epigénèse probabiliste et développement cerisy ... en ligne/15-09 cerisy/120... · les approches...
TRANSCRIPT

Epigénèse Probabiliste et Développement
Cerisy, Septembre 2015

Epigénèse Probabiliste et Développement
� François Jouen, Laboratoire Cognitions Humaine et Artificielle, EPHE
� Michèle Molina, Laboratoire Psychologie des Actions Motrices et Langagières, UCBN

Les approches classique du développement
� Tirent leur origine de la thermo dynamique, la science de l’équilibre des systèmes physiques (équilibration et stades)
� Le développement conçu comme un système linéaire, c’est-à-dire un système prédictif
� Un système normatif (les QDs et QIs )
� Une statistique de la norme et non de la variance (le non traitement des différences individuelles)
� Un ordre des stades invariable et universel
� La génétique comme fonction explicative

Le développement comme un système complexe
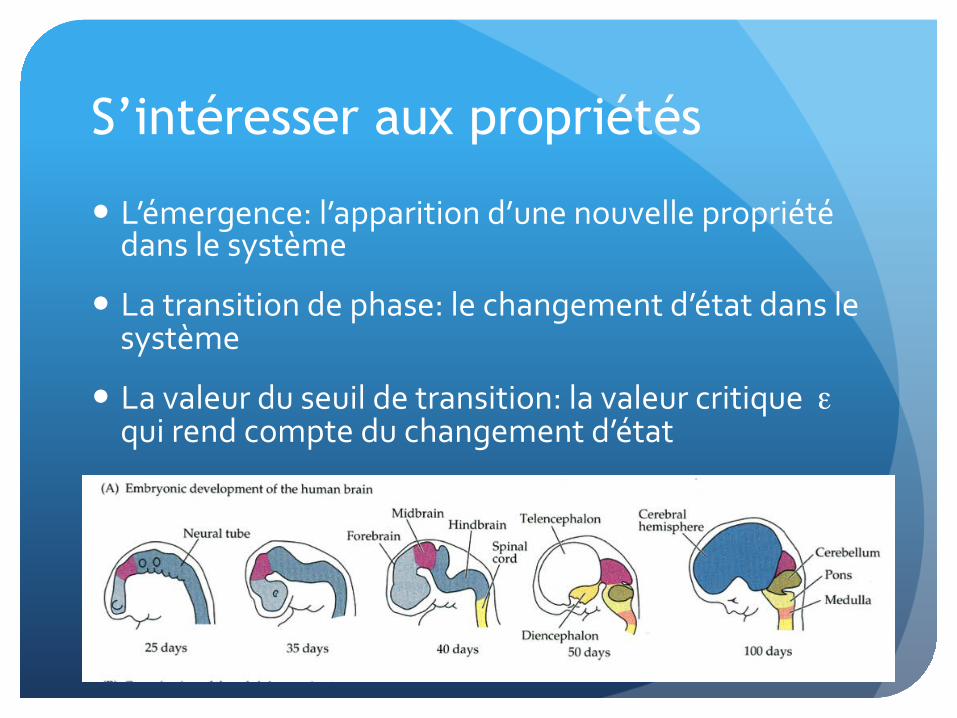
S’intéresser aux propriétés
� L’émergence: l’apparition d’une nouvelle propriété dans le système
� La transition de phase: le changement d’état dans le système
� La valeur du seuil de transition: la valeur critique ε qui rend compte du changement d’état

Que nous apprend l’embryologie?
� L’ADN est à l’origine de la variabilité génique: la ségrégation aléatoire des chromosomes maternels et paternels crée une diversité génique quasi illimitée
� Le développement embryofœtal est une suite d’événements spatiaux et temporels qui permettent la constitution de formes de plus en plus complexes (du zygote à l’individu)
� La régulation de ce système est assurée par des facteurs génétiques et des facteurs épignétiques
� Les facteurs génétiques sont les gènes régulateurs du développement (précoces et nécessaires pour déterminer le plan du corps)
� Les facteurs épigénétiques sont externes et sont des modulateurs de l’expression génique (les stimulations externes dans le cas de la synaptogénèse)

Variabilité phénotypique
© Tanner, 1972
Quelle proximité génique entre ces deux individus?

La TSGN d’Edelman (1987): les mécanismes de sélection

Constitution de la neuro-anatomie comme processus stochastique

La neurogenèse
� La corticogenèse repose sur l’ensemble de processus (multiplication, migration, différenciation, apoptose, synaptogenèse et myélinogenèse) regroupé sous le terme de neurogenèse
� Elle assure la mise en place d’un ensemble de neurones (100 milliards) dont chacun apparaît comme une entité originale qui se distingue des autres par sa forme et le nombre de ses connexions (1000 à 10 000 par neurone)

Neurones et densité post-synaptique
. © CNRS Photothèque / Daniel Choquet
© CNRS Images / Bordeaux Imaging Center / Sébastien Marais / Daniel Choquet

Les conséquences sur le fonctionnement du SNC
� La mise en place de la neuro-‐anatomie depend de l’activité mise en œuvre pour intégrer les effets de l’environnement
� Une organisation redondante des systèmes neuronaux (multimodalité)
� Une organisation massivement distribuée des réseaux de neurones (plasticité)
� Une énorme variabilité topobiologique (rend compte des différences phénotypiques et rend possible le principe de sélection)

Sélection et expérience: Gottlieb 1991
� Expériences sur les canards:
� Groupe 1: environnement sonore fixe (la fondamentale)
� Groupe 2: environnement sonore variable (varia=on autour de la fondamentale)
� Groupe 1: pas de spécificité au cri de l’espèce
� Groupe 2: spécificité au cri de l’espèce et pas de différence avec un groupe témoin
� Mise en évidence de la fonc/on développementale de la variabilité

Sélection et expérience: Gottlieb 1991
� Canards avec environnement sonore variable d’une autre espèce (poule):
� Perte de la réponse à la spécificité au cri de l’espèce canard
� Acquisi=on de la réponse d’une autre espèce
� Mise en évidence de l’effet de l’environnement sur la mise en place d’une fonc/on

Gottlieb 1991: Un effet de l’environnement sur la structure
• ADN—mARN—Protéines. On peut u=liser protéines pour mesurer l’expression des gènes.
• Deux groupes de canards: • contrôle dans le noir sans s=mula=on • expérimental soit incubateur éclairé soit enregistrement des cris des congénères.
• Résultats: modifica=on significa=ve de la synthèse de protéines dans les régions audi=ves et visuelles.
• S/mula/on a modifié l’expression génique

Un effet complexe de l’environnement: modification phénotypique
� Des cellules clonées (lymphocytes T régulateurs (T-reg)) de souris
� Expression phénotypique (expression de la GFP (Green Fluorescent Protein)) différenciée selon la densité des cellules dans les microgrilles.
© Andras Paldi (EPHE)

Un effet partagé parmi les espèces Especes Signal Résultat
Nematodes Absence ou présence de nourriture
Modification de l’expression du gène daf-7
Reptiles température Modification du sexe
Rats Stimulation tactile Modification de C-Fos (donc de l’induction cellulaire)
Rats Nutrition prénatale ADN cérébral et nombre de neurones
Rats Séparation précoce de la mère Modification à vie du corticotropin-releasing factor
Humains Stress psychologique Activité ARNm réduite dans le récepteur IL-2 (diminution de la défense immunitaire)

Que nous apprennent ces travaux?
� Le développement des individus à l’intérieur d’une espèce donnée ne suit pas un cours invariant ou inévitable. Plus spécifiquement la séquence et le résultat du développement individuel sont probables et dépendent de l’environnement
� Tous les systèmes sensoriels commencent à fonc/onner pendant qu’ils maturent encore (pendant les processus de division cellulaire, de migra=on, de croissance et de différen=a=on), de telle façon que chaque système contribue à son développement prénatal mais également à son développement postnatal

Un autre enseignement à tirer
� Il existe au cours de l’embryogenèse une bidirectionalité des relations entre structures et fonctions
� La bidirectionnalité est une conséquence de la nature probabiliste de l’épigénèse: elle implique des effets réciproques dans la relation structure-‐fonction.
� Si la fonction dépend de la structure, la fonction peut, elle aussi, modifier le développement des structures périphériques ou centrales qui la soutiennent.

Une nouvelle conception de l’activité des gènes et du développement(Gottlieb)

Le problème de la naissance
� Néoténie: 5/6 de la croissance du cerveau se déroule après la naissance
� Rupture épistémique: passage de l’organisme (biologique) au sujet (cognitif)?
� la mise en place des structures au cours de l’embryogenèse, largement déterminée sur le plan génétique, rend possible, après la naissance, la construction des connaissances
� Mais dans ce cas, quid de la continuité transnatale motrice (de Vries, Visser & Prechtl, 1982), sensorielle, et mnésique (Lecanuet, Granier-Deferre & Schaal, 1993)?
� Quid de la plasticité neuronale observée durant la gestation (Gazzaniga, Ivry & Mangun, 1998 ) ?

Une hypothèse différente
� Les mécanismes épigénétiques permettent la construction d’une neuro-anatomie spécifique: un substrat neural initialement plastique, peu différencié, qui présente une extrême variabilité et une redondance
� La neurogenèse est un système dépendant d’expériences
� La synaptogenèse est un système en attente d’expériences qui reste ouvert aux entrées environnementales
� Les processus de sélection sont de même nature avant et après la naissance
� Les processus de sélection permettent de donner un sens à l’univers

Redondance et multimodalité
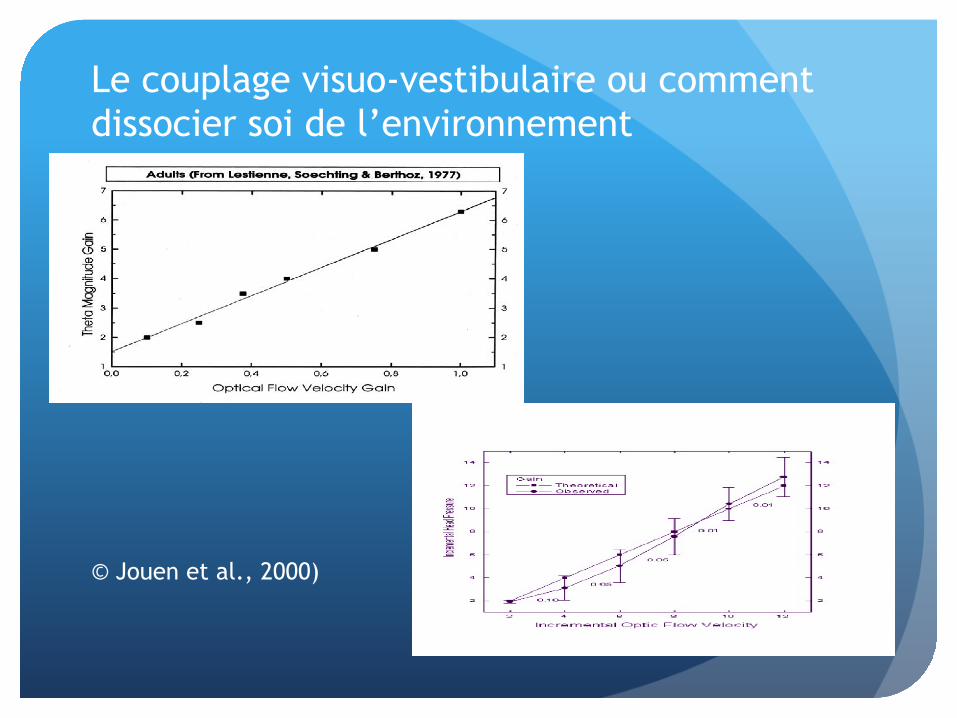
Le couplage visuo-vestibulaire ou comment dissocier soi de l’environnement
© Jouen et al., 2000)

Le colliculus ou comment adresser le même point de l’espace
© Stein et Meredith, 1993

Les structures élémentaires de la connaissance: Alegria et Noirot, 1978

Apprentissage précoce
� Système de sélection
� Utilise la neuro anatomie
� Rapide (3 repas)
� Se construit sur la base de réponses embryonnaires

Hindawi Publishing CorporationInternational Journal of PediatricsVolume 2012, Article ID 848596, 5 pagesdoi:10.1155/2012/848596
Research ArticlePrenatal Mouth Movements: Can We Identify Co-Ordinated FetalMouth and LIP Actions Necessary for Feeding?
Nadja Reissland,1 Claire Mason,1 Benoist Schaal,2 and Karen Lincoln3
1 Department of Psychology, University of Durham, South Road, Durham DH1 2NR, UK2 Developmental Ethology and Cognitive Psychology Group, Center for Smell, Taste, and Food Science, CNRS (UMR 6265),Universite de Bourgogne, 21078 Dijon Cedex, France
3 The James Cook University Hospital, Middlesbrough TS4 3BW, UK
Correspondence should be addressed to Nadja Reissland, [email protected]
Received 27 March 2012; Accepted 23 May 2012
Academic Editor: Katsumi Mizuno
Copyright © 2012 Nadja Reissland et al. This is an open access article distributed under the Creative Commons AttributionLicense, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properlycited.
Observations of prenatal movement patterns of mouth and lips essential for feeding could have the potential for an assessment ofthe readiness to feed after birth. Although there is some research on sucking per se, we know very little about prenatal preparatorymovements for sucking, namely, the ability to co-ordinate opening the mouth widely and then pursing the lips as if around ateat or nipple in utero. The purpose of the present study was to test two hypotheses using an adapted version of the Facial ActionCoding Scheme: first that mouth stretch (AU 27) will be followed by lip pucker (AU 18), and second that these coordinatedmovement patterns will increase as a function of gestational age. Fifteen healthy fetuses were scanned four times between 24 and36 weeks gestation using 4D ultrasound visualization. Results showed a decreased number of mouth stretches with increasing fetalage. Contrary to our expectations, we did not find an increase in movement patterns of mouth stretch followed by lip puckerin preparation for feeding ex utero. The results are discussed in terms of sensory triggers in utero required to elicit preparatorymovements for feeding ex utero.
1. Introduction
In order to feed from the breast or an artificial teat, new-borninfants need to be able to orally grasp and suck. Nutritivesucking has been examined in preterm infants [1] as ameans of evaluating developmental risk for poor motormaturity [2]. The difficulty of many preterm infants to suckfrom the breast or a bottle has been attributed to theirunderdeveloped motor abilities due to their premature birth[3]. However, others suggest that it might be caused byneurological problems, such as not being able to co-ordinatebreathing, sucking, and swallowing [4]. Medoff-Cooper et al.[5] examined the relationship between the pattern of suckingbehavior in preterm infants and neurodevelopmental out-comes during the first year of life and found that suckingbehavior in preterm infants is an indication of their psy-chomotor and mental development.
The majority of fetal movement patterns develop duringthe first half of pregnancy [6] and progress to mature forms
after birth [7–9]. Observations of prenatal movement pat-terns of mouth and lips essential for later feeding abilitycould have the potential for an assessment of the readinessto suck after birth [10]. Research indicates that by 24 weeksof menstrual age, the fetus responds to palmar stimulationwith mouth opening [11]. Humphrey [11] used direct obser-vation of externalized human fetuses placed in a warm fluidbath showing a link between manual and oral activity. Insum, although some research [8–10] has addressed suckingbehavior, such as, tongue movements and swallowing, less isknown about prenatal preparatory movements for sucking[10], specifically the ability to coordinate mouth and lipmovements in utero. In one study [10], fetal movementswere observed in clusters so that, for example, mouthingwas defined as “rhythmic open-and-close mouth movementswithout significant fluid or tongue movement” (page 68). Incontrast to this work, the current study identified two specificmovements, namely, the “pursing of lips” (AU 18) whichdoes do not include inferior maxilla movement, whereas
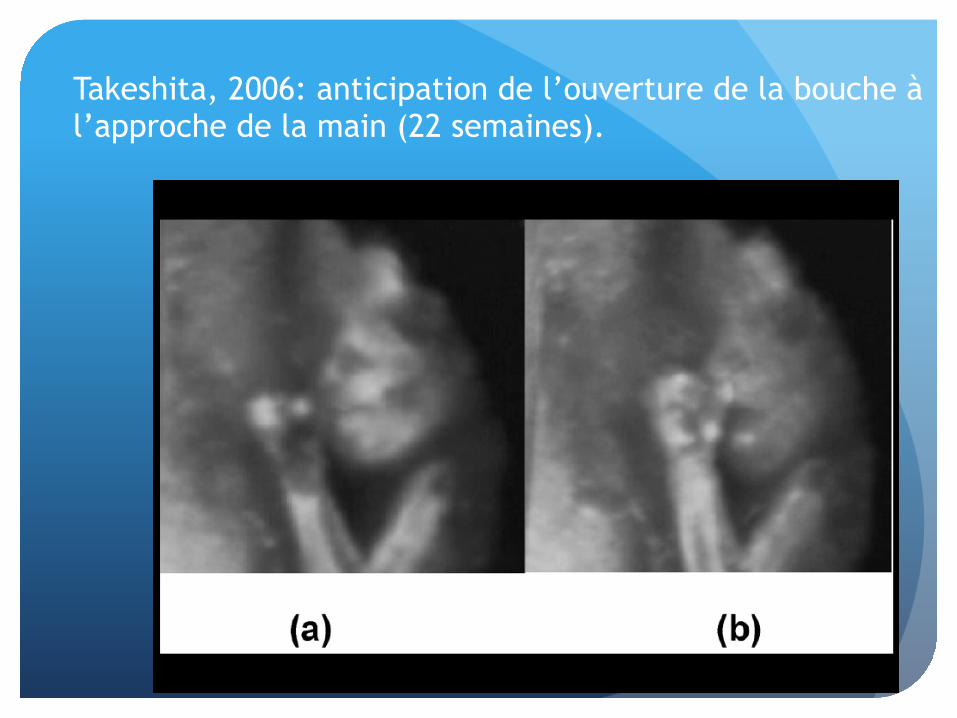
Takeshita, 2006: anticipation de l’ouverture de la bouche à l’approche de la main (22 semaines).

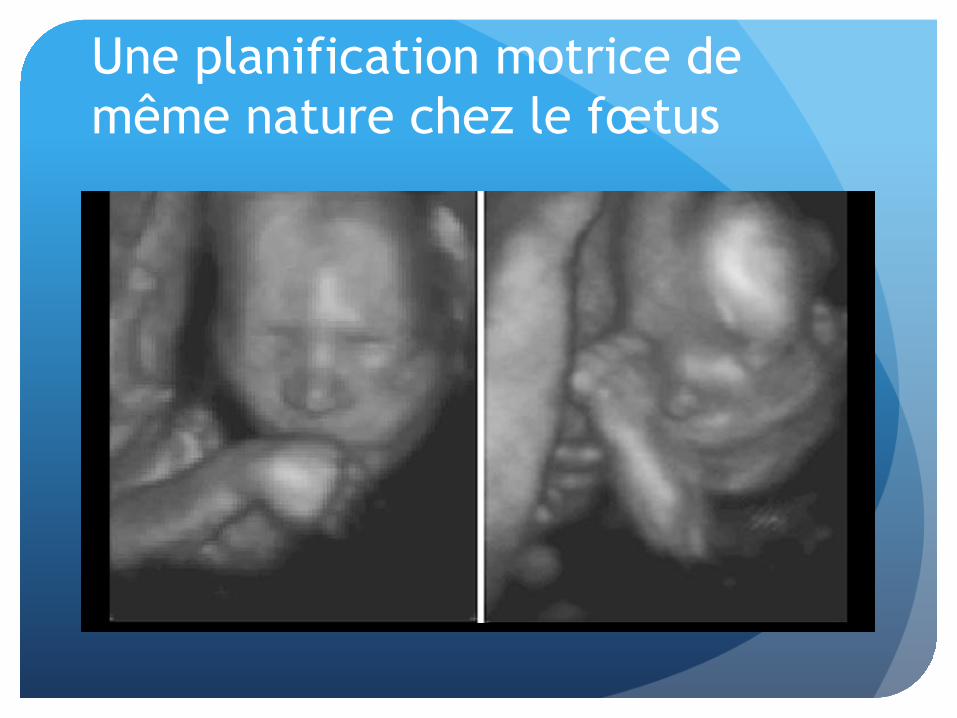
Une planification motrice de même nature chez le fœtus

le connectome ou la corticalisation néonatale
© CNRS Photothèque / Université de Strasbourg / Antoine Grigis
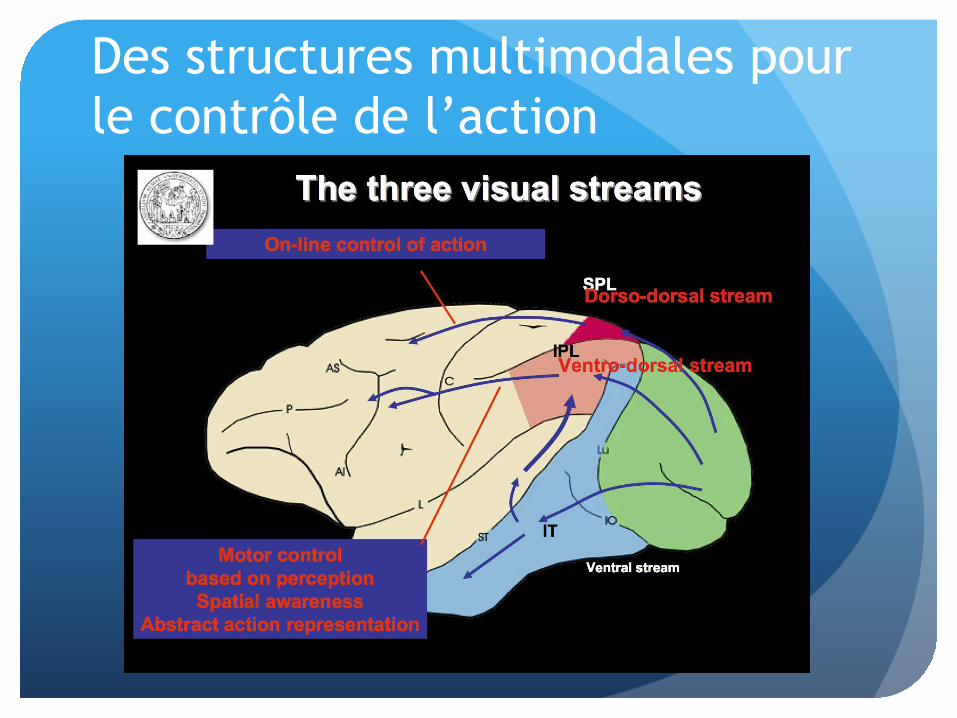
Des structures multimodales pour le contrôle de l’action

La voie ventro-dorsale: F4-VIP
34

L’aire pré-motrice F4
35
� Aire somato-centrée
� Capture manuelle (bras)
� Orientation de la tête

La voie ventro-dorsale: F5-AIP
36

L’aire pré-motrice F5
M2 Approche Pluridisciplinaire du Handicap Neurologique de l’adulte
37
3 classes de neurones • Neurones moteurs • Des neurones sensori-
moteurs • Neurones
canoniques • Neurones miroirs

© Meltzoff and Moore, 1977
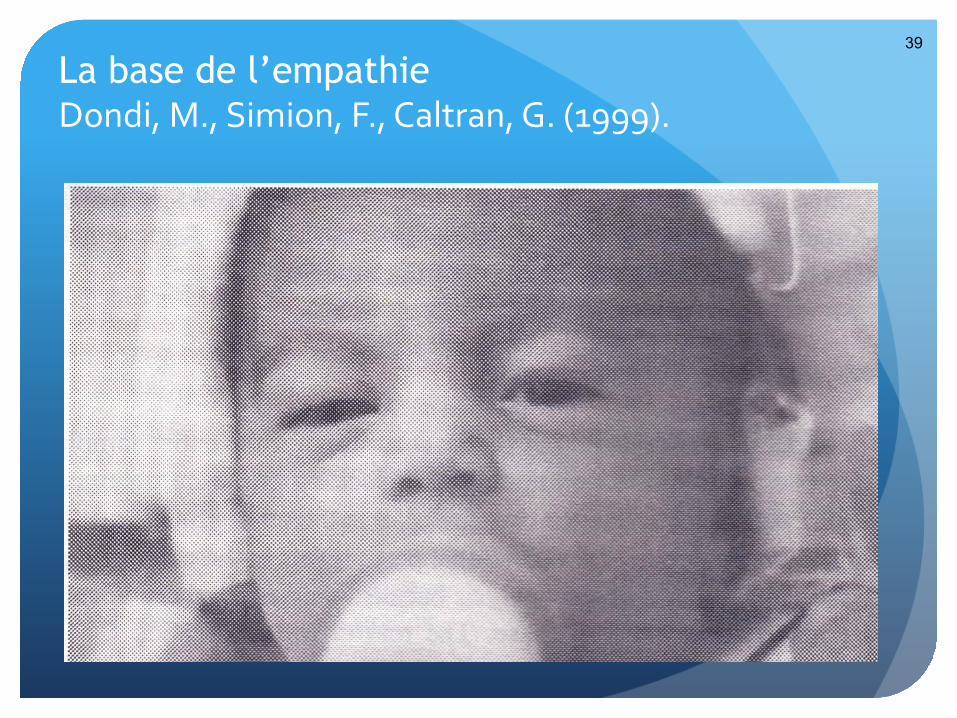
La base de l’empathie Dondi, M., Simion, F., Caltran, G. (1999).
39

40

Neurones miroirs et apprentissage
41
L’expérimentateur réalise face à un nouveau-né âgé de quelques heures un mouvement avec ses doigts (E Nagy)

Et le nouveau-né imite
Crédits : E. Nagy
42

Variabilité à tous les niveaux
� Au niveau des processus topobiologiques biochimiques qui sont responsables de la construction du SNC.
� Au niveau des connexions entre neurones.
� Au niveau des relations entre structures neuro anatomiques
� Un même message entrant peut sélectionner des organisations connexionnelles internes différentes et cependant conduire à une relation entrée-sortie identique, c’est-à-dire au même comportement de l’organisme (Changeux)
43

Variabilité motrice
Nouveau-né à terme Prématuré né à 28 semaines
© Hadders-Algra et al., 1997
44

Réflexes et variabilité
§ Durée (seconde) § Amplitude (volt) § Fréquence (Hz)
© Molina et Jouen, 1998
45

Le réflexe d’agrippement palmaire chez le nouveau-né
Les réflexes « archaïques » sont modulables © Molina et Jouen, 1998
46
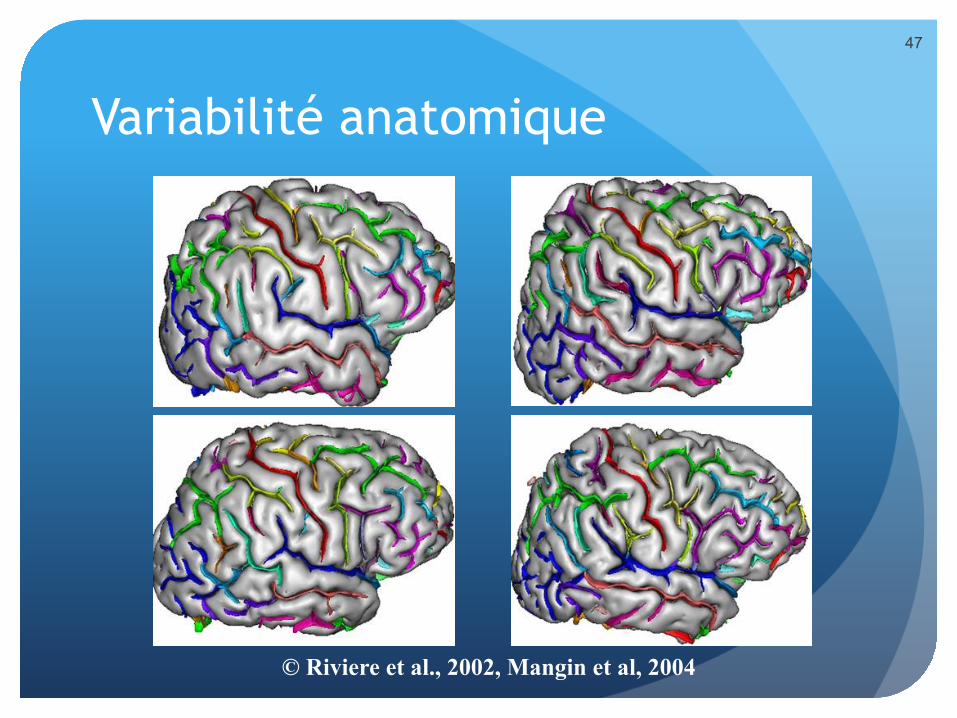
Variabilité anatomique
© Riviere et al., 2002, Mangin et al, 2004
47

Variabilité fonctionnelle: les sujets se ressemblent-ils?
© Wei et al, NeuroImage 2004.
48

La neuroanatomie permet la plasticité
© CNRS Photothèque / IMN / Bruno Bontempi

Plasticité neuronale post-natale
50
Crédits: Purves et al. Neurosciences, de Boeck Editeur

Plasticité et sélection : « other-race effect »
� Capacité à identifier les visages d’autres ethnies
� Présente à la naissance
� Disparaît progressivement entre 3 et 9 mois (Kelly et al. 2007)

Plasticité et sélection: le langage
� Mise en place durant l’embryogenèse
� Capacité à traiter toutes les langues à la naissance
� Langue maternelle: premier mois de vie
� Maintien de 2 réseaux si bilinguisme
� 3 mois: activation des structures langagières dans la partie gauche du cerveau (comme adulte)

Plasticité et sélection: le langage
53
© Egerton et Brynner, 2001

Des effets drastiques …
IRMf chez 389 enfants de 4 à 22 ans (de ménages vivant en dessous du seuil fédéral de pauvreté) les volumes régionaux de la matière grise de ces enfants sont de 8 à 10 % en dessous de la norme de développement dans les lobes frontaux et temporaux. « Notre travail suggère que les structures cérébrales spécifiques de l’apprentissage sont vulnérables aux conditions environnementales de la pauvreté, telles que le stress, le manque de stimulation et des problèmes de nutrition. »

Mais compensables à long terme! 55

Un interaction gènes environnement complexe
56

Merci de votre attention