developmental control of cav1.2 l-type calcium channel splicing by fox proteins
TRANSCRIPT

MOLECULAR AND CELLULAR BIOLOGY, Sept. 2009, p. 4757–4765 Vol. 29, No. 170270-7306/09/$08.00�0 doi:10.1128/MCB.00608-09Copyright © 2009, American Society for Microbiology. All Rights Reserved.
Developmental Control of CaV1.2 L-Type Calcium ChannelSplicing by Fox Proteins�†
Zhen Zhi Tang,1 Sika Zheng,1 Julia Nikolic,1 and Douglas L. Black1,2*Howard Hughes Medical Institute, University of California at Los Angeles,1 and Department of Microbiology,
Immunology, and Molecular Genetics, University of California at Los Angeles,2 Los Angeles, California 90095
Received 9 May 2009/Returned for modification 9 June 2009/Accepted 17 June 2009
CaV1.2 voltage-gated calcium channels play critical roles in the control of membrane excitability, geneexpression, and muscle contraction. These channels show diverse functional properties generated by alterna-tive splicing at multiple sites within the CaV1.2 pre-mRNA. The molecular mechanisms controlling thissplicing are not understood. We find that two exons in the CaV1.2 channel are controlled in part by membersof the Fox family of splicing regulators. Exons 9* and 33 confer distinct electrophysiological properties on thechannel and show opposite patterns of regulation during cortical development, with exon 9* progressivelydecreasing its inclusion in the CaV1.2 mRNA over time and exon 33 progressively increasing. Both exonscontain Fox protein binding elements within their adjacent introns, and Fox protein expression is induced incortical neurons in parallel with the changes in CaV1.2 splicing. We show that knocking down expression of Foxproteins in tissue culture cells has opposite effects on exons 9* and 33. The loss of Fox protein increases exon9* splicing and decreases exon 33, as predicted by the positions of the Fox binding elements and by the patternof splicing in development. Conversely, overexpression of Fox1 and Fox2 proteins represses exon 9* andenhances exon 33 splicing in the endogenous CaV1.2 mRNA. These effects of Fox proteins on exons 9* and 33can be recapitulated in transfected minigene reporters. Both the repressive and the enhancing effects of Foxproteins are dependent on the Fox binding elements within and adjacent to the target exons, indicating thatthe Fox proteins are directly regulating both exons. These results demonstrate that the Fox protein family isplaying a key role in tuning the properties of CaV1.2 calcium channels during neuronal development.
CaV1.2 L-type voltage-gated calcium channels are widelydistributed in brain, heart, smooth muscle, and endocrine cellsand play essential roles in gene expression, muscle contraction,and hormone release (6, 13, 16, 39, 47). These channels arecomposed of three subunits, with the �1 subunit being thelargest and incorporating the conduction pore, the voltagesensor and gating apparatus, as well as sites for channel regu-lation by second messengers, drugs, and toxins (Fig. 1A) (9, 14,17). This CaV1.2 subunit is subject to extensive alternativesplicing that generates multiple functionally distinct isoforms(1, 29, 33, 49, 62). At least twenty of the 56 exons in the humanCaV1.2 transcript are alternatively spliced (29, 50, 51, 55). Inparticular, alternative exon 9* within the cytoplasmic I-II loopand exon 33 within the IVS3-IVS4 transmembrane segmentsconfer different electrophysiological and pharmacologicalproperties on the channel and exhibit tissue-specific differ-ences in inclusion (30, 31, 54, 55). Changes in exon 9* (alsonamed exon 9A) splicing are seen in human arterial smoothmuscle cells that have developed atherosclerosis (57) and inhypertrophied cardiomyocytes of spontaneously hypertensiverats (56). Alternative exons 9* and 33 are conserved acrossvertebrate species demonstrating their functional importance
to the CaV1.2 channel. However, the molecular mechanismscontrolling their splicing have not been studied.
Members of the Fox protein family, homologs of the Femi-nizing on the X gene product from Caenorhabditis elegans (21,41, 48), regulate the splicing of many neuron- and muscle-specific splicing events (22, 40, 59, 63–66). There are threemammalian family members, Fox1 (A2BP1), Fox2 (RBM9),and Fox3 (hnrbp3), each containing a nearly identical RNA-binding domain that recognizes the hexanucleotide elementUGCAUG (2). These proteins bind the introns adjacent totheir target exons where they generally repress splicing whenbound upstream of the exon but enhance splicing from a down-stream binding site (22, 40, 59, 63, 64, 66). In addition to theRNA-binding domain, all three proteins have similar N andC-terminal domains that are extensively modified by alterna-tive promoter use and alternative splicing to produce a largefamily of related proteins. Fox1 is expressed in neurons andmuscle, and Fox3 is expressed only in neurons (22, 23, 36, 59).Fox2 shows somewhat broader expression, being found in em-bryonic stem cells and in the embryo in addition to neuronsand muscle (3, 44, 63).
We identified multiple (U)GCAUG elements in the se-quence surrounding CaV1.2 exons 9* and 33 of human, rat,mouse, chicken, and other species. We show here that themammalian Fox proteins have opposite effects on the alterna-tive splicing of exons 9* and 33. In developing mouse cortex,these exons change their pattern of splicing in parallel withincreases in Fox protein expression. This establishes roles forthe Fox proteins in tuning CaV1.2 channel physiology duringdevelopment.
* Corresponding author. Mailing address: Howard Hughes MedicalInstitute, University of California, Los Angeles, Los Angeles, CA90095-1662. Phone: (310) 794-7644. Fax: (310) 206-8623. E-mail:[email protected].
† Supplemental material for this article may be found at http://mcb.asm.org/.
� Published ahead of print on 29 June 2009.
4757
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 04
Feb
ruar
y 20
22 b
y 12
4.25
.181
.138
.

MATERIALS AND METHODS
Cell culture, treatments, and transfections. N2a and HEK293T cells weregrown according to the American Type Culture Collection-recommended pro-tocols. Myoblast C2C12 cells were grown in high-glucose Dulbecco modifiedEagle medium (Invitrogen) with 1.5 g of sodium bicarbonate/liter and 1 mMsodium pyruvate, supplemented with 10% fetal bovine serum. Transfectionswere done with Lipofectamine 2000 (Invitrogen) according to the manufacturer’sinstructions.
Plasmids. The Dup-rat E9* and Dup-rat E33 reporter minigenes were con-structed by amplifying rat genomic DNA containing exons 9* and 33 with flank-ing intron sequences from rat genomic DNA using Pfu DNA polymerase andprimers designed from the GenBank genomic sequence NW_047696. The PCRfragments were cloned into the template plasmid pDUP4-1 (37, 38) between theApaI and BglII sites that are engineered into 5� and 3� ends of the amplicons.The primers are presented in Table S1 in the supplemental material (primers 1to 4). Mutations in Fox binding elements were generated by using the
QuikChange site-directed mutagenesis kit (Stratagene) and followed by subclon-ing into pDUP4-1 using the ApaI and BglII sites. All constructs were confirmedby sequencing. The Fox1 and Fox2 cDNA expression plasmids were constructedin pcDNA3.1 (Invitrogen) as described previously (59). Each carried an N-terminal FLAG tag.
shRNAs for knockdown of Fox proteins. Short hairpin RNAs (shRNAs)pBlsH1Fox-1 and pBlsH1Fox-2 were designed to target the 3� untranslatedregion of mouse Fox1 or Fox2 genes. The template vector was pBlsH1 and thedesign of the shRNA was similar to what was described previously (12). Theoligonucleotides used were primers 9 to 12 in Table S1 in the supplementalmaterial. The shRNA expression plasmids pBlsH1Fox-1 and pBlsH1Fox-2 weretransfected into C2C12 and N2A cells grown in six-well plates using Lipo-fectamine 2000 (Invitrogen). The efficiency of knockdown was verified by West-ern blot.
Reverse transcription-PCR (RT-PCR) and primer extension. CytoplasmicRNA was isolated from cells or mouse embryonic cortexes with an RNeasyminikit (Qiagen) and reverse transcribed with random hexamers and SuperscriptIII (Invitrogen) according to the manufacturer’s instruction. Splice productswere PCR amplified (25 cycles for N2A cells and 29 cycles for C2C12 cells) usinga 32P-labeled primer in the 3� constitutive exon and an unlabeled primer in the5� constitutive exon. The primers (primers 5 to 8) are listed in Table S1 in thesupplemental material. Primer extensions were performed as described previ-ously (28, 38), using a 32P-labeled DUP3 reverse primer (5�-AACAGCATCAGGAGTGGACAGATCCC-3�). The annealing temperature was 55°C. The primergives rise to a 230-bp primer extension product when the middle exon 9* or exon33 from the minigene is not included. The length of the exon 9*-included productis 305 bp and exon 33-included product is 263 bp. 32P-labeled PCR and primerextension products were separated by 8% polyacrylamide–7.5 M urea denaturinggels. Gels were dried, visualized by using a Typhoon phosphorimager (Amer-sham Biosciences), and quantified by using ImageQuant TL software (Amer-sham Biosciences).
Antibodies and Western blotting. Anti-Fox2 antibody was obtained from Be-thel Laboratories. Pan-anti-Fox RRM antibody was raised against an 86-amino-acid peptide sequence containing the Fox1 RRM and 3 amino acids upstreamand 12 amino acids downstream. The peptide sequence is PKRLHVSNIPFRFRDPDLRQMFGQFGKILDVEIIFNERGSKGFGFVTFENSADADRAREKLHGTVVEGRKIEVNNATARVMTNKKM. This sequence is identical in thehuman and mouse Fox1 and Fox2 proteins except for the last amino acid. It hasfive amino acid changes in Fox3. The antibody was affinity purified on theantigen. Its specificity was confirmed by probing blots of recombinant Fox1 and2 proteins and native Fox proteins in cell lysates before and after Fox proteinknockdown.
The mouse monoclonal anti-Fox1 antibody was raised against bacterially ex-pressed recombinant mouse Fox1 fused with glutathione S-transferase. Immu-nization in BALB/c mice was done according to standard protocols (27). My-eloma and spleen cell fusion was carried out by Susan Oh at the CaltechMonoclonal Antibody Facility using HL-1 myeloma cells. Hybridomas wereinitially screened by enzyme-linked immunosorbent assay using the original an-tigen. Positive hybridomas supernatants were retested on Western blots of totalprotein from HEK293T cells transfected with Flag-tagged mouse Fox1. One ofthe positive clones, 1D10, was subsequently cloned by limiting dilution. Theanti-Fox3 rabbit polyclonal antibody was raised against the Fox3 peptide CQTPVPPEHGMTLYT at Pacific Immunology and purified from rabbit serum onpeptide coupled to Sulfo-Link resin (Pierce).
Tissue culture cell lysates for Western blot were isolated in radioimmunopre-cipitation assay buffer (150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate,0.1% sodium dodecyl sulfate, and 50 mM Tris at pH 8.0) as previously described(11). Whole-cell lysates were resolved on 4 to 20% Tris-glycine gels (Invitrogen)in Tris-glycine-SDS running buffer. The proteins were transferred onto Protrannitrocellulose membranes (Whatman, Germany) or Immobilon-FL polyvinyli-dene difluoride membranes (Millipore). Membranes were probed under stan-dard conditions with anti-Fox RNA recognition motif (RRM; 1:10,000), anti-Fox1 (1:5,000), anti-Fox2 (1:1,000; Bethyl Laboratories, Inc.), anti-Fox3(1:3,000), anti-Flag (1:3,000; Sigma), and anti-glyceraldehyde phosphate dehy-drogenase (anti-GAPDH; 1:200,000; Research Diagnostics, Inc.) antibodies.Blots were then probed with horseradish peroxidase-conjugated secondary anti-bodies (1:10,000; GE Healthcare) and developed by using Femto Supersignalreagents (Pierce Biotechnology) and Kodak BioMax XAR film. For blotting withfluorophore-conjugated secondary antibodies, the polyvinylidene difluoridemembranes were probed with ECL Plex Cy3-conjugated goat anti-mouse (1:5,000; GE Healthcare) and Cy5-conjugated goat anti-rabbit (1:2,500; GE Health-care) secondary antibodies, dried, and scanned on a Typhoon PhosphorImager
FIG. 1. (A) Diagram of the CaV1.2 subunit. The protein is com-posed of four membrane-spanning domains (I to IV), with each do-main consisting of six transmembrane segments (S1 to S6). Proteinsegments encoded by alternatively spliced exons are indicated by blackor hatched boxes. Exons 1a, 1b, and 1c derive from three alternativepromoters (15, 42, 45); exons 8a/8, 21/22, and 31/32 are spliced in amutually exclusive manner (51, 61, 62, 67); and exons 9*, 10*, 33, 45*,and 45 are alternative cassette exons (5, 18, 19, 24, 25). (B and C)RT-PCR assay of changes of exons 9* and 33 in embryonic mousecortex from embryonic day 12 (E12) to day 18 (E18). Exon 9* gradu-ally decreases in inclusion (B) and exon 33 gradually increases indeveloping mouse cortex (C). The PCR amplified region for exon 9*encompasses exons 9 through 10. The exon 9* included band is 354 bpand the exon 9* skipped band is 279 bp. The PCR-amplified region forexon 33 encompasses exons 30, 31, 32, 33, and 34. The exon 33 in-cluded and excluded fragments are 260 and 227 bp. The values forpercent exon 9* or exon 33 inclusion are the upper band intensitydivided by the summed intensities of upper and lower bands. Approx-imately 7 to 10 embryonic cortices were pooled for RNA extraction ineach group.
4758 TANG ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 04
Feb
ruar
y 20
22 b
y 12
4.25
.181
.138
.

(GE Healthcare). Quantification of fluorescent signals was performed by usingImageQuant TL software (Amersham Biosciences).
Immunostaining. Brain tissues from 18-day-old mouse embryos (CD-1 mice;Charles River Laboratories, Inc.) were cryosectioned at 10 �m in thickness.Sections attached to glass slides were fixed with 4% paraformaldehyde (pH 7) for20 min, washed three times with phosphate-buffered saline (PBS) with 0.1%Triton (PBST), and permeabilized with 0.5% Triton in PBS for 10 min at roomtemperature. Sections were then blocked in PBST, 2% bovine serum albumin(Sigma-Aldrich), and 3% normal goat serum for 1 h. The samples were incu-bated with primary antibodies overnight at 4°C. Primary antibodies were used atthe following concentrations: anti-Fox RRM (1:500), anti-Fox1 (1:1,000), anti-Fox2 (1:200; Bethyl Laboratories, Inc.), anti-NeuN (1:100; Chemicon), anti-MAP2 (1:200; Chemicon), and anti-GFAP (1:1,000; Chemicon). After beingwashed three times with PBST, sections were incubated with Alexa 568 (red)-conjugated anti-rabbit or anti-mouse immunoglobulin G, and Alexa 488 (green)-conjugated anti-mouse or anti-rabbit immunoglobulin G (1:1,000; MolecularProbes) secondary antibodies at room temperature for 1 h. Sections were rinsedthree times with PBST and mounted with ProLong Gold antifade reagent withDAPI (4�,6�-diamidino-2-phenylindole; Invitrogen). Control staining performedin parallel without adding primary antibodies yielded no labeling.
RESULTS
CaV1.2 exons 9* and 33 show divergent regulation in devel-oping cortex coinciding with the expression of Fox proteins.Exon 9* of CaV1.2 modifies the cytoplasmic I-II loop of theprotein, where it alters the voltage-dependent gating of thechannel (Fig. 1A) (31, 54). We find that in mouse embryoniccortex (E12) this exon is included in 25% of the CaV1.2mRNA but is progressively repressed over development,reaching only 4% inclusion by embryonic day 18 (Fig. 1B). Thisbehavior is in contrast to CaV1.2 exon 33. Exon 33 modifies thelinker separating the domain IV transmembrane segments S3and S4 and alters both the voltage dependence of the channeland its sensitivity to dihydropyridine drugs (30, 55). At embry-onic day 12, exon 33 is 63% included in the cortical CaV1.2mRNA. Unlike exon 9*, this splicing progressively increases tonear 100% inclusion over the next 6 days of development(Fig. 1C).
We have found that many proteins involved in calcium sig-naling and homeostasis exhibit alternative splicing that is reg-ulated by members of the Fox protein family (our unpublisheddata). In mammals, there are three members of this group ofRNA-binding proteins Fox1 (A2BP1), Fox2 (RBM9), and Fox3(hnrbp3). Examining CaV1.2 exon 9* in the rat and mouse, wefound potential Fox binding sites (UGCAUG) in three posi-tions, upstream and downstream of the exon and within theexon itself (Fig. 2A). For exon 33, we found one Fox recogni-tion site 111 nucleotides downstream of the 5� splice site. In thehuman CaV1.2 genes, these elements are also present althoughwith some variation (see Fig. S1 and S2 in the supplementalmaterial). Thus, both exons 9* and 33 are candidates for Foxprotein regulation.
It was previously shown that in the adult nervous system, theFox proteins are specifically expressed in postmitotic neurons(36, 59). We next examined the expression of the three Foxproteins during the developmental period that exons 9* and 33change in splicing. Using antibodies to each individual Foxprotein, as well as an antibody that reacts with all three pro-teins, we probed immunoblots of the proteins from the embry-onic cortex (Fig. 2C). We found that each protein is at bestweakly expressed at E12 but is induced over the next 6 days,showing strong expression by E18. Thus, the induction of Fox
protein expression during cortical development parallels thechanges in exon 9* and exon 33 splicing.
To identify cells in the developing brain that express the Foxproteins, sections of mouse cortex from embryonic day 18 wereimmunostained with �-Fox RRM, �-Fox1, and �-Fox2 anti-bodies and costained for the neuronal markers MAP2 orNeuN. We found that Fox proteins are strongly expressed inthe postmitotic neurons of the developing cortical plate but areonly weakly expressed in the proliferating cells of the interme-diate zone and subventricular zone (Fig. 2D, E, and F). Theexpression in the undifferentiated cells is primarily Fox2, afinding in agreement with its earlier induction during develop-ment and with its known expression in neuronal progenitorcells (63). As seen at higher magnification of the postmitoticcells in the outer cortical layers, nearly all cells are expressingboth the Fox proteins and the terminal differentiation markersNeuN and MAP2. However, there is some heterogeneity in theamount of staining and cells can be identified expressing pri-marily one protein or another. This heterogeneity is resolvedduring postnatal development since nearly all mature neuronsin the cortex express both Fox1 and Fox2 proteins (59; data notshown).
Fox proteins repress CaV1.2 exon 9* and activate exon 33. Ifthe Fox proteins expressed during cortical development areinducing changes in CaV1.2 exon splicing, it is predicted thatthey would act to repress exon 9* and to enhance exon 33. Totest whether these exons are indeed affected by Fox proteins,we performed RNA interference knockdown and protein over-expression experiments and measured the response of CaV1.2exon splicing. The N2a (Neuro-2a) mouse neuroblastoma cellline expresses CaV1.2 transcripts and Fox2 protein, but verylittle Fox1 or Fox3. This line is amenable to relatively efficientRNA interference knockdown and exhibits a neuronal pheno-type for the splicing of many gene transcripts. Transfectingthese cells with an shRNA expression construct targeting Fox1had no effect on the total Fox protein levels as measured withthe Fox RRM antibody (Fig. 3A, lanes 1 and 2). Expression ofan shRNA targeting Fox2 nearly eliminates Fox protein ex-pression, as did coexpression of the Fox1 and Fox2 shRNAs(Fig. 3A, lanes 3 and 4). In the control cells, exon 9* is includedin 48% of the endogenous CaV1.2 mRNA, as measured byRT-PCR. This is unchanged by the Fox1 shRNA (Fig. 3B,lanes 1 and 2). In contrast, the loss of Fox2 leads to an increasein exon 9* splicing to 59%, indicating a repressive effect ofFox2 on exon 9* splicing. The opposite is seen with exon 33.This exon is 85% included in the CaV1.2 mRNA in controltreated N2A cells (Fig. 3C lanes 1 and 2). Depletion of Fox2reduces exon 33 inclusion to ca. 50% (Fig. 3C, lanes 3 and 4).
The Fox2 depletion experiments indicate that Fox proteinsrepress exon 9* and enhance exon 33, as predicted by the timecourse of expression and splicing during development. To con-firm this, we performed Fox protein overexpression experi-ments. Untreated N2A cells show high levels of exon 33 inclu-sion, making it difficult to observe strong enhancement by Foxproteins. Instead, we used C2C12 myoblast cells that showlower basal levels of exon 33 inclusion. C2C12 cells were trans-fected with Fox1 or Fox2 expression plasmids or with controlvector. Immunoblot analysis indicated that in the transfectedcells, the Fox protein was increased by 55 to 60% over thecontrol cells (Fig. 4A). Exon 9* shows 33% inclusion in the
VOL. 29, 2009 Fox PROTEINS REGULATE L-TYPE CALCIUM CHANNEL SPLICING 4759
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 04
Feb
ruar
y 20
22 b
y 12
4.25
.181
.138
.

vector treated cells but is repressed to 24% inclusion by Fox1expression and to 15% inclusion by Fox2 (Fig. 4B, lanes 2 and3). Again, the opposite pattern is observed for exon 33. Fox1expression moderately increases exon 33 splicing from 64 to70%, whereas Fox2 has a stronger effect, increasing inclusion
to 85% (Fig. 4C, lanes 2 and 3). Thus, both Fox protein de-pletion and overexpression indicate that these regulators re-press exon 9* but enhance exon 33 splicing in the CaV1.2mRNA.
We also measured the dose response of exon 9* splicing to
FIG. 2. (A and B) Mouse genomic sequences encompassing exons 9* and 33. Exonic sequences are in uppercase and intronic sequence inlowercase. The exonic or intronic Fox binding elements “tgcatg” are underlined. (C) Immunoblots of protein in developing cortex show increasingFox protein expression between embryonic day 12 (E12) to embryonic day 18 (E18). The anti-Fox1, anti-Fox2, and anti-Fox3 antibodies eachrecognize an individual Fox protein, and the anti-Fox RRM antibody binds to the RRM found in all three proteins. Approximately 7 to 10embryonic cortices were pooled for protein extraction at each time point. The pooled cortices were weighed and homogenized in radioimmuno-precipitation assay buffer at 1:20 (wt/vol), yielding samples of approximately equal protein concentration. The final loading volumes of corticallysates were adjusted to obtain similar GAPDH immunoblot signals. �, anti. (D to F) Immunostaining of coronal sections from E18 mouse cortexwas visualized on an epifluorescence microscope (Eclipse-TE2000S; Nikon), at 10�10-fold magnification to cover the cortical plate (CxP),intermediate zone/subventricular zone (IZ/SVZ), and ventricle (V) regions within a same image. Fox RRM, Fox1 and Fox2 staining are shown inred (far left panels), while NeuN and MAP2 are shown in green (inner left panels). The overlaid staining of Fox and the neuronal markers (NeuNand MAP2) are shown in the inner right panels. The boxes in the overlay panels indicate the approximate regions shown at higher resolution inthe far right panels, which were examined at 10�40-fold magnification by confocal microscope (LSM 510 META; Zeiss). As seen previously (59),Fox protein staining is primarily nuclear (data not shown). (D) Costaining of Fox proteins with the postmitotic neuronal marker NeuN. Foxproteins are expressed in NeuN-positive layers of postmitotic neurons but not in proliferating cells of the intermediate zone/subventricular zone.The majority of neurons are costained by anti-Fox RRM and NeuN, with some stained only by anti-Fox RRM. (E) Immunostaining for Fox1 andthe neuronal marker MAP2. Both Fox1 and MAP2 staining is observed in the postmitotic neuronal layers. In the enlarged panel, the nuclear Fox1staining is surrounded by cytoplasmic MAP2. (F) Immunostaining for Fox2 and NeuN. Similar to anti-Fox RRM and anti-Fox1 staining, Fox2staining is found in postmitotic neurons.
4760 TANG ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 04
Feb
ruar
y 20
22 b
y 12
4.25
.181
.138
.

increasing amounts of transfected Fox1 or Fox2 plasmid in theN2a cells (Fig. 4D and E). Increasing amounts of Fox1 plasmidprogressively reduced exon 9* splicing from 46 to 24%. Simi-larly increasing levels of Fox2 expression reduced exon 9*inclusion from 44 to 27%. These exons respond to even smallchanges in the concentration of either Fox protein.
Regulation of exons 9* and 33 is dependent on Fox bindingelements. There are many examples of RNA-binding proteinsaffecting the expression of other regulators, which could in turnaffect the splicing of their own target exons. If CaV1.2 exons 9*and 33 are direct targets of the Fox proteins, then their splicingshould be dependent on Fox binding sites within the adjacentRNA. Several studies have indicated that the position of a Foxbinding site relative to a target exon will determine whetherthe protein will enhance or repress splicing. Upstream bindingsites are repressive, but downstream Fox sites generally act asenhancers (22, 59, 64). This is in agreement with what weobserve for exon 33, which contains a downstream Fox bindingsite and is enhanced by the protein. However, it is more diffi-cult to predict the behavior of exon 9*, which contains bothupstream and downstream Fox sites, as well as a Fox site in theexon itself. To demonstrate a direct role for the Fox proteins inthe splicing of these exons and to determine which Fox bindingsites are most important for their regulation, we constructedminigene splicing reporters for each exon. Each exon wascloned into the splicing reporter construct pDUP4-1 along withseveral hundred nucleotides of flanking intron sequence, in-
cluding all the most likely Fox binding sites (Fig. 5A and 6A;also see Fig. S3 in the supplemental material).
When expressed from the minigene in HEK293T cells, exon9* is 89% included in the RNA (Fig. 5B). Cotransfection of aFox1 or Fox2 expression plasmid strongly repressed this splic-ing in a dose-dependent manner, with exon 9* inclusion re-duced to 5 or 6% at the final titration point (Fig. 5B and C).
FIG. 3. Depletion of Fox proteins increases exon 9* and decreasesexon 33 splicing in N2a cells. (A) Immunoblot of Fox proteins in cellstreated with control vector (lane 1) and cells treated with short hairpinvectors targeting Fox1, Fox2, or both (lanes 2, 3, and 4). GAPDHexpression is monitored as the loading control. (B) RT-PCR assay ofexon 9* splicing in the control or Fox-depleted cells. (C) RT-PCRassay of exon 33 splicing in the control or Fox-depleted cells. �, anti.
FIG. 4. Overexpression of Fox1 and Fox2 proteins represses exon9* but enhances exon 33 splicing. (A) Immunoblot of Fox proteins inC2C12 cells transfected with Fox1 and Fox2 expression plasmids. Thebands for endogenous Fox proteins and coexpressed tagged Fox1 andFox2 proteins are indicated. The total Fox expression (endogenousplus recombinant) was normalized to the amount of GAPDH andindicated below. (B) RT-PCR assay of exon 9* in C2C12 cells trans-fected with pcDNA3.1 control vector or with Fox1 and Fox2 expressionplasmids. (C) RT-PCR assay of exon 33 in the same cells. (D) RT-PCRassay of exon 9* in N2a cells after transfection with increasing amountsof Fox1 and Fox2 expression plasmids. (E) Immunoblot of Fox1 and 2proteins in cells transfected with increasing doses of Fox1 and Fox2encoding plasmids. GAPDH expression served as the loading control.Note that this blot was exposed for a short time to avoid saturating thesignal from the transiently expressed protein and measure its increas-ing expression with increasing plasmid. The lower levels of endogenousFox2 in N2A cells can be observed upon longer exposure (see Fig. 3).�, anti.
VOL. 29, 2009 Fox PROTEINS REGULATE L-TYPE CALCIUM CHANNEL SPLICING 4761
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 04
Feb
ruar
y 20
22 b
y 12
4.25
.181
.138
.
Thus, in the minigene context the exon is highly sensitive to theFox protein concentration.
Each of the putative Fox binding elements upstream (u),within (e), or downstream (d) of exon 9* was mutated eithersingly or in combination with the other elements (Fig. 5A andD). These mutant minigenes were cotransfected with the Fox2expression plasmid or with a vector control; the splicing wasassayed by primer extension, and Fox protein expression wasassayed by immunoblotting. The wild-type minigene splicingwas reduced from 93% exon 9* inclusion in the vector controlto 14% inclusion with Fox2 coexpression (Fig. 5D lanes 1 and2). Exons carrying mutations in either the upstream or theexonic Fox sites were much less responsive to Fox2 protein,
with exon inclusion being reduced to 52 or 55% with Fox2expression (Fig. 5D, lanes 3 to 6). Combining the upstream andexonic mutations eliminates nearly all response to the Foxprotein; exon inclusion decreases from 97 to 94% with Fox2expression (Fig. 5D, lanes 9 and 10). Mutation of the down-stream element had little effect on the Fox response, with exoninclusion decreasing to 20% upon Fox2 expression (Fig. 5D,lanes 7 and 8). Combining this mutation with the upstream andexonic mutations also had only minimal effects (Fig. 5D, lanes11 to 16). Thus, the Fox-dependent repression of exon 9* ishighly dependent on Fox-binding elements within the exon andthe upstream intron. This repression is likely a direct effect ofthe Fox protein acting on the CaV1.2 RNA.
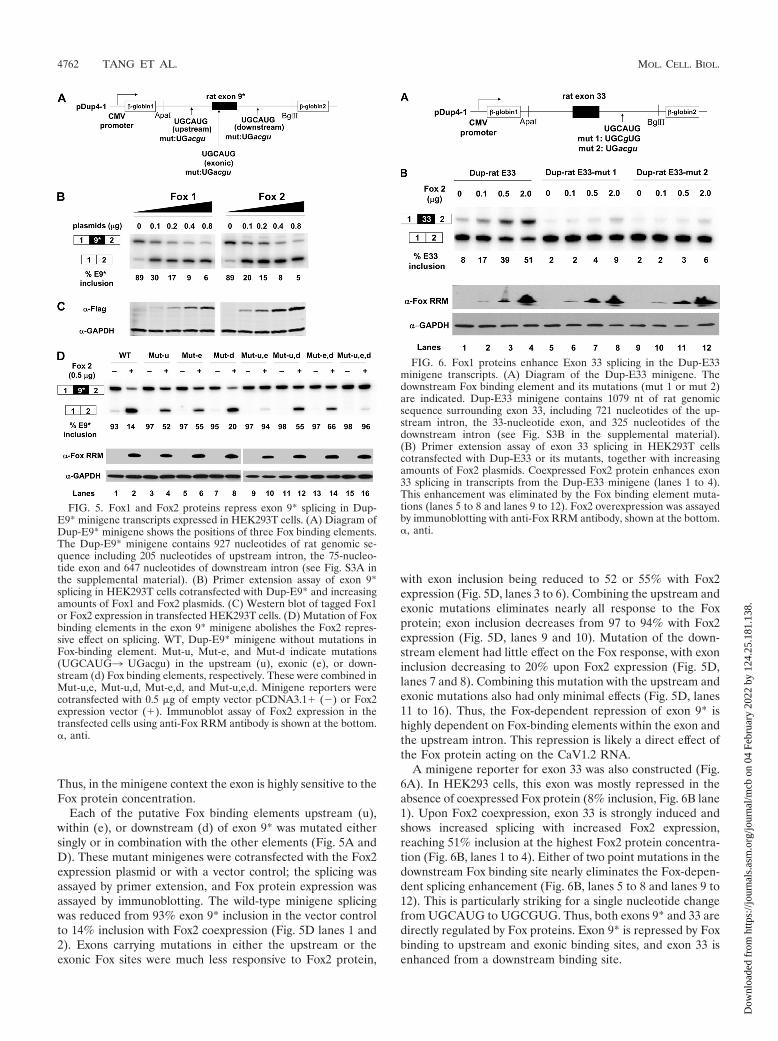
A minigene reporter for exon 33 was also constructed (Fig.6A). In HEK293 cells, this exon was mostly repressed in theabsence of coexpressed Fox protein (8% inclusion, Fig. 6B lane1). Upon Fox2 coexpression, exon 33 is strongly induced andshows increased splicing with increased Fox2 expression,reaching 51% inclusion at the highest Fox2 protein concentra-tion (Fig. 6B, lanes 1 to 4). Either of two point mutations in thedownstream Fox binding site nearly eliminates the Fox-depen-dent splicing enhancement (Fig. 6B, lanes 5 to 8 and lanes 9 to12). This is particularly striking for a single nucleotide changefrom UGCAUG to UGCGUG. Thus, both exons 9* and 33 aredirectly regulated by Fox proteins. Exon 9* is repressed by Foxbinding to upstream and exonic binding sites, and exon 33 isenhanced from a downstream binding site.
FIG. 5. Fox1 and Fox2 proteins repress exon 9* splicing in Dup-E9* minigene transcripts expressed in HEK293T cells. (A) Diagram ofDup-E9* minigene shows the positions of three Fox binding elements.The Dup-E9* minigene contains 927 nucleotides of rat genomic se-quence including 205 nucleotides of upstream intron, the 75-nucleo-tide exon and 647 nucleotides of downstream intron (see Fig. S3A inthe supplemental material). (B) Primer extension assay of exon 9*splicing in HEK293T cells cotransfected with Dup-E9* and increasingamounts of Fox1 and Fox2 plasmids. (C) Western blot of tagged Fox1or Fox2 expression in transfected HEK293T cells. (D) Mutation of Foxbinding elements in the exon 9* minigene abolishes the Fox2 repres-sive effect on splicing. WT, Dup-E9* minigene without mutations inFox-binding element. Mut-u, Mut-e, and Mut-d indicate mutations(UGCAUG3 UGacgu) in the upstream (u), exonic (e), or down-stream (d) Fox binding elements, respectively. These were combined inMut-u,e, Mut-u,d, Mut-e,d, and Mut-u,e,d. Minigene reporters werecotransfected with 0.5 �g of empty vector pCDNA3.1� (�) or Fox2expression vector (�). Immunoblot assay of Fox2 expression in thetransfected cells using anti-Fox RRM antibody is shown at the bottom.�, anti.
FIG. 6. Fox1 proteins enhance Exon 33 splicing in the Dup-E33minigene transcripts. (A) Diagram of the Dup-E33 minigene. Thedownstream Fox binding element and its mutations (mut 1 or mut 2)are indicated. Dup-E33 minigene contains 1079 nt of rat genomicsequence surrounding exon 33, including 721 nucleotides of the up-stream intron, the 33-nucleotide exon, and 325 nucleotides of thedownstream intron (see Fig. S3B in the supplemental material).(B) Primer extension assay of exon 33 splicing in HEK293T cellscotransfected with Dup-E33 or its mutants, together with increasingamounts of Fox2 plasmids. Coexpressed Fox2 protein enhances exon33 splicing in transcripts from the Dup-E33 minigene (lanes 1 to 4).This enhancement was eliminated by the Fox binding element muta-tions (lanes 5 to 8 and lanes 9 to 12). Fox2 overexpression was assayedby immunoblotting with anti-Fox RRM antibody, shown at the bottom.�, anti.
4762 TANG ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 04
Feb
ruar
y 20
22 b
y 12
4.25
.181
.138
.

DISCUSSION
Developmental regulation of calcium channel physiologythrough changes in splicing. We found that the Fox proteinscontrol two exons in the CaV1.2 pre-mRNA. The induction ofFox protein expression during neurogenesis leads to the pro-gressive repression of exon 9* and the progressive inclusion ofexon 33. Both of these exons have been shown to affect theelectrophysiological properties CaV1.2 channels (29). Exon 33encodes an 11-amino-acid insert into the IVS3-IVS4 extracel-lular loop, adjacent to IVS4 voltage sensor region of the chan-nel. Lipscombe et al. have noted that changes in this linker mayalter the movement of the adjoining S4 helix and thus influencethe kinetics of channel activation (33). Studies of L-type, P/Q-type, and N-type calcium channels, as well as of potassiumchannels have all shown that the IVS3-IVS4 linker is an im-portant region in coupling voltage-sensing to channel opening(10, 20, 32–34, 55, 58).
When expressed in HEK293T cells, CaV1.2 channels con-taining the exon 9* encoded peptide exhibit a �10-mV hyper-polarized shift (left shift) in their current/voltage relationshipcurve compared to channels lacking the 9* peptide (31, 55).The exclusion of exon 33 in channels that contain exon 9*(CaV1.2�E9*�E33) shifts both the activation and the inacti-vation potential of the channel by 8 to 10 mV in the samehyperpolarized direction (30, 54). The exclusion of exon 33also makes the channel more sensitive to nifedipine inhibition.Thus, both the decreased splicing of exon 9* and the increasedsplicing of exon 33, seen in response to Fox proteins, shouldlead to channels with a more positive depolarizing potential foractivation and inactivation. The effect of these changes oncellular activity will need examination in model systems withbetter defined physiology and in the context of single cells.Although the cellular consequences of these changes in chan-nel properties are not known, it is clear that the Fox proteinsare playing significant roles in fine-tuning calcium channelphysiology during neuronal development and likely in maturecells.
In addition to the CaV1.2 channel, there are three otherL-type calcium channel genes: CaV1.1, CaV1.3, and CaV1.4(14, 17, 33). The gene structures of these pore-forming sub-units are highly conserved and exhibit alternative splicing inmany of the same regions. The CaV1.1, CaV1.3, and CaV1.4transcripts also contain alternative exons in the IVS3-IVS4linker region similar to CaV1.2 exon 33 (4, 26, 43, 51, 55, 58).Interestingly, Fox binding elements ((U)GCAUG) are founddownstream of all of these IVS3-IVS4 exons (Fig. 7) (64).CaV1.3 also contains an alternatively spliced 26-amino-acidexon 9a in the I-II loop similar to CaV1.2 exon 9* (26, 46). This9a exon has one TGCATG and one GCATG element in itsdownstream intron, suggesting regulation by Fox proteins butperhaps in the opposite direction than exon 9* of CaV1.2. Wealso found conserved Fox sites adjacent to alternative exons inother types of calcium channels, as well as exons in calciumtransport proteins. These observations indicate that beyondtheir alteration of the L-type calcium channel, the Fox proteinsare likely affecting other aspects of calcium signaling and cal-cium ion homeostasis in excitable cells.
An alternatively spliced exon is generally under the controlof multiple splicing regulators, and it is likely that other splic-
ing factors are affecting exons 9* and 33 (7, 8, 35, 37, 60). TheFox binding sites studied here are within larger highly con-served intronic regions that presumably include binding sitesfor other proteins. Indeed, we found that the PTB/nPTB splic-ing factors also affect these exons, although less strongly (Z. Z.Tang and D. L. Black, unpublished data). Notably, CaV1.2channels containing exon 9* and excluding exon 33 are ex-pressed in smooth muscle cells that also express Fox1 andFox2. In this cellular context, there must be other factorsdetermining the use of these exons. In addition, there are atleast 12 other CaV1.2 alternative exons, and many more pro-tein regulators will be needed to generate the complex splicingpatterns seen for this gene (1, 54, 55). Of particular interest arethe factors controlling exons 8a and 8, which are mutated in theTimothy Syndrome (52, 53) and factors controlling exons al-tered in cardiovascular disease.
ACKNOWLEDGMENTS
We thank Andrey Damianov and Peter Stoilov for the anti-FoxRRM antibody and the pBlsH1Fox-1 and pBlsH1Fox-2 plasmid con-structs.
This study was supported in part by NIH grant RO1 GM49662 toD.L.B. D.L.B. is an Investigator of the Howard Hughes Medical In-stitute.
REFERENCES
1. Abernethy, D. R., and N. M. Soldatov. 2002. Structure-functional diversity ofhuman L-type Ca2� channel: perspectives for new pharmacological targets.J. Pharmacol. Exp. Ther. 300:724–728.
2. Auweter, S. D., R. Fasan, L. Reymond, J. G. Underwood, D. L. Black, S.Pitsch, and F. H. Allain. 2006. Molecular basis of RNA recognition by thehuman alternative splicing factor Fox-1. EMBO J. 25:163–173.
3. Baraniak, A. P., J. R. Chen, and M. A. Garcia-Blanco. 2006. Fox-2 mediatesepithelial cell-specific fibroblast growth factor receptor 2 exon choice. Mol.Cell. Biol. 26:1209–1222.
4. Barry, E. L., F. A. Gesek, S. C. Froehner, and P. A. Friedman. 1995. Multiplecalcium channel transcripts in rat osteosarcoma cells: selective activation ofalpha 1D isoform by parathyroid hormone. Proc. Natl. Acad. Sci. USA92:10914–10918.
5. Biel, M., P. Ruth, E. Bosse, R. Hullin, W. Stuhmer, V. Flockerzi, and F.Hofmann. 1990. Primary structure and functional expression of a high-voltage activated calcium channel from rabbit lung. FEBS Lett. 269:409–412.
6. Bito, H., K. Deisseroth, and R. W. Tsien. 1997. Ca2�-dependent regulationin neuronal gene expression. Curr. Opin. Neurobiol. 7:419–429.
7. Black, D. L. 2003. Mechanisms of alternative pre-messenger RNA splicing.Annu. Rev. Biochem. 72:291–336.
8. Blencowe, B. J. 2006. Alternative splicing: new insights from global analyses.Cell 126:37–47.
FIG. 7. Alternative cassette exons in the IVS3-IVS4 regions of thefour human L-type calcium channel genes all contain conserved Foxbinding elements in downstream intron. Alternative exons are in up-percase and the 300 nucleotides of downstream intronic sequence is inlowercase. There are no “tgcatg” or “gcatg” elements in the 200 nu-cleotides of intronic sequence upstream these alternative exons.
VOL. 29, 2009 Fox PROTEINS REGULATE L-TYPE CALCIUM CHANNEL SPLICING 4763
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 04
Feb
ruar
y 20
22 b
y 12
4.25
.181
.138
.

9. Bodi, I., G. Mikala, S. E. Koch, S. A. Akhter, and A. Schwartz. 2005. TheL-type calcium channel in the heart: the beat goes on. J. Clin. Investig.115:3306–3317.
10. Bourinet, E., T. W. Soong, K. Sutton, S. Slaymaker, E. Mathews, A. Monteil,G. W. Zamponi, J. Nargeot, and T. P. Snutch. 1999. Splicing of �1A subunitgene generates phenotypic variants of P- and Q-type calcium channels. Nat.Neurosci. 2:407–415.
11. Boutz, P. L., G. Chawla, P. Stoilov, and D. L. Black. 2007. MicroRNAsregulate the expression of the alternative splicing factor nPTB during muscledevelopment. Genes Dev. 21:71–84.
12. Boutz, P. L., P. Stoilov, Q. Li, C. H. Lin, G. Chawla, K. Ostrow, L. Shiue, M.Ares, Jr., and D. L. Black. 2007. A posttranscriptional regulatory switch inpolypyrimidine tract-binding proteins reprograms alternative splicing in de-veloping neurons. Genes Dev. 21:1636–1652.
13. Cartin, L., K. M. Lounsbury, and M. T. Nelson. 2000. Coupling of Ca2� toCREB activation and gene expression in intact cerebral arteries from mouse:roles of ryanodine receptors and voltage-dependent Ca2� channels. Circ.Res. 86:760–767.
14. Catterall, W. A., E. Perez-Reyes, T. P. Snutch, and J. Striessnig. 2005.International Union of Pharmacology. XLVIII. Nomenclature and struc-ture-function relationships of voltage-gated calcium channels. Pharmacol.Rev. 57:411–425.
15. Cheng, X., J. Liu, M. Asuncion-Chin, E. Blaskova, J. P. Bannister, A. M.Dopico, and J. H. Jaggar. 2007. A novel Ca(V)1.2 N terminus expressed insmooth muscle cells of resistance size arteries modifies channel regulation byauxiliary subunits. J. Biol. Chem. 282:29211–29221.
16. Dolmetsch, R. E., U. Pajvani, K. Fife, J. M. Spotts, and M. E. Greenberg.2001. Signaling to the nucleus by an L-type calcium channel-calmodulincomplex through the MAP kinase pathway. Science 294:333–339.
17. Ertel, E. A., K. P. Campbell, M. M. Harpold, F. Hofmann, Y. Mori, E.Perez-Reyes, A. Schwartz, T. P. Snutch, T. Tanabe, L. Birnbaumer, R. W.Tsien, and W. A. Catterall. 2000. Nomenclature of voltage-gated calciumchannels. Neuron 25:533–535.
18. Fearon, I. M., G. Varadi, S. Koch, I. Isaacsohn, S. G. Ball, and C. Peers.2000. Splice variants reveal the region involved in oxygen sensing by recom-binant human L-type Ca2� channels. Circ. Res. 87:537–539.
19. Graf, E. M., M. Bock, J. F. Heubach, I. Zahanich, S. Boxberger, W. Richter,J. H. Schultz, and U. Ravens. 2005. Tissue distribution of a human Ca v 1.2�1 subunit splice variant with a 75-bp insertion. Cell Calcium 38:11–21.
20. Hans, M., A. Urrutia, C. Deal, P. F. Brust, K. Stauderman, S. B. Ellis, M. M.Harpold, E. C. Johnson, and M. E. Williams. 1999. Structural elements indomain IV that influence biophysical and pharmacological properties ofhuman �1A-containing high-voltage-activated calcium channels. Biophys. J.76:1384–1400.
21. Hodgkin, J., J. D. Zellan, and D. G. Albertson. 1994. Identification of acandidate primary sex determination locus, fox-1, on the X chromosome ofCaenorhabditis elegans. Development 120:3681–3689.
22. Jin, Y., H. Suzuki, S. Maegawa, H. Endo, S. Sugano, K. Hashimoto, K.Yasuda, and K. Inoue. 2003. A vertebrate RNA-binding protein Fox-1 reg-ulates tissue-specific splicing via the pentanucleotide GCAUG. EMBO J.22:905–912.
23. Kiehl, T. R., H. Shibata, T. Vo, D. P. Huynh, and S. M. Pulst. 2001. Iden-tification and expression of a mouse ortholog of A2BP1. Mamm. Genome12:595–601.
24. Klockner, U., G. Mikala, J. Eisfeld, D. E. Iles, M. Strobeck, J. L. Mershon,A. Schwartz, and G. Varadi. 1997. Properties of three COOH-terminal splicevariants of a human cardiac L-type Ca2�-channel �1-subunit. Am. J. Physiol.272:H1372–H1381.
25. Koch, W. J., A. Hui, G. E. Shull, P. Ellinor, and A. Schwartz. 1989. Char-acterization of cDNA clones encoding two putative isoforms of the alpha 1subunit of the dihydropyridine-sensitive voltage-dependent calcium channelisolated from rat brain and rat aorta. FEBS Lett. 250:386–388.
26. Kollmar, R., L. G. Montgomery, J. Fak, L. J. Henry, and A. J. Hudspeth.1997. Predominance of the alpha1D subunit in L-type voltage-gated Ca2�
channels of hair cells in the chicken’s cochlea. Proc. Natl. Acad. Sci. USA94:14883–14888.
27. Lane, H. A. 1988. Antibodies: a laboratory manual. Cold Spring HarborLaboratory Press, Cold Spring Harbor, NY.
28. Lee, J. A., Y. Xing, D. Nguyen, J. Xie, C. J. Lee, and D. L. Black. 2007.Depolarization and CaM kinase IV modulate NMDA receptor splicingthrough two essential RNA elements. PLoS Biol. 5:e40.
29. Liao, P., T. F. Yong, M. C. Liang, D. T. Yue, and T. W. Soong. 2005. Splicingfor alternative structures of Cav1.2 Ca2� channels in cardiac and smoothmuscles. Cardiovasc. Res. 68:197–203.
30. Liao, P., D. Yu, G. Li, T. F. Yong, J. L. Soon, Y. L. Chua, and T. W. Soong.2007. A smooth muscle Cav1.2 calcium channel splice variant underlieshyperpolarized window current and enhanced state-dependent inhibition bynifedipine. J. Biol. Chem. 282:35133–35142.
31. Liao, P., D. Yu, S. Lu, Z. Tang, M. C. Liang, S. Zeng, W. Lin, and T. W.Soong. 2004. Smooth muscle-selective alternatively spliced exon generatesfunctional variation in Cav1.2 calcium channels. J. Biol. Chem. 279:50329–50335.
32. Lin, Z., Y. Lin, S. Schorge, J. Q. Pan, M. Beierlein, and D. Lipscombe. 1999.Alternative splicing of a short cassette exon in �1B generates functionallydistinct N-type calcium channels in central and peripheral neurons. J. Neu-rosci. 19:5322–5331.
33. Lipscombe, D., J. Q. Pan, and A. C. Gray. 2002. Functional diversity inneuronal voltage-gated calcium channels by alternative splicing of Ca(v) �1.Mol. Neurobiol 26:21–44.
34. Mathur, R., J. Zheng, Y. Yan, and F. J. Sigworth. 1997. Role of the S3–S4linker in Shaker potassium channel activation. J. Gen. Physiol. 109:191–199.
35. Matlin, A. J., F. Clark, and C. W. Smith. 2005. Understanding alternativesplicing: toward a cellular code. Nat. Rev. Mol. Cell. Biol. 6:386–398.
36. McKee, A. E., E. Minet, C. Stern, S. Riahi, C. D. Stiles, and P. A. Silver.2005. A genome-wide in situ hybridization map of RNA-binding proteinsreveals anatomically restricted expression in the developing mouse brain.BMC Dev. Biol. 5:14.
37. Modafferi, E. F., and D. L. Black. 1999. Combinatorial control of a neuron-specific exon. RNA 5:687–706.
38. Modafferi, E. F., and D. L. Black. 1997. A complex intronic splicing enhancerfrom the c-src pre-mRNA activates inclusion of a heterologous exon. Mol.Cell. Biol. 17:6537–6545.
39. Moosmang, S., P. Lenhardt, N. Haider, F. Hofmann, and J. W. Wegener.2005. Mouse models to study L-type calcium channel function. Pharmacol.Ther. 106:347–355.
40. Nakahata, S., and S. Kawamoto. 2005. Tissue-dependent isoforms of mam-malian Fox-1 homologs are associated with tissue-specific splicing activities.Nucleic Acids Res. 33:2078–2089.
41. Nicoll, M., C. C. Akerib, and B. J. Meyer. 1997. X-chromosome-countingmechanisms that determine nematode sex. Nature 388:200–204.
42. Pang, L., G. Koren, Z. Wang, and S. Nattel. 2003. Tissue-specific expressionof two human Ca(v)1.2 isoforms under the control of distinct 5� flankingregulatory elements. FEBS Lett. 546:349–354.
43. Perez-Reyes, E., X. Y. Wei, A. Castellano, and L. Birnbaumer. 1990. Molec-ular diversity of L-type calcium channels. Evidence for alternative splicing ofthe transcripts of three non-allelic genes. J. Biol. Chem. 265:20430–20436.
44. Ponthier, J. L., C. Schluepen, W. Chen, R. A. Lersch, S. L. Gee, V. C. Hou,A. J. Lo, S. A. Short, J. A. Chasis, J. C. Winkelmann, and J. G. Conboy. 2006.Fox-2 splicing factor binds to a conserved intron motif to promote inclusionof protein 4.1R alternative exon 16. J. Biol. Chem. 281:12468–12474.
45. Saada, N. I., E. D. Carrillo, B. Dai, W. Z. Wang, C. Dettbarn, J. Sanchez, andP. Palade. 2005. Expression of multiple CaV1.2 transcripts in rat tissuesmediated by different promoters. Cell Calcium 37:301–309.
46. Safa, P., J. Boulter, and T. G. Hales. 2001. Functional properties of Cav1.3(�1D) L-type Ca2� channel splice variants expressed by rat brain and neu-roendocrine GH3 cells. J. Biol. Chem. 276:38727–38737.
47. Sinnegger-Brauns, M. J., A. Hetzenauer, I. G. Huber, E. Renstrom, G.Wietzorrek, S. Berjukov, M. Cavalli, D. Walter, A. Koschak, R. Waldschutz,S. Hering, S. Bova, P. Rorsman, O. Pongs, N. Singewald, and J. Striessnig.2004. Isoform-specific regulation of mood behavior and pancreatic beta celland cardiovascular function by L-type Ca2� channels. J. Clin. Investig. 113:1430–1439.
48. Skipper, M., C. A. Milne, and J. Hodgkin. 1999. Genetic and molecularanalysis of fox-1, a numerator element involved in Caenorhabditis elegansprimary sex determination. Genetics 151:617–631.
49. Snutch, T. P., W. J. Tomlinson, J. P. Leonard, and M. M. Gilbert. 1991.Distinct calcium channels are generated by alternative splicing and are dif-ferentially expressed in the mammalian CNS. Neuron 7:45–57.
50. Soldatov, N. M. 1994. Genomic structure of human L-type Ca2� channel.Genomics 22:77–87.
51. Soldatov, N. M. 1992. Molecular diversity of L-type Ca2� channel transcriptsin human fibroblasts. Proc. Natl. Acad. Sci. USA 89:4628–4632.
52. Splawski, I., K. W. Timothy, N. Decher, P. Kumar, F. B. Sachse, A. H. Beggs,M. C. Sanguinetti, and M. T. Keating. 2005. Severe arrhythmia disordercaused by cardiac L-type calcium channel mutations. Proc. Natl. Acad. Sci.USA 102:8086–8096.
53. Splawski, I., K. W. Timothy, L. M. Sharpe, N. Decher, P. Kumar, R. Bloise,C. Napolitano, P. J. Schwartz, R. M. Joseph, K. Condouris, H. Tager-Flusberg, S. G. Priori, M. C. Sanguinetti, and M. T. Keating. 2004. Ca(V)1.2calcium channel dysfunction causes a multisystem disorder including ar-rhythmia and autism. Cell 119:19–31.
54. Tang, Z. Z., X. Hong, J. Wang, and T. W. Soong. 2007. Signature combina-torial splicing profiles of rat cardiac- and smooth-muscle Cav1.2 channelswith distinct biophysical properties. Cell Calcium 41:417–428.
55. Tang, Z. Z., M. C. Liang, S. Lu, D. Yu, C. Y. Yu, D. T. Yue, and T. W. Soong.2004. Transcript scanning reveals novel and extensive splice variations inhuman L-type voltage-gated calcium channel, Cav1.2 �1 subunit. J. Biol.Chem. 279:44335–44343.
56. Tang, Z. Z., P. Liao, G. Li, F. L. Jiang, D. Yu, X. Hong, T. F. Yong, G. Tan,S. Lu, J. Wang, and T. W. Soong. 2008. Differential splicing patterns ofL-type calcium channel Ca(v)1.2 subunit in hearts of spontaneously hyper-tensive rats and Wistar Kyoto rats. Biochim. Biophys. Acta 1783:118–130.
57. Tiwari, S., Y. Zhang, J. Heller, D. R. Abernethy, and N. M. Soldatov. 2006.
4764 TANG ET AL. MOL. CELL. BIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 04
Feb
ruar
y 20
22 b
y 12
4.25
.181
.138
.

Atherosclerosis-related molecular alteration of the human CaV1.2 calciumchannel �1C subunit. Proc. Natl. Acad. Sci. USA 103:17024–17029.
58. Tuluc, P., N. Molenda, B. Schlick, G. J. Obermair, B. E. Flucher, and K.Jurkat-Rott. 2009. A Ca(V)1.1 Ca2� channel splice variant with high con-ductance and voltage-sensitivity alters EC coupling in developing skeletalmuscle. Biophys. J. 96:35–44.
59. Underwood, J. G., P. L. Boutz, J. D. Dougherty, P. Stoilov, and D. L. Black.2005. Homologues of the Caenorhabditis elegans Fox-1 protein are neuronalsplicing regulators in mammals. Mol. Cell. Biol. 25:10005–10016.
60. Wang, Z., and C. B. Burge. 2008. Splicing regulation: from a parts list ofregulatory elements to an integrated splicing code. RNA 14:802–813.
61. Welling, A., Y. W. Kwan, E. Bosse, V. Flockerzi, F. Hofmann, and R. S.Kass. 1993. Subunit-dependent modulation of recombinant L-type calciumchannels: molecular basis for dihydropyridine tissue selectivity. Circ. Res. 73:974–980.
62. Welling, A., A. Ludwig, S. Zimmer, N. Klugbauer, V. Flockerzi, and F.Hofmann. 1997. Alternatively spliced IS6 segments of the �1C gene deter-
mine the tissue-specific dihydropyridine sensitivity of cardiac and vascularsmooth muscle L-type Ca2� channels. Circ. Res. 81:526–532.
63. Yeo, G. W., N. G. Coufal, T. Y. Liang, G. E. Peng, X. D. Fu, and F. H.Gage. 2009. An RNA code for the FOX2 splicing regulator revealed bymapping RNA-protein interactions in stem cells. Nat. Struct. Mol. Biol.16:130–137.
64. Zhang, C., Z. Zhang, J. Castle, S. Sun, J. Johnson, A. R. Krainer, and M. Q.Zhang. 2008. Defining the regulatory network of the tissue-specific splicingfactors Fox-1 and Fox-2. Genes Dev. 22:2550–2563.
65. Zhou, H. L., A. P. Baraniak, and H. Lou. 2007. Role for Fox-1/Fox-2 inmediating the neuronal pathway of calcitonin/calcitonin gene-related pep-tide alternative RNA processing. Mol. Cell. Biol. 27:830–841.
66. Zhou, H. L., and H. Lou. 2008. Repression of prespliceosome complexformation at two distinct steps by Fox-1/Fox-2 proteins. Mol. Cell. Biol.28:5507–5516.
67. Zuhlke, R. D., A. Bouron, N. M. Soldatov, and H. Reuter. 1998. Ca2�
channel sensitivity toward the blocker isradipine is affected by alternativesplicing of the human �1C subunit gene. FEBS Lett. 427:220–224.
VOL. 29, 2009 Fox PROTEINS REGULATE L-TYPE CALCIUM CHANNEL SPLICING 4765
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 04
Feb
ruar
y 20
22 b
y 12
4.25
.181
.138
.