conserved genetic programs in insect and mammalian brain development
TRANSCRIPT

Conserved genetic programsin insect and mammalianbrain developmentFrank Hirth and Heinrich Reichert*
Summary
In recent years it has become evident that the developmental regulatory genesinvolved in patterning the embryonic body plan are conserved throughout theanimal kingdom. Striking examples are the orthodenticle (otd/Otx) gene family andthe Hox gene family, both of which act in the specification of anteroposteriorpolarity along the embryonic body axis. Studies carried out in Drosophila andmouse now demonstrate that these genes are also involved in the formation of theinsect and mammalian brain; the otd/Otx genes are involved in rostral braindevelopment and the Hox genes are involved in caudal brain development. Thesestudies also show that the genes of the otd/Otx family can functionally replaceeach other in cross-phylum rescue experiments and indicate that the geneticmechanisms underlying pattern formation in insect and mammalian brain develop-ment are evolutionarily conserved. BioEssays 21:677–684, 1999.r 1999 John Wiley & Sons, Inc.
IntroductionUntil recently, the brains of insects and vertebrates have beenconsidered to be unrelated. Indeed, at the overt anatomicallevel, the two brain types appear to be radically different, anda comparison of their organization in the adult does not revealany obvious brain structure that might be homologous orotherwise related in evolutionary terms (Fig. 1). However,remarkable similarities become apparent if one considers themolecular anatomical level of regulatory gene expression,which gives rise to these brain structures during embryogen-esis. Homologous regulatory genes are being identified thatcontrol regionalization, patterning, and identity in embryonicbrain development in a comparable manner in insects andvertebrates. Among these are the homeotic or Hox genes andthe genes belonging to the otd/Otx family. The aim of thisreview is to highlight recent experimental evidence fromDrosophila and mouse, which indicates that not only the
structure and expression, but also the function of these genesin embryonic brain development is highly conserved. Thesefindings are discussed in the context of a common evolution-ary origin of the insect and mammalian brain.
Embryonic development of the brain in insectsand vertebratesThe insect brain is composed of an anterior supraesophagealganglion and a posterior subesophageal ganglion. The supra-esophageal ganglion is subdivided into the protocerebrum(PC or b1), the deutocerebrum (DC or b2), and the tritocere-brum (TC or b3), whereas the subesophageal ganglion issubdivided into the mandibular (MD or s1), maxillary (MX ors2), and labial (LB or s3) neuromeres.(1–4) The anteriorembryonic brain of Drosophila derives from the procephalicneurogenic region, which is specified to become neuroecto-derm through genetic interactions during gastrulation.(5) Theposterior embryonic brain of Drosophila derives from therostral-most ventral neurogenic region and is specified in amanner similar to that of the ventral nerve cord.(6) Within thecephalic neuroectoderm, progenitor cells called neuroblastsdelaminate to form proliferative clusters comprising neuro-blasts and their progeny, which subsequently establish thedeveloping Drosophila brain.
In vertebrates, inductive interactions between germlayersduring gastrulation cause an early regionalization of the
Institute of Zoology, University of Basel, Rheinsprung 9, CH-4051Basel, Switzerland.Funding agencies: Swiss NSF; EU BIOTECH program.*Correspondence to: Heinrich Reichert, Institute of Zoology, Universityof Basel, Rheinsprung 9, CH-4051 Basel, Switzerland. E-mail:[email protected]
Review articles
BioEssays 21:677–684, r 1999 John Wiley & Sons, Inc. BioEssays 21.8 677

developing anterior neural tube. This leads to a rostrocaudalsubdivision of the anterior neural tube into the rostral fore-brain (prosencephalon or telencephalon/diencephalon) andmidbrain (mesencephalon) regions and into the caudal hind-brain region (rhombencephalon or metencephalon/myelen-cephalon). The developing hindbrain reveals a clear meta-meric organization based on rhombomeres with distinctcytoarchitectures. Its regional diversity is achieved through aprocess of segmentation that bears a superficial resem-blance to segmentation of Drosophila; seven to eight rhombo-meres form by internal subdivision and have a pair-wiseorganization with compartment-like properties.(7) Althoughthe segmental organization of the vertebrate prosencephalonis still debated, comparative studies on the restricted ex-pression domains of regulatory genes in the developingprosencephalon suggest that this region, like the hindbrain, issubdivided into neuromeres known as prosomeres(P1–P6).(8,9)
Homeotic genes and posterior brain developmentin DrosophilaThe homeotic or Hox genes encode for homeodomain tran-scription factors. In Drosophila they are arranged along thechromosome in two gene clusters known as the Antennape-dia (ANT-C) and Bithorax (BX-C) complex. There exists acorrelation between the relative position of the genes withinthe cluster and their spatial and temporal expression patternin the body; genes located towards the 38 end are expressedmore anteriorly and earlier than genes located towards the 58
end. This has been termed spatial and temporal colinearity. Itappears to be related to a conserved functional hierarchyamong the members of the homeotic gene clusters, in thatmore posterior acting genes impose their developmental
specificities upon anterior acting genes, a fact called ‘‘pheno-typic suppression.’’(10)
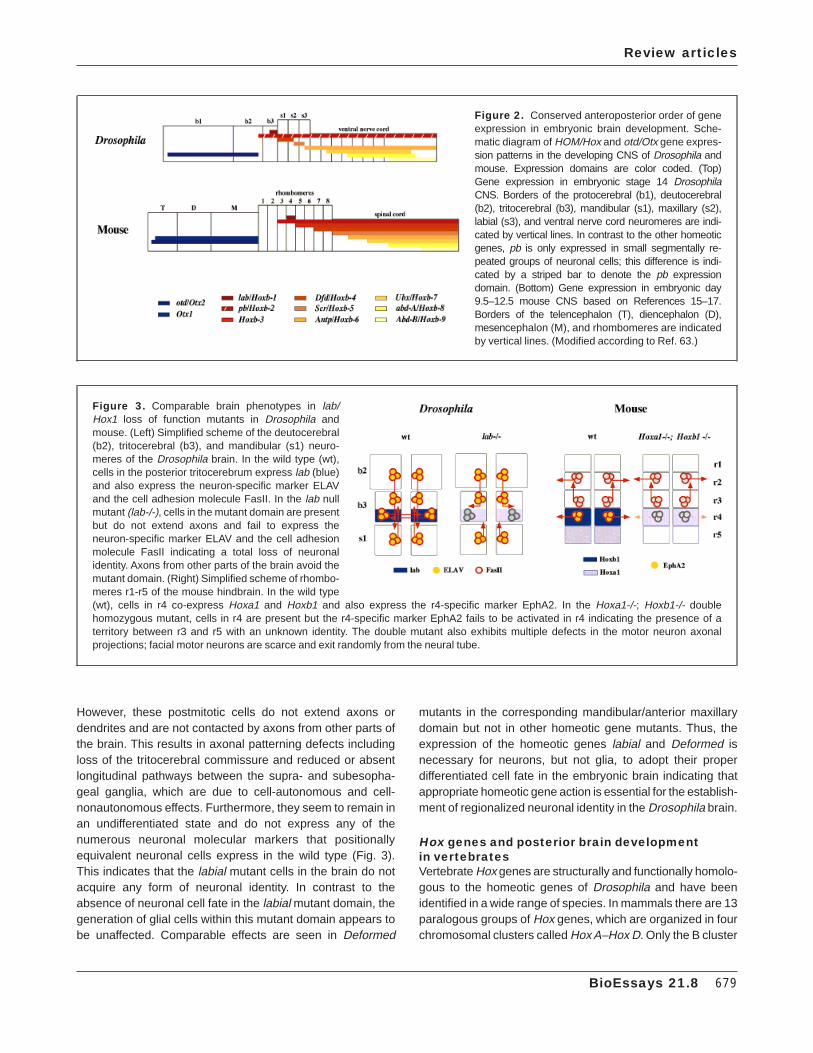
Homeotic genes are expressed in the developing brainand ventral nerve cord of Drosophila in an ordered set ofdomains and their anterior expression boundaries usuallycoincide with neuromere compartment boundaries (Fig. 2).Their relative anteroposterior order of expression in thedeveloping CNS does not strictly fulfill the criterium of spatialcolinearity because the anterior expression boundary of labialis posterior to that of proboscipedia.(11) Interestingly, in embry-onic epidermal structures of Drosophila, spatial colinearity ofhomeotic gene expression is perfectly preserved.(12) Thismeans that the relative order of homeotic gene expression ismore similar in the CNS of the fly and the mouse (see below),than in the CNS and the epidermis of the fly. Most of thehomeotic genes expressed in the Drosophila brain havediscrete, nonoverlapping expression domains and do notextend posteriorly along the CNS.
Mutational inactivation of either of the homeotic geneslabial or Deformed causes severe axonal patterning defectsin the embryonic brain.(11) Thus, in labial null mutants,regionalized axonal projection defects occur in the posteriortritocerebrum. These axonal projection defects are not due todeletions in the affected neuromere. The neural progenitorcells, which give rise to the affected tritocerebral cells, arepresent and correctly located in the mutant domain. Further-more, the postmitotic progeny of these progenitors, the cellsthat are fated to become neurons in the wild type, are alsogenerated and correctly located in the mutant tritocerebraldomain. Indeed, they remain present and correctly located inthe mutant embryonic brain throughout subsequent embryo-genesis and do not appear to be eliminated by cell death.
Figure 1. Adult brains of Drosophila and mouse. Coronal sections. The Drosophila brain is shown within the head and associated withthe compound eyes. The section of the mouse brain is stained with cresyl violet and is at 21.50 bregman.
Review articles
678 BioEssays 21.8
However, these postmitotic cells do not extend axons ordendrites and are not contacted by axons from other parts ofthe brain. This results in axonal patterning defects includingloss of the tritocerebral commissure and reduced or absentlongitudinal pathways between the supra- and subesopha-geal ganglia, which are due to cell-autonomous and cell-nonautonomous effects. Furthermore, they seem to remain inan undifferentiated state and do not express any of thenumerous neuronal molecular markers that positionallyequivalent neuronal cells express in the wild type (Fig. 3).This indicates that the labial mutant cells in the brain do notacquire any form of neuronal identity. In contrast to theabsence of neuronal cell fate in the labial mutant domain, thegeneration of glial cells within this mutant domain appears tobe unaffected. Comparable effects are seen in Deformed
mutants in the corresponding mandibular/anterior maxillarydomain but not in other homeotic gene mutants. Thus, theexpression of the homeotic genes labial and Deformed isnecessary for neurons, but not glia, to adopt their properdifferentiated cell fate in the embryonic brain indicating thatappropriate homeotic gene action is essential for the establish-ment of regionalized neuronal identity in the Drosophila brain.
Hox genes and posterior brain developmentin vertebratesVertebrate Hox genes are structurally and functionally homolo-gous to the homeotic genes of Drosophila and have beenidentified in a wide range of species. In mammals there are 13paralogous groups of Hox genes, which are organized in fourchromosomal clusters called Hox A–Hox D. Only the B cluster
Figure 2. Conserved anteroposterior order of geneexpression in embryonic brain development. Sche-matic diagram of HOM/Hox and otd/Otx gene expres-sion patterns in the developing CNS of Drosophila andmouse. Expression domains are color coded. (Top)Gene expression in embryonic stage 14 DrosophilaCNS. Borders of the protocerebral (b1), deutocerebral(b2), tritocerebral (b3), mandibular (s1), maxillary (s2),labial (s3), and ventral nerve cord neuromeres are indi-cated by vertical lines. In contrast to the other homeoticgenes, pb is only expressed in small segmentally re-peated groups of neuronal cells; this difference is indi-cated by a striped bar to denote the pb expressiondomain. (Bottom) Gene expression in embryonic day9.5–12.5 mouse CNS based on References 15–17.Borders of the telencephalon (T), diencephalon (D),mesencephalon (M), and rhombomeres are indicatedby vertical lines. (Modified according to Ref. 63.)
Figure 3. Comparable brain phenotypes in lab/Hox1 loss of function mutants in Drosophila andmouse. (Left) Simplified scheme of the deutocerebral(b2), tritocerebral (b3), and mandibular (s1) neuro-meres of the Drosophila brain. In the wild type (wt),cells in the posterior tritocerebrum express lab (blue)and also express the neuron-specific marker ELAVand the cell adhesion molecule FasII. In the lab nullmutant (lab-/-), cells in the mutant domain are presentbut do not extend axons and fail to express theneuron-specific marker ELAV and the cell adhesionmolecule FasII indicating a total loss of neuronalidentity. Axons from other parts of the brain avoid themutant domain. (Right) Simplified scheme of rhombo-meres r1-r5 of the mouse hindbrain. In the wild type(wt), cells in r4 co-express Hoxa1 and Hoxb1 and also express the r4-specific marker EphA2. In the Hoxa1-/-; Hoxb1-/- doublehomozygous mutant, cells in r4 are present but the r4-specific marker EphA2 fails to be activated in r4 indicating the presence of aterritory between r3 and r5 with an unknown identity. The double mutant also exhibits multiple defects in the motor neuron axonalprojections; facial motor neurons are scarce and exit randomly from the neural tube.
Review articles
BioEssays 21.8 679

comprises homologs of all of the Drosophila homeotic genes.As in the case of Drosophila, spatial and temporal colinearityalso holds for the vertebrate Hox genes, and more posterioracting genes impose their developmental specificities onanterior acting genes, a fact termed ‘‘posterior preva-lence.’’(10,13,14)
In the developing vertebrate hindbrain, Hox gene expres-sion precedes rhombomere formation and becomes progres-sively restricted during subsequent embryogenesis. The Hoxgene expression patterns form an ordered set of domainswith a two-rhombomere periodicity, and the genes locatedsequentially 38 in the cluster are positioned successively in ananterior direction along the anteroposterior axis coincidingwith rhombomeric borders. This relative anteroposterior orderof Hox gene expression in the developing hindbrain of allvertebrates, including man, is virtually identical to the relativeanteroposterior order of homeotic gene expression in theDrosophila CNS (Fig. 2).(11,15–19)
Single loss-of-function mutations in the mouse demon-strate that the primary site of action of Hox genes generallymaps to their anterior-most expression domain. However,mutant phenotypes of particular paralogs differ markedlysuggesting synergy or functional compensation among paralo-gous Hox genes.(20,21) This is most evident for the labialhomologs Hoxa1 and Hoxb1.(22,23) During gastrulation, Hoxa1and Hoxb1 are activated in the neural ectoderm and byheadfold stage (7.5–7.75 dpc) their expression patterns havereached a sharp anterior boundary coinciding with the pre-sumptive rhombomere 3/4 (r3/r4) border. During rhombo-mere boundary formation, anterior r4 expression of Hoxa1does not persist whereas Hoxb1 expression is maintained inr4 at high levels.(16,17) Functional inactivation of Hoxa1 resultsin segmentation defects leading to a reduced size of r4 andr5, defects in motor neuron axonal projections, and defects inthe location of cell bodies of the trigeminal and facial/vestibulo-acoustic nerve, but does not alter normal r4 identity.(21–23) Incontrast, loss of Hoxb1 has no influence on the size of r4 butfails to maintain normal r4 identity. Thus, r4-specific facialbranchiomotor and contralateral vestibulo-acoustic efferentneurons are incorrectly specified and fail to migrate into theirproper positions resulting in an atypical motor nucleus andloss of the facial nerve.(24,25) In addition, Hoxb1 loss-of-function causes a partial transformation of r4 into an r2identity,(24,25) whereas Hoxa1 gain-of-function causes a par-tial transformation of r2 into an r4 identity.(26,27)
In Hoxa1-/-; Hoxb1-/- double loss-of-functions mutants, r4shows a reduced size as in Hoxa1 homozygous mutant mice,but r4-specific markers fail to be activated indicating thepresence of a territory between r3 and r5 with an unknownidentity. Hoxa1-/-; Hoxb1-/- double homozygotes also exhibita reduced number of facial motor neurons, which appear toexit randomly from the neural tube without fasciculatingtowards distinct exit points (Fig. 3).(22,23) Together these
results suggest that in addition to their individual roles, Hoxa1and Hoxb1 synergize in the specification of r4 identity and inthe patterning of cranial nerves during vertebrate hindbraindevelopment. This is very similar to the role of the Hoxa1/Hoxb1 homolog labial in specifying segmental neuronalidentity during embryonic brain development of Drosophila.
The otd gene and anterior brain developmentin DrosophilaThe cephalic gap gene otd encodes a homeodomain transcrip-tion factor that is required for head development and segmen-tal patterning in Drosophila. The first otd transcripts appear atearly blastoderm stages covering a broad circumferentialstripe in the anterior region of the embryo, that includes theanlagen of several cephalic segments as judged from blasto-derm fate maps.(28–31) During gastrulation, expression of otdbecomes progressively restricted to the procephalic region. Asecond domain of otd expression appears at the ventralmidline at embryonic stage 9–10 in a longitudinal stripe ofmesectodermal cells that will generate a mixed population ofventral nerve cord neurons and glia.(28–30)
During neuroectoderm formation, the procephalic otd ex-pression domain resolves into a smaller patch correspondingto most of the protocerebral and an adjacent part of thedeutocerebral brain anlagen. All of the neuroblasts thatsegregate from this region transiently express otd. Duringsubsequent brain regionalization and patterning, neuronalotd expression in neurons is observed throughout most of theprotocerebrum and adjacent deutocerebrum (Fig. 2). Interest-ingly, otd expression is not observed in the anteriormostprotocerebral region.(32,33)
This spatially restricted expression domain correlates wellwith the site of action of otd in patterning the rostral brain ofDrosophila.(32,33) Homozygous null mutation of otd results inthe deletion of the protocerebral anlage due to defectiveneuroectoderm specification in these regions. Most protocere-bral neuroblasts and some deutocerebral neuroblasts areabsent in the mutant, and in consequence a dramaticallyreduced embryonic brain is formed (Fig. 4).(33) This regional-ized absence of brain neuroblasts in otd mutants correlateswith the loss or reduction in expression of the lethal of scutegene in the otd mutant domain; the proneural gene lethal ofscute is thought to be required for neurectodermal cells toacquire the competence to form neuroblasts.(32) In addition todefects in the anterior brain, mutational inactivation of otdalso affects the development of visual and mechanosensorystructures and causes midline defects in the ventral nervecord resulting in deranged connectives and fused commis-sural axon tracts.(28,32,34)
Evidence for differential timing requirements in the actionof otd in anterior brain vs. ventral CNS development isprovided by genetic rescue experiments. Ubiquitous overex-pression of otd in a null mutant background is able to restore
Review articles
680 BioEssays 21.8

both the brain defects and the defects in the ventral nervecord.(35) However, the time window during which otd expres-sion is sufficient to rescue the brain defect is clearly aftergastrulation (embryonic stage 7–8), but before neuroblastdelamination. The early blastoderm expression of otd is notrequired for brain development. To rescue the ventral nervecord defects, otd must be expressed even later (embryonicstage 10–11). This supports the idea, that depending on thespatio-temporal pattern of its expression, the otd gene fulfillsdifferent functions during embryonic development.
Ubiquitous overexpression of otd in a wild type back-ground results in the generation of ectopic or transformedneuronal structures such as dramatically increased brainlobes, ectopic head ganglionic structures, as well as fusedand enlarged ventral ganglia.(35) Interestingly, some of theseectopic head ganglia and enlarged ventral ganglia showectopic expression of the brain specific homeobox gene,(36)
which is normally only expressed in the developing protocere-brum. This suggests that otd expression might be able toinduce a partial protocerebral identity of these ectopic struc-tures and, thus, supports the notion that the otd gene, like thehomeotic genes, might be able to specify the identity of CNSstructures.
The Otx genes and anterior brain developmentin vertebratesBased on homology between homeobox sequences, ho-mologs of the Drosophila otd gene have been isolated invarious vertebrates including zebrafish, mouse, and hu-man.(37) In mouse, the two vertebrate homologs, Otx1 andOtx2, are expressed in the developing head and brain innested and overlapping domains. Otx1 transcripts first ap-pear at approximately 8 dpc, whereas Otx2 expression canbe detected earlier at the prestreak stage (5.5 dpc) within thevisceral endoderm and throughout the epiblast from which alltissues of the developing mouse derive. At 7–7.5 dpc thedomain of Otx2 expression becomes restricted to the anteriorregion of the embryo, which includes the neuroectoderm ofthe prosencephalon and mesencephalon, as well as anteriormesoendodermal regions.(38,39) By 9.5–10 dpc, the Otx2 geneis expressed in a broad domain that spans the developingforebrain and midbrain with its posterior expression boundarycorresponding to the presumptive midbrain-hindbrain bound-ary (Fig. 2).(40) Otx1 expression is nested within this Otx2expression domain (Fig. 2) and subsequently becomes spa-tially and temporally restricted to the developing cortex andcerebellum.(41) Interestingly, the domain of Otx2 expressiondoes not include the most anterior brain region comprisingthe optic chiasm and optic recess.(38,39)
These dynamic and spatially restricted expression do-mains correlate with the sites of action of Otx1 and Otx2 inrostral brain patterning. Heterozygous Otx2 mutations showbrain abnormalities and craniofacial malformations which are
reminiscent of otocephaly,(42) whereas homozygous Otx2mutation leads to the deletion of brain regions anterior to r3(Fig. 4) due to defective anterior neuroectoderm specifica-tion.(42–44) As has been shown recently using murine chimericembryos(45) and a knock-in mouse model in which Otx2 isreplaced with Otx1,(46) these defects can be related to afunctional role of Otx2 in the visceral endoderm at the onset ofgastrulation where it is required for anterior neural plateinduction. In contrast to the early functional requirement ofOtx2, no apparent defects in early brain development arecaused by Otx1 mutation. However, later in development lossof function of Otx1 affects telencephalic temporal and perirhi-
Figure 4. Conserved expression and function of the otd/Otx2 genes in embryonic brain development. Schematicdiagram of otd and Otx2 gene expression patterns and otd andOtx2 mutant phenotypes in the developing CNS of Drosophilaand mouse. (Top) otd gene expression in the wild type (wt) andbrain phenotype of otd null mutant in embryonic stage 14Drosophila CNS. Borders of the protocerebral (b1), deutocere-bral (b2), tritocerebral (b3), mandibular (s1), maxillary (s2),labial (s3), as well as some of the ventral nerve cord neuro-meres are indicated by vertical lines. (Bottom) Otx2 geneexpression in the wild type (wt) and brain phenotype of Otx2homozygous null mutant in embryonic day 12.5 mouse CNSbased on References 38–40. Borders of the telencephalon(T), diencephalon (D), mesencephalon (M), and rhombo-meres are indicated by vertical lines.
Review articles
BioEssays 21.8 681

nal areas, the hippocampus, and mesencephalic as well ascerebellar brain structures and also causes epilepsy.(47,48) Inaddition, the development of eye and inner ear is impaired.
The Otx1 mutant phenotype and the overlapping expres-sion domain of Otx1 with that of Otx2 suggest synergy orfunctional compensation among the Otx1/2 genes compa-rable to paralogous Hox gene function. This is indeed thecase since Otx1 and Otx2 cooperate to specify correct braindevelopment in a dosage dependent manner. Thus, doubleheterozygous embryos (Otx11/-; Otx21/-) show a severereduction of both the mesencephalon and the posteriordiencephalon with an expansion of r1.(49) Mice carrying onlyone functional copy of Otx2 (Otx1-/-; Otx21/-) show arepatterning of the developing brain in that the caudaldiencephalon and mesencephalon are molecularly and mor-phologically transformed into an enlarged metencephalon. Inaddition, in these mice a structure comparable to a midbrain-hindbrain-boundary is positioned in a region corresponding tothe presumptive prosomere 2, and the dorsal telencephalonacquires mesencephalic molecular features.(50) These datademonstrate essential roles of the Otx1/2 genes in morpho-genesis and pattern formation in anterior brain developmentin vertebrates, and in this sense are clearly reminiscent of therole of the otd gene during anterior embryonic brain develop-ment in Drosophila.
Functional conservation of otd/Otx genesin brain developmentTaken together, investigations of early morphogenesis andpatterning in the embryonic brains of Drosophila and mousereveal developmental mechanisms that are strikingly similar,and suggest an evolutionary conservation of Hox and otd/Otxgenes in embryonic brain development that extends beyondgene structure to patterned expression and function. A directexperimental demonstration of this functional conservationhas recently been carried out for otd/Otx genes in patterningof the rostral brain.(35,51) In these experiments, the humanOTX1 and OTX2 genes have been introduced and overex-pressed in Drosophila otd null mutants(35) and, conversely,the murine Otx1 gene has been replaced with the Drosophilaotd gene.(51)
In Drosophila, the human OTX genes, like the endog-enous otd gene, are able to rescue the brain defects as wellas the midline defects observed in otd null mutants. Inaddition, ubiquitous overexpression in wild type embryosresults in ectopic neural structures, regardless whether hu-man OTX or fly otd are overexpressed.(35) Similarly, in themouse, the Drosophila otd gene rescues many of the braindefects seen in the Otx1-/- mutant; a full rescue of corticogen-esis and epilepsy, as well as a partial rescue of eye defectsand other brain abnormalities (but not inner ear defects) isobserved.(51) Moreover, Drosophila otd is able to partiallyreplace Otx1 in its cooperative interactions with Otx2 for
correct brain patterning. Thus, the anatomical and molecularbrain defects of Otx1-/-; Otx21/- embryos were rescued byDrosophila otd in a dose-dependent manner with high effi-ciency for telencephalic structures, intermediate efficiency forthe posterior diencephalon and poor efficiency for the mesen-cephalon.(51)
Vertebrate and Drosophila otd/Otx gene products sharestructural homology, which is confined mainly to the homeodo-main suggesting that this domain is crucial in selecting at avery high stringency the same target sequence(s). Thisimplies that the extensive functional equivalence of theotd/Otx genes may be due to conserved developmentalgenetic circuits with common functional features that arecontrolled by the homeodomain. However, although thefunctional equivalence of the otd/Otx genes in brain develop-ment is extensive, it is not complete. For example, the otdgene does not complement Otx1 function in inner ear develop-ment nor does it complement the dosage dependent Otx1function in establishing midbrain structures including themidbrain-hinbrain-boundary.(51) These differences in the func-tional features of the two genes may result predominantlyfrom regulatory changes due to coding sequences outsidethe homeobox which for Drosophila otd and vertebrate Otxgenes differ markedly.(39)
Several other examples have recently been reported onthe evolutionary conservation of developmental regulatorygenes in vertebrates and invertebrates at the structural,functional, and regulatory levels. A striking example are theeyeless/Pax6 genes and the regulatory elements in thelabial/Hoxb1 enhancer regions, which are interchangeableacross different phyla.(52–55) The functional equivalence of theotd/Otx genes now demonstrates for the first time that theconcept of conserved gene action across different phyla alsoapplies to embryonic brain development.
ConclusionsRecent investigations on the mechanisms controlling insectand mammalian brain development show that homologousregulatory genes belonging to the Hox and otd/Otx genefamilies control regionalization, patterning, and identity duringearly embryonic brain development in a strikingly similar way.Thus, Hox genes are expressed co-linearly in the developingcaudal brain of Drosophila and mouse and are required forsegmental neuronal identity. The otd/Otx genes are ex-pressed in the anterior brain of Drosophila and mouse andare required for entire rostral brain formation. These resultssuggest that the genetic programs underlying embryonicbrain development are evolutionarily conserved and providefurther evidence for the idea that an extensive region of theanterior-posterior axis of the insect and vertebrate body plan,including the brain, is homologous in a developmental geneticsense.(4,37,56–58)
Review articles
682 BioEssays 21.8

This implies that the insect and vertebrate brain derived bydescent from a common ancestral brain and it also impliesthat a common molecular ground plan of brain developmenthad been established before the diversification of the protos-tome and deuterostome lineages.(56,58,59) In this sense, theconserved genetic mechanisms controlling insect and verte-brate brain development might reflect the previously existinggenetic circuits acting in the development of a commonancestral brain. Beyond the identification of these ancestralfunctional features, it will be interesting to determine if someconserved structural features previously existing in a com-mon ancestral brain are still present in both insect andvertebrate brain development. For example, recent datasuggests that the midbrain/isthmic region may not only bevertebrate-specific implicating a tripartite organization of theancestral chordate brain.(60) It will be of particular interest todetermine if the Drosophila brain also contains a crypticmidbrain/isthmic region or if such a structure is induceable.(61)
Moreover, it will now be important to identify which othergenes are involved in the molecular genetic circuits for braindevelopment and to elucidate their structural organizationand functional interactions. The identification of downstreamtarget genes through manipulative genetics and functionalgenomic techniques and the dissection of gene action throughintergenic and intragenic chimeras should allow us to uncoverthe specificity and structural implications of genetic circuitswith common regulatory and functional features both ininsects and vertebrates. But most exciting will be the attemptto decode the genetic circuits, which have uncommon molecu-lar features in insects and vertebrates(62) and which contrib-ute to the obvious differences between the fly’s brain and ourown.
AcknowledgmentsWe thank Antonio Simeone, Beate Hartmann, Martin Herzig,and Ronny Leemans for comments on the manuscript. Wealso thank Robert Williams for permission to use data fromthe Mouse Brain Atlas Project (http://mickey.utmem.edu/atlas/atlas.html).
References1. Therianos S, Leuzinger S, Hirth F, Goodman CS, Reichert H. Embryonic
development of the Drosophila brain: formation of commissural anddescending pathways. Development 1995;121:3849–3860.
2. Younossi-Hartenstein A, Nassif C, Green P, Hartenstein V. Early neurogen-esis of the Drosophila brain. J Comp Neurol 1996;370:313–329.
3. Campos-Ortega JA, Hartenstein V. The embryonic development of Dro-sophila Melanogaster. Berlin: Springer-Verlag; 1997.
4. Reichert H, Boyan G. Building a brain: developmental insights in insects.Trends Neurosci 1997;20:258–264.
5. Jurgens G, Hartenstein V. The terminal regions of the body pattern. In:Bate M, Martinez-Arias A, editors. The development of Drosophila. NewYork: Cold Spring Harbor Laboratory Press; 1993. p 687–746.
6. Doe CQ, Skeath JB. Neurogenesis in the insect central nervous system.Curr Opin Neurobiol 1996;6:18–24.
7. Lumsden A, Krumlauf R. Patterning the vertebrate neuraxis. Science1996;274:1109–1115.
8. Rubenstein JLR, Martinez S, Shimamura K, Puelles L. The embryonicvertebrate forebrain: the prosomeric model. Science 1994;266:578–580.
9. Rubenstein JLR, Shimamura K, Martinez S, Puelles L. Regionalization ofthe prosencephalic neural plate. Annu Rev Neurosci 1998;21:445–477.
10. Duboule D, Morata G. Colinearity and functional hierarchy among genesof the homeotic complexes. Trends Genet 1994;10:358–364.
11. Hirth F, Hartmann B, Reichert H. Homeotic gene action in embryonic braindevelopment of Drosophila. Development 1998;125:1579–1589.
12. Kaufman TC, Seeger MA, Olsen G. Molecular and genetic organization ofthe antennapedia gene complex of Drosophila melanogaster. Adv Genet1990;27:309–362.
13. Ruddle FH, Bartels JL, Bentley KL, Kappen C, Murtha MT, Pendleton JW.Evolution of Hox genes. Annu Rev Genet 1994;28:423–442.
14. Capecchi MR. The role of Hox genes in hindbrain development. In: CowanWM, Jessell TM, Zipursky SL, editors. Molecular and cellular approachesto neural development. Oxford: Oxford University Press; 1997. p 334–355.
15. Graham A, Papalopulu N, Krumlauf R. The murine and Drosophilahomeobox gene complexes have common features of organization andexpression. Cell 1989;57:367–378.
16. Wilkinson DG, Bhatt S, Cook M, Boncinelli E, Krumlauf R. Segmentalexpression of Hox-2 homeobox-containing genes in the developingmouse hindbrain. Nature 1989;341:405–409.
17. Hunt P, Gulisano M, Cook M, Sham M-H, Faiella A, Wilkinson D, BoncinelliE, Krumlauf R. A distinct Hox code for the branchial region of thevertebrate head. Nature 1991;353:861–864.
18. Vieille-Grosjean I, Hunt P, Gulisano M, Boncinelli E, Thorogood P. Bran-chial HOX gene expression and human craniofacial development. DevBiol 1997;183:49–60.
19. Prince VE, Moens CB, Kimmel CB, Ho RK. Zebrafish hox genes:expression in the hindbrain region of wild-type and mutants of thesegmentation gene, valentino. Development 1998;125:393–406.
20. Maconochie M, Nonchev S, Morrison A, Krumlauf R. Paralogous Hoxgenes: function and regulation. Annu Rev Genet 1996;30:529–556.
21. Rijli FM, Gavalas A, Chambon P. Segmentation and specification in thebranchial region of the head: the role of the Hox selector genes. Int J DevBiol 1998;42:393–401.
22. Studer M, Gavalas A, Marshall H, Ariza-McNaughton L, Rijli FM, ChambonP, Krumlauf R. Genetic interactions between Hoxa1 and Hoxb1 reveal newroles in regulation of early hindbrain patterning. Development 1998;125:1025–1036.
23. Gavalas A, Studer M, Lumsden A, Rijli FM, Krumlauf R, Chambon P. Hoxa1and Hoxb1 synergize in patterning the hindbrain, cranial nerves andsecond pharyngeal arch. Development 1998;125:1123–1136.
24. Goddard JM, Rossel M, Manley NR, Capecchi MR. Mice with targeteddisruption of Hoxb1 fail to form the motor nucleus of the VIIth nerve.Development 1996;122:3217–3228.
25. Studer M, Lumsden A, Ariza-McNaughton L, Bradley A, Krumlauf R.Altered segmental identity and abnormal migration of motor neurons inmice lacking Hoxb1. Nature 1996;384:630–634.
26. Alexandre D, Clarke JDW, Oxtoby E, Yan Y-L, Jowett T, Holder N. Ectopicexpression of Hoxa1 in the zebrafish alters the fate of the mandibular archneural crest and phenocopies a retinoic acid-induced phenotype. Devel-opment 1996;122:735–746.
27. Zhang M, Kim H-J, Marshall H, Gendron-Maguire M, Lucas AD, Baron A,Gudas LJ, Krumlauf R, Grippo JF. Ectopic Hoxa1 induces rhombomeretransformation in mouse hindbrain. Development 1994;120:2431–2442.
28. Finkelstein R, Smouse D, Capaci TM, Spradling AC, Perrimon N. Theorthodenticle gene encodes a novel homeodomain protein involved in thedevelopment of the Drosophila nervous system and ocellar visual struc-tures. Genes Dev. 1990;4:1516–1527.
29. Finkelstein R, Perrimon N. The orthodenticle gene is regulated by bicoidand torso and specifies Drosophila head development. Nature 1990;346:485–488.
30. Cohen SM, Jurgens G. Mediation of Drosophila head development bygap-like segmentation genes. Nature 1990;346:482–485.
31. Jurgens G, Lehmann R, Schardin M, Nusslein-Volhard C. Segmentalorganization of the head in the embryo of Drosophila melanogaster. Ablastoderm fate map of the cuticle structures of the larval head. Roux’sArch Dev Biol 1986;195:359–377.
Review articles
BioEssays 21.8 683

32. Younossi-Hartenstein A, Green P, Liaw G-W, Rudolph K, Lengyel J,Hartenstein V. Control of early neurogenesis of the Drosophila brain by thehead gap genes tll, otd, ems, and btd. Dev. Biol. 1997;182:270–283.
33. Hirth F, Therianos S, Loop T, Gehring WJ, Reichert H, Furukubo-TokunagaK. Developmental defects in brain segmentation caused by mutations ofthe homeobox genes orthodenticle and empty spiracles in Drosophila.Neuron 1995;15:769–778.
34. Schmidt-Ott U, Gonzalez-Gaitan M, Jackle H, Technau G. Number,identity, and sequence of the Drosophila head segments as revealed byneural elements and their deletion patterns in mutants. Proc Natl Acad SciUSA 1994;91:8363–8367.
35. Leuzinger S, Hirth F, Gerlich D, Acampora D, Simeone A, Gehring WJ,Finkelstein R, Furukubo-Tokunaga K, Reichert H. Equivalence of the flyorthodenticle gene and the human OTX genes in embryonic braindevelopment of Drosophila. Development 1998;125:1703–1710.
36. Jones B, McGinnis W. A new Drosophila homeobox gene, bsh, isexpressed in a subset of brain cells during embryogenesis. Development1993;117:793–806.
37. Simeone A. Otx1 and Otx2 in the development and evolution of themammalian brain. EMBO J 1998;17:6790–6798.
38. Simeone A, Acampora D, Gulisano M, Stornaiuolo A, Boncinelli E. Nestedexpression domains of four homeobox genes in the developing rostralbrain. Nature 1992;358:687–690.
39. Simeone A, Acampora D, Mallamaci A, Stornaiuolo A, D’Apice MR, NigroV, Boncinelli E. A vertebrate gene related to orthodenticle contains ahomeodomain of the bicoid class and demarcates anterior neuroecto-derm in the gastrulating mouse embryo. EMBO J. 1992;12:2735–2747.
40. Millet S, Bloch-Gallego E, Simeone A, Alvarado-Mallart R-M. The caudallimit of Otx2 gene expression as a marker of the midbrain/hindbrainboundary: a study using in situ hybridisation and chick/quail homotopicgrafts. Development 1996;122:3785–3797.
41. Frantz GD, Weimann JM, Levin ME, McConnell S. Otx1 and Otx2 definelayers and regions in developing cerebral cortex and cerebellum. JNeurosci 1994;14:5725–5740.
42. Matsuo I, Kuratani S, Kimura C, Takeda N, Aizawa S. Mouse Otx2functions in the formation and patterning of rostral head. Genes Dev1995;9:2646–2658.
43. Acampora D, Mazan S, Lallemand Y, Avantaggiato V, Maury M, SimeoneA, Brulet, P. Forebrain and midbrain regions are deleted in Otx2-/- mutantsdue to a defective anterior neuroectoderm specification during gastrula-tion. Development 1995;121:3279–3290.
44. Ang S-L, Jin O, Rhinn M, Daigle N, Stevenson L, Rossant J. A targetedmouse Otx2 mutation leads to severe defects in gastrulation and forma-tion of axial mesoderm and to deletion of rostral brain. Development1996;122:243–252.
45. Rhinn M, Dierich A, Shawlot W, Behringer RR, Le Meur M, Ang S-L.Sequential roles for Otx2 in visceral endoderm and neuroectoderm forforebrain and midbrain induction and specification. Development 1998;125:845–856.
46. Acampora D, Avantaggiato V, Tuorto F, Briata P, Corte G, Simeone A.Visceral endoderm-restricted translation of Otx1 mediates recovery ofOtx2 requirements for specification of anterior neural plate and normalgastrulation. Development 1998;125:5091–5104.
47. Acampora D, Mazan S, Avantaggiato V, Barone P, Tuorto F, Lallemand Y,Brulet P, Simeone A. Epilepsy and brain abnormalities in mice lacking theOtx1 gene. Nat Genet 1996;14:218–222.
48. Suda Y, Matsuo I, Kuratani S, Aizawa S. Otx1 function overlaps with Otx2in development of mouse forebrain and midbrain. Genes Cells 1996;1:1031–1044.
49. Suda Y, Matsuo I, Aizawa S. Cooperation between Otx1 and Otx2 genes indevelopmental patterning the rostral brain. Mech Dev 1997;69:125–141.
50. Acampora D, Avantaggiato V, Tuorto F, Simeone A. Genetic control ofbrain morphogenesis through Otx gene dosage requirement. Develop-ment 1997;124:3639–3650.
51. Acampora D, Avantaggiato V, Tuorto F, Barone P, Reichert H, Finkelstein R,Simeone A. Murine Otx1 and Drosophila otd genes share conservedgenetic functions required in invertebrate and vertebrate brain develop-ment. Development 1998;125:1691–1702.
52. Halder G, Callaerts P, Gehring WJ. Induction of ectopic eyes by targetedexpression of the eyeless gene in Drosophila. Science 1995;267:1788–1792.
53. Popperl H, Bienz M, Studer M, Chan S-K, Aparicio S, Brenner S, Mann RS,Krumlauf R. Segmental expression of Hoxb1 is controlled by a highlyconserved autoregulatory loop dependent upon exd/pbx. Cell 1995;81:1031–1042.
54. Chan S-K, Popperl H, Krumlauf R, Mann RS. An extradenticle-inducedconformational change in a HOX protein overcomes an inhibitory functionof the conserved hexapeptide motif. EMBO J. 1996;15:2476–2487.
55. Chan S-K, Ryoo H-D, Gould A, Krumlauf R, Mann RS. Switching the in vivospecificity of a minimal Hox-responsive element. Development 1997;124:2007–2014.
56. De Robertis EM, Sasai Y. A common plan for dorsoventral patterning inBilateria. Nature 1996;380:37–40.
57. Thor S. The genetics of brain development: conserved programs in fliesand mice. Neuron 1995;15:975–977.
58. Arendt D, Nubler-Jung K. Common ground plans in early brain develop-ment in mice and flies. BioEssays 1996;18:255–259.
59. De Robertis EM. The ancestry of segmentation. Nature 1997;387:25–26.60. Wada H, Saiga H, Satoh N, Holland PWH. Tripartite organization of the
ancestral chordate brain and the antiquity of placodes: insights fromascidian Pax-2/5/8, Hox and Otx genes. Development 1998;125:1113–1122.
61. Ang S-L. The brain organization. Nature 1996;380:25–26.62. Goldowitz D, Hamre K. The cells and molecules that make a cerebellum.
Trends Neurosci 1998;21:375–382.63. Sharman AC, Brand M. Evolution and homology of the nervous system:
cross-phylum rescues of otd/Otx genes. Trends Genet 1998;14:211–214.
Review articles
684 BioEssays 21.8