hyperphagia, polydipsia, and other effects of hypothalamic lesions in the white-throated sparrow
TRANSCRIPT

HYPERPHAGIA, POLYDIPSIA, AND OTHER EFFECTS OF HYPOTHALAMIC LESIONS IN THE WHITE-THROATED SPARROW, ZONOTRICHIA ALBICOLLIS
WAYNE J. KUENZEL
AND
CARL W. HELMS
Department of Zoology University of Georgia Athens, Georgia 30601
Many populations of migratory birds show a conspicuous increase in body weight due to the deposition of fat prior to or during migra- tion (King and Famer 1965). Kuenzel and Helms (1967) recently demonstrated that elec- trolytic lesions, properly placed in the ventro- medial area of the hypothalamus of the White- throated Sparrow (Zonotrichia albicollb) led to marked weight gains. These increases were comparable in rate and magnitude to normal premigratory increases found in captive repre- sentatives of this species in spring, implicating the ventromedial hypothalamic region in the regulation or control of vernal premigratory hyperphagia and fattening.
These studies have been continued on the White-throated Sparrow. The results presented in this paper verify and extend the earlier find- ings showing that lesion-induced weight in- creases are repeatable phenomena, and that these gains are based upon increased food intake or hyperphagia leading to a massive fat deposition. However, increased food intake in lesioned birds is often accompanied by an increased water intake or polydipsia and varia- bly associated with disturbances of several facets of the annual cycle, including Zugun- ruhe or nocturnal activity, gonadal develop- ment, and molt.
METHODS
White-throated Sparrows were captured in central Pennsylvania during fall migration in 1965 and in northeastern Georgia during fall migration in the following year. Individuals were color-banded and placed singly in cages (Hendryx SB-Special) in indoor aviaries (in Pennsylvania in 1965, Georgia in 1966 ) in which mean air temperature varied between 15-21°C mean relative humidity ranged be- tween 25-50 per cent, and photoperiod matched that normal to the season and local- ity. All birds were given food (“layena” in Pennsylvania, “pro-lay eggena” in Georgia ) and water ad Zibitum. Individuals were cage- adapted for at least 30 days before surgery.
SURGICAL TECHNIQUES
Location of the ventromedial hypothalamic area. We employed a stereotaxic instrument (Baltimore Instrument Co.) with a headholder modified for small birds. To date, no stereotaxic atlas of the brain of small passerine birds is available. It was therefore necessary to devise a means for relating stereotaxic coordinates with designated loci of the hypothalamus.
Figure 1 shows the skull of a White-throated Spar- row positioned in the stereotaxic headholder. The theoretical axes (two horizontal and one vertical) shown pass through a common point which represents the horizontal zero reference. The posterior region of the skull was fixed by inserting the ear bars into the external auditory canals. The mouth bar, posi- tioned at the same level as the ear bars and directly under the lower mandible at the commissural angle of the bill, was used as the anterior fixation point of the skull. The vertical zero was taken at the point perpendicular to the horizontal plane where the elec- trode tip touched the exposed skull.
A group of skulls with intact brains was fcnestrated and fixed in 10 per cent buffered formalin. The dorsal, posterior region of the skull was carefully re- moved and the heads were oriented in the headholder as described above. A razor blade was inserted into the electrode carrier. Successive 0.61.0-mm macro- cross-sections of the brain were cut proceeding an- teriorly from the cerebellum, placed on slides, cover- slipped, and viewed unstained under a microscope until sectioning reached the ventromedial hypothalamic area. The razor blade was then removed and replaced with an electrode which was positioned in the exposed area of the brain. Coordinates were recorded (Kuenzel 1966). Lesions produced as described below were then placed medially and bilaterally throughout the hypothalamic volume in a series of birds (Group P, see below). Refined coordinates yielding hyperphagia or other effects are given in table 1 for all birds in- cluded in this report.
Constructing microelectrodes. Stainless steel insect pins (Wards #l ) were used as electrodes. After being dinoed successivelv in dilute sulfuric acid. dis- tilled water, and acetone; they were coated with three layers of insl-x ( Insl-X Products Corp.). Before and after each operation the insulation of each electrode was checked for leakage by passing a current through the electrode while it was immersed in egg albumin.
Lesi0nin.g. A bird was weighed, then injected intra- muscularly with Equithesin (Jensen-Salsbery Labora- tories). The dosage rate was computed from the quantity normally given to chickens, 2.5 ml/kg (Lumb 1963). The bird was then fitted into the headholder of the stereotaxic instrument and the skin covering the skull was cut and retracted. The electrode was posi- tioned at preselected coordinates and lowered until it
The Condor, 72:66-75, 1970 66

EFFECTS OF HYPOTHALAMIC LESIONS 67
FIGURE 1. Skull of White-throated Sparrow in stereotaxic headholder with theoretical axes and zero reference I indicated.
touched the top of the exposed skull. A small hole was then made through the bone at this point using a dentist’s drill and the electrode was lowered to a predetermined depth. A single midline anodal lesion (1 ma, 15 set) or, in several birds of the first series, bilateral anodal lesions (1 ma, 8 set) were administered to each bird. Following operation, lesioned birds were returned to their individual cages.
EXPERIMENTAL GROUPS
Data to be presented in this paper were taken from three experimental series. Croup PI (24 birds lesioned between 31 October 1965 and 9 January 1966) con- tained 11 birds (9 Q 0, 2 $ 8) that responded with im- mediate and conspicuous weight gains (P,,) and 13 birds showing no weight change (P,,). Results from this group of Pennsylvania birds have been given by Kuenzel and Helms (1967) and will not be repeated in detail. Group G, (30 birds lesioned between 12 and 15 February 1967) contained 6 responders (3 0 0,3 d 8) or lesioned-experimentals (GE) and 24 non-responders or lesioned-controls (Gc). This group of Georgia birds will receive major attention. Group Gz (11 birds from group (G,) relesioned between 22 and 23 April 1967) contained 5 experimentals (4 0 0, 1 6 ) (Gz~) and 6 controls (G~c).
Body weight ( & 0.02g) and visible fat were deter- mined, using the fat class units of Helms and Drury ( 1960). Fat class 6 was added to this scheme and used for the following condition: the mounded furcular ( or claviculo-corocoid ), lateral thoracic, and transverse abdominal fat organs (see King and Farner 1965) en-
I
FIGURE 2. Metabolism-activity cage. The beaker at the left of the cage contains a weighed amount of food of known moisture content and caloric value which is used to fill the food cup daily. The wire bottom under the cage prevents the bird from trampling and mixing food and feces, facilitating the later separation of these materials which collect in the aluminum pan. Plastic skirts around the lower half of the cage retard losses of food and fecal material. The graduated water fountain on the right of the cage supplies water ad libitum and allows consumption to be determined ( kO.2 ml). Two movable perches counterweighted to the left of the cage rest on microswitches at the right and sample locomotor activity. Dimensions of the cage are 16 inches (depth) x 8% inches (width) x 10% inches (height).
croach upon and begin to overlie the pectoral muscles which may contain sufficient fat to appear conspicu- ously lightened in color. One GE bird had a fat index of 0.7 (extracted lipid/dry fat-free weight) for the pectorial muscle. Note, however, that fatty muscle has been seen only in lesioned birds and may be the result of diabetes insipidus in some of the individuals (Lepkovsky and Yasuda 1966). Conspicuous muscle lightening is not required for a designation of fc 6.
Molt and cloaca1 development were noted weekly (or biweekly in some PI birds). Nocturnal activity was recorded daily for some individuals in all three groups. Nine of the G birds (four GIE and five Gc) were placed in metabolism-activity cages (fig. 2). Energy balance determinations of metabolism were recorded on a weekly basis by methods similar to those used by King ( 1961). Water consumption was read weekly from the graduated water fountains shown in figure 2 and corrected for evaporative loss. Sub- cutaneous and peritoneal lipids of four of the fattest
TABLE 1. Stereotaxic coordinates.”
Horizontal coordinates
Lateral scale (mm) Rostrocaudal scale (mm)
Successful coordinates Bilateral lesions -co.25 -2.0 Single lesions &O.OO -2.0 to -2.5
mean = -2.25 Unsuccessful coordinates
All lesions > +0.5, < -0.5 >-1.5, G-3.0
D See text for explanation of zero references.
Vertical coordinates
Depth scale (mm)
6.0 6.0 to 8.0 mean = 7.0
39.0, G5.5
68 WAYNE J. KUENZEL AND CARL W. HELMS
4o t I
I- i FEB MAR APR MAY JUN
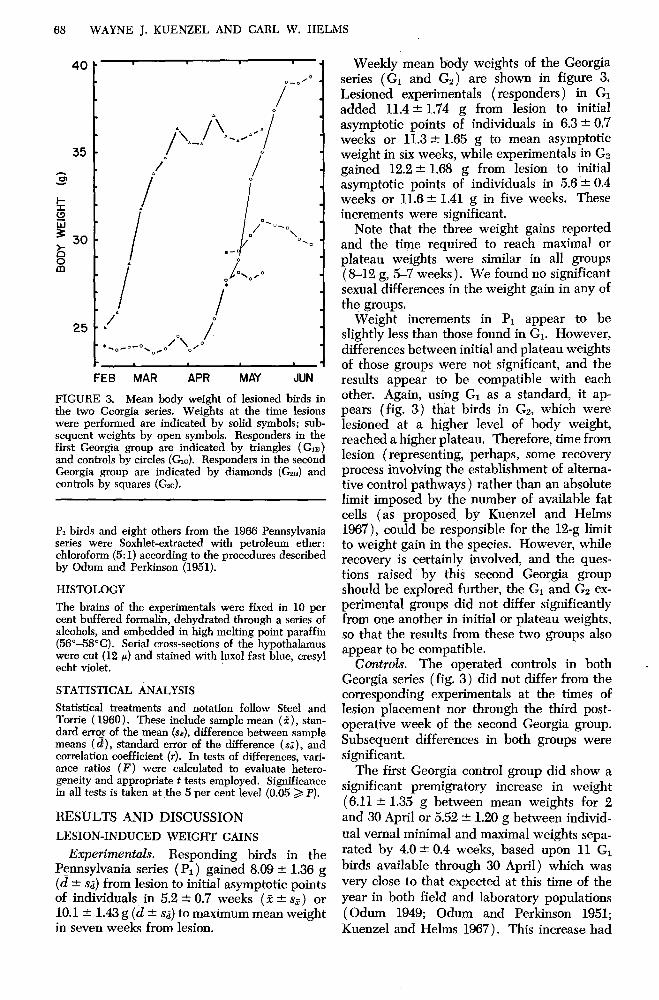
FIGURE 3. Mean body weight of lesioned birds in the two Georgia series. Weights at the time lesions were performed are indicated by solid symbols; sub- sequent weights by open symbols. Responders in the first Georgia group are indicated by triangles (G,) and controls by circles (Gc). Responders in the second Georgia group are indicated by diamonds (G,n) and controls by squares (G*c).
P1 birds and eight others from the 1966 Pennsylvania series were Soxhlet-extracted with petroleum ether: chloroform (5: I) according to the procedures described by Odum and Perkinson (1951).
HISTOLOGY
The brains of the experimentals were fixed in 10 per cent buffered formalin, dehydrated through a series of alcohols, and embedded in high melting point paraffin (56”-58”C). Serial cross-sections of the hypothalamus were cut (I2 p) and stained with 1~x01 fast blue, cresyl echt violet.
STATISTICAL ANALYSIS
Statistical treatments and notation follow Steel and Torrie ( 1960). These include sample mean (?), stan- dard erroz of the mean (sr), difference between sample means (d), standard error of the difference (si), and correlation coefficient (7). In tests of differences, vari- ance ratios (F) were calculated to evaluate hetero- geneity and appropriate t tests employed. Significance in all tests is taken at the 5 per cent level (0.05 > P).
RESULTS AND DISCUSSION
LESION-INDUCED WEIGHT GAINS
Expmhentals. Responding birds in the Pennsylvania series (P,) gained 8.09 2 1.36 g (d * 8;) from lesion to initial asymptotic points of individuals in 5.2 + 0.7 weeks (1 k q) or 10.1 2 1.43 g (d * s,J to maximum mean weight in seven weeks from lesion.
Weekly mean body weights of the Georgia series (Gi and Gs) are shown in figure 3. Lesioned experimentals (responders) in GI added 11.4 + 1.74 g from lesion to initial asymptotic points of individuals in 6.3 + 0.7 weeks or 11.3 f 1.65 g to mean asymptotic weight in six weeks, while experimentals in GP gained 12.2 & 1.68 g from lesion to initial asymptotic points of individuals in 5.6 + 0.4 weeks or 11.6 + 1.41 g in five weeks. These increments were significant.
Note that the three weight gains reported and the time required to reach maximal or plateau weights were similar in all groups (8-12 g, %7 weeks). We found no significant sexual differences in the weight gain in any of the groups.
Weight increments in P1 appear to be slightly less than those found in Gi. However, differences between initial and plateau weights of those groups were not significant, and the results appear to be compatible with each other. Again, using G1 as a standard, it ap- pears (fig. 3) that birds in G2, which were lesioned at a higher level of body weight, reached a higher plateau. Therefore, time from lesion (representing, perhaps, some recovery process involving the establishment of altema- tive control pathways) rather than an absolute limit imposed by the number of available fat cells (as proposed by Kuenzel and Helms 1967), could be responsible for the 12-g limit to weight gain in the species. However, while recovery is certainly involved, and the ques- tions raised by this second Georgia group should be explored further, the Gi and Gs ex- perimental groups did not differ significantly from one another in initial or plateau weights, so that the results from these two groups also appear to be compatible.
C0nt~ol.s. The operated controls in both Georgia series (fig. 3) did not differ from the corresponding experimentals at the times of lesion placement nor through the third post- operative week of the second Georgia group. Subsequent differences in both groups were significant.
.
The first Georgia control group did show a significant premigratory increase in weight (6.11 -t 1.35 g between mean weights for 2 and 30 April or 5.52 it 1.20 g between individ-
ual vernal minimal and maximal weights sepa- rated by 4.0 2 0.4 weeks, based upon 11 G1
birds available through 30 April) which was very close to that expected at this time of the year in both field and laboratory populations (Odum 1949; Odum and Perkinson 1951; Kuenzel and Helms 1967). This increase had

EFFECTS OF HYPOTHALAMIC LESIONS 69
FIGURE 4. Representative indoor winter (left), migratory (middle ), and lesion-induced (right) levels of depot fat. Dissections of individuals with fat classes of 2, 4, and 6 (from left to right) are shown: upper picture, skin opened ventrally; middle picture, carcass with skin and some subcutaneous fat removed; lower picture, thoracic and abdominal regions with sternum and ventral abdominal walI (and peritoneal fat) re- moved. Note the absence of excessive fat around the heart of the most obese individual dissected (see Odum and Perkinson 1951).
been essentially completed by birds in the sec- ond group at the time of lesion placement.
LESION-INDUCED FAT DEPOSITION
Based upon changes in visible fat, Kuenzel and Helms (1967) suggested that fat deposi- tion was primarily responsible for the gains in body weight observed in Pennsylvania birds.
In the first Georgia series, lesion and initial asymptotic fat class ( fc) values for individuals averaged 2.79 and 5.17 respectively. Using the equation relating body weight and fat class given by Kuenzel and Hehns ( 1967), the pre-
dieted weight difference in Gi birds would have been 10.7 g (observed = 11.4 g). In the second Georgia series, lesion and initial asymp- totic fat values for individuals averaged re- spectively 3.72 and 5.60 fc. Using the same equation, the expected weight difference in Ga birds would have been 10.4 g (observed = 12.2 g). This agreement demonstrates that the bulk of the weight increment in lesioned birds is attributable to fat deposition.
However, to illustrate the magnitude of fat that can be added following hypothalamic lesions, and to contrast it with two “normal” levels of depot lipid, four of the fattest birds in the Pennsylvania series, along with four cap- tives judged to contain vernal migratory levels of fat and four with low reserves typical of non-responding birds in winter, were selected and dissected. Representatives are shown in figure 4. The skin and feathers containing sub- cutaneous fat and the ventral abdominal wall containing peritoneal fat were removed from these 12 birds (see Odum and Perkinson 1951), dried, and extracted.
Results of this selection and extraction are summarized in table 2. This table also contains predicted total fat values for each group of birds extracted. The first of these predictions is based upon the equation given by Kuenzel and Helms ( 1967). The second is based upon an unpublished recalculation of the 1951 data of Odum and Perkinson (log,, TF = 0.811 + 0.480 log,, SF * 0.0824, where TF = total fat and SF = sectional fat in the abdominal wall, calculated for birds with TF > 2.0 g) . It is ap- parent that the extrapolation of this equation to high fat levels is in error, however, since the fat in the skin and peritoneum alone of the lesioned birds nearly equals the estimate. The third prediction is based upon the equation given by Odum and Perkinson ( 1951) for sub- cutaneous lipid. The final column of this table, based upon median values of predicted total fat, suggests a relative measure of depot lipid in the three groups that were extracted. The excessive amounts of fat in the selected lesioned birds are evident.
ENERGETICS OF WEIGHT GAIN AND WATER CONSUMPTION IN LESIONED RESPONDERS
Weight gain and fat deposition. Weekly means of body weight of the first Georgia group are plotted in figure 5 so that differ- ences between those birds used for energy balance study and others in the series can be compared. Since none of these differences were significant, the individuals employed ap- pear to constitute a representative sample of the total group.

70 WAYNE J. KUENZEL AND CARL W. HELMS
TABLE 2. Sectional fat extractions.
Normal control fat Normal migratory fat Lesioned-induced fat
n Wet weight (g) (f z!z SE)” Visible fat (median fc)b Extracted fat (g)
Peritoneal (i 2 ss) Subcutaneous ( f & SE)
Predicted’ total fat (g) based upon: Visible Peritoneal Subcutaneous
Weight increase ( g) following lesioning Approx. g lipid/100 g body weight
4 4 4 25.18 c 0.67 32.10 f 0.35 41.48 f 1.45
2.38 4.66 5.38
0.0935 + 0.0133 0.0758 -+ 0.158
0.791 f 0.0838 2.79 f 0.322
2.90 ? 0.55 7.60 2 0.489
0.9 6.2 2.1 5.8 2.0 6.3
7.9 19.3
14.1 10.8 16.3 14.08 34.2
a Total live weight of birds before dissection. b fc = fat class units of Helms and Dnny ( 1960 ). e See text for derivation of predictions.
In this sample, as in the total group, both lesioned-responders and non-responders under- went a dynamic phase of weight gain (im- mediately following lesion placement in the former group, and between the seventh and eleventh week following lesion placement in
FEB MAR APR MAY FIGURE 5. Mean body weight of lesioned birds in the first Georgia series ( G) . Data are separated into two groups in this figure: birds used for energy bal- ance determinations of metabolism (solid circles on the lines) ; and others in the group (plain lines ). Up- per pair of lines represent data from experimentals ( responders,) ; lower lines, controls (non-responders ) .
the latter group). The response of the experi- mental birds was certainly due to the “proper” placement of the lesion. Following the dy- namic phase, responders maintained a plateau weight with only relatively minor fluctuations through the remainder of the experiment. This static phase represents a period in which weight (hence fat) is regulated at an abnor- mally high level as an effect or consequence of pathological interference with the neural regulating machinery of the individual. The response of the control birds, housed under seasonally normal photoperiods, constituted a normal response to the proximate timing action of increasing or increased day length (Wolfson 1954,1959; Farner 1955). Note that this phase is not shown by the lesioned-experimentals (but see below) who are, supposedly, already at their maximum fat levels (Kuenzel and Helms 1967).
The weight responses of the experimentals and controls as discussed above are replotted in figure 6 with data from the 14-week metab- olism experiment. To facilitate the following presentation of the energetic data plotted in this figure, the body weight curves of the ex- perimental and control birds will be used for reference.
Specifically, body weight records of experi- mentals were divided into two phases: phase a (E, ), lesion-induced dynamic phase of weight gain; and phase b ( Eb), lesion-induced static phase or plateau of weight regulation. The body weight records of controls were divided into three phases: phase a (C,), non- stimulated static phase of weight regulation; phase b (CJ, photo-induced dynamic phase of weight gain; and phase c (C,), photo-induced “static” phase of weight regulation.
Nocturnal activity, water consumption, and energetics. The results of the energy balance

25 -
FEB MAR APR MAY
FIGURE 6. Mean weight, activity, water consump- tion, and metabolism of lesioned-responders (solid circles on the lines) and lesioned-controls (plain lines) in the first Georgia series (G). BW = body weight. See text for explanation of phases a, b, and c. Solid triangles beneath data points denote the median weeks used in testing differences between the several re- sponse phases. NA = nocturnal activity; WC = water consumption; GE = gross energy; ME = metabolizable energy; EE = excretory energy; and UC = utilization coefficient.
EFFECTS OF HYPOTHALAMIC LESIONS 71
portion of this study are given in figure 6 and table 3 (body weight and nocturnal activity are omitted from the table during dynamic phases). Appropriate tests (determined by the homogeneity or heterogeneity of the samples) were made between means for the median week in each response phase (indicated in fig. 6 by solid triangles). Only significant differences determined by this valid but weak testing procedure will be mentioned in the following discussion of, results.
Body weight was significantly higher in the lesion-induced static phase of the experimental birds ( Eb) than in controls before (C,) or after (C,) vernal fat deposition (respective d = 11.7 * 1.52 g, and 6.14 * 1.26 g). During the photo-induced static phase of weight regu- lation (C, ) controls were significantly heavier than before their dynamic phase ( C,* ) when photoperiod was not overtly stimulating (d = 5.57 c 1.08 g).
Nocturnal activity recordings did not begin until 26 March, but initial values of lf%200 counts/hr are not unusual for indoor captives of this species. Nocturnal activity for unoper- ated White-throated Sparrows held indoors during the winter phase of the annual cycle averaged 111 ‘- 24.4 counts/hr, with a coeffi- cient of variation of 188 per cent (72 observa- tions of weekly means, unpublished data). Activity of control birds increased when photo- induced weight gain was about half completed (normal in unoperated controls). This occurred during the week of 23 April. White-throated Sparrows held outdoors in central Pennsyl- vania began nocturnal activity on 19.9 April * 2.3 days (17 records for males, unpublished observations). Activity peaked in early May
TABLE 3. Body weight, nocturnal activity, energy balance, and water consumption” after lesion placement.
Experimentals CO~tIOlS
E&~$$~S E, Static phase C, Static phase C, Dynamic phase
(photo-induced) C, Static phase
No. determinations 24 32 30 25 15 Body weight (g) 35.06 ” 0.4494 22.97 & 0.3544 28.21 t 0.3444 Nocturnal activity
(counts/hr) 46.6 k 21.4 598. f 102. Gross energy
( kcal/day ) 20.60 & 0.5290 18.86 rt 0.05805 15.27 r 0.2588 19.41 & 0.7655 21.04 f 0.9515 Metabolizable
energy (kcal/day) 15.71 -t 0.4082 14.56 ? 0.2818 11.47 r 0.1886 14.51 e 0.5306 15.57 -c 1.5561 Excretory energy
(kcal/day ) 4.893 * 0.1750 4.300 -+ 0.07829 3.80 -c 0.08268 4.902 ‘- 0.2418 5.472 k 0.4163 Utilization
coefficient ( % ) 76.3 & 0.522 77.1 r 0.312 75.2 & 0.246 74.9 -c 0.288 74.41 & 0.778 Water consumption
( ml/day 1 32.1 + 2.35 22.6 ? 0.873 6.05 ? 0.199 7.21 k 0.262 6.29 k 0.978
a.+*s*.

72 WAYNE J. KUENZEL AND CARL W. HELMS
FIGURE 7. Trace drawings from cross-sections of brains of White-throated Sparrows made using a camera lucida. Lesions (mean widths and lengths respectively = .32 and .59 mm) found in extremely polydipsic birds (A and B) and hyperphagic birds with mild polydipsia (C and D) are indicated by vertical parallel lines. ANT COM = anterior commissure; N ROT = nucleus rotundus; III = third ventricle; SUP DES = supraoptic decussation; OP CH = optic chiasma.
at about 600 counts/hr, as did the activity of unoperated captives held indoors (unpublished observations). The control pattern of activity appears to be representative of typical pre- migratory and migratory behavior in captives. The experimental birds were, however, in- active at night and only one bird showed typi- cal Zugunmhe or nocturnal activity (beginning on 17 April). This behavior of lesioned birds will be explored below.
Water consumption increased gradually in controls, averaging 5.2 ml/day during week 1 and 9.4 ml/day when the experiment was terminated. It was significantly higher during the dynamic phase (C,) than during the first six weeks (C,) of the experiment (1.37 f 0.590 ml/day), but differences between the periods C, and C, (3.24 k 2.19 ml/day) and between C, and Cb (1.87 5 2.20 ml/day) were insignificant. Consumption during C, was more variable, with one individual increasing his consumption to 17.8 ml/day by mid-May. Drinking was sig-
nificantly correlated with gross energy intake or feeding (r = +0.8).
In contrast, lesioned responders showed a
marked polydipsia, drinking more than their body weight per day for the first three weeks ( 35.0 ml/day). In the Eb phase, however, con- sumption dropped significantly (-13.9 k 5.44 ml/day) and appears to have been approaching normal levels toward the end of the experiment. Nevertheless, water consumption of responders was significantly above that of non-responders throughout the experiment. Moreover, based upon three responders and five controls main- tained on record until the following 17 Decem- ber, initial responders appeared to retain an elevated water intake of about 7 ml/day over control levels. Therefore the lesion-damage to control mechanisms was probably permanent. As expected, polydipsic birds were polyuric. Water consumption of responders was also significantly correlated with food intake (r = +0.6).
Metabolic estimates plotted in figure 6 in- clude gross energy (the caloric value of food ingested), excretory energy (the caloric value of urinary and fecal wastes), and metabolizable energy (calculated by subtracting excretory from gross energy) or the calories available to

EFFECTS OF HYPOTHALAMIC LESIONS 73
the organism, including those supposedly lost through the calorigenic effect or specific dy- namic action (King 1961; King and Farner 1961). Excretory energy will not be discussed in detail.
Gross and metabolizable energy were highest and reached comparable values in responders and controls during the lesion- and photo- induced dynamic phases, respectively. These values decreased in experimentals as the Eb plateau was approached. They leveled out in controls with the attainment of migratory fat reserves. Reductions in gross (4.02 +- 1.49 kcal/day ) and metabolizable energy (-3.09 + 1.23 kcal/day ) were significant in experi- mentals between the dynamic and static phases. Increases in gross (5.02 + 1.65 kcal/ day) and metabolizable energy (3.50 f 1.15 kcal/day) were significant in controls between C, and C, phases. Metabolizable energy was higher in E, birds than in C, ones (1.29 2 0.394 kcal/day). With the tests used, other differ- ences were not detected. Lesion- and photo- induced hyperphagia evident in these data will be discussed below.
Utilization coefficients were calculated by dividing metabolizable energy by gross energy and multiplying the result by 100. There were no significant differences between groups or phases in this variable. Calories deposited as fat, therefore, must have come from increased food intake and not from gross alterations in the way in which food was utilized by the organism ( King 1961) ,
LOCATION OF LESIONS
The brains of experimental birds were exam- ined for the location of lesions. Generally speaking, a lesion which effected a fattening response was found to be at the base of the third ventricle below the anterior commissure, and above the optic chiasma and supraoptic decussation at an anterior-posterior level be- tween the supraoptic nuclei and median emi- nence.
A difference in lesion placement was noticed between those birds which were extremely polydipsic with delayed hyperphagia and those which immediately became hyperphagic with only mild polydipsia. It can be seen that two birds (A and B of fig. 7) representative of the former group had lesions which disrupted the supraoptic decussation as well as a few fibers of the optic chiasma. These lesion placements suggest that neurosecretory products from the supraoptic nuclei were blocked from entering the posterior pituitary. This in turn would prevent one of these secretory products, anti- diuretic hormone ( ADH), from eventually
1 I
25 1 FEB MAR APR MAY JUN
FIGURE 8. Mean nocturnal activity and body weight of lesioned-responders with recorded activity. Data are separated on the basis of a conspicuous nocturnal activity (plain lines) or no nocturnal activity (solid circles on the lines). Those points, to the left of the first triangle are from the first Georgia group (G) only. Between the first and second triangle, data from G and the Pennsylvania series ( P1) are combined. To the right of them second triangle, data from all three groups (PI, G, and GI) are given.
reaching the kidney tubules for the active re- absorption of tissue fluids (see McFarland 1959; Anand 1961; Wayner 1964). The polyuric response typical of these birds supports this hypothesis. In contrast two birds which were representative of the second group showing immediate hyperphagia but mild polydipsia had shallower lesions. In these two birds (C and D of fig. 7) the supraoptic decussation was intact.
DEPOT FAT AND NOCTURNAL ACTIVITY
Data from all lesioned responders in the Penn- sylvania and Georgia series for which we have activity records were used to plot figure 8 which illustrates what appears to be a dichot- omy within this group in activity and weight during the period of spring migration. Note that these records begin with representatives of only the first Georgia group ( G1), but that Pennsylvania individuals (which were lesioned earlier) are added to the means in the week after the first solid triangle at the base of the graph, and that the second Georgia group (which was lesioned later) is added on the week after the second solid triangle. All test- ing was done only within or between the same or corresponding groups of individuals.
A total of eight responders showed normal

74 WAYNE J. KUENZEL AND CARL W. HELMS
nocturnal activity, beginning in mid- to late April and peaking at 666-700 counts/hr in early May. Late March weights of this group were high as a result of the lesion placement (about 33 g) but during April this group added 3.11 2 1.34 g. This was a significant increment although only half of that added by controls (over 6 g, see fig. 6). Moreover, from the onset of conspicuous nocturnal activity to early June, members of this group lost a significant amount of weight (-3.33 * 1.06 g) paralleling results found in studies of both indoor and outdoor captives of this species (unpublished data). These lesioned birds showing nocturnal activity appeared to possess rudiments of the normal seasonal patterns of weight and fat variation.
However, 10 responders showed no noctur- nal activity through spring migration. The changes in body weight between the sixth and seventeenth week of record (2.16 f 1.69 g) were not significant. No convincing signs of either a photo-induced dynamic phase or trace of a loss in weight in late spring were evident in this experimental group.
The two groups of experimental birds did not differ in weight at lesion (1.92 * 1.42 g). The group with no nocturnal activity was sig- nificantly heavier (3.77 * 1.43 g) than the group with normal nocturnal activity by the sixth week, suggesting a more marked and rapid response to lesioning and perhaps a more profound (or extensive) effect of the surgical procedure in the former group.
GONADAL EFFECI’S
Four of five males in the Georgia series that responded to lesioning with conspicuous weight gain showed no signs of cloaca1 de- velopment (see Wolfson 1952) through the nor- mal breeding season. One male showed a func- tional development. Disruption of gonadal cycles is to be expected following certain ven- tromedial hypothalamic lesions (Lepkovsky and Yasuda X%6), especially if the nucleus of origin of the tubero-infundibular tract or the entry of the tubero-infundibular fibers into the median eminence is destroyed (Wilson 1967).
MOLT
Of 10 of the Georgia experimentals held through postnuptial molt, seven had no molt or a molt restricted to some body feathers and a few of the flight feathers, while three under- went massive and virtually simultaneous re- placement of all contour feathers. However, the accelerated molt may not have been sig- nificant since, for unknown reasons, many
controls underwent a similarly abnormal period of feather replacement. On the other hand, it does appear that lesions can inhibit normal feather replacement in some birds. This is verified by examination of prenuptial molt in the first Georgia group. Less than half of the responders (two of six) showed signs of sig- nificant feather replacement, while 91 per cent of the non-responders molted (range of weeks, 26 February-S May; median and mode-week ending on 21 April).
SYNDROME INDUCED BY VENTROMEDIAL HYPOTHALAMIC LESIONS
Records of weight, water consumption, activity, cloaca1 development, and molt (through the post-nuptial period) were available for five experimental males in the Georgia series (Gr and Gz). Four of these birds were hyperphagic and polydipsic and showed little or no noc- turnal activity, cloaca1 development, or molt. One male was hyperphagic and polydipsic but did have normal nocturnal activity and gonadal development and an accelerated molt. Similar records were available for three responding females. None showed conspicuous cloaca1 de- velopment, but this is not unusual in captives. One was hyperphagic and polydipsic, showed no nocturnal activity, and had a reduced molt. The other two were hyperphagic, but not poly- dipsic, had no nocturnal activity, and under- went an accelerated molt. We have found other individuals, including two G1 and one Gz controls that were polydipsic but not hy- perphagic.
Although these samples are small, they do indicate that the effects of ventromedial lesions in the White-throated Sparrow are variable. Several ethophysiological events may be dis- rupted but these disturbed functions are not invariably found together in the same indi- viduals. We assume that these differences in response are related to exact placement and extent of destruction by electrocautery. This assumption is currently under further investi- gation.
SUMMARY
Destruction of the ventromedial area of the hypothalamus in the White-throated Sparrow by electrocautery produced a variety of etho- physiological changes. A conspicuous hyper- phagia resulted in a dynamic phase of fat deposition and weight gain taking 5-7 weeks to attain plateau levels 8-12 g above those existing before surgery. Food intake was slightly reduced but remained elevated at plateau. Polyuria and polydipsia accompanied, in many individuals, the dynamic phase of

EFFECTS OF HYPOTHALAMIC LESIONS 75
weight gain. Water consumption was reduced KING, J. R., AND D. S. FARNER. 1965. Studies of fat
with time but appeared to remain permanently deposition in migratory birds. Ann. New York
above control levels. A suppression of further Acad. Sci. 131( 1) :42%440.
KUENZEL, W. J. 1966. Production of obesity in the
weight changes, nocturnal activity, cloaca1 de- velopment, and molt, occurred.
It is possible that some part of the central “clock,” timing and synchronizing the annual KUENZEL, W. J., AND C. W. HELMS. 1967. Obesity
cycle, may be located in the ventromedial hy- produced in a migratory bird by hypothalamic lesions. Bioscience 17:395-396.
White-throated Sparrow, Zonotrichia albicollis, by electrocauterization of the ventromedial hy- pothalamus. M. S. thesis, Bucknell Univ., Lewis- burg, Pennsylvania. 71 p.
pothalamus. LEPKOVSKY, S., AND M. YASUDA. 1966. Hypothalamic lesions. growth. and bodv comuosition of male
ACKNOWLEDGMENTS chickens, Poultry Sci. 45:582-588. LUMB, WILLIAM V. 1963. Small animal anesthesia.
Lea and Febiger, Philadelphia. MCFARLAND, L. 1959. Effects of an electrolytic
lesion in the avian hypothalamo-hypophyseal tract. Anat. Rec. 133:411. -( Abstr.). _
ODW. E. P. 1949. Weight variations in winterine
We would like to thank the staff of the Freas-Rooke Computing Center for their assistance; Mrs. Shirley Marshall for caloric determinations; S. D. Fretwell and J. T. McGinnis for statistical advice; S. D. Fret- well, J. E. Harclerode, and J. R. King for their critical evaluation of the manuscript; and L. F. Wilson for permission to collect birds on his property. This work was supported by Bucknell University, the University of Georgia, and with grants AM 06798 and HE 08294 ( USPHS ) and Environmental Health Training Grant TO1 ES06074 ( USPHS ). Portions of this study were undertaken while Helms was on an NSF Science Faculty Fellowship.
White-throated Sparrows in relation to tempera: ture and migration. Wilson Bull. 61:3-14.
ODUM, E. P., AND J. D. PERKINSON, JR. 1951. Rela- tion of lipid metabolism to migration in birds. Seasonal variation in body lipids of the migratory White-throated Sparrow. Physiol. Zool. 24:21& 229.
LITERATURE CITED
STEEL, R. G., AND 1. H. TORRIE. 1966. Princinles and procedures-of statistics. McGraw-Hill, New York.
ANAND, B. D. 1961. Nervous regulation of food in- take. Physiol. Rev. 41: 677-708.
FARNER, D. S. 1955. The annual s.timulus for migra- tion: experimental and physiological aspects. p. 198-237. In A. Wolfson [ea.] Recent studies in avian biology. Univ. of Illinois Press, Urbana.
HELMS, C. W., AND W. H. DRURY, JR. 1966. Winter and migratory weight and fat: field studies on some North American buntings. Bird-Banding 31: l-40.
KING, J. R. 1961. The bioenergetics of vernal pre- migratory fat deposition in the White-crowned &arrow. Condor 63:128-142.
KING,- J. R., AND D. S. FARNER. 1961. Energy metabolism, thermoregulation, and body tem- perature, p. 215-288. In A. J. Marshall [ed.] Biology and comparative physiology of birds. Vol. II. Acad. Press, New York and London.
WAYNN~~~ M.& [ed.] 1964. Thirst. Macmillan Co.,
WILSON, F. E. 1967. The tubero-infundibular neu- ron system: a component of the photoperiodic con- trol mechanism of the White-crowned Snarrow, Zorwtrichia bucophrys gambelii. Z. Zellforsch: 82: l-24.
WOLFSON, A. 1952. The cloaca1 protuberance. Bird- Banding 23: 159-165.
WOLFSON, A. 1954. Weight and fat deposition in relation to spring migration in transient White- throated Sparrows. Auk 71:413X34.
WOLFSON, A. i959. Ecologic and physiologic factors in the regulation of spring migration and repro- ductive cycles in birds. D. 38-70. In A. Gorbman [ea.] Comparative endocrinology. Wiley, New York.
Accepted for publication 9 December 1968.