cxcr2-dependent mucosal neutrophil influx protects · pdf filecxcr2-dependent mucosal...
TRANSCRIPT

of May 21, 2018.This information is current as
Lesion-Forming Bacterial PathogenDiarrhea Caused by an Attaching/EffacingInflux Protects against Colitis-Associated CXCR2-Dependent Mucosal Neutrophil
Hanson, Declan F. McCole and Lars EckmannMartina E. Spehlmann, Sara M. Dann, Petr Hruz, Elaine
http://www.jimmunol.org/content/183/5/3332doi: 10.4049/jimmunol.0900600August 2009;
2009; 183:3332-3343; Prepublished online 12J Immunol
MaterialSupplementary
0.DC1http://www.jimmunol.org/content/suppl/2009/08/12/jimmunol.090060
Referenceshttp://www.jimmunol.org/content/183/5/3332.full#ref-list-1
, 27 of which you can access for free at: cites 59 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

CXCR2-Dependent Mucosal Neutrophil Influx Protects againstColitis-Associated Diarrhea Caused by an Attaching/EffacingLesion-Forming Bacterial Pathogen1
Martina E. Spehlmann, Sara M. Dann, Petr Hruz, Elaine Hanson, Declan F. McCole,and Lars Eckmann2
Enteropathogenic Escherichia coli (EPEC) is a major cause of diarrheal disease in young children, yet symptoms and duration arehighly variable for unknown reasons. Citrobacter rodentium, a murine model pathogen that shares important functional featureswith EPEC, colonizes mice in colon and cecum and causes inflammation, but typically little or no diarrhea. We conductedgenome-wide microarray studies to define mechanisms of host defense and disease in C. rodentium infection. A significant fractionof the genes most highly induced in the colon by infection encoded CXC chemokines, particularly CXCL1/2/5 and CXCL9/10,which are ligands for the chemokine receptors CXCR2 and CXCR3, respectively. CD11b� dendritic cells were the major pro-ducers of CXCL1, CXCL5, and CXCL9, while CXCL2 was mainly induced in macrophages. Infection of gene-targeted micerevealed that CXCR3 had a significant but modest role in defense against C. rodentium, whereas CXCR2 had a major andindispensable function. CXCR2 was required for normal mucosal influx of neutrophils, which act as direct antibacterial effectors.Moreover, CXCR2 loss led to severe diarrhea and failure to express critical components of normal ion and fluid transport,including ATPase �2-subunit, CFTR, and DRA. The antidiarrheal functions were unique to CXCR2, since other immune defectsleading to increased bacterial load and inflammation did not cause diarrhea. Thus, CXCR2-dependent processes, particularlymucosal neutrophil influx, not only contribute to host defense against C. rodentium, but provide protection against infection-associated diarrhea. The Journal of Immunology, 2009, 183: 3332–3343.
E nteropathogenic Escherichia coli (EPEC)3 is an importantcause of diarrheal disease and mortality worldwide, espe-cially among infants and young children in the developing
world (1). EPEC is distinguished from other diarrheagenic E. coliby its ability to form attaching/effacing (A/E) lesions characterizedby intimate attachment to the epithelium and localized microvillusloss, combined with a lack of classical enterotoxins. The infectionis accompanied by watery diarrhea, fever, vomiting, and abdomi-nal pain, and it can lead to significant mortality. The mechanismsgoverning infection-associated diarrhea are not well understood,but they appear to be multifactorial (2). A decrease in the absorp-tive surface epithelium, disruption of tight junctions and epithelialbarrier function, and release of secretagogues by inflammatorycells have been proposed to mediate diarrhea, although the relativeimportance of these and possibly other mechanisms remains un-known (2–4).
Progress in elucidating the pathogenesis of EPEC infection hasbeen hampered by the dearth of suitable animal models. Instead,model pathogens with characteristics similar to EPEC have beenemployed widely, including rabbit EPEC and Citrobacter roden-tium in mice. C. rodentium causes A/E lesions and expresses keyvirulence genes, including intimin, espA, espB, and tir, that arefunctionally exchangeable with the corresponding genes in EPEC.Infection of adult mice with C. rodentium leads to self-limitingepithelial hyperplasia and mucosal inflammation (5–8). Effectivehost defense against the pathogen requires CD4� T cells, B cells,mast cells, and neutrophils (9–11). IgG Abs contribute to bacterialclearance, whereas secretory IgA or IgM plays a limited role inhost defense (12, 13). Several immunoregulatory cytokines, in-cluding IFN-�, TNF-�, IL-6, IL-12, IL-17, and IL-22, control in-nate or adaptive immune responses against the bacteria, since micelacking the respective cytokines have a reduced ability to clearinfection (14–19). However, the histologically apparent complex-ity of the host response that accompanies infection (5, 7) suggeststhat numerous other gene products are likely to be involved inorchestrating mucosal defense against the bacteria.
Expression analysis of large numbers of genes by microarrayshas reached a technical maturity that now allows routine applica-tion to many biological systems. To date, such expression studieshave mostly been conducted in cell culture models, as they allowtight control over experimental conditions. However, a number ofin vivo applications have demonstrated the utility of microarrays indefining previously elusive biological processes in complex phys-iological settings (20–22). Based on such prior successes, we setout to examine the global gene expression profile associated withhost defense against the model pathogen, C. rodentium. Our ex-pression studies and the mechanistic pursuit of the findings unex-pectedly revealed a novel role of CXC chemokines and neutrophils
Department of Medicine, University of California, San Diego, La Jolla, CA 92093
Received for publication February 23, 2009. Accepted for publication July 4, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants DK70867,DK35108, AI56075, and RR17030, and the University of California, San Diego Di-gestive Diseases Research Development Center (DK80506). M.E.S. was supported bya fellowship from the MFG Educative Science, S.M.D. by a fellowship from theCrohn’s and Colitis Foundation of America, and P.H. by a fellowship from the SwissNational Science Foundation (SSMBS; PASMA 114623).2 Address correspondence and reprint requests to Dr. Lars Eckmann, Department ofMedicine, University of California, San Diego, 9500 Gilman Drive, La Jolla, CA92093. E-mail address: [email protected] Abbreviations used in this paper: EPEC, enteropathogenic Escherichia coli; A/E,attaching-and-effacing; Isc, short-circuit current; MPO, myeloperoxidase.
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0900600
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

in controlling not only bacterial load but also the diarrheal re-sponse to infection, thus providing new insights into the impor-tance of host components in determining diarrhea after infectionwith an A/E lesion-forming enteric pathogen.
Materials and MethodsMice and infection protocol
All wild-type and gene-targeted mice of either gender were obtained fromThe Jackson Laboratory and were at least 8 wk old for experiments. Forinfections, C. rodentium (a gift from Dr. B. Vallance, British ColumbiaChildren’s Hospital, Vancouver, Canada) was grown overnight in Luria-Bertani broth at 37°C, harvested by centrifugation, and resuspended infresh broth at 2.5 � 109/ml. Mice were infected by oral gavage with 200�l (5 � 108 bacteria) of the suspension. To determine bacterial numbers inthe stool, fecal pellets were collected from individual mice, weighed, andhomogenized in 5 ml of PBS. For enumerating bacteria in spleen and liver,each organ was homogenized in 2 ml of PBS. Serial dilutions of the ho-mogenates were plated onto MacConkey agar, and the number of CFU wasdetermined after overnight incubation at 37°C. The detection limit of theCFU assay was 103 CFU/g feces and �101 CFU per liver or spleen. Theidentity of representative colonies was verified by PCR analysis of the C.rodentium espB gene as described (12). All animal studies were reviewedand approved by the University of California, San Diego Institutional An-imal Care and Use Committee.
Histological analysis
Organs were removed and fixed in 10% phosphate-buffered formalin(Fisher Scientific) for 24 h. Colons were opened longitudinally,cleaned, and processed as “Swiss rolls” before fixation. Fixed tissueswere embedded in paraffin, and 5-�m sections were prepared andstained with H&E. For immunohistological studies, tissues were frozenin OCT compound (Sakura Finetek), cut on a cryomicrotome, air-dried,fixed in acetone, and incubated with 0.3% H2O2 for 20 min at roomtemperature to inactivate endogenous peroxidase. After blocking withPBS containing 2% rabbit serum (Abcam), sections were incubatedovernight at 4°C in the same buffer containing 1 �g/ml rabbit anti-mouse myeloperoxidase (MPO; Abcam), a 1/1000 dilution of rabbitanti-C. rodentium (from B. Vallance) (12), or 1 �g/ml normal rabbitIgG (Jackson ImmunoResearch Laboratories) as a control. Sectionswere washed, stained for 30 min at room temperature with 2 �g/mlHRP-conjugated goat anti-rabbit IgG (Vector Laboratories, Burlin-game, CA), developed with 3,3�-diaminobenzidine/H2O2 (Vector Lab-oratories), and counterstained with Gill’s hematoxylin (Fisher Scien-tific). Alternatively, sections were stained with 3 �g/ml Cy3-conjugatedgoat anti-rabbit IgG (R&D Systems) and mounted in media containing4�,6�-diamidino-2-phenylindole (Vector Laboratories).
Microarray and real-time PCR analysis
RNA was isolated from whole colon tissue using TRIzol reagent (Invitro-gen). RNA quality was assessed with an Agilent Technologies bioanalyzer.Total RNA was processed and hybridized to CodeLink Mouse Whole Ge-nome Bioarrays (carrying 34,790 unique probes; GE Healthcare). Biotin-labeled first-strand cDNA from 2.5 to 5 �g of total RNA was used tohybridize arrays in 6� standard saline citrate phosphate/EDTA/30% for-mamide at 25°C for 18 h. Hybridized slides were washed and stained withstreptavidin-Alexa 647 conjugate (Invitrogen/Molecular Probes). The pro-cessed slides were scanned at 10 �m resolution with an Axon 4000B Scan-ner (Molecular Devices). Positive and negative bacterial control geneswere spotted on the CodeLink arrays, and specific bacterial mRNA wasused to spike the cDNA synthesis reaction. Background correction wasperformed by the CodeLink software. For each array, RNA was pooledfrom four to six mice. Filtering was conducted by deleting probes that hadlow or ambiguous values. Numbers were normalized by dividing raw in-tensity values by medians for each chip. The complete microarray data setsare available on the National Center for Biotechnology Information GeneExpression Omnibus (www.ncbi.nlm.nih.gov/geo) under accession no.GSE16847.
For PCR analysis, RNA was treated with Turbo DNase-free (Ambion)to remove contaminating DNA. Reverse transcription was performed usingthe High Capacity cDNA Reverse Transcription kit (Applied Biosystems)and real-time PCR amplification using the Mesa Green 2� SYBR mix(Eurogentec). Amplification of the expected single products was confirmedon 1% agarose gels stained with ethidium bromide. Primers and PCR prod-
uct sizes are listed in supplemental Table I.4 Relative changes in targetmRNA levels were calculated by the 2��Ct method, with GAPDH as thereference standard.
Isolation of epithelial and lamina propria cells
The colon was opened, incubated for 10 min at room temperature in PBSwith 5% FCS and 1 mM DTT to remove mucus, and cut into 5-mm pieces.Tissue pieces were incubated in HBSS (without Ca and Mg) with 5 mMEDTA, 5% FCS, 10 mM HEPES (pH 7.3), and 1 mM DTT for 2 � 20 minat 37°C with constant shaking. Detached epithelial cells were passedthrough a nylon mesh strainer (100-�m pore size; Fisher Scientific) andcollected by centrifugation. The remaining tissue pieces were washed andincubated twice at 37°C for 30 min in RPMI 1640 medium containing 1mg/ml collagenase D (Roche Applied Science) and 100 �g/ml DNase I(Worthington Biochemical). After each incubation, mixtures were vortexedfor 30 s, and suspensions were passed through a 40-�m cell strainer. Cellsfrom both digestions were combined and collected by centrifugation.
Flow cytometry
Cell suspensions were stained with optimal concentrations of fluoro-chrome-conjugated Abs against mouse CD11b, CD11c, CD45, Gr-1 (allfrom eBioscience), and/or CXCR2 (R&D Systems) for 30 min on ice. Cellswere washed and fixed with 2% paraformaldehyde in PBS before analysison a BD Biosciences FACSCalibur flow cytometer. Cell sorting of stainedbut unfixed cells was performed on a MoFlo high-speed cell sorter (Beck-man Coulter). Macrophages were identified as CD45�CD11b�CD11c�
cells. Dendritic cells were identified as CD45�CD11c� cells and furthersubdivided into CD11b� and CD11b� subsets. All other leukocytes wereidentified as CD45�CD11b�CD11c� cells.
Immunoblotting
Tissues were lysed for 30 min on ice in a buffer containing 150 mM NaCl,5 mM KCl, 10 mM HEPES, 0.5 mM EDTA, 0.2 mM EGTA, 1 mM sodiumfluoride, 1 mM vanadate, 0.05% Nonidet P-40, 1 mM DTT, and a proteaseinhibitor cocktail (Roche Applied Science). After centrifugation to removedebris, equal protein amounts (20 �g) were boiled for 10 min in loadingbuffer (50 mM Tris (pH 6.8), 100 mM DTT, 2% SDS, 40% glycerol, and0.2% bromophenol blue) and size-separated by electrophoresis on a 10%Tris-glycine polyacrylamide gel (Bio-Rad Laboratories). Proteins wereelectrotransferred to a polyvinylidene difluoride membrane, which wasblocked and stained overnight at 4°C with rabbit anti-mouse DRA/SLC26A3 (Santa Cruz Biotechnology) in a buffer of 10 mM Tris (pH 7.6),5% BSA, and 0.05% Tween 20, followed by incubation with HRP-conju-gated goat anti-rabbit IgG and detection by ECL (GE Healthcare).
Neutrophil killing assay
A suspension of bone marrow cells, obtained from the femurs of C57BL/6mice, was layered on top of two Histopaque layers (densities of 1.083 and1.119 g/ml) (Sigma-Aldrich) and centrifuged at 1200 rpm for 30 min. Cellsat the interface between the two Histopaque layers were collected andwashed in HBSS. Neutrophil purity was �90% as determined by Giemsastaining. Cells (5 � 107/ml) were pretreated for 1 h with mouse TNF-�and/or IFN-� (50 ng/ml each) (both from PeproTech), and C. rodentiumwas added at a multiplicity of infection of 1:1 in 100 �l of RPMI 1640medium. After incubation at 37°C for up to 6 h with light agitation, cultureswere diluted 1/10 in water to lyse the neutrophils, and serial dilutions wereplated onto MacConkey agar.
Analysis of colonic cytokine production
Colon pieces (1 � 1 mm) were placed into 12-well plates in 500 �l ofRPMI 1640 medium containing 50 �g/ml each streptomycin and penicillin(both from Sigma-Aldrich) and incubated for 6 h at 37°C. Supernatantswere centrifuged and frozen at �80°C. In some experiments, colon tissueswere homogenized in PBS with 0.1% Tween 20 and a cocktail of proteaseand phosphatase inbibitors (Roche Applied Science), centrifuged to re-move debris, and supernatants were frozen at �80°C. Cytokine levels weredetermined by ELISA (R&D Systems) and were normalized to tissueweight.
ELISA
Blood was collected by tail vein bleeding, diluted 1/10 into PBS containing2 mM EDTA, centrifuged, and stored at �80°C until use. C. rodentium,
4 The online version of this article contains supplemental material.
3333The Journal of Immunology
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

grown overnight in Luria-Bertani broth, were washed extensively in PBS,and 108 bacteria were added in 50 �l per well of a 96-well polystyreneplate. After overnight air drying, bacteria were fixed for 5 min at roomtemperature with 0.15% glutaraldehyde in 0.15 M phosphate buffer (pH7.0), followed by blocking with 0.15 M glycine in 15 mM phosphate buffer(pH 7.0), and plates were incubated overnight at 4°C with PBS containing5% nonfat dry milk and 0.5% Tween 20. After incubation with serumsamples for 2 h, HRP-conjugated goat Abs against mouse IgG or IgM(SouthernBiotech) were applied to the plates for 1 h at room temperature.Bound peroxidase was visualized with tetramethylbenzidine-H2O2 in ace-tate buffer, and reactions were stopped with sulfuric acid and read at450 nm.
MPO analysis
Colon pieces were snap-frozen in liquid nitrogen and stored at �80°C.Tissues were homogenized in hexadecyltrimethylammonium bromidebuffer (Sigma-Aldrich), cleared by centrifugation, and the supernatant wasadded to a solution containing o-dianisidine (Sigma-Aldrich) and H2O2.Absorbance of the colorimetric reaction was measured at 600 nm. PurifiedMPO of known activity was used as a standard. MPO activity is expressedin units per gram of wet tissue.
Fecal water and ion transport studies
For fecal water analysis, stool was collected for 1 h from individualmice, weighed, dried for 24 h at 95°C, and weighed again. Percentageof fecal water was calculated by dividing the difference of dry and totalweight by the total weight. For electrophysiological studies, a 3-cmsegment of mid-distal colon was stripped of the muscle layer and cutinto smaller sections that were then mounted on Ussing chamber inserts(Warner Instruments) with a window area of 0.07 cm2. Tissues werebathed in a modified oxygenated Ringer’s solution (140 mM Na�, 5.2mM K�, 1.2 mM Ca�, 0.8 mM Mg�, 120 mM Cl�, 25 mM HCO3
�, 2.4mM H2PO4
�, 0.4 mM HPO42�, and 10 mM glucose) at 37°C. The
tissues were short-circuited by an automated voltage clamp, and thecurrent (Isc) required to obtain zero potential difference across the tis-sues was monitored as an indication of net active ion transport. Todetermine transepithelial resistance, a defined voltage peak was tran-siently applied across the tissue, the resulting current was measured,and resistance was calculated by Ohm’s law.
Statistical analysis
Data are expressed as means � SEM. Differences were analyzed by t test,with p values of �0.05 considered significant. CFU counts were log10
transformed, and means and SEs of the mean were calculated from the logvalues. Samples without detectable C. rodentium colonies were assigned alog10 value equivalent to half of the detection limit of the CFU assay.Differences between CFU counts were evaluated by Mann-Whitney rank-sum test, with p values of �0.05 considered statistically significant. Resultsfrom males and females were combined, as no significant differences weredetected between the genders in bacterial colonization or mucosal re-sponses after infection.
ResultsGlobal gene expression analysis of the colonic response toC. rodentium reveals prominent induction of chemokines
To broaden the understanding of the pathogenesis of entericinfections with A/E lesion-forming bacteria, we used a murineinfection model with the prototypic pathogen, C. rodentium (8),and conducted a microarray analysis as an unbiased approach toobtain a genome-wide gene expression profile of the colonicresponse to infection. Intragastric administration of C. roden-tium to normal C57BL/6 mice leads to peak infection levels by1 wk, and bacterial clearance after 2–3 wk (12) (see Figs. 3Aand 4A below). Accordingly, we extracted total RNA from thecolon of infected mice at different times after infection (1–3 wk)and related expression levels to a parallel cohort of uninfectedmice. The microarray studies revealed a wide spectrum of ex-pression changes, whose exact characteristics were dependenton the selection criteria applied for determining significant andmeaningful changes. To select suitable criteria, we first evalu-ated the reliability of the microarray data by assessing mRNAlevels at different times after infection for 20 representative
genes with real-time PCR as an independent, sensitive, andhighly specific approach. Comparison of microarray and real-time PCR data showed a strong positive correlation, withgreater changes in expression showing better reproducibilitythan lesser changes (Fig. 1A). Based on this observation, wesubsequently focused on genes whose expression levels were�4-fold altered at any point after infection.
The microarrays employed in our studies carried 34,790 probescovering the entire murine genome. Of these, 577 probes (1.7%),representing the same number of unique genes, revealed a �4-foldincrease (326 genes) or decrease (251 genes) in expression at anytime after C. rodentium infection (Fig. 1B, supplemental Table II,and data not shown). Most changes were observed at weeks 1 and2, while relatively fewer changes were seen after 3 wk. Increasesdominated over decreases by ratios ranging from 2.2:1 after 1 wkto 1.5:1 after 3 wk. Furthermore, increased expression of most ofthe genes was confined to a limited period after infection, withonly few genes showing significant increases throughout the 3-wkperiod and practically no genes exhibiting a biphasic expressionpattern (Fig. 1B and supplemental Table II). Taken together, theseresults suggest that the initial host response to infection is charac-terized by induction of an array of “early response” genes, whilethe later clearance and healing phase is dominated by induction ofa different set of genes.
Because we sought to elucidate the events important in the ini-tiation of host defense against the bacteria, we focused on thosegenes for which the microarrays had revealed the earliest andgreatest relative increase in expression (Table I and supplementalTable II). Of the top 20 genes in this category, 4 were CXC che-mokines. Most of the other highly induced genes encode productswith diverse functions, ranging from acute phase reactants
FIGURE 1. Microarray analysis of colonic gene expression after C. ro-dentium infection. Wild-type (C57BL/6J) mice were infected orally with5 � 108 C. rodentium, or left uninfected. Total colon RNA was extractedafter 1, 2, and 3 wk and subjected to microarray analysis. In parallel,mRNA levels of representative genes were assayed by real-time PCR.mRNA levels are expressed as ratio of the levels in infected over unin-fected mice. A, Correlation of microarray data and real-time PCR results.Each datum point represents one gene at one time point. B, Venn diagramof all genes with a �4-fold increase in expression at any time afterinfection.
3334 CXCR2 AND CXCR3 IN PATHOGENESIS OF C. rodentium INFECTION
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

(leucine-rich �2-glycoprotein 1 and serum amyloid A3) and calci-um-binding proteins with diverse intracellular and extracellular func-tions (S100A8 and S100A9) (23) to ion pumps (Na�/K� ATPase,�-subunit) and lectins with antimicrobial activity (Reg3�, Reg3�, andReg3�) (24).
Since chemokines were highly overrepresented among themost highly induced genes (i.e., they made up 20% of the top20 genes, but only represent 0.15% of all predicted murinegenes) and since they are critical chemoattractants and activa-tors of leukocyte subsets, we focused our further analysis on theentire group of chemokines represented on the microarrays. Adistinct chemokine expression pattern was observed, with thehighest early induction (�10-fold on week 1) observed forCXCL1, CXCL2, CXCL5, and CXCL9 (Table I), and modestearly induction for CXCL3 (5.7-fold), CXCL10 (7.6-fold), andCXCL11 (4.8-fold). Limited or no early induction (�4-fold)was observed for the other CXC chemokines or for most CCchemokines, with the partial exception of CCL7 (6.2-fold) andCCL19 (4.4-fold). These data show that rapid induction of adistinct subset of CXC chemokines is a prominent feature of thecolonic host response to C. rodentium.
Increased colonic CXCL expression after C. rodentium infection
To further investigate chemokine functions in host defense againstC. rodentium, we focused on the two most highly induced groupsof CXC chemokines, CXCL1/2/5 and CXCL9/10/11. These che-mokines are cognate ligands for only two CXC chemokine recep-tors, CXCR2 and CXCR3, respectively, suggesting that the ligandinteractions with these two receptors may be important in antibac-terial host defense. Time course studies revealed a rapid inductionof at least five of the six chemokines in the colon within 3 daysafter infection and maximal expression by 7–14 days (Fig. 2A).Marked, albeit lower, increases were also observed in jejunum,mesenteric lymph nodes, liver, and spleen, indicating that the che-mokine responses were not limited to the site of infection (Fig.2B). Furthermore, protein production followed mRNA changes, asdemonstrated by ELISA in supernatants of short-term colon ex-plant cultures for CXCL5 and CXCL9 as representatives of eachchemokine group (Fig. 2C).
We next examined the cellular source of the chemokines afterinfection. Epithelial cells and subsets of lamina propria cells wereisolated and analyzed by real-time PCR for chemokine expression.CXCL1, CXCL5, and CXCL9 were most highly induced inCD11b� dendritic cells, while CXCL2 was mainly induced inmacrophages (Fig. 2D). CXCL5 was also modestly induced in ep-ithelial cells and macrophages, and CXCL9 in epithelial cells. Incontrast, CD11b� dendritic cells or any other CD45� leukocyteswere not a significant source of the four chemokines after infection(Fig. 2D). Thus, CD11b� dendritic cells and macrophages werethe major chemokine responders to C. rodentium.
CXCR3 in host defense against C. rodentium
As a next step toward elucidating the role of specific CXCchemokines in defense against C. rodentium, we determined theimpact of the loss of ligand/receptor interactions on the patho-genesis of infection. For this, we employed mice with targeteddeletions of the CXC chemokine receptors, CXCR2 andCXCR3, as several of their respective cognate ligands wereinduced after infection. Mice lacking CXCR3 are fertile andhealthy under standard husbandry conditions (25). Upon oral C.rodentium challenge, CXCR3-deficient mice had significantlyincreased fecal CFU at 14 and 17 days after infection (Fig. 3A),times that represent the middle to late bacterial clearance phase.No significant differences in bacterial load were observed at 7days or 21 days (Fig. 3A), indicating that CXCR3 was dispens-able in the early innate control and ultimate eradication of thebacteria. Furthermore, CXCR3-deficient mice had significantlyhigher bacterial numbers in liver and spleen at 2 wk, but sur-prisingly also after 1 wk, at which time fecal CFU did not differbetween knockout and wild-type mice (Fig. 3A).
Histological analysis of the colon revealed that CXCR3-defi-cient mice had modestly increased submucosal and mucosal infil-tration with inflammatory cells, and greater epithelial ulcerationafter 2 wk (Fig. 3B), which was accompanied by elevated levels ofthe neutrophil marker MPO in colon homogenates (Fig. 3C).Moreover, the liver showed disseminated foci of necrotic tissue inCXCR3-deficient but not wild-type mice after infection (Fig. 3D).These and the CFU data suggest that CXCR3-dependent processes
Table I. Induction of colonic gene expression after C. rodentium infectiona
No.Fold
Change Accession No. Gene Protein
1 183 NM_009114 S100a9 S100 calcium-binding protein A9 (calgranulin B)2 136 NM_013650 S100a8 S100 calcium-binding protein A8 (calgranulin A)3 80 NM_029796 Lrg1 Leucine-rich �2-glycoprotein 14 35 NM_009141 Cxcl5 Chemokine (C-X-C motif) ligand 5 (LIX)5 35 NM_011315 Saa3 Serum amyloid A36 28 NM_008392 Irg1 Immunoresponsive gene 17 22 NM_009140 Cxcl2 Chemokine (C-X-C motif) ligand 2 (MIP-2)8 21 NM_011259 Reg3a Regenerating islet-derived 3�9 18 NM_008361 Il1b IL-1�10 17 NM_011260 Reg3g Regenerating islet-derived 3�11 17 NM_010819 Clec4d C-type lectin domain family 4, member d12 17 NM_013415 Atp1b2 Na�/K�-ATPase �2-subunit13 16 NM_001033199 AI747448 Calcium-activated chloride channel14 15 NM_001146073 Hexdc Hexosaminidase (glycosyl hydrolase family 20)15 14 NM_008176 Cxcl1 Chemokine (C-X-C motif) ligand 1 (KC)16 13 NM_011036 Reg3b Regenerating islet-derived 3�17 12 NM_009896 Socs1 Suppressor of cytokine signaling 118 12 NM_008599 Cxcl9 Chemokine (C-X-C motif) ligand 9 (MIG)19 11 NM_008489 Lbp LPS-binding protein20 11 NM_175449 Fam26f Hypothetical protein LOC215900
a Total RNA was extracted from the colon of C. rodentium-infected (week 1) and uninfected C57BL/6J mice and subjected to microarray analysis.Levels of mRNA are expressed as ratio of infected over uninfected tissues (fold change) and are sorted in decreasing order of induction for the top 20genes.
3335The Journal of Immunology
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

not only help to control bacterial clearance in the colon, but haveadditional functions in containing systemic infection and tissuedamage.
Analysis of Ab titers against C. rodentium demonstrated thatCXCR3 deficiency significantly delayed induction of specific IgGAbs, whereas specific IgM production was not affected (Fig. 3E).Delayed antibacterial IgG responses may partly explain the de-layed bacterial clearance in the absence of CXCR3, since IgG con-tributes to immune defense against the bacteria (12, 13). Takentogether, these studies show that CXCR3 has a significant but lim-ited role in host defense against C. rodentium.
Loss of CXCR2 severely compromises C. rodentium clearancebut not development of specific Ab responses
Upon realization that CXCR3 was of limited importance in anti-bacterial defense, we turned to CXCR2, the other chemokine re-ceptor whose cognate ligands were strongly and rapidly inducedafter infection. Mice lacking CXCR2 are fertile and generallyhealthy, although they display disturbances in constitutive neutro-phil trafficking (26). After oral C. rodentium infection, CXCR2-deficient mice had bacterial numbers comparable to wild-type con-trols after 1 wk, but subsequently displayed a marked defect incontrolling and eradicating the infection. They continued to havedetectable fecal CFU counts for at least 4 wk, whereas wild-typemice had cleared the bacteria by 3 wk (Fig. 4A). In parallel, in-creased bacterial numbers were found in liver and spleen ofCXCR2-deficient mice (Fig. 4A).
Histologically, CXCR2-deficient mice exhibited increased signsof colonic inflammation and ulceration after 2 wk, while infection-associated epithelial hyperplasia normally found in wild-type micewas attenuated in the knockout mice (Fig. 4B). No pathologicalchanges were observed in the liver of either CXCR2-deficient orwild-type mice (Fig. 4C), suggesting that CXCR2, unlike CXCR3,had no unique role in preventing infection-associated liver dam-age. Immunohistological staining demonstrated that CXCR2-defi-
cient mice had greater bacterial colonization of the colon surfaceand deeper infiltration of C. rodentium into colonic crypts (Fig.4D). In some crypts of knockout mice, bacteria were foundthroughout the entire crypt including the base, whereas bacterialcolonization was always limited to the colon surface in wild-typemice (Fig. 4D).
In contrast to CXCR3 knockout mice, CXCR2-deficient micewere not impaired in their ability to mount specific Ab re-sponses against the bacteria, since they had elevated (ratherthan decreased) levels of antibacterial IgM and IgG relative towild-type mice after 2 wk and no significant differences after 3wk (Fig. 4E). Taken together, these results indicate that CXCR2plays an important and indispensable role in mucosal defenseagainst C. rodentium.
CXCR2 deficiency abolishes infection-associated influx ofneutrophils as critical effector cells
Given the key role of CXCR2 in host defense against C. roden-tium, we began to explore the mechanism by which the receptormediates its physiologic functions. Flow cytometric analysisshowed that the number of CXCR2� cells in the colon increasedby 9-fold 1 wk after infection of wild-type mice (Fig. 5A). Thevast majority (95%) of these cells were positive for the neutrophilmarker Gr-1. Conversely, a large proportion (84%) of Gr-1� cellswere also CXCR2� (Fig. 5A). Consistent with the flow cytometri-cally detected influx of neutrophils, levels of the neutrophil-spe-cific marker MPO increased 8-fold in the colon of infected mice(Fig. 5B).
In contrast to wild-type mice, neutrophil numbers did not in-crease in the colon of infected CXCR2-deficient mice, as deter-mined by flow cytometry for Gr-1� cells and MPO assay (Fig. 5B).Immunohistological staining confirmed that CXCR2-deficientmice were severely impaired in colonic neutrophil influx, whereaswild-type mice exhibited marked mucosal infiltration in the sub-epithelial surface and pericryptal regions (Fig. 5C). These data
FIGURE 2. Chemokine expressionin response to C. rodentium infection.C57BL/6J mice were infected with C.rodentium or left uninfected (week 0),and mRNA expression was analyzedby real-time PCR for the indicated che-mokines in the colon (A) or the indi-cated organs (B). Expression levels areshown relative to uninfected controls.Data are means � SEM (n � 4). C,Explants of whole colon tissue werecultured for 6 h, and levels of CXCL5and CXCL9 in the supernatants wereexamined by ELISA. Data aremeans � SEM (n � 4). D, The indi-cated cell types were isolated from thecolon of infected (2 wk) and uninfectedmice by differential detachment (epi-thelial cells) or FACS (leukocyte sub-sets) and examined for CXC chemo-kine expression by real-time PCR. Dataare shown as ratio of mRNA levels incells from infected over uninfectedmice. �, p � 0.05 by t test comparedwith the levels in uninfected mice.
3336 CXCR2 AND CXCR3 IN PATHOGENESIS OF C. rodentium INFECTION
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

demonstrate that CXCR2 is critical for neutrophil influx into thecolon in response to C. rodentium infection.
Because CXCR2 deficiency led to a loss of normal mucosalneutrophil recruitment, we asked whether these cells can act asdirect effectors against the bacteria. Neutrophils were isolatedfrom the bone marrow and tested for their ability to kill C.rodentium. Resting neutrophils could not kill the bacteria overa 6-h incubation period (Fig. 5D). However, stimulation withTNF-� induced significant killing capacity in the neutrophils,which was further enhanced by costimulation with IFN-� (Fig.5D). Both cytokines are known to be important in host defenseagainst C. rodentium (14, 15). These results indicate that acti-vated neutrophils kill C. rodentium, which can explain their rolein antibacterial defense (11).
Critical role of CXCR2 in protection against infection-associateddiarrhea
In addition to impaired bacterial clearance, we observed thatinfection of CXCR2-deficient mice, but not wild-type controls,was accompanied by marked diarrhea. Thus, analysis of fecalwater content showed that diarrhea began within 7 days afterinfection and persisted for 2–3 wk, whereas no evidence ofincreased fecal water content was observed in wild-type mice atany time after infection (Fig. 6A). The underlying mechanismsof diarrhea were local in nature, since electrophysiologicalstudies of explanted colonic mucosa in Ussing chambers dem-onstrated reversal of the normally positive baseline Isc in wild-type mice to a negative baseline Isc in infected CXCR2-deficient
mice (Fig. 6B). These results are consistent with a loss of nor-mal sodium and/or chloride uptake mechanisms in the colon,which can contribute to diarrhea by removing the driving forcefor passive water absorption from the lumen. In contrast, trans-mucosal resistance was not significantly different betweenknockout or wild-type mice before or after infection (data notshown), indicating that the epithelium was not differentiallycompromised by infection.
To define the molecular basis of the apparent defects in iontransport in CXCR2-deficient mice, we analyzed the expressionof key ion transporters in the colon. Wild-type mice showedmarked increases in mRNA expression for ATPase (�2-sub-unit), CFTR, and DRA, and modest increases for NHE3 andPAT1 upon infection (Fig. 6C). In contrast, CXCR2-deficientmice were severely impaired in the induction of these ion trans-porters. Immunoblots confirmed that DRA was strongly inducedin wild-type mice, but only weakly in their CXCR2-deficientlittermates in response to infection (Fig. 6D). Expression levelsof the epithelium-specific product villin were not different be-tween the groups (data not shown), indicating that differentialloss of epithelial cells was not responsible for the observeddifferences in ion transporter expression. Because deficiency ofDRA and NHE3 can cause diarrhea due to a failure to absorbchloride or sodium ions from the lumen, respectively (27, 28),these data strongly suggest that the failure to normally induceexpression of these ion transporters is responsible for the in-fection-induced diarrhea in the absence of CXCR2.
FIGURE 3. C. rodentium infectionof CXCR3-deficient mice. Cxcr3�/�
(E) and wild-type mice (F) were orallyinfected with C. rodentium. A, Bacterialnumbers in stool, liver, and spleen weredetermined by CFU assay. Data aremeans � SEM (n 20). �, p � 0.05 byrank sum test compared with wild-typemice at the same time. B, Paraffin sec-tions of the colon were prepared andstained with H&E. Photographs weretaken with a �20 lens and processedequally for inclusion in the figure. C,MPO activity in the colon of infectedmice (2 wk) was determined by enzy-matic assay, and was normalizedagainst tissue weight. MPO activity inboth groups of uninfected mice was�0.01 U/g. D, H&E-stained paraffinsections of the liver show thatCxcr3�/� mice, but not wild-type mice,had distinct necrotic areas (arrows). E,Serum was collected at the indicatedtimes after infection, and IgM and IgGAbs against C. rodentium were deter-mined by ELISA with whole bacteriaas Ag. Data are means � SEM (n � 6mice/group). In C and E, �, p � 0.05 byt test compared with wild-type mice.
3337The Journal of Immunology
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Neutrophil influx rather than bacterial load determines diarrheaphenotype
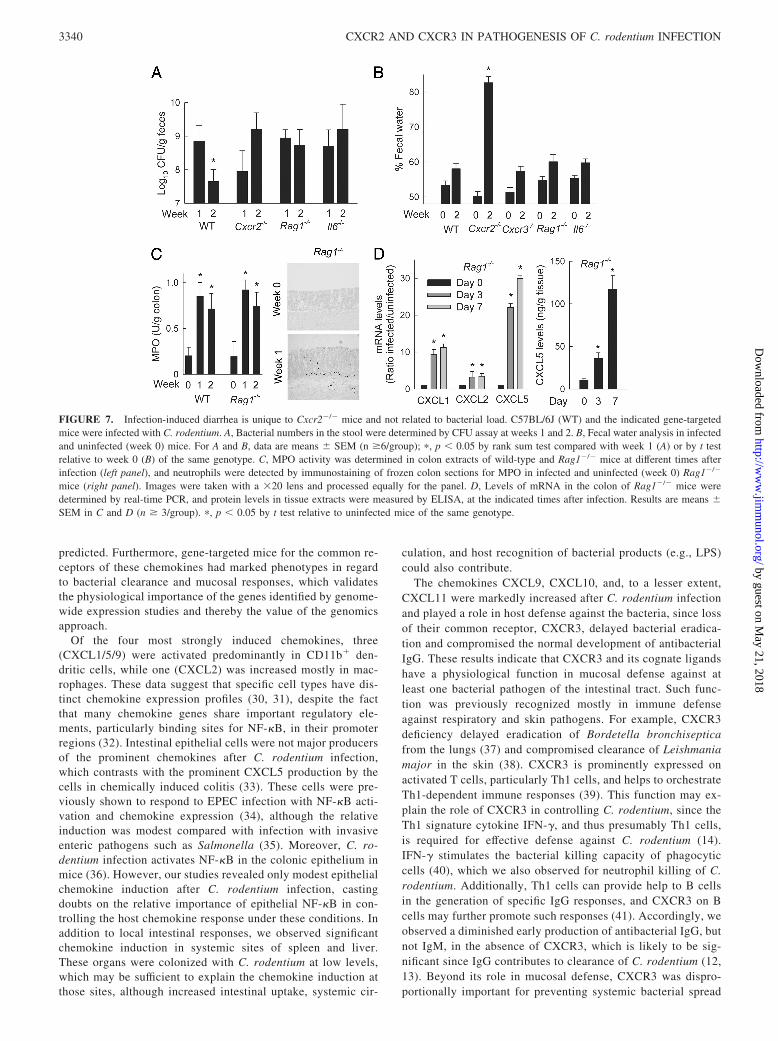
The electrophysiological alterations were not simply related to theincreased bacterial load in CXCR2-deficient compared withwild-type mice, since infection of mice lacking RAG1, IL-6, orCXCR3 also led to high bacterial colonization after 2 wk (Figs. 3Aand 7A), but these mice showed no signs of diarrhea after infection(Fig. 7B). In stark contrast to CXCR2-deficient mice, the threegroups of knockout mice without diarrhea had increased neutrophilnumbers and/or MPO levels in the colon after C. rodentium infec-tion (Figs. 3C and 7C) (16), suggesting that neutrophils can protectagainst infection-associated diarrhea beyond their antimicrobialfunction and regardless of the underlying immune defect in thedifferent mice. Furthermore, mucosal neutrophil influx in RAG1-deficient mice was associated with increased colonic mRNA ex-pression for CXCL1, CXCL2, and CXCL5 and increased proteinproduction of at least one of these chemokines, CXCL5 (Fig. 7D).These data highlight that local chemokine induction and the ac-companying CXCR2-dependent neutrophil influx represent innateresponses to infection.
DiscussionInfections with EPEC are a major worldwide cause of diarrhealdisease in children, yet the pathophysiologic mechanisms of
disease and host defense are not well understood. Much isknown about the interactions between the bacteria and hostcells, particularly regarding their ability to adhere to epithelialcells (29), but these bacteria do not produce classical entero-toxins, suggesting that a complex cross-talk between bacteriaand host cells, and the ensuing host responses, are important fordetermining the disease manifestations. The analysis of suchresponses often proceeds by testing individual host factors andprocesses in hypothesis-driven approaches. Important progresshas been made with these approaches, particularly when com-bined with the commonly used murine infection model with theA/E lesion-forming pathogen C. rodentium. For example, thecytokines IFN-�, TNF-�, and IL-6 were shown to have indis-pensable and distinct roles in host defense against C. rodentium(14 –16). However, these studies were limited to a single hostfactor, precluding evaluation of their relative overall impor-tance and interdependence with other host factors and pro-cesses. As an alternative, discovery-driven approaches are be-coming increasingly prominent in obtaining a broader genomicview of the host response to infection. Our data and those inother infection models (20, 22) demonstrate that such genomicapproaches can yield unbiased information about dominant fea-tures of the host response to infection. For example, analysis ofsusceptible and resistant strains of mice yielded key differences
FIGURE 4. Importance of CXCR2in host defense against C. rodentium.Cxcr2�/� mice (E) and wild-type mice(F) were infected with C. rodentium. A,Bacterial numbers in stool, liver, andspleen were determined by CFU assay.Data are means � SEM (n 16). �,p � 0.05 by rank sum test comparedwith wild-type mice at the same time. Band C, H&E-stained paraffin sections ofcolon and liver. D, Paraffin sections ofthe colon were prepared and stained byindirect immunofluorescence with Absagainst C. rodentium (dark gray) andcounterstained with a nuclear stain(light gray). Photographs in B–D weretaken with a �20 lens and processedequally for each panel. E, Serum wascollected and tested by ELISA for IgMand IgG Abs against C. rodentium us-ing whole bacteria as Ag. Data aremeans � SEM (n � 10/group). �, p �0.05 by t test compared with wild-typemice.
3338 CXCR2 AND CXCR3 IN PATHOGENESIS OF C. rodentium INFECTION
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

in the colonic expression of ion transporters after C. rodentiuminfection (20). We found here that a specific array of chemo-kines were among the most strongly induced genes in response
to C. rodentium infection. Although chemokines are often earlyresponse genes in bacterial infections, the particular set of che-mokines induced by C. rodentium could not have been easily
FIGURE 5. Neutrophil involvement in host defense against C. rodentium. A, C57BL/6 mice were infected with C. rodentium or left uninfected (week0). Lamina propria cells were isolated from the colon and examined by flow cytometry for CXCR2 and Gr-1 expression. Numbers in the quadrants indicatethe percentage of each cell population relative to total lamina propria cells. B, Time course analysis of Gr-1� cells (left panel) and MPO activity (rightpanel) in the colon of wild-type (F) and Cxcr2�/� mice (E). Data are means � SEM (n � 4/group). �, p � 0.05 by t test compared with wild-type mice.C, Frozen sections of the colon were prepared and stained by an indirect immunoperoxidase technique with an Ab against MPO (dark gray), andcounterstained with hematoxylin (light gray). Enlargements of the mid-crypt regions are shown in the insets. Images were taken with a �20x lens andprocessed equally for either the low- or high-magnification panels. D, Neutrophils were prepared from the bone marrow of wild-type mice and werepreincubated for 1 h with TNF-�, IFN-�, or TNF-� plus IFN-�, or left unstimulated. C. rodentium was added at a multiplicity of infection of 1:1, and totalbacterial numbers in the cultures after hypotonic cell lysis were determined by CFU assay at the indicated times. Data are means � SEM (n 3/group)of a representative experiment (�, p � 0.05 by t test relative to untreated neutrophils). Two additional experiments yielded similar results.
FIGURE 6. Severe diarrhea in C. rodentium-infectedCxcr2�/� mice. Cxcr2�/� mice (open symbols andbars) and wild-type mice (closed symbols and bars)were infected with C. rodentium. A, Fecal water contentwas determined at the indicated times after infection.Data are means � SEM (n 6/group). �, p � 0.05 byt test relative to wild-type mice. B, Electrophysiologicalanalysis of net ion transport (baseline short-circuit cur-rent, Isc) in Ussing chamber-mounted, muscle-strippedcolon segments from infected and uninfected (week 0)mice. Data are means � SEM (n 6/group). �, p �0.05 by t test. C, Expression levels of the indicated iontransporters and pumps in the colon of infected (week 2)and uninfected mice were determined by real-time PCRand are shown as ratios of infected over uninfectedmice. Data are means � SEM (n 3/group). �, p �0.05 by t test. D, Immunoblot analysis of DRA andGAPDH in infected and uninfected (week 0) mice. Foreach lane, colon extracts from four to six mice werepooled before analysis.
3339The Journal of Immunology
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
predicted. Furthermore, gene-targeted mice for the common re-ceptors of these chemokines had marked phenotypes in regardto bacterial clearance and mucosal responses, which validatesthe physiological importance of the genes identified by genome-wide expression studies and thereby the value of the genomicsapproach.
Of the four most strongly induced chemokines, three(CXCL1/5/9) were activated predominantly in CD11b� den-dritic cells, while one (CXCL2) was increased mostly in mac-rophages. These data suggest that specific cell types have dis-tinct chemokine expression profiles (30, 31), despite the factthat many chemokine genes share important regulatory ele-ments, particularly binding sites for NF-�B, in their promoterregions (32). Intestinal epithelial cells were not major producersof the prominent chemokines after C. rodentium infection,which contrasts with the prominent CXCL5 production by thecells in chemically induced colitis (33). These cells were pre-viously shown to respond to EPEC infection with NF-�B acti-vation and chemokine expression (34), although the relativeinduction was modest compared with infection with invasiveenteric pathogens such as Salmonella (35). Moreover, C. ro-dentium infection activates NF-�B in the colonic epithelium inmice (36). However, our studies revealed only modest epithelialchemokine induction after C. rodentium infection, castingdoubts on the relative importance of epithelial NF-�B in con-trolling the host chemokine response under these conditions. Inaddition to local intestinal responses, we observed significantchemokine induction in systemic sites of spleen and liver.These organs were colonized with C. rodentium at low levels,which may be sufficient to explain the chemokine induction atthose sites, although increased intestinal uptake, systemic cir-
culation, and host recognition of bacterial products (e.g., LPS)could also contribute.
The chemokines CXCL9, CXCL10, and, to a lesser extent,CXCL11 were markedly increased after C. rodentium infectionand played a role in host defense against the bacteria, since lossof their common receptor, CXCR3, delayed bacterial eradica-tion and compromised the normal development of antibacterialIgG. These results indicate that CXCR3 and its cognate ligandshave a physiological function in mucosal defense against atleast one bacterial pathogen of the intestinal tract. Such func-tion was previously recognized mostly in immune defenseagainst respiratory and skin pathogens. For example, CXCR3deficiency delayed eradication of Bordetella bronchisepticafrom the lungs (37) and compromised clearance of Leishmaniamajor in the skin (38). CXCR3 is prominently expressed onactivated T cells, particularly Th1 cells, and helps to orchestrateTh1-dependent immune responses (39). This function may ex-plain the role of CXCR3 in controlling C. rodentium, since theTh1 signature cytokine IFN-�, and thus presumably Th1 cells,is required for effective defense against C. rodentium (14).IFN-� stimulates the bacterial killing capacity of phagocyticcells (40), which we also observed for neutrophil killing of C.rodentium. Additionally, Th1 cells can provide help to B cellsin the generation of specific IgG responses, and CXCR3 on Bcells may further promote such responses (41). Accordingly, weobserved a diminished early production of antibacterial IgG, butnot IgM, in the absence of CXCR3, which is likely to be sig-nificant since IgG contributes to clearance of C. rodentium (12,13). Beyond its role in mucosal defense, CXCR3 was dispro-portionally important for preventing systemic bacterial spread
FIGURE 7. Infection-induced diarrhea is unique to Cxcr2�/� mice and not related to bacterial load. C57BL/6J (WT) and the indicated gene-targetedmice were infected with C. rodentium. A, Bacterial numbers in the stool were determined by CFU assay at weeks 1 and 2. B, Fecal water analysis in infectedand uninfected (week 0) mice. For A and B, data are means � SEM (n �6/group); �, p � 0.05 by rank sum test compared with week 1 (A) or by t testrelative to week 0 (B) of the same genotype. C, MPO activity was determined in colon extracts of wild-type and Rag1�/� mice at different times afterinfection (left panel), and neutrophils were detected by immunostaining of frozen colon sections for MPO in infected and uninfected (week 0) Rag1�/�
mice (right panel). Images were taken with a �20 lens and processed equally for the panel. D, Levels of mRNA in the colon of Rag1�/� mice weredetermined by real-time PCR, and protein levels in tissue extracts were measured by ELISA, at the indicated times after infection. Results are means �SEM in C and D (n � 3/group). �, p � 0.05 by t test relative to uninfected mice of the same genotype.
3340 CXCR2 AND CXCR3 IN PATHOGENESIS OF C. rodentium INFECTION
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

to liver and spleen, and in protecting against necrotic liver dam-age. Such spread may occur secondarily to increased mucosaldamage and epithelial barrier loss, although none was apparenthistologically and deficiency in at least one of the key CXCR3ligands, CXCL10, is associated with enhanced rather than di-minished epithelial barrier function in the colon (42). Instead,CXCR3 may exert a unique immunological function in the liveror spleen. For example, the receptor is required for effectiverecruitment of effector T cells to the liver and is found on mostT cells in different inflammatory conditions of the liver (43). Ifhepatic T cell-dependent defenses fail to develop normally inthe absence of CXCR3, it may lead to enhanced bacterial pro-liferation and hepatic necrosis. Despite the importance ofCXCR3 in controlling C. rodentium in intestine and liver, it wasultimately not required for eradication, emphasizing that redun-dancy of immune mechanisms in general, and chemokines inparticular, is central to the development of robust host defensesagainst pathogenic microbes (32).
The chemokines CXCL1, CXCL2, and CXCL5 were amongthe most highly induced genes in the C. rodentium infectionmodel. All three are cognate ligands for CXCR2, and deficiencyof the receptor compromised bacterial clearance, indicating thatthese chemokines constitute a critical component of the intes-tinal defense against the bacteria. Similarly, CXCR2 was pre-viously shown to be indispensable in immune defense againstpathogenic bacteria in other tissues, including Pseudomonasaeruginosa and Nocardia asteroides in the lungs (44, 45), anduropathogenic E. coli in the urinary tract (46). The protectivefunctions of CXCR2 are by no means universal, since the re-ceptor is not required for clearance of influenza virus in thelungs (47), and is in fact detrimental in models of septic peri-tonitis (48). CXCR2 is strongly expressed on neutrophils, andits cognate ligands are potent neutrophil chemoattractants. Con-sistent with this, we found that most Gr-1� cells that invadedthe mucosa in response to C. rodentium were positive forCXCR2. Conversely, practically all CXCR2� cells were Gr-1�
neutrophils. Importantly, loss of CXCR2 abrogated neutrophilinflux into the colon mucosa, indicating that the receptor and itscognate ligands were critical for orchestrating this process.Neutrophil attraction to sites of infection and damage mediatedby CXCR2 and its ligands is dependent on the tissue contextand stimulation conditions (49, 50). Irrespective of the means ofchemoattraction, neutrophils typically exert their defense func-tions by direct microbial killing of obligate extracellular mi-crobes or facultative intracellular microbes during transient pas-sage between host cells (51). C. rodentium and EPEC residepredominantly outside host cells in the intestinal lumen and atthe epithelial surface, although they can invade host cells andlow numbers are found deeper in the mucosa and in systemicsites. Infiltrating neutrophils were localized around the cryptsthroughout the colonic mucosa and were able to kill C. rodentiumwhen activated, suggesting that they contribute to antibacterial de-fense by forming an active defense barrier against bacterial spreadacross the crypts (11), similar to their function in protecting againstsystemic spread of orally ingested Salmonella (52, 53). Moreover,lack of CXCR2 and the accompanying mucosal neutrophil influxled to heavy and deep bacterial colonization of the crypts, sug-gesting that neutrophils are critical for preventing the build-up ofbacterial repertoires in the crypts. Such microbial colonies in thecrypts are presumably protected against removal in the luminalbulk flow, which could allow them to serve as seed sites for con-tinued luminal infection. We note that neutrophils are necessaryfor mucosal defense against C. rodentium (11), but they are notsufficient since mice lacking T and B cells or IL-6 have high
mucosal neutrophil numbers but exhibit bacterial clearance defects(10, 12, 16).
Watery diarrhea and the accompanying dehydration are hall-marks of EPEC-induced disease in humans. Diarrhea can occurwithin hours of infection, suggesting that bacterial factors drivethese early events (54). Disruption of epithelial tight junctions,increased paracellular permeability, dysregulation of criticalion transporters, and loss of microvilli have been reported tooccur rapidly in response to translocation of EPEC effector pro-teins into host cells (2– 4, 55). However, diarrhea can be vari-able and persist over extended periods up to weeks, suggestingthat host factors are likely to be decisive in later stages of thehuman disease. Infection is accompanied by influx of inflam-matory cells that can release secretagogues, and it leads to mu-cosal up-regulation of receptors whose activation causes fluidsecretion (56). The importance of host factors in determininginfection-induced disease is clearly demonstrated by the findingthat the genetic background of certain strains of mice can renderthem susceptible to massive diarrhea and dehydration-relatedmortality (57). Differential disease susceptibility in these miceis accompanied by altered expression of key solute transporters,particularly DRA and aquaporin 8 (20), although the underlyingmechanisms are not understood. Our data show that CXCR2-dependent processes are critical determinants of infection-in-duced diarrhea, providing strong support for the role of hostfactors in controlling disease outcome upon infection with anA/E lesion-forming pathogen. These murine findings suggestthat similar principles may also underlie the variable diseasemanifestations in human EPEC infections. Mechanistically,CXCR2 was required in the mice for induction of key elementsof ion transport pathways, including DRA, PAT1, and ATP1�2,which may be interpreted as a compensatory induction of fluiduptake processes in the colon to counteract any inflammation-associated increase in secretagogue activities. Loss of DRAcauses a chloride-losing chronic diarrhea in patients and gene-targeted mice (27), suggesting that the failure to induce DRAnormally in CXCR2-deficient mice is probably important in thediarrhea observed in these mice. Although our studies were notdesigned to examine the mechanisms by which CXCR2 mediatessolute transporter induction, one could speculate that neutrophilshave a key function, as these were the main CXCR2-expressingcells after infection. Their absence in CXCR2-deficient mice wasaccompanied by diarrhea, while their presence in several othermodels with high bacterial colonization levels was consistentlyassociated with protection against diarrhea. These observationssuggest that neutrophils have a role in preventing infection-asso-ciated diarrhea that goes beyond their function in antibacterial de-fense. Such protective neutrophil functions have also been dem-onstrated in different models of acute colitis (58). Thus,neutrophils must be viewed as multifunctional cells that contributeto intestinal inflammation and diarrhea under some conditions(59), but also protect against microbial growth and entry, and theaccompanying diarrheal manifestations in other circumstances.Exploiting these protective functions may suggest an avenue formanaging chronic diarrheal diseases in humans caused by EPECand other A/E lesion-causing pathogens.
AcknowledgmentsWe thank Yolanda Andersen, Dustin Hammond, John J. Quinlan III, andLucia Hall for expert technical support.
DisclosuresThe authors have no financial conflicts of interest.
3341The Journal of Immunology
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

References1. Clarke, S. C., R. D. Haigh, P. P. Freestone, and P. H. Williams. 2003. Virulence
of enteropathogenic Escherichia coli, a global pathogen. Clin. Microbiol. Rev.16: 365–378.
2. Chen, H. D., and G. Frankel. 2005. Enteropathogenic Escherichia coli: unravel-ling pathogenesis. FEMS Microbiol. Rev. 29: 83–98.
3. Hodges, K., N. M. Alto, K. Ramaswamy, P. K. Dudeja, and G. Hecht. 2008. Theenteropathogenic Escherichia coli effector protein EspF decreases sodium hydro-gen exchanger 3 activity. Cell. Microbiol. 10: 1735–1745.
4. Gill, R. K., A. Borthakur, K. Hodges, J. R. Turner, D. R. Clayburgh, S. Saksena,A. Zaheer, K. Ramaswamy, G. Hecht, and P. K. Dudeja. 2007. Mechanism un-derlying inhibition of intestinal apical Cl/OH exchange following infection withenteropathogenic E. coli. J. Clin. Invest. 117: 428–437.
5. Johnson, E., and S. W. Barthold. 1979. The ultrastructure of transmissible murinecolonic hyperplasia. Am. J. Pathol. 97: 291–313.
6. Schauer, D. B., and S. Falkow. 1993. Attaching and effacing locus of aCitrobacter freundii biotype that causes transmissible murine colonic hyperpla-sia. Infect. Immun. 61: 2486–2492.
7. Mundy, R., T. T. MacDonald, G. Dougan, G. Frankel, and S. Wiles. 2005.Citrobacter rodentium of mice and man. Cell. Microbiol. 7: 1697–1706.
8. Borenshtein, D., M. E. McBee, and D. B. Schauer. 2008. Utility of theCitrobacter rodentium infection model in laboratory mice. Curr. Opin. Gastro-enterol. 24: 32–37.
9. Bry, L., and M. B. Brenner. 2004. Critical role of T cell-dependent serum anti-body, but not the gut-associated lymphoid tissue, for surviving acute mucosalinfection with Citrobacter rodentium, an attaching and effacing pathogen. J. Im-munol. 172: 433–441.
10. Simmons, C. P., S. Clare, M. Ghaem-Maghami, T. K. Uren, J. Rankin, A. Huett,R. Goldin, D. J. Lewis, T. T. MacDonald, R. A. Strugnell, et al. 2003. Central rolefor B lymphocytes and CD4� T cells in immunity to infection by the attachingand effacing pathogen Citrobacter rodentium. Infect. Immun. 71: 5077–5086.
11. Lebeis, S. L., B. Bommarius, C. A. Parkos, M. A. Sherman, and D. Kalman.2007. TLR signaling mediated by MyD88 is required for a protective innateimmune response by neutrophils to Citrobacter rodentium. J. Immunol. 179:566–577.
12. Maaser, C., M. P. Housley, M. Iimura, J. R. Smith, B. A. Vallance, B. B. Finlay,J. R. Schreiber, N. M. Varki, M. F. Kagnoff, and L. Eckmann. 2004. Clearanceof Citrobacter rodentium requires B cells but not secretory immunoglobulin A(IgA) or IgM antibodies. Infect. Immun. 72: 3315–3324.
13. Yoshida, M., K. Kobayashi, T. T. Kuo, L. Bry, J. N. Glickman, S. M. Claypool,A. Kaser, T. Nagaishi, D. E. Higgins, E. Mizoguchi, et al. 2006. Neonatal Fcreceptor for IgG regulates mucosal immune responses to luminal bacteria. J. Clin.Invest. 116: 2142–2151.
14. Higgins, L. M., G. Frankel, I. Connerton, N. S. Goncalves, G. Dougan, andT. T. MacDonald. 1999. Role of bacterial intimin in colonic hyperplasia andinflammation. Science 285: 588–591.
15. Goncalves, N. S., M. Ghaem-Maghami, G. Monteleone, G. Frankel, G. Dougan,D. J. Lewis, C. P. Simmons, and T. T. MacDonald. 2001. Critical role for tumornecrosis factor � in controlling the number of lumenal pathogenic bacteria andimmunopathology in infectious colitis. Infect. Immun. 69: 6651–6659.
16. Dann, S. M., M. E. Spehlmann, D. C. Hammond, M. Iimura, K. Hase, L. J. Choi,E. Hanson, and L. Eckmann. 2008. IL-6-dependent mucosal protection preventsestablishment of a microbial niche for attaching/effacing lesion-forming entericbacterial pathogens. J. Immunol. 180: 6816–6826.
17. Simmons, C. P., N. S. Goncalves, M. Ghaem-Maghami, M. Bajaj-Elliott,S. Clare, B. Neves, G. Frankel, G. Dougan, and T. T. MacDonald. 2002. Impairedresistance and enhanced pathology during infection with a noninvasive, attach-ing-effacing enteric bacterial pathogen. Citrobacter rodentium, in mice lackingIL-12 or IFN-�. J. Immunol. 168: 1804–1812.
18. Zheng, Y., P. A. Valdez, D. M. Danilenko, Y. Hu, S. M. Sa, Q. Gong,A. R. Abbas, Z. Modrusan, N. Ghilardi, F. J. de Sauvage, and W. Ouyang. 2008.Interleukin-22 mediates early host defense against attaching and effacing bacte-rial pathogens. Nat. Med. 14: 282–289.
19. Ishigame, H., S. Kakuta, T. Nagai, M. Kadoki, A. Nambu, Y. Komiyama,N. Fujikado, Y. Tanahashi, A. Akitsu, H. Kotaki, et al. 2009. Differential roles ofinterleukin-17A and -17F in host defense against mucoepithelial bacterial infec-tion and allergic responses. Immunity 30: 108–119.
20. Borenshtein, D., R. C. Fry, E. B. Groff, P. R. Nambiar, V. J. Carey, J. G. Fox, andD. B. Schauer. 2008. Diarrhea as a cause of mortality in a mouse model ofinfectious colitis. Genome Biol. 9: R122.
21. Costello, C. M., N. Mah, R. Hasler, P. Rosenstiel, G. H. Waetzig, A. Hahn, T. Lu,Y. Gurbuz, S. Nikolaus, M. Albrecht, et al. 2005. Dissection of the inflammatorybowel disease transcriptome using genome-wide cDNA microarrays. PLoS Med.2: e199.
22. Kobayashi, K. S., M. Chamaillard, Y. Ogura, O. Henegariu, N. Inohara,G. Nunez, and R. A. Flavell. 2005. Nod2-dependent regulation of innate andadaptive immunity in the intestinal tract. Science 307: 731–734.
23. Foell, D., H. Wittkowski, T. Vogl, and J. Roth. 2007. S100 proteins expressed inphagocytes: a novel group of damage-associated molecular pattern molecules.J. Leukocyte Biol. 81: 28–37.
24. Cash, H. L., C. V. Whitham, and L. V. Hooper. 2006. Refolding, purification, andcharacterization of human and murine RegIII proteins expressed in Escherichiacoli. Protein Expression Purif. 48: 151–159.
25. Hancock, W. W., B. Lu, W. Gao, V. Csizmadia, K. Faia, J. A. King, S. T. Smiley,M. Ling, N. P. Gerard, and C. Gerard. 2000. Requirement of the chemokinereceptor CXCR3 for acute allograft rejection. J. Exp. Med. 192: 1515–1520.
26. Cacalano, G., J. Lee, K. Kikly, A. M. Ryan, S. Pitts-Meek, B. Hultgren,W. I. Wood, and M. W. Moore. 1994. Neutrophil and B cell expansion in micethat lack the murine IL-8 receptor homolog. Science 265: 682–684.
27. Schweinfest, C. W., D. D. Spyropoulos, K. W. Henderson, J. H. Kim,J. M. Chapman, S. Barone, R. T. Worrell, Z. Wang, and M. Soleimani. 2006.Slc26a3 (dra)-deficient mice display chloride-losing diarrhea, enhanced colonicproliferation, and distinct up-regulation of ion transporters in the colon. J. Biol.Chem. 281: 37962–37971.
28. Schultheis, P. J., L. L. Clarke, P. Meneton, M. L. Miller, M. Soleimani,L. R. Gawenis, T. M. Riddle, J. J. Duffy, T. Doetschman, T. Wang, et al. 1998.Renal and intestinal absorptive defects in mice lacking the NHE3 Na�/H� ex-changer. Nat. Genet. 19: 282–285.
29. Frankel, G., and A. D. Phillips. 2008. Attaching effacing Escherichia coli andparadigms of Tir-triggered actin polymerization: getting off the pedestal. Cell.Microbiol. 10: 549–556.
30. Volpe, E., G. Cappelli, M. Grassi, A. Martino, A. Serafino, V. Colizzi,N. Sanarico, and F. Mariani. 2006. Gene expression profiling of human macro-phages at late time of infection with Mycobacterium tuberculosis. Immunology118: 449–460.
31. Zaharik, M. L., T. Nayar, R. White, C. Ma, B. A. Vallance, N. Straka, X. Jiang,J. Rey-Ladino, C. Shen, and R. C. Brunham. 2007. Genetic profiling of dendriticcells exposed to live- or ultraviolet-irradiated Chlamydia muridarum revealsmarked differences in CXC chemokine profiles. Immunology 120: 160–172.
32. Viola, A., and A. D. Luster. 2008. Chemokines and their receptors: drug targetsin immunity and inflammation. Annu. Rev. Pharmacol. Toxicol. 48: 171–197.
33. Kwon, J. H., A. C. Keates, P. M. Anton, M. Botero, J. D. Goldsmith, andC. P. Kelly. 2005. Topical antisense oligonucleotide therapy against LIX, anenterocyte-expressed CXC chemokine, reduces murine colitis. Am. J. Physiol.289: G1075–G1083.
34. Savkovic, S. D., A. Koutsouris, and G. Hecht. 1997. Activation of NF-�B inintestinal epithelial cells by enteropathogenic Escherichia coli. Am. J. Physiol.273: C1160–C1167.
35. Elewaut, D., J. A. DiDonato, J. M. Kim, F. Truong, L. Eckmann, andM. F. Kagnoff. 1999. NF-�B is a central regulator of the intestinal epithelial cellinnate immune response induced by infection with enteroinvasive bacteria. J. Im-munol. 163: 1457–1466.
36. Wang, Y., G. S. Xiang, F. Kourouma, and S. Umar. 2006. Citrobacter rodentium-induced NF-�B activation in hyperproliferating colonic epithelia: role of p65(Ser536) phosphorylation. Br. J. Pharmacol. 148: 814–824.
37. Widney, D. P., Y. Hu, A. K. Foreman-Wykert, K. C. Bui, T. T. Nguyen, B. Lu,C. Gerard, J. F. Miller, and J. B. Smith. 2005. CXCR3 and its ligands participatein the host response to Bordetella bronchiseptica infection of the mouse respi-ratory tract but are not required for clearance of bacteria from the lung. Infect.Immun. 73: 485–493.
38. Rosas, L. E., J. Barbi, B. Lu, Y. Fujiwara, C. Gerard, V. M. Sanders, andA. R. Satoskar. 2005. CXCR3�/� mice mount an efficient Th1 response but failto control Leishmania major infection. Eur. J. Immunol. 35: 515–523.
39. Bonecchi, R., G. Bianchi, P. P. Bordignon, D. D’Ambrosio, R. Lang, A. Borsatti,S. Sozzani, P. Allavena, P. A. Gray, A. Mantovani, and F. Sinigaglia. 1998.Differential expression of chemokine receptors and chemotactic responsivenessof type 1 T helper cells (Th1s) and Th2s. J. Exp. Med. 187: 129–134.
40. Fu, Y. K., S. Arkins, Y. M. Li, R. Dantzer, and K. W. Kelley. 1994. Reductionin superoxide anion secretion and bactericidal activity of neutrophils from agedrats: reversal by the combination of � interferon and growth hormone. Infect.Immun. 62: 1–8.
41. Park, M. K., D. Amichay, P. Love, E. Wick, F. Liao, A. Grinberg, R. L. Rabin,H. H. Zhang, S. Gebeyehu, T. M. Wright, et al. 2002. The CXC chemokinemurine monokine induced by IFN-� (CXC chemokine ligand 9) is made byAPCs, targets lymphocytes including activated B cells, and supports antibodyresponses to a bacterial pathogen in vivo. J. Immunol. 169: 1433–1443.
42. Sasaki, S., H. Yoneyama, K. Suzuki, H. Suriki, T. Aiba, S. Watanabe,Y. Kawauchi, H. Kawachi, F. Shimizu, K. Matsushima, et al. 2002. Blockade ofCXCL10 protects mice from acute colitis and enhances crypt cell survival. Eur.J. Immunol. 32: 3197–3205.
43. Zeremski, M., L. M. Petrovic, L. Chiriboga, Q. B. Brown, H. T. Yee,M. Kinkhabwala, I. M. Jacobson, R. Dimova, M. Markatou, and A. H. Talal.2008. Intrahepatic levels of CXCR3-associated chemokines correlate with liverinflammation and fibrosis in chronic hepatitis C. Hepatology 48: 1440–1450.
44. Moore, T. A., M. W. Newstead, R. M. Strieter, B. Mehrad, B. L. Beaman, andT. J. Standiford. 2000. Bacterial clearance and survival are dependent on CXCchemokine receptor-2 ligands in a murine model of pulmonary Nocardia aster-oides infection. J. Immunol. 164: 908–915.
45. Tsai, W. C., R. M. Strieter, B. Mehrad, M. W. Newstead, X. Zeng, andT. J. Standiford. 2000. CXC chemokine receptor CXCR2 is essential for protec-tive innate host response in murine Pseudomonas aeruginosa pneumonia. Infect.Immun. 68: 4289–4296.
46. Svensson, M., H. Irjala, C. Svanborg, and G. Godaly. 2008. Effects of epithelialand neutrophil CXCR2 on innate immunity and resistance to kidney infection.Kidney Int. 74: 81–90.
47. Wareing, M. D., A. L. Shea, C. A. Inglis, P. B. Dias, and S. R. Sarawar. 2007.CXCR2 is required for neutrophil recruitment to the lung during influenza virusinfection, but is not essential for viral clearance. Viral Immunol. 20: 369–378.
48. Ness, T. L., C. M. Hogaboam, R. M. Strieter, and S. L. Kunkel. 2003. Immu-nomodulatory role of CXCR2 during experimental septic peritonitis. J. Immunol.171: 3775–3784.
3342 CXCR2 AND CXCR3 IN PATHOGENESIS OF C. rodentium INFECTION
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

49. Johnston, R. A., J. P. Mizgerd, and S. A. Shore. 2005. CXCR2 is essential formaximal neutrophil recruitment and methacholine responsiveness after ozone ex-posure. Am. J. Physiol. 288: L61–L67.
50. Coates, N. J., and S. R. McColl. 2001. Production of chemokines in vivo inresponse to microbial stimulation. J. Immunol. 166: 5176–5182.
51. Appelberg, R. 2007. Neutrophils and intracellular pathogens: beyond phagocy-tosis and killing. Trends Microbiol. 15: 87–92.
52. Vassiloyanakopoulos, A. P., S. Okamoto, and J. Fierer. 1998. The crucial role ofpolymorphonuclear leukocytes in resistance to Salmonella dublin infections ingenetically susceptible and resistant mice. Proc. Natl. Acad. Sci. USA 95:7676–7681.
53. Cheminay, C., D. Chakravortty, and M. Hensel. 2004. Role of neutrophils inmurine salmonellosis. Infect. Immun. 72: 468–477.
54. Levine, M. M., E. J. Bergquist, D. R. Nalin, D. H. Waterman, R. B. Hornick,C. R. Young, and S. Sotman. 1978. Escherichia coli strains that cause diarrhoeabut do not produce heat-labile or heat-stable enterotoxins and are non-invasive.Lancet 1: 1119–1122.
55. McNamara, B. P., A. Koutsouris, C. B. O’Connell, J. P. Nougayrede,M. S. Donnenberg, and G. Hecht. 2001. Translocated EspF protein from enter-
opathogenic Escherichia coli disrupts host intestinal barrier function. J. Clin.Invest. 107: 621–629.
56. Hecht, G., J. A. Marrero, A. Danilkovich, K. A. Matkowskyj, S. D. Savkovic,A. Koutsouris, and R. V. Benya. 1999. Pathogenic Escherichia coli increase Cl-secretion from intestinal epithelia by upregulating galanin-1 receptor expression.J. Clin. Invest. 104: 253–262.
57. Borenshtein, D., P. R. Nambiar, E. B. Groff, J. G. Fox, and D. B. Schauer. 2007.Development of fatal colitis in FVB mice infected with Citrobacter rodentium.Infect. Immun. 75: 3271–3281.
58. Kuhl, A. A., H. Kakirman, M. Janotta, S. Dreher, P. Cremer, N. N. Pawlowski,C. Loddenkemper, M. M. Heimesaat, K. Grollich, M. Zeitz, et al. 2007. Aggra-vation of different types of experimental colitis by depletion or adhesion blockadeof neutrophils. Gastroenterology 133: 1882–1892.
59. Madara, J. L., T. W. Patapoff, B. Gillece-Castro, S. P. Colgan, C. A. Parkos,C. Delp, and R. J. Mrsny. 1993. 5�-Adenosine monophosphate is the neutrophil-derived paracrine factor that elicits chloride secretion from T84 intestinal epi-thelial cell monolayers. J. Clin. Invest. 91: 2320–2325.
3343The Journal of Immunology
by guest on May 21, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from