detection and identification of human pathogenic ...dna.kdna.ucla.edu/simpsonlab/lab...
TRANSCRIPT

JOBNAME: JEP 82#3 96 PAGE: 1 SESS: 3 OUTPUT: Wed Jun 5 02:28:21 1996/xypage/worksmart/tsp000/69001j/4pu
Detection and Identification of Human Pathogenic Leishmania andTrypanosoma Species by Hybridization of PCR-Amplified
Mini-exon Repeats
ANTHEA RAMOS,* DMITRI A. MASLOV,†,1 OCTAVIO FERNANDES,‡ DAVID A. CAMPBELL,§ AND
LARRY SIMPSON,*,†,2
*Howard Hughes Medical Institute, †Department of Biology, and §Medical Microbiology and Immunology, Universityof California, Los Angeles, California, U.S.A.; and ‡Departmento de Patologia, Universidade do Estado do Rio de
Janeiro, Rio de Janeiro, Brazil
RAMOS, A., MASLOV, D. A., FERNANDES, O., CAMPBELL, D. A., AND SIMPSON, L. 1995. Detectionand identification of human pathogenicLeishmaniaandTrypanosomaspecies by hybridization ofPCR-amplified mini-exon repeats.Experimental Parasitology82, 242–250. A single pair of PCRprimers within a conserved region of the mini-exon repeat was used to amplify the repeats from 10species of pathogenicLeishmaniabelonging to four major clinical groups and also from three speciesof Trypanosoma.Oligonucleotide hybridization probes for the detection and identification of thePCR-amplified repeats were constructed from alignments of mini-exon intron and intergenic se-quences. The probes generated from mini-exon intergenic regions of theL. (V.) braziliensis, L.(L.)donovani,and L. (L.) mexicanaspecies hybridized specifically to their cognate groups withoutdiscriminating between the species within the groups. The probes forL. (L.) major and L. (L.)aethiopicawere species-specific, while theL. (L.) tropica probe also hybridized with theL. (L.)aethiopicamini-exon repeat. The mini-exon intron-derived probes forT. cruzi, T. rangeli,andT.bruceiwere species-specific. This method involving the detection of specific PCR-amplified productsproduced using a single primer set represents a novel sensitive and specific assay for multipletrypanosomatid species and groups.© 1996 Academic Press, Inc.
INDEX DESCRIPTORS ANDABBREVIATIONS: Leishmania; Trypanosoma; oligonucleotide probe; Ply-merase chain reaction (PCR); mini-exon gene repeat.
INTRODUCTION
PathogenicLeishmaniaand Trypanosomaspecies are the causal agents of several impor-tant human and animal diseases, such as dermaland visceral leishmaniasis, Chagas’ disease, andAfrican sleeping sickness. Rapid detection andidentification of pathogenic trypanosomatids inpatients, animal reservoirs, or insect vectorsrepresent an important problem. PCR amplifi-cation of specific parasite DNA sequences hasthe advantage over serological assays of provid-ing direct evidence for the presence of parasitesin the host and also, in view of the inherent
sequence polymorphisms of the parasite ge-nome, of allowing a classification of the speciesor possibly even the strain of the parasite.Several methods utilizing PCR amplification
of parasite nuclear DNA or kinetoplastminicircle DNA target sequences have been re-ported forTrypanosoma cruzi(Gonzalezet al.1984; Sturmet al. 1989; Britto et al. 1995;Winckeret al. 1994; Avilaet al. 1993; Laskayet al.1991) and pathogenicLeishmaniaspecies(Wirth and Pratt 1982; Lopeset al.1984; Lopesand Wirth 1986; Wirthet al. 1986; Rodgersetal. 1990; Howardet al. 1991; Ulianaet al.1994). A common feature of these methods isthe use of specific primer sets and reactions foreach particular parasite or species. The use ofmultiple PCR amplifications substantially in-creases the cost and difficulty of analysis. Mul-tiplex PCR, a method in which a mixture ofprimer sets is used, with each specific for theDNA from a particular organism, should theo-
1 Present affiliation: Department of Biology, Universityof California, Riverside, CA 92521.
2 To whom reprint requests and correspondence shouldbe addressed at Howard Hughes Medical Institute, UCLA,6-780 MacDonald Building, 675 Circle Drive S., Box951662, Los Angeles, CA 90095-1662. Fax: (310) 206-8967. E-mail: [email protected].
EXPERIMENTAL PARASITOLOGY82, 242–250 (1996)ARTICLE NO. 0031
2420014-4894/96 $18.00Copyright © 1996 by Academic Press, Inc.All rights of reproduction in any form reserved.

JOBNAME: JEP 82#3 96 PAGE: 2 SESS: 2 OUTPUT: Wed Jun 5 02:28:21 1996/xypage/worksmart/tsp000/69001j/4pu
retically overcome this problem and allow thedetection of multiple etiological agents in thesame patient, animal, or insect with a singleassay. However, attempts in our laboratory tocoamplify several sequences (T. cruziminicircle DNA, hepatitis B DNA, cytomega-lovirus DNA, HIV DNA) led to a substantialloss of sensitivity and specificity, apparently asa result of interactions of heterologous primersand templates (L. Simpson, D.A. Maslov, andA. Ramos, data not shown).The mini-exon genes of the trypanosomatid
protozoa represent a good target for a PCR-based multiple detection and identification as-say, which can overcome the inherent problemsof multiplex PCR. Mini-exon-derived RNA isinvolved in trans-splicing of mRNA, a processrestricted to the kinetoplastid protozoa andsome worms (De Langeet al.1983, 1986; Aga-bian 1990; Murphyet al. 1986; Sutton andBoothroyd 1986; Campbellet al. 1984; Nilsen1993). Mini-exon genes are absent from the ver-tebrate hosts and invertebrate vectors, but arepresent as multiple tandem repeats in the para-site genomes. Each repeat contains a highlyconserved 39-nt exon sequence, a moderatelyvariable 55- to 101-nt transcribed intron se-quence, and a highly variable 250- to 1350-ntnontranscribed intergenic region. The lengthand nucleotide sequence of the mini-exon re-peat was shown to vary between differentgroups of pathogenicLeishmaniaspecies(Hassanet al.1993; Fernandeset al.1994) andbetween the trypanosomesT. cruziandT. ran-geli (Murthy et al.1992). In this paper we showthat specific and sensitive detection of mini-exon repeats PCR-amplified from pathogenicLeishmaniaandTrypanosomaspecies using theconserved exon sequence can be accomplishedusing specific capture oligomers derived fromthe polymorphic intron and intergenic regions.
MATERIALS AND METHODS
Strains of parasites and growth conditions.The strainsof LeishmaniaandTrypanosomaand their sources are asfollows: L. (L.) amazonensisPH8 (Ralph Lainson, InstitutoEvandro Chagas, Belem, Brazil), JOS (R. McMaster, Uni-versity of British Columbia, Canada), M2269 (G. Grimaldi,FIOCRUZ, Brazil);L. (L.) mexicanaM379 (D. McMahon-
Pratt, Yale University, U.S.A.), BEL21 (G. Grimaldi);L.(V.) braziliensisM2903 (D. McMahon-Pratt), LT300 (R.McMaster); L. (L.) chagasiBA3 (R. McMaster);L. (V.)panamensisWR120 (D. McMahon-Pratt);L. (V.) guyanen-sis M4147 (G. Grimaldi);L. (L.) aethiopica L100 (G.Grimaldi); L. (L.) major LT252 (S. Beverley, Harvard Uni-versity, U.S.A.), NIH-S (R. McMaster), A2 (R. McMaster);L. (L.) tropicaOD (G. Grimaldi);L. (L.) donovaniDD8 (G.Grimaldi), MRC-74 (D. McMahon-Pratt);L. (L.) infantumITMAP263 (K. Stuart, Seattle Biomedical Research Insti-tute, U.S.A.), IPTI (G. Grimaldi);T. (T.) brucei427 (Camp-bell et al. 1984);T. (S.) cruzi Sylvio (Dvoraket al. 1980,1982), MT01, MT03, MT05-26 (J. Coura, FIOCRUZ, Bra-zil); T. (H.) rangeliEv26 (N. Saravia, CIDEIM, Colombia)(Holguin et al. 1987), MT02 and MT04 (J. Coura).Leishmaniapromastigotes were grown at 26°C in
HOMEM (Berenset al. 1976) medium supplemented with10% heat-inactivated fetal bovine serum (Gemini Bio-Products, Calabasas, CA).T. bruceiprocyclics were grownin SDM-79 medium (Brun and Schoenenberger 1979). Epi-mastigotes ofT. cruzi and T. rangeliwere grown in LITmedium (Castellaniet al. 1967) supplemented with 10mg/ml hemin and 10% serum.Isolation of DNA.Cells from 10-ml stationary phase cul-
tures were pelleted at 2000g for 10 min and washed with anequal volume of SET (10 mM Tris–HCl, pH 8.0, 150 mMNaCl, 100 mM EDTA). The final pellet was suspended in0.5 ml SET and lysed with 2% Sarcosyl and 0.5 mg/mlpronase at 65°C for 30 min. The lysate was phenol–chloroform extracted and the DNA was precipitated with anequal volume of isopropanol.Oligonucleotides.The oligonucleotides shown in Fig. 1
were used in this study.PCR amplification.Reaction mixtures contained 20 mM
Tris–HCl, pH 8.3, 1.5 mM MgCl2, 25 mM KCl, 0.05%Tween-20, 0.1 mg/ml bovine serum albumin, 10% dimeth-ylsulfoxide (forLeishmaniaspecies only), 250mM of eachdNTP, 20mM of each amplification primer and 2.5 U ofTaqDNA polymerase. Conditions included initial denatur-ation at 95°C for 5 min followed by 5 cycles at 95°C for 1min, 45°C for 30 sec, 65°C for 1 min and 35 cycles at 95°Cfor 1 min, 50°C for 30 sec, 72°C for 1 min, and a finalextension at 72°C for 10 min.Blotting and hybridization.Aliquots (4–10%) of the am-
plified material were separated on 2% agarose gels andblotted onto a nylon filter (Micron Separations). Oligonu-cleotides were labeled by [g-32P]ATP and T4 polynucleo-tide kinase to specific activities of 108–109 cpm/mg. Hy-bridizations in Figs. 2 and 4 were performed at 37°C in 6×SSC, 1× Denhard’s solution, 0.1% SDS, and 100mg/ml ofsonicated salmon sperm DNA in rotating bottles. Filterswere washed briefly in 3× SSC (1× SSC contains 150 mMNaCl and 15 mM sodium citrate) at room temperature fol-lowed by washes in 3M tetramethylammonium chloride(Fisher), 50 mM Tris–HCl, pH 8.0, 2 mM EDTA, 0.5% SDSat 55°C. High-stringency washes were performed in 2×SSC, 0.1% SDS at 65°C. Hybridizations in Fig. 3 were
PATHOGENICLeishmaniaAND TrypanosomaSPECIES 243

JOBNAME: JEP 82#3 96 PAGE: 3 SESS: 2 OUTPUT: Wed Jun 5 02:28:21 1996/xypage/worksmart/tsp000/69001j/4pu
FIG. 1. Oligonucleotides used for amplification and hybridization of mini-exon repeats. The alignments usedfor the construction of primer sequences are shown. Dots indicate matches with the sequence of one repeat.Degenerate nucleotides in the primers are indicated by (X/X). Lb,L. brasiliensis;Lg, L. guyanensis;Lp, L.panamensis;Ld, L. donovani;Li, L. infantum;Lc, L. chagasi;Laz,L. amazonensis;Lmx, L. mexicana;Lae,L.aethiopica;Ltrop, L. tropica; Lmj, L. major.
244

JOBNAME: JEP 82#3 96 PAGE: 4 SESS: 4 OUTPUT: Wed Jun 5 02:28:21 1996/xypage/worksmart/tsp000/69001j/4pu
performed at 37°C overnight in BLOTTO (Johnsonet al.1984). The filter was washed for 1 hr at 65°C in 0.5×SSC/0.1% SDS.
RESULTS
Amplification of Mini-exon Repeats
Previous work showed that a single pair ofoverlapping oligonucleotides from the con-served exon region of the mini-exon repeat (S-1629 and S-1630 in Fig. 1) could be used toamplify mini-exon repeats from a broad rangeof kinetoplastid protozoa (Fernandeset al.
1994; Murthy et al. 1992). We found in thiswork that the optimal amplification of mini-exon repeats from various pathogenicLeish-maniaspecies required the addition of 10% di-methysulfoxide to the reaction mixture. Thisconcentration of DMSO, however, adversely af-fected the mini-exon amplification from severalTrypanosomaspecies. Therefore, two separatePCR reactions were performed to obtain thepanel of PCR products shown in Fig. 2.The size of the amplified mini-exon repeat
was consistent for each group of species and, in
FIG. 2. Hybridization of PCR-amplified mini-exon repeats from a panel of trypanosomatids with group-specific and species-specific oligonucleotide probes. (A) Ethidium-stained. Lane M,fX174/HaeIII size stan-dard; 1,L. brasiliensisM2903; 2,L. panamensisWR120; 3,L. guyanensisM4147; 4,L. amazonensisPH8; 5,L. mexicanaM379; 6,L. aethiopicaL100; 7,L. major LT252; 8,L. tropicaOD; 9, unknown; 10,L. donovaniMRC-74; 11,L. infantumITMAP263; 12,T. brucei427; 13,T. rangeliEv26; 14,T. cruziSylvio; 15,Trypano-plasma horreliPg-JH. (B) Hybridized with S-1593. (C) Hybridized with S-1595. (D) Hybridized with S-1698.
PATHOGENICLeishmaniaAND TrypanosomaSPECIES 245

JOBNAME: JEP 82#3 96 PAGE: 5 SESS: 2 OUTPUT: Wed Jun 5 02:28:21 1996/xypage/worksmart/tsp000/69001j/4pu
some cases, for each species (Fernandeset al.1994). Three species representing the NewWorld viscerotropicLeishmania(subgenusVi-annia)—L. brasiliensis, L. panamensis,andL.guyanensis—showed a 260-nt monomeric re-peat (expected size from Fernandeset al.(1994), 253–261 bp). Multimers are due to thegenomic tandem arrangement of the repeats.Two representative species of the New WorlddermotropicLeishmania—L. mexicanaand L.amazonensis—generated products of 320 and340 nt, respectively (expected size, 316–332bp). In three Old World dermotropic species (L.aethiopica, L. major,andL. tropica), the prod-ucts were 440–460 nt in size (expected size,436–460 bp), which is similar to the size of theproducts from Old World viscerotropic species(L. donovaniand L. infantum) (expected size,458, 411 bp). An unknown species ofLeish-mania yielded a product of 330 nt, which isclose to that obtained for a species from theL.mexicanagroup. %All three trypanosome spe-cies generated PCR products of the expectedsize: 1350 nt forT. brucei,860 nt forT. rangeli,and 600 nt forT. cruzi (expected sizes fromMurthy et al. (1992) are 1350 forT. brucei(Campbellet al. 1984), 858 forT. rangeli,and582–609 forT. cruzi).
Specificity of the Oligonucleotide Probes
A.Leishmaniaspecies.The probes forLeish-mania were designed on the basis of the se-quence alignments of Fernandeset al. (1994).Since the mini-exon gene introns of all speciesof Leishmaniaare highly conserved, probes forspecific groups were derived from the less con-served intergenic regions. Several gels identicalto the one shown in Fig. 2A were blotted andhybridized with labeled oligonucleotide probes.%The moderately degenerate oligonucleotideS-1593 and the less degenerate S-1698 weredesigned for the New World viscerotropicVi-annia or the Old World viscerotropicL. dono-vani groups, respectively (Fig. 1). As shown inFigs. 2B and 2D, both probes hybridized only totheir cognate groups.The degenerate oligonucleotide S-1595 (Fig.
1) specifically hybridized as predicted to theNew World dermotropicL. mexicanaPCRproduct (Fig. 2C). In addition, this probe hy-bridized to the product from an unknownLeish-mania species and this identification was alsoconsistent with the sizes of the amplified mini-exon repeats (Fernandeset al. 1994).Intergenic regions from species belonging to
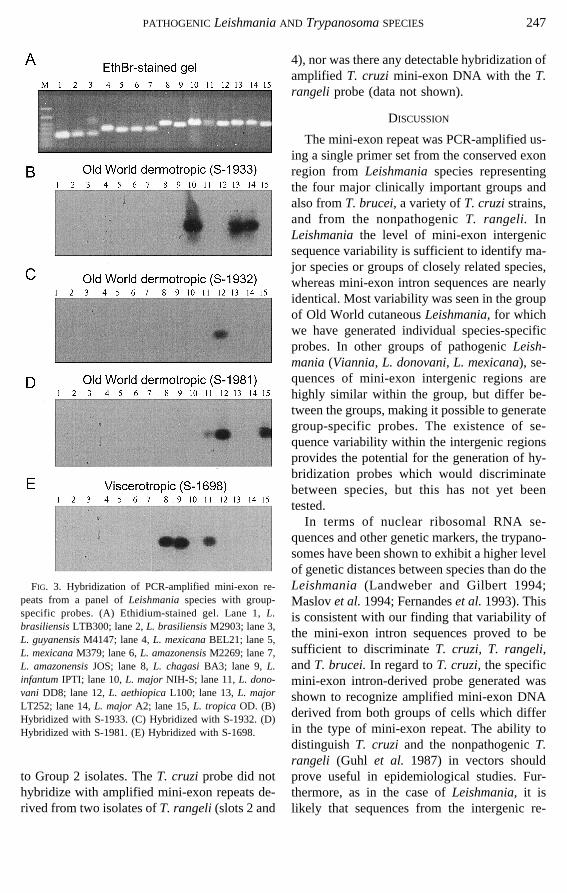
the Old World cutaneousLeishmaniagroup areless conserved (Fernandeset al. 1994), andhence no group-specific probe could be derived.Instead, we attempted to design species-specificprobes for these parasites using variable inter-genic sequences (Fig. 1). As shown in Fig. 3,under stringent hybridization conditions, theoligonucleotide S-1933 produced a specific sig-nal with three different isolates ofL. major(Fig.3B), and the oligonucleotide S-1932 hybridizedonly with L. aethiopica(Fig. 3C). However, theoligonucleotide S-1981, which was designed asa probe forL. tropica,hybridized also to theL.aethiopicaPCR product (Fig. 3D). A weak hy-bridization signal was also seen with one isolateof L. donovani,and this signal was even stron-ger under nonstringent wash conditions (datanot shown). However, differential identificationof L. tropicaandL. donovanican be made witholigonucleotide S-1698 (Fig. 3E).B.Trypanosomaspecies.Oligonucleotides
S-2005, S-1598, and S-1607, which were de-rived from equivalent regions of the respectivemini-exon intron sequences ofT. cruzi, T. ran-geli, andT. brucei(Fig. 1), hybridized only toPCR products from their cognate species (Figs.4A, 4B, 4C).We have found that isolates ofT. cruzi fall
within two major groups with regard to the se-quences of the mini-exon repeats obtained byPCR amplification (D. Campbell and O. Fern-andes, in preparation). Therefore, the specificityof the S-2005 probe was examined with severalT. cruzi isolates. The slot–blot experimentshown in Fig. 4D shows that, under nonstrin-gent hybridization conditions, this probe reactedwith amplified mini-exon repeats derived fromisolates from both major groups ofT. cruzi,al-though to a variable extent. The most inten-sively hybridizing slots (1, 3, and 16) belonged
RAMOS ET AL.246
JOBNAME: JEP 82#3 96 PAGE: 6 SESS: 2 OUTPUT: Wed Jun 5 02:28:21 1996/xypage/worksmart/tsp000/69001j/4pu
to Group 2 isolates. TheT. cruziprobe did nothybridize with amplified mini-exon repeats de-rived from two isolates ofT. rangeli(slots 2 and
4), nor was there any detectable hybridization ofamplified T. cruzimini-exon DNA with theT.rangeli probe (data not shown).
DISCUSSION
The mini-exon repeat was PCR-amplified us-ing a single primer set from the conserved exonregion from Leishmaniaspecies representingthe four major clinically important groups andalso fromT. brucei,a variety ofT. cruzistrains,and from the nonpathogenicT. rangeli. InLeishmaniathe level of mini-exon intergenicsequence variability is sufficient to identify ma-jor species or groups of closely related species,whereas mini-exon intron sequences are nearlyidentical. Most variability was seen in the groupof Old World cutaneousLeishmania,for whichwe have generated individual species-specificprobes. In other groups of pathogenicLeish-mania(Viannia, L. donovani, L. mexicana), se-quences of mini-exon intergenic regions arehighly similar within the group, but differ be-tween the groups, making it possible to generategroup-specific probes. The existence of se-quence variability within the intergenic regionsprovides the potential for the generation of hy-bridization probes which would discriminatebetween species, but this has not yet beentested.In terms of nuclear ribosomal RNA se-
quences and other genetic markers, the trypano-somes have been shown to exhibit a higher levelof genetic distances between species than do theLeishmania (Landweber and Gilbert 1994;Maslovet al.1994; Fernandeset al.1993). Thisis consistent with our finding that variability ofthe mini-exon intron sequences proved to besufficient to discriminateT. cruzi, T. rangeli,andT. brucei.In regard toT. cruzi,the specificmini-exon intron-derived probe generated wasshown to recognize amplified mini-exon DNAderived from both groups of cells which differin the type of mini-exon repeat. The ability todistinguishT. cruzi and the nonpathogenicT.rangeli (Guhl et al. 1987) in vectors shouldprove useful in epidemiological studies. Fur-thermore, as in the case ofLeishmania,it islikely that sequences from the intergenic re-
FIG. 3. Hybridization of PCR-amplified mini-exon re-peats from a panel ofLeishmaniaspecies with group-specific probes. (A) Ethidium-stained gel. Lane 1,L.brasiliensisLTB300; lane 2,L. brasiliensisM2903; lane 3,L. guyanensisM4147; lane 4,L. mexicanaBEL21; lane 5,L. mexicanaM379; lane 6,L. amazonensisM2269; lane 7,L. amazonensisJOS; lane 8,L. chagasiBA3; lane 9,L.infantumIPTI; lane 10,L. majorNIH-S; lane 11,L. dono-vani DD8; lane 12,L. aethiopicaL100; lane 13,L. majorLT252; lane 14,L. major A2; lane 15,L. tropicaOD. (B)Hybridized with S-1933. (C) Hybridized with S-1932. (D)Hybridized with S-1981. (E) Hybridized with S-1698.
PATHOGENICLeishmaniaAND TrypanosomaSPECIES 247

JOBNAME: JEP 82#3 96 PAGE: 7 SESS: 4 OUTPUT: Wed Jun 5 02:28:21 1996/xypage/worksmart/tsp000/69001j/4pu
gions ofT. cruzicould be used to identify indi-vidual isolates or specific groups, and this isbeing tested.These results constitute an initial step toward
the development of a PCR-based detection–identification system useful for clinical and epi-demiological applications. The next step is tolink covalently the capture oligonucleotidesspecific for each trypanosomatid group or spe-cies to microtiter wells and to employ nonra-dioactive methods for the detection of hybrid-ized PCR-amplified mini-exon DNA (Murthyetal. 1992; Galindoet al. 1993). This work is inprogress.
ACKNOWLEDGMENTS
We thank D. McMahon-Pratt for supplying several
strains of trypanosomatids used in this study. We also thankR. Lainson, G. Grimaldi, S. Beverley, J. Dvorak, N. Saravia,J. Coura, R. McMaster, and K. Stuart for contributingstrains used in this work. Grant support was provided byUNDP/World Bank/WHO Special Programme for Researchand Training in Tropical Diseases (L.S.). D.A.C. is a Bur-roughs Wellcome New Investigator in Molecular Parasitol-ogy.
REFERENCES
AGABIAN, N. 1990.Transsplicing of nuclear pre-mRNAs.Cell 61, 1157–1160.
AVILA , H. A., PEREIRA, J. B., THIEMANN, O., DE PAIVA , E.,DEGRAVE, W., MOREL, C. M., AND SIMPSON, L. 1993. De-tection of Trypanosoma cruziin blood specimens ofchronic chagasic patients by polymerase chain reactionamplification of kinetoplast minicircle DNA: Compari-son with serology and xenodiagnosis.Journal of ClinicalMicrobiology31, 2421–2426.
FIG. 4. Hybridization of PCR-amplified mini-exon repeats from a panel of trypanosomatids with species-specific oligonucleotide probes forTrypanosomaspecies. The gels in A–C are identical to that in Fig. 2. Seelegend to Fig. 2 for identification of lanes. (A) Hybridized with S-1607.(B) Hybridized with S-1598. (C)Hybridized with S-2005; an identical result was obtained using probe S-1597. (D) Slot-blot of amplifiedmini-exon DNA from strains ofT. cruzibelonging to the two major groups with different mini-exon sequencesand from two strains ofT. rangelihybridized with S-2005. Slots 2, 4,T. rangeli;slots 3, 16, 26,T. cruziGroup2; slots 1, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 17, 19, 20, 21, 22, 23, 24, 25,T. cruziGroup 1; slot 27, negativehybridization control.
RAMOS ET AL.248

JOBNAME: JEP 82#3 96 PAGE: 8 SESS: 2 OUTPUT: Wed Jun 5 02:28:21 1996/xypage/worksmart/tsp000/69001j/4pu
BERENS, R. L., BRUN, R., AND KRASSNER, S. M. 1976. Asimple monophasic medium for axenic culture of hemo-flagellates.Journal of Parasitology62, 360–365.
BRITTO, C., AND CARDOSO, M. A., MONTEIROVANNI, C. M.,HASSLOCHER-MORENO, A., XAVIER, S. S., OELEMANN, W.,SANTORO, A., PIRMEZ, C., MOREL, C. M., AND WINCKER,P. 1995. Polymerase chain reaction detection ofTrypano-soma cruziin human blood samples as a tool for diagno-sis and treatment evaluation.Parasitology110,241–247.
BRUN, R., AND SCHOENENBERGER, M. 1979. Cultivation andin vitro cloning of procyclic culture forms ofTrypano-soma bruceiin semi-defined medium.Acta Tropica36,289–292.
CAMPBELL, D., THORNTON, D., AND BOOTHROYD, J. 1984.Apparent discontinuous transcription ofTrypanosomabrucei variant surface antigen genes.Nature 311, 350–355.
CASTELLANI, O., RIBEIRO, L. V., AND FERNANDES, J. F. 1967.Differentiation ofTrypanosoma cruziin culture.Journalof Protozoology14, 447–451.
DE LANGE, T., LIU, A. Y., VAN DERPLOEG, L., BORST, P.,TROMP, M. C., AND VAN BOOM, J. H. 1983. Tandem rep-etition of the 59 mini-exon of variant surface glycoproteingenes: A multiple promoter for VSG transcription.Cell34, 891–900.
DE LANGE, T., MICHELS, P., VEERMAN, H., CORNELISSEN, A.,AND BORST, P. 1986. Many trypanosome messengerRNAs share a common 59 terminal sequence.NuclearAcids Research12, 3777–3790.
DVORAK, J. A., HARTMAN, D. L., AND MILES, M. A. 1980.Trypanosoma cruzi:Correlation of growth kinetics to zy-modeme type in clones derived from various sources.Journal Protozoology27, 472–474.
DVORAK, J. A., HALL , T. E., CRANE, M. S., ENGEL, J. C.,MCDANIEL, J. P.,AND URIEGAS, R. 1982.Trypanosomacruzi: flow cytometric analysis. I. Analysis of total DNA/organism by means of mithramycin-induced fluores-cence.Journal of Protozoology29, 430–437.
FERNANDES, A. P., NELSON, K., AND BEVERLEY, S. M. 1993.Evolution of nuclear ribosomal RNAs in kinetoplastidprotozoa: Perspectives on the age and origins of parasit-ism. Proceedings of the National Academy of SciencesUSA.90, 11608–11612.
FERNANDES, O., MURTHY, V. K., KURATH, U., DEGRAVE,W. M., AND CAMPBELL, D. A. 1994. Mini-exon genevariation in human pathogenicLeishmaniaspecies.Mol-ecules and Biochemistry Parasitology66, 261–271.
GALINDO, I., RANGEL-ALDAO, R., AND RAMIREZ, J. 1993. Acombined polymerase chain reaction colour developmenthybridization assay in a microtitre format for the detec-tion of Clostridium spp. Applied Microbiology Biotech-nology39, 553–557.
GONZALEZ, A., PREDIGER, E., HUECAS, M., NOGUEIRA, N.,AND LIZARDI, P. 1984. Minichromosomal repetitive DNAin Trypanosoma cruzi:Its use in a high-sensitivity para-site detection assay.Proceedings of the National Acad-emy of Sciences USA11, 3356–3360.
GUHL, F., GOLD, J. W. M., MARINKELLE, C. J., JARAMILLO ,C. A., AND BRIDGE, D. 1987. ClinicalTrypanosoma ran-geli infections as a complication of Chagas’ disease.Parasitology94, 475–484.
HASSAN, M. Q., GHOSH, A., GHOSH, S. S., GUPTA, M., BASU,D., MALLIK , K. K., AND ADHYA, S. 1993. Enzymatic am-plification of mini-exon-derived RNA gene spacers ofLeishmania donovani:Primers and probes for DNA di-agnosis.Parasitology107,509–517.
HOLGUIN, A. F., SARAVIA , N. G., AND D’A LESSANDRO, A.1987. Lack of enzyme polymorphism inTrypanosomarangelistocks from sylvatic and domiciliary transmissioncycles in Colombia.American Journal of Tropical Medi-cine and Hygiene36, 53–58.
HOWARD, M. K., KELLY, J. M., LANE, R. P., AND MILES,M. A. 1991. A sensitive repetitive DNA probe that isspecific to theLeishmania donovanicomplex and its useas an epidemiological and diagnostic reagent.MolecularBiochemistry Parasitology44, 63–72.
JOHNSON, P. A., GAUTSCH, J. W., SPORTMAN, J. R.,AND EL-DER, J. H. 1984. Improved technique utilizing nonfat drymilk for analysis of proteins and nucleic acids transferredto nitrocellulose.Genetic Analysis and Technique1, 3–8.
LANDWEBER, L. F., AND GILBERT, W. 1994. Phylogeneticanalysis of RNA editing: A primitive genetic phenom-enon.Proceedings of the National Academy of SciencesUSA91, 918–921.
LASKAY, T., KIESSLING, R., RINKE DEWIT, T. F.,AND WIRTH,D. F. 1991. Generation of species-specific DNA probesfor Leishmania aethiopica. Molecular Biochemistry andParasitology44, 279–286.
LOPES, U. G., MOMEN, H., GRIMALDI , G. J., MARZOCHI,M. C., PACHECO, R. S.,AND MOREL, C. M. 1984. Schiz-odeme and zymodeme characterization ofLeishmaniainthe investigation of foci of visceral and cutaneous leish-maniasis.Journals of Parasitology70, 89–98.
LOPES, U. G.,AND WIRTH, D. F. 1986. Identification of vis-ceral Leishmaniaspecies with cloned sequences of ki-netoplast DNA.Molecular Biochemistry and Parasitol-ogy20, 77–84.
MASLOV, D. A., AVILA , H. A., LAKE, J. A.,AND SIMPSON, L.1994. Evolution of RNA editing in kinetoplastid proto-zoa.Nature365,345–348.
MURPHY, W., WATKINS, K., AND AGABIAN, N. 1986. Iden-tification of a novel Y branch structure as an intermediatein trypanosome mRNA processing: Evidence for transsplicing.Cell 47, 517–525.
MURTHY, V. K., DIBBERN, K. M., AND CAMPBELL, D. A.1992. PCR amplification of mini-exon genes differenti-atesTrypanosoma cruzifrom Trypanosoma rangeli. MolCell Probes6, 237–243.
NILSEN, T. W. 1993.Trans-splicing of nematode premes-senger RNA.Annual Review of Microbiology47, 413–440.
RODGERS, M. R., POPPER, S. J.,AND WIRTH, D. F. 1990. Am-plification of kinetoplast DNA as a tool in the detection
PATHOGENICLeishmaniaAND TrypanosomaSPECIES 249

JOBNAME: JEP 82#3 96 PAGE: 9 SESS: 2 OUTPUT: Wed Jun 5 02:28:21 1996/xypage/worksmart/tsp000/69001j/4pu
and diagnosis ofLeishmania. Experimental Parasitology71, 267–275.
STURM, N. R., DEGRAVE, W., MOREL, C., AND SIMPSON, L.1989. Sensitive detection and schizodeme classificationof Trypanosoma cruzicells by amplification of kineto-plast minicircle DNA sequences: use in diagnosis of Cha-gas disease.Molecular Biochemistry Parasitology33,205–214.
SUTTON, R., AND BOOTHROYD, J. 1986. Evidence for transsplicing in trypanosomes.Cell 47, 527–535.
ULIANA , S. R., NELSON, K., BEVERLEY, S. M., CAMARGO,E. P.,AND FLOETER-WINTER, L. M. 1994. DiscriminationamongstLeishmaniaby polymerase chain reaction andhybridization with small subunit ribosomal DNA derivedoligonucleotides.Journal of Eukaryotic Microbiology41,324–330.
WINCKER, P., BRITTO, C., PEREIRA, J. B., CARDOSO, M. A.,
OELEMANN, W., AND MOREL, C. M. 1994. Use of a sim-plified polymerase chain reaction procedure to detectTrypanosoma cruziin blood samples from chronic cha-gasic patients in a rural endemic area.American Journalof Tropical Medicine and Hygiene51, 771–777.
WIRTH, D., ROGERS, W., BARKER, R., DOURADO, H., SUE-SEBANG, S., AND ALBUQUERQUE, B. 1986. Leishmaniasisand malaria: New tools for epidemiological analysis.Sci-ence234,975–979.
WIRTH, D. F., AND PRATT, D. M. 1982. Rapid identificationof Leishmaniaspecies by specific hybridization of ki-netoplast DNA in cutaneous lesions.Proceedings of theNational Academy of Sciences USA79, 6999–7003.
Received 31 August 1995; accepted with revision 16 No-vember 1995
RAMOS ET AL.250