dna damage,gadd153expression, and cytotoxicity in plateau-phase renal proximal tubular epithelial...
TRANSCRIPT

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 341, No. 2, May 15, pp. 300–308, 1997Article No. BB979969
DNA Damage, gadd153 Expression, and Cytotoxicityin Plateau-Phase Renal Proximal Tubular EpithelialCells Treated with a Quinol Thioether1
Jeongmi K. Jeong,2 Erik Dybing,* Erik Søderlund,* Gunnar Brunborg,* Jorn A. Holme,*Serrine S. Lau, and Terrence J. Monks3
Division of Pharmacology and Toxicology, College of Pharmacy, University of Texas at Austin, Austin, Texas 78712;and *Department of Environmental Medicine, National Institute of Public Health, P.O. Box 4404 Torshov,N-0403, Oslo, Norway
Received January 29, 1997
Although the addition of 3-aminobenzamide following2-Br-bis-(GSyl)HQ exposure has no effect on the re-2-Bromo-bis-(glutathion-S-yl)hydroquinone [2-Br-moval of DNA SSB, it causes a slight but significantbis-(GSyl)HQ] causes DNA single-strand breaks (SSB),increase in gadd153 expression and cell viability, indi-causes growth arrest, induces the expression ofcating that activation of poly(ADP-ribose)polymerasegadd153 (a gene inducible by growth arrest and DNAmay exacerbate toxicity. Finally, aurintricarboxylicdamage), and decreases histone H2B mRNA in log-acid did not prevent DNA SSB or cytotoxicity in 2-Br-phase renal proximal tubular epithelial cells (LLC-bis-(GSyl) HQ-treated LLC-PK1 cells, implying that ac-PK1). Renal epithelial cells in vivo normally exhibit ativation of endonucleases does not play a role in theselow mitotic index, therefore experiments in both pla-processes. q 1997 Academic Pressteau- and log-phase cells are necessary for a compre-
Key Words: quinone thioethers; reactive oxygen spe-hensive understanding of the stress response to 2-Br-cies; hydrogen peroxide; DNA damage; growth arrest;bis-(GSyl)HQ. In the present article we demonstratestress response; gadd153.that not all features of the stress response in log-phase
cells are reproduced in plateau-phase cells. Thus, al-though 2-Br-bis-(GSyl)HQ causes concentration andtime-dependent increases in DNA SSB, and increasesthe expression of gadd153, histone H2B mRNA levels Reactive oxygen species (ROS)4 may be involved inare unaltered in plateau-phase cells. The relationship the initiation and progression of a variety of humanbetween reactive oxygen species, DNA damage, gene diseases (1). Of particular importance is the interac-expression, and cytotoxicity was also investigated. tion of ROS with DNA, which results in a spectrum ofOur findings suggest that (i) 2-Br-bis-(GSyl)HQ-medi- structural alterations, including strand scission, baseated DNA damage in LLC-PK1 cells is mediated by the
pair mutations, deletions, rearrangements, and inser-generation of H2O2; (ii) DNA damage, either directlytions (2). In addition to the formation of ROS as nor-or indirectly, contributes to cell death; and (iii) DNAmal by-products of cellular metabolism, ROS are alsodamage, either directly or indirectly, provides the ini-frequently formed during the process of chemical me-tial signal for gadd153 expression. In addition, DNAtabolism (3). In this case, chemicals are converted torepair is rapid in LLC-PK1 cells, and the DNA-repairphenolic and polyphenolic metabolites. The latter caninhibitors 1-b-D-arabinofuranosylcytosine and hy-undergo oxidation to the corresponding quinone, fol-droxyurea have no effect on the amount of DNA SSB.
4 Abbreviations used: Ara-C, 1-b-D-arabinofuranosylcytosine;1 This work was made possible by Grant ES 07359 (T.J.M.) fromthe National Institute of Environmental Health Science. 2 -Br-bis - (GSyl)HQ, 2-bromo-bis - (glutathion -S -yl)hydroquinone;
DMEM, Dulbecco’s modified Eagle’s medium; EBSS, Earl’s balanced2 Present address: Division of Toxicology, M.I.T., 77 MassachusettsAve., 56-638, Cambridge, MA 02139-4307. salt solution; FBS, fetal bovine serum; Hepes, N-[2-hydroxyethyl]-
piperazine-N *-[2-ethanesulfonic acid]; ROS, reactive oxygen species;3 To whom correspondence should be addressed. Fax: (512) 471-5002. E-mail: [email protected]. SSB, single-strand breaks; O•0
2 , superoxide anion.
300 0003-9861/97 $25.00Copyright q 1997 by Academic Press
All rights of reproduction in any form reserved.
AID ABB 9969 / 6b33$$$421 04-21-97 10:35:13 arcas

301QUINOL THIOETHER-MEDIATED DNA DAMAGE AND CYTOTOXICITY
lowed by redox cycling with the concomitant genera- icity linked)? (iii) Are gadd153 expression and DNAdamage related?tion of semiquinone radicals and ROS (4). In this se-
quence, quinones undergo an enzyme-catalyzed(NADPH–cytochrome-P450 reductase, NADH–cyto- MATERIALS AND METHODSchrome-b5 reductase, or NADH–ubiquinone oxidore-
Chemicals and probes. Catalase (from bovine liver, thymol-free;ductase) one-electron reduction (4–9) to the semiqui- 15,840 U/mg), deferoxamine mesylate, bathocuproine disulfonic acidnone radical which can reduce molecular oxygen to (disodium salt), 1,10-phenanthroline, neutral red, 1-b-D-arabinofura-produce superoxide anion (O•0
2 ) with the regeneration nosylcytosine (Ara-C), hydroxyurea, 3-aminobenzamide, and aurin-tricarboxylic acid (ammonium salt) were obtained from Sigma Chem-of the quinone. The cytotoxicity of quinones involvesical Co. (St. Louis, MO). All other chemicals were purchased commer-the iron-catalyzed Haber–Weiss reaction, in whichcially and were of the highest grade available. 2-Br-bis-(GSyl)HQ
O•02 undergoes dismutation to form hydrogen peroxide was synthesized according to established methodology (19). Plasmid
(H2O2), and reduces Fe3/ to Fe2/. The H2O2 then reacts pBluescript containing a 600-bp fragment of gadd153 cDNA waskindly provided by Dr. Niki J. Holbrook (National Institute on Aging,with Fe2/ to generate the hydroxyl radical, which isGerontology Research Center, MD). The cDNA probe for histone H2Bprobably the reactive species responsible for DNAwas purchased from Oncor (Gaithersburg, MD). Dr. Sue Fisher (M.damage. Quinones are also electrophiles and they re- D. Anderson Cancer Center, TX) provided 7S ribosomal RNA cDNA.
act readily with cellular nucleophiles, particularly Procedures for cDNA probe preparation have been described else-where (16).protein and nonprotein sulfhydryls. Although the con-
Cell culture and treatment conditions. LLC-PK1 cells (Americanjugation of quinones with glutathione is usually asso-Type Culture Collection, Rockville, MD) are a renal proximal tubularciated with the process of detoxication, quinone thio-epithelial cell line derived from the New Hampshire Minipig, andethers retain the ability to redox cycle and generate were maintained in Dulbecco’s modified Eagle’s medium (DMEM)
ROS (10, 11) thereby facilitating quinone-induced tox- containing 10% fetal bovine serum (FBS) (Atlanta Biologicals, Nor-icity (12–14). cross, GA) and high glucose without pyruvate (GIBCO BRL,
Bethesda, MD) at 377C in a humidified incubator containing 5% CO2.In a model of quinone thioether-mediated toxicity,For all experiments, cells between passages 185 and 200 were used.2-bromo-bis-(glutathion-S-yl)hydroquinone [2-Br-bis-Cells were seeded at a density of 2 1 106 cells/100-mm dish and were
(GSyl)HQ] causes DNA single-strand breaks (SSB) in used after 7 days of growth, with medium changes on the fourthlog-phase renal tubular epithelial cells (LLC-PK1) (15) day. Cultures were washed twice with Earl’s balanced salt solution
(EBSS; concentrations in mM: 0.8 MgSO4r7H2O, 116.3 NaCl, 10.0and induces the expression of gadd153 (16), a stress-NaHCO3, 1.0 NaH2PO4rH2O, 5.4 KCl, 5.5 D-glucose, 20 Hepes, andresponse gene activated by DNA damage and growth1.8 CaCl2r2H2O) and were treated with 2-Br-bis-(GSyl)HQ in EBSS.
arrest. In preconfluent cell cultures, the cells are ac- For the gadd153 expression studies, cells were washed twice andtively synthesizing DNA and undergoing cell division, treated with the conjugates in DMEM (with Hepes, pH 7.4). Catalase
(10 U/ml), Ara-C (0.1 mM), and hydroxyurea (2 mM) were added toraising the possibility that the DNA in these cells iscell cultures concomitant with 2-Br-bis-(GSyl)HQ for 2 h. 3-Amino-more accessible, and therefore more sensitive, to ROS.benzamide (10 mM) was added to the cell cultures for 1 h immediatelyFollowing chemical challenge in vivo, the proportion following conjugate exposure (posttreatment). Deferoxamine mesy-
of cells in growth phase increases as the tissue at- late (10 mM), bathocuproine disulfonic acid (disodium salt) (10 mM),and 1,10-phenanthroline (0.2 mM) were added to cultures for 1 h andtempts to repair the initial injury, and log-phase cellsremoved prior to exposure of the cells to 2-Br-bis-(GSyl)HQ for 2 h.provide an important model for delineating the nature
Determination of cytotoxicity. Cytotoxicity was determined withof the stress response under these conditions. Becausethe neutral red assay and was performed by modification (15) ofrenal epithelial cells in vivo normally exhibit a lowpublished procedures (20, 21). Cells were washed twice with DMEM
mitotic index, experiments in both plateau- and log- containing Hepes without FBS and treated with 1 mL of 2-Br-bis-phase cells are necessary to fully understand the com- (GSyl)HQ (0, 50, 100, 200, 400, and 600 mM) for varying periods of
time (0, 0.5, 1, 2, and 5 h) in DMEM containing Hepes without FBS,plete nature of the stress response to 2-Br-bis-pH 7.4. After exposure to 2-Br-bis-(GSyl)HQ, the culture medium(GSyl)HQ. We report that not all features of the stresswas removed and the cells were incubated with neutral red (0.25response in log-phase cells are reproduced in plateau- mg/ml) at 377C in an incubator with 5% CO2 for 1 h. Neutral red
phase cells. was removed from the cells and cells were rinsed with washing/fixation solution (1% formaldehyde and 1% calcium). Neutral red,The cytotoxicity of 2-Br-bis-(GSyl)HQ is dependentwhich accumulates within lysosomes, was then extracted by a 15-upon the formation of H2O2 (15) and because there ap-min incubation with extraction solution (1% glacial acetic acid inpear to be differences in the association between H2O2, 50% ethanol) at room temperature. The absorbance of neutral red
DNA damage, growth inhibition, and cytotoxicity, de- was measured at 540 nm (Shimadzu UV-160). Each experiment wasperformed in quadruplicate and repeated at least three times.pending upon the cell system under investigation (17,
Northern blot analysis. Total RNA was isolated using RNAzol18), we extended our studies to ask the following ques-(Cinna/Biotecx Laboratory International, Inc., Friendswood, TX) andtions. (i) Is 2-Br-bis-(GSyl)HQ-mediated DNA damagethe procedure of Chomczynski and Sacchi (22). For Northern blotcaused by ROS (are DNA damage and ROS related)? analysis, RNA was electrophoresed in 1.0% agarose gel containing
(ii) Is the DNA damage of sufficient magnitude that it formaldehyde and was transferred to Zeta Probe membranes (Bio-Rad, Richmond, CA). cDNA probes were labeled with [32P]dCTP (ICN,contributes to cell death (are DNA damage and cytotox-
AID ABB 9969 / 6b33$$$421 04-21-97 10:35:13 arcas

302 JEONG ET AL.
thereafter. When cells sustain DNA damage, a batteryof genes is activated, including gadd153, which encodesa 19-kDa protein related to the CCAAT/enhancer-bind-ing protein family of transcription factors (26).Gadd153 probably acts as a negative regulator of theactivity of other transcription factors, playing an im-portant role in cell cycle or cell growth control. In-creases in gadd153 mRNA were observed at concentra-tions of 2-Br-bis-(GSyl)HQ of 200 mM and above (Fig.2). The expression of gadd153 was increased 30 minafter exposure of cells to 200 mM 2-Br-bis-(GSyl)HQ,and the increased expression was sustained for at least
FIG. 1. Concentration- and time-dependent formation of DNA sin- 5 h (Fig. 2). Interestingly, despite the upregulation ofgle-strand breaks in renal tubular epithelial cells following exposure
a gene involved in negative growth control, expressionto 2-Br-bis-(glutathion-S-yl)hydroquinone. LLC-PK1 cells were ex-of histone H2B mRNA, a replication-dependent histoneposed to either (A) varying concentrations of 2-Br-bis-(GSyl)HQ (0–
600 mM) for 30 min or (B) 400 mM 2-Br-bis-(GSyl)HQ for between 0 usually tightly coupled to DNA synthesis, was un-and 2 h, and DNA single-strand breaks were determined by alkaline changed (Fig. 3). The effect of 2-Br-bis-(GSyl)HQ onelution as described under Materials and Methods. the viability of LLC-PK1 cells was both concentration
and time dependent (data not shown). Significant (P õ0.05) decreases in cell viability were observed 30 min
Costa Mesa, CA) by the random-primer method (23) using a kit (Boeh- after exposure of cells to 200 mM 2-Br-bis-(GSyl)HQ,ringer Mannheim, Mannheim, Germany). All blots were hybridized and this concentration produced a 50% decrease in cellin 0.25 M Na2HPO4, pH 7.2, and 7% SDS, overnight at 657C. Blots
viability after 2 h. No further decreases in viabilitywere washed twice with 25 mM Na2HPO4, pH 7.2, and 5% SDS atoccurred in the next 3 h (data not shown).657C for 30 min and twice with 25 mM Na2HPO4, pH 7.2, and 1% SDS
at 657C for 30 min. All blots were rehybridized with the cDNA probe Are endonucleases, poly(ADP-ribosylation), and DNAfor 7S rRNA, to correct for potential variations in RNA loading, by repair involved in the response to 2-Br-bis-(GSyl)HQ?stripping the original probe with 0.11 SSC and 0.5% SDS at 957C for
To examine whether endonuclease(s) are involved in0.5–1 h. Autoradiography was performed using Kodak XAR-5 film(Sigma Chemical Co.) exposed for between 1 h and 5 days, depending the DNA damage caused by 2-Br-bis-(GSyl)HQ, LLC-on the probe, at 0707C with intensifying screens.
Quantitation of autoradiograms. The data from Northern blotanalysis were quantitated by densitometer (Imaging Research, Inc.,Ontario, Canada). All densitometric values were normalized to 7SrRNA obtained from the same analysis.
Determination of DNA single-strand breaks by alkaline filter elu-tion. The induction of DNA SSB in LLC-PK1 cells was measuredwith an automated alkaline filter elution system (24) based on themethod of Kohn and co-workers (25). After exposure, cells werewashed once with 5 ml cold EBSS, and 2 ml EBSS was added to thecell culture flasks. Approximately 1–21 106 cells were removed fromthe flasks by scraping with a rubber policeman, and cell sampleswere loaded onto 25-mm polycarbonate filters (2 mm; Nucleopore,Cambridge, UK). After lysing and deproteinization, DNA was eluted(0.03 ml/min) with 20 mM Na2EDTA at pH 12.50 { 0.04. Two-hourfractions were collected for a total of 16 h, and DNA was determinedfluorimetrically with the Hoechst 33258 dye.
RESULTS
2-Br-bis-(GSyl)HQ-mediated DNA single-strandbreaks, gadd153 and histone H2B expression, and cyto-toxicity. 2-Br-bis-(GSyl)HQ causes the concentration-and time-dependent formation of single-strand breaksin DNA in plateau-phase cultures of renal tubular epi- FIG. 2. Time- and concentration-dependent effect of 2-Br-bis-(glu-
tathion-S-yl)hydroquinone on gadd153 mRNA expression in renalthelial cells (Fig. 1). DNA SSB were detected at a con-tubular epithelial cells. LLC-PK1 cells were exposed to either (A)centration of 100 mM 2-Br-bis-(GSyl)HQ and were max-200 mM 2-Br-bis-(GSyl)HQ for between 0 and 5 h or (B) varyingimal at 400 mM. At 400 mM 2-Br-bis-(GSyl)HQ, DNA concentrations (0–600 mM) of 2-Br-bis-(GSyl)HQ for 2 h. Total RNA
SSB were detected as early as 15 min after exposure, was extracted, separated, and examined by Northern blot analysis.The level of 7S rRNA was used as the internal control.were maximal at 60 min, and decreased significantly
AID ABB 9969 / 6b33$$$422 04-21-97 10:35:13 arcas
303QUINOL THIOETHER-MEDIATED DNA DAMAGE AND CYTOTOXICITY
findings, neither gadd153 mRNA levels nor cytotoxicity(data not shown) were influenced by the addition ofAra-C and hydroxyurea.
A role for hydrogen peroxide and hydroxyl radical in2-Br-bis-(GSyl)HQ-mediated DNA SSB, gadd-153 ex-pression, and cytotoxicity. To determine whetherH2O2 and the hydroxyl radical are involved in the for-mation of DNA SSB and the induction of gadd153 inplateau-phase LLC-PK1 cells, we investigated the ef-fects of transition metal ion chelators (the copper ionchelator, bathocuproine; the iron chelator, deferoxa-mine; and the divalent metal ion chelator, 1,10-phenan-throline) and catalase (10 U/ml) on DNA SSB andgadd153 expression. Pretreatment of LLC-PK1 cellswith deferoxamine (Fig. 6) or cotreatment with catalase(Fig. 7) decreased 2-Br-bis-(GSyl)HQ-mediated DNASSB and gadd153 induction. Both catalase and defer-oxamine (10 mM) protected LLC-PK1 cells from 2-Br-
FIG. 3. Concentration- and time-dependent effect of 2-Br-bis-(glu- bis-(GSyl)HQ-mediated cytotoxicity (Figs. 6 and 7). Intathion-S-yl)hydroquinone on histone H2B mRNA expression in re- contrast to deferoxamine, neither bathocuproine (10nal tubular epithelial cells. LLC-PK1 cells were exposed to either (A) mM) nor 1,10-phenanthroline (0.2 mM) protected cellsvarying concentrations of 2-Br-bis-(GSyl)HQ (0–500 mM) for 2 h or
against 2-Br-bis-(GSyl)HQ-mediated DNA SSB (Fig.(B) 200 mM 2-Br-bis-(GSyl)HQ for between 0 and 5 h. Total RNA was6A) or cytotoxicity (Fig. 6B). However, 1,10-phenan-extracted, separated, and examined by Northern blot analysis. The
level of 7S rRNA was used as the internal control. throline did attenuate increases in gadd153 mRNA ex-pression (Fig. 6B).DISCUSSION
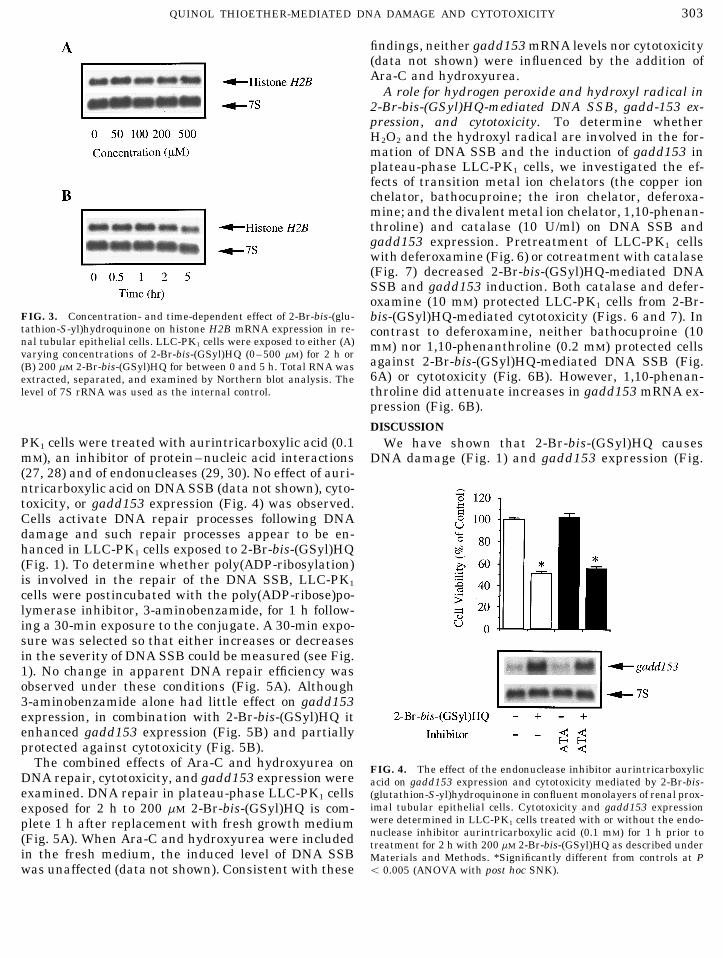
PK1 cells were treated with aurintricarboxylic acid (0.1 We have shown that 2-Br-bis-(GSyl)HQ causesmM), an inhibitor of protein–nucleic acid interactions DNA damage (Fig. 1) and gadd153 expression (Fig.(27, 28) and of endonucleases (29, 30). No effect of auri-ntricarboxylic acid on DNA SSB (data not shown), cyto-toxicity, or gadd153 expression (Fig. 4) was observed.Cells activate DNA repair processes following DNAdamage and such repair processes appear to be en-hanced in LLC-PK1 cells exposed to 2-Br-bis-(GSyl)HQ(Fig. 1). To determine whether poly(ADP-ribosylation)is involved in the repair of the DNA SSB, LLC-PK1
cells were postincubated with the poly(ADP-ribose)po-lymerase inhibitor, 3-aminobenzamide, for 1 h follow-ing a 30-min exposure to the conjugate. A 30-min expo-sure was selected so that either increases or decreasesin the severity of DNA SSB could be measured (see Fig.1). No change in apparent DNA repair efficiency wasobserved under these conditions (Fig. 5A). Although3-aminobenzamide alone had little effect on gadd153expression, in combination with 2-Br-bis-(GSyl)HQ itenhanced gadd153 expression (Fig. 5B) and partiallyprotected against cytotoxicity (Fig. 5B).
The combined effects of Ara-C and hydroxyurea on FIG. 4. The effect of the endonuclease inhibitor aurintricarboxylicDNA repair, cytotoxicity, and gadd153 expression were acid on gadd153 expression and cytotoxicity mediated by 2-Br-bis-examined. DNA repair in plateau-phase LLC-PK1 cells (glutathion-S-yl)hydroquinone in confluent monolayers of renal prox-
imal tubular epithelial cells. Cytotoxicity and gadd153 expressionexposed for 2 h to 200 mM 2-Br-bis-(GSyl)HQ is com-were determined in LLC-PK1 cells treated with or without the endo-plete 1 h after replacement with fresh growth mediumnuclease inhibitor aurintricarboxylic acid (0.1 mM) for 1 h prior to(Fig. 5A). When Ara-C and hydroxyurea were included treatment for 2 h with 200 mM 2-Br-bis-(GSyl)HQ as described under
in the fresh medium, the induced level of DNA SSB Materials and Methods. *Significantly different from controls at Põ 0.005 (ANOVA with post hoc SNK).was unaffected (data not shown). Consistent with these
AID ABB 9969 / 6b33$$$422 04-21-97 10:35:13 arcas

304 JEONG ET AL.
FIG. 5. The effect of 3-aminobenzamide on DNA single-strand breaks, gadd153 expression, and cytotoxicity mediated by 2-Br-bis-(glutath-ion-S-yl)hydroquinone in confluent monolayers of renal proximal tubular epithelial cells. (A) DNA single-strand breaks were measured inLLC-PK1 cells treated with [h] or without [¬] 3-aminobenzamide (10 mM) for 1 h after incubation with 200 mM 2-Br-bis-(GSyl)HQ (s) for30 min; [l] untreated controls. (B) Cytotoxicity and gadd153 expression were determined as described under Materials and Methods.*Significantly different from controls at P õ 0.005. †Significantly different from 2-Br-bis-(GSyl)HQ-treated cells at P õ 0.005 (ANOVA withpost hoc SNK).
2) in confluent cultures of LLC-PK1 cells, indicating of LLC-PK1 cells (Fig. 3), perhaps in support of theapparent rapid DNA repair. Thus, not all features ofthat DNA damage is not dependent on DNA replica-
tion and cell division. 2-Br-bis-(GSyl)HQ also causes the molecular stress response to 2-Br-bis-(GSyl)HQ-mediated DNA damage in log-phase LLC-PK1 cellsDNA fragmentation and induces gadd153 mRNA in
preconfluent cultures of renal proximal tubular epi- are reproduced in plateau-phase cells. The extent towhich signals generated in response to the inhibitionthelial cells (15, 16). The gadd153 mRNA is always
rapidly and highly induced by DNA damaging agents, of DNA synthesis in a S-phase cell are shared by thenormal S–G2 transition is not known (33).whereas the response to growth arrest appears
slower and less intense (31). Thus, the intensity of Within the context of demonstrating that the growthstatus influences the genomic response of LLC-PK1the genomic (gadd153) stress response to DNA dam-
age in log-phase and plateau-phase LLC-PK1 cells is cells to 2-Br-bis-(GSyl)HQ, we have also shown for thefirst time that (i) 2-Br-bis-(GSyl)HQ-mediated DNAsimilar. Consistent with the gadd153 response, the
induction of hsp70 mRNA by 2-Br-bis-(GSyl)HQ in damage in LLC-PK1 cells is mediated by the generationof H2O2 (Figs. 6 and 7); (ii) DNA damage, either directlylog-phase and plateau-phase LLC-PK1 cells is equiv-
alent (data not shown). or indirectly, contributes to cell death (Figs. 6 and 7);and (iii) DNA damage, either directly or indirectly, pro-Enhancement of DNA repair processes occurs in
LLC-PK1 cells after exposure to 2-Br-bis-(GSyl)HQ vides the initial signal for gadd153 expression (Figs. 6and 7). Thus, the formation of DNA SSB in LLC-PK1(Figs. 1 and 5). Cells respond to DNA damage by
initiating the transcription of a battery of genes in- cells treated with 2-Br-bis-(GSyl)HQ is completely pre-vented by catalase (Fig. 7A) with corresponding in-volved in the DNA repair process (31). The initial
response includes the arrest of cell cycle progression, creases in cell viability (Fig. 7B), indicating that H2O2
is necessary for both processes and that DNA damagepresumably to facilitate DNA repair prior to DNAreplication (G1 arrest) and cell division (G2 arrest). and cytotoxicity are associated. The relationship be-
tween H2O2, DNA damage, and cytotoxicity may beConsistent with this view, scheduled DNA synthesis(16) and histone mRNA levels (32) decline rapidly both cell-type and cell-cycle dependent. For example,
DNA damage induced following exposure of log-phasein preconfluent LLC-PK1 cells treated with 2-Br-bis-(GSyl)HQ. In contrast, histone H2B mRNA is unaf- CHO cells to menadione did not correlate with the inhi-
bition of cell growth (17), whereas in exponentiallyfected by 2-Br-bis-(GSyl)HQ in confluent monolayers
AID ABB 9969 / 6b33$$$423 04-21-97 10:35:13 arcas

305QUINOL THIOETHER-MEDIATED DNA DAMAGE AND CYTOTOXICITY
FIG. 6. The effect of transition metal ion chelators on DNA single-strand breaks, gadd153 expression, and cytotoxicity caused by 2-Br-bis-(glutathion-S-yl)hydroquinone in confluent monolayers of renal proximal tubular epithelial cells. (A) DNA single strand breaks weremeasured in LLC-PK1 cells treated with [¬] or without [s] bathocuproine (10 mM), deferoxamine (10 mM), or 1,10-phenanthroline (0.2 mM)for 1 h prior to exposure for 2 h to 200 mM 2-Br-bis-(GSyl)HQ; [l] untreated controls. (B) Cytotoxicity and gadd153 expression weredetermined as described under Materials and Methods. *Significantly different from controls at P õ 0.005. †Significantly different from 2-Br-bis-(GSyl)HQ-treated cells at P õ 0.005 (ANOVA with post hoc SNK).
growing MCF-7 human breast cancer epithelial cells, blastic cells (36). The gadd153 gene may also be subjectto redox modulation (37) and is induced in HeLa cellsmenadione-induced DNA damage correlated well with
cytotoxicity and growth inhibition (18, 34). exposed to H2O2 (38, 39). In contrast, low concentra-tions of H2O2 serve as a competence-like factor in quies-DNA damage in LLC-PK1 cells is accompanied by an
increase in the expression of gadd153 (Fig. 2). Control cent Balb/3T3 cells (40), and 50 mM H2O2 stimulatedgrowth in MCF-10A cells (34). Thus, the response tolevels of gadd153 mRNA are low in postconfluent LLC-
PK1 cells, and the generation of H2O2 by 2-Br-bis- H2O2 appears to be both concentration and cell-typedependent. In LLC-PK1 cells exposed to 2-Br-bis-(GSyl)HQ may result in damage to DNA or other cellu-
lar molecules, providing a signal for gadd153 expres- (GSyl)HQ, catalase prevents the induction of gadd153(Fig. 7B), indicating that H2O2 causes growth inhibi-sion. H2O2 has been shown to induce a senescence-like
growth arrest in human diploid fibroblasts (35) and to tory damage in renal proximal tubular epithelial cells.Because H2O2 is a precursor to the highly destructiveact as a negative regulator of growth in mouse osteo-
AID ABB 9969 / 6b33$$$423 04-21-97 10:35:13 arcas

306 JEONG ET AL.
FIG. 7. Effect of catalase on DNA single-strand breaks, gadd153 expression, and cytotoxicity caused by 2-Br-bis-(glutathion-S-yl)hydroqui-none in confluent monolayers of renal proximal tubular epithelial cells. (A) DNA single-strand breaks were measured in LLC-PK1 cellstreated with [¬] or without [s] catalase (10 units/ml) and 200 mM 2-Br-bis-(GSyl)HQ; [l] untreated controls. (B) Cytotoxicity and gadd153expression were determined as described under Materials and Methods. *Significantly different from controls at P õ 0.005 (ANOVA withpost hoc SNK).
hydroxyl radical, and because this reaction requires cells exposed to 2-Br-bis-(GSyl)HQ, the H2O2 is gener-ated intracellularly (probably at localized sites), andthe availability of transition metal ions, we investi-
gated the role of transition metal ions in 2-Br-bis- some fraction of it will diffuse to the extracellular com-partment dependent upon the efficiency of its degrada-(GSyl)HQ-mediated DNA SSB formation. Neither the
copper ion chelator bathocuproine nor the divalent tion by intracellular catalase and GSH peroxidase.DNA SSB initiate the activation of poly(ADP-ribo-metal ion chelator 1,10-phenanthroline prevents 2-Br-
bis-(GSyl)HQ-mediated DNA SSB in LLC-PK1 cells se)polymerase (44, 45), an enzyme involved in DNArepair. However, DNA repair is unchanged in cells(Fig. 6A). In contrast, the iron chelator deferoxamine
prevents both 2-Br-bis-(GSyl)HQ-mediated DNA SSB treated with the poly(ADP-ribose)polymerase inhibitor3-aminobenzamide, following exposure to 2-Br-bis-and gadd153 expression (Fig. 6). Attenuation of
gadd153 expression is therefore not dependent upon (GSyl)HQ (Fig. 5A). The efficiency of DNA repair inLLC-PK1 cells may well mask the inhibitory effects ofthe removal of H2O2 per se, but on preventing the gen-
eration of hydroxyl radical. Thus, although ROS are 3-aminobenzamide, which, although able to delay therepair of H2O2-induced DNA SSB, does not prevent re-implicated as intracellular messengers in a variety of
systems (41), H2O2 is unlikely to directly serve as a pair (46). In contrast, decreases in cell viability causedby 2-Br-bis-(GSyl)HQ are partially reversed by 3-amino-messenger for the induction of gadd153. Indeed, oxida-
tive stress and DNA damage regulate gadd153 tran- benzamide (Fig. 5B). Inhibition of poly(ADP-ribose)-polymerase also blocks cell death by other DNA-dam-scription by different pathways (42).
When LLC-PK1 cell suspensions are bathed in a solu- aging agents, such as UV light and N-methyl-N *-nitro-nitrosoguanidine (44) and 1,2-dibromo-3-chloropropanetion of 1 mM H2O2, the subsequent DNA fragmentation
is inhibited by aurintricarboxylic acid (43), suggesting (47). The activation of poly(ADP-ribose)polymerase canbe toxic to cells due to severe depletion of NAD/ andthat endonucleases are activated in this system. How-
ever, aurintricarboxylic acid has no effect on 2-Br-bis- ATP for poly(ADP-ribosylation) following DNA damage(44, 48). Thus, 3-aminobenzamide probably protects(GSyl)HQ-mediated DNA SSB (data not shown),
gadd153 expression, or cytotoxicity (Fig. 4). Thus, the against 2-Br-bis-(GSyl)HQ-mediated cytotoxicity by at-tenuating depletions in NAD/ and ATP.cellular and molecular responses to ROS also appear
contingent upon their site of generation. In LLC-PK1 Ara-C plus hydroxyurea efficiently block nucleotide
AID ABB 9969 / 6b33$$$423 04-21-97 10:35:13 arcas

307QUINOL THIOETHER-MEDIATED DNA DAMAGE AND CYTOTOXICITY
of Foreign Compounds (Anders, M. W., and Dekant, W., Eds.),excision repair and ligation (49). Increases in the accu-pp. 183–210, Academic Press, New York.mulation of DNA strand breaks are observed in UV-
14. Monks, T. J., and Lau, S. S. (1994) in Handbook of Experimentalirradiated HeLa (50) and L1210 cells (51) and theyPharmacology, Vol. 112, Conjugation–Deconjugation Reactionsaccumulate in all phases of the cell cycle (mitosis, G1, in Drug Metabolism and Toxicity (Kauffman, F. C., Ed.), pp.
and S phase) (50). Ara-C and hydroxyurea, however, 459–509, Springer-Verlag, Berlin.do not increase DNA SSB after exposure to X-ray- 15. Mertens, J. J. W. M., Gibson, N. W., Lau, S. S., and Monks, T. J.treated (52) or 2-Br-bis-(GSyl)HQ-treated LLC-PK1 (1995) Arch. Biochem. Biophys. 320, 51–58.cells (data not shown). Thus, the lack of effect of Ara- 16. Jeong, J. K., Stevens, J. L., Lau, S. S., and Monks, T. J. (1996)
Mol. Pharmacol. 50, 592–598.C plus hydroxyurea, combined with the rapid removal17. Cantoni, O., Fiorani, M., Cattabeni, F., and Bellomo, G. (1991)of transient DNA SSB is consistent with the view that
Biochem. Pharmacol. 42, S220–S222.the DNA damage induced by 2-Br-bis-(GSyl)HQ is due18. Nutter, L. M., Ngo, E. O., Fisher, G. R., and Gutierrez, P. L.to the formation of the hydroxyl radical, resembling
(1992) J. Biol. Chem. 267, 2474–2479.that observed after X-ray treatment. Interestingly,19. Lau, S. S., Jones, T. W., Sioco, R., Hill, B. A., Pinon, R. K., andtreatment of LLC-PK1 cells with Ara-C and hydroxy-
Monks, T. J. (1990) Toxicology 64, 291–31.urea alone did not induce gadd153 expression (data not20. Borenfreund, E., and Puerner, J. A. (1984) J. Tissue Cultureshown), consistent with data showing that increases in
Methods 9, 7–9.gadd153 mRNA result from the formation of DNA SSB21. Babich, H., and Borenfreund, E. (1990) ATLA 18, 129–144.and/or subsequent growth arrest, rather than from a22. Chomczynski, P., and Sacchi, N. (1987) Anal. Biochem. 162, 156–direct inhibition of replicative DNA synthesis (53).
159.In summary, the stress response to 2-Br-bis-
23. Feinberg, A. P., and Vogelstein, B. (1985) Anal. Biochem. 137,(GSyl)HQ in confluent monolayer cultures of renal 266–267.proximal tubular epithelial cells differs from that seen 24. Kohn, K. W., Ewig, R. A. G., Erickson, L. C., and Zwelling, L. A.in log-phase cells. In addition, the formation of DNA (1981) in DNA Repair: A Laboratory Manual of Research Proce-SSB, gadd153 induction, and cytotoxicity are all modu- dures (Friedberg, E. C., and Hanawalt, P. C., Eds.), Vol. 1, Part
B, pp. 379–401, Dekker, New York.lated by agents which prevent the formation of the hy-25. Brunborg, G., Holme, J. A., Soderlund, E. J., Omichinski, J. G.,droxyl radical. The nature of the cellular and molecular
and Dybing, E. (1988) Anal. Biochem. 174, 522–536.responses to H2O2 or H2O2-generating quinones ap-26. Hocke, G. M., Barry, D., and Fey, G. H. (1992) Mol. Cell. Biol.pears to be cell-type and cell-cycle dependent. The site
12, 2282–2292.at which the cell is exposed to H2O2 may also determine27. Gonzalez, R. G., Haxo, R. S., and Schleich, T. (1980) Biochemis-the nature of the stress response.
try 19, 4299–4303.
28. Blumenthal, T., and Landers, T. A. (1973) Biochem. Biophys.REFERENCES Res. Commun. 55, 680–688.
29. Hallick, R. B., Chelm, B. K., Gray, P. W., and Orozco, E. M., Jr.1. Halliwell, B., and Cross, C. E. (1994) Environ. Health Perspect.(1977) Nucleic Acids Res. 4, 3055–3064.102(Suppl. 10), 5–12.
30. McConkey, D. J., Hartzell, P., Nicotera, P., and Orrenius, S.2. Wiseman, H., and Halliwell, B. (1996) Biochem. J. 313, 17–29.(1989) FASEB J. 3, 1843–1849.3. Leadon, S. A., Stampfer, M. R., and Bartley, J. (1988) Proc. Natl.
31. Fornace, A. J., Jr., Nebert, D. W., Hollander, M. C., Leuthy, J. D.,Acad. Sci. USA 85, 4365–4368.Papathanasiou, M., Fargnoli, J., and Holbrook, N. J. (1989) Mol.4. Monks, T. J., Hanzlik, R. P., Cohen, G. M., Ross, D., and Gra-Cell. Biol. 9, 4196–4203.ham, D. G. (1992) Toxicol. Appl. Pharmacol. 112, 2–16.
32. Jeong, J. K., Lau, S. S., and Monks, T. J. (1997) J. Biol. Chem.5. Kappus, H. (1986) Biochem. Pharmacol. 35, 1–6.272, 7511–7518.6. Smith, M. T., Evans, C. G., Thor, H., and Orrenius, S. (1985) in
33. Marzluff, W. F., and Hanson, R. J. (1993) in Control of Messen-Oxidative Stress (Sies, H., Ed.), pp. 91–113, Academic Press,ger RNA Stability (Belasco, J., and Brawerman, G., Eds.), pp.London.267–290, Academic Press, San Diego.7. Wilson, I., Wardman, P., Lin, T., and Sartorelli, A. C. (1986) J.
34. Djuric, Z., Everett, C. K., and Luongo, D. (1993) Free RadicalMed. Chem. 29, 1381–1384.Biol. Med. 14, 541–547.8. Brunmark, A., and Cadenas, E. (1988) Free Radical Biol. Med.
35. Chen, Q., and Ames, B. N. (1994) Proc. Natl. Acad. Sci. USA 91,7, 435–477.4130–4134.9. O’Brien, P. J. (1991) Chem. Biol. Interact. 80, 1–41.
36. Shibanamu, M., Arata, S., Murata, M., and Nose, K. (1995) Exp.10. Wefers, H., and Sies, H. (1983) Arch. Biochem. Biophys. 224,Cell Res. 218, 132–136.568–578.
37. Chen, Q., Yu, K., Holbrook, N. J., and Stevens, J. L. (1992) J.11. Brown, P. C., Dulik, D. M., and Jones, T. W. (1991) Arch. Bio-Biol. Chem. 267, 8207–8212.chem. Biophys. 285, 187–196.
38. Luethy, J. D., and Holbrook, N. J. (1992) Cancer Res. 52, 1–10.12. Monks, T. J., and Lau, S. S. (1992) CRC Crit. Rev. Toxicol. 22,39. Fornace, A. J., Jr. (1992) Annu. Rev. Genet. 26, 507–526.243–270.
13. Monks, T. J., and Lau, S. S. (1994) in Advances in Pharmacology, 40. Shibanamu, M., Kuroki, T., and Nose, K. (1990) Oncogene 5,1025–1032.Vol. 27, Conjugation-Dependent Carcinogenicity and Toxicology
AID ABB 9969 / 6b33$$$423 04-21-97 10:35:13 arcas

308 JEONG ET AL.
41. Powis, G., Briehl, M., and Oblong, J. (1995) Pharm. Ther. 68, 48. Gaal, J. C., Smith, K. R., and Pearson, C. K. (1987) Trends Bio-chem. Sci. 12, 129–130.149–173.
42. Luethy, J. D., and Holbrook, N. J. (1994) Cancer Res. 54(Suppl.), 49. Gedik, C. M., and Collins, A. R. (1991) Mutat. Res. 254, 231–237.1902s–1906s. 50. Downes, C. S., and Collins, A. R. (1982) Nucleic Acids Res. 10,
43. Ueda, N., and Shah, S. V. (1992) J. Clin. Invest. 90, 2593–2597. 5357–5368.44. Berger, N. A. (1985) Radiat. Res. 101, 4–15. 51. Kufe, D. W., Weichselbaum, R., Egan, E. M., Dahlberg, W., and
Fram, R. J. (1984) Mol. Pharmacol. 25, 322–326.45. Shall, S. (1984) Nucleic Acids Symp. Series. 13, 143–191.52. Brunborg, G., Holme, J. A., and Hongslo, J. K. (1995) Mutat. Res.46. Palomba, L., Guidarelli, A., Cattabeni, F., and Cantoni, O. (1995)
342, 157–170.Eur. J. Pharmacol. 291, 167–173.47. Holme, J. A., Søderlund, E. J., Brunborg, G., Lag, M., Nelson, 53. Leuthy, J. D., and Holbrook, N. J. (1988) Proc. Natl. Acad. Sci.
USA 85, 8800–8804.S. D., and Dybing, E. (1991) Cell Biol. Toxicol. 7, 413–432.
AID ABB 9969 / 6b33$$$424 04-21-97 10:35:13 arcas