenon also occurred in the cultures of V. anguillarum strain PF430-3 and phage
KVP40 at high cell densities, detachment of cells from aggregates were disseminating into free-living
variants, which co-existed with phage KVP40.
Potential application of treatments that promoted dispersal of aggregated cells into the planktonic phase
seems attractive, as the results described in previous work indicated that even the use of pro- and anti-QS
therapies that function by modulating bacterial chemical communication circuits to weaken the biofilm and
lead the sessile cells into planktonic phase, where they could be efficiently killed (70-72). Also, it seems
likely that the antibiotics used for biofilm infection could be significantly reduced if biofilms have already
partially been disintegrated (73). Thus, detachment of biofilms will ultimately improve the therapeutic
effect of biofilm control.
As discussed above, comparisons of differences among V. cholerae, P. aeruginosa and E. coli strains reveal a
wide range of biofilm formation patterns, all linked to QS signaling molecules (AI-1 or AI-2) (47, 64, 74).
These interspecific differences are likely dependent on the internal environment and related to the amount
of competition and the necessity for dispersal.
Intraspecific variations in biofilm formation have also been observed. The biofilm formation of V.
anguillarum strains PF430-3 and BA35 in a flow cell device (75) were highly different with respect to
structure and stability (Figure 7). For example, an increase in biofilm formation associated with the pellicle
on the top of tubes were observed in strain PF430-3; On the contrary, the biofilm stability of V. anguillarum

20
strain BA35 is relatively poor, as no surface pellicle in the liquid cultures could be found. According to the in
vitro study of strain BA35, biofilm formation decreased after 2 days. The decrease in the level of biofilm
could also be partially attributed to an earlier onset of the dispersal phase because of a rapid increase in
biofilm formation, which has been observed in Hosseinidoust’s study (76). In general, biofilm stability is
closely correlated with the EPS production (77), as previously visualized by use of fluorescently labeled
lectins carbohydrate-containing EPS in P. aeruginosa biofilms (78). Perhaps, EPS production in V.
anguillarum strain PF430-3 contributed to the formation of the characteristic colonies, which prevented
early dispersing. A similar phenomenon was also mentioned in Fong’s V. cholerae study (79), by deleting
vps genes which is required for biofilm formation, many of the mutants exhibited reduced capacity to
produce VPS and biofilm formation.
Figure 7. Bacterial biofilm formation of V. anguillarum strain PF430-3 and BA35 after 24 h, samples were stained with LIVE/DEAD
BacLight viability stain (Invitrogen) and visualized by confocal scanning light microscope.
3.3. Heterogeneity in biofilms
Bacterial biofilms are physiologically and spatially heterogeneous; even within a single species, differential
spatial and temporal gene expression occurs due to chemical gradients and adaptation to the local
environmental conditions (80-82). The expression of different extracellular molecules could change the
surface antigen properties of the cells and thereby promote the formation and maintenance of different
types of multicellular communities during the biofilm infection, including different stages between “on”
and “off” status (83). These strategies generally were recognized by the function of enabling the individual
cells to evade immune system or predator (84). For example, a subset of the population in the planktonic
phase may accumulate mutations that enable them to produce more EPS, grow slower, become non-motile
and form aggregates (80). These aggregates from the mutants may increase the adhesion capacity to the
substratum and function as a physical barrier that protects against attacks and enhances the survival of the
entire community. The remaining planktonic members of the community could contribute to different

21
benefits to the community, such as protozoa grazing, phage attack or others may be involved in cellular
communication (47, 85, 86). Thus, functional specialization may be promoted through mutations or gene
regulations and could also stimulate multicellular development (86).
Despite an increasing knowledge of biofilm structure and properties, many key questions still remain to be
answered about essential factors regulating biofilm formations. Several environmental conditions have
been proposed to influence biofilm structure. For instance, Kreft’s study (87) has shown that the stability of
a biofilm is closely correlated with the EPS production as well as pilus and flagella, i.e. high EPS production
resulted in a solid structure of biofilm. Similarly, Nagaoka reported that a high level of EPS production
increased viscosity of the medium (88). Moreover, in Reisner’s E. coli K-12 biofilm models, the presence of
transfer constitutive IncF plasmids were essential in inducing biofilm forming structures (89). It should be
noted that the discussion here is limited to the single-species biofilm formation; additionally, solid-air
biofilms and liquid-gas biofilms would probably have their distinct characteristics, which would influence
cellular communications (90).
3.4. Diagnosis of biofilms
Detection of bacterial biofilms of V. anguillarum, V. cholerae, S. aureus and P. aeruginosa are pivotal in
many diagnostic decisions. In addition, the presence of multi-drug resistance V. anguillarum, V. cholerae
and P. aeruginosa, and methicillin-resistant S. aureus (MRSA) can pose intractable problems. These
pathogens all form culturable biofilms, which can be studied in the laboratory, and much of the progress in
our understanding of biofilm structure, regulation and treatment relates to studies of such pathogens (91,
92). Additionally, the problems of biofilms cannot be neglected, since most biofilm samples are either
conditionally viable environmental cells (CVEC) or viable but non-culturable (VBNC). Due to these CVECs
and VBNCs, traditional methods are inefficient in detecting and identifying certain biofilms. However,
molecular methods offer a way to overcome these shortages in traditional methods. Most nucleic acid-
based molecular methods for the detection and identification of bacteria begin with the extraction of DNA
and/or RNA from the sample to be analyzed or directly identified by fluorescence in situ hybridization (FISH)
(93). This extraction and thus diagnosis will be more efficient, and will yield more precise quantification, if
the nucleic acids have not been degraded by chemical preservatives or by endonuclease enzymes (91). Also,
recent studies have shown that using enrichment media containing autoinducers (AIs) improves
resuscitation of dormant V. cholerae in environmental water samples and S. enterica serovar typhimurium
and enterohemorrhagic E. coli (92, 94-96).

22
3.5. Treatment of vibriosis in aquaculture
Traditionally, pathogens in aquaculture have been treated with antibiotics. For instance, quinolones is still
the first drug of choice for vibriosis in aquaculture environments. This may explain the oxolinic acid (OA), a
4-quinolone resistance found among Vibrio spp. strains isolated from diseased Atlantic cod (97). Moreover,
bacterial infections mediated via biofilm formation are persistent and difficult to treat with traditional
methods, due to low antibiotic penetration and non- or extremely slow-growing physiological state (98).
Besides, a severe problem in the aquaculture industry these days is that even without any apparent disease
symptoms, antibiotics are still used routinely and prophylactically, due to lack of sanitary barriers (99). The
unconsumed fish pellets and fish feces, may add to excess antibiotic residues in the sediments at the
bottom of raising pen (99). For instance, in Thailand, it has been reported by Holmström and colleagues
that of 76 shrimp farmers, 56 used antibiotics (100), with more than 10 different antibiotics used
prophylactically. Table 1 is an overview of the major classes of antibiotics used in the aquaculture industry
adapted from Defoirdt’s study (101). Serious concerns have been raised with respect to the development of
bacterial antibiotic resistances and horizontal gene transfer to human pathogenic bacteria, via antibiotic
residues. As discussed above, antibiotic use has to be restricted and alternatives for treating bacterial
infection must be implemented.

23
Table 1 (101)
The different classes of antibiotics used in aquaculture, their importance for human medicine and examples of (multi)resistant
pathogenic bacteria isolated from aquaculture settings.
Drug class
Importance for
human medicine Example Resistant bacteria
Multiple
resistance? Isolated from
Aminoglycosides Critically important Streptomycin Edwardsiella ictulari Yes Diseased striped catfish
(Pangasianodon
hypophthalmus), Vietnam
Amphenicols Important Florfenicol Enterobacter spp. and
Pseudomonas spp.
Yes Freshwater salmon farms,
Chile
Beta-lactams Critically important Amoxicillin Vibrio spp., Aeromonas
spp. and Edwardsiella
tarda
Yes
Different aquaculture
settings, Australia
Beta-lactams Critically important Ampicillin Vibrio harveyi Yes Shrimp farms and coastal
waters, Indonesia
Fluoroquinolones Critically important Enrofloxacin Tenacibaculum
maritimum
Yes Diseased turbot
(Scophthalmus maximus)
and sole (Solea
senegalensis), Spain and
Portugal
Macrolides Critically important Erythromycin Salmonella spp. Yes Marketed fish, China
Nitrofurans Critically important Furazolidone Vibrio anguillarum Yes Diseased sea bass and sea
bream, Greece
Nitrofurans Important Nitrofurantoin Vibrio harveyi Yes Diseased penaeid shrimp,
Taiwan
Quinolones Critically important Oxolinic acid Aeromonas spp.,
Pseudomonas spp. and
Vibrio spp.
Yes Pond water, pond sediment
and tiger shrimp (Penaeus
monodon), Philippines
Sulphonamides Important Sulphadiazine Aeromonas spp. Yes Diseased katla (Catla catla),
mrigel (Cirrhinus mrigala)
and punti (Puntius spp.),
India
Tetracyclines Highly important Tetracycline Aeromonas hydrophila Yes Water from mullet and
tilapia farms, Egypt
Tetracyclines Highly important Oxytetracycline Aeromonas salmonicida Yes Atlantic salmon (Salmo
salar) culture facilities,
Canada

24
4. Alternative disease control in aquaculture
Multiple commercial vaccines are currently used to protect fish against outbreaks of vibriosis; all of these
vaccines consist of inactivated strains of both V. anguillarum serotypes O1 and O2 (102). However,
outbreaks of vibriosis caused by serotype O2 have not been completely prevented (22). Moreover, at the
larval stage, fish are more vulnerable to V. anguillarum infections, and the vaccine is largely ineffective as
their immune system is not fully developed. Therefore, alternative antibacterial strategies are required to
be identified and developed.
Global regulators such as autoinducer-1 (AI-1) and autoinducer-2 (AI-2) inhibitors of bacterial physiology
have emerged as a potential approach to manipulate bacteria as discussed above (103, 104). These AIs not
only have important roles in biofilm formation, virulence and antibiotic resistance, but also have secondary
functions, which can be used as pro- or anti-QS/antibiotic/phage therapy synergy for both acute and
persistent infection treatment in the future (103, 105-108). All these characteristics can be taken into
consideration for the development of future treatment strategies for bacterial infections in aquaculture.
4.1. Bacteriophages
Bacteriophages are viruses that parasitize bacteria. Phages are the most abundant microorganisms in the
ecosystem, with total numbers estimated to be more than 1030 (109). Bacteriophages are ubiquitous, and
are found in marine and freshwater, soils, as well as the intestinal tracts of animals, and are estimated to
be on the order of 107–109 per gram dry weight of soils and feces or per milliliter of seawater (109). The
ecological functions of bacteriophages also involves playing important roles in e.g. structuring bacterial
diversity and succession in the ocean, promoting biogeochemical element cycling and as key drivers of
horizontal gene transfer (110).
4.2. Phage classification
Bacteriophages are classified by the International Committee on Taxonomy of Viruses (ICTV) based on their
morphology and types of nucleic acid. More than 5500 phages have been examined by electronic
microscopy, ~96% are tailed (111). These tailed phages, which belong to the order of Caudovirales, can be
divided into 3 families (Myoviridae, Siphoviridae, and Podoviridae) (Figure 8). The differences among these
three families are: a long or short contractile tail (Myoviridae), a long non-contractile tail (Siphoviridae), and
a short, non-contractile tail (Podoviridae).

25
Figure 8. Transmission electron microscopy (TEM) images of selected vibriophages from three different families, scale bar indicates
200 nm. Phage KVP40, ΦH20, and Φ4-7 belong to Myoviridae, Siphoviridae, and Podoviridae, respectively (18).
4.3. Bacteriophage life cycle
Since phages need bacterial hosts to replicate, phage infection may lead to bacterial cell death (lytic life
cycle) or to a lysogenic relationship, where both genomes of phage and host replicate together. Specifically,
in the lytic life cycle, phage progenies are released from lysis of the bacterial host. New progenies then
continue to infect more hosts. In the lysogenic life cycle, temperate phage DNA is integrated into host
chromosomes and replicates along with cell division (Figure 9). However, this is not always permanent and
phages can be induced from the lysogenic to the lytic life cycle, which is dependent upon environmental
signals and the number of infected phages per cell (112), such as DNA damage, UV exposure, mitomycin C,
hydrogen peroxide and high MOI, by causing an SOS response (113). Additionally, phages are known to play
a critical role in the evolution of pathogenic bacterial species, and as it is particularly true for V. cholerae
(114). For example, a major virulence factor of V. cholerae, the cholera toxin (CT), is encoded by ctxAB in
the lysogenic phage CTXΦ (115). Likewise, cryptic prophages also were shown to help bacteria cope with
adverse environments, such as cell growth, antibiotic resistance, early biofilm formation, as well as
environmental stresses (116, 117).

26
Figure 9. Schematic picture of phage lytic and lysogenic life cycles. Phage adsorption is the first key step in phage proliferation; in
order to efficiently bind to the bacterial receptor, phage receptor-binding protein (RBP) is required to specifically interact with
bacterial cell surface receptor to enable intracellular DNA injection. In the lytic life cycle, phage DNA replicated separately from
host genome, resulting in the destruction of the infected cell and its membrane. In the lysogenic life cycle, phage DNA is integrated
into the host genome and can be transferred to daughter cells, until the lytic cycle is induced. However, a portion of these induced
cells could enter an abortive lytic cycle by losing prophage and becoming non-lysogens (curing) (112).
4.4. Vibriophages
Although viruses have been enumerated, isolated and characterized from the marine environment, there is
little information about the specific phage types. Previous seasonal and spatial studies about vibriophages
were carried out mainly based on the V. parahaemolyticus hosts, such as vibriophages isolated from the
Strait of Georgia (Vancouver, Canada) (118) and Tampa Bay (Florida, USA) (119), as well as V. cholerae
prophages such as K139 (120), and filamentous phage CTXΦ, which has been shown to be linked to the
bacterial pathogenicity, CT production (121).
Among the best characterized vibriophages is bacteriophage KVP40. KVP40 was originally isolated from
polluted seawater off the coast of Japan using V. parahaemolytcius as the host (122). Phage KVP40 is a
novel T4-like virulent vibriophage, with a broad-host-range, belonging to the Myoviridae family, and the
genome size is around 244 kb, with a G+C content of 42.6%, and a National Center for Biotechnology
Information (NCBI) accession number of B_KVP40 AY283928 (123). Vibriophage KVP40 is known to cause
infection through the universal outer membrane protein K (OmpK) and has previously been shown to infect

27
more than 8 Vibrio species, including V. anguillarum, V. parahaemolyticus, V. harveyi, V. natriegens, V.
cholerae, as well as Photobacterium leiognathi (Table 2) (122, 124).
Table 2. Adapted from Inoue (124)
Distribution of OmpK or its homologs among Vibrio and Photobacterium strains
Species Strain Phage sensitivity a Phage adsoprtion
b OmpK-like protein
c
Phage-resistant mutants
d
V. parahaemolyticus 1010 + + + OmpK-
VIB4 - + +
VIB8 + + +
VIB9 + + + OmpK
d
V. alginolyticus VIB20 + + +
VIB31 + + + OmpK
-
VIB33 - + +
V. natriegens VIB30 + + - OmpK
d, OmpK
-
V. cholerae non-O:1 VIB5 + + +
V. mimicus VIB13 + + + OmpK
d, OmpK
-,
OmpK+
V. anguillarum I VIB35 + + + OmpKd
VIB36 + + + OmpK+
V. splendidus II VIB37 + + + OmpK-
V. fluvialis I VIB7 + + +
VIB18 + + + OmpK
-
V. vulnifics VIB6 - - + V. harveyi VIB26 - + + V. campbellii VIB27 - - + V. nereis VIB29 - - + V. pelagius I VIB38 - - + V. costicola VIB39 - + + P. leiognathi PHO1 + + + P. angusturn PHO2 - + -
a Sensitivity to KVP40 tested by plaque formation.
b More than 10% adsorption of KVP40 as compared with 1010.
c OmpK or its
homologs detected by immunoblotting. d Phenotypes of KVP40-resistant mutants isolated. The OmpK
-like protein was defective
(OmpK-), partially defective (OmpK
d), or produced normally (OmpK
+).

28
4.5. Application of phages to control fish pathogens: Phage therapy
Due to their efficient lysis, lytic phages can potentially be used against bacterial infection, and are much
more specific than commonly used antibiotics. Therefore, by using phage therapy, a specific bacteriophage
could theoretically be chosen to target a specific pathogen. Because of their host specificity, they would not
affect beneficial bacteria (e.g. gut flora), thus reducing the chances of opportunistic infections (125). In
2006, the United States Food and Drug Administration (FDA) approved using bacteriophages on ready-to-
eat meat and poultry products in order to kill the Listeria monocytogenes bacteria, giving these products
GRAS status (Generally Recognized as Safe) (126). Therefore, properly developed phage products can be
invaluable in controlling bacterial infections in various settings.
Recently, there has been a growing number of studies on phage therapy, focusing on a variety of fish
pathogens including A. salmonicida in brook trout, Yersinia ruckeri in salmon, Lactococcus garvieae in
yellowtail, P. plecoglossicida in ayu, F. psychrophilum in rainbow trout, V. anguillarum in Atlantic salmon
and V. harveyi in shrimp (127-134). These studies have demonstrated the potential of specific phages to
significantly control pathogen density and, in some cases, reduce fish mortality. For instance, Silva’s study
of V. anguillarum and vibriophage showed that the larvae mortality in the infected and treated group was
similar to normal levels and significantly lower than the infected but not treated group (135). Moreover,
according to Lomelí-Ortega’s study, lytic phage A3S and Vpms1 were also effective to reduce larvae
mortality caused by V. parahaemolyticus (136). Similarly, in Vinod’s field trail experiments, treatment with
bacteriophage improved larval survival and brought about decline in luminescent V. harveyi counts in
hatchery tanks (133).
In addition, studies from V. cholerae (114) have also shown that lytic vibriophages may play an essential
role in modulating the seasonal dynamics of cholera in endemic areas, such as the Ganges Delta region of
Bangladesh and India. Specifically, the number of cholera patients increased whenever the number of lytic
vibriophages in the water decreased, and cholera epidemics tended to end concurrent with large increases
in the concentration of vibriophages in the water (114, 137), indicating that lytic vibriophages specific for V.
cholerae may limit the severity of cholera outbreaks by killing susceptible bacteria present in the reservoir.
Due to their selective killing of susceptible strains, vibriophages are also believed to play a key role in the
emergence of new V. cholerae pandemic serogroups or clones (114, 138, 139).
Therefore, it seems a promising strategy to apply vibriophages to gain control of vibriosis infections in fish
used for aquaculture. However, successful application of phage therapy in the treatment of vibriosis
requires a detailed understanding of phage-host interactions in both planktonic and biofilm forms,

29
especially with regards to anti-phage defense mechanisms in the bacterial hosts, specifically, bacterium V.
anguillarum in this Ph.D. study.

30
5. Bacteriophage resistance mechanisms
Predation pressure from bacteriophages is substantial, mainly because their abundance outnumbers
microbial cells by an estimated 10-fold in most natural environments (140). Accordingly, bacteriophage lytic
infection imposes a strong selection for bacterial mutations/tolerance providing reduced phage
susceptibilities or resistance against phage infection. Bacteria have evolved a wide range of different
resistance/tolerance mechanisms including 1), preventing phage adsorption; 2), cutting phage nucleic acid;
3), abortive infection through altruistic suicide; and 4), QS-mediated receptor down-regulation, which can
make the host immune to the viral infection (141). Similarly, phages can also overcome bacterial resistance
by adapting to new receptors, battling restriction-modification systems, evading CRISPR-Cas systems, and
escaping abortive-infection mechanisms (142).
5.1. Phage receptor modification
Adsorption is a key step recognition between phage receptor-binding protein and phage receptors on the
sensitive host cells (143). Most of phage receptors are presented on the bacterial cell walls, such as LamB
for phage lambda, the prion outer membrane protein F and C (OmpF and OmpC) for phage T2 and T4, and
moreover, the flagella protein for phage phi (144-147). A recent study showed that ϕCb13 and ϕCbK
actively interact with the flagellum of the bacterium Caulobacter crescentus and subsequently attach to
receptors on the cell pole. Using the flagella or pili to initiate contact with the host cell, increases the
likelihood of attachment and successful infection (148). Similarly, we noticed that a large fraction of cells in
the BA35+ΦH20 cultures were found to be either non-motile or with impaired mobility, which may imply
that flagella play an important role in phage-host interactions. Furthermore, a specific polysaccharide was
also implicated as an essential component in adsorption, for instance, phage kh and host L. lactis subsp.
cremoris KH, as well as phage 1358 and L. lactis (149, 150). Additionally, according to Seed’s study, V.
cholerae lipopolysaccharide O1 antigen functions as a major target of phage ICP1 (151). By using phase
variation of O antigen biosynthesis, V. cholerae cells can easily generate variable expression of surface
components, which is generally thought to help these organisms evade the immune system and phage
predation (151). However, little is known about the requirements of these phage receptors and it is unclear
if the polysaccharide was acting as a receptor or if it was facilitating reversible phage binding to a secondary
receptor (152, 153).
Mutating phage receptors or producing EPS to block the interaction between phage receptor and phage
receptor-binding protein to prevent phage adsorption can be the first step that bacterial cells exhibit in
developing resistance/tolerance to avoiding infection (154). Previous studies in Vibrio hosts, showed that
mutations or modifications of the outer membrane protein K (OmpK), led to resistance to vibriophage

31
KVP40 (124). Similarly, in Paper III, ompK in-frame deletion also proved that OmpK acts as a phage binding
receptor in V. anguillarum strain PF430-3 (Figure 10). Furthermore, mutations in outer membrane protein A
(OmpA) of E. coli K-12 appear to play a role in inhibiting phage infection (155). Phage receptors, in addition
to the phage attachment, were involved in the bacterial nutrient intakes, which may be responsible for the
morphological changes of the colonies with small size, known as small-colony variants (SCVs) and fitness
costs, as have been previously demonstrated experimentally, such as reduced abilities to take up specific
nutrients (156) or reduced competitive abilities in general (157, 158). For example, mutation in the LamB
phage receptor caused the inability to transport long chain maltodextrin across the outer membrane (159).
In Paper II, phage ΦH20-resistant mutant of strains BA35 was found to have reduced ability to utilize a
number of carbon resources in the BIOLOG GN2 test.
Additionally, Park (160) found that bacteriophage-resistant mutants of P. plecoglossicida lacked virulence
for ayu. In addition, in a successful phage therapy experiment of E. coli infection in mice and calves, the
resistant mutants isolated from the treatment were the less virulent k-1 type mutants (161). In Paper III, the
function of phage KVP40 binding receptor OmpK still remained unknown, but, it has been suggested to be
involved in bile salt resistance and iron acquisition (162). Studies on vaccines have shown that OmpK could
also function as a subunit vaccine against V. anguillarum infection (162), because the outer membrane has
been reported as selectively permeable or serves as a receptor as well as adhesions in most Gram-negative
bacteria for colonization (163, 164).
Figure 10. Spot assay for testing host specificity of phage KVP40 against wild-type, QS mutants (ΔvanT and ΔvanO) and ompK
mutants (ΔompK, ΔvanT ΔompK, and ΔvanO ΔompK) as well. Five μl serial 100-fold dilutions of phage KVP40 lysate (1, 109; 2, 10
7; 3,
105; 4; 2, 10
3 PFU ml
-1) were shown by spot titration onto top agar lawns of the indicated strains (Paper III).

32
The role of spatial refuge in stabilizing bacteria-phage interactions has been observed in many ecosystems,
especially, micro-colonies and biofilms, as discussed in previous studies (165-167), and even in the marine
environments, such as marine snow and sediments. Because of the low dispersal rates in the
heterogeneous environments, by creating ephemeral refuge may directly/indirectly block phage receptor
from phage attack by non-mutation-based mechanisms (165-168); as also shown in V. anguillarum strain
PF430-3 protected against phage KVP40 infection by increasing cell aggregation and biofilm formation,
allowing coexistence rather than coevolution, finally promoting the stability of phage-host systems by
reducing the risk of lytic phage attacks.
5.2. Cutting phage nucleic acids
Once the nucleic acid has been injected into the bacterial cell, the restriction-modification (R-M) systems
protect the bacterium by cutting invading DNA into pieces (169). When unmethylated phage DNA enters a
cell harboring R-M systems, restriction enzymes, thereby, rapidly degrade the foreign genetic material
functioning as a prokaryotic immune system (170). For instance, in Bacillus subtilis Marburg nonB mutated
strain, nonsense mutation on ydiB was found to be related to the restriction system targeting sequence of
BsuMR, which was identical to XhoI (CTCGAG) (171, 172). However, according to Krüger’s study (173), R-M
systems are not always perfect, for instance, phages and plasmids can acquire host modifications to avoid
restriction endonuclease, which highlights an evolutionary arms race between bacterial host and phages
(142).
5.3. CRISPR/Cas bacterial immune system cleaves bacteriophage DNA
Another mechanism recently described is CRISPR (Clustered Interspaced Short Palindromic Repeats) and
the CRISPR-associated (cas) genes system, in which a CRISP-cas loci was identified composing of 21-48 bp
direct repeats interspaced by non-repetitive spacers (26-72 bp) of similar length (174, 175). By using this
immunity system to acquire at least one new repeat-spacer unit at the 5’ end of the repeat-spacer region of
a CRISPR locus that targets foreign nucleic acids, bacteria can efficiently protect themselves from including
phage DNA and plasmids (175, 176).
5.4. Abortive infection (ABi) system
The study of the ABi system began 50 years ago, and even now, the mechanisms underlying the infection
are still not completely understood (141). The bacterial cell can increase the chances of its own population
survival by using the abortive infection system, where phage infection leads to the death of infected
bacterial cells. The system is characterized by a normal infection start (i.e., the phage adsorbs and injects its
DNA into the host cell), followed by an interruption of the replication, transcription or translation, leading
to the release of little or no new phage progenies (141, 177). Recent studies in B. subtilis showed that the

33
Marburg strain remained resistant to phage SP10 due to a NonA-mediated aborted infection system, acted
as a second layer of protection against phage SP10 infection, specifically, the overexpression of nonA gene
terminated cell growth with reduced efficiency of colony formation and respiration activity (172).
5.5. Regulation of phage-host interactions by extracellular signaling molecules
A recent study has demonstrated a new mechanism of phage resistance in which, bacteria can coordinate
their receptor gene expression upon the environmental QS signal, to avoid the risk of infection at high cell
densities (45). Since phages require a host to replicate, it follows that the predation pressure is relatively
higher at a high cell density status compared to sparsely populated environments. Hence, if bacterial hosts
could regulate their anti-phage mechanisms based on cell densities, they could easily reduce their
susceptibilities to infection during high cell densities, while avoiding the metabolic burden of maintaining
elevated anti-phage defenses during growth at low cell densities. For instance, E. coli possesses the ability
to use AHLs to reduce its susceptibility to at least two phages, phage λ and phage χ (45). In the phage λ case,
phage receptor LamB was shown 40% down-regulation compared to the untreated controls without AHLs
(45). In addition, high concentration of universal communication molecule AI-2 (100 μM) was demonstrated
to induce virulence genes and transfer them via phage release in E. faecalis V583ΔABC (178). Intriguingly, a
recent study from Hargreaves (179) identified Clostridium difficile phage θCDHM1 encoded an intact QS
operon. Similarly, QS-associated genes have also been identified in several phages of Psedomonas spp (180,
181), and function as acylhydrolase to break down AHLs, was identified in the Iodobacter-phage ϕPLPE
(182). Although it remains unknown whether any of these genes are of functional importance, it certainly
opens the exciting possibility that some phages may actively interfere with the QS in the bacterial
communities.
5.6. Implications of phage protection mechanisms: Phage-host coexistence and co-evolution
As complexity arises within populations of bacterial resistant mutants, it may help mutants survive better
or have more offspring. If so, this complexity will be favored by natural selection and spread through the
bacterial population. However, most mutations with phenotypic effects are harmful, such as reducing the
competitiveness of the mutant strains, as most phage receptors involve nutrient intake or pathogenicity, a
receptor-deficient mutant will have a slower growth rate or reduced virulence (160, 161, 183-185). That is,
phage resistant mutants with those traits will tend to be wiped out before reproducing, taking the
deleterious traits out of bacterial communities. Therefore, the non-mutation defense mechanisms among
these phage hosts may suggest phage-host co-existence interactions, rather than the classical phage-host
co-evolutionary arms race, known as Red-Queen theory.

34
The Red-Queen hypothesis was first formed by Van Valen in order to explain the “law of extinction” (186).
According to the previous phage-host interaction studies, virulent phages managing to coexist with their
bacterial host leads to continuous variations and selections towards the adaptation of bacterial hosts by
evolving resistance to current phages and phage evolve to counter resistance. It has been reported that the
arms race has a huge impact on global nutrient cycling, on climate, on the evolution of the biosphere, and
on the evolution of virulence pathogens (187). However, this theory was also criticized because the
evolution rates between phage and host are not symmetrical. Recent studies showed that, in soil, phages
seem ahead of the bacterial hosts in the evolutionary arms race (188). In the natural environment, at each
stage of the arms race, one could become extinct, and without one, phage and bacteria coexistence,
therefore, does not even exist. For instance, one of the typical examples of “Cheshire Cat” ecological
dynamics is the haploid phase of Emiliania huxleyi provided an escape mechanism that involved separation
of meiosis from sexual fusion in time, to ensure that genes of dominant diploid clones were passed on to
the next generation in a virus-free environment (189). Thus, it is important to understand both evolutionary
and non-evolutionary mechanisms (such as gene regulation) that can regulate phage-host interactions in
the future.
Uncovering anti-phage defense mechanisms is essential for understanding phage-host dynamics and for
application of phages in disease control, as they reflect the remarkable diverse interactions between
bacterial hosts and viruses and play a key role as agents shaping microbial community structure. As the
predation pressure from phages is a key determinant of the size and composition of bacterial populations,
understanding of the potential factors that govern phage-bacterial interactions will be important in any
context where the goal is to control the growth of a microbial population including, for example, the
treatment of bacterial infections, development of effective probiotics, production of cultured dairy
products, or manipulation of the human microbiome to prevent or treat life-style diseases.

35
Aims of this Ph.D. project
The aim of this Ph.D. project was to investigate the interactions between vibriophages and their hosts V.
anguillarum, and to ultimately improve the efficiency of phage therapy.
The objectives of this project were:
To isolate and characterize vibriophages and V. anguillarum and investigate the occurrence of phage
susceptibility patterns. (Paper I)
To explore phage-host interactions in two V. anguillarum strains, and the resistance/tolerance mechanisms
responsible for the different outcomes of these two different systems. (Paper II)
To determine whether AHLs-mediated Quorum Sensing regulates anti-phage defenses in V. anguillarum
strain PF430-3. (Paper III)

36
A summary of this Ph.D. project
Paper I
Vibriophages and their interactions with the fish pathogen Vibrio anguillarum
The use of phage therapy to control bacterial pathogens may offer an effective method to reduce vibriosis
infection in fish or shellfish maintained in aquaculture. Phage therapy is a promising alternative to the use
of antibiotics and thus avoids the risks associated with antibiotics abuse and overuse. However, a detailed
understanding of phage-host interactions is needed to evaluate the potential of this technique.
As the first approach, we collected and isolated 24 V. anguillarum strains and 13 Vibrio sp. strains, which
represented considerable temporal (20 years) and geographic (9 countries) variations. Irrespective of their
origins of isolation, further characterizations based on phylogenetic (16S rRNA) and physiological (BIOLOG
GN2 and API 20NE) fingerprints, revealed that they were quite similar.
Ten phages were isolated from Danish water samples with different Vibrio hosts. In addition to the Danish
phage isolates, a broad-host-range vibriophage KVP40 was also included in this study. Phages were further
characterized by genome sizes, morphologies, and host-range analysis. Together, 11 phages represented
three different families (Myoviridae, Siphoviridae, and Podoviridae), which covered all the host ranges of 37
Vibrio strains.
Despite the occurrence of unique susceptibility patterns of the individual host isolates, key phenotypic
properties related to phage susceptibility are distributed worldwide and have been maintained in the
global Vibrio community for decades. Unfortunately, the phage susceptibility pattern of the isolates did not
show any relation to the physiological relationships, demonstrating that similar phage susceptibility
patterns may occur across a broad phylogenetic spectrum and with large physiological differences among
Vibrio communities.
Subsequently, culture experiments were conducted with two V. anguillarum strains (BA35 and PF430-3)
and their corresponding phages (ΦH20 and KVP40), representing strong lytic potential initially followed by
rapid regrowth of phage-resistant and phage-tolerant subpopulations, which is an obstacle to successful
phage therapy. Different bacteriophage resistance/tolerance mechanisms underlined by these two specific
phage-host interactions were not revealed in this study.

37
Paper II
Vibriophages differentially influence biofilm formation by Vibrio anguillarum strains
In this study, we explored in vitro phage-host interactions in two V. anguillarum strains (BA35 and PF430-3)
and their corresponding phages (ΦH20, Siphoviridae and KVP40, Myoviridae), respectively, during growth in
the settings of micro-colonies, biofilms, as well as the free-living phase.
We sought to figure out the resistance/tolerance mechanisms responsible for the different phenomena
between these two phage-host systems.
For strain BA35 and phage ΦH20, a strong phage control of the phage-sensitive population and subsequent
selection for the phage-resistant mutants with fitness cost, implicates a co-evolutionary arms race may
exist between bacterial hosts and their predators. This then serves as a potential mechanism to promote
diversity of resistant subpopulations and phage ΦH20 through stable co-evolution and mutations.
For strain PF430-3 and phage KVP40, addition of phage KVP40 to strain PF430-3 cultures resulted in an
increase in the levels of biofilm/micro-colony/aggregate formation, especially, during the initial stage. By
creating physical spatial refuge between host and parasite, the phage can potentially allow long-term co-
existence between strain PF430-3 and phage KVP40. In addition, we were unable to isolate any single
colonies with full and permanent resistance to phage KVP40. Hence, it may be speculated that the
development of this transient protection is due to the fact that phage KVP40 infects more than 8 Vibrio
species using phage receptor OmpK, which is widely distributed in the Vibrio community. Thus, resistance
mutations are either very rare or have significantly negative or even lethal effects on host fitness. In that
case, development of alternative strategies to genetic mutations in the OmpK receptor would be of strong
selective advantage, perhaps contributing to the evolution of the protection mechanism.
Overall, the specific mechanisms of phage resistance/tolerance were not identified in this study between
these two phage-host systems, and further studies are needed to continue to investigate the mechanism of
resistance/tolerance towards these two phages. Taken together, these data demonstrated highly variable
phage protection mechanisms among Vibrio communities, emphasizing the challenges of using phages to
control vibriosis in aquaculture, and adding to the complex roles of phages as drivers of prokaryotic
diversity and population dynamics.

38
Paper III
Quorum Sensing determines the choice of anti-phage defense strategy in Vibrio anguillarum
Since phage and host have a long co-existence and co-evolutionary development, it is generally believed
that bacteria can effectively defend against phage predation by using various means and with limited
fitness cost to cope with further selection pressure. Alternating different protection strategies at different
stages perhaps lessens the metabolic burden, facilitating specific subpopulations to prevail in phage-host
systems. As bacterial densities vary overtime from sparsely populated environments to highly dense
bacterial populations, phage predation pressure is not constant. By altering their anti-phage activities upon
population density (phase-dependent defense), bacteria could actively reduce their susceptibility to
infection during growth at high cell densities. Vibriophage KVP40 is known to cause infection through the
universal outer membrane protein K (OmpK) and has previously been shown to infect more than 8 Vibrio
species; by using spatial refuge and a quorum sensing-mediated receptor down-regulation would decrease
the metabolic burden in these Vibrio communities.
In this study, we revealed the roles of spatial refuge and cell-cell signaling in shaping phage-host
interactions. Two V. anguillarum PF430-3 QS mutants were constructed, ΔvanT-locked in low cell density
and ΔvanO-locked in high cell density, allowing us to explore the role of QS for protection against phage
infection. Specifically, ΔvanT became more susceptible to phage KVP40 by enhancing biofilm formation,
and thus creating spatial refuge, providing protection to the host against phage infection. In contrast,
ΔvanO became more resistant by down-regulating the phage receptor OmpK to reduce its susceptibility to
phage infection with no bacterial aggregates. Further, wild-type strain culture experiments showed a strong
negative correlation between AHLs production in the supernatant and phage receptor ompK gene
expression, which was consistent with previous results. The data presented here, therefore, suggest that
VanO negatively regulates the expression of VanT, which in turn directly or indirectly represses the
expression of essential phage receptor OmpK (Figure 11).
Figure 11. Models for the mechanism and function of QS regulation and biofilm formation and phage receptor ompK expression in
V. anguillarum. Solid arrows represent positive effects, while solid T bars represent negative effects.

39
Future perspectives
The studies presented in this Ph.D. dissertation provide new insights into phage-host interactions of the
important fish pathogen V. anguillarum and their vibriophages, shedding light on the relationships between
phage infection pressure, biofilm development, quorum sensing and bacteriophage resistance/tolerance
mechanisms in V. anguillarum. However, there are lots of unsolved issues in this study, which raise
questions regarding future experiments.
Paper I
Thirty seven Vibrio strains were collected with considerable temporal (20 years) and geographical (9
countries) differences in their origins of isolation, which provides us with knowledge on spatial and
temporal distributions of the pathogen which may prove useful when assessing global outbreaks of
vibriosis. However, these strains are still insufficiently characterized. For instance, phage susceptibility
patterns of the isolates did not show any similarities to the physiological relationships obtained from
BIOLOG GN2 and 16S rRNA profiles, and thus further studies linking phage susceptibility to phenotypic and
genotypic characteristics would be relevant.
First, by selecting vibriophage resistant mutants among the potential hosts, we could identify cellular
components that are involved in the bacterial receptor for the specific phages, as previously shown in
Davidson’s study in B. anthracis (gamma) phage γ receptor GamR (190). Further studies will be focused on
potential phage receptor identification and will determine whether there is any correlation between phage
susceptibilities and phage receptor types.
Secondly, one of the most important issues in phage therapy is the ability to obtain phages capable of
infecting the causal pathogen; hence, the isolation step is a critical element. The isolation of vibriophages is
considered relatively difficult, and there is a need for new and more efficient isolation methods.
Thirdly, physical and chemical factors, such as temperature, acidity, and ions have shown to affect phage
persistence (191). A better understanding of storage and stability of phage stock, i.e. mechanisms and rates
of phage decay, are also essential for a successful future phage therapy, not only for those interested in
pharmaceutical and agricultural applications, but also for laboratory phage studies.
Lastly, most bacterial pathogens contain prophages or phage remnants integrated into bacterial
chromosomes (192). As these prophages may carry virulence genes, it is essential to obtain a better
understanding of the content and genetic properties of prophages integrated in V. anguillarum strains.
During the lytic phage cycle, prophage induction can be triggered, which makes lytic phage stock

40
contaminated with the induced prophages that were previously resident in the host strains. Thereby, the
data we obtained from the host range assay could be related to the prophage from the host of the lytic
phage. The argument of prophage here suggests that phage host range can be optimized by using prophage
free hosts for lytic phage proliferation in the future. We are currently whole genome sequencing our full
collection of V. anguillarum and vibriophages, which will allow such a detailed analysis of prophage
contents.

41
Paper II
The studies from Paper II demonstrated that two closely related V. anguillarum strains BA35 and PF430-3
displayed highly variable phage protection mechanisms, during growth in the settings of micro-colonies,
biofilms, as well as the free-living phase.
In general, biofilm stability is closely correlated with EPS production (87), and we hypothesized that EPS
production in strain PF430-3 contributed to the formation of the characteristic micro-colonies, which
protected the strain PF430-3 from phage infection by creating spatial refuge. Therefore, further
experiments on EPS quantification and identification remain to be carried out. Previous studies on P.
aeruginosa have shown fluorescently labelled lectins could be used in combination with epifluorescence
microscopy, allowing the visualization and characterization of carbohydrate-containing EPS in biofilm
matrices (78). This could lead us to the next step in determining and quantifying EPS production. Moreover,
vps in-frame deletion mutants could be constructed to test whether phage-induced biofilm/aggregate
formation is mediated by EPS production at the genetic level. Additionally, to better understand the genetic
regulatory network, bacterial transcriptomes could provide us with the bacterial strain's immediate genetic
response to phage exposure, enabling an assessment of the transcriptional pattern, such as genes involved
in polysaccharide production and flagella assembly (193). Furthermore, it is relevant to determine whether
phage KVP40-induced aggregates are related to host strain PF430-3 or to the phage KVP40 and could be
tested by combining phage KVP40 with susceptible V. anguillarum strains which have been characterized in
Paper I.
As previously mentioned, isolating vibriophages is tedious and isolation of a natural vibriophage which is
both specific to a certain host and expresses a relevant depolymerase is thus likely very low. Therefore,
using synthetic biology to engineer our broad-host-range phage KVP40 to express the most effective EPS-
degrading enzymes specific to the Vibrio targeting biofilms may be a way forward, as has been successfully
conducted in an E. coli study (194). This would allow us to develop a diverse library of biofilm-dispersing
phages rather than rely on isolating vibriophages from the environment.
Albeit the specific mechanism of resistance of strain BA35 was not identified in Paper II, the observation
that a large fraction of phage resistant mutants of BA35 were found to be either non-motile or having
severely impaired motility, which implies that flagella may play an important role in the infection of V.
anguillarum strain BA35 by phage ΦH20. This was consistent with recent observations that the flagellum
was involved in the initial infection of C. crescentus and S. enterica serovar typhimurium phages (148, 195).
Future work on whether phage-resistant mutants BA35 have mutations or down-regulations in genes
involved in both flagella and potential phage receptors should be carried out. Lastly, time-shift experiments

42
(196) will be conducted to gain new insights into the underlying processes of antagonistic co-evolution
between BA35 and phage ΦH20, if any.

43
Paper III
It is well known that mutations to phage resistance frequently have maladaptive pleiotropic effects (197,
198). It should be advantageous to avoid maintaining constantly elevated anti-phage defenses, if bacteria
would benefit from subjecting their anti-phage defenses to QS-control. Thus, future studies will focus on
whether QS-mediated OMP down-regulation gives rise to a reduction in relative fitness to elucidate the
regulatory pathway connecting QS-signaling to OMP down-regulation in V. anguillarum. Then it should be
possible to construct mutants, in which regulation of the relevant OMPs is uncoupled from QS, without
affecting the remainder of the QS regulon. The relative fitness of these mutants relative to the otherwise
isogenic parent strains will be measured in the absence of vibriophage.
Ultimately, to fully characterize the regulatory pathway underlying QS-mediated OmpK down-regulation
mechanisms, which have not been identified in Paper III, we will determine whether QS transcription factor
VanT binds directly to the promoter of putative targets, such as ompK and vps. Specifically, we will set up a
VanT Electrophoretic mobility shift assay (EMSA) experiment to test our hypothesis. Using this approach
would allow us to potentially expand the currently modest number of described targets mediated by VanT
in the literature.
Finally, the evidence that QS regulates phage receptor expression may potentially be used actively by
quenching QS signaling, hence preventing receptor down-regulation. However, in the case of V. cholerae, at
low cell densities, when the concentrations of the extracellular AIs are low, the expressions of virulence
factor toxin-co-regulated pilus, CT and biofilm formation are activated; at high cell densities, the
accumulation of two QS autoinducers (CAI-1 and AI-2) repress the expression of virulence factors and
biofilm formation (43, 104). If the close-related V. anguillarum strains behave similarly, it would be
problematic to synergize phage therapy and quorum quenching to prevent vibriosis infection. Further, it
would be interesting to target potential phage receptors which are not regulated by QS signaling molecules,
and by combining specific phages and QS molecules (CAI-1 and AI-2) may offer a synergistic antimicrobial
effect to prevent vibriosis infection in aquaculture industries.

44
References
1. FAO. 2014. The state of world fisheries and aquaculture. FAO Rome, Italy. 2. Bondad-Reantaso MG, Subasinghe RP, Arthur JR, Ogawa K, Chinabut S, Adlard R, Tan Z, Shariff M.
2005. Disease and health management in Asian aquaculture. Vet Parasitol 132:249-272. 3. Fryer JL, Hedrick RP. 2003. Piscirickettsia salmonis: a Gram-negative intracellular bacterial
pathogen of fish. J Fish Dis 26:251-262. 4. Nematollahi A, Decostere A, Pasmans F, Haesebrouck F. 2003. Flavobacterium psychrophilum
infections in salmonid fish. J Fish Dis 26:563-574. 5. Smith P, Davey S. 1993. Evidence for the competitive exclusion of Aeromonas salmonicida from fish
with stress-inducible furunculosis by a fluorescent pseudomonad. J Fish Dis 16:521-524. 6. Gratzek JB. 1981. An overview of ornamental fish diseases and therapy. J Small Anim Pract 22:345-
366. 7. Woo PT, Leatherland JF, Bruno DW. 2011. Fish diseases and disorders, vol 3. CABI. 8. Naylor RL, Goldburg RJ, Primavera JH, Kautsky N, Beveridge MC, Clay J, Folke C, Lubchenco J,
Mooney H, Troell M. 2000. Effect of aquaculture on world fish supplies. Nature 405:1017-1024. 9. Vázquez FJS, Muñoz-Cueto JA. 2014. Biology of European sea bass. CRC Press. 10. Austin B, Austin DA. 2007. Bacterial fish pathogens: disease of farmed and wild fish. Springer. 11. Frans I, Michiels CW, Bossier P, Willems KA, Lievens B, Rediers H. 2011. Vibrio anguillarum as a
fish pathogen: virulence factors, diagnosis and prevention. Journal of Fish Diseases 34:643-661. 12. Larsen JL, Pedersen K, Dalsgaard I. 1994. Vibrio anguillarum serovars associated with vibriosis in
fish. J Fish Dis 17:259-267. 13. Larsen JL, Rasmussen HB, Dalsgaard I. 1988. Study of Vibrio anguillarum strains from different
sources with emphasis on ecological and pathobiological properties. Appl Environ Microbiol 54:2264-2267.
14. Harrell LW, Novotny AJ, Schiewe M, Hodgins HO. 1976. Isolation and description of two vibrios pathogenic to Pacific salmon. Fishery Bulletin 74:447-449.
15. MacDonell M, Colwell R. 1985. Phylogeny of the Vibrionaceae, and recommendation for two new genera, Listonella and Shewanella. Syst Appl Microbiol 6:171-182.
16. Thompson FL, Thompson CC, Dias GM, Naka H, Dubay C, Crosa JH. 2011. The genus Listonella MacDonell and Colwell 1986 is a later heterotypic synonym of the genus Vibrio pacini 1854 (Approved Lists 1980)-a taxonomic opinion. Int J Syst Evol Microbiol 61:3023-3027.
17. Naka H, Dias GM, Thompson CC, Dubay C, Thompson FL, Crosa JH. 2011. Complete genome sequence of the marine fish pathogen Vibrio anguillarum harboring the pJM1 virulence plasmid and genomic comparison with other virulent strains of V. anguillarum and V. ordalii. Infect Immun 79:2889-2900.
18. Tan D, Gram L, Middelboe M. 2014. Vibriophages and their interactions with the fish pathogen Vibrio anguillarum. Appl Environ Microbiol 80:3128-3140.
19. Rodríguez JM, López-Romalde S, Beaz R, Alonso M, Castro D, Romalde JL. 2006. Molecular fingerprinting of Vibrio tapetis strains using three PCR-based methods: ERIC-PCR, REP-PCR and RAPD. Dis Aquat Organ 69:175-183.
20. Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. 2000. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol 38:1008-1015.
21. Skov MN, Pedersen K, Larsen JL. 1995. Comparison of Pulsed-Field Gel Electrophoresis, Ribotyping, and Plasmid Profiling for Typing of Vibrio anguillarum Serovar O1. Appl Environ Microbiol 61:1540-1545.
22. Toranzo A, Santos Y, Barja J. 1996. Immunization with bacterial antigens: Vibrio infections. Dev Biol Stand 90:93-105.

45
23. Rasmussen HB. 1987. Subgrouping of lipopolysaccharide O antigens from Vibrio anguillarum serogroup O2 by immunoelectrophoretic analyses. Curr Microbiol 16:39-42.
24. Pereira CS, Thompson JA, Xavier KB. 2013. AI-2-mediated signalling in bacteria. FEMS Microbiol Rev 37:156-181.
25. Miller MB, Bassler BL. 2001. Quorum sensing in bacteria. Annu Rev Microbiol 55:165-199. 26. Fuqua C, Parsek MR, Greenberg EP. 2001. Regulation of gene expression by cell-to-cell
communication: acyl-homoserine lactone quorum sensing. Annu Rev Genet 35:439-468. 27. Fuqua WC, Winans SC, Greenberg EP. 1994. Quorum sensing in bacteria: the LuxR-LuxI family of
cell density-responsive transcriptional regulators. J Bacteriol 176:269-275. 28. Ng WL, Bassler BL. 2009. Bacterial quorum-sensing network architectures. Annu Rev Genet 43:197-
222. 29. Ruby EG. 1996. Lessons from a cooperative, bacterial-animal association: the Vibrio fischeri-
Euprymna scolopes light organ symbiosis. Annu Rev Microbiol 50:591-624. 30. Engebrecht J, Silverman M. 1984. Identification of genes and gene products necessary for bacterial
bioluminescence. Proc Natl Acad Sci U S A 81:4154-4158. 31. Schaefer AL, Val DL, Hanzelka BL, Cronan JE, Jr., Greenberg EP. 1996. Generation of cell-to-cell
signals in quorum sensing: acyl homoserine lactone synthase activity of a purified Vibrio fischeri LuxI protein. Proc Natl Acad Sci U S A 93:9505-9509.
32. Stevens AM, Dolan KM, Greenberg EP. 1994. Synergistic binding of the Vibrio fischeri LuxR transcriptional activator domain and RNA polymerase to the lux promoter region. Proc Natl Acad Sci U S A 91:12619-12623.
33. Croxatto A, Pride J, Hardman A, Williams P, Camara M, Milton DL. 2004. A distinctive dual-channel quorum-sensing system operates in Vibrio anguillarum. Mol Microbiol 52:1677-1689.
34. Croxatto A, Chalker VJ, Lauritz J, Jass J, Hardman A, Williams P, Camara M, Milton DL. 2002. VanT, a homologue of Vibrio harveyi LuxR, regulates serine, metalloprotease, pigment, and biofilm production in Vibrio anguillarum. J Bacteriol 184:1617-1629.
35. Weber B, Lindell K, El Qaidi S, Hjerde E, Willassen NP, Milton DL. 2011. The phosphotransferase VanU represses expression of four qrr genes antagonizing VanO-mediated quorum-sensing regulation in Vibrio anguillarum. Microbiology 157:3324-3339.
36. Weber B, Croxatto A, Chen C, Milton DL. 2008. RpoS induces expression of the Vibrio anguillarum quorum-sensing regulator VanT. Microbiology 154:767-780.
37. Milton DL, Chalker VJ, Kirke D, Hardman A, Cámara M, Williams P. 2001. The LuxM Homologue VanM from Vibrio anguillarum Directs the Synthesis of N-(3-Hydroxyhexanoyl) homoserine Lactone and N-Hexanoylhomoserine Lactone. J Bacteriol 183:3537-3547.
38. Bassler BL, Greenberg EP, Stevens AM. 1997. Cross-species induction of luminescence in the quorum-sensing bacterium Vibrio harveyi. J Bacteriol 179:4043-4045.
39. Costerton JW, Stewart PS, Greenberg EP. 1999. Bacterial biofilms: a common cause of persistent infections. Science 284:1318-1322.
40. Bassler BL, Wright M, Showalter RE, Silverman MR. 1993. Intercellular signalling in Vibrio harveyi: sequence and function of genes regulating expression of luminescence. Mol Microbiol 9:773-786.
41. Bassler BL, Losick R. 2006. Bacterially speaking. Cell 125:237-246. 42. Li YH, Tang N, Aspiras MB, Lau PCY, Lee JH, Ellen RP, Cvitkovitch DG. 2002. A quorum-sensing
signaling system essential for genetic competence in Streptococcus mutans is involved in biofilm formation. J Bacteriol 184:2699-2708.
43. Zhu J, Miller MB, Vance RE, Dziejman M, Bassler BL, Mekalanos JJ. 2002. Quorum-sensing regulators control virulence gene expression in Vibrio cholerae. Proc Natl Acad Sci U S A 99:3129-3134.
44. Quinones B, Dulla G, Lindow SE. 2005. Quorum sensing regulates exopolysaccharide production, motility, and virulence in Pseudomonas syringae. Mol Plant Microbe Interact 18:682-693.

46
45. Hoyland-Kroghsbo NM, Maerkedahl RB, Svenningsen SL. 2013. A quorum-sensing-induced bacteriophage defense mechanism. MBio 4:e00362-00312.
46. Ghosh D, Roy K, Williamson KE, Srinivasiah S, Wommack KE, Radosevich M. 2009. Acyl-Homoserine Lactones Can Induce Virus Production in Lysogenic Bacteria: an Alternative Paradigm for Prophage Induction. Appl Environ Microbiol 75:7142-7152.
47. Hammer BK, Bassler BL. 2003. Quorum sensing controls biofilm formation in Vibrio cholerae. Mol Microbiol 50:101-104.
48. Solano C, Echeverz M, Lasa I. 2014. Biofilm dispersion and quorum sensing. Curr Opin Microbiol 18:96-104.
49. Hall-Stoodley L, Costerton JW, Stoodley P. 2004. Bacterial biofilms: from the natural environment to infectious diseases. Nat Rev Microbiol 2:95-108.
50. Croxatto A, Lauritz J, Chen C, Milton DL. 2007. Vibrio anguillarum colonization of rainbow trout integument requires a DNA locus involved in exopolysaccharide transport and biosynthesis. Environ Microbiol 9:370-382.
51. Lindell K, Fahlgren A, Hjerde E, Willassen N-P, Fällman M, Milton DL. 2012. Lipopolysaccharide O-antigen prevents phagocytosis of Vibrio anguillarum by rainbow trout (Oncorhynchus mykiss) skin epithelial cells. PloS one 7:e37678.
52. Stewart PS. 2002. Mechanisms of antibiotic resistance in bacterial biofilms. Int J Med Microbiol 292:107-113.
53. Flemming H-C, Wingender J. 2010. The biofilm matrix. Nat Rev Microbiol 8:623-633. 54. Whitchurch CB, Tolker-Nielsen T, Ragas PC, Mattick JS. 2002. Extracellular DNA required for
bacterial biofilm formation. Science 295:1487-1487. 55. Seper A, Fengler VH, Roier S, Wolinski H, Kohlwein SD, Bishop AL, Camilli A, Reidl J, Schild S. 2011.
Extracellular nucleases and extracellular DNA play important roles in Vibrio cholerae biofilm formation. Mol Microbiol 82:1015-1037.
56. Mann EE, Rice KC, Boles BR, Endres JL, Ranjit D, Chandramohan L, Tsang LH, Smeltzer MS, Horswill AR, Bayles KW. 2009. Modulation of eDNA release and degradation affects Staphylococcus aureus biofilm maturation. PLoS One 4:e5822.
57. Thomas VC, Thurlow LR, Boyle D, Hancock LE. 2008. Regulation of autolysis-dependent extracellular DNA release by Enterococcus faecalis extracellular proteases influences biofilm development. J Bacteriol 190:5690-5698.
58. Carrolo M, Frias MJ, Pinto FR, Melo-Cristino J, Ramirez M. 2010. Prophage spontaneous activation promotes DNA release enhancing biofilm formation in Streptococcus pneumoniae. PLoS One 5:e15678.
59. Gödeke J, Paul K, Lassak J, Thormann KM. 2010. Phage-induced lysis enhances biofilm formation in Shewanella oneidensis MR-1. ISME J 5:613-626.
60. Watnick PI, Kolter R. 1999. Steps in the development of a Vibrio cholerae El Tor biofilm. Mol Microbiol 34:586-595.
61. Donlan RM, Costerton JW. 2002. Biofilms: survival mechanisms of clinically relevant microorganisms. Clin Microbiol Rev 15:167-193.
62. Monroe D. 2007. Looking for chinks in the armor of bacterial biofilms. PLoS Biol 5:e307. 63. Oliver A, Canton R, Campo P, Baquero F, Blazquez J. 2000. High frequency of hypermutable
Pseudomonas aeruginosa in cystic fibrosis lung infection. Science 288:1251-1254. 64. Singh PK, Schaefer AL, Parsek MR, Moninger TO, Welsh MJ, Greenberg EP. 2000. Quorum-sensing
signals indicate that cystic fibrosis lungs are infected with bacterial biofilms. Nature 407:762-764. 65. Pratt LA, Kolter R. 1998. Genetic analysis of Escherichia coli biofilm formation: roles of flagella,
motility, chemotaxis and type I pili. Mol Microbiol 30:285-293. 66. Cooksey K. 1992. Extracellular polymers in biofilms, p 137-147, Biofilms-science and technology.
Springer.

47
67. Welch M, Mikkelsen H, Swatton JE, Smith D, Thomas GL, Glansdorp FG, Spring DR. 2005. Cell–cell communication in Gram-negative bacteria. Mol BioSyst 1:196-202.
68. Zhu J, Mekalanos JJ. 2003. Quorum sensing-dependent biofilms enhance colonization in Vibrio cholerae. Dev Cell 5:647-656.
69. Finkelstein RA, Boesman-Finkelstein M, Chang Y, Häse C. 1992. Vibrio cholerae hemagglutinin/protease, colonial variation, virulence, and detachment. Infect Immun 60:472-478.
70. Purevdorj-Gage B, Costerton WJ, Stoodley P. 2005. Phenotypic differentiation and seeding dispersal in non-mucoid and mucoid Pseudomonas aeruginosa biofilms. Microbiology 151:1569-1576.
71. Nadell CD, Xavier JB, Levin SA, Foster KR. 2008. The evolution of quorum sensing in bacterial biofilms. PLoS Biol 6:e14.
72. Hong SH, Hegde M, Kim J, Wang X, Jayaraman A, Wood TK. 2012. Synthetic quorum-sensing circuit to control consortial biofilm formation and dispersal in a microfluidic device. Nat Commun 3:613.
73. Sillankorva S, Neubauer P, Azeredo J. 2010. Phage control of dual species biofilms of Pseudomonas fluorescens and Staphylococcus lentus. Biofouling 26:567-575.
74. Barrios AFG, Zuo RJ, Hashimoto Y, Yang L, Bentley WE, Wood TK. 2006. Autoinducer 2 controls biofilm formation in Escherichia coli through a novel motility quorum-sensing regulator (MqsR, B3022). J Bacteriol 188:305-316.
75. Nielsen MW, Sternberg C, Molin S, Regenberg B. 2011. Pseudomonas aeruginosa and Saccharomyces cerevisiae Biofilm in Flow Cells. Jove-Journal of Visualized Experiments doi:Unsp E2383Doi 10.3791/2383:e2383-e2383.
76. Hosseinidoust Z, Tufenkji N, van de Ven TG. 2013. Formation of biofilms under phage predation: considerations concerning a biofilm increase. Biofouling 29:457-468.
77. Kreft JU, Picioreanu C, Wimpenny JW, van Loosdrecht MC. 2001. Individual-based modelling of biofilms. Microbiology 147:2897-2912.
78. Strathmann M, Wingender J, Flemming HC. 2002. Application of fluorescently labelled lectins for the visualization and biochemical characterization of polysaccharides in biofilms of Pseudomonas aeruginosa. J Microbiol Methods 50:237-248.
79. Fong JC, Syed KA, Klose KE, Yildiz FH. 2010. Role of Vibrio polysaccharide (vps) genes in VPS production, biofilm formation and Vibrio cholerae pathogenesis. Microbiology 156:2757-2769.
80. Whiteley M, Bangera MG, Bumgarner RE, Parsek MR, Teitzel GM, Lory S, Greenberg EP. 2001. Gene expression in Pseudomonas aeruginosa biofilms. Nature 413:860-864.
81. Bassler BL. 1999. How bacteria talk to each other: regulation of gene expression by quorum sensing. Curr Opin Microbiol 2:582-587.
82. Stewart PS, Franklin MJ. 2008. Physiological heterogeneity in biofilms. Nat Rev Microbiol 6:199-210. 83. Murphy TF, Kirkham C. 2002. Biofilm formation by nontypeable Haemophilus influenzae: strain
variability, outer membrane antigen expression and role of pili. BMC Microbiol 2:7. 84. Branda SS, Vik Å, Friedman L, Kolter R. 2005. Biofilms: the matrix revisited. Trends Microbiol
13:20-26. 85. Matz C, McDougald D, Moreno AM, Yung PY, Yildiz FH, Kjelleberg S. 2005. Biofilm formation and
phenotypic variation enhance predation-driven persistence of Vibrio cholerae. Proc Natl Acad Sci U S A 102:16819-16824.
86. Stoodley P, Sauer K, Davies DG, Costerton JW. 2002. Biofilms as complex differentiated communities. Annu Rev Microbiol 56:187-209.
87. Kreft J-U, Picioreanu C, Wimpenny JW, van Loosdrecht MC. 2001. Individual-based modelling of biofilms. Microbiology 147:2897-2912.
88. Nagaoka H, Ueda S, Miya A. 1996. Influence of bacterial extracellular polymers on the membrane separation activated sludge process. Water Sci Technol 34:165-172.
89. Reisner A, Haagensen JA, Schembri MA, Zechner EL, Molin S. 2003. Development and maturation of Escherichia coli K-12 biofilms. Mol Microbiol 48:933-946.

48
90. Parsek MR, Greenberg EP. 2005. Sociomicrobiology: the connections between quorum sensing and biofilms. Trends Microbiol 13:27-33.
91. Costerton JW, Post JC, Ehrlich GD, Hu FZ, Kreft R, Nistico L, Kathju S, Stoodley P, Hall‐Stoodley L, Maale G. 2011. New methods for the detection of orthopedic and other biofilm infections. FEMS Immunol Med Microbiol 61:133-140.
92. Faruque SM, Biswas K, Udden SMN, Ahmad QS, Sack DA, Nair GB, Mekalanos JJ. 2006. Transmissibility of cholera: In vivo-formed biofilms and their relationship to infectivity and persistence in the environment. Proc Natl Acad Sci U S A 103:6350-6355.
93. Costerton W, Veeh R, Shirtliff M, Pasmore M, Post C, Ehrlich G. 2003. The application of biofilm science to the study and control of chronic bacterial infections. J Clin Invest 112:1466-1477.
94. Bari SM, Roky MK, Mohiuddin M, Kamruzzaman M, Mekalanos JJ, Faruque SM. 2013. Quorum-sensing autoinducers resuscitate dormant Vibrio cholerae in environmental water samples. Proc Natl Acad Sci U S A 110:9926-9931.
95. Reissbrodt R, Rienaecker I, Romanova JM, Freestone PP, Haigh RD, Lyte M, Tschape H, Williams PH. 2002. Resuscitation of Salmonella enterica serovar typhimurium and enterohemorrhagic Escherichia coli from the viable but nonculturable state by heat-stable enterobacterial autoinducer. Appl Environ Microbiol 68:4788-4794.
96. Ayrapetyan M, Williams TC, Oliver JD. 2014. Interspecific quorum sensing mediates the resuscitation of viable but nonculturable vibrios. Appl Environ Microbiol 80:2478-2483.
97. Colquhoun DJ, Aarflot L, Melvold CF. 2007. gyrA and parC Mutations and associated quinolone resistance in Vibrio anguillarum serotype O2b strains isolated from farmed Atlantic cod (Gadus morhua) in Norway. Antimicrob Agents Chemother 51:2597-2599.
98. Drenkard E. 2003. Antimicrobial resistance of Pseudomonas aeruginosa biofilms. Microbes Infect 5:1213-1219.
99. Cabello FC. 2006. Heavy use of prophylactic antibiotics in aquaculture: a growing problem for human and animal health and for the environment. Environ Microbiol 8:1137-1144.
100. Holmström K, Gräslund S, Wahlström A, Poungshompoo S, Bengtsson BE, Kautsky N. 2003. Antibiotic use in shrimp farming and implications for environmental impacts and human health. Int J Food Sci Technol 38:255-266.
101. Defoirdt T, Sorgeloos P, Bossier P. 2011. Alternatives to antibiotics for the control of bacterial disease in aquaculture. Curr Opin Microbiol 14:251-258.
102. SANTOS RYUSC, ESTEVEZ TAUSC, BARJA PJLUSC. 2000. ANTI-$i(VIBRIO ANGUILLARUM) VACCINE (GAVA-3) FOR THE PREVENTION OF THE VIBRIOSIS DISEASE IN THE TURBOT AND SALMONIDAE, AND PREPARATION PROCESS. Google Patents.
103. O'Loughlin CT, Miller LC, Siryaporn A, Drescher K, Semmelhack MF, Bassler BL. 2013. A quorum-sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proc Natl Acad Sci U S A 110:17981-17986.
104. Higgins DA, Pomianek ME, Kraml CM, Taylor RK, Semmelhack MF, Bassler BL. 2007. The major Vibrio cholerae autoinducer and its role in virulence factor production. Nature 450:883-886.
105. Vendeville A, Winzer K, Heurlier K, Tang CM, Hardie KR. 2005. Making'sense'of metabolism: autoinducer-2, LuxS and pathogenic bacteria. Nat Rev Microbiol 3:383-396.
106. Surette MG, Bassler BL. 1998. Quorum sensing in Escherichia coli and Salmonella typhimurium. Proc Natl Acad Sci U S A 95:7046-7050.
107. Burmølle M, Thomsen TR, Fazli M, Dige I, Christensen L, Homøe P, Tvede M, Nyvad B, Tolker‐Nielsen T, Givskov M. 2010. Biofilms in chronic infections-a matter of opportunity-monospecies biofilms in multispecies infections. FEMS Immunol Med Microbiol 59:324-336.
108. Pei R, Lamas-Samanamud GR. 2014. Inhibition of biofilm formation by t7 bacteriophages producing quorum-quenching enzymes. Appl Environ Microbiol 80:5340-5348.
109. Dublanchet A, Patey O. 2008. Bacterial infection-update on phage therapy. Med Mal Infect 38:407-409.

49
110. Middelboe M. 2008. Microbial disease in the sea: effects of viruses on carbon and nutrient cycling. Princeton University Press, Princeton, NJ.
111. Ackermann H-W. 2007. 5500 Phages examined in the electron microscope. Arch Virol 152:227-243. 112. Oppenheim AB, Kobiler O, Stavans J, Court DL, Adhya S. 2005. Switches in bacteriophage lambda
development. Annu Rev Genet 39:409-429. 113. Roberts JW, Roberts CW. 1975. Proteolytic cleavage of bacteriophage lambda repressor in
induction. Proc Natl Acad Sci U S A 72:147-151. 114. Faruque SM, Naser IB, Islam MJ, Faruque AS, Ghosh AN, Nair GB, Sack DA, Mekalanos JJ. 2005.
Seasonal epidemics of cholera inversely correlate with the prevalence of environmental cholera phages. Proc Natl Acad Sci U S A 102:1702-1707.
115. Waldor MK, Mekalanos JJ. 1996. Lysogenic conversion by a filamentous phage encoding cholera toxin. Science 272:1910-1914.
116. Wang X, Kim Y, Ma Q, Hong SH, Pokusaeva K, Sturino JM, Wood TK. 2010. Cryptic prophages help bacteria cope with adverse environments. Nat Commun 1:147.
117. Wang X, Kim Y, Wood TK. 2009. Control and benefits of CP4-57 prophage excision in Escherichia coli biofilms. ISME J 3:1164-1179.
118. Comeau AM, Chan AM, Suttle CA. 2006. Genetic richness of vibriophages isolated in a coastal environment. Environ Microbiol 8:1164-1176.
119. Kellogg CA, Rose JB, Jiang SC, Thurmond JM, Paul JH. 1995. Genetic Diversity of Related Vibriophages Isolated from Marine Environments around Florida and Hawaii, USA. Mar Ecol Prog Ser 120:89-98.
120. Reidl J, Mekalanos JJ. 1995. Characterization of Vibrio cholerae bacteriophage K139 and use of a novel mini‐transposon to identify a phage-encoded virulence factor. Mol Microbiol 18:685-701.
121. Lin W, Fullner KJ, Clayton R, Sexton JA, Rogers MB, Calia KE, Calderwood SB, Fraser C, Mekalanos JJ. 1999. Identification of a Vibrio cholerae RTX toxin gene cluster that is tightly linked to the cholera toxin prophage. Proc Natl Acad Sci U S A 96:1071-1076.
122. Matsuzaki S, Tanaka S, Koga T, Kawata T. 1992. A broad-host-range vibriophage, KVP40, isolated from sea water. Microbiol Immunol 36:93-97.
123. Miller ES, Heidelberg JF, Eisen JA, Nelson WC, Durkin AS, Ciecko A, Feldblyum TV, White O, Paulsen IT, Nierman WC, Lee J, Szczypinski B, Fraser CM. 2003. Complete genome sequence of the broad-host-range vibriophage KVP40: comparative genomics of a T4-related bacteriophage. J Bacteriol 185:5220-5233.
124. Inoue T, Matsuzaki S, Tanaka S. 1995. A 26-kDa outer membrane protein, OmpK, common to Vibrio species is the receptor for a broad-host-range vibriophage, KVP40. FEMS Microbiol Lett 125:101-105.
125. Fortuna W, Miedzybrodzki R, Weber-Dabrowska B, Gorski A. 2008. Bacteriophage therapy in children: facts and prospects. Med Sci Monit 14:RA126-132.
126. Lang LH. 2006. FDA approves use of bacteriophages to be added to meat and poultry products. Gastroenterol 131:1370.
127. Imbeault S, Parent S, Lagacé M, Uhland CF, Blais JF. 2006. Using bacteriophages to prevent furunculosis caused by Aeromonas salmonicida in farmed brook trout. J Aquat Anim Health 18:203-214.
128. Stevenson R, Airdrie D. 1984. Isolation of Yersinia ruckeri bacteriophages. Appl Environ Microbiol 47:1201-1205.
129. Nakai T. 2003. Bacteriophage control of Pseudomonas plecoglossicida infection in ayu Plecoglossus altivelis. Dis Aquat Organ 53:33-39.
130. Stenholm AR, Dalsgaard I, Middelboe M. 2008. Isolation and characterization of bacteriophages infecting the fish pathogen Flavobacterium psychrophilum. Appl Environ Microbiol 74:4070-4078.

50
131. Castillo D, Higuera G, Villa M, Middelboe M, Dalsgaard I, Madsen L, Espejo R. 2012. Diversity of Flavobacterium psychrophilum and the potential use of its phages for protection against bacterial cold water disease in salmonids. J Fish Dis 35:193-201.
132. Nakai T, Sugimoto R, Park K, Matsuoka S, Mori K-i, Nishioka T, Maruyama K. 1999. Protective effects of bacteriophage on experimental Lactococcus garvieae infection in yellowtail. Dis Aquat Organ 37:33-41.
133. Vinod MG, Shivu MM, Umesha KR, Rajeeva BC, Krohne G, Ka-Unasagar I, Karunasagar I. 2006. Isolation of Vibrio harveyi bacteriophage with a potential for biocontrol of luminous vibriosis in hatchery environments. Aquaculture 255:117-124.
134. Higuera G, Bastías R, Tsertsvadze G, Romero J, Espejo RT. 2013. Recently discovered Vibrio anguillarum phages can protect against experimentally induced vibriosis in Atlantic salmon, Salmo salar. Aquaculture 392:128-133.
135. Silva YJ, Costa L, Pereira C, Mateus C, Cunha A, Calado R, Gomes NC, Pardo MA, Hernandez I, Almeida A. 2014. Phage therapy as an approach to prevent Vibrio anguillarum infections in fish larvae production. PLoS One 9:e114197.
136. Lomelí-Ortega CO, Martínez-Díaz SF. 2014. Phage therapy against Vibrio parahaemolyticus infection in the whiteleg shrimp (Litopenaeus vannamei) larvae. Aquaculture 434:208-211.
137. Faruque SM, Mekalanos JJ. 2012. Phage-bacterial interactions in the evolution of toxigenic Vibrio cholerae. Virulence 3:556-565.
138. Jaiswal A, Koley H, Ghosh A, Palit A, Sarkar B. 2013. Efficacy of cocktail phage therapy in treating Vibrio cholerae infection in rabbit model. Microbes Infect 15:152-156.
139. Jensen MA, Faruque SM, Mekalanos JJ, Levin BR. 2006. Modeling the role of bacteriophage in the control of cholera outbreaks. Proc Natl Acad Sci U S A 103:4652-4657.
140. Suttle CA. 2005. Viruses in the sea. Nature 437:356-361. 141. Labrie SJ, Samson JE, Moineau S. 2010. Bacteriophage resistance mechanisms. Nat Rev Microbiol
8:317-327. 142. Samson JE, Magadan AH, Sabri M, Moineau S. 2013. Revenge of the phages: defeating bacterial
defences. Nat Rev Microbiol 11:675-687. 143. Rakhuba DV, Kolomiets EI, Dey ES, Novik GI. 2010. Bacteriophage Receptors, Mechanisms of
Phage Adsorption and Penetration into Host Cell. Pol J Microbiol 59:145-155. 144. Samuel AD, Pitta TP, Ryu WS, Danese PN, Leung EC, Berg HC. 1999. Flagellar determinants of
bacterial sensitivity to chi-phage. Proc Natl Acad Sci U S A 96:9863-9866. 145. Hantke K. 1978. Major outer membrane proteins of E. coli K12 serve as receptors for the phages T2
(protein Ia) and 434 (protein Ib). Mol Gen Genet 164:131-135. 146. Randall-Hazelbauer L, Schwartz M. 1973. Isolation of the bacteriophage lambda receptor from
Escherichia coli. J Bacteriol 116:1436-1446. 147. Lindberg AA. 1973. Bacteriophage receptors. Annu Rev Microbiol 27:205-241. 148. Guerrero-Ferreira RC, Viollier PH, Ely B, Poindexter JS, Georgieva M, Jensen GJ, Wright ER. 2011.
Alternative mechanism for bacteriophage adsorption to the motile bacterium Caulobacter crescentus. Proc Natl Acad Sci U S A 108:9963-9968.
149. Valyasevi R, Sandine WE, Geller BL. 1990. The Bacteriophage Kh Receptor of Lactococcus lactis Subsp. cremoris Kh Is the Rhamnose of the Extracellular Wall Polysaccharide. Appl Environ Microbiol 56:1882-1889.
150. McCabe O, Spinelli S, Farenc C, Labbé M, Tremblay D, Blangy S, Oscarson S, Moineau S, Cambillau C. 2015. The targeted recognition of Lactococcus lactis phages to their polysaccharide receptors. Mol Microbiol.
151. Seed KD, Faruque SM, Mekalanos JJ, Calderwood SB, Qadri F, Camilli A. 2012. Phase variable O antigen biosynthetic genes control expression of the major protective antigen and bacteriophage receptor in Vibrio cholerae O1. PLoS Pathog 8:e1002917.

51
152. Geller BL, Ngo HT, Mooney DT, Su P, Dunn N. 2005. Lactococcal 936-species phage attachment to surface of Lactococcus lactis. J Dairy Sci 88:900-907.
153. Mahony J, Kot W, Murphy J, Ainsworth S, Neve H, Hansen LH, Heller KJ, Sorensen SJ, Hammer K, Cambillau C, Vogensen FK, van Sinderen D. 2013. Investigation of the relationship between lactococcal host cell wall polysaccharide genotype and 936 phage receptor binding protein phylogeny. Appl Environ Microbiol 79:4385-4392.
154. Forde A, Fitzgerald GF. 1999. Analysis of exopolysaccharide (EPS) production mediated by the bacteriophage adsorption blocking plasmid, pCI658, isolated from Lactococcus lactis ssp cremoris HO2. Int Dairy J 9:465-472.
155. Morona R, Klose M, Henning U. 1984. Escherichia coli K-12 outer membrane protein (OmpA) as a bacteriophage receptor: analysis of mutant genes expressing altered proteins. J Bacteriol 159:570-578.
156. Luckey M, Nikaido H. 1980. Specificity of diffusion channels produced by lambda phage receptor protein of Escherichia coli. Proc Natl Acad Sci U S A 77:167-171.
157. Harcombe WR, Bull JJ. 2005. Impact of phages on two-species bacterial communities. Appl Environ Microbiol 71:5254-5259.
158. Kay MK, Erwin TC, McLean RJ, Aron GM. 2011. Bacteriophage ecology in Escherichia coli and Pseudomonas aeruginosa mixed-biofilm communities. Appl Environ Microbiol 77:821-829.
159. Ferenci T, Schwentorat M, Ullrich S, Vilmart J. 1980. Lambda receptor in the outer membrane of Escherichia coli as a binding protein for maltodextrins and starch polysaccharides. J Bacteriol 142:521-526.
160. Park SC, Shimamura I, Fukunaga M, Mori KI, Nakai T. 2000. Isolation of bacteriophages specific to a fish pathogen, Pseudomonas plecoglossicida, as a candidate for disease control. Appl Environ Microbiol 66:1416-1422.
161. Smith HW, Huggins MB. 1983. Effectiveness of phages in treating experimental Escherichia coli diarrhoea in calves, piglets and lambs. J Gen Microbiol 129:2659-2675.
162. Hamod MA, Nithin MS, Shukur YN, Karunasagar I, Karunasagar I. 2012. Outer membrane protein K as a subunit vaccine against V. anguillarum. Aquaculture 354:107-110.
163. Nikaido H. 2003. Molecular basis of bacterial outer membrane permeability revisited. Microbiol Mol Biol Rev 67:593-656.
164. Rodkhum C, Hirono I, Stork M, Di Lorenzo M, Crosa JH, Aoki T. 2006. Putative virulence-related genes in Vibrio anguillarum identified by random genome sequencing. J Fish Dis 29:157-166.
165. Schrag SJ, Mittler JE. 1996. Host-parasite coexistence: The role of spatial refuges in stabilizing bacteria-phage interactions. Am Nat 148:348-377.
166. Brockhurst MA, Buckling A, Rainey PB. 2006. Spatial heterogeneity and the stability of host-parasite coexistence. J Evol Biol 19:374-379.
167. Heilmann S, Sneppen K, Krishna S. 2012. Coexistence of phage and bacteria on the boundary of self-organized refuges. Proc Natl Acad Sci U S A 109:12828-12833.
168. Brockhurst MA, Rainey PB, Buckling A. 2004. The effect of spatial heterogeneity and parasites on the evolution of host diversity. Proc Biol Sci 271:107-111.
169. Tock MR, Dryden DT. 2005. The biology of restriction and anti-restriction. Curr Opin Microbiol 8:466-472.
170. Pingoud A, Fuxreiter M, Pingoud V, Wende W. 2005. Type II restriction endonucleases: structure and mechanism. Cell Mol Life Sci 62:685-707.
171. Matsuoka S, Arai T, Murayama R, Kawamura F, Asai K, Sadaie Y. 2004. Identification of the nonA and nonB loci of Bacillus subtilis Marburg permitting the growth of SP10 phage. Genes Genet Syst 79:311-317.
172. Yamamoto T, Obana N, Yee LM, Asai K, Nomura N, Nakamura K. 2014. SP10 Infectivity Is Aborted after Bacteriophage SP10 Infection Induces nonA Transcription on the Prophage SPβ Region of the Bacillus subtilis Genome. J Bacteriol 196:693-706.

52
173. Krüger D, Bickle TA. 1983. Bacteriophage survival: multiple mechanisms for avoiding the deoxyribonucleic acid restriction systems of their hosts. Microbiol Rev 47:345.
174. Barrangou R, Fremaux C, Deveau H, Richards M, Boyaval P, Moineau S, Romero DA, Horvath P. 2007. CRISPR provides acquired resistance against viruses in prokaryotes. Science 315:1709-1712.
175. Deveau H, Garneau JE, Moineau S. 2010. CRISPR/Cas system and its role in phage-bacteria interactions. Annu Rev Microbiol 64:475-493.
176. Garneau JE, Dupuis ME, Villion M, Romero DA, Barrangou R, Boyaval P, Fremaux C, Horvath P, Magadan AH, Moineau S. 2010. The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA. Nature 468:67-71.
177. Chopin MC, Chopin A, Bidnenko E. 2005. Phage abortive infection in lactococci: variations on a theme. Curr Opin Microbiol 8:473-479.
178. Rossmann FS, Racek T, Wobser D, Puchalka J, Rabener EM, Reiger M, Hendrickx A, Diederich A-K, Jung K, Klein C. 2015. Phage-mediated Dispersal of Biofilm and Distribution of Bacterial Virulence Genes Is Induced by Quorum Sensing. PLoS Pathog 11:e1004653-e1004653.
179. Hargreaves KR, Kropinski AM, Clokie MR. 2014. What does the talking?: quorum sensing signalling genes discovered in a bacteriophage genome. PloS one 9:e85131.
180. Kropinski AM. 2000. Sequence of the genome of the temperate, serotype-converting, Pseudomonas aeruginosa bacteriophage D3. J Bacteriol 182:6066-6074.
181. Adriaenssens EM, Mattheus W, Cornelissen A, Shaburova O, Krylov VN, Kropinski AM, Lavigne R. 2012. Complete genome sequence of the giant Pseudomonas phage Lu11. J Virol 86:6369-6370.
182. Leblanc C, Caumont-Sarcos A, Comeau AM, Krisch HM. 2009. Isolation and genomic characterization of the first phage infecting Iodobacteria: ϕPLPE, a myovirus having a novel set of features. Environ Microbiol Rep 1:499-509.
183. Lenski RE. 1988. Dynamics of interactions between bacteria and virulent bacteriophage, p 1-44, Advances in microbial ecology. Springer.
184. Lennon JT, Khatana SAM, Marston MF, Martiny JB. 2007. Is there a cost of virus resistance in marine cyanobacteria? ISME J 1:300-312.
185. Smith HW, Huggins MB. 1982. Successful Treatment of Experimental Escherichia Coli Infections in Mice Using Phage-Its General Superiority over Antibiotics. J Gen Microbiol 128:307-318.
186. Van Valen L. 1977. The red queen. Am Nat:809-810. 187. Stern A, Sorek R. 2011. The phage-host arms race: shaping the evolution of microbes. Bioessays
33:43-51. 188. Lapchin L, Guillemaud T. 2005. Asymmetry in host and parasitoid diffuse coevolution: when the red
queen has to keep a finger in more than one pie. Front Zool 2:4. 189. Frada M, Probert I, Allen MJ, Wilson WH, de Vargas C. 2008. The “Cheshire Cat” escape strategy of
the coccolithophore Emiliania huxleyi in response to viral infection. Proc Natl Acad Sci U S A 105:15944-15949.
190. Davison S, Couture-Tosi E, Candela T, Mock M, Fouet A. 2005. Identification of the Bacillus anthracis (gamma) phage receptor. J Bacteriol 187:6742-6749.
191. Jończyk E, Kłak M, Międzybrodzki R, Górski A. 2011. The influence of external factors on bacteriophages-review. Folia Microbiol 56:191-200.
192. Brussow H, Canchaya C, Hardt WD. 2004. Phages and the evolution of bacterial pathogens: from genomic rearrangements to lysogenic conversion. Microbiol Mol Biol Rev 68:560-602, table of contents.
193. Conway T, Schoolnik GK. 2003. Microarray expression profiling: capturing a genome-wide portrait of the transcriptome. Mol Microbiol 47:879-889.
194. Lu TK, Collins JJ. 2007. Dispersing biofilms with engineered enzymatic bacteriophage. Proc Natl Acad Sci U S A 104:11197-11202.
195. Choi Y, Shin H, Lee JH, Ryu S. 2013. Identification and characterization of a novel flagellum-dependent Salmonella-infecting bacteriophage, iEPS5. Appl Environ Microbiol 79:4829-4837.

53
196. Gaba S, Ebert D. 2009. Time-shift experiments as a tool to study antagonistic coevolution. Trends Ecol Evol 24:226-232.
197. Lenski RE. 1988. Experimental studies of pleiotropy and epistasis in Escherichia coli. I. Variation in competitive fitness among mutants resistant to virus T4. Evolution:425-432.
198. Lenski RE, Levin BR. 1985. Constraints on the Coevolution of Bacteria and Virulent Phage-a Model, Some Experiments, and Predictions for Natural Communities. Am Nat 125:585-602.

54
Accompanying papers

Published Ahead of Print 7 March 2014. 10.1128/AEM.03544-13.
2014, 80(10):3128. DOI:Appl. Environ. Microbiol. Demeng Tan, Lone Gram and Mathias Middelboe the Fish Pathogen Vibrio anguillarumVibriophages and Their Interactions with
http://aem.asm.org/content/80/10/3128Updated information and services can be found at:
These include:
SUPPLEMENTAL MATERIAL Supplemental material
REFERENCEShttp://aem.asm.org/content/80/10/3128#ref-list-1at:
This article cites 46 articles, 12 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from
on A
pril 29, 2014 by Copenhagen U
niversity Libraryhttp://aem
.asm.org/
Dow
nloaded from

Vibriophages and Their Interactions with the Fish Pathogen Vibrioanguillarum
Demeng Tan,a Lone Gram,b Mathias Middelboea
Marine Biological Section, University of Copenhagen, Helsingør, Denmarka; Department of Systems Biology, Technical University of Denmark, Kongens Lyngby, Denmarkb
Vibrio anguillarum is an important pathogen in aquaculture, responsible for the disease vibriosis in many fish and invertebratespecies. Disease control by antibiotics is a concern due to potential development and spread of antibiotic resistance. The use ofbacteriophages to control the pathogen may offer a non-antibiotic-based approach to reduce vibriosis. A detailed understandingof the phage-host interaction is needed to evaluate the potential of phages to control the pathogen. In this study, we examinedthe diversity and interactions of 11 vibriophages, 24 V. anguillarum strains, and 13 Vibrio species strains. Together, the hostranges of the 11 phages covered all of the tested 37 Vibrio sp. host strains, which represented considerable temporal (20 years)and geographical (9 countries) differences in their origins of isolation. Thus, despite the occurrence of unique susceptibility pat-terns of the individual host isolates, key phenotypic properties related to phage susceptibility are distributed worldwide andmaintained in the global Vibrio community for decades. The phage susceptibility pattern of the isolates did not show any rela-tion to the physiological relationships obtained from Biolog GN2 profiles, demonstrating that similar phage susceptibility pat-terns occur across broad phylogenetic and physiological differences in Vibrio strains. Subsequent culture experiments with twophages and two V. anguillarum hosts demonstrated an initial strong lytic potential of the phages. However, rapid regrowth ofboth phage-resistant and phage-sensitive cells following the initial lysis suggested that several mechanisms of protection againstphage infection had developed in the host populations.
Vibriosis is one of the most prevalent and devastating diseasesin marine aquaculture, causing substantial mortality and
economic losses in both fish and shellfish cultures worldwide (1).The disease is caused by the marine pathogen Vibrio (Listonella)anguillarum, which infects �50 different species of fish and shell-fish, with severe implications for the marine fish-rearing industry(1, 2). Vaccines against vibriosis have been developed and havebeen widely successful in grown fish (3). However, the larval stageis especially vulnerable to V. anguillarum infections, as the im-mune system is not fully developed and vaccines are not effective.Traditionally, antibiotics have been widely used in antimicrobialprophylaxis and treatment of vibriosis in aquaculture, despiteconcern about the development and dispersal of antibiotic resis-tance in the pathogen community. Tetracycline and quinolonesare still the first drugs of choice, and multiple cases of antibioticresistance have been reported for V. anguillarum in aquacultureenvironments (4, 5). Therefore, there is a need for the develop-ment of alternative methods to control and prevent V. anguilla-rum infection in aquaculture.
Bacteriophages are natural and abundant biological entitiesin the marine environment, playing important roles in, e.g., struc-turing bacterial diversity and succession in the ocean and promot-ing biogeochemical element cycling, and being key drivers ofhorizontal gene transfer (6). The strong lytic potential of bacterio-phages against specific bacterial hosts has led to an increasing in-terest in the therapeutic use of lytic phages to control pathogenicbacterial infections in aquaculture. There has been a growingnumber of studies on phage therapy focusing on a variety of fishpathogens, including Aeromonas salmonicida in brook trout, Yer-sinia ruckeri in salmon, Lactococcus garvieae in yellowtail, Pseu-domonas plecoglossicida in ayu, Flavobacterium psychrophilum inrainbow trout, V. harveyi in shrimp, and V. anguillarum in salmon(7–14). These studies have demonstrated the potential of specificphages to significantly control pathogen density and, in some
cases, reduce fish mortality. Therefore, there is growing evidencethat phages can be applied to control diseases in aquaculture.
The use of bacteriophages to control specific pathogens is com-plicated by the phenotypic and genotypic complexity of bothphage and host communities, which may consist of multiple co-existing strains with different host range and susceptibility pat-terns (14, 15). In order to establish efficient phage control of adiverse host community, a large and well-characterized collectionof phages is required which, in combination, covers a broad rangeof hosts.
In this study, a collection of Vibrio sp. strains and vibriophageswas isolated from Danish aquaculture farms and characterizedalong with a collection of 20 V. anguillarum strains and one lytic V.anguillarum phage of different temporal and geographic origins.Patterns in phage and host phenotype, genotype, morphology,and phage-host ranges and interactions were studied to establish acollection of well-characterized vibriophages and to evaluate theirpotential for controlling the pathogen.
MATERIALS AND METHODSBacterial strains. The collection consisted of 20 V. anguillarum strainsprovided by Technical University of Denmark (DTU, Denmark); V.anguillarum strain PF430-3, derived from strain PF4, which was originally
Received 29 October 2013 Accepted 5 March 2014
Published ahead of print 7 March 2014
Editor: K. E. Wommack
Address correspondence to Mathias Middelboe, [email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/AEM.03544-13.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AEM.03544-13
3128 aem.asm.org Applied and Environmental Microbiology p. 3128 –3140 May 2014 Volume 80 Number 10
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

isolated in Chile (16) but was reisolated after long-term (2 weeks) cultur-ing; and 16 Vibrio sp. strains recently isolated from Danish aquaculturefarms (Table 1). The 16 recent Danish isolates were obtained from 18water samples collected from two fish farms (Venøsund and Maximus,Denmark) between May 2012 and August 2012. Aliquots of water samples(200 ml) were mixed with 200 ml 2� LB medium (12106; MO-BIO) andagitated at room temperature for 24 h. For isolation of Vibrio sp. strains,100-�l subsamples from serial dilutions were plated on thiosulfate citratebile salt sucrose agar (TCBS; 221872; Difco) incubated at 30°C for 24 h.Single colonies were then isolated and subsequently pure cultured by re-peated streaking on TCBS agar. These isolates were then further charac-terized to determine if they were V. anguillarum (see below). All of thestrains were kept at �80°C in LB medium with sterile glycerol (15%,vol/vol).
16S rRNA sequencing. Bacterial genomic DNA was extracted using acommercial genomic DNA purification kit (Clontech). The 16S rRNAgene was amplified by universal primers 27f (5=-AGAGTTTGATCCTGGCTCAG-3=) and 1492r (5=-GGTTACCTTGTTACGACTT-3=). PCR wasperformed in a 24-�l reaction mixture, which contained genomic DNAtemplate, 10� PCR buffer, 2 mM deoxynucleoside triphosphates
(dNTPs), primers (12.5 �M each), MgCl2 (25 mM), hot start polymerase,and water. PCR amplification was carried out on a thermal cycler (AppliedBiosystems) for 5 min at 95°C for denaturation, followed by 35 cycles of 45s at 95°C, 1 min at 51°C, and 2 min at 72°C and final extension at 72°C for7 min. PCRs were verified using 1% agarose gels stained with ethidiumbromide. PCR products were subsequently sequenced in both forwardand reverse directions (Beijing Genomic Institute [BGI]).
A phylogenetic analysis based on neighbor joining was performed on16S rRNA gene sequences. The original sequences were manually cor-rected, trimmed, and aligned using the BioEdit sequence alignment tool(http://www.mbio.ncsu.edu/bioedit/bioedit.html). Each of the compiledsequences were compared to the available databases using the basic localalignment search tool (BLAST) to determine phylogenetic affiliations.Mega 5.1 software (http://www.megasoftware.net/) was used to align all37 sequences. Overhanging ends were removed to allow direct compari-son of sequences with the same length (695 bp). The phylogenetic rela-tionship was bootstrapped 1,000 times, and the gaps were treated by pair-wise deletion.
Phenotypic analysis of bacterial strains. Biochemical tests were per-formed using an API 20NE system (bioMérieux) according to the instruc-
TABLE 1 The 37 Vibrio sp. strains used in the study
Numbera Strain Source BLAST result (accession no.) Homology (%)API 20NEcode
BIOLOGGN2c
16S phylogeneticgroupd
Referenceor source
1* 87-9-117 Rainbow trout, Finland Vibrio anguillarum (CP006699.1) 96 7577745 1 1 24, 492* BA35 Sockeye salmon, USA Vibrio anguillarum (CP006699.1) 95 7577745 1 1 493* 51/82/2 Rainbow trout, Germany Vibrio anguillarum (CP006699.1) 97 7577745 1 1 24, 494* T265 Salmon, UK Vibrio anguillarum (CP006699.1) 97 7577745 1 1 24, 495* 91-8-178 Turbot, Norway Vibrio anguillarum (CP006699.1) 97 7577745 1 1 24, 496* 87-9-116 Rainbow trout, Finland Vibrio anguillarum (CP006699.1) 97 7577745 1 1 24, 497* 601/90 Sea bass, Italy Vibrio anguillarum (CP006699.1) 96 7577745 1 1 This study8* 178/90 Sea bass, Italy Vibrio anguillarum (CP006699.1) 96 7577745 1 1 24, 499* 261/91 Sea bass, Italy Vibrio anguillarum (CP006699.1) 97 7577645 1 1 24, 4910b PF430-3 Salmon, Chile Vibrio anguillarum (KF150778.1) 96 7477745 1 1 17
11* A023 Turbot, Spain Vibrio anguillarum (CP006699.1) 95 7577345 1 1 4912* 4299 Unknown Vibrio anguillarum (KF150778.1) 96 5576744 1 1 5013* LMG 12010 Unknown Vibrio anguillarum (CP006699.1) 95 7577344 1 1 4914* 850610–1/6a Rainbow trout, Denmark Vibrio anguillarum (CP006699.1) 97 7577745 1 1 4915* 90-11-287 Water sample, Denmark Vibrio anguillarum (CP006699.1) 96 7577745 1 1 24, 5116* 91-7-154 Trout, Denmark Vibrio anguillarum (CP006699.1) 97 7577745 1 1 4917* S2 2/9 Rainbow trout, Denmark Vibrio anguillarum (FJ824662.1) 98 7577745 1 1 4918* 90-11-286 Rainbow trout, Denmark Vibrio anguillarum (KF150778.1) 97 7576745 1 1 4919* HWU53 Rainbow trout, Denmark Vibrio anguillarum (CP006699.1) 97 7576745 1 1 4920* 6018/1 Rainbow trout, Denmark Vibrio anguillarum (CP006699.1) 97 7576745 1 1 49
21* 9014/8 Rainbow trout, Denmark Vibrio anguillarum (CP006699.1) 96 7577745 1 1 4922 VA1 Salmon, Denmark Vibrio anguillarum (KF150778.1) 96 7577745 1 1 This study23 VA2 Salmon, Denmark Vibrio anguillarum (KF150778.1) 95 7577745 1 1 This study24 VA3 Salmon, Denmark Vibrio anguillarum (KF150778.1) 96 7576745 1 1 This study25 VSP1 Salmon, Denmark Vibrio splendidus (KC884584.1) 96 7430044 2 2 This study26 VSP2 Salmon, Denmark Vibrio cyclitrophicus (KC185413.1) 97 7520044 2 2 This study27 VSP3 Salmon, Denmark Vibrio cyclitrophicus (KF488567.1) 97 7430044 2 2 This study28 VSP8 Denmark Vibrio cyclitrophicus (KF488567.1) 95 7430044 2 2 This study29 VSP10 Salmon, Denmark Vibrio cyclitrophicus (KC185413.1) 97 7430044 2 2 This study30 VSP12 Denmark Vibrio cyclitrophicus (KF488567.1) 96 7430044 2 2 This study
31 VSP4 Sole, Denmark Vibrio shilonii (JX999944.1) 95 7420044 2 3 This study32 VSP5 Sole, Denmark Vibrio sp. (EU517619.1) 97 7476744 3 3 This study33 VSP6 Denmark Vibrio sp. (FM957468.1) 96 7476744 3 3 This study34 VSP7 Denmark Vibrio sp. (FM957468.1) 97 7476744 3 3 This study35 VSP9 Denmark Vibrio sp. (KC178920.1) 97 7476744 3 3 This study36 VSP11 Denmark Vibrio sp. (KC178920.1) 97 7476744 3 3 This study37 VSP13 Denmark Vibrio sp. (FM957471.1) 97 7476744 3 3 This study
a Strains marked by an asterisk were provided by Technical University of Denmark (DTU).b The strain PF430-3 was provided by the Hellenic Centre for Marine Research.c Group numbers according to the BIOLOG GN2 dendrogram.d Group numbers according to 16S rRNA phylogenetic tree (see Fig. S1 in the supplemental material).
Isolation and Characterization of Vibriophages
May 2014 Volume 80 Number 10 aem.asm.org 3129
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

tions of the manufacturer. The inoculum was distributed into test stripswhich were then incubated at 30°C for 48 h. Biochemical reactions wereread based on color development, translated into numerical codes, andinterpreted with the API 20NE database.
The sensitivity of the strains to the vibriostatic compound agent 0129(2,4-diamino-6,7-diisopropylpteridine phosphate) (Oxoid) was deter-mined by the disk diffusion method (17) at both 10 and 150 �g per diskwith incubation for 24 h at 30°C.
Biolog GN2 MicroPlates (Biolog) containing 95 different carbonsources were used to test the ability of the strains to use different carbonsources. Briefly, pure cultures of bacterial strains were grown on LB agarplates for 24 h at 30°C and subsequently transferred to 0.9% saline buffer(9 g NaCl in 1 liter distilled water) and washed twice to remove medium(8,000 � g for 10 min). From this bacterial suspension (ca. 108 CFUml�1), 100-�l aliquots were inoculated into each well in duplicate platesand incubated in the dark at 30°C for 48 h. Data were analyzed based onvisual inspection of tetrazolium violet development as an indicator ofmetabolic activity.
Bacteriophage isolation. The water samples for bacterial isolationwere also used for propagation of phages for subsequent isolation andpurification of new vibriophages. Four different protocols were used (18).(i) A 100-ml water sample was sterile filtered (0.22 �m; Millipore) andmixed with 100 ml 2� LB medium containing a mixture of 5 randomVibrio strains (Table 1) to stimulate proliferation of phages specific to theadded strains. After 24 h of incubation at room temperature, bacteria inthe lysate were killed by chloroform (20 �l ml�1), followed by centrifu-gation (10,000 � g, 10 min) to remove bacterial debris. (ii) One-hun-dred-ml unfiltered water samples were mixed with 100 ml 2� LB brothand incubated at room temperature for 24 h to stimulate proliferation ofphages specific to cooccurring Vibrio species host strains in the watersample. Aliquots of enrichment samples (50 ml) were then treated withchloroform and centrifuged as described above, and the supernatant wasmixed with individual Vibrio strains (Table 1) in a second round of en-richment as described above. (iii) Five-ml water samples were filtered(0.22 �m pore size), and the filtrate was diluted and used directly forplaque assay (19) without previous enrichment. (iv) Two-hundred-mlwater samples were filtered (0.22 �m), and phages were then concentratedusing 30-kDa centrifugation filters (Amicon ultra 15 protein; Millipore)and stored at 4°C until further use.
The presence of Vibrio-specific phages in the water samples describedabove was tested against all 37 Vibrio sp. strains by spotting 5-�l sampleson lawns of individual strains using the double-layer agar method (18). Inthe case of cell lysis in the spotted area, phages were scraped off the plateand transferred to SM buffer (50 mM Tris-Cl, pH 7.5, 99 mM NaCl, 8 mMMgSO4, 0.01% gelatin). These phage concentrates were then diluted, andthe presence of phages was verified by plaque assay (19). For purificationof individual phages, single plaques were isolated with a sterile pipet andtransferred to SM buffer. This procedure was repeated at least three timesto ensure purification of individual phages.
To obtain high-titer phage stocks, 5 ml SM buffer was added to plateswith confluent lysis, and the top agar was shredded with an inoculationloop. The mixture was transferred to a sterile tube and incubated on ashaker for at least 2 h at room temperature to release phage particles andthen centrifuged (10,000 � g, 10 min, 4°C). The supernatant was filteredthrough a 0.22-�m membrane (Millipore), and the phage stock wasstored at 4°C in the dark. In addition to the phage isolates obtained fromDanish aquaculture farms, a broad-host-range vibriophage (KVP40) (20)was kindly provided by the Hellenic Centre for Marine Research.
Phage host range and EOP. The host range of each phage isolate wasdetermined using a spot assay against all of the available Vibrio sp. strains(Table 1). Aliquots of 5 �l phage stock (phage titer between 106 and 109
PFU ml�1) were spotted on lawns of the potential host strain prepared as4 ml top agar inoculated with 500 �l bacterial cultures in mid-log phase (ca.108 CFU ml�1). After 24 h of incubation at 30°C, the spots were assessed bythe clarity of plaques and scored as transparent, turbid, or no inhibition. To
provide a quantitative measure of the phage lytic potential, the efficiency ofplating (EOP) for each phage on each phage-sensitive strain was determinedby measurement of the number of PFU produced on individual hosts from agiven phage stock, as quantified by plaque assay (19).
Purification of bacteriophage genome DNA. Aliquots (500 �l) ofhigh-titer (ca. 109 PFU ml�1) bacteriophage stocks were further concen-trated by centrifugation (1,000 � g, 25 min) in 30-kDa ultrafiltrationAmicon Microcon spin devices (Millipore), and then the membrane waswashed three times by 50 �l 100� Tris-EDTA buffer (TE buffer; 10 mmolTris-Cl, 1 mmol EDTA, pH 8) (1,000 � g for 18 min each). After thewashing, the phages were eluted with 20 �l 100� TE buffer by centrifu-gation of the inverted membrane into a new tube (300 � g for 5 min). Todenature phage proteins and release the phage DNA, the concentrated phageswere heated to 70°C for 10 min and then cooled on ice for subsequent deter-mination of phage genome size. To confirm the concatemer phenomena of anumber of phages (phage �H2, �H8, and �H20), additional extraction ofDNA from these phages was performed using a QIAprep spin M13 kit(Qiagen) according to the manufacturer’s instructions.
Phage genome size. The bacteriophage genome sizes were determinedby pulsed-field gel electrophoresis (PFGE) using the CHEF-DRIII pulsed-field electrophoresis system (Bio-Rad) (14). Phage DNA was run using 1%agarose gels in 0.5� Tris-borate-EDTA (TBE) buffer (10� TBE, 890 mMTris, 20 mM EDTA, 890 mM boric acid, pH 8.3) with two different in-strument settings for small and large genome sizes. For the large genomesizes (�50 kb), switch times were ramped from 50 to 90 s for 24 h and avoltage of 6 V/cm, whereas switch times ranged from 0.5 to 5 s and a totalrun time of 15 h for small genomes (�50 kb). An 8- to 48-kb molecularsize DNA marker and a Lambda DNA ladder (Bio-Rad) were used asstandards. The gels were stained with 0.01% (vol/vol) SYBR green I (In-vitrogen) in 0.5� TBE for 1 h in the dark and then washed with distilledwater for 30 min prior to visualization of the bands using the ChemiDocMP imaging system (Bio-Rad).
Bacteriophage morphology by TEM. For transmission electron mi-croscopy (TEM) analysis of phage isolates, the phage stocks were furtherconcentrated by ultracentrifugation (100,000 � g, 90 min, 20°C; 70.1Ti;Beckman), followed by resuspension of the pellet in 100 �l SM buffer.Formvar-carbon-coated copper grids were placed on top of the phageconcentrates, allowing the phages to adsorb for 20 min. The grids werethen stained with 10 �l 2% sodium phosphotungstate (pH 7.4; 0.02-�m-pore-size filters) for 2 min. Excess stain was removed by touching the edgeof the grids with filter paper, and the grids were washed with several dropsof distilled water and allowed to dry on the filter paper for 15 min. Thegrids were observed with a JEM-2100 transmission electron microscope(JEOL) operated at 80 kV.
One-step growth curve. To characterize phage life cycle characteris-tics, one-step growth experiments were performed for two of the phageisolates using marine broth (MB; 2216; Difco) at a low multiplicity ofinfection (MOI; i.e., the initial ratio of phage particles to host cells), fol-lowing a protocol modified from that of Ellis and Delbrück (21). Forphage �H20, 1 ml of exponentially growing bacteria (108 CFU ml�1) wasmixed with 100 �l lytic phage stock (108 PFU ml�1; MOI of 0.1) andincubated at room temperature for 20 min to allow the phages to adsorbto the host according to procedures described by Hidaka and Tokushige(22). Bacteria were then pelleted (7,000 � g, 10 min), and the nonad-sorbed phages in the supernatant were discarded. The pellet was thenresuspended in 1 ml MB and diluted to an appropriate phage titer (104
PFU ml�1). Two 100-�l subsamples of culture were collected at regularintervals for 60 min for enumeration. In one subsample, free phages wereimmediately quantified by plaque assay (18). The other subsample waspretreated with 10 �l chloroform to measure the release of intracellularphages and then centrifuged at 10,000 � g for 1 min prior to phage enu-meration. For phage KVP40, the same procedure was applied, except thatfor this phage, the stock was diluted to 107 PFU ml�1, reducing the MOIto 0.01.
Tan et al.
3130 aem.asm.org Applied and Environmental Microbiology
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

Susceptibility of V. anguillarum to phage infection at differentMOIs. The lytic potentials of two selected phages, �H20 and KVP40,against V. anguillarum were examined at MOIs of 0.1, 1, and 10 in liquidMB cultures during infection of V. anguillarum BA35 and PF430-3, re-spectively. Serial dilutions of the phage stocks (ca. 107 to 109 PFU ml�1)were inoculated in duplicate 50-ml mid-log-phase cultures of host bacte-ria in MB at a density of �107 CFU ml�1. For each experiment, duplicatecontrol cultures without phage addition were also established. The effectsof phage lysis on host cell density were monitored by regular measure-ments of the optical density at 600 nm (OD600) during the 24-h incuba-tions. Phage abundance was quantified in one of the experiments (MOI,0.1) from the number of PFU obtained on lawns of host strains by plaqueassay (19) periodically over 24 h. At various time points, individual colo-nies were isolated from the cultures on TCBS plates and subsequentlypurified through 2 to 3 rounds of reisolation of single colonies to deter-mine changes in phage susceptibility in the host community during incu-bation.
Statistical analysis. For all of the dendrograms, UPGMA (un-weighted-pair group method with arithmetic mean) construction exper-iments and Dice coefficients were calculated between pairs of each strainor phage and grouped by UPGMA and 100 bootstrap replicates using thedendrogram construction utility DendroUPGMA (Biochemistry andBiotechnology Department, Rovira i Virgili University, Tarragona, Spain)(http://genomes.urv.cat/UPGMA/index.php).
RESULTSIsolation of Vibrio strains. A total of 16 Vibrio sp. strains isolatedfrom Danish aquaculture were susceptible to phage KVP40; there-fore, these 16 strains were tentatively classified as closely relatedVibrio sp. strains. Of the total of 37 strains examined, 35 weresensitive to the vibriostatic agent O129 at both 10 and 150 �g,whereas two isolates (VSP8 and VSP9) were resistant to O129 atconcentrations of up to 150 �g. The 37 Vibrio sp. strains used inthis study covered a considerable spatial variability, with isolatesoriginating from the Mediterranean Sea (Spain and Italy), the Bal-tic Sea (Finland and Germany), the North Sea (Norway andUnited Kingdom), the Atlantic Ocean (United States), the southPacific Ocean (Chile), and inner Danish waters; the range in timeof isolation covered �20 years (23).
Identification of Vibrio strains. Based on 16S rRNA gene sim-ilarity, 24 of the isolates were presumptively identified as V.anguillarum according to NCBI BLAST analysis (NCBI BLAST;http://www.ncbi.nlm.nih.gov/BLAST/) (Table 1), including theentire DTU collection, the Chilean strain (PF430-3), and 3 Danishwater isolates (VA1, VA2, and VA3), whereas the remaining 13 Dan-ish isolates yielded identification as V. cyclitrophicus, V. slendidus, V.shilonii, and Vibrio spp. (Table 1). A phylogenetic tree based on the16S rRNA gene sequence analysis showed three main groups:group 1 (V. anguillarum), containing 20 DTU strains, strainPF430-3, and three Danish V. anguillarum isolates (VA1, VA2,and VA3); group 2 (V. splendidus and V. cyclitrophicus), contain-ing six Danish water isolates (VSP1, VSP2, VSP3, VSP8, VSP10,and VSP12); and group 3 (V. shilonii and Vibrio spp.), containingthe remaining strains (VSP4, VSP5, VSP6, VSP7, VSP9, VSP11,and VSP13) (see Fig. S1 in the supplemental material).
API 20NE fingerprinting. Eleven different identities (Table 1)were generated from the 37 strains using API 20NE test. For all ofthe strains, positive results were obtained for potassium nitrate,D-glucose, esculin ferric citrate, 4-nitrophenyl -D-galactopyra-noside, malic acid, and oxidase reactions, and negative reactionswere found for urea, capric acid, adipic acid, and phenylacetic
acid, whereas the remaining 11 biochemical tests gave various re-sults (data not shown).
Biolog fingerprint. The phenotypic diversity of and relation-ship between the 37 strains were explored by studying the differ-ences in carbon resource utilization, which showed a clustering ofthe strains into three primary phenotypic groups (Fig. 1). Cluster1 contained 24 strains, including all of the strains in the DTUcollection, the PF430-3 strain, and three of the Danish isolates(VA1, VA2, and VA3), whereas the remaining 13 Danish isolateswere grouped into clusters 2 and 3 (Fig. 1). Overall, the number ofsubstrates used varied from 54 (strain BA35) to 22 (strain VSP10)of the 95 tested substrates, with the highest number of availablesubstrates found in cluster 1 (33 to 54) and the lowest in cluster 2(22 to 38). All of the 22 substrates used by strain VSP10 could alsobe used by all other strains, suggesting that the utilization of these22 substrates was a fundamental property of Vibrio strains.
Bacteriophage enrichment and isolation. Ten different bac-teriophage isolates (Table 2) were obtained from Danish watersusing different enrichment protocols. Phages �H20, �H8,�S4-7, and �S4-18 were isolated by following protocol I, using amixture of Vibrio sp. strains as hosts for enrichment, whereasphages �H1, �H2, �H4, �H5, �H7, and �2E-1 were isolatedusing a single Vibrio sp. strain for enrichment (protocol II).
Phage genome sizes, as determined by PFGE, showed largedifferences among the 11 phages and covered a 20-fold range,from 11 kb to 244 kb (Table 2). An interesting phenomenon wasthe systematic observation of several distinct bands appearing onthe PFGE gels for phage �H20, �H8, and �H2 despite up to 5rounds of phage purification from a single plaque. For example,PFGE analysis of phage �H20 revealed seven bands, ranging from50 to 291 kb, with each band size increasing by 50 kb, and phage�H8, for which two bands of 50 kb and 100 kb, respectively, wereobserved (Fig. 2; also see Fig. S2 in the supplemental material).
Host range and EOP. The host range and lytic potential of the11 vibriophages were tested against 24 V. anguillarum and 13Vibrio sp. strains by spotting 5 �l of a serially diluted phage stockonto top agar containing the potential host strain. Together, thecollection of phages was able to infect all 37 strains; however, therewere large differences in host range pattern and lytic efficiencyamong the phages (Fig. 3). Six phages (�H2, �H8, �H20, �S4-7,�S4-18, and �2E-1) exhibited a narrow host range, infecting onlythe host used for its isolation and perhaps one or two additionalstrains. Phages �H1, �H7, �H4, �H5, and KVP40, on the otherhand, were characterized by broader host ranges covering bothlocal host isolates and isolates obtained from geographically dis-tant locations. Conversion of the phage host range pattern into adendrogram using the Dice coefficients comparing algorithmshowed some distinct clusters of the phages and bacterial strains(Fig. 3). Generally, phages with the same genome size had similarhost ranges, for example, the �H1 and �H7 pair and the �H4 and�H5 pair, which had pairwise identical genome sizes and alsoclustered together according to host range (Fig. 3). Interestingly,the phages with the largest genomes, i.e., �H1, �H7 (194 kb), andKVP40 (244 kb), also had the largest host range, infecting many ofthe same hosts, despite the fact that they were isolated from Dan-ish (�H1 and �H7) and Japanese (KVP40) locations (24). Phagesusceptibility pattern, on the other hand, did not show obviousconnection to the other genotypic or phenotypic relationships.
Isolation and Characterization of Vibriophages
May 2014 Volume 80 Number 10 aem.asm.org 3131
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

Phage morphology. Phage morphology was examined byTEM for all 11 phages (Table 2 and Fig. 4). According to theInternational Committee on Taxonomy of Viruses (ICTV), threemorphological groups of phages were represented by the 11 phag-
es: Siphoviridae (�H8 and �H20), with flexible tails, Podoviridae(�H2, �2E-1, and �S4-18), with short tails, and Myoviridae(�H1, �H4, �H5, �H7, �S4-7, and KVP40), possessing an ico-sahedral head and contractile tails with tail fibers and relatively
FIG 1 Phenotype dendrogram derived by UPGMA cluster analysis of all 37 strains in the Biolog GN2 profiles. The data were scored and compared by using Dicecoefficient algorithm. The scale bar indicates the dissimilarity.
Tan et al.
3132 aem.asm.org Applied and Environmental Microbiology
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

large genomes (25) (Fig. 4 and Table 2). Interestingly, somevibriophages with different genome sizes, e.g., �H2 and �S4-18,belong to the same family.
Phage-host interactions. Life cycle characteristics were deter-mined for phages �H20 and KVP40 from one-step growth curvesduring incubation with their host strains BA35 and PF430-3, re-spectively. Despite different morphologies and genome sizes,phages �H20 and KVP40 had quite similar burst sizes (70 and 80PFU per cell, respectively) and identical eclipse (20 min) and la-tent periods (25 min).
The parallel long-term (24 h) infection experiments withphages �H20 and KVP40 and the hosts BA35 and PF430-3, re-spectively, at different MOIs also showed similar lytic potential of
the two phages, with a strong dependence on the initial MOI forphage control of the host population (Fig. 5). In batch cultures atan MOI of 0.01, both phage KVP40 and �H20 showed rapidpropagation during the first 10 h of incubation, resulting in phagetiters stabilizing around 1010 and 109 PFU ml�1, respectively (Fig.5). The host growth rates in the control cultures were 0.241 0.002 h�1 and 0.231 0.007 h�1 for PF430-3 and BA35, respec-tively, and both strains reached stationary phase after 10 h. In thephage-amended cultures, phages were able to control growth ofthe host for 5 to 6 h followed by regrowth of the host population.
Accordingly, decreasing the initial MOI resulted in a delayedand less pronounced decrease in the OD600 (Fig. 6). Regrowthoccurred in all cultures, and after 20 h the OD600 stabilized at 0.6 to0.7, corresponding to 65 to 75% of the final OD600 in the controlcultures. In addition to the OD600 measurements, bacterial iso-lates were obtained during the incubations for further analysis ofchanges in phage susceptibility in the host population (Fig. 6). Inthe BA35-plus-�H20 cultures, the occurrence of the bacterio-phage-resistant isolates varied considerably over time and be-tween treatments. According to the spot test results, the emer-gence of the bacteriophage-resistant bacteria was faster at highthan at low MOI. Within the first hour after incubation, resistantstrains were isolated at an MOI of 10, whereas resistant pheno-types were not detected at MOIs of 1 and 0.1. At high MOI, onlyresistant isolates were obtained after 7 h of incubation. At thelower MOIs, resistant isolates dominated during the regrowthphase (7 h). However, partially sensitive bacteria (i.e., reducedsensitivity compared to the sensitive wild-type strain) were iso-lated from 8 h onwards and constituted 40 to 50% of the isolatesafter 24 h. All of the resistant and partially sensitive strains isolatedfrom the phage-amended BA35 culture after 24 h formed smallercolonies with lower growth rates on TCBS plates than those seenin the control cultures without phages. For the partially sensitiveand resistant BA35-derived isolates, a second round of purifica-tion and subsequent susceptibility test were carried out. Interest-ingly, all of the partially sensitive strains returned to their sensitivephenotype after purification in the absence of phages, whereas allof the completely resistant isolates remained resistant.
In the parallel experiment with V. anguillarum strain PF430-3and phage KVP40, no completely resistant isolates were obtainedfrom any of the treatments during the incubation, and the colony
TABLE 2 Method of isolation, morphological characteristics, and genome size of vibriophages isolated from Danish aquaculture farms and phageKVP40 (Japan)
Family Phage Isolation protocolaGenomesize (kb) Host
Head diam(nm)
Head length(nm)
Tail diam(nm)
Tail length(nm)
Referenceor source
Myoviridae KVP40 Unknown 242 PF430-3 76 120 30 120 32Myoviridae �H1 II 194 4299 107 129 19 233 This studyMyoviridae �H7 II 194 90-11-286 117 129 22 226 This studyMyoviridae �S4-7 I 145 VSP10 81 80 26 58 This studyMyoviridae �H4 II 95 90-11-287 72 80 22 229 This studyMyoviridae �H5 II 95 51/82/2 63 77 24 222 This studySiphoviridae �H8 I 50 T265 114 127 19 237 This studySiphoviridae �H20 I 50 BA35 54 65 12 144 This studyPodoviridae �S4-18 I 45 VA1 63 64 12 13 This studyPodoviridae �2E-1 II 42 VSP8 53 46 8 10 This studyPodoviridae �H2 II 11 A023 89 82 15 32 This studya I, phage isolation method using a single enrichment step with a mixture of potential host strains; II, phage isolation method using two steps of enrichments with a single strain.For further information, see Materials and Methods.
FIG 2 PFGE analysis of selected phage genomes with concatemers (��2,��8, and ��20), with switch times ranging from 0.5 to 5 s and a total runtime of 15 h.
Isolation and Characterization of Vibriophages
May 2014 Volume 80 Number 10 aem.asm.org 3133
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

morphology did not change over time. Instead, all of the isolateswere sensitive or partially sensitive strains, and after 8 h all ob-tained isolates were fully sensitive to KVP40. In contrast to thecontrol cultures, visible biofilm was formed in the KVP40-amended cultures, and some isolates formed aggregates whensubsequently grown in liquid culture. For the aggregating andpartially sensitive PF430-3-derived isolates, a second and thirdround of purification and subsequent spot assay showed that allstrains returned to the original phenotype and were fully sensitiveto phage KVP40.
DISCUSSION
Successful application of phage therapy in the treatment of vibri-osis requires detailed knowledge of the diversity and distributionof the phage susceptibility properties of Vibrio sp. pathogens as-sociated with vibriosis as well as a collection of well-characterizedphages that covers host diversity. In this study, we characterized 11
vibriophages and 37 Vibrio sp. strains (Table 1 and 2), coveringconsiderable temporal and spatial variability. Further, two V. an-guillarum strains, BA35 and PF430-3, and the specific phages�H20 and KVP40 were selected for more detailed studies of phagelytic potential and phage-host interactions.
Diversity of V. anguillarum strains. The V. anguillarumstrains examined in the present study (Table 1) showed close sim-ilarity according to 16S rRNA gene sequences irrespective of timeor place of isolation. Thirty-seven of the isolates which groupedtogether covered a period of isolations of �20 years and a widerange of geographical locations (Chile, Spain, Italy, Germany,United Kingdom, Norway, Finland, Denmark, and the UnitedStates) and fish species (turbot, trout, sea bass, and salmon). Thissuggests that the 16S rRNA gene phylogeny of V. anguillarum ishighly uniform and stable in time and space. Thus, the method isunable to discriminate between potentially pathogenic and non-pathogenic V. anguillarum strains or resolve differences in V.
FIG 3 Host ranges of 11 phages (columns) against 37 Vibrio sp. strains (rows). Black boxes indicate clear plaques, gray boxes indicate turbid plaques, and whiteboxes indicate no infection. Numbers express the efficiency of plating, i.e., the fraction of infectivity of the phage stock on a given host compared to the infectivityon the original host of isolation (isolation efficiency of the host was 100%). Dendrograms were derived by UPGMA cluster analysis. The data were scored andcompared by using the Dice coefficient algorithm. The scale bar indicates the dissimilarity.
Tan et al.
3134 aem.asm.org Applied and Environmental Microbiology
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

anguillarum communities across spatial and temporal scales. Sim-ilarly, genetic fingerprinting methods (enterobacterial repetitive-element intergenic consensus sequence [ERIC] PCR) showed thatVibrio sp. isolates from coastal British Columbia did not groupaccording to geography, suggesting genetic homogeneity amongthe environmental strains (26). Previous studies have, however,revealed a large degree of microheterogeneity in 16S rRNA geneswithin Vibrio sp. strains (27), limiting the use of 16S rRNA genesfor phylogenetic studies in Vibrionaceae. Higher taxonomic reso-lution has been obtained using multilocus sequence analyses(MLSA) of six protein-coding genes (28); however, this methodalso focuses on variations in housekeeping genes and is applicablemainly for identification of Vibrio strains at the species level.
The V. anguillarum isolates clustered in distinct groups basedon the Biolog GN2 profiles and differed from the other identifiedclusters of Vibrio strains. In contrast, the phage susceptibility pat-tern of the V. anguillarum isolates did not show any connection tothe species or physiological relationships obtained from Biologprofiles. In fact, phage susceptibility patterns did not even groupaccording to the species clusters defined for the 37 strains. Thissuggests that phenotypic traits related to sensitivity to phages arenot resolved by the methods used in this study. This supportsprevious studies of Cellulophaga baltica (29), V. parahaemolyticus(15), F. psychrophilum (14), and Escherichia coli (30), which haveall demonstrated a high degree of phenotypic diversity in hostsusceptibility at the strain level that was uncoupled from the over-all genetic relationships among the host isolates. Thus, our resultsconfirm highly variable patterns in phage sensitivity even withinrelatively narrow phylogenetic and phenotypic groups of bacteria,and at the same time they demonstrate similar susceptibility pat-terns across a broad phylogenetic scale in Vibrio spp., emphasizingthe complexity of phage-host interactions in this group.
As observed for the physiological fingerprints, there was over-lap in the phage susceptibility patterns for V. anguillarum strainsisolated across the large temporal and spatial scales and from dif-ferent fish species represented in the current study. For example,the strains 90-11-286 and VA3, which were isolated from Danishrainbow trout and salmon farms, respectively, with 20 years be-tween isolations, were both infected by the same 3 phages (�H1,�H7, and KVP40). Similarly, strains VIB88 and BA35, isolated
from rainbow trout farms and salmon farms in Germany and theUnited States, respectively, both were infected by phages �H4 and�H5, isolated from a Danish turbot farm 20 years later. This dem-onstrates that despite the occurrence of unique susceptibility pat-terns of individual isolates, key phenotypic properties related tophage susceptibility are distributed worldwide and maintained inthe global V. anguillarum community for decades.
Phage isolation and diversity. The 10 vibriophages isolatedfrom Danish fish farms all had unique host ranges when testedagainst the collection of 37 Vibrio strains. All three major morpho-logical families, Myoviridae, Podoviridae, and Siphoviridae, wererepresented, and the phages ranged in genome size from 11 to 194kb, distributed in 6 distinct groups (Table 2). The phage isolatesshowed large differences in host specificity, and together the Dan-ish phages were able to infect and lyse 30 of the 37 strains, coveringmost of the phylogenetic, geographical, and temporal variationthat characterized the host isolates. This is interesting, as it sug-gests that vibriophage infective properties are not restricted tolocal cooccurring host communities but rather widely distributedacross large spatial and temporal scales, as also suggested by pre-vious observations of an ocean-scale distribution of geneticallyrelated vibriophages (31).
The two broad-host-range phages isolated in this study (�H1and �H7) both belonged to the Myoviridae family and had rela-tively large genomes (194 kb). This is consistent with the hostrange pattern observed for F. psychrophilum phages, where thelarge-genome phages (�90 kb) had substantially broader hostranges than small-genome phages (�12 kb) (29). In accordancewith this, the previously isolated large-genome vibriophagesKVP40 (244 kb) (32) and �pp2 (246 kb) (33) also have very broadhost ranges covering several species of Vibrio. KVP40 is known tocause infection through the common outer membrane proteinOmpK and has previously been shown to infect eight Vibrio spe-cies, including V. anguillarum, V. parahaemolyticus, and V. chol-erae (20). The presence of several copies of genes encoding pro-teins associated with phage tail or tail fibers in the KVP40 genomesuggested an increased flexibility in host range adaptation, in-creasing host range (32). Despite the fact that KVP40 was origi-nally isolated in Japan �20 years ago (24), it still infected approx-imately 40% of the Vibrio sp. isolates obtained from Danish
FIG 4 TEM pictures of selected phages. Phages were stained with 2% sodium phosphotungstate. Scale bar, 200 nm.
Isolation and Characterization of Vibriophages
May 2014 Volume 80 Number 10 aem.asm.org 3135
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

aquaculture, confirming its cross-species host range, and it main-tained infective properties across large temporal and spatial scales.Thus, the characteristic of the new phage isolates �H1 and �H7adds to the emerging view of a global distribution of large-ge-nome, broad-host-range vibriophages.
Generally, vibriophages were widespread in Danish aquacul-ture, as phages were isolated from all of the investigated farmsindependently of outbreaks of vibriosis. However, the fact thatphages could not be isolated without previous propagation in en-richment cultures indicated that densities of specific vibriophages
FIG 5 (A) Optical density (OD600) in cultures of V. anguillarum BA35 amended with phage �H20 at MOIs of 0 (control), 0.1, 1, and 10 and a correspondingabundance of PFU ml�1 in the treatment at an MOI of 0.1. (B) Optical density (OD600) in cultures of V. anguillarum PF430-3 amended with phage KVP40 atMOIs of 0 (control), 0.1, 1, and 10 and the corresponding PFU abundance in the treatment at an MOI of 0.1. Error bars represent standard deviations from allexperiments carried out in duplicate.
Tan et al.
3136 aem.asm.org Applied and Environmental Microbiology
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

FIG 6 Phage susceptibility properties of V. anguillarum isolates obtained from cultures enriched with phages at different MOIs. (A to C) V. anguillarum strainBA35 plus phage �H20 at an MOI of 10 (A), 1 (B), or 0.1 (C). (D to F) V. anguillarum strain PF430-3 plus phage KVP40 at an MOI of 10 (D), 1 (E), or 0.1 (F).“Sensitive” indicates that isolates were fully sensitive to the phage. “Resistant” indicates that isolates were not susceptible to phage �H20 infection in the spot test.“Partially resistant” indicates that isolates had a reduced susceptibility to the phage. “Aggregates” indicates that isolates were forming aggregates, which protectedagainst phage infection.
Isolation and Characterization of Vibriophages
May 2014 Volume 80 Number 10 aem.asm.org 3137
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

were relatively low in the fish farms. Some samples from which Vibriosp. hosts were isolated did not harbor their corresponding phages,suggesting that specific hosts and lytic phages did not always co-exist in the same environment. However, it should be noted thatthe filtration of the water samples and chloroform treatment priorto phage isolation may have resulted in the loss of phages (18, 34).
In all isolate-based studies of phage diversity, the results arebiased by the choice of host strains used for isolation, as eachindividual strain will only target a subset of the infective phagespresent in the sample. In the current study, broad-host-rangephages �H1 and �H7 were isolated using strains from the globalVibrio species collection, whereas the more host-specific phages,such as �2E-1, �S4-18, and �S4-7, were isolated using a cooc-curring Vibrio sp. host obtained from the same sample. As theevolution of a broad host range in phages is believed to be accom-panied by a fitness cost, i.e., a reduced infective efficiency in itsoriginal hosts (35), it is likely that the observed differences inphage isolation pattern reflect different phage properties. There-fore, we speculate that highly specific phages are efficient, oppor-tunistic phages which propagate rapidly in response to the growthof their specific hosts and will dominate in samples where theirhost is present. Consequently, these phages are the most likely tobe isolated using cooccurring hosts as target strains. Broad-host-range phages, on the other hand, may follow a K-strategy (36),maintaining lower densities but for longer periods; therefore, theymay be more likely to be pulled out from a sample using non-cooccurring or rare hosts. Due to these differences in isolationpattern, we do not know to what extent the isolated phages arerepresentative of the local phage community.
PFGE provides a good discrimination for phage genome sizesand clearly demonstrates a large variability in genome size withinthe group of vibriophages. Several phage genomes, including�H20, �H8, and �H2, yielded several bands in PFGE analysis,representing multiplications of its own genome size. These bandsmay represent concatemers of the phage genome produced duringDNA replication, as previously demonstrated in phage (37).Also, concatemer formation of phage DNA has been observed as aresult of DNA cleavage during heating and subsequent reassemblyat ambient temperature (38). The possibility of coinfection ofother phages still cannot be ruled out; however, we did not seedifferent phage morphotypes during TEM analysis of single iso-lates. A previous report of different phage morphotypes associatedwith multiple bands on a PFGE gel, however, suggested that coin-fection of more than one phage may be an explanation for theoccurrence of several genome sizes on a PFGE gel of a purifiedphage (29). Further genomic analyses are required to elucidate theorigin of the multiple bands obtained in the PFGE analyses.
Phage-host interactions. The in vitro bacterial challenge assaywith V. anguillarum strains BA35 and PF430-3 demonstrated sig-nificant but temporary phage control of both strains which wasstrongly dependent on the initial MOI. Even at the highest MOI,the host bacteria were not completely eliminated and regrowth ofcells following phage infection suggested the emergence of phage-resistant strains after 5 to 6 h of incubation. Lytic bacteriophageshave previously been shown to select for the emergence of resis-tant strains in culture studies with marine heterotrophic bacteria(39–41), where phage-resistant strains rapidly replaced the sensi-tive wild type following exposure to phages.
Isolation and characterization of bacterial isolates during theexperiments confirmed that completely resistant strains, and
strains with reduced sensitivity, dominated the bacterial popula-tion after 7 h in the culture containing strain BA35 and phage�H20. Interestingly, however, phage-resistant strains were notisolated after exposure of strain PF430-3 to phage KVP40, as hasalso been observed in other phage infection studies with hostssuch as Salmonella enterica (42), S. enterica serovar Oranienburg(43), and E. coli O157:H7 (44). In the current experiment withstrain PF430-3, phage exposure led to the formation of bacterialaggregates, and all of the isolated colonies had maintained fullsensitivity to the phage after purification. These results suggestthat different mechanisms were responsible for the regrowth ofcells in the presence of phages following lysis of the initial sensitivepopulation. The BA35 and phage �H20 interaction results in asuccession toward phage-resistant strains, whereas the strainPF430-3 apparently is protected against phage KVP40 infection byaggregation. The underlying mechanism providing bacterial pro-tection against phages via aggregation, allowing the coexistence oflytic phages and sensitive strains, is not revealed in the presentstudy. However, previous studies have shown that phage-inducedlysis can promote the formation of bacterial biofilms (45), whichmay offer protection against phage infection via the exopolysac-charide (EPS) matrix covering the biofilm (46) or the presence ofbacterial refuges in the spatially heterogeneous environment (47).More recently, Høyland-Kroghsbo et al. (48) found that N-acyl-L-homoserine lactone (AHL) quorum-sensing signals in E. colicaused a transient reduction in the phage adsorption rate andsuggested this as a mechanism allowing coexistence of lytic phagesand sensitive hosts. Such mechanisms may also have contributedto the observed coexistence of sensitive strains of PF430-3 and thelytic phage KVP40 in the current study.
Potential for phage therapy in aquaculture. One of the mainchallenges in using bacteriophages to control pathogens in aqua-culture is to cover the diversity and temporal and spatial variationsin pathogen composition, as well as the phage resistance that mayrapidly develop. A detailed characterization of phage propertiesand understanding of phage-host interactions are essential re-quirements for the successful application of phage-based patho-gen control. The local occurrence of both specific and broad-host-range phages which, in combination, were able to infect all of thetested host strains, covering large geographical and temporalscales, suggested that vibriophage-host systems are ubiquitouslydistributed. In a phage therapy context this is promising, as itsuggests the possibility of establishing a phage library with broadapplication against V. anguillarum communities worldwide.However, the fast development of phage resistance during expo-sure to specific phages and the indication that different mecha-nisms provided resistance in different hosts also emphasized thatthe susceptibility to phages is a dynamic property in the Vibrio sp.hosts. Consequently, phage resistance mechanisms and dynamicsand the implications for host properties are important to addressin future exploration of phage control of V. anguillarum in aqua-culture.
ACKNOWLEDGMENTS
This study was supported by the Danish Strategic Research Council(ProAqua; project 12-132390), by a grant from the Danish Council forIndependent Research (FNU 09-072829), and by the EU-IRSES-fundedproject AQUAPHAGE.
We thank Pantelis Katharios, Hellenic Centre for Marine Research, for
Tan et al.
3138 aem.asm.org Applied and Environmental Microbiology
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

kindly providing V. anguillarum PF430-3 and vibriophage KVP40 andJette Melchiorsen for excellent technical support.
REFERENCES1. Frans I, Michiels CW, Bossier P, Willems K, Lievens B, Rediers H. 2011.
Vibrio anguillarum as a fish pathogen: virulence factors, diagnosis andprevention. J. Fish Dis. 34:643– 661. http://dx.doi.org/10.1111/j.1365-2761.2011.01279.x.
2. Ransom DP. 1978. Bacteriologic, immunologic and pathogenic studies ofVibrio spp. pathologic to salmonids. Ph.D. dissertation. Oregon State Uni-versity, Corvallis, OR.
3. Joosten PHM, Tiemersma E, Threels A, Caumartin Dhieux C, Rom-bout JHWM. 1997. Oral vaccination of fish against Vibrio anguillarumusing alginate microparticles. Fish Shellfish Immunol. 7:471– 485. http://dx.doi.org/10.1006/fsim.1997.0100.
4. Aoki T, Satoh T, Kitao T. 1987. New tetracycline resistance determinanton R plasmids from Vibrio anguillarum. Antimicrob. Agents Chemother.31:1446. http://dx.doi.org/10.1128/AAC.31.9.1446.
5. Stamm JM. 1989. In vitro resistance by fish pathogens to aquaculturalantibacterials, including the quinolones difloxacin (A-56619) and sara-floxacin (A-56620). J. Aquat. Anim. Health 1:135–141. http://dx.doi.org/10.1577/1548-8667(1989)001�0135:IVRBFP�2.3.CO;2.
6. Middelboe M. 2008. Microbial disease in the sea: effects of viruses oncarbon and nutrient cycling. Princeton University Press, Princeton, NJ.
7. Park SC, Nakai T. 2003. Bacteriophage control of Pseudomonas plecoglos-sicida infection in ayu Plecoglossus altivelis. Dis. Aquat. Org. 53:33–39.http://dx.doi.org/10.3354/dao053033.
8. Castillo D, Higuera G, Villa M, Middelboe M, Dalsgaard I, Madsen L,Espejo R. 2012. Diversity of Flavobacterium psychrophilum and the poten-tial use of its phages for protection against bacterial cold water disease insalmonids. J. Fish Dis. 35:193–201. http://dx.doi.org/10.1111/j.1365-2761.2011.01336.x.
9. Nakai T, Sugimoto R, Park K, Matsuoka S, Mori K, Nishioka T,Maruyama K. 1999. Protective effects of bacteriophage on experimentalLactococcus garvieae infection in yellowtail. Dis. Aquat. Org. 37:33– 41.http://dx.doi.org/10.3354/dao037033.
10. Vinod M, Shivu M, Umesha K, Rajeeva B, Krohn G, Karunasagar I,Karunasagar I. 2006. Isolation of Vibrio harveyi bacteriophage with apotential for biocontrol of luminous vibriosis in hatchery environments.Aquaculture 255:117–124. http://dx.doi.org/10.1016/j.aquaculture.2005.12.003.
11. Imbeault S, Parent S, Lagacé M, Uhland CF, Blais J-F. 2006. Usingbacteriophages to prevent furunculosis caused by Aeromonas salmonicidain farmed brook trout. J. Aquat. Anim. Health 18:203–214. http://dx.doi.org/10.1577/H06-019.1.
12. Stevenson R, Airdrie D. 1984. Isolation of Yersinia ruckeri bacterio-phages. Appl. Environ. Microbiol. 47:1201–1205.
13. Higuera G, Bastías R, Tsertsvadze G, Romero J, Espejo RT. 2013.Recently discovered Vibrio anguillarum phages can protect against exper-imentally induced vibriosis in Atlantic salmon, Salmo salar. Aquaculture392-395:128 –133. http://dx.doi.org/10.1016/j.aquaculture.2013.02.013.
14. Stenholm AR, Dalsgaard I, Middelboe M. 2008. Isolation and charac-terization of bacteriophages infecting the fish pathogen Flavobacteriumpsychrophilum. Appl. Environ. Microbiol. 74:4070 – 4078. http://dx.doi.org/10.1128/AEM.00428-08.
15. Comeau AM, Chan AM, Suttle CA. 2006. Genetic richness of vibrio-phages isolated in a coastal environment. Environ. Microbiol. 8:1164 –1176. http://dx.doi.org/10.1111/j.1462-2920.2006.01006.x.
16. Silva-Rubio A, Avendano-Herrera R, Jaureguiberry B, Toranzo AE,Magarinos B. 2008. First description of serotype O3 in Vibrio anguillarumstrains isolated from salmonids in Chile. J. Fish Dis. 31:235–239. http://dx.doi.org/10.1111/j.1365-2761.2007.00878.x.
17. Lorian V. 2005. Antibiotics in laboratory medicine. Lippincott Williams& Wilkins, Philadelphia, PA.
18. Middelboe M, Chan AM, Bertelsen SK. 2010. Isolation and life cyclecharacterization of lytic viruses infecting heterotrophic bacteria and cya-nobacteria, p 118 –133. In Wilhelm SW, Weinbauer MG, Suttle CA (ed),Manual of aquatic viral ecology. ASLO, Waco, TX.
19. Adams MH. 1959. Bacteriophages. Interscience Publishers, NewYork, NY.
20. Inoue T, Matsuzaki S, Tanaka S. 1995. A 26-kDa outer membraneprotein, OmpK, common to Vibrio species is the receptor for a broad-
host-range vibriophage, KVP40. FEMS Microbiol. Lett. 125:101–105.http://dx.doi.org/10.1111/j.1574-6968.1995.tb07342.x.
21. Ellis EL, Delbrück M. 1939. The growth of bacteriophage. J. Gen. Physiol.22:365–384. http://dx.doi.org/10.1085/jgp.22.3.365.
22. Hidaka T, Tokushige A. 1978. Isolation and characterization of Vibrioparahaemolyticus bacteriophages in sea water. Mem. Fac. Fish. KagoshimaUniv. 27:79 –90.
23. Skov MN, Pedersen K, Larsen JL. 1995. Comparison of pulsed-field gelelectrophoresis, ribotyping, and plasmid profiling for typing of Vibrioanguillarum serovar O1. Appl. Environ. Microbiol. 61:1540 –1545.
24. Matsuzaki S, Tanaka S, Koga T, Kawata T. 1992. A broad-host-rangevibriophage, KVP40, isolated from sea water. Microbiol. Immunol. 36:93–97. http://dx.doi.org/10.1111/j.1348-0421.1992.tb01645.x.
25. Comeau AM, Tremblay D, Moineau S, Rattei T, Kushkina AI, TovkachFI, Krisch HM, Ackermann HW. 2012. Phage morphology recapitulatesphylogeny: the comparative genomics of a new group of myoviruses. PLoSOne 7:e40102. http://dx.doi.org/10.1371/journal.pone.0040102.
26. Comeau AM, Suttle CA. 2007. Distribution, genetic richness and phagesensitivity of Vibrio spp. from coastal British Columbia. Environ. Micro-biol. 9:1790 –1800. http://dx.doi.org/10.1111/j.1462-2920.2007.01299.x.
27. Jensen S, Frost P, Torsvik VL. 2009. The nonrandom microheterogene-ity of 16S rRNA genes in Vibrio splendidus may reflect adaptation toversatile lifestyles. FEMS Microbiol. Lett. 294:207–215. http://dx.doi.org/10.1111/j.1574-6968.2009.01567.x.
28. Pascual J, Macián MC, Arahal DR, Garay E, Pujalte MJ. 2010. Multi-locus sequence analysis of the central clade of the genus Vibrio by using the16S rRNA, recA, pyrH, rpoD, gyrB, rctB and toxR genes. Int. J. Syst. Evol.Microbiol. 60:154 –165. http://dx.doi.org/10.1099/ijs.0.010702-0.
29. Holmfeldt K, Middelboe M, Nybroe O, Riemann L. 2007. Largevariabilities in host strain susceptibility and phage host range governinteractions between lytic marine phages and their Flavobacteriumhosts. Appl. Environ. Microbiol. 73:6730 – 6739. http://dx.doi.org/10.1128/AEM.01399-07.
30. Fischer CR, Yoichi M, Unno H, Tanji Y. 2004. The coexistence ofEscherichia coli serotype O157:H7 and its specific bacteriophage in contin-uous culture. FEMS Microbiol. Lett. 241:171–177. http://dx.doi.org/10.1016/j.femsle.2004.10.017.
31. Kellogg CA, Rose JB, Jiang SC, Thurmond JM, Paul JH. 1995. Geneticdiversity of related vibriophages isolated from marine environmentsaround Florida and Hawaii, USA. Mar. Ecol. Prog. Ser. 120:89 –98. http://dx.doi.org/10.3354/meps120089.
32. Miller ES, Heidelberg JF, Eisen JA, Nelson WC, Durkin AS, CieckoA, Feldblyum TV, White O, Paulsen IT, Nierman WC. 2003. Com-plete genome sequence of the broad-host-range vibriophage KVP40:comparative genomics of a T4-related bacteriophage. J. Microbiol.185:5220 –5233. http://dx.doi.org/10.1128/JB.185.17.5220-5233.2003.
33. Lin YR, Lin CS. 2012. Genome-wide characterization of Vibrio phage�pp2 with unique arrangements of the mob-like genes. BMC Genomics13:224. http://dx.doi.org/10.1186/1471-2164-13-224.
34. Roberts LM, Dunker AK. 1993. Structural changes accompanying chlo-roform-induced contraction of the filamentous phage fd. Biochemistry32:10479 –10488. http://dx.doi.org/10.1021/bi00090a026.
35. Duffy S, Turner PE, Burch CL. 2006. Pleiotropic costs of niche expansionin the RNA bacteriophage �6. Genetics 172:751–757. http://dx.doi.org/10.1534/genetics.105.051136.
36. Southwood T, May R, Hassell M, Conway G. 1974. Ecological strategiesand population parameters. Am. Nat. 108:791– 804. http://dx.doi.org/10.1086/282955.
37. Waterbury PG, Lane MJ. 1987. Generation of lambda phage concatemersfor use as pulsed field electrophoresis size markers. Nucleic Acids Res.15:3930 –3930. http://dx.doi.org/10.1093/nar/15.9.3930.
38. Sun M, Louie D, Serwer P. 1999. Single-event analysis of the packaging ofbacteriophage T7 DNA concatemers in vitro. Biophys. J. 77:1627–1637.http://dx.doi.org/10.1016/S0006-3495(99)77011-6.
39. Middelboe M. 2000. Bacterial growth rate and marine virus-host dynam-ics. Microb. Ecol. 40:114 –124. http://dx.doi.org/10.1007/s002480000050.
40. Riemann L, Grossart HP. 2008. Elevated lytic phage production as aconsequence of particle colonization by a marine Flavobacterium (Cellu-lophaga sp.). Microb. Ecol. 56:505–512. http://dx.doi.org/10.1007/s00248-008-9369-8.
41. Middelboe M, Holmfeldt K, Riemann L, Nybroe O, Haaber J. 2009.Bacteriophages drive strain diversification in a marine Flavobacterium:implications for phage resistance and physiological properties. Environ.
Isolation and Characterization of Vibriophages
May 2014 Volume 80 Number 10 aem.asm.org 3139
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

Microbiol. 11:1971–1982. http://dx.doi.org/10.1111/j.1462-2920.2009.01920.x.
42. Kim M, Ryu S. 2011. Characterization of a T5-like coliphage, SPC35, anddifferential development of resistance to SPC35 in Salmonella entericaserovar Typhimurium and Escherichia coli. Appl. Environ. Microbiol. 77:2042–2050. http://dx.doi.org/10.1128/AEM.02504-10.
43. Kocharunchitt C, Ross T, McNeil D. 2009. Use of bacteriophages asbiocontrol agents to control Salmonella associated with seed sprouts. Int.J. Food Microbiol. 128:453– 459. http://dx.doi.org/10.1016/j.ijfoodmicro.2008.10.014.
44. Sheng H, Knecht HJ, Kudva IT, Hovde CJ. 2006. Application of bacte-riophages to control intestinal Escherichia coli O157:H7 levels in rumi-nants. Appl. Environ. Microbiol. 72:5359 –5366. http://dx.doi.org/10.1128/AEM.00099-06.
45. Gödeke J, Paul K, Lassak J, Thormann KM. 2011. Phage-induced lysisenhances biofilm formation in Shewanella oneidensis MR-1. ISME J.5:613– 626. http://dx.doi.org/10.1038/ismej.2010.153.
46. Labrie SJ, Samson JE, Moineau S. 2010. Bacteriophage resistance mech-anisms. Nat. Rev. Microbiol. 8:317–327. http://dx.doi.org/10.1038/nrmicro2315.
47. Heilmann S, Sneppen K, Krishna S. 2012. Coexistence of phage andbacteria on the boundary of self-organized refuges. Proc. Natl. Acad. Sci.U. S. A. 109:12828 –12833. http://dx.doi.org/10.1073/pnas.1200771109.
48. Høyland-Kroghsbo NM, Mærkedahl RB, Svenningsen SL. 2013. A quo-rum-sensing-induced bacteriophage defense mechanism. mBio 4:e00362-12. http://dx.doi.org/10.1128/mBio.00362-12.
49. Pedersen K, Gram L, Austin D, Austin B. 1997. Pathogenicity ofVibrio anguillarum serogroup O1 strains compared to plasmids, outermembrane protein profiles and siderophore production. J. Appl. Micro-biol. 82:365–371. http://dx.doi.org/10.1046/j.1365-2672.1997.00373.x.
50. Schrøder MB, Ellingsen T, Mikkelsen H, Norderhus EA, Lund V.2009. Comparison of antibody responses in Atlantic cod (Gadusmorhua L.) to Vibrio anguillarum, Aeromonas salmonicida and Francisellasp. Fish Shellfish Immunol. 27:112–119. http://dx.doi.org/10.1016/j.fsi.2008.11.016.
51. Hjelm M, Bergh Ø, Riaza A, Nielsen J, Melchiorsen J, Jensen S, DuncanH, Ahrens P, Birkbeck H, Gram L. 2004. Selection and identification ofautochthonous potential probiotic bacteria from turbot larvae (Scophthal-mus maximus) rearing units. Syst. Appl. Microbiol. 27:360 –371. http://dx.doi.org/10.1078/0723-2020-00256.
Tan et al.
3140 aem.asm.org Applied and Environmental Microbiology
on April 29, 2014 by C
openhagen University Library
http://aem.asm
.org/D
ownloaded from

Figure S1. Neighbour-joining tree based on 16S rRNA gene sequencing showing the phylogenetic
relationship between 37 strains listed in table 1 and V. harveyi (AY747308.1, NCBI) and V. parahaemolyticus
(FJ594056.1, NCBI). Phylogenetic relationships were bootstrapped 1,000 times. The scale bar represents
the number of nucleotide substitutions per site.
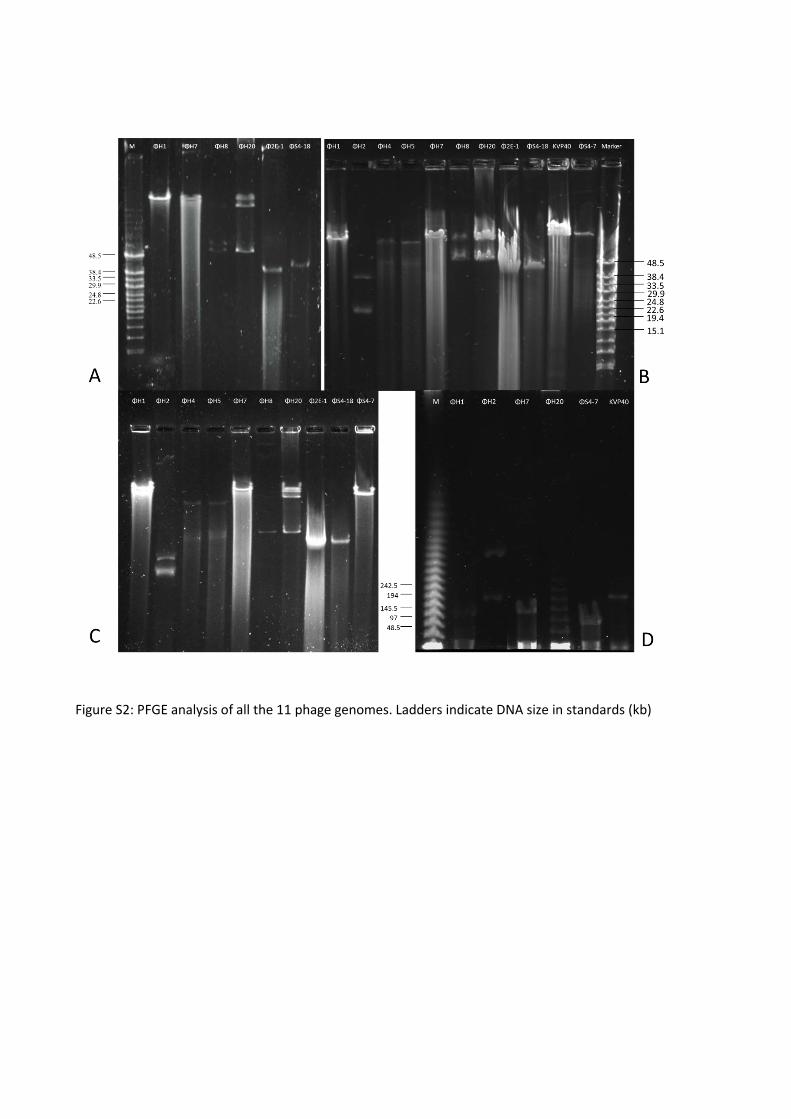
Figure S2: PFGE analysis of all the 11 phage genomes. Ladders indicate DNA size in standards (kb)

1
Vibriophages differentially influence biofilm formation by Vibrio anguillarum strains 1
Demeng Tan, Amalie Dahl and Mathias Middelboe* 2
Marine Biological Section, University of Copenhagen, Helsingør, Denmark 3
4
*correspondence: [email protected] 5
Running title: The interaction of vibriophages and biofilms 6
7
8

2
Abstract 9
Vibrio anguillarum is an important pathogen in marine aquaculture, responsible for vibriosis. 10
Bacteriophages can potentially be used to control bacterial pathogens; however, successful 11
application of phages requires a detailed understanding of phage-host interactions at both free-12
living and surface-associated growth conditions. In this study, we explored in vitro phage-host 13
interactions in two different strains of V. anguillarum (BA35 and PF430-3) during growth in micro-14
colonies, biofilms, and free-living cells. Two vibriophages, ΦH20 (Siphoviridae) and KVP40 15
(Myoviridae), had completely different effects on the biofilm development. Addition of phage ΦH20 16
to strain BA35 showed efficient control of biofilm formation as well as in liquid cultures. The 17
interactions between BA35 and ΦH20 were thus characterized by a strong phage control of the 18
phage-sensitive population and subsequent selection for phage-resistant mutants. Addition of phage 19
KVP40 to strain PF430-3 resulted in increased biofilm development, especially during the early stage. 20
Subsequent experiments in liquid cultures showed that addition of phage KVP40 stimulated the 21
aggregation of host cells, which protected the cells against phage infection. By the formation of 22
biofilms, strain PF430-3 created spatial refuges that protected the host from phage infection and 23
allowed coexistence between phage-sensitive cells and lytic phage KVP40. Together the results 24
demonstrate highly variable phage protection mechanisms in two closely related V. anguillarum 25
strains, thus emphasizing the challenges of using phages to control vibriosis in aquaculture, and 26
adding to the complex roles of phages as drivers of prokaryotic diversity and population dynamics. 27
28
29
30
31
32

3
Introduction 33
Vibrio anguillarum is a marine pathogenic bacterium causing vibriosis, a fatal hemorrhagic 34
septicemia, which contributes to significant mortalities in fish and shellfish aquaculture worldwide 35
(1-3). The persistence of Vibrio pathogens in aquaculture has been attributed to their ability to form 36
biofilms with increased tolerance to disinfectants and antibiotics (4, 5). Moreover, the first stage of 37
infection involves biofilm-like micro-colonies in the skin tissue, causing chronic infection (5). 38
Recently, bacteriophages have been suggested as potential agents of pathogen control in 39
aquaculture, and the controlling effects of phages have been explored for a number of fish 40
pathogens (6-8). Successful application of phages to reduce vibriosis-related mortality has been 41
demonstrated (9, 10). The capabilities of some phages to produce depolymerases, which hydrolyze 42
extracellular polymers in bacterial biofilms, have made the use of bacteriophages particularly 43
relevant in the treatment of biofilm-forming pathogens, as demonstrated in biofilms of 44
Pseudomonas aeruginosa (11), Escherichia coli (12), and Staphylococcus aureus (13). 45
Apart from the potential physical and chemical barrier provided by biofilms, the use of 46
bacteriophages to control pathogens is challenged by the development and selection for phage-47
resistant or phage-tolerant subpopulations. A broad range of resistance mechanisms have evolved in 48
bacteria (14) and the lytic infection of phage-susceptible bacteria has shown to drive a functional 49
and genetic diversification of the host population based on the rapid emergence of phage-resistant 50
mutants (15, 16). A deeper understanding of temporal and spatial dynamics of phage-biofilm 51
interactions and the development of phage-resistance in these systems is therefore essential for 52
assessing the potential of using phages in disease control. 53
In this study, we quantified the influence of phages on biofilm formation in two strains of V. 54
anguillarum, both during the initial stages of attachment and micro-colony formation and in more 55
developed biofilms. The results showed large intraspecific differences in phage-biofilm interactions 56

4
of two V. anguillarum phage-host systems, and revealed the presence of highly different protective 57
mechanisms against phage infection, adding further complexity to the use of phages to control 58
vibriosis. 59
Materials and Methods 60
Bacterial strains and bacteriophages 61
Bacteriophages and hosts used in this study are listed in Table 1. Two V. anguillarum strains were 62
used in this study: strain PF430-3, originally isolated from salmonid aquaculture in Chile (17) and 63
strain BA35, originally isolated from sockeye salmon in USA (18). Phage KVP40 which infects PF430-3 64
is a broad-host-range phage originally isolated from Japan (19, 20) and phage ΦH20, infecting BA35, 65
was isolated from Danish aquaculture (21). 66
Effects of phages on initial bacterial attachment and micro-colony formation 67
To quantify the effect of phages on micro-colony formation of the two strains, 10 µl of diluted mid-68
log-phase cultures (106 CFU ml-1) was filtered onto 20+ replicate 25 mm 0.2 µm polycarbonate filters 69
and incubated on LB agar in 6-well plates (22). Two ml phage stock (ca. 108 PFU ml-1) was added to 70
half the filters, the other half serving as controls with addition of 2 ml SM buffer (50 mM Tris-Cl, pH 71
7.5, 100 mM NaCl, 8 mM MgSO4, 0.01% gelatin). Duplicate filters of phage amended and controls 72
were collected every hour for 8 h, transferred to a 25 mm filtration manifold (Millipore) and stained 73
with 0.5 % SYBR Gold (Invitrogen) for 10 min, followed by 3 x rinsing with Milli-Q water. The filters 74
were then mounted on a microscope slide and stored frozen until quantification by epifluorescence 75
microscopy using Cell M image analysis software (Olympus) for determination of colony area. 76
Phage-biofilm interactions 77
Effects of phage ΦH20 and KVP40 on the formation and disruption of V. anguillarum biofilms were 78
monitored for the two strains BA35 and PF430-3 in two different experimental approaches: In the 79

5
pre-treated experiment, duplicate sets of polypropylene plastic tubes (Sarstedt) with 5 ml marine 80
broth (MB, Difco) were inoculated with 100 μl overnight bacterial culture and 100 μl phage stock 81
(final concentration 107 CFU ml-1 and 106 PFU ml-1, multiplicity of infection, MOI=0.1) simultaneously. 82
In the post-treated experiments, the bacteria were allowed to grow in 5 ml MB for 10 days after 83
inoculation to establish a biofilm. After 10 day incubation, the liquid was removed and the tubes 84
were rinsed with MB. Then 5 ml MB was added along with 100 μl phage stock (final concentration 85
106 PFU ml-1), and the tubes were incubated for another 7-8 days. 86
In both experiments, the tubes were incubated at 30 °C and duplicate tubes were sacrificed daily for 87
measurement of biofilm formation, and density of free-living bacteria and phages. Measurements of 88
OD600nm were used to estimate free-living bacterial abundance and determination of phage 89
concentration in the free-living phase was done by plaque assay (23). Biofilm was quantified 90
according to O'Toole (24) with some modifications. Briefly, the liquid was removed, and tubes were 91
rinsed twice with artificial seawater (ASW, Sigma). Then 6 ml 0.4% crystal violet (Sigma) was added 92
to each tube and after 15 min, stain was removed. Tubes were washed with tap water in order to 93
remove excess stain and left to dry for 5 min. Six ml of 33% acetic acid (Sigma) was added and left 94
for 5 min to allow the stain to dissolve. The absorbance was measured at OD595nm. 95
In addition to the measurements above, we also quantified the biofilm-associated infective phages 96
in the post-treated experiment after disruption of the biofilm by sonication. First, the liquid was 97
removed and the tube rinsed 3 times with ASW. Then, 6 ml ASW was added to completely cover the 98
biofilm, and the tube was sonicated for 10 s at 80 amplitude (Pulse on 1 s and Pulse off 1 s, Misonix 99
sonication bath) to dislodge bacteria and phages from the biofilm matrix. Serial dilutions were 100
performed and phage concentration was quantified by plaque assay (23). 101
Phage-host interactions in the free-living phase: detection of aggregate formation 102
The effects of phages ΦH20 and KVP40 on the growth and aggregate formation of their respective 103
host strains, BA35 and PF430-3, during free-living growth in liquid cultures were examined. 104

6
Overnight cultures were inoculated in 50 ml MB with their respective phage at a MOI of 0.1. The 105
flasks were incubated at room temperature with agitation; samples were collected for 106
determination of the presence of cell aggregates by epifluorescence microscopy. For epifluorescence 107
microscopic analysis of cell aggregation, subsamples were filtered onto 0.45 μm polycarbonate 108
membrane filters (Whatman) and stained with SYBR Gold (0.5%, Invitrogen) for 10 min. Bacteria and 109
phages on the filters were distinguished based on their dimensions and/or their relative brightness. 110
Phage susceptibility and physiological fingerprint of bacterial isolates 111
In order to determine changes in phage susceptibility, a total of ~180 cells were isolated from phage-112
treated micro-colony experiments, pre-treated biofilm experiments and free-living samples and 113
analyzed for phage susceptibility pattern. For colony isolation, samples were in all cases plated onto 114
Thiosulfate Citrate Bile Salt Sucrose agar (TCBS, Difco) and incubated for 24 h. From these plates, 115
single colonies were isolated and purified by 3 rounds of re-isolation and then transferred to 4 ml 116
MB for subsequent phage susceptibility analysis. Phage susceptibility of the isolated strains was 117
determined by spot tests (21). 118
BIOLOG GN2 Micro-Plates (BIOLOG) containing 95 different carbon sources were used to test the 119
ability of selected phage-amended isolates to use different substrates following manufacturer’s 120
instructions (21). For strain PF430-3, the procedure was slightly modified as bacterial lysate 121
(bacterial culture subjected to phage-induced lysis) and aggregates were collected by centrifugation 122
at a lower speed (3,000×g, 5 min), and washed twice with 0.9% saline buffer. 123
Adsorption experiments with fluorescently labelled phages 124
Phage adsorption to selected isolates was tested by adding SYBR Gold labelled phages to cultures 125
followed by visual inspection phage binding to the cells using epifluorescence microscopy, following 126
the protocol of Kunisaki (25) with some modifications. Aliquots of 600 μl phage lysate were digested 127
by addition of DNase I (1 μl DNase I +3 μl RDD buffer, Qiagen) at 37°C for 2 h and then stained with 128

7
SYBR Gold (final concentration 5×, Invitrogen) overnight at 4 °C. Ten μl chloroform was then added 129
to inactivate DNase I. The phage stock was then further filtered through a 30 kDa ultrafiltration spin 130
devices (Millipore) at 1,000×g for 90 min to remove the free SYBR Gold. The SYBR Gold-labelled 131
phages were added to the mid-log wild-type cells and phage lysate, respectively, at a MOI of 132
approximately 10 and incubated at room temperature for 20 min. Samples were pelleted at 133
12,000×g for 3 min, and the pellet was re-suspended in a small volume of SM buffer, mixed with pre-134
heated 0.5% agarose (w/v) (Bio-rad) around 45°C and transferred to gel-coated slide glass. 135
Statistical analysis 136
All statistics were performed using the Student's paired t-test (two-tailed) (OriginPro 8.6) to 137
determine the significance of the differences between phage amended and control cultures. 138
Differences were considered to be significant if P<0.05. 139
Results 140
Initial biofilm formation and short-term effects of phages 141
The two V. anguillarum strains formed micro-colonies with highly different morphologies in the 142
absence of their corresponding phages. Strain BA35 formed simple single-layered micro-colonies, 143
where individual cells could easily be identified whereas strain PF430-3 formed complex 3-144
dimensional volcano-shaped structures (Fig. 1). Phage ΦH20 showed an efficient control of micro-145
colonies BA35 according to the microscope observations, reducing total colony area from 41,000 146
µm2 per mm2 filter on the control filters to 5,000 µm2 per mm2 filter after 6 h incubation, and leaving 147
mainly single cells on the filters following phage exposure (data not shown). For strain PF430-3, the 148
individual colonies were unaffected by phage exposure, whereas single cells were lysed by phage 149
KVP40, leaving mainly colonies on the filters 6 h post addition (Fig. 1). 150
Effects of phages on biofilm formation 151

8
Establishment and growth of biofilm produced by the two V. anguillarum strains BA35 and PF430-3 152
in cultures pre-treated with phage ΦH20 and KVP40, respectively, was examined over a long-term 153
experiment in parallel to control cultures without phages. The OD600nm of free-living bacteria in the 154
control cultures reached maximum values of 0.72±0.01 and 0.98±0.01 for BA35 and PF430-3, 155
respectively, after 2 days (Fig. 2A and 2C). A gradual minor decrease was then observed for BA35, 156
whereas the density of PF430-3 decreased to ~50 % of the maximum value at day 9 (Fig. 2A and 2C). 157
The presence of the phages had different effects on the free-living cell density of the two strains. 158
For strain BA35, the density reached a level of 70 % of the control cultures after 2 days and remained 159
at this level (only significantly lower at day 3 (p=0.025) and 6 (p=0.009)) throughout the experiment 160
(Fig. 2A). In the cultures with strain PF430-3, addition of phage KVP40 significantly reduced the 161
optical density of the culture (0.008≤p≤0.027 when comparing the OD values at individual time 162
points from day 2 onwards) to approximately 25% of the corresponding values in the control 163
cultures (Fig. 2C). Phage concentration reached peaks of around 1.3×1010±3.5×108 and 164
3.4×108±7.1×106 PFU ml-1 for ΦH20 and KVP40, respectively at day 1 (Fig. 2A and2C). Interestingly, 165
in the PF430-3+KPV40 cultures, phage concentrations decreased significantly (p=0.007) from day 1 166
to day 2 then stabilizing around 106 PFU ml-1 (Fig. 2C). Likewise, the concentrations phage ΦH20 167
decreased significantly (p=0.0055) from day 1 to day 5 and then remained at a titer above 168
2.2×109±2.8×108 PFU ml-1 (Fig. 2A). 169
The biofilm development in response to phage addition also showed completely different patterns in 170
the two strains with either inhibition (BA35) or stimulation (PF430-3) of biofilm formation relative to 171
control cultures. In the BA35+ΦH20 pre-treated cultures, phage ΦH20 effectively prevented biofilm 172
formation, causing a 2-fold decrease in the biofilm biomass relative to the control level (Fig. 2B). In 173
the PF430-3+KVP40 pre-treated cultures, phage KVP40 addition resulted in almost a 3-fold increase 174
relative to the control (Fig. 2D). In the control cultures, there was a steady increase in biofilm density 175

9
for strain PF430-3, whereas a fast initial establishment of biofilm of strain BA35 was gradually 176
reduced during the 7-day period (Fig. 2B and 2D). 177
To assess the effect of phages on fully matured biofilms, 10-day biofilms were challenged with their 178
corresponding phages measuring the same parameters as above. As in the pre-treatment 179
experiments, phage addition significantly reduced the BA35 biofilm (3-4 fold reduction) within 2 180
days (p=0.046, when comparing OD values from day 1 and 2) and slightly stimulated the production 181
of biofilm in PF430-3, relative to controls (Fig. 3B and 3D). 182
For the free-living populations, the input of nutrients associated with the addition of phages (and 183
corresponding addition of medium in the controls) caused an increase in bacterial concentration in 184
the control cultures (Fig. 3A and 3C). Similar to the pre-treated cultures, the OD600nm decreased over 185
time in the control cultures, with the strongest decrease observed in the PF430-3 cultures. The 186
presence of phages, however, imposed a significant 4-5 fold decrease in concentration of free-living 187
cells in both experiments (0.003≤p≤0.046, when comparing the OD values at individual time points 188
from day 2 onwards), except for day 5 (Fig. 3A and 3C). 189
The concentration of free-living phages reached a peak around 3×109±2.8×107 and 5.8×108±2.8×107 190
PFU ml-1 for ΦH20 and KVP40, respectively, after 24 h (Fig. 3A and 3C) followed by a gradual 191
decrease in the concentration of phage KVP40 (Fig. 3C). The biofilm-associated concentration of 192
phage KVP40 constituted less than 10% of the free-living population and decreased dramatically 193
after the peak (Fig. 3D). 194
Effects of phage KVP40 on bacterial aggregation in liquid cultures 195
In the BA35 and BA35+ΦH20 cultures, no aggregate formation was observed and phage addition 196
reduced the density of free-living cells relative to the control as a result of cell lysis (Fig. 4). Motility 197
of BA35 seemed to be affected by phage addition as cells in the phage-added cultures had reduced 198
or lost motility after 24 h (Supplementary video 1 and 2). In the PF430+KVP40 cultures, on the other 199

10
hand, aggregates of multi-layered cell clusters embedded in a matrix were observed after 24 h (Fig. 4 200
and Supplementary video 3 and 4). 201
Bacteriophage resistance and physiological fingerprint 202
For strain BA35, all the isolates obtained from the micro-colony experiment were resistant to phage 203
infection and approximately 30 % of the free-living and biofilm-associated isolates were sensitive to 204
ΦH20 (Table S1). The resistant colonies were small-colony variants on TCBS plates. In contrast, all 205
the isolates from KVP40-exposed PF430-3 cultures were fully or partially susceptible to phage KVP40 206
and no completely KVP40-resistant mutants were isolated (Table S1). Colony morphologies of PF430-207
3 isolates were similar to the wild-type PF430-3. Spot assays showed that all partially resistant 208
isolates regained full sensitivity to phage KVP40 after purification by re-streaking of single colonies 209
on agar plates. 210
The phenotypic diversity and implications of resistance for the metabolic properties of the isolates 211
following phage exposure were explored by obtaining a metabolic fingerprint of selected isolates 212
using BIOLOG GN2 assay (Fig. S1). For strain BA35, the phage-resistant isolate had lost the ability to 213
use 10 of the 54 substrates that could be metabolized by wild-type BA35 cells, corresponding to a 19% 214
reduction of the capacity substrate utilization. In contrast, none of the PF430-3 isolates obtained 215
after phage exposure showed any changes in physiological fingerprint according to the BIOLOG 216
profile (Fig. S1). 217
To further examine the mechanisms of resistance/tolerance against phages in the two strains, SYBR 218
Gold-labelled phages ΦH20 and KPV40 were incubated with wild-type cells and phage lysate. 219
Fluorescently labeled phage ΦH20 attached to the sensitive cells of the strain BA35 (Fig. 5A), 220
whereas no phage-attachment was observed in phage-resistant mutants (Fig. 5B). In the parallel 221
experiment with fluorescently labelled KVP40 phages, the phages were immobilized in the 222
aggregates and did not attach to the cell surface (Fig. 5C and 5D). 223

11
Discussion 224
Phage effects on micro-colony formation and initial biofilm development 225
The microscopic observations of micro-colony formation of cells attached to a polycarbonate filter 226
surface revealed that the two strains formed highly different types of micro-colonies, with large 227
implications for the phage-host interactions. The phage-resistant isolates obtained from BA35 228
cultures exposed to ΦH20 maintained resistance upon re-culturing, suggesting that resistance was a 229
result of genetic mutations which prevented phage infection. In contrast to the strong lytic effect of 230
ΦH20 and corresponding reduction in micro-colony formation of strain BA35, phage KVP40 did not 231
significantly affect the size of individual PF430-3 micro-colonies. A possible explanation could be that 232
the complex 3-dimensional structure of PF430-3 micro-colonies provided protection from phage 233
infection by creating physical barriers, once formed. 234
Long-term effects of phages on biofilm formation from liquid cultures of different V. anguillarum 235
strains 236
The strong controlling effects of phage ΦH20 on the strain BA35 biofilm observed both in the pre-237
treated and post-treated conditions supported the microscopic observations that the BA35 biofilm is 238
a relatively simple structure which is easily accessible for phage adsorption. Also in the free-living 239
phase, phage ΦH20 was able to control the phage susceptible BA35 population, but at these 240
conditions, phage-resistant mutants replaced the sensitive population within the first 24 h. The 241
interactions between ΦH20 and strain BA35 thus seemed very similar for attached and free-living 242
cells, suggesting that the flat and smooth biofilm structure did not offer protection against phage 243
infection. 244
In contrast, addition of phage KVP40 to strain PF430-3 resulted in an enhanced biofilm formation in 245
both experiments, and particularly during the early stage of biofilm development. At the same time, 246
the free-living bacterial population was efficiently controlled and maintained at an OD level of <25 % 247

12
of the maximum density in the control cultures, in both the pre-treated and post-treated PF430-3 248
experiments. This suggested a coupling between the phage-mediated decrease in density of free-249
living cells and the corresponding cell aggregate formation and subsequent increase in biofilm 250
formation. 251
Potential mechanisms of phage protection in V. anguillarum strains 252
While the dominance of phage resistant strains following phage exposure suggested mutational 253
changes as the mechanism of phage protection in strain BA35, the interaction between strain PF430-254
3 and phage KVP40 pointed towards aggregation and biofilm formation as a protection against 255
phage infection. This mechanism of protection was supported by a number of observations: 1), 256
addition of fluorescently labelled KVP40 phage to aggregates and free-living cells of PF430-3 showed 257
that KVP40 was trapped in the aggregate matrix and did not adsorb to the host cells embedded in 258
the aggregates; 2), the complex 3-dimensional structure of the micro-colonies observed for PF430-3 259
was also consistent with the hypothesis that the aggregation provided a refuge for phage-sensitive 260
bacteria (26-28); and 3), the presence of a fraction of free-living phage-sensitive cells outside the 261
aggregates may in part be explained by the entrapment of phages in the aggregates which reduced 262
the phage encounter rate, possibly allowing co-existence of small populations of free-living phages 263
and free-living sensitive cells. 264
In combination with the reduced phage-sensitivity found in a number of the isolates, the 265
immobilization of free-living cells in aggregates and subsequent attachment to surfaces provides an 266
efficient refuge for cells exposed to phage attack, thus adding to the multiple and complex 267
protection mechanisms found in bacteria (14, 29, 30). Whether the aggregation is a direct response 268
to phage exposure, e.g. through quorum sensing (QS)-mediated gene regulation which is well known 269
from V. anguillarum (31-33), or is an indirect effect of phage-lysis of free-living cells, thus adding a 270
strong positive selection pressure for aggregate-forming phenotypes needs to be elucidated (30). 271

13
It should be noted that successful phage amplification depends not only on the host cell lysis, but 272
also on the phages able to bind to the susceptible bacterium. Polysaccharide depolymerases have 273
been reported in various phages, which allow phages to access phage receptors in biofilm-associated 274
cells (34), Consequently, the generality of protection mechanisms observed in the present study is 275
unknown, as other phages may be able to infect the biofilm-forming strain PF430-3. It is often 276
assumed that bacteriophage resistance is associated with a cost (35, 36) due to reduced abilities to 277
take up specific nutrients (37) and reduced competitive abilities in general (38, 39). The BIOLOG 278
fingerprint of resistant isolates of strain BA35 supported such a link between phage-resistance and 279
reduced physiological performance, as the resistant mutants had a reduced ability to utilize a 280
number of substrates. The absence of observed physiological changes in the phage-exposed isolates 281
of strain PF430-3 relative to the wild-type strain , on the other hand supported the suggestion that 282
phage tolerance in strain PF430-3 was not caused by mutational changes but rather was due to 283
protection by aggregation. It should be emphasized however, other fitness costs than those resolved 284
by the BIOLOG assay may have been present in strain PF430-3. 285
Interestingly, the free-living isolates obtained from KVP40-amended PF430-3 cultures, which had 286
reduced phage susceptibility, all recovered full sensitivity after re-growth in the absence phage 287
KVP40. This indicated that the phage tolerance was a transient phenotype, as has also been 288
observed in other phage infection studies including hosts such as V. cholerae (40), S. enterica (41), S. 289
oranienburg (42), and E. coli O157:H7 (41, 43), and suggests that down-regulation of phage receptor 290
gene expression may also play a role in phage susceptibility in strain PF430-3 (44). 291
In conclusion, our data demonstrated completely different mechanisms of protection against phages 292
in two strains of V. anguillarum and showed that these intraspecific differences strongly influenced 293
the outcome of the phage-host interactions with respect to 1) the ability of phages to control free-294
living host populations; 2) the ability of phages to control the development and stability of biofilm; 3) 295
the phage-driven diversification of bacterial populations; and 4) the co-existence and co-evolution of 296

14
phages and hosts. However, additional studies are needed to explore whether the different 297
responses to phage exposure are general properties of the strains or are linked to the specific 298
phages. 299
Implications for phage-control of the pathogen 300
In a phage therapy context, the interaction between strain BA35 and phage ΦH20 suggests that 301
phage control of strain BA35 potentially would prove useful, since the phage efficiently inhibited 302
biofilm formation both on short and longer time scale as well as proved efficient in disrupting 303
already established biofilms. The phage-derived stimulation of biofilm formation in strain PF430-3 304
may, on the other hand, suggests that phage KVP40 would be inefficient in controlling this strain in 305
aquaculture, and perhaps rather stimulate its pathogenic impact by inducing biofilm formation. 306
Our study thus confirms that successful application of phage therapy in the treatment of biofilm 307
requires detailed understanding of phage-host interactions, and emphasizes that the complexity and 308
diversity of phage-host interactions even within the same species of pathogen is a challenge for 309
future use of phages in disease control. 310
Acknowledgements 311
This study was supported by grants from the Danish Strategic Research Council (ProAqua project 12-312
132390), the Danish Council for Independent Research (FNU 09-072829) and the EU-IRSES-funded 313
project AQUAPHAGE (269175). We thank Pantelis Katharios, Hellenic Centre for Marine Research, 314
for kindly providing V. anguillarum PF430-3 and vibriophage KVP40 and Lone Gram, Technical 315
University of Denmark, for kindly providing V. anguillarum BA35. Jeanett Hansen is acknowledged 316
for technical support. 317
References 318
1. Frans I, Michiels CW, Bossier P, Willems KA, Lievens B, Rediers H. 2011. Vibrio anguillarum 319 as a fish pathogen: virulence factors, diagnosis and prevention. J Fish Dis 34:643-661. 320

15
2. Evelyn T. 1971. First records of vibriosis in Pacific salmon cultured in Canada, and taxonomic 321 status of the responsible bacterium, Vibrio anguillarum. J Fish Res Board Can 28:517-525. 322
3. Larsen JL, Pedersen K, Dalsgaard I. 1994. Vibrio anguillarum serovars associated with 323 vibriosis in fish. J Fish Dis 17:259-267. 324
4. Hall-Stoodley L, Costerton JW, Stoodley P. 2004. Bacterial biofilms: from the natural 325 environment to infectious diseases. Nat Rev Microbiol 2:95-108. 326
5. Lindell K, Fahlgren A, Hjerde E, Willassen NP, Fallman M, Milton DL. 2012. 327 Lipopolysaccharide O-antigen prevents phagocytosis of Vibrio anguillarum by rainbow trout 328 (Oncorhynchus mykiss) skin epithelial cells. PLoS One 7:e37678. 329
6. Nakai T, Sugimoto R, Park K, Matsuoka S, Mori K, Nishioka T, Maruyama K. 1999. 330 Protective effects of bacteriophage on experimental Lactococcus garvieae infection in 331 yellowtail. Dis. Aquat. Org. 37:33-41. 332
7. Imbeault S, Parent S, Lagacé M, Uhland CF, Blais J-F. 2006. Using bacteriophages to prevent 333 furunculosis caused by Aeromonas salmonicida in farmed brook trout. J. Aquat. Anim. Health 334 18:203-214. 335
8. Stenholm AR, Dalsgaard I, Middelboe M. 2008. Isolation and characterization of 336 bacteriophages infecting the fish pathogen Flavobacterium psychrophilum. Appl Environ 337 Microbiol 74:4070-4078. 338
9. Karunasagar I, Shivu MM, Girisha SK, Krohne G, Karunasagar I. 2007. Biocontrol of 339 pathogens in shrimp hatcheries using bacteriophages. Aquaculture 268:288-292. 340
10. Vinod M, Shivu M, Umesha K, Rajeeva B, Krohn G, Karunasagar I, Karunasagar I. 2006. 341 Isolation of Vibrio harveyi bacteriophage with a potential for biocontrol of luminous vibriosis 342 in hatchery environments. Aquaculture 255:117-124. 343
11. Fu W, Forster T, Mayer O, Curtin JJ, Lehman SM, Donlan RM. 2010. Bacteriophage cocktail 344 for the prevention of biofilm formation by Pseudomonas aeruginosa on catheters in an in 345 vitro model system. Antimicrob Agents Chemother 54:397-404. 346
12. Doolittle MM, Cooney JJ, Caldwell DE. 1995. Lytic infection of Escherichia coli biofilms by 347 bacteriophage T4. Can J Microbiol 41:12-18. 348
13. Sass P, Bierbaum G. 2007. Lytic activity of recombinant bacteriophage phi11 and phi12 349 endolysins on whole cells and biofilms of Staphylococcus aureus. Appl Environ Microbiol 350 73:347-352. 351
14. Labrie SJ, Samson JE, Moineau S. 2010. Bacteriophage resistance mechanisms. Nat Rev 352 Microbiol 8:317-327. 353
15. Martiny JB, Riemann L, Marston MF, Middelboe M. 2014. Antagonistic coevolution of 354 marine planktonic viruses and their hosts. Ann Rev Mar Sci 6:393-414. 355
16. Freese E. 1959. The specific mutagenic effect of base analogues on phage T4. J Mol Biol 356 1:87-105. 357
17. Silva-Rubio A, Avendano-Herrera R, Jaureguiberry B, Toranzo AE, Magarinos B. 2008. First 358 description of serotype O3 in Vibrio anguillarum strains isolated from salmonids in Chile. J 359 Fish Dis 31:235-239. 360
18. Pedersen K, Gram L, Austin DA, Austin B. 1997. Pathogenicity of Vibrio anguillarum 361 serogroup O1 strains compared to plasmids, outer membrane protein profiles and 362 siderophore production. J Appl Microbiol 82:365-371. 363
19. Inoue T, Matsuzaki S, Tanaka S. 1995. A 26-kDa outer membrane protein, OmpK, common 364 to Vibrio species is the receptor for a broad-host-range vibriophage, KVP40. FEMS Microbiol 365 Lett 125:101-105. 366
20. Miller ES, Heidelberg JF, Eisen JA, Nelson WC, Durkin AS, Ciecko A, Feldblyum TV, White O, 367 Paulsen IT, Nierman WC, Lee J, Szczypinski B, Fraser CM. 2003. Complete genome sequence 368 of the broad-host-range vibriophage KVP40: comparative genomics of a T4-related 369 bacteriophage. J Bacteriol 185:5220-5233. 370

16
21. Tan D, Gram L, Middelboe M. 2014. Vibriophages and their interactions with the fish 371 pathogen Vibrio anguillarum. Appl Environ Microbiol 80:3128-3140. 372
22. Hojberg O, Binnerup SJ, Sorensen J. 1997. Growth of silicone-immobilized bacteria on 373 polycarbonate membrane filters, a technique to study microcolony formation under 374 anaerobic conditions. Appl Environ Microbiol 63:2920-2924. 375
23. Adams MH. 1959. Bacteriophages. Interscience Publishers, New York, N.Y. 376 24. O'Toole GA, Pratt LA, Watnick PI, Newman DK, Weaver VB, Kolter R. 1999. Genetic 377
approaches to study of biofilms. Methods Enzymol 310:91-109. 378 25. Kunisaki H, Tanji Y. 2010. Intercrossing of phage genomes in a phage cocktail and stable 379
coexistence with Escherichia coli O157:H7 in anaerobic continuous culture. Appl Microbiol 380 Biotechnol 85:1533-1540. 381
26. Heilmann S, Sneppen K, Krishna S. 2012. Coexistence of phage and bacteria on the 382 boundary of self-organized refuges. Proc Natl Acad Sci U S A 109:12828-12833. 383
27. Schrag SJ, Mittler JE. 1996. Host-parasite coexistence: The role of spatial refuges in 384 stabilizing bacteria-phage interactions. American Naturalist 148:348-377. 385
28. Brockhurst MA, Buckling A, Rainey PB. 2006. Spatial heterogeneity and the stability of host-386 parasite coexistence. J Evol Biol 19:374-379. 387
29. Samson JE, Magadan AH, Sabri M, Moineau S. 2013. Revenge of the phages: defeating 388 bacterial defences. Nat Rev Microbiol 11:675-687. 389
30. Koskella B, Brockhurst MA. 2014. Bacteria-phage coevolution as a driver of ecological and 390 evolutionary processes in microbial communities. FEMS Microbiol Rev 38:916-931. 391
31. Croxatto A, Chalker VJ, Lauritz J, Jass J, Hardman A, Williams P, Camara M, Milton DL. 2002. 392 VanT, a homologue of Vibrio harveyi LuxR, regulates serine, metalloprotease, pigment, and 393 biofilm production in Vibrio anguillarum. J Bacteriol 184:1617-1629. 394
32. Croxatto A, Pride J, Hardman A, Williams P, Camara M, Milton DL. 2004. A distinctive dual-395 channel quorum-sensing system operates in Vibrio anguillarum. Mol Microbiol 52:1677-396 1689. 397
33. Milton DL. 2006. Quorum sensing in vibrios: complexity for diversification. Int J Med 398 Microbiol 296:61-71. 399
34. Hughes KA, Sutherland IW, Jones MV. 1998. Biofilm susceptibility to bacteriophage attack: 400 the role of phage-borne polysaccharide depolymerase. Microbiology 144 ( Pt 11):3039-3047. 401
35. Lenski RE. 1988. Experimental studies of pleiotropy and epistasis in Escherichia coli. II. 402 Compensation for maldaptive effects associated with resistance to virus T4. Evolution:433-403 440. 404
36. Lenski RE. 1988. Experimental studies of pleiotropy and epistasis in Escherichia coli. I. 405 Variation in competitive fitness among mutants resistant to virus T4. Evolution:425-432. 406
37. Luckey M, Nikaido H. 1980. Specificity of diffusion channels produced by lambda phage 407 receptor protein of Escherichia coli. Proc Natl Acad Sci U S A 77:167-171. 408
38. Harcombe WR, Bull JJ. 2005. Impact of phages on two-species bacterial communities. Appl 409 Environ Microbiol 71:5254-5259. 410
39. Kay MK, Erwin TC, McLean RJ, Aron GM. 2011. Bacteriophage ecology in Escherichia coli and 411 Pseudomonas aeruginosa mixed-biofilm communities. Appl Environ Microbiol 77:821-829. 412
40. Seed KD, Faruque SM, Mekalanos JJ, Calderwood SB, Qadri F, Camilli A. 2012. Phase 413 variable O antigen biosynthetic genes control expression of the major protective antigen and 414 bacteriophage receptor in Vibrio cholerae O1. PLoS Pathog 8:e1002917. 415
41. Kim M, Ryu S. 2011. Characterization of a T5-like coliphage, SPC35, and differential 416 development of resistance to SPC35 in Salmonella enterica serovar typhimurium and 417 Escherichia coli. Appl Environ Microbiol 77:2042-2050. 418
42. Kocharunchitt C, Ross T, McNeil D. 2009. Use of bacteriophages as biocontrol agents to 419 control Salmonella associated with seed sprouts. Int J Food Microbiol 128:453-459. 420

17
43. Sheng H, Knecht HJ, Kudva IT, Hovde CJ. 2006. Application of bacteriophages to control 421 intestinal Escherichia coli O157:H7 levels in ruminants. Appl Environ Microbiol 72:5359-5366. 422
44. Høyland-Kroghsbo NM, Mærkedahl RB, Svenningsen SL. 2013. A quorum-sensing-induced 423 bacteriophage defense mechanism. mBio 4. 424
45. Matsuzaki S, Tanaka S, Koga T, Kawata T. 1991. A broad-host-range vibriophage, KVP40, 425 isolated from sea water. Microbiol. Immunol. 36:93-97. 426
427

18
Figure Legends 428
Figure 1. Fluorescent microscopic images of micro-colony formation of strain BA35 (left two columns) 429 and PF430-3 (right two columns) at different time points in the absence and presence of phage 430 ΦH20 and KVP40, respectively. Samples were stained with 0.5% SYBR Gold for 10 min. 431
Figure 2. Culture density and biofilm formation of V. anguillarum strain BA35 and PF430-3 in cultures 432 pre-treated with phage ΦΗ20 and KVP40, respectively, and control cultures. A) Optical density 433 (OD600nm, left y-axis) of free-living cells (strain BA35) in the presence and absence of phage ΦΗ20, 434 and the corresponding free-living phage ΦΗ20 concentration (PFU ml-1, right y-axis); B) Strain BA35 435 biofilm formation in the presence and absence of phage ΦΗ20, quantified by crystal violet (OD595nm, 436 left y-axis); C) Optical density (OD600nm, left y-axis) of free-living cells (strain PF430-3) in the presence 437 and absence of phage KVP40 and the corresponding free-living phage KVP40 concentration of (PFU 438 ml-1, right y-axis) ; and D) Strain PF430-3 biofilm formation in the presence and absence of phage 439 KVP40, quantified by crystal violet (OD595nm, left y-axis). Error bars represent the actual ranges from 440 all experiments carried out in duplicate. 441
Figure 3. Culture density and biofilm formation of V. anguillarum strain BA35 and PF430-3 in 442 cultures post-treated with phage ΦΗ20 and KVP40 respectively, and control cultures. A) Optical 443 density of free-living cells (strain BA35) (OD600nm, left y-axis) in the presence and absence of phage 444 ΦΗ20, and the corresponding phage free-living ΦΗ20 concentration of (PFU ml-1); B) Strain BA35 445 biofilm formation in the presence and absence of phage ΦΗ20, quantified by crystal violet (OD595nm, 446 left y-axis) and corresponding biofilm-associated phage concentration (PFU ml-1, right y-axis); C) 447 Optical density (OD600nm, left y-axis) of free-living cells (strain PF430-3) in the presence and absence 448 of phage KVP40, and the corresponding free-living phage KVP40 concentration of (PFU ml-1, right y-449 axis); and D) Strain PF430-3 biofilm formation in the presence and absence of phage KVP40, 450 quantified by crystal violet (OD595nm, left y-axis) and the corresponding biofilm-associated phage 451 concentration of (PFU ml-1, right y-axis). Error bars represent the actual ranges from all experiments 452 carried out in duplicate. 453
Figure 4. Fluorescence microscopic examination of aggregate formation in V. angillarum strains BA35 454 and PF430-3 in the presence and absence of phages. No aggregate formation was observed in BA35 455 and BA35+ΦH20 cultures, and phage addition reduced the concentration of free-living cells relative 456 to the control . In the PF430+KVP40 cultures, phage-induced aggregates were observed. 457
Figure 5. Visualization of phage adsoprtion by SYBR Gold-labelled phages under phase-contrast 458 epifluorescence microscope. A, wild-type BA35 cells with SYBR Gold-labelled phage ΦH20 (phase-459 contrast+fluorescence); B, BA35 phage lysate with SYBR Gold-labelled phage ΦH20 (phase-460 contrast+fluorescence); C, PF430-3 phage lysate with SYBR Gold-labelled phage KVP40 (phase-461 constrast); and D, PF430-3 phage lysate with SYBR Gold-labelled phage KVP40 (phase-462 contrast+fluorescence). 463
464


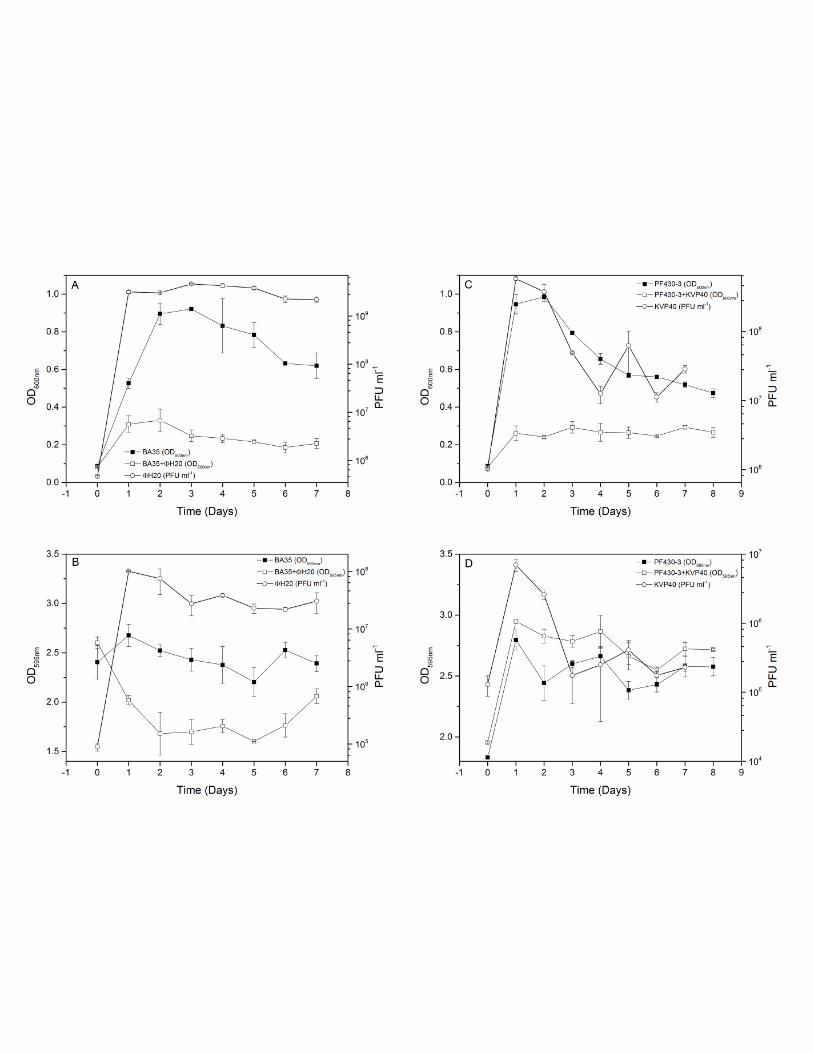



Table 1. Bacteriophages and hosts Phage Bacterial host (source) Phage receptor Genome Family Phage source KVP40 PF430-3 (Chile) (17) outer membrane
protein K (OmpK) 244 kb Myoviridae seawater, Japan (19, 20)
ΦH20 BA35 (USA) (18) unknown 50 kb Siphoviridae seawater, Denmark (21)
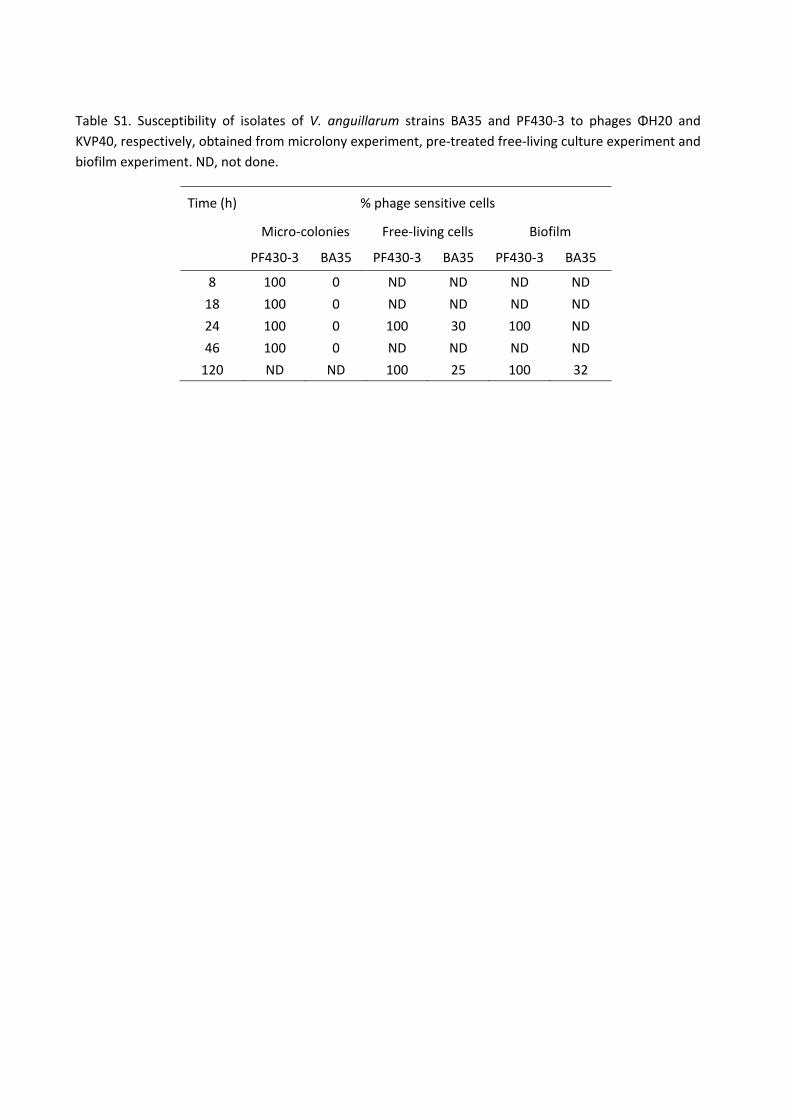
Table S1. Susceptibility of isolates of V. anguillarum strains BA35 and PF430-3 to phages ΦH20 and
KVP40, respectively, obtained from microlony experiment, pre-treated free-living culture experiment and
biofilm experiment. ND, not done.
Time (h) % phage sensitive cells
Micro-colonies Free-living cells Biofilm
PF430-3 BA35 PF430-3 BA35 PF430-3 BA35
8 100 0 ND ND ND ND
18 100 0 ND ND ND ND
24 100 0 100 30 100 ND
46 100 0 ND ND ND ND
120 ND ND 100 25 100 32
Supplementary Figure S1. Carbon substrate utilization of different V. anguillarum strains (BA35 and PF430-
3). BA35 WT, BA35 wild-type; BA35 R, BA35 resistant isolate; PF430-3 WT, PF430-3 wild-type; PF430-3 S,
PF430-3 Sensitive isolate; and PF430-3 L, PF430-3 bacteria and bacteriophage lysate.

1
Quorum sensing determines the choice of anti-phage defense strategy in Vibrio anguillarum 1
Demeng Tana, Sine Lo Svenningsenb, Mathias Middelboea* 2
3
aMarine Biological Section, bSection for Biomolecular Sciences, Department of Biology, University 4
of Copenhagen, Copenhagen, Denmark 5
*Correspondence: [email protected] 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21

2
Abstract 22
Selection for phage resistance is a key driver of bacterial diversity and evolution, and phage-host 23
interactions may therefore have strong influence on the genetic and functional dynamics of 24
bacterial communities. In this study, we found that an important, but so far largely overlooked, 25
determinant of the outcome of phage-bacterial encounters in the fish pathogen Vibrio 26
anguillarum is bacterial cell-cell communication, known as quorum sensing. Specifically, V. 27
anguillarum PF430-3 cells locked in the low-cell-density state (ΔvanT mutant) express high levels 28
of the phage receptor OmpK; resulting in a high susceptibility to phage KVP40, but achieve 29
protection from infection by enhanced biofilm formation. By contrast, cells locked in the high-cell-30
density state (ΔvanΟ mutant) are almost completely unsusceptible due to quorum-sensing-31
mediated down-regulation of OmpK expression. The phenotypes of the two quorum sensing 32
mutant strains are accurately reflected in the behavior of wild-type V. anguillarum, which (i) 33
displays increased OmpK expression in aggregated cells compared to free-living variants in the 34
same culture, (ii) displays a clear inverse correlation between ompK mRNA levels and the 35
concentration of N-acyl homoserine lactone quorum-sensing signals in the culture medium, and (iii) 36
survive mainly by one of these two defense mechanisms, rather than by genetic mutation to 37
phage resistance. Taken together, our results demonstrate that V. anguillarum employs quorum 38
sensing information to choose between two complementary anti-phage defense strategies. 39
Further, the prevalence of non-mutational defense mechanisms in strain PF430-3 suggests highly 40
flexible adaptations to KVP40 phage infection pressure, possibly allowing the long-term co-41
existence of phage and host. 42
43
44
45
46
47
48

3
Importance 49
Comprehensive knowledge on bacterial anti-phage strategies and their regulation is essential for 50
understanding the role of phages as drivers of bacterial evolution and diversity. In an applied 51
context, development of successful phage-based control of bacterial pathogens also requires 52
detailed understanding of the mechanisms of phage protection in pathogenic bacteria. Here we 53
demonstrate for the first time the presence of QS-regulated phage defense mechanisms in the fish 54
pathogen V. anguillarum and provide evidence that QS regulation allows V. anguillarum to 55
alternate between different phage protection mechanisms depending on population cell density. 56
Further, our results demonstrate the prevalence of non-mutational defense mechanisms in the 57
investigated V. anguillarum strain, which allow flexible adaptations to a dynamic phage infection 58
pressure. 59
60
61
62
63
64
65
66
67
68
69
70
71
72

4
Introduction 73
Vibrio anguillarum is a marine pathogenic bacterium which causes vibriosis in numerous fish and 74
shellfish species leading to high mortalities and economic losses in aquaculture worldwide (1). The 75
use of bacteriophages to control bacterial infections in aquaculture has gained increased attention 76
the past years and successful application of phages to reduce vibriosis-related mortality has been 77
demonstrated (2). Development of a phage-based treatment is, however, challenged by the wide 78
variety of anti-phage defense strategies observed in bacterial hosts (3). There is clear evidence 79
that genetic mutation, normally causing disruption or modification of phage receptors in the host 80
membrane, plays an important role in preventing phage infection in some cases (4-8). Such 81
genetic changes may impose a fitness cost to the host cell as disruption of phage receptors can 82
reduce the uptake of certain substrates (9-11). Alternative defense mechanisms which do not 83
involve mutational changes have been described and also play a role in V. anguillarum (Tan et al., 84
unpublished). Aggregate formation and production of exopolysaccharides was suggested to 85
provide protection against infection in V. anguilllarum strain PF430-3 (7, Tan et al., unpublished). 86
Recently, a more flexible protection mechanism was discovered in Escherichia coli, involving a 87
temporary down-regulation of phage receptor production in response to N-acyl-L-homoserine 88
lactone (AHL) cell-cell signaling molecules (12). This mechanism is controlled by quorum sensing 89
(QS), i.e., the ability of bacteria to regulate gene expression according to population density via 90
the production and subsequent detection of extracellular signaling molecules (13). 91
Little is still known about the mechanisms of phage protection in natural Vibrio communities. The 92
universal outer membrane protein K (OmpK) has previously been shown to be the infection site 93
for vibriophage KVP40, which infects more than 8 Vibrio species, including V. anguillarum, V. 94
parahaemolyticus, V. harveyi and V. cholerae (14, 15). Mutations in this protein have been 95
reported in V. parahaemolyticus R4000 upon exposure to KVP40 (14), but were not detected in 96
KVP40-amended cultures of V. anguillarum PF430-3 (7, Tan et al., unpublished). Thus, other 97
mechanisms for preventing infection by KVP40 in the Vibrio community may also be prevalent. 98
AHL-mediated QS circuits have been identified in many Vibrio species including V. anguillarum (13, 99
16, 17). V. anguillarum controls QS-regulated genes via the transcription factor VanT, which is 100
activated in response to extracellular signaling molecules (18, 19). At low autoinducer 101

5
concentrations (i.e. low cell density), the response regulator VanO becomes activated by 102
phosphorylation, and represses the expression of VanT. At high cell densities, autoinducer 103
concentrations increase and bind to membrane-bound receptors (18, 19). At a certain threshold 104
concentration, VanO is dephosphorylated, and VanT expression is induced, allowing gene 105
regulation within the QS regulon (18, 19). Several N-Acyl homoserine lactone autoinducers have 106
been identified in stationary-phase V. anguillarum spent culture supernatant (20). 107
As densities of bacterial populations vary from sparsely populated environments to highly dense 108
populations in nutrient-rich environments, and phages require a bacterial host in order to 109
multiply, phage predation pressure is not constant and may be expected to correlate with the 110
density of the bacterial host population, among other factors. Thus, bacteria could potentially 111
benefit from altering their anti-phage strategies depending on the perceived population density, 112
thereby minimizing the metabolic burden often associated with resistance by genetic mutation (3, 113
9, 21, 22). 114
In this study, we identified a potent anti-phage defense mechanism in V. anguillarum, and show 115
that different anti-phage strategies prevail at different population densities. At high cell density 116
conditions, QS-mediated down-regulation of the OmpK receptor reduced phage adsorption and 117
rendered individual cells almost unsusceptible to phage infection. At low cell density conditions, 118
on the other hand, OmpK expression was unaffected by QS and the individual cells were fully 119
susceptible to infection. However, at these conditions, we have shown in a previous study that 120
aggregation of the cells prevents phage from reaching the OmpK receptor (7, Tan et al., 121
unpublished). In neither case was phage protection associated with ompK mutation. Overall, the 122
study shows that QS controls the choice of anti-phage defense strategy in the examined V. 123
anguillarum strain PF430-3 suggesting the presence of dynamic, temporary adaptations to phage 124
infection pressure, while still securing the ability to produce a functional OmpK receptor. In a 125
phage therapy context, these results are highly relevant, as a combination of phage-based and 126
anti-QS targeted treatments could enhance the efficiency of phage control of vibriosis. 127
Results 128
We have recently demonstrated that addition of phage KVP40 to V. anguillarum strain PF430-3 129
resulted in increased cell aggregation and biofilm formation, which provided protection against 130

6
phage infection (7, Tan et al., unpublished). However, at high cell densities, detachment of cells 131
from aggregates into free-living variants were observed, which co-existed with phages (Tan et al., 132
unpublished), suggesting that alternative defense mechanisms also played a role under these 133
conditions. Isolation and subsequent re-culturing of these free-living cells in the absence of KVP40 134
showed that the re-cultured cells had regained sensitivity to phage KVP40 (7), suggesting that the 135
initial protection was not caused by mutational changes but rather a temporary protection 136
mechanism. To test if an extracellular factor might be involved in the regulation of the phage-137
induced aggregation phenotype, we examined the effect of adding cell-free spent culture fluid 138
from high-cell-density cultures of strain PF430-3 to freshly inoculated cultures of the same strain 139
in the presence or absence of phage KVP40 by phase-contrast microscopy (Fig. 1, left panels). 140
Intriguingly, the phage-induced aggregation phenotype of PF430-3 was completely inhibited by 141
the presence of the cell-free spent culture fluid, suggesting that an extracellular factor(s), possibly 142
a QS signaling molecule, is involved in the regulation of phage defense in V. anguillarum PF430-3. 143
Synthetic AHL-induced down-regulation of phage production 144
V. anguillarum have been shown to produce at least three different AHL QS autoinducers, namely 145
N-(3-hydroxyhexanoyl) homoserine lactone, N-(3-oxodecanoyl) homoserine lactone, and N-146
hexanoylhomoserine lactone (20). To further evaluate the role of QS for phage-host interactions in 147
V. anguillarum, the effect of synthetic AHL addition on phage KVP40 production was quantified in 148
strain PF430-3 (Fig. S2). The results showed that the presence of synthetic AHL autoinducers in the 149
medium reduced phage KVP40 production by 1.5 to 2-fold after 2 h relative to control cultures 150
without AHL addition (Fig. S2, squares). In parallel cultures grown without phage addition, cell 151
growth was unaffected by AHL addition (Fig. S2, circles). 152
KVP40 efficiency of plating on QS mutants of PF430-3 153
In order to examine the effects of QS on the interaction between phage KVP40 and V. anguillarum 154
PF430-3, two otherwise isogenic QS mutants were constructed which represent cell behavior at 155
high (ΔvanO) or low (ΔvanT) cell densities, respectively. Since the phosphorylated VanO protein 156
represses the QS regulator VanT at low cell densities, ΔvanO displays full expression of VanT at all 157
densities, and is therefore locked in a high-cell-density phenotype (18). By contrast, a ΔvanT 158
mutant has lost the ability to regulate QS-associated functions and is therefore locked in a low-159

7
cell-density phenotype. Use of the two extreme QS-phenotypes of ΔvanT and ΔvanO mutants in 160
the current study allowed us to explore the role of QS-mediated phage protection in V. 161
anguillarum PF430-3. We first tested the infectivity of phage KVP40 on the wild-type V. 162
angullarum PF430-3 strain, and the constructed mutants ΔvanT, ΔvanO, ΔompK, ΔvanT ΔompK, 163
and ΔvanO ΔompK by examining the efficiency of plating (EOP, the number of plaques obtained on 164
each host from a given phage input). Relative to the wild-type strain, the phage susceptibility of 165
the ΔvanT mutant had increased by 53% (p< 0.05, n=2), and the plaque morphology had changed 166
from turbid to clear (Fig. S3A). Additionally, the ΔvanO mutant had become less susceptible to 167
phage infection with a reduction of 37%, (p<0.01, n=2) relative to wild-type (Fig. S3B). As expected, 168
ΔompK, ΔvanT ΔompK, and ΔvanO ΔompK had all become resistant to KVP40 confirming that 169
ΟmpK is the phage receptor (Fig. S3Α). 170
Phage-host interactions in liquid cultures 171
Further assessments of QS-mediated regulation of phage infectivity in 10 h infection experiments 172
with phage KVP40 and 3 hosts (wild-type, ΔvanT, and ΔvanO), showed strong effects of the 173
mutational changes on phage-host interactions. Phage KVP40 reduced cell density in the wild-type 174
and ΔvanT cultures within 10 h incubation by 60% and 75%, respectively, relative to the OD of 175
control cultures without phages, with significantly lower OD values in the ΔvanT culture than in 176
the wild-type culture (0.62 and 0.88 after 10 h, respectively) (p<0.01, n=2; Fig. 2A). In the ΔvanO 177
culture, on the other hand, OD was not significantly affected by phage addition (Fig. 2A). In 178
accordance with these results, we observed rapid phage propagation in the wild-type and ΔvanT 179
cultures where KVP40 abundance stabilized at ~1010 PFU ml-1, whereas the phage production was 180
~100-fold lower in the ΔvanO culture (Fig. 2B). We note that the minimum OD values of 0.33 and 181
0.15 after 5 h in the phage-treated wild-type and ΔvanT cultures, respectively, was followed by a 182
regrowth of cells during the remainder of the incubation (Fig. 2A), despite the abundance of 183
KVP40 phage. 184
Aggregation of cells in the two QS mutants following phage KVP40 exposure was assessed by 185
phase-contrast microscopy (Fig. 1, center and right panels; see also Fig. S1). Similar to the wild-186
type shown in Figure 1, addition of phages led to increased aggregation in the ΔvanT mutant 187
cultures. In fact, at 24 h most cells occurred in large aggregates in the ΔvanT mutant cultures, 188

8
while a fraction of free-living cells was still detectable in the wild-type culture (Fig. 1 and Fig. S1). 189
In the ΔvanO culture, on the other hand, phage addition did not result in formation of aggregates 190
(Fig. 1 and Fig. S1), suggesting that the QS pathway is involved in the regulation of the phage-191
induced aggregation phenotype. Furthermore, the addition of cell-free spent culture fluid, which 192
prevented aggregation of the wild-type culture, did not prevent aggregation of the ΔvanT mutant 193
cultures upon exposure to phage, suggesting that the inhibitory factor present in the cell-free 194
culture fluid prevents aggregation via the QS pathway (Fig. 1). However, addition of the synthetic 195
AHL autoinducers did not mimic the inhibitory effect of adding cell-free spent culture fluid on 196
phage-induced aggregation in wild-type PF430-3 (data not shown). 197
Effects of phage KVP40 and QS on biofilm formation 198
Based on the indications of phage-driven stimulation of biofilm formation in the short term liquid 199
culture experiments, a more systematic quantification of the effects of phage KVP40 on biofilm 200
formation in the three strains was performed. In the control cultures without phages, the biofilm 201
formation of the wild-type and ΔvanT strains increased for 5 days and then remained constant at a 202
level of OD595nm = 1.5-1.75, as measured by crystal violet staining of the biofilm. In ΔvanO cultures, 203
the biofilm developed at a slower rate but reached the same end point as the other strains after 8 204
days (Fig. S4A). Pretreatment of the cultures with KVP40 enhanced biofilm formation in all 3 205
strains. The OD595nm in wild-type and ΔvanT cultures reached values of 2.8 after 3 days, whereas 206
OD595nm in the KVP40-treated ΔvanO cultures increased gradually during the incubation to a 207
maximum of 2.2 after 8 days (Fig. S4A). Phage concentration in the liquid above the biofilm 208
increased rapidly and stabilized around 109 PFU ml-1 in both wild-type and ΔvanT cultures after 1 209
day. In the ΔvanO+KVP40 cultures, phage concentration increased at a slower pace until day 3, 210
where it also stabilized at ~109 PFU ml-1 (Fig. S4B). 211
Together, the results from these experiments demonstrate a reduced susceptibility to phage 212
KVP40 in the ΔvanO mutant despite the fact that this mutant aggregates less and forms biofilm 213
more slowly than the wild type, suggesting that V. anguillarum PF430-3 may obtain protection 214
against KVP40 via a separate mechanism in the high-cell-density QS mode mimicked by the ΔvanO 215
mutant. 216
Phage adsorption rate 217

9
To address the question of how ΔvanO mutant cells reduce their susceptibility to KVP40, we first 218
measured the rate of adsorption of KVP40 to the wild-type and QS mutants host cells. The 219
adsorption rate of wild-type, ΔvanT, and ΔvanO was calculated as 6.7x10-10 ± 8.49x10-11 ml-1 min-1, 220
8.1x10-10 ± 9.2x10-11 ml-1 min-1, and 3.8×10-10 ± 1.4x10-11ml-1 min-1, respectively. Thus, the rate of 221
adsorption of phage KVP40 to ΔvanO was significantly lower than that observed for the wild-type 222
(p<0.04, n=2) and ΔvanT (p<0.02, n=2) strains. 223
ompK expression in cultures of wild type and QS mutant cells 224
To further examine the mechanism underlying the differences in adsorption rates among wild-225
type and QS-mutants (ΔvanT and ΔvanO), and to link these differences to QS regulation, we 226
quantified the expression of phage KVP40 receptor ompK in 9 different cultures (wild-type, ΔvanT, 227
ΔvanO, wild-type+KVP40, ΔvanT+KVP40, ΔvanO+KVP40, ΔompK, ΔvanT ΔompK, and ΔvanO ΔompK) 228
by quantitative real-time PCR (qPCR). The relative ompK expression levels of wild-type and ΔvanT 229
strains were approximately 4 times higher than the relative ompK expression in the ΔvanO strain 230
suggesting a down-regulation of ompK, when the cells are locked in the regulatory state mimicking 231
high cell densities (Fig. 3A). Purification and separation of outer membrane proteins by SDS-PAGE 232
(Fig. 3B) using three ompK mutants (ΔompK, ΔvanT ΔompK, and ΔvanO ΔompK) as negative 233
controls confirmed the presence of the OmpK receptor (26 kDa) in wild-type and ΔvanT strains 234
and the QS-mediated OmpK receptor down-regulation in the ΔvanO strain (14). In agreement with 235
the gene expression data, OmpK appeared to be slightly more abundant in the ΔvanT strain than 236
in the wild-type strain. SDS-PAGE outer membrane protein analysis also demonstrated that other 237
outer membrane proteins than OmpK, which potentially function as phage receptors, were down 238
regulated in the QS mutant ΔvanO as well (Fig. 3B, OmpK-indicated by arrowhead). 239
ompK mRNA levels in the free-living and aggregated cell fractions 240
We have shown above that the ΔvanT mutant displays increased aggregation, and contains high 241
levels of ompK, whereas the ΔvanO mutant displays very little aggregation, but instead reduces its 242
susceptibility to phage KVP40 by expressing low levels of OmpK. Next, we verified whether these 243
two phenotypes are also correlated in the wild-type PF430-3 strain. We fractionated cultures 244
(wild-type+KVP40, ΔvanT+KVP40 and ΔvanO+KVP40) into a free-living and an aggregate fraction 245
by centrifugation, and compared relative ompK mRNA levels in the two fractions using qPCR (Fig. 246

10
4). The analysis revealed that ompK gene expression varied significantly between aggregated and 247
free-living cells in the wild-type strain, with approximately 2-fold higher ompK mRNA levels in the 248
bacteria located in aggregates than in the fraction containing free-living cells (Fig. 4). Moreover, it 249
should be noted that the applied centrifugation procedure does not result in complete separation 250
of the aggregates from the free-living cells; hence the difference in ompK mRNA levels between 251
the two fractions is likely underestimated. Interestingly, ompK mRNA levels were also higher in the 252
aggregated fraction of the ΔvanO mutant cells than in the corresponding free-living fraction, 253
although the ompK levels in both fractions were still much reduced compared to the wild-type or 254
ΔvanT cultures. Thus, even in the absence of a functional QS pathway, free-living cells express less 255
ompK mRNA than aggregated cells, which could either reflect the existence of a redundant VanO-256
independent regulatory mechanism, or simply reflect a phage-mediated enrichment of cells with 257
low ompK mRNA expression among the surviving free-living cells. Unfortunately, we were unable 258
to collect free-living cells from ΔvanT cultures grown in the presence of phage, as any free-living 259
cells were lysed by the phage and the remaining cells were all aggregated. However, in agreement 260
with the data shown in Figure 3A (ompK mRNA levels in the presence of phage KVP40), ompK 261
mRNA levels were up-regulated in the ΔvanT aggregates even compared to the wild-type 262
aggregated fraction. 263
Temporal changes in ompK expression and AHLs production in wild type V. anguillarum PF430-3 264
In order to determine whether ompK expression was in fact repressed by a cell density-dependent 265
factor secreted by the V. angullarum PF430-3 wild-type strain, simultaneous measurements of 266
ompK expression and extracellular AHL levels were performed during growth of the wild-type 267
strain in batch culture in the absence of phages. Interestingly, phage receptor ompK expression 268
and AHL levels were significantly negatively correlated (r=0.811, p≤0.001, n=2) during the first 12 h. 269
Thus, in accordance with the previous experiment, ompK expression was 3-4 times higher at the 270
low cell density around 2 h, than at high-cell density at 6 h (Fig. 5). AHL production reached a 271
maximum at 6 h and decreased after 8 h, concomitant with an increase in ompK expression. After 272
16 h a sharp decline in ompK expression was observed. 273
Examination of potential fitness costs in ΔompK mutants 274

11
Throughout our studies of the interaction of phage KVP40 with V. anguillarum strain PF430-3, we 275
did not once observe a single case of resistance to phage infection due to mutation of ompK. By 276
contrast, cells that survived exposure to phage KVP40 always multiplied to produce offspring that 277
had regained phage sensitivity (7, Tan et al., unpublished), Therefore, we speculate that mutation 278
of ompK may be associated with a significant fitness cost. The physiological role of OmpK is not 279
known, but BIOLOG GN2 physiological fingerprints of the wild-type strain and the ΔompK mutant 280
showed that the ΔompK mutant lost the ability to utilize glucose-1-phosphate, glucose-6-281
phosphate, cis-aconitic acid, L-alaninamide, and L-alanine as growth substrates, suggesting that 282
indeed OmpK may play an important role for V. anguillarum metabolism (Fig. S5). We were, 283
however, unable to detect a change in the relative abundance of the ΔompK mutant compared to 284
wild-type after 8h of growth in mixed culture. Thus, loss of the OmpK receptor does not directly 285
affect the competitive ability of strain PF430-3 under our experimental conditions (Fig. S6). 286
Discussion 287
Given the recent findings that bacteria may employ QS to reduce phage receptor expression 288
during conditions of high infection risk (12), we aimed at exploring the role of QS in regulating 289
susceptibility to the broad-host-range phage KVP40 in V. anguillarum strain PF430-3 through 290
down-regulation of the phage receptor OmpK. The reduced susceptibility to phage KVP40 in V. 291
anguillarum strain PF430-3 after addition of cell-free supernatant or synthetic AHLs, provided the 292
first indications of a QS-regulated mechanism of phage protection. This was further supported by 293
our subsequent studies of phage susceptibility in the two constructed V. anguillarum QS mutants, 294
ΔvanT and ΔvanO. These results showed that phage susceptibility and adsorption was reduced in 295
the ΔvanO and enhanced in ΔvanT relative to the wild type, directly confirming that QS played a 296
key role in the protection against KVP40 infection. The >100–fold higher phage production and 3-297
fold lower bacterial density in cultures of ΔvanT relative to ΔvanO after phage addition thus 298
emphasized that QS signaling in the high-cell-density phenotype mediated an efficient protection 299
against phage infection. 300
Measurements of the ompK expression further resolved the underlying mechanisms of QS-301
mediated phage protection. Thus, the significant (4-fold) reduction in ompK expression in the 302
ΔvanO and the verification that an OmpK receptor deficient mutant (ΔompK) was resistant to 303

12
phage KVP40 provided direct evidence for QS-mediated down-regulation of ompK expression as an 304
important mechanism for protection against phage infection in V. anguillarum. 305
Interestingly, our results also showed that down-regulation of ompK expression was not the only 306
QS-regulated phage defense mechanism in the investigated V. anguillarum strain. Cell aggregation 307
and formation of a biofilm in response to phage addition in the wild-type strain suggested that the 308
transformation from a free-living life form to growth in a biofilm was also a mechanism of 309
protection against phage infection, as also supported by a previous study (Tan et al., unpublished). 310
The exact mechanism by which phages induce cell aggregation, however, remains to be discovered. 311
While the production of extracellular polymers by the host to create a physical barrier against 312
phage infection has been suggested previously (3, 23), the current results suggest that this 313
mechanism is QS-controlled in V. anguillarum. Addition of phage KVP40 strongly stimulated cell 314
aggregation and biofilm formation in the wild-type and low-cell-density mutant (ΔvanT), whereas 315
no aggregation was observed in the high-cell-density mutant (ΔvanO). Hence the data suggest that 316
cell aggregation and biofilm formation is an important phage defense mechanism at low cell 317
densities where QS regulation of ompK expression is inefficient. Consequently, we show that V. 318
anguillarum PF430-3 alternates between these two types of phage defense mechanisms and that 319
QS regulates the choice of strategy by up-regulating one mechanism (removal of OmpK receptor) 320
while down-regulating another mechanism (cell aggregation) at high cell densities. Whether the 321
overall reduction in OmpK receptors at high-cell-density conditions results in a general decrease in 322
the number of receptors per cell in the population, thus reducing the encounter rate between 323
phages and OmpK receptors, and/or if OmpK down-regulation generates a fraction of cells 324
completely without receptors (i.e. resistant cells) is not known. However, the presence of turbid 325
plaques in the ΔvanO mutant upon exposure to KVP40 in spot assays indicated that some of the 326
cells were, in fact, temporarily fully resistant to the phage. The presence of two distinct phage 327
defense mechanisms in wild-type cells was confirmed by the observation that ompK expression 328
was reduced in free-living cells relative to cells embedded in aggregates. Together with the 329
significant negative correlation between extracellular AHL concentration and ompK expression 330
during growth of the wild-type strain in batch cultures, this demonstrated the dynamic nature of 331
phage defense and emphasized that the obtained results were not restricted to the extreme 332
phenotypes of the QS mutants. Interestingly, ompK expression was in fact enhanced in phage-333

13
amended wild-type and ΔvanT cultures relative to control cultures without phages. OmpK is a 334
porin-like protein which has been suggested to be involved in bile salt resistance, as well as iron 335
acquisition (24). We show here that OmpK also plays a role for the ability of V. angulillarum to use 336
glucose-1-phosphate, glucose-6-phosphate, cis-aconitic acid, L-alaninamide, and L-alanine as 337
growth substrates. We speculate therefore that up-regulation of ompK in aggregated cells may be 338
an adaptation to stimulate nutrient uptake in an aggregate environment where supply of specific 339
nutrients may be limited, or for enhancing the bile salt resistance of the cell aggregates inside the 340
host. However, in the short-term competition experiment we carried out, the ΔompK mutant was 341
able to reach the same density as the wild-type strain in a mixed culture. Further studies are 342
needed to determine the physiological role(s) of OmpK and hence the cost of its repression or loss. 343
We note that in the wild-type cultures of V. anguillarum PF430-3, AHLs accumulate as the culture 344
grows to high cell densities, but they largely disappear later in stationary phase (Fig. 5). This 345
phenomenon has been observed previously in cultures of Yersinia pseudotuberculosis and 346
Pseudomonas aeruginosa, grown in LB medium, and was found to be caused by pH-dependent 347
lactonolysis (25). We found, however, that the pH of the marine broth cultures used here 348
remained stable around 7.6 for at least 13 hours (data not shown). Alternative explanations for 349
the disappearance of the AHLs in stationary phase are the possible production of an AHL lactonase 350
enzyme as reported in Bacillus spp. (26) or the uptake of AHLs for use as a source of energy, 351
carbon and nitrogen, as reported for Variovorax paradoxus (27). Additional studies are required to 352
determine the mechanism responsible for AHL removal in V. anguillarum strain PF430-3. 353
One surprising observation from the current study was that QS reduced biofilm formation (Fig. S4). 354
Biofilm formation is in many bacterial pathogens found to be up-regulated by QS and stimulated at 355
high cell density promoting virulence (28). This has also been observed in V. anguillarum (strain 356
NB10), where a low-cell-density mutant (ΔvanT) showed significantly lower biofilm formation than 357
the wild-type strain (29).The reason for this contrasting effect of QS on biofilm formation in PF430-358
3 is not clear. V. anguillarum strain NB10 showed limited susceptibility to phage KVP40 and 359
produced turbid plaques (data not shown), suggesting that KVP40 is not an important predator on 360
that strain; hence other anti-phage strategies may be favored. VanT expression is known to 361
regulate physiological responses required for survival and stress response (18), and in the 362

14
investigated strain PF430-3, this seems to include shifting from aggregation to ompK down 363
regulation in response to increased cell density. In addition, since total protease activity correlates 364
with VanT expression in V. anguillarum strain NB10 (29), it may be speculated that at high cell 365
densities, when VanT is fully expressed, VanT activates protease activity to promote the release of 366
cells from aggregates, in a manner similar to the action of the VanT homologue HapR in V. 367
cholerae, where QS also down-regulates biofilm formation (30, 31). By QS-mediated ompK gene 368
regulation, aggregate-associated bacteria may thus have evolved a mechanism of detaching a 369
subpopulation of free-living cells with reduced phage susceptibility which can survive in phage 370
dense environments, thus allowing further spreading of vibriosis infections. In any case, the results 371
support previous indications of large differences in phage defense strategies among different 372
strains of V. anguillarum (Tan et al., unpublished) and emphasize the need for further exploring 373
whether the mechanisms described here are a general phenomenon in bacteria or are limited to a 374
subset of V. anguillarum strains. 375
Phages are known to be key drivers of bacterial evolution by selecting for phage-resistant mutants, 376
and it has been proposed that phage-host interactions lead to either an arms race dynamics of 377
antagonistic host and phage co-evolution, or a fluctuating selection dynamics involving frequency-378
dependent selection for rare host and phage genotypes (32, 33). In this study, however, we found 379
evidence for an alternative to these scenarios, as the defense mechanisms identified do not 380
involve mutational changes or complete elimination of phage susceptible host cells. The described 381
anti-phage mechanisms in V. anguillarum thus represent more flexible adaptations to dynamic 382
changes in phage and host densities, adding to the complexity of phage-host co-evolutionary 383
interactions and phage-driven genetic and phenotypic changes in bacterial populations. Since 384
mutational changes often result in a loss of fitness for the host (9, 10), the mechanisms described 385
here represent a potential fitness advantage for the host. It may be speculated that the 386
development of this temporary protection mechanism in V. anguillarum PF430-3 is related to the 387
fact that the OmpK receptor is widely conserved among Vibrio and Photobacterium species, and 388
thus probably executes an important function(s) in the cell. It is likely, therefore, that mutations in 389
ompK could have significantly negative effects on host fitness in natural habitats, in which case 390
development of alternative phage-defense strategies that leave the ompK gene intact would be of 391
strong selective advantage. Further studies are needed, however, to confirm this hypothesis. 392

15
Successful application of phage therapy in the treatment of vibriosis requires detailed knowledge 393
of the phage-host interactions and the regulation of anti-phage strategies in Vibrio. Our results 394
add to the suite of known phage defense mechanisms and their regulation in marine bacteria and 395
further emphasizes that the complexity of phage-host interactions pose a challenge for future use 396
of phages in disease control. On the other hand, the evidence that QS regulates phage receptor 397
expression may potentially be used actively by quenching QS signaling, hence preventing receptor 398
down-regulation. In support of that, QS inhibitors have been shown to impede expression of 399
virulence factors (34) and recently the addition of modified T7 phages producing quorum-400
quenching enzymes have resulted in inhibition of biofilm formation, suggesting that the use of QS-401
inhibitors may be a promising strategy in future antimicrobial therapy (35). 402
Materials and Methods 403
Bacterial strains and bacteriophages 404
The bacterial strain V. anguillarum PF430-3 was originally isolated from salmonid aquaculture in 405
Chile (36), and phage KVP40 which infects PF430-3 is a broad-host-range phage originally isolated 406
from Japan (37) (Table S1). Basic characterization of phage KVP40 and its interactions with PF430-407
3 has been provided recently (7). All the constructed mutant strains derived from V. anguillarum 408
PF430-3 wild-type strain and plasmids used in this study are listed in supplementary Table S1. E. 409
coli S17-1 (λpir) was used as the donor strain for transfer of plasmid DNA into V. anguillarum 410
PF430-3 by conjugation (29, 38). Antibiotics were used at the following concentration: 100 μg ml-1 411
ampicillin, 25 μg ml-1 chloramphenicol (for E. coli), and 5 μg ml-1 chloramphenicol (for V. 412
anguillarum) (29, 38). 413
DNA manipulation and mutant construction 414
For mutant constructions, in-frame deletions were made in the vanT, vanO, and ompK genes by 415
allelic exchange as first described by Milton and Croxatto (29, 38). The upstream DNA sequence of 416
the vanT gene was amplified from wild-type V. anguillarum PF430-3 by PCR using primers vT1 and 417
vT2, which introduced a BglII site. The downstream DNA sequence of vanT was amplified using 418
primers vT3 and vT4, which introduced a SacI site. These two fragments, which contain a 15-nt 419
overlap of identical sequence, were used as template for a second PCR using primers vT1 and vT4. 420

16
The PCR product was digested with BglII and SacI and cloned into the BglII and SacI sites of pDM4, 421
creating pDM4vanT. In-frame deletion of vanT in PF430-3 was confirmed by PCR using primers vT1 422
and vT4 and subsequent gel electrophoresis. To construct plasmid pDM4vanO and pDM4ompK, 423
and vanO and ompK mutants, we used the same method described for vanT above. The primers 424
are listed in supplementary Table S2. 425
Examination of fitness loss in the ompK mutant 426
In an attempt to search for potential functions of phage receptor OmpK and hence implications of 427
its down regulation or loss, BIOLOG GN2 Micro-Plates (BIOLOG) containing 95 different carbon 428
sources were used to test the ability of wild-type strain and ΔompK mutant to use different 429
substrates following the manufacturer’s instructions. Further, the potential reduction in 430
competitive abilities of the ΔompK mutant was examined in competition assays. Briefly, wild-type 431
and ΔompK bacterial cells were mixed in a ratio of 1:1 in sterilized seawater and then quantified 432
over time. Subsamples were collected every 1-2 h for 8 h and diluted and plated on LB plates. 433
Subsequently, 20 colonies were picked from each time point and identified as either wild-type or 434
ΔompK by PCR with Primers 1340 F and 1340R listed in Table S2. 435
Effects of cell-free supernatant enrichment on KVP40 phage infection 436
Cell-free supernatant from wild-type V. anguillarum PF430-3 cultures was prepared from a mid-437
log-phase culture of V. anguillarum PF430-3 in Marine Broth (MB) , which was centrifuged 438
(10,000×g for 10 min), sterile filtered (0.2 μm, Millipore) and subsequently added to freshly 439
inoculated cultures of wild-type, ΔvanT and ΔvanO, with and without phage KVP40, for 440
examination of the potential effects of V. anguillarum PF430-3 -produced autoinducer molecules 441
on the lytic effects of the phage KVP40. Images were obtained after 24 h incubation. 442
Effects of synthetic AHLs on phage production in V. anguillarum PF430-3 wild type 443
Synthetic AHLs (obtained from Sigma and Nottingham University), N-hexanoylhomoserine lactone 444
(C6-HSL), N-(3-hydroxyhexanoyl)homoserine lactone (3-hydroxy-C6-HSL) and N-(3-445
Oxododecanoyl)-L-homoserine lactone (3-oxo-C10-HSL) were added to V. anguillarum PF430-3 to 446
examine their effects on phage susceptibility. Briefly, the AHLs were dissolved in acidified ethyl 447
acetate (0.1% v/v acetic acid), mixed and added to a glass test tube to a final concentration of 10 448

17
μM and placed at room temperature for complete evaporation of the solvent, as described 449
previously (12, 39). For the control cultures without AHLs, test tubes containing only ethyl acetate 450
were prepared in parallel. Overnight cultures of wild-type cells were pelleted and resuspended, 451
and aliquots of 100 μl cell suspension were transferred to 5 ml 3% NaCl buffer in tubes containing 452
AHLs or the solvent control, and incubated at 30 °C for 1 h. The phage susceptibility assay was 453
then initiated by addition of phage KVP40 at an average phage input (API) of 1. Parallel control 454
cultures without phages were also established. Samples were collected every hour for 7 h for 455
quantification of possible effects of AHL on cell growth in the absence of phage (colony forming 456
units, CFU) and on phage production (plaque forming units, PFU). 457
Phage-host interactions and effects on cell aggregation in wild-type and QS-mutants 458
The susceptibility of each strain (wild-type, ΔvanT, ΔvanO, ΔompK, ΔvanT ΔompK, and ΔvanO 459
ΔompK) to phage KVP40 was gauged by the efficiency of plating. Briefly, a fixed quantity of phage 460
obtained from the same phage stock (KVP40) was mixed with identically grown bacterial cells of 461
each genotype, and the plaque forming units were quantified by plaque assay (40). The 462
experiments were performed in triplicate. 463
The lytic potential of phage KVP40 against wild-type and QS-mutants (ΔvanT and ΔvanO) was 464
tested at an API of 1 in duplicate 100 ml liquid MB cultures and parallel control cultures without 465
phage. The effect of phage-mediated lysis on host cell density was monitored by regular OD600nm 466
measurement over the 10 h incubation. Phage concentration was quantified by plaque assay 467
periodically (40). 468
For visual inspection of cell densities and aggregate formation, additional aliqouts were collected 469
at 24 h, and visualized by phase-contrast microscopy using an oil immersion objective (Olympus 470
BX61). The images shown in Figure 1 and Figure S1 represent random fields on the microscope 471
slide from the 24 h samples. 472
Effects of phage addition on biofilm formation in wild-type and QS-mutants 473
Biofilm formation was monitored in wild-type, ΔvanT, and ΔvanO cultures following addition of 474
phage KVP40 using methods described previously (41) with some modifications. Briefly, 13 ml 475
polypropylene plastic tubes filled with 5 ml MB were inoculated with 100 μl overnight bacterial 476

18
inoculum and 100 μl phage stock (108 CFU ml-1 and 106 PFU ml-1, API=0.01) and incubated without 477
shaking along with parallel control cultures without phages. For each experiment, duplicate tubes 478
were examined daily for quantification of phage abundance in the liquid phase (40) and biofilm 479
biomass was quantified by crystal violet staining using a standard protocol (42). For biofilm 480
quantification, the liquid was removed, and tubes were rinsed twice with artificial seawater (ASW, 481
Sigma). The biofilm was stained with 0.4% crystal violet (Sigma) for 15 min and the tubes were 482
washed with tap water to remove excess stain. An aliquot (6 ml) of 33% acetic acid (Sigma) was 483
added and left for 5 min to allow the stain to dissolve. The absorbance was measured at OD595nm. 484
Bacteriophage adsorption rate 485
The rates of adsorption of phage KVP40 to wild-type, ΔvanT, and ΔvanO cells were determined by 486
mixing phage KVP40 and exponentially growing host cultures at an API of 0.01 and incubating at 487
room temperature for 32 min with agitation. Aliquots were removed periodically, centrifuged at 488
18,000×g at 4°C for 2 min, and the supernatants were immediately diluted to prevent additional 489
adsorption. The un-adsorbed phage particles were quantified by plaque assay (40). 490
Outer membrane protein K (OmpK) preparation and SDS-PAGE analysis 491
OMP preparation was made essentially according to Wang (43) with some modifications. Briefly, 492
cells were pelleted from 25 ml overnight cultures of all the strains (wild-type, ΔvanT, ΔvanO, 493
ΔompK, ΔvanT ΔompK, and ΔvanO ΔompK), resuspended in 4.5 ml water, and sonicated on ice 494
(100 amplitude, 3 min). To solubilize the cytoplasmic membranes, sarkosyl (N-lauroyl sarcosine) 495
was added to a final concentration of 2%, and incubated at room temperature for 30 min. To 496
pellet the outer membranes, the mixture was ultra-centrifuged (400,000 rpm for 1 h at 4 °C, SW 497
55 Ti, Beckman). The pellet was washed with ice-cold water and ultra-centrifuged again (400,000 498
rpm for 30 min at 4 °C, SW 55 Ti, Beckman). The pellet was resuspended in 100 μl 100 mM Tris-HCl 499
(pH 8) and 2% SDS buffer, and the proteins were separated on a 10% SDS-polyacrylamide gel and 500
stained with Coomassie blue. 501
Fractionation of V. anguillarum cultures in aggregate and free-living cell fractions 502

19
In order to examine ompK gene expression patterns in aggregates and free-living cells, respectively, 503
cultures were fractionated by centrifugation and ompK expression was determined in the 504
aggregate and non-aggregate fractions. Cultures of V. anguillarum wild-type and QS mutants were 505
grown in MB in the presence of phage KVP40 at an API of 1 at RT for 12 h, and the aggregates 506
were collected by centrifugation (1,000×g, 10 min). The supernatant was transferred to a new 507
tube and centrifugation was repeated. After the second round of centrifugation, the free-living 508
cells in the supernatant was transferred to a new tube and collected by centrifugation at 10,000×g 509
for 10 min. Pellets of cell aggregates and free-living cells were collected and stored at -80 °C for 510
subsequent RNA extraction. 511
AHL signal levels in cultures of wild-type V. anguillarum PF430-3. 512
An overnight culture of the wild-type strain was diluted in MB to the final OD600nm of 0.04 and 513
grown at 30 °C for 20 h with agitation. Samples were taken every 2 h for determination of AHL 514
production and for extraction of RNA for ompK mRNA quantification. AHL biosynthesis was 515
assayed using E. coli SP436 reporter strain (44). E. coli SP436 harbors a plasmid-borne fusion of the 516
V. fischeri lux operon to gfp, and responds to AHL by expressing green fluorescence protein (GFP) 517
as described previously (44). Briefly, 50 μl cell-free supernatant was mixed with 150 μl LB medium 518
and 10 μl of an overnight culture of SP436 (100 μg ml-1 ampicillin), and further incubated at 30 °C 519
for 5 h with agitation using a FLUOstar Omega microtiter plate reader (BMG Labtech) with gain 520
setting of 1150. 521
RNA extraction and cDNA synthesis 522
For quantification of ompK expression in the wild-type and QS mutants (ΔvanT and ΔvanO), RNA 523
was extracted from cells pelleted after 9 h incubation in MB for the experiments shown in Figure 5, 524
or as described above for the experiments shown in Figure 3 and 4. Total RNA was extracted using 525
Trizol (Invitrogen) and chloroform extraction as described previously (45). Genomic DNA was 526
removed by adding DNase I according to the manufacturer’s protocol (Fermentas). RNA was 527
stored at -80 °C until further use. 528
Reverse transcription was performed with the Thermo Scientific Revert Aid First Strand cDNA 529
Synthesis Kit as described by the manufacturer (Thermo). The first cDNA strand was obtained 530

20
using random hexamer primers with 1000 ng of DNase I-treated RNA. Control samples (-RT) were 531
treated identically except that the reverse transcriptase enzyme was omitted. 532
Real-time PCR gene expression quantification 533
The relative expression levels of ompK and recA (as the endogenous control) were determined by 534
quantitative RT-PCR performed in a CFX96 real-time PCR detection system (Bio-Rad), using 535
SsoAdvanced SYBR Green Supermix (Bio-Rad), and the gene-specific primers listed in 536
supplementary Table S2 (46). qPCR reactions were set up in 25 μl containing 0.2 μM of primers, 537
and SYBR Green qPCR mix with the following program, 30 s at 95 °C for denaturation, followed by 538
40 cycles of (4 s at 95 °C, 30 s at 55°C). Melting curve analysis was performed from 65 °C to 95 °C 539
with an increment of 0.5 °C for 5 s. The comparative CT method was used for relative 540
quantification of RNA (47). 541
Acknowledgements 542
The study was supported by the Danish Council for Strategic Research (ProAqua project 12-132390) 543
and the EU-IRSES-funded project AQUAPHAGE (269175). We thank Debra Milton, Umeå University, 544
for providing plasmid pDM4 and E. coli S1-17 and Pantelis Katharios, Hellenic Centre for Marine 545
Research, for providing V. anguillarum PF430-3 and vibriophage KVP40. We are grateful to Nina 546
Molin Høyland-Kroghsbo for many helpful discussions. 547
References 548
1. Frans I, Michiels CW, Bossier P, Willems KA, Lievens B, Rediers H. 2011. Vibrio anguillarum as a 549 fish pathogen: virulence factors, diagnosis and prevention. J Fish Dis 34:643-661. 550
2. Vinod M, Shivu M, Umesha K, Rajeeva B, Krohn G, Karunasagar I, Karunasagar I. 2006. Isolation of 551 Vibrio harveyi bacteriophage with a potential for biocontrol of luminous vibriosis in hatchery 552 environments. Aquaculture 255:117-124. 553
3. Labrie SJ, Samson JE, Moineau S. 2010. Bacteriophage resistance mechanisms. Nat Rev Microbiol 554 8:317-327. 555
4. Luria SE, Delbruck M. 1943. Mutations of Bacteria from Virus Sensitivity to Virus Resistance. 556 Genetics 28:491-511. 557
5. Morona R, Klose M, Henning U. 1984. Escherichia coli K-12 outer membrane protein (OmpA) as a 558 bacteriophage receptor: analysis of mutant genes expressing altered proteins. J Bacteriol 159:570-559 578. 560
6. Bassford PJ, Jr., Diedrich DL, Schnaitman CL, Reeves P. 1977. Outer membrane proteins of 561 Escherichia coli. VI. Protein alteration in bacteriophage-resistant mutants. J Bacteriol 131:608-622. 562
7. Tan D, Gram L, Middelboe M. 2014. Vibriophages and their interactions with the fish pathogen 563 Vibrio anguillarum. Appl Environ Microbiol 80:3128-3140. 564

21
8. Guglielmotti DM, Reinheimer JA, Binetti AG, Giraffa G, Carminati D, Quiberoni A. 2006. 565 Characterization of spontaneous phage-resistant derivatives of Lactobacillus delbrueckii 566 commercial strains. Int J Food Microbiol 111:126-133. 567
9. Lenski RE. 1988. Experimental studies of pleiotropy and epistasis in Escherichia coli. I. Variation in 568 competitive fitness among mutants resistant to virus T4. Evolution:425-432. 569
10. Lenski RE, Levin BR. 1985. Constraints on the Coevolution of Bacteria and Virulent Phage - a Model, 570 Some Experiments, and Predictions for Natural Communities. American Naturalist 125:585-602. 571
11. Szmelcman S, Hofnung M. 1975. Maltose transport in Escherichia coli K-12: involvement of the 572 bacteriophage lambda receptor. J Bacteriol 124:112-118. 573
12. Hoyland-Kroghsbo NM, Maerkedahl RB, Svenningsen SL. 2013. A quorum-sensing-induced 574 bacteriophage defense mechanism. MBio 4:e00362-00312. 575
13. Miller MB, Bassler BL. 2001. Quorum sensing in bacteria. Annu Rev Microbiol 55:165-199. 576 14. Inoue T, Matsuzaki S, Tanaka S. 1995. A 26-kDa outer membrane protein, OmpK, common to 577
Vibrio species is the receptor for a broad-host-range vibriophage, KVP40. FEMS Microbiol. Lett. 578 125:101-105. 579
15. Inoue T, Matsuzaki S, Tanaka S. 1995. Cloning and sequence analysis of Vibrio parahaemolyticus 580 ompK gene encoding a 26-kDa outer membrane protein, OmpK, that serves as receptor for a broad-581 host-range vibriophage, KVP40. FEMS Microbiol. Lett. 134:245-249. 582
16. Bassler BL, Greenberg EP, Stevens AM. 1997. Cross-species induction of luminescence in the 583 quorum-sensing bacterium Vibrio harveyi. J Bacteriol 179:4043-4045. 584
17. Milton DL, Hardman A, Camara M, Chhabra SR, Bycroft BW, Stewart GS, Williams P. 1997. 585 Quorum sensing in Vibrio anguillarum: characterization of the vanI/vanR locus and identification of 586 the autoinducer N-(3-oxodecanoyl)-L-homoserine lactone. J Bacteriol 179:3004-3012. 587
18. Weber B, Lindell K, El Qaidi S, Hjerde E, Willassen NP, Milton DL. 2011. The phosphotransferase 588 VanU represses expression of four qrr genes antagonizing VanO-mediated quorum-sensing 589 regulation in Vibrio anguillarum. Microbiology 157:3324-3339. 590
19. Croxatto A, Pride J, Hardman A, Williams P, Camara M, Milton DL. 2004. A distinctive dual-channel 591 quorum-sensing system operates in Vibrio anguillarum. Mol Microbiol 52:1677-1689. 592
20. Milton DL, Chalker VJ, Kirke D, Hardman A, Camara M, Williams P. 2001. The LuxM homologue 593 VanM from Vibrio anguillarum directs the synthesis of N-(3-hydroxyhexanoyl)homoserine lactone 594 and N-hexanoylhomoserine lactone. J Bacteriol 183:3537-3547. 595
21. Lenski RE. 1988. Experimental studies of pleiotropy and epistasis in Escherichia coli. II. 596 Compensation for maldaptive effects associated with resistance to virus T4. Evolution:433-440. 597
22. Middelboe M, Holmfeldt K, Riemann L, Nybroe O, Haaber J. 2009. Bacteriophages drive strain 598 diversification in a marine Flavobacterium: implications for phage resistance and physiological 599 properties. Environ Microbiol 11:1971-1982. 600
23. Heilmann S, Sneppen K, Krishna S. 2012. Coexistence of phage and bacteria on the boundary of 601 self-organized refuges. Proc Natl Acad Sci U S A 109:12828-12833. 602
24. Hamod MA, Bhowmick PP, Shukur YN, Karunasagar I, Karunasagar I. 2014. Iron shortage and bile 603 salts play a major role in the expression of ompK gene in Vibrio anguillarum. Pol J Microbiol 63:27-604 31. 605
25. Yates EA, Philipp B, Buckley C, Atkinson S, Chhabra SR, Sockett RE, Goldner M, Dessaux Y, Camara 606 M, Smith H, Williams P. 2002. N-acylhomoserine lactones undergo lactonolysis in a pH-, 607 temperature-, and acyl chain length-dependent manner during growth of Yersinia 608 pseudotuberculosis and Pseudomonas aeruginosa. Infect Immun 70:5635-5646. 609
26. Dong YH, Gusti AR, Zhang Q, Xu JL, Zhang LH. 2002. Identification of quorum-quenching N-acyl 610 homoserine lactonases from Bacillus species. Appl Environ Microbiol 68:1754-1759. 611
27. Leadbetter JR, Greenberg E. 2000. Metabolism of acyl-homoserine lactone quorum-sensing signals 612 by Variovorax paradoxus. J Bacteriol 182:6921-6926. 613

22
28. Davies DG, Parsek MR, Pearson JP, Iglewski BH, Costerton JW, Greenberg EP. 1998. The 614 involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 280:295-298. 615
29. Croxatto A, Chalker VJ, Lauritz J, Jass J, Hardman A, Williams P, Camara M, Milton DL. 2002. VanT, 616 a homologue of Vibrio harveyi LuxR, regulates serine, metalloprotease, pigment, and biofilm 617 production in Vibrio anguillarum. J Bacteriol 184:1617-1629. 618
30. Jobling MG, Holmes RK. 1997. Characterization of hapR, a positive regulator of the Vibrio cholerae 619 HA/protease gene hap, and its identification as a functional homologue of the Vibrio harveyi luxR 620 gene. Mol Microbiol 26:1023-1034. 621
31. Hammer BK, Bassler BL. 2003. Quorum sensing controls biofilm formation in Vibrio cholerae. Mol 622 Microbiol 50:101-104. 623
32. Martiny JB, Riemann L, Marston MF, Middelboe M. 2014. Antagonistic coevolution of marine 624 planktonic viruses and their hosts. Ann Rev Mar Sci 6:393-414. 625
33. Koskella B, Brockhurst MA. 2014. Bacteria–phage coevolution as a driver of ecological and 626 evolutionary processes in microbial communities. FEMS microbiology reviews. 627
34. O'Loughlin CT, Miller LC, Siryaporn A, Drescher K, Semmelhack MF, Bassler BL. 2013. A quorum-628 sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proc Natl Acad 629 Sci U S A 110:17981-17986. 630
35. Pei R, Lamas-Samanamud GR. 2014. Inhibition of biofilm formation by t7 bacteriophages producing 631 quorum-quenching enzymes. Appl Environ Microbiol 80:5340-5348. 632
36. Silva-Rubio A, Avendano-Herrera R, Jaureguiberry B, Toranzo AE, Magarinos B. 2008. First 633 description of serotype O3 in Vibrio anguillarum strains isolated from salmonids in Chile. J Fish Dis 634 31:235-239. 635
37. Matsuzaki S, Tanaka S, Koga T, Kawata T. 1992. A broad-host-range vibriophage, KVP40, isolated 636 from sea water. Microbiol Immunol 36:93-97. 637
38. Milton DL, OToole R, Horstedt P, WolfWatz H. 1996. Flagellin A is essential for the virulence of 638 Vibrio anguillarum. J Bacteriol 178:1310-1319. 639
39. Ghosh D, Roy K, Williamson KE, Srinivasiah S, Wommack KE, Radosevich M. 2009. Acyl-640 homoserine lactones can induce virus production in lysogenic bacteria: an alternative paradigm for 641 prophage induction. Appl Environ Microbiol 75:7142-7152. 642
40. Adams MH. 1959. Bacteriophages. Interscience Publishers, New York, N.Y. 643 41. Hosseinidoust Z, Tufenkji N, van de Ven TG. 2013. Formation of biofilms under phage predation: 644
considerations concerning a biofilm increase. Biofouling 29:457-468. 645 42. O'Toole GA, Pratt LA, Watnick PI, Newman DK, Weaver VB, Kolter R. 1999. Genetic approaches to 646
study of biofilms. Methods Enzymol 310:91-109. 647 43. Wang S-Y, Lauritz J, Jass J, Milton DL. 2003. Role for the major outer-membrane protein from 648
Vibrio anguillarum in bile resistance and biofilm formation. Microbiology 149:1061-1071. 649 44. Burmolle M, Hansen LH, Oregaard G, Sorensen SJ. 2003. Presence of N-acyl homoserine lactones 650
in soil detected by a whole-cell biosensor and flow cytometry. Microb Ecol 45:226-236. 651 45. Svenningsen SL, Waters CM, Bassler BL. 2008. A negative feedback loop involving small RNAs 652
accelerates Vibrio cholerae's transition out of quorum-sensing mode. Genes Dev 22:226-238. 653 46. Crisafi F, Denaro R, Genovese M, Yakimov M, Genovese L. 2013. Application of relative real-time 654
PCR to detect differential expression of virulence genes in Vibrio anguillarum under standard and 655 stressed growth conditions. J Fish Dis. 656
47. Schmittgen TD, Livak KJ. 2008. Analyzing real-time PCR data by the comparative CT method. Nature 657 protocols 3:1101-1108. 658
659
660

23
661
662
663
Figure Legends 664
Figure 1. Visualization by phase-contrast microscopy of V. anguillarum strains PF430-3 (wild-type, 665 ΔvanT and ΔvanO) in the presence or absence of phage KVP40 in either fresh Marine Broth or cell-666 free spent culture fluid obtained from a mid-log phase culture of wild-type V. anguillarum PF430-3. 667
Figure 2. (A) Optical density (OD600nm) of cultures of V. anguillarum wild-type and QS mutants 668 (ΔvanT and ΔvanO) in the presence or absence of phage KVP40 at API of 1 were measured at 1 h 669 intervals over a 10-h period incubation. (B) Corresponding abundance of PFU ml-1 were quantified 670 by plaque assay over a 10-h period incubation in wild-type+KVP40, ΔvanT+KVP40 and 671 ΔvanO+KVP40 cultures, respectively. Error bars represent standard deviations from all 672 experiments carried out in duplicate. 673
Figure 3. (A) ompK gene expression of wild type and QS mutants (ΔvanT and ΔvanO) in the 674 presence or absence of phage KVP40. (B) SDS-PAGE analysis of the outer membrane protein K 675 (OmpK) with Coomassie blue staining. Lane M, protein marker is shown on the right; lane 1-6, 676 wild-type, ΔvanT, ΔvanO, ΔompK, ΔompK ΔvanT, and ΔompK ΔvanO, respectvely. Phage receptor 677 ompK mRNA transcript levels were quantified by real-time PCR. “Relative gene expression” 678 corresponds to the level of ompK mRNA after normalization to the level of recA mRNA in the same 679 sample. Error bars represent standard deviations (duplicate samples). 680
Figure 4. Fractionation experiment. For bacterial aggregates (A), samples were harvested at 681 1,000×g for 10 min. For free-living bacteria (F), samples were harvested at 10,000×g for 10 min. 682 Phage receptor ompK mRNA transcript levels were quantified by real-time PCR. Relative gene 683 expression was normalized against the gene expression level of recA. Error bars 684 represent standard deviations (duplicate samples). 685
Figure 5. AHL assay and ompK expression quantification in the wild-type strain over time. Aliquots 686 of cultures were withdrawn every 2 h during growth for measurement of AHL production and 687 ompK gene expression by RT-PCR as described in the legend to Figure 3. Error bars 688 represent standard deviations (duplicate samples). 689

Figure 1

Figure 2

Figure 3
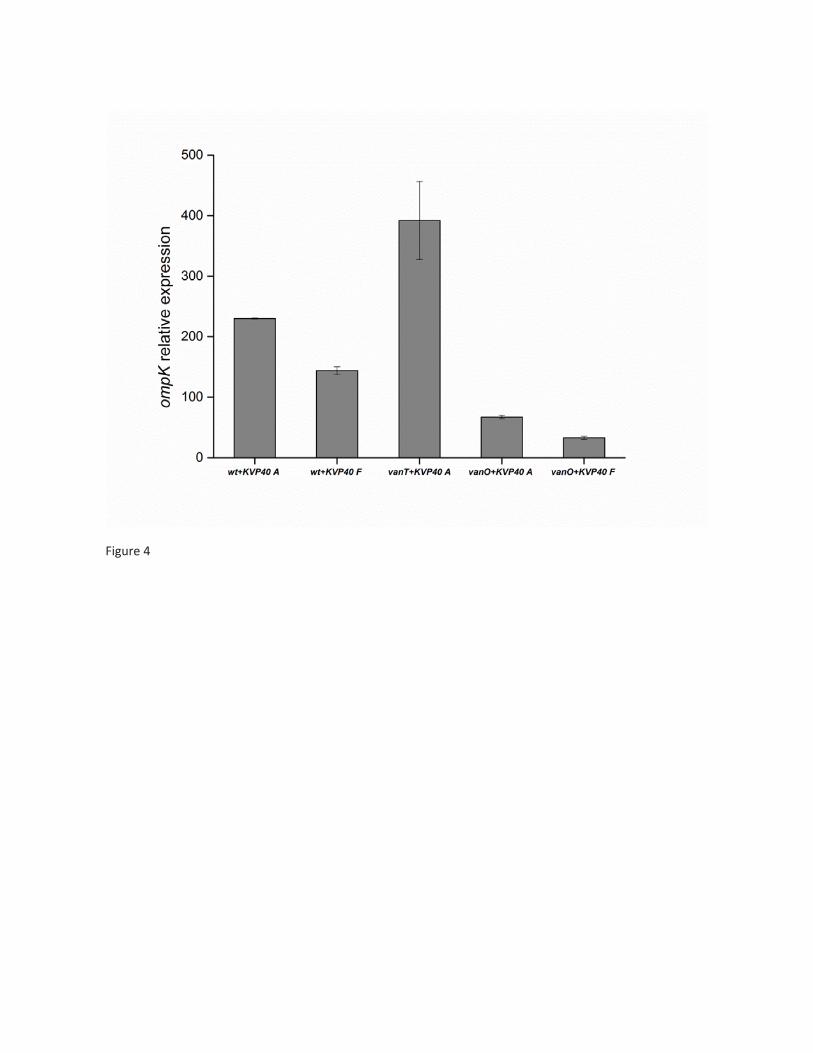
Figure 4

Figure 5

TABLE S1. Bacterial strains, bacteriophage and plasmids
Strains, phage or plasmids Genotype or relevant markers
E. coli S17-1 thi pro hsdR hsdM+recA RP4-2-Tc::Mu-Km::Tn7 λpir SY327 Δ(lac-pro) argE(Am) rif malA recA56 λpir SP436 Apr
V. anguillarum PF430-3 wt, isolated from Chile ΔvanT ΔvanT ΔvanO ΔvanO ΔompK ΔompK
ΔompK ΔvanT ΔompK ΔvanT ΔompK ΔvanT ΔompK ΔvanT
Plasmids
pDM4 Cmr; suicide vector with an R6K origin (pir requiring) and sacBR genes from
Bacillus subtilis pDTvanT Cmr; pDM4 derivative containing vanT fused in-frame pDTvanO Cmr; pDM4 derivative containing vanO fused in-frame
pDT ompK Cmr; pDM4 derivative containing ompK fused in-frame
pAHL-GFP Apr, luxR-PluxI-gfpmut3 Phage KVP40 Genome 244 kb, Myoviridae, isolated from Japan

Table S2 Primers
Name Primers (5'-3) qPCR (gene)
recA F-CTAGTCGAAATTTTTTCGACACAGC R-GGCTCGTTAAATTTTTTATCGACTCTT
ompK F-TAGAGATGGAATTTGGCGG R-TTTGTCACTGCTTGGGTT
vanT deletion vT1 F-TTTAGATCTCATCACTTCAACTGTATGTTTCATACTGTTC vT2 R-AAGTTAAGCCTAGTGCATGAGTTGTTAATCCTTGCCAATGAC vT3 F-CACTAGGCTTAACTTCCTAGTCTC vT4 R-TTTGAGCTCCGGTTTGGCTATCGAGATGG
vanO deletion vO1 F-TTTAGATCTGAGATCGCCAAACTAGAAGGG vO2 R-TCTCTATGAGTTCTCGTCAGGTTGCATATTCTAGCCATT vO3 F-TTTGAGCTCCGGTTTGGCTATCGAGATGG Vo4 R-TTTGAGCTCCGGTTTGGCTATCGAGATGG
ompK deletion oK1 F-TTTAGATCTGTAAGCCCCTAATTGAGT oK2 R-TAGAGAGTCCCCTTTTAATCGAACCTAAGTTTTGGCAAATG oK3 F-AAAGGGGACTCTCTATGTTTTGTCATAATTC oK4 R-TTTGAGCTCACAATCTTAACCAACATCCC
1340F F-TCGCCCAAAGCAACCCAA 1340R R-CCGTAGCTTCATTTTCCCTT

Figure S1. Examination of phage-host interactions (wild-type+KVP40, ΔvanT+KVP40, and ΔvanO+KVP40) in marine broth under phase-contrast microscopy after 24 h incubation. Images represent random fields obtained from microscopic slides prepared after 24 h incubation of the indicated V. anguillarum strain with phage KVP40.

Figure S2. Phage KVP40 production and bacterial growth upon exposure to synthetic AHLs. Shown are phage concentrations in the cell-free supernatant of cultures incubated with (closed symbols) or without (open symbols) 10 μm AHLs over the course of 7 h (squares). AHLs were added at time = -1h, and phage were added at an API of 1 at time = 0 h. To determine whether the AHLs affected bacterial growth in the absence of phage, the number of colony forming units (CFU) in parallel cultures +/- AHL but without phage addition were quantified (circles). Error bars represent standard deviations of experiments carried out in duplicate.

Figure S3. (A) Spot assay for testing host specificity of phage KVP40 against wild type, QS mutants (ΔvanT and ΔvanO) and ompK mutants (ΔompK, ΔvanT ΔompK, and ΔvanO ΔompK) as well. Five μl serial 100-fold dilutions of phage KVP40 lysate (1, 109; 2, 107; 3, 105; 4; 103 PFU ml-1) were shown by spot titration onto top agar lawns of the indicated strains. (B) Plaque assay quantification by using wild type and QS-mutants (ΔvanT and ΔvanO) as host, respectively. Error bars represent standard deviations (duplicate samples).

Figure S4. (A) Biofilm formation in V. anguillarum wild-type and QS mutants (ΔvanT and ΔvanO) pre-treated with phage KVP40 (open symbols) and in controls without phage KVP40 addition (closed symbols) quantified by crystal violet (OD595nm) staining during 8 days incubation. (B) Viable phage KVP40 recovered from free-living phases of wt+KVP40, ΔvanT+KVP40 and ΔvanO+KVP40, respectively, were quantified by plaque assay for 7 days. Error bars represent standard deviations from all experiments carried out in duplicate. It should be noted that the quantification of KVP40 most likely represent an underestimation of the actual phage production as only detached phage particles were counted. It is likely therefore, that the actual phage concentration was higher in the wild-type and ΔvanT cultures than in the ΔvanO cultures as more phages were likely trapped in the more developed biofilm of those cultures.

Figure S5. Carbon substrate utilization (BIOLOG GN2) between wild-type strain and ΔompK mutant. +, positive reaction at 24 h; -, negative reaction after 48 h.
C-source
wild-type
Δompk
Bromosuccinic Acid + +L-Histidine + +D-Fructose + +
β-Methyl- D-Glucoside + +Inosine + +Dextrin + +
D-Psicose + +Citric Acid + +
Uridine + +Thymidine + +Tween 40 + +Tween 80 + +α-D-Glucose + +D-Sorbitol + +
D,L-Lactic Acid + +L-Proline + +Sucrose + +
D-Gluconic Acid + +N-Acetyl-DGlucosamine + +
α-D-Lactose + +D-Trehalose + +L-Asparagine + +
L-Aspartic Acid + +L-Serine + +Maltose + +
L-Glutamic Acid + +D-Mannitol + +
D,L-α-Glycerol Phosphate + +D-Cellobiose + +D-Mannose + +
Mono-Methyl-Succinate + +Succinic Acid + +
Cis-Aconitic Acid + -Glucose-6-Phosphate + -
L-Alaninaide + -L-Alanine + -
Glucose-1-Phosphate + -D-Galactose + +
Glycerol + +D,L-α-Glycerol Phosphate + +
Hydroxy-L-Proline + +α-Keto Glutaric Acid + +
D-Alanine + +L-Alanylglycine + +
L-Threonine + +Propionic Acid + +
Acetic Acid + +Urocanic Acid + +

Figure S6. Competition assay between wild-type (balck) and ΔompK (grey) at a ratio of 1 was performed. Colonies were picked at intervals and subsequently identified by PCR. Each bar represents (percent plotted in y axis) the fraction of the mixture.