does dietary hyperphagia contradict the lipostatic theory?
TRANSCRIPT

Neuroscience & Biobehavioral Reviews. Vol. 14, pp. 117-123. © Pergamon Press plc, 1990. Printed in the U.S.A. 0149-7634/90 $3.00 + .00
Does Dietary Hyperphagia Contradict the Lipostatic Theory?
I S R A E L R A M I R E Z
Monell Chemical Senses Center, 3500 Market Street, Philadelphia, PA 19104-3308
Rece ived 29 June 1989
RAMIREZ, I. Does dietary hyperphagia contradict the lipostatic theory? NEUROSCI BIOBEHAV REV 14(2) 117-123, 1990.--It has frequently been suggested that body weight or fat somehow exerts an inhibitory influence on food intake in a way that acts to maintain a stable body weight or fat. The principal evidence supporting this idea is that animals that have been induced to overeat and become overweight by various means, eat less than control rats when they a.~." peranitt,d to eat freely. If the degree of suppression of appetite by overweight is as large as several experiments suggest, then dietary hyperphagia should be self-limiting. Any overeating induced by dietary treatments should disappear after animals become moderately overweight. Animals fed some kinds of hyperhagia-promoting diets do show this pattern. However, animals fed other kinds of diets do not show this pattern, and with most diets, dietary hyperphagia continues for extended periods. This implies that either 1) overweight does not suppress appetite as much as suggested by various authorities, 2) dietary manipulations can override normal regulatory mechanisms, or 3) certain diets induce irreversible changes in body fat that are not evident from changes in body weight.
Lipostatic theory Body weight Food intake Appetite Diet
IT is widely thought that energy intake, body weight and/or body fat are controlled in a homeostatic fashion because animals increase food intake after a fast and decrease intake after being overfed [e.g., (2, 11, 12, 19, 20, 36, 47, 57, 59)]. This concept is the basis of the lipostatic theory of food intake (20). Although the mechanisms involved remain obscure, the importance of homeostatic controls has not been seriously questioned. At f'trst sight, the finding that certain diets can cause animals to overeat and become obese (31, 38, 49) would seem to cast doubt on the importance of homeostatic mechanisms. It is therefore surprising that detailed discussions of this issue have not been published.
The purpose of the present review is to examine whether studies of dietary hyperphagia and obesity are inconsistent with current conceptions of the homeostatic control of energy intake, body weight, and body fat. This will be done by developing specific semiquantitative predictions based on studies of the suppression of appetite produced by overfeeding. Analysis of the results of these studies indicates that any overeating induced in rats by cafeteria (supermarket), high-fat, high-sugar or wet diets should abate within 2-6 weeks. The results of experiments involving dietary hyperphagia frequently contradict this expecta- tion. Theories about how diets stimulate energy intake are dis- cussed elsewhere (31, 38, 49).
The present review focuses on food intake as a controller of body weight and body fat. Changes in energy expenditure also influence body weight and body fat (31, 38, 54) but there are not enough data available to evaluate the role of energy expenditure [see (11, 18, 54) for differing views].
QUANTITATIVE RELATIONSHIP BETWEEN OVERWEIGHT AND APPETITE
In order to predict how long diets may induce overeating, it is
necessary to know the magnitude of the suppressive effects of overweight on food intake. Animals that have been forced to overeat and gain excess weight will undereat when they are allowed to eat freely (2, 5, 12-14, 25, 26, 32, 36, 39, 42, 44, 46, 55). Overeating may be induced in various ways including: gavage, insulin injections, electrical brain stimulation, supermar- ket diets, etc. Comparison of the degree of overweight or obesity with food intake should reveal the quantitative relationship be- tween these two variables.
A number of factors complicate the interpretation of such experiments. One could plausibly argue that some methods of inducing overeating make animals ill; for example, insulin injec- tions at a dose sufficient to induce overeating, commonly kill a fraction of the animals (36). Alternatively, forced overeating could induce adaptive changes that favor increased intake (14,42). Animals that have been induced to overeat by feeding them certain diets (high-fat, cafeteria, wet, etc.) might reduce intake when returned to control diets because plain rat chow may seem boring to a rat that has been eating cookies and chocolate [sensory contrast effect (39)]. Nevertheless, two studies (14,39) that include controls for these complicating "factors suggest that over- weight strongly suppresses appetite.
Representative Studies
Five published data sets are described below. All of them are derived from experiments on rats; other species have not been given much attention. These studies were chosen because they illustrate nature of the technical problems in this area and how they might be minimized. In each study, the methods and results were reported sufficiently completely to permit the present reanalysis. These studies employed a wide range of methods of forcing rats to overeat (insulin, gavage, cafeteria diet, brain stimulation). Similar
117

118 RAMIREZ
TABLE 1
STUDIES OF THE SUPPRESSION OF FOOD INTAKE BY OVERWEIGHT*
Peak Weightt Food Intaket Peak Body Fatt g kcal/day g No.
Ramirez and Friedman (36)
Low Dose Insulin Minimum 1-Day Intake
Control 376 --+ 8 96 ± 4 Insulin 412 ± 12 41 ± 23 % of Control 110% 43%
Suppression per 1% excess weight:[: 5.7%
Rogers (39) First Day's Intake
Restricted Cafeteria 307 ± 9 37 --- 6 Ad Lib Cafeteria 385 ±- 15 25 ± 5 % of Control 125% 68%
Suppression per 1% excess weight:~ 1.3%
Steffens (55) First Day's Intake
Control 400 79 Brain Stimulated 554 0 % of Control 139% 0%
Suppression per 1% excess weight:~ 2.60
Rothwell and Stock (44)
Tube fed 75% of ad lib intake for 22 days
Control 440 ± 13 Tube fed 498 ± 14 % of Control 113%
Suppression per 1% excess weight:~
Harris, Kasser and Martin (14)
7 10
4-8 8
Five-Day Intake
77 --- 3 109 ± 12 12 47 --- 1 158 --- 12 12
61% 145%
3.0%
First Day's Intake 100% Tube fed 260 57 32 6 160% Tube fed 327 37 73 6 % of Control 126% 65% 228%
Suppression per 1% excess weight:l: 1.3%
*Means _+ SE were taken from the tables or figures of the references cited. t% of Control was computed by dividing the value for the overweight group by the value of the control group. ~This was computed by dividing (% of control food intake - 100) by (100- % control body weight).
results have been reported in other publications (2, 5, 8, 13, 25-27, 41-43, 45, 46). Relevant statistics derived from tables or figures are summarized in Table 1.
Ramirez and Friedman. Rats were given protamine zinc insulin injections for ten days and then allowed to recover (36). Data from the low dose series (culminating in 6 units/rat/day) was used because this level of insulin treatment produced minimal mortal- ity. Only data from normal (i.e., nondiabetic) rats are used in this article. Maximal hyperphagia did not appear immediately after cessation of insulin treatment because of the long duration of action of protamine zinc insulin. Therefore, maximal weight is contrasted with minimum daily food intake; this occurred on the second day after the last injection.
Rogers. Rats were made to overeat by feeding them a cafeteria diet for 26 days (39). This study attempted to control for "sensory
contrast effects" (plain chow may seem insipid if preceded by highly palatable foods) by restricting the intake of some of the cafeteria rats until they weighed the same as control rats. This is an excellent control although food restriction could produce rats that are obese (increased body fat) but not overweight (29). The degree of hyperphagia was obtained by computing the difference between the intake of the ad lib cafeteria rats from the restricted cafeteria rats during the first day both groups consumed the control diet.
Steffens. Rats were induced to overeat by electrical hypotha- lamic stimulation 3 times a day for 21 days (55). Body weights for control rats were given but not food intakes. For the purposes of this article, daily food intake of experimental rats prior to electrical stimulation provides a satisfactory control.
Rothwell and Stock. Rats were fed by gavage (tube fed) for 21-24 days (44). Only data from Experiment 1C was used because

LIPOSTATIC THEORY 119
TABLE 2
FORMULAS FOR SIMPLE MODEL
Body Weight Gain = (difference between control and experimental groups in energy intake)/[(9 kcal/g fat)*(1.4 kcal eaten/kcal stored)]
Body Weight Loss=(difference between control and experimental groups in energy intake)/(9 kcal/g fat)
Food Intake=flood intake in control group)*(stimulation by diet) - (% overweight)*(l.6% suppression per 1% overweight)
it employed the largest number of subjects and provided the most detailed data. Daily food intake was not reported for the period that rats underate. The present analysis relies on a 5-day block; this probably underestimates maximal suppression of food intake because the degree of suppression of food intake declines as overweight declines. Data from the cafeteria diet experiments in the same report were not used here because of the possibility of contrast effects.
Harris, Kasser and Martin. Rats were tube fed for 21 days either 100% or 160% of their baseline intake (14). Two factors are probably responsible for the finding that the degree of hypophagia observed was less than in the other studies. After the rats were allowed to eat freely, body weight and body fat was persistently elevated in the rats fed 160% of their baseline food intake. The rats that were tube fed 100% of their baseline food intake, ate less food after tube feeding than they did prior to tube feeding.
DISCUSSION Increasing body weight to 110-126% of control values de-
creased food intake to 32-68% of control values, an impressive result. Increasing body weight by 39% completely suppressed food intake in Steffens study! In order to facilitate comparisons between experiments, Table 1 gives the estimated degree of suppression of food intake per 1% overweight. Thus, making a rat 1% overweight is expected to reduce food intake by 1.3-5.7% (Table 1). Van Itallie and Kissileff analyzed a different set of data and (59) reached a similar conclusion. The exact biochemical mechanisms (endocrine, neural, or metabolic) are not crucial for the present discussion. Only the magnitude of the relationship is needed for the predictions described below.
PREDICTIONS If being overweight suppresses energy intake, then dietary
hyperphagia should dissipate with time. The expected duration of hyperphagia may be calculated from the observation that increas- ing body weight by 1% decreases energy intake by 1.3-5.7% (see Table 1). Experiments involving dietary hyperphagia commonly stimulate energy intake by 8-50% (31, 38, 49). This increases body weight by 10-20% within 2-6 weeks. Therefore, hyper- phagia should be greatly attenuated within this time.
In order to make this line of reasoning more explicit, a simple mathematical model was devised based on a set of realistic assumptions (see Table 2). Control rat body weight and intake were derived from mean control values in Table 1. It is assumed that increasing body weight by 1% decreases food intake by 1.6%, which is the average of the five studies in Table 1. Application of this model to hypothetical animals that were 10 or 40% overweight shows that food intake is initially depressed and gradually ap- proaches control level (Fig. 1). Body weight falls over the same period (Fig. 1). These results are in general agreement with most published studies (2, 5, 13, 25-27, 44, 46) but see (8, 41--43, 45).
This same model was used to predict how much the intake of
c~
m c~ 0
7 0
6 0
5 0
4 0
3 0
2 0
1 0
0
- - 4 0 % . . . . 1 0 % . . . . . C o n t r o l O v e r w e t g h t O v e r w e i g h t
i ~ --~
i i I i i i i i
1 2 3 4 8 6 7 8
Weeks
5 1 0
4 9 0
i 470 45O
4 3 0
. ~ 4 1 0
mO 3 9 0
37O
3 6 0 ' J
0 1 2
I I I I I I
3 4 6 6 7 8
Weeks
FIG. 1. Behavior of a simple model for the appetite suppressive effects of overweight. Hypothetical animals are set at 0, 10, or 40% overweight on the first day. Control rats are assumed to have reached a stable weight. Data were computed on a daily basis but reported as weekly means (see Table 2).
rats would be suppressed by offering them diets that initially stimulate intake by 20% or 50%. It is assumed that excess intake leads to increased fat deposition with an efficiency of 1 kcal/fat deposited for each 1.4 kcal/food ingested (28); this figure was compared to some previous studies (32,34) and found to be adequate for low-fat diets. Thus, increasing the intake of a rat that normally eats 70 kcal/day to 105 kcal would increase body fat by 2.8 g; for a 365 g rat this 0.8% increase in body weight would decrease the next day's intake by 1.3 kcal.
Calculations for a eight-week prriod are shown in Fig. 2. It may be seen that hyperphagia is substantially reduced within a couple of weeks. The greater the initial level of hyperphagia, the more rapidly it declines during the first week. Some other sets of assumptions were examined: e.g., type of function relating over- weight and appetite suppression, level of food intake, etc. Unless one assumes that small degrees of overweight do not suppress appetite, the overall results are similar over a range of plausible sets of assumptions. Russek (47) described a more complex model that gave results similar to those seen in Fig. 2 (see his Fig. 19).
In certain situations, high-fat or high-sucrose diets may induce overweight without increasing energy intake (31, 38, 54). In these

120 RAM IREZ
A
"1~ 0 m
a~
0 , ¢
- - 5 0 % - - - 2 0 % Increase Increase
Cont ro l
110
100
90
80
70
6O 0
I I I i I I I i
1 2 3 4 5 6 7 8
W e e k s
4 3 0
4 2 0
4 1 0
4O0
39O
38O
37O
36O
38( 0
I I I I I I I I
1 2 3 4 5 6 7 8
W e e k s
FIG. 2. The same as Fig. 1 except that rats were offered diets that stimulate energy intake by 0. 20, or 50% on the first day.
cases, the present line of reasoning suggests that rats fed such diets should eventually eat substantially less energy than control rats.
Similar arguments have been made by others for dietary hyperphagia (2, 7, 11), hypothalamic lesions (19,24), ovarian hormones (60) and anorectic drugs (57). The major innovation in the present work is the semiquantitative approach which allows one to determine how long an experiment is needed to detect the predicted disappearance of hyperphagia.
Additional Contributing Factors
Other factors also act to suppress dietary hyperphagia. It has been suggested that feeding animals a monotonous diet tends to suppress intake (39,40). Indeed, the rationale behind the use of supermarket and cafeteria diets is that frequently switching foods should increase energy intake by reducing monotony. The impor- tance of variety in the control of long-term intake has neither been verified nor refuted [see (38, 40, 43)].
Rats are capable of learning the relationship between the energy content of a food and its taste and smell (3). There is not sufficient data on this factor in the control of energy intake to permit one to predict how much learning may influence intake of hyperphagia-promoting diets. However, learning should cause intake of hyperphagia-promoting diets to decrease with time.
On the other hand, adipocyte number can be increased by certain diets (8-10, 15, 21). This might cause rats fed obesity- promoting diets to remain heavier or fatter than control rats after the obesity-promoting diet is withdrawn, as has been observed in some studies (8, 41-43, 45). In other studies, however, dietary- obese animals lost all their excess body weight or body fat when returned to the control diet (27, 48, 50, 56). Any tendency for weight changes to be irreversible might prolong dietary hyper- phagia indefinitely. Hill et al. (15) found that dietary obesity was fully reversible after 17 weeks but not after 30 weeks of feeding a high-fat diet; this would suggest that irreversible changes are important only for very long studies. The importance of irrevers- ible changes cannot be evaluated until more is known about the factors that determine whether obesity is fully reversible.
ARE THE PREDICTIONS CONFIRMED?
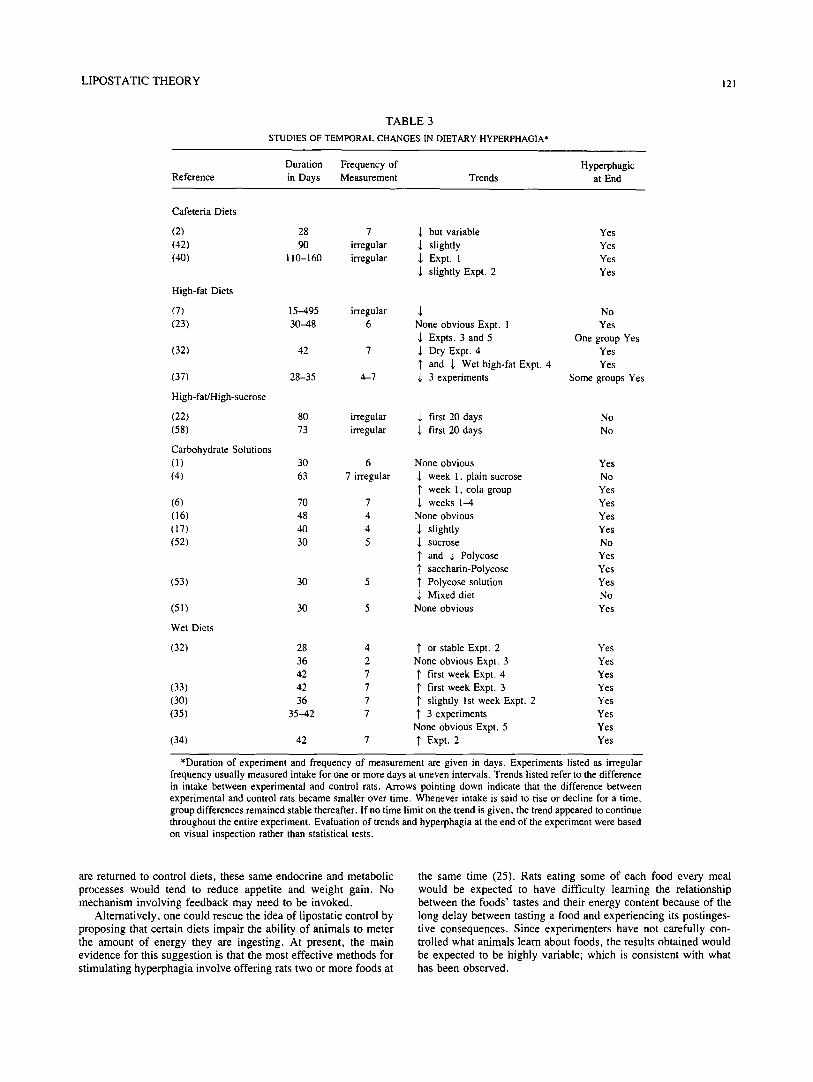
Experiments involving dietary hyperphagia and obesity do not uniformly show the expected temporal effects. Table 3 summa- rizes the results that have been obtained by various investigators. Experiments were selected for inclusion in Table 3 if they provided enough data for one to observe systematic temporal changes over 15-160 days. Experiments employing weanling rats were excluded because young rats are sometimes resistant to dietary hyperphagia (34). Experiments in which animals became overweight or obese without showing hyperphagia, were also excluded in order to simplify the discussion [see (54)].
On the whole, the observed declines are not dramatic. In a majority of the reports, dietary hyperphagia declined within 2-6 weeks (Table 3) but some hyperphagia and/or excess weight gain usually persis~:'J for as long as the experiments were conducted. In other experin:ents, energy intake showed no regular change over time despite prolonged excess weight gain (Table 3). Occasion- ally, energy intake increased over the first few days; the reasons for this increase are unknown but might include neophobia to unfamiliar foods or physiological adaptation to the diets.
In general, experiments involving high-fat diets and sugar solutions show the expected decrease whereas experiments involv- ing Polycose solutions and wet diets do not. It is not obvious why this should be so.
Unexpectedly, rats fed an experimental diet eventually came to eat less energy than control rats in at least two experiments (7,52). Presumably, energy efficiency was increased by these diets.
Evidence that hyperphagia decreases with time does not estab- lish that lipostatic mechanisms are responsible for the observed decrease. At least part of the decrease must be attributable to such things as learning to reduce intake of calorically dense foods, habituation, and physiological adaptations. Experimental tests capable of discriminating among these possibilities are certainly possible but they have not been done. The best that can be said at present is that some types of hyperphagia-promoting diets produce the expected results.
CONCLUSIONS
The findings reported here suggest that some forms of dietary hyperphagia persist longer than would be expected if lipostatic mechanisms were operating. Furthermore, some forms of dietary hyperphagia consistently fail to show the expected reduction in intake over time.
One possible explanation is that overweight or excess body fat does not directly suppress appetite. Perhaps force-feeding animals stimulates metabolic or endocrine mechanisms that suppress ap- petite directly without acting via a lipostat (7). Certain diets may alter metabolic and endocrine mechanisms in such a way as to favor the development of obesity (25); when dietary=obese animals
LIPOSTATIC THEORY 121
TABLE 3
STUDIES OF TEMPORAL CHANGES IN DIETARY HYPERPHAGIA*
Duration Frequency of Hyperphagic Reference in Days Measurement Trends at End
Cafeteria Diets
(2) 28 7 ,~ but variable Yes (42) 90 irregular ,l, slightly Yes (40) 110-160 irregular ~, Expt. 1 Yes
,I, slightly Expt. 2 Yes
High-fat Diets
(7) (23)
(32)
(37)
High-fat/High-sucrose
(22) (58)
Carbohydrate Solutions (1) (4)
(6) (16) (17) (52)
(53)
(51)
Wet Diets
(32)
(33) (30) (35)
(34)
15-495 irregular ~, No 30--48 6 None obvious Expt. 1 Yes
-l, Expts. 3 and 5 One group Yes 42 7 ,l, Dry Expt. 4 Yes
1' and ~, Wet high-fat Expt. 4 Yes 28-35 4-7 ~, 3 experiments Some groups Yes
80 irregular ~, first 20 days No 73 irregular -1, first 20 days No
30 6 None obvious Yes 63 7 irregular ~ week 1, plain sucrose No
1' week 1, cola group Yes 70 7 ~, weeks 1-4 Yes 48 4 None obvious Yes 40 4 ,~ slightly Yes 30 5 $ sucrose No
~" and ~, Polycose Yes 1' saccharin-Polycose Yes
30 5 1' Polycose solution Yes ~, Mixed diet No
30 5 None obvious Yes
28 4 I" or stable Expt. 2 Yes 36 2 None obvious Expt. 3 Yes 42 7 1' first week Expt. 4 Yes 42 7 1' first week Expt. 3 Yes 36 7 1' slightly 1st week Expt. 2 Yes
35-42 7 1' 3 experiments Yes None obvious Expt. 5 Yes
42 7 ~' Expt. 2 Yes
*Duration of experiment and frequency of measurement are given in days. Experiments listed as irregular frequency usually measured intake for one or more days at uneven intervals. Trends listed refer to the difference in intake between experimental and control rats. Arrows pointing down indicate that the difference between experimental and control rats became smaller over time. Whenever intake is said to rise or decline for a time, group differences remained stable thereafter. If no time limit on the trend is given, the trend appeared to continue throughout the entire experiment. Evaluation of trends and hyperphagia at the end of the experiment were based on visual inspection rather than statistical tests.
are returned to control diets, these same endocrine and metabolic processes would tend to reduce appetite and weight gain. No mechanism involving feedback may need to be invoked.
Alternatively, one could rescue the idea of lipostatic control by proposing that certain diets impair the ability of animals to meter the amount of energy they are ingesting. At present, the main evidence for this suggestion is that the most effective methods for stimulating hyperphagia involve offering rats two or more foods at
the same time (25). Rats eating some of each food every meal would be expected to have difficulty learning the relationship between the foods ' tastes and their energy content because of the long delay between tasting a food and experiencing its postinges- tive consequences. Since experimenters have not carefully con- trolled what animals learn about foods, the results obtained would be expected to be highly variable; which is consistent with what has been observed.

122 RAMIREZ
Another approach to saving the idea of lipostatic control is to argue that temporary overweight is not fully reversible. Perhaps being slightly overweight for only a few weeks regularly produces lasting increases in body fat that may not be evident if only body weight is measured. This approach rescues the lipostatic hypoth- esis by weakening it.
Experimental tests o f these hypotheses could improve our understanding of the possible existence and importance of lipo- static mechanisms. It would be helpful if more future studies reported data on the time course of dietary effects. Most studies
report only total intake over the entire experinaental period if they report intake at all.
ACKNOWLEDGEMENTS
This work was supported by a National Science Foundation grant (BNS 87-19309), National Institutes of Health support (Biomedical Research Support Grant RR05825-07), and the Monell Chemical Senses Center. Highly skilled technical assistance was provided by Charlene Connelly and William McLaughlin. M. Tordoff and Peter J. Rogers provided useful suggestions on earlier versions of this manuscript.
R E F E R E N C E S
I. Ackroff, K.; Sclafani, A. Sucrose-induced hyperphagia and obesity in rats fed a macronutrient self-selection diet. Physiol. Behav. 44: 181-187; 1988. 24.
2. Armitage, G.; Hervey, G. R.; Rolls, B. J.; Rowe, E. A.; Tobin, G. The effects of supplementation of the diet with highly palatable foods 25. upon energy balance in the rat. J. Physiol. 342:229-251: 1983.
3. Booth, D. A. Conditioned satiety in the rat. J. Comp. Physiol. 26. Psychol. 81:457--471: 1972.
4. Bukowiecki, L. J.; Lupien, J.; Foll~a, N.; Jahjah, L. Effects of sucrose, caffeine, and cola beverages on obesity, cold resistance, and 27. adipose tissue cellularity. Am. J. Physiol. 244:R500-R507; 1983.
5. Carpenter, R. G.; Grossman, S. P. Reversible obesity and plasma fat metabolites. Physiol. Behav. 30:51-55; 1983. 28.
6. Castonguay, T. W.; Hirsch, E.; Collier, G. Palatability of sugar solutions and dietary selection? Physiol. Behav. 27:7-12; 1981. 29.
7. Faust, I. M. Signals from adipose tissue. In: Cioffi, L. A.; James, W. P. T.; ltallie, T. B., eds. The body weight regulatory system: Normal 30. and disturbed mechanisms. New York: Raven Press: 1981:39-43.
8. Faust, I. M. Nutrition and the fat cell. Int. J. Obes. 16:314-321; 1986. 31. 9. Faust, I. M.; Johnson, P. R.; Stern, J. S.; Hirsch, J. Diet-induced
adipocyte number in adult rats: a new model of obesity. Am. J. 32. Physiol. 235:E279-E286; 1978.
10. Faust, I. M.; Miller, W. H., Jr.; Sclafani, A.; Aravich, P. F.; Triscari, 33. J.; Sullivan, A. C. Diet-dependent hyperplastic growth of adipose tissue in hypothalamic obese rats. Am. J. Physiol. 247:R1038-RI046; 34. 1984.
11. Flatt, J. P. Dietary fat, carbohydrate balance, and weight mainte- 35. nance: effect of exercise. Am. J. Clin. Nutr. 45:296-306; 1987.
12. Friedman, M. I.; Ramirez, I. Relationship of fat metabolism to food 36. intake. Am. J. Clin. Nutr. 42:1093-1098; 1985.
13. Geary, N.; Grrtschel, H.; Petry, H.-P.; Scharrer, E. Meal patterns and body weight changes during insulin hyperphagia and postinsulin 37. hypophagia. Behav. Neural Biol. 31:435-442; 1981.
14. Harris, R. B. S.; Kasser, T. R.; Martin, R. J. Dynamics of recovery of body composition after overfeeding, food restriction or starvation 38. of mature female rats. J. Nutr. 116:2536-2546; 1986.
15. Hill, J. O.; Dorton, J.; Sykes, M. N.; DiGirolamo, M. Reversal of 39. dietary obesity is influenced by its duration and severity. Int. J. Obes. 13:711-722; 1989.
16. Hirsch, E.; Ball, E.; Godkin, L. Sex differences in the effects of 40. voluntary activity on sucrose-induced obesity. Physiol. Behav. 29: 253-262: 1982.
17. Hirsch, E.; Walsh, M. Effect of limited access to sucrose on 41. overeating and patterns of feeding. Physiol. Behav. 25:129-134; 1982.
18. Keesey, R. E. Physiological regulation of body weight and the issue 42. of obesity. Med. Clin. North Am. 73:15-27; 1989.
19. Keesey, R. E. A set-point analysis of the regulation of body weight. In: Stunkard, A. J., ed. Obesity. Philadelphia: W. B. Saunders; 43. 1980:144-165.
20. Kennedy, G. C. The role of depot fat in the hypothalamic control of food intake in the rat. Proc. R. Soc. Lond. (Biol.) 140:578-592; 1953. 44.
21. Klyde, B. J.; Hirsch, J. Increased cellular proliferation in adipose tissue of adult rats fed a high-fat diet. J. Lipid Res. 20:705-715; 1979.
22. Levin, B. E.; Triscari, J.; Sullivan, A. C. Relationship between 45. sympathetic activity and diet-induced obesity in two rat strains. Am. J. Physiol. 245:R367-R371; 1983. 46.
23. Lucas, F.; Ackroff, K.; Sclafani, A. Dietary fat-induced hyperphagia
in rats as a function of fat type and physical form. Physiol. Behav. 45:937-946; 1989. Morrison, S. D. The hypothalamic syndrome in rats. Fed. Proc. 36:139-142; 1977. Panksepp, J.; Nance, D. M. Insulin, glucose and hypothalamic regulation of feeding. Physiol. Behav. 9:447-451; 1972. Panksepp, J.; Pollack, A.; Krost, K.; Meeker, R.; Ritter, M. Feeding in response to repeated protamine zinc insulin injections. Physiol. Behav. 14:487--493; 1975. Peckham, S. C.; Entenman, C.; Carroll, H. W. The influence of a hypercaloric diet on gross body and adipose tissue composition in the rat. J. Nutr. 77:187-197; 1962. Pullar, J. D.; Webster, A. J, F. The energy cost of fat and protein deposition in the rat. Br. J. Nutr. 37:355-363; 1977. Ramirez, I. Hypophagia following dietary obesity. Physiol. Behav. 38:95-98; 1986. Ramirez, I. Diet texture, moisture and starch type in dietary obesity. Physiol. Behav. 41:149-154; 1987. Ramirez I. When does sucrose increase appetite and adiposity? Appetite 9:1-19; 1987. Ramirez, I. Feeding a liquid diet increases energy intake, weight gain and body fat in rats. J. Nutr. 117:2127-2134; 1987. Ramirez I. Overeating, overweight and obesity induced by an unpreferred diet. Physiol. Behav. 43:501-506; 1988. Ramirez I, Resistance to dietary hyperphagia in juvenile rats. J. Nutr. 119:1333-1339; 1989. Ramirez I. Stimulation of energy intake and growth by saccharin in rats. J. Nutr. 120:123-133; 1990. Ramirez I.; Friedman, M. I. Metabolic concomitants of hypophagia during recovery from insulin-induced obesity in rats. Am. J. Physiol. 245:E21 I-E219; 1983. Ramirez, I.; Friedman, M. I. Dietary hyperphagia in rats: Role of fat, carbohydrate, and energy content. Physiol. Behav. 47(6): in press; 1990. Ramirez, I.; Tordoff, M. G.; Friedman, M. I. Dietary obesity and hyperphagia: What causes them? Physiol. Behav. 45:163-168; 1989. Rogers, P. J. Returning "cafeteria-fed" rats to a chow diet. Negative contrast and effects of obesity on feeding behavior. Physiol. Behav. 35:493-499; 1985. Rogers, P. J.; Blundell, J. E. Meal patterns and food selection during the development of obesity in rats fed a cafeteria diet. Neurosci. Biobehav. Rev. 8:441-453; 1984. Rolls, B. J.; Rowe, E. A. Exercise and the development and persistence of dietary obesity in male and female rats. Physiol. Behav. 23:241-247; 1979. Rolls, B. J.; Rowe, E. A.; Turner, R. C. Persistent obesity in rats following a period of consumption of a mixed, high energy diet. J. Physiol. 298:415-427; 1980. Rolls, B. J.; van Duijvenvoorde, P. M.; Rowe, E. A. Variety in the diet enhances intake in a meal and contributes to the development of obesity in the rat. Physiol. Behav. 31:21-27; 1983. Rothwell, N. J.; Stock, M. J. Regulation of energy balance in two models of reversible obesity in the rat. J. Comp. Physiol. Psychol. 93:1024--1034; 1979. Rowe, E. A.; Rolls, B. J. Effects of environmental temperature on dietary obesity and growth in rats. Physiol. Behav. 28:219-226; 1982. Rowland, N.; Engle, D. J. Hypothalamic hyperphagia prevented by prior subdiaphragmatic vagotomy: Insulin hyperphagia is unaffected.

L I P O S T A T I C T H E O R Y 123
Physiol. Behav. 21:685-689; 1978. 47. Russek, M. Semi-quantitative simulation of food intake control and
weight regulation. In: Booth, D. A., ed. Hunger models: Computable theory of feeding control. New York: Academic Press; 1978:195-226.
48. Salmon, D. M. W.; Flatt, J. P. Effect of dietary fat content on the incidence of obesity among ad libitum fed mice. Int. J. Obes. 9:443--449; 1985.
49. Sclafani, A. Carbohydrate taste, appetite, and obesity: A overview. Neurosci. Biobehav. Rev. 11:131-153; 1987.
50. Sclafani, A.; Gorman, A. N. Effects of age, sex, and prior body weight on the development of dietary obesity in adult rats. Physiol. Behav. 18:1021-1026; 1977.
51. Sclafani, A.; Vigorito, M.; Pfeiffer, C. L. Starch-induced overeating and overweight in rats: Influence of starch type and form. Physiol. Behav. 42:409--415; 1988.
52. Sclafani, A.; Xenakis, S. Sucrose and polysaccharide induced obesity in the rat. Physiol. Behav. 32:169-174; 1984.
53. Sclafani, A.; Xenakis, S. Influence of diet form on the hyperphagia- promoting effect of polysaccharide in rats. Life Sci. 34:1253-1259;
1984. 54. Slattery, J. M.; Potter, R. M. Hyperphagia: A necessary precondition
to obesity? Appetite 6:133-142; 1985. 55. Steffens, A. B. Influence of reversible obesity on eating behavior,
blood glucose, and insulin in the rat. Am. J. Physiol. 228:1738-1744; 1975.
56. Stephens, D. N. Growth and the development of dietary obesity in adulthood of rats which have been undernourished during develop- ment. Br. J. Nutr. 44:215-227; 1980.
57. Stunkard, A. J. Anorectic agents lower a body weight set point. Life Sci. 30:2043-2055; 1982.
58. Triscari, J.; Nauss-Karol, B. E.; Levin, B. E.; Sullivan, A. C. Changes in lipid metabolism in diet-induced obesity. Metabolism 34:580-587; 1985.
59. Van itallie, T. B.; Kissileff, H. R. Physiology of energy intake: an inventory control model. Am. J. Clin. Nutr. 42:914-923; 1985.
60. Wade, G. N. Sex hormones, regulatory behaviors, and body weight. Adv. Stud. Behav. 6:201-279; 1976.