computational simulation of fracture repair in stem cell
TRANSCRIPT

University of Calgary
PRISM: University of Calgary's Digital Repository
Graduate Studies The Vault: Electronic Theses and Dissertations
2012-09-28
Computational Simulation of Fracture Repair in Stem
Cell Seeded Defects under Different Mechanical
Loading
Nasr, Saghar
Nasr, S. (2012). Computational Simulation of Fracture Repair in Stem Cell Seeded Defects under
Different Mechanical Loading (Unpublished master's thesis). University of Calgary, Calgary, AB.
doi:10.11575/PRISM/25397
http://hdl.handle.net/11023/242
master thesis
University of Calgary graduate students retain copyright ownership and moral rights for their
thesis. You may use this material in any way that is permitted by the Copyright Act or through
licensing that has been assigned to the document. For uses that are not allowable under
copyright legislation or licensing, you are required to seek permission.
Downloaded from PRISM: https://prism.ucalgary.ca

UNIVERSITY OF CALGARY
Computational Simulation of Fracture Repair in Stem Cell Seeded Defects under Different
Mechanical Loading
by
Saghar Nasr
A THESIS
SUBMITTED TO THE FACULTY OF GRADUATE STUDIES
IN PARTIAL FULFILMENT OF THE REQUIREMENTS FOR THE
DEGREE OF MASTER OF SCIENCE
DEPARTMENT OF MECHANICAL AND MANUFACTURING ENGINEERING
CALGARY, ALBERTA
September 2012
© Saghar Nasr 2012

i
Abstract
Mechanical factors play a key role in regulation of tissue regeneration during skeletal healing,
but the underlying mechanisms are not fully understood. The objective of the current study was
to explore the role of mechanical factors on tissue differentiation during fracture healing, using a
biphasic mechanoregulatory algorithm. The first specific objective was to investigate the effect
of mechanical loading on a stem cell seeded collagenous scaffold in a one-dimensional confined
compression configuration. Both empirical and computational data suggest that mechanical
stimulation of the scaffold may be an effective way to initiate differentiation pathways prior to
implantation for tissue engineering applications. The second objective was to predict the
development of differentiated tissues in a tibia burr-hole fracture murine model with
computational mechanoregulatory algorithms. The computational and experimental studies will
be used simultaneously in future studies to further develop mechanoregulatory models with a
more robust quantitative base.

ii
Acknowledgements
Blessed Be His Name
First I must thank Dr. Neil A. Duncan for his tremendous dedication and guidance throughout
the duration of this investigation. Without his support, this work would not have been possible.
I want to thank my amazing parents, sister, brother, grandparents, and my uncle, Shahriar, for
their love and unwavering support. My thanks are also extended to Drs. Leonard Hills and
Mojtaba Kazemi for their encouragements and valuable comments.
This research was supported by a team grant from the Canadian Institutes of Health Research in
Skeletal Regenerative Medicine, the Natural Sciences and Engineering Research Council and the
Canada Research Chair in Orthopaedic Bioengineering.

iii
Table of Contents
Abstract ........................................................................................................................... i Acknowledgements ......................................................................................................... ii
Table of Contents ........................................................................................................... iii List of Tables ................................................................................................................. vi
List of Figures and Illustrations .................................................................................... viii List of Symbols, Abbreviations and Nomenclature ....................................................... xix
CHAPTER ONE: INTRODUCTION .............................................................................. 1 1.1 Background and motivation ................................................................................... 1
1.2 Bone ...................................................................................................................... 4 1.2.1 Bone repair .................................................................................................... 9
1.2.1.1 Biological stages of fracture healing ................................................... 10 1.2.1.2 Source of progenitor cells for fracture healing .................................... 13
1.3 Problem statement and rationale .......................................................................... 14 1.4 Thesis objectives ................................................................................................. 22
1.5 Thesis overview .................................................................................................. 26
CHAPTER TWO: MECHANOREGULATION ALGORITHMS OF TISSUE
DIFFERENTIATION IN BONE .......................................................................... 28 2.1 Pauwels theory .................................................................................................... 29
2.2 Interfragmentary strain theory ............................................................................. 31 2.3 Mechanostat theory, (Frost, 1987) ....................................................................... 32
2.4 Computational simulations of tissue differentiation ............................................. 34 2.4.1 Single solid phase model (Carter‟s theory, 1988) ......................................... 34
2.4.2 Single solid phase model (Claes and Heigele, 1999) .................................... 37 2.4.3 Single solid phase model (Gardner et al., 2000) ........................................... 38
2.4.4 Biphasic model (Kuiper et al., 1996-2000) .................................................. 39 2.4.5 Biphasic model (Prendergast et al., 1997). ................................................... 41
2.4.6 Biphasic model (Sandino and Lacroix, 2011)............................................... 43 2.4.7 Models with biological factors ..................................................................... 45
CHAPTER THREE: DEVELOPMENT AND VERIFICATION OF THE FINITE
ELEMENT MODEL ............................................................................................ 61
3.1 Bone mechanics .................................................................................................. 61 3.1.1.1 Osteoporotic bone mechanics ............................................................. 65
3.1.2 Bone structure and optimisation .................................................................. 69 3.1.3 Mechanical behaviour of cortical bone ........................................................ 72
3.1.4 Mechanical behaviour of cancellous bone .................................................... 75 3.2 Soft tissue biphasic theory ................................................................................... 76
3.2.1 Kinematics .................................................................................................. 78 3.2.2 Conservation of mass .................................................................................. 80
3.2.3 Conservation of linear momentum ............................................................... 81 3.3 Finite element model of mechanoregulation ........................................................ 85

iv
3.3.1 Adaptive mechanoregulation algorithm ....................................................... 85 3.3.1.1 User defined subroutine: USDFLD..................................................... 88
3.3.1.2 Smoothing process ............................................................................. 91 3.3.1.3 Diffusion of progenitor cells............................................................... 92
3.4 Verification of the implemented model ................................................................ 99 3.4.1 Results ...................................................................................................... 102
3.4.2 Discussion ................................................................................................. 107 3.5 Fracture healing case studies ............................................................................. 108
3.5.1 An axisymmetric idealized murine model .................................................. 109 3.5.1.1 Results ............................................................................................. 111
3.5.1.2 Discussion ........................................................................................ 113 3.5.2 A 3D idealised model of murine tibia ........................................................ 114
3.5.2.1 Results ............................................................................................. 115 3.5.2.2 Discussion ........................................................................................ 117
3.6 Summary of the computational analyses ............................................................ 119
CHAPTER FOUR: COLLAGENOUS SCAFFOLD UNDER CONFINED COMPRESSION
........................................................................................................................... 122 4.1 Confined compression loading device description.............................................. 122
4.2 Computational modelling of the system ............................................................. 126 4.2.1 The cell base and the top lid ...................................................................... 127
4.2.2 The collagen gel and the porous plug ......................................................... 127 4.2.3 The silicone rings ...................................................................................... 129
4.3 Validation of the computational model against the experimental results ............. 131 4.3.1 The square-cross-section ring .................................................................... 131
4.3.2 The circular-cross-section ring................................................................... 132 4.3.3 The system excluding the collagenous gel ................................................. 134
4.3.4 The system including the collagenous gel .................................................. 136 4.4 Mechanical behaviour of the collagen gel in confined compression: cyclic loading143
4.5 Prediction of tissue differentiation in confined compression .............................. 149 4.6 Summary ........................................................................................................... 154
CHAPTER FIVE: TISSUE DIFFERENTIATION IN A BURR-HOLE FRACTURE
MODEL IN A MURINE TIBIA ......................................................................... 156
5.1 Introduction ....................................................................................................... 157 5.2 Reconstruction of a murine tibia ........................................................................ 158
5.2.1 Importing and preparing the data (ScanIP module) .................................... 158 5.2.2 Image processing (ScanIP module) ............................................................ 158
5.2.3 Creating the volumetric model, assigning material properties and mesh generation
(ScanFE module) ....................................................................................... 163
5.2.4 Convergence study .................................................................................... 167 5.3 Verification of the generated FE model of the intact tibia .................................. 168
5.4 Development of the burr-hole fracture model .................................................... 170 5.4.1 Selection of the decay length model........................................................... 173
5.5 Tissue differentiation predictions within the burr-hole fracture .......................... 178

v
5.5.1 Investigation of axial compression load ..................................................... 182 5.5.2 Influence of cell diffusivity rate ................................................................. 187
5.5.3 Influence of fracture position ..................................................................... 190 5.5.3.1 Different mechanical stimuli with the same cell origins .................... 190
5.5.3.2 Different mechanical stimuli and cell origins .................................... 193 5.5.4 Influence of reduced mechanical properties ............................................... 195
5.5.5 Influence of bending load .......................................................................... 199 5.6 Summary and discussion ................................................................................... 201
CHAPTER SIX: CONCLUSIONS, LIMITATIONS AND FUTURE DIRECTIONS .. 209 6.1 Summary and conclusions ................................................................................. 209
6.2 Limitations ........................................................................................................ 213 6.3 Future directions ................................................................................................ 217
REFERENCES ........................................................................................................... 219
APPENDIX A: USER DEFINED SUBROUTINE: USDFLD ..................................... 231

vi
List of Tables
Table 2.1. Mechanoregulatory stimulus for tissue differentiation (Prendergast et al. 1997). ....... 42
Table 2.2. Biphasic model prediction of tissue differentiation by Sandino and Lacroix (2011)... 44
Table 2.3. Summary of the mechanoregulatory algorithms of musculoskeletal tissue
differentiation. ................................................................................................................... 59
Table 3.1. The quantitative commutative tomography outcomes at the distal woman tibia
(Macdonald et al. 2011). .................................................................................................... 69
Table 3.2. Estimation of normal stress under axial, bending and torsional loads. ....................... 71
Table 3.3. Poroelastic tissue material properties (Isaksson et al. 2006)....................................... 88
Table 3.4. Dependence of material properties on field variable (FV) in the fracture site............. 90
Table 3.5. Applying the smoothing process to the algorithm. ..................................................... 92
Table 3.6. Similarity between mass diffusion and heat transfer equations. ................................. 95
Table 3.7. Element type, poroelastic and thermal properties of the tissues of a human. .............. 96
Table 3.8. Dimensions of the axisymmetric FE model of an ovine tibia. .................................. 101
Table 3.9. Geometry of the murine tibia proximal section (Windahl et al. 1999, Geris et al.
2004). .............................................................................................................................. 110
Table 3.10. Material properties used for the study (Rho, Ashman and Turner 1993, Isaksson
et al. 2006). ..................................................................................................................... 111
Table 4.1. Poroelastic properties of the porous plug and the collagen gel (Isaksson et al.
2006, Tromas et al. 2012). ............................................................................................... 129
Table 4.2. The geometry of the silicone rings. ......................................................................... 130
Table 4.3. The predicted equilibrium strains, fluid velocities and pore pressure at the peak
loading in the top, middle and bottom sample elements. .................................................. 149
Table 4.4. Poroelastic tissue material properties (Isaksson et al. 2006)..................................... 150
Table 5.1. The relationship between the mechanical properties and mass density (Rho et al.
1995). .............................................................................................................................. 165
Table 5.2. The range of greyscale based material properties of the murine tibia. ...................... 166

vii
Table 5.3. Convergence study for three mesh densities. ........................................................... 168
Table 5.4. Elements numbers for different parts of the fracture model. .................................... 171
Table 5.5. Summary of the variables for the parametric studies. .............................................. 180
Table 5.6. Element numbers and material properties used for the current study (Rho et al.
1993, Isaksson et al. 2006). .............................................................................................. 181
Table 5.7. Mechanical properties used for an osteoporotic murine tibia (Li and Aspden
1997b, Lacroix 2000, Macdonald et al. 2011, Sun et al. 2008, Isaksson et al. 2006). The
values can be contrasted to those for healthy bone in Table 5.6. ....................................... 197

viii
List of Figures and Illustrations
Figure 1.1. Cortical and trabecular bone shown in a µCT cross-sectional view of a murine
tibia. ....................................................................................................................................5
Figure 1.2. µCT images of vertebrae from two patients: (a) Normal bone in a 74 year old
woman (b) osteoporotic bone in a 94 year old woman (Cole et al. 2010). ........................... 10
Figure 1.3. Fracture healing stages: (a) inflammation, (b) callus differentiation, (c)
ossification, (d) remodelling (Bailon-Plaza and van der Meulen 2001)............................... 13
Figure 1.4 The essential tissues involved in fracture healing (Einhorn 2005). ............................ 14
Figure 1.5. (a) Sheep tibia subjected to torsion. (b) Histological section showing the
cartilaginous formation in the interfragmentary gap and the bony tissue in the internal
and external callus (Bishop et al. 2006).............................................................................. 17
Figure 1.6. (a) Lateral loading of the left knee, (b) µCT images of burr-holes in control
group with delayed healing, and (c) loaded group with promoted healing. Bar = 1 mm
(Zhang and Yokota 2011). ................................................................................................. 19
Figure 1.7. µCT images demonstrating the healing process and bone regeneration during 4
weeks: (a) in the control group, (b) collagenous scaffold seeded with ES cell-derived
osteoblasts implanted into the fracture site (Taiani 2012). Bar = 1 mm. ............................. 19
Figure 1.8. Both experimental and computational studies are required to fully understand the
tissue behaviour under mechanical loads and develop mechanoregulatory algorithms
(Epari, Duda and Thompson 2010). ................................................................................... 22
Figure 1.9. Objectives of the current study based on the in vitro collagenous scaffold and in
vivo burr-hole murine model. ............................................................................................ 23
Figure 1.10. Location of the burr-hole in the medial aspect of the tibia: (a) reconstructed FE
model based on the in vivo study, (b) µCT image of the in vivo model (Taiani 2012). A
section through the long axis of the burr-hole looking proximally: (c) reconstructed FE
model based on the in vivo study and (d) µCT image (Taiani 2012). .................................. 25
Figure 2.1. Types of the mechanoregulatory algorithms. ........................................................... 29
Figure 2.2. A schematic of Pauwels' theory, octahedral shear stress and hydrostatic pressure
used as biophysical stimuli (Pauwels 1960). ...................................................................... 30
Figure 2.3. Strain tolerance of different types of tissues (Tubbs 1981). ...................................... 31
Figure 2.4. Graph shows the mechanostat theory proposed by Frost (Frost, 2003). The
relationship between the tissue strain level and change in bone mass is presented. AW,
MOW and POW refer to the dead zone, mild overload and excessive load, respectively.

ix
MESm and MESp are the bone modelling and the micro-damage thresholds,
respectively. Fx is the bone fracture strength. These setpoints vary between individuals
and are hypothesised to be genetically determined. ............................................................ 33
Figure 2.5. The effect of mechanical loading and vascularity on bone differentiation (Carter
et al. 1988). ....................................................................................................................... 36
Figure 2.6. Carter s me hano iology theory. rin ipal tensile strain and hydrostati stress
history are the key iophysi al stimuli (Carter and eaupr 2001). .................................... 37
Figure 2.7. Single phase model introduced by Claes et al. (1999) in which hydrostatic
pressure and strain are the key biophysical stimuli. ............................................................ 38
Figure 2.8. Biphasic model introduced by Kuiper et al. (2000a), using fluid shear strain and
stress as the biophysical stimuli. ........................................................................................ 40
Figure 2.9. Proposed biphasic algorithm by Lacroix et al. (2002) in which octahedral shear
strain and fluid velocity are the biophysical stimuli. ........................................................... 42
Figure 2.10. Biphasic model introduced by Sandino and Lacroix (2011) in which fluid shear
stress and octahedral shear strain were used as biophysical stimuli. ................................... 44
Figure 2.11. (a) The relationship between the sprout branching and the sprout length, (b) the
rate of sprout growth as a function of mechanical stimulus (Checa and Prendergast
2009). ................................................................................................................................ 51
Figure 2.12. Mechanoregulatory algorithm proposed by Checa and Prendergast (2009)
simulating tissue differentiation by both the local mechanical environment and the
presence of oxygen from nearby blood vessels................................................................... 51
Figure 2.13. The mechanobiological algorithm proposed by Isaksson et al. (2008) simulating
tissue differentiation according to local mechanical environment, cellular events and
matrix production. ............................................................................................................. 53
Figure 2.14. Mechanoregulatory algorithm proposed by Sandino and Lacroix (2011)
simulating tissue differentiation based on mechanical stimuli, cellular events and
angiogenesis. ..................................................................................................................... 55
Figure 2.15. Schematic of the algorithm used by Nagel and Kelly (2010) to incorporate
collagen fibre orientations.................................................................................................. 57
Figure 3.1. Summary of model development and verification. ................................................... 61
Figure 3.2. The force-displacement plot representing bone behaviour (Cole et al. 2010). ........... 62

x
Figure 3.3. Relationship between the mineral content and bone mechanical properties: (a) by
increasing the mineral density, the stiffness increases, (b) while an increase in the bone
mineral content leads to more brittleness (Currey 2012)..................................................... 64
Figure 3.4. Variation of bone mass in men and women across the lifespan (McDonnell et al.
2007). ................................................................................................................................ 65
Figure 3.5. The material properties of a femoral osteoporotic (OP) bone were compared to
normal bone: (a) bone stiffness, (b) the yield strength, (3) bone density, and (d) the
absorbed energy of the bone decreases in osteoporotic bone (Li and Aspden 1997a). ......... 66
Figure 3.6. The radiographs of the proximal humerus in (a) a 87-year-old man with low bone
mineral density and, (b) a 65-year-old man with higher mineral density (Tingart et al.
2003). ................................................................................................................................ 67
Figure 3.7. The graph demonstrates the relationship between porosity and age for men and
women (McCalden et al. 1993). ......................................................................................... 68
Figure 3.8. The relationship between stiffness and density of the subchondral bone plate (Li
and Aspden 1997b). ........................................................................................................... 68
Figure 3.9. Excessive torque may cause a spiral fracture (a), tension causes transverse
fracture (b), oblique fracture may be created by compression (c), and butterfly fragments
may result from bending (d) (Giotakis and Narayan 2007). ................................................ 70
Figure 3.10. Variation in the size and mass distribution affects the bending stiffness of bone.
The middle figure represents a young bone with thicker cortical shell, whereas the bone
with thinner cortical shaft, at the right, represents an adult bone. In the older bone,
resorption of the inner surface and apposition of the outer surface decreased the bone
thickness. The bone mass was distributed further from the bending plane compared to
the young bone. ................................................................................................................. 72
Figure 3.11. The stress-strain curve illustrates that bone is stiffer in the longitudinal direction.
Young‟s modulus is the same in tension and ompression, whereas in ompression one
has more strength (Kutz 2003). .......................................................................................... 73
Figure 3.12. Young‟s modulus and ultimate stress of tibial cortical bone decrease with
increasing age. The rate of decrease is higher for ultimate stress compared to the
modulus (Burstein et al. 1976). .......................................................................................... 74
Figure 3.13. The response of cortical bone under different strain rates (Kutz 2003). .................. 75
Figure 3.14. The stress-strain plot for a load-unload-reload trabecular sample. The loading
starts at point 1, it is unloaded at point 2 and reloaded at point 3. The initial Young‟s
modulus from the linear section of the reloading (3-4) is the same as the original

xi
Young‟s modulus (1-2) at the beginning, but reduces quickly to residualE . A permanent
residual strain residualε will be developed (Keaveny, Wachtel and Kopperdahl 1999). .......... 76
Figure 3.15. (a) Movement of fluid when tissue is under a free draining confined
compression test, (b) ramp deformation is applied (increased with a linear ramp to B and
remained constant from B to E, (b) the flow exudes immediately after the deformation is
applied and the stress reaches its maximum amount (B). The fluid continues to flow.
The tissue reaches an equilibrium point and the stress decreases and reaches a plateau
(E) (Mow et al. 1980). ....................................................................................................... 78
Figure 3.16. Proposed biphasic algorithm by Prendergast et al. (1997); strain and fluid
velocity are the biophysical stimuli. ................................................................................... 87
Figure 3.17. The iterative model used to simulate fracture repair. .............................................. 88
Figure 3.18. A partial input file showing how the field variable was defined in the input file. .... 91
Figure 3.19. The profiles of (a) temperature and (b) fluid velocity are shown under the same
boundary condition show the same pattern (Cussler 2009). ................................................ 95
Figure 3.20. (a) Axisymmetric model of a human tibia, the radius of the cortical and bone
marrow are 15 and 9 mm (at the left). The cortical, bone marrow and callus are shown in
red, grey and green, respectively, (b) three origins for progenitor cells are shown.
Arrows indicate the cell origins (at the right). .................................................................... 96
Figure 3.21. (a) Cell concentration within the callus at two time points: 0.5s and 1 s, (b) cell
concentration over time in three sample elements, (c) the cell concentration at two time
points through the callus. ................................................................................................... 97
Figure 3.22. Schematic of the implemented algorithm to predict tissue distribution. The
coupled diffusive-poroelastic analysis for the mechanical stimulus and cell
concentration were obtained for each element (ABAQUS). ............................................... 99
Figure 3.23. The axisymmetric model of an ovine tibia with 3mm fracture gap and an
external callus: (a) Isaksson et al. (2006), and (b) the present study: bone marrow (in
red), cortical bone (in orange) and callus (in blue) are modelled. ..................................... 101
Figure 3.24. Prediction of fracture healing in the present study during 50 steps (days).
Cortical bone was subjected to a 300 [N] axial compression load (1 Hz). ......................... 104
Figure 3.25. The change of fluid flow [ m/s] over time under the cortical shaft and callus tip
during fracture healing. Cortical bone was subjected to a 300 N axial compression
loading (1 Hz). ................................................................................................................ 105
Figure 3.26. Comparison of the two simulations during fracture healing in the first steps.
Cortical bone was subjected to a 300 N axial compression loading (1 Hz): (a) the present

xii
study, and (b) Isaksson et al. (2006). In both models, intramembranous ossification
occurred at the callus tip and periosteum. Also, fibrous tissue and cartilaginous tissue
can be found in the same zones. ....................................................................................... 105
Figure 3.27. Comparison of the two simulations during fracture healing at step 50, cortical
bone was under 300 N axial compression loading (1 Hz): (a) the present study, (b)
Isaksson et al. (2006). The amount of fibrous tissue decreased significantly under the
cortical shaft. The distribution of mature and intermediate bone distribution is almost
identical. .......................................................................................................................... 106
Figure 3.28. Overall similar healing patterns were observed over time under a 300 [N] axial
compression load in (a) the present study, and (b) Isaksson et al. (2006). ......................... 107
Figure 3.29. (a) Axisymmetric FE model of a murine tibia (bone marrow in red, cortical bone
in grey and callus in blue), (b) the amplitude of the applied cyclic axial compression
loads. ............................................................................................................................... 109
Figure 3.30. Predicted tissue differentiation in the present study under (a) 0.5 N, (b) 1 N and
(c) 2 N axial compression load (1 Hz). ............................................................................. 112
Figure 3.31. Predicted mechanical stimuli for a sample element under the cortical shaft
during the healing process for three axial compression loading magnitudes. .................... 113
Figure 3.32. a) Axisymmetric FE model of a murine tibia. The inner and outer diameter of
cortical bone (gray) and external callus (blue) are 1, 1.5 and 2.4 mm, respectively, b)
Predicted tissue differentiation in the present study, c) CT images from Gardner et al.
(2006) for different load magnitudes (1 Hz). .................................................................... 114
Figure 3.33. The sample elements considered within three regions of the callus (external and
internal callus, and the interfragmentary gap). ................................................................. 116
Figure 3.34.Prediction of mechanical stimuli at three sample points, when bone is subjected
to axial torsion. The sample element under the cortical shaft has the highest mechanical
stimuli. ............................................................................................................................ 116
Figure 3.35. (a) A 3D FEM of a murine tibia with 0.4 mm gap, predicted tissue
differentiation in the model under: (b) Axial torsion (8 degree, 1 Hz), (c) Axial torsion
& compression (8 degrees, 0.45 MPa, 1 Hz). ................................................................... 117
Figure 3.36. (a) Sheep tibial section subjected to torsion (histological slide) (Bishop et al.
2006), (b) predicted tissue differentiation subjected to torsion (day 15, in the present
study). ............................................................................................................................. 118
Figure 3.37. Summary of the computational simulations in the current study. .......................... 121
Figure 4.1. FX-4000 ™ Flex ell ® ompression plus ™ system. ............................................. 123

xiii
Figure 4.2. Schematic of the Flexcell system cross-section in the uncompressed and
compressed configurations............................................................................................... 124
Figure 4.3. Parts of the modified system: (1) cell base, (2) square ring, (3) porous plug, (4)
round ring, (5) lid, and (6) fixed lid.................................................................................. 125
Figure 4.4. The modified system to conduct confined compression test. .................................. 126
Figure 4.5. The parts added to the modified Flexcell system. ................................................... 127
Figure 4.6. The force-displacement curve of the rings obtained from a uniaxial compression
test (Olesja Hazenbiller, M.Sc. Student, University of Calgary). ...................................... 131
Figure 4.7. The axial displacement [mm] of the axisymmetric model of the square ring under
30 N axial compression load at t=30 s. The inner and outer diameters are 10.7 mm and
12.7 mm, respectively, and the height is 1 mm. ................................................................ 132
Figure 4.8. The axial displacement [mm] of the axisymmetric model of the round ring under
30 N axial compression load at t=30 s. The inner and outer diameters are 6 mm and 10
mm, respectively, and the radius of the cross section is 2 mm. ......................................... 133
Figure 4.9. The axial displacement [mm] of the axisymmetric FE model of the system
without gel under 30 N axial compression load at t=30 s. The diameter of the porous
plug is 12.7 mm and the height is 3.175 mm. ................................................................... 135
Figure 4.10. Comparison of load-displacement curves between the FE and experimental
results under axial compression load applied at a rate of 1 N/s. ........................................ 136
Figure 4.11. The axisymmetric FE model of the modified Flexcell system. ............................. 137
Figure 4.12. (a) The contact surface at the beginning of the analysis, (b) if no adjustment
zone was used, ( ) the adjustment „a‟ was defined and the nodes within the zone were
moved onto the master surface (ABAQUS v6.11 user manual). ....................................... 138
Figure 4.13. Model predictions of the force-displacement curve for the whole system
including the collagen gel under confined compression: ramp load with the rate of 1 N/s. 140
Figure 4.14. Distribution of axial strain (EE2) and fluid velocity (FLVEL, [mm/s]), within
the collagenous scaffold under 20 N axial compression load at t=20 s.............................. 141
Figure 4.15. Change of fluid velocity over time within four sample elements of the
collagenous scaffold. The scaffold was loaded at a rate of 1 N/s. Four sample elements
are shown through the depth of the collagen (at the right). ............................................... 142
Figure 4.16. Change of axial strain over time within four sample elements of the collagenous
scaffold The scaffold was loaded at a rate of 1 N/s. Four sample elements are shown
through the depth of the collagen (at the right). ................................................................ 142

xiv
Figure 4.17. The load-displacement behaviour of the gel can be observed by comparing: (1)
the whole model including the gel, and (2) the whole model without the gel. The
deformation of the square ring decreased, while the deformation of the round ring was
independent from the gel and did not change. .................................................................. 143
Figure 4.18. The amplitude of the applied cyclic axial compression loads. The pressure,
ranging from 5-20 [kPa], was applied to the cell base with a frequency of 1 Hz. .............. 144
Figure 4.19. Model prediction for the axial strain during confined compression: cyclic
loading (P=5 kPa, 1 Hz). Compressive strain in top element under axial compression. ..... 145
Figure 4.20. Prediction of axial strain at the peak loading in three selected sample elements in
the collagenous scaffold in confined compression under cyclic loading of P=5 kPa (1
Hz). ................................................................................................................................. 145
Figure 4.21. Prediction of octahedral shear strain at the peak loading in three selected sample
elements in the collagenous scaffold in confined compression under cyclic loading of
P=5 kPa (1 Hz). ............................................................................................................... 146
Figure 4.22. Prediction of fluid velocity at the peak loading in the top sample element in the
collagenous scaffold in confined compression under cyclic loading of P=5 kPa (1 Hz). ... 146
Figure 4.23. Prediction of pore pressure at the peak loading in three selected sample elements
in the collagenous scaffold in confined compression under cyclic loading of P=5 kPa (1
Hz). ................................................................................................................................. 147
Figure 4.24. Comparison of axial strains at the peak loading in the top sample element in the
collagenous scaffold in confined compression under different cyclic applied loads of
P=5, 10, 20 kPa (1 Hz). ................................................................................................... 148
Figure 4.25. The gel was subjected to 10 kPa pressure (1 Hz). The hysteresis of the stress-
strain curve shows the effects of the interstitial flow and viscous dissipation (the graph
shows the first 33 seconds of analysis). ............................................................................ 148
Figure 4.26. Schematic of tissue differentiation algorithm. ...................................................... 151
Figure 4.27. Prediction of fluid velocity at the peak loading in three sample elements in the
collagenous scaffold during tissue differentiation (P = 20 kPa, 1 Hz). .............................. 152
Figure 4.28. Mechanical stimuli in a sample element at the superficial layer for a 5 kPa and a
20 kPa (1 Hz) compressive pressure applied to the system. .............................................. 153
Figure 4.29. (a) Gel before and after loading, (b) FE prediction of tissue differentiation (a 20
kPa compressive stress, 1 Hz). ......................................................................................... 154
Figure 4.30. Compressive load significantly influenced chondrogenesis (Col 2-day 15),
(Olesja Hazenbiller, M.Sc. student, University of Calgary). ............................................. 154

xv
Figure 5.1. Overview of this chapter. ....................................................................................... 157
Figure 5.2. Top view of sample holder is located fitted within the diameter of the field of
view (orange lines), (b) schematic of the sample holder used for scanning the murine
tibia (µCT 35, User Manual). ........................................................................................... 159
Figure 5.3. Cortical and trabecular bone could be visually distinguished in the µCT cross-
sectional view of a murine tibia. ...................................................................................... 161
Figure 5.4. Bone geometry before and after applying the recursive Gaussian filter. ................. 162
Figure 5.5. Cross-sectional view showing the cortical bone, bone marrow and trabecular
bone. ............................................................................................................................... 163
Figure 5.6. Illustrated are the masks for five different locations: (a) proximal tibia (b)
proximal tibial diaphysis, (c) tibial crest diaphysis, (d) midshaft, (e) distal tibia; Red,
blue and yellow masks represent cortical, trabecular and bone marrow, respectively........ 163
Figure 5.7. Lateral, medial, posterior, and anterior views of the finite element model of the
reconstructed murine tibia................................................................................................ 166
Figure 5.8. (a) Volume image, (b) greyscale data, (c) segmented mask, (d) isolated
segmented mask, (e) smoothed mask (recursive Gaussian filter), (f) mesh generation of
the extracted volume. ....................................................................................................... 167
Figure 5.9. Three zones that were compared: zone 1 (proximal tibia), zone 2 (tibial crest),
zone 3 (distal tibia). ......................................................................................................... 169
Figure 5.10. Force-strain relations measured by (a) Stadelmann et al. (2009), and (b)
predicted in the present study. .......................................................................................... 169
Figure 5.11. The distribution of principal strains when the tibia was subjected to a 10 N axial
compression load. ............................................................................................................ 170
Figure 5.12. Location of the burr-hole in the medial aspect of the tibia: (a) FE model, (b)
experimental fracture model (Taiani 2012). A section through the long axis of the burr-
hole: (c) FE model, (d) experimental fracture model (Taiani 2012). A section through
the frontal plane of the fractured tibia: (e) FE model, (f) experimental fracture model
(Taiani 2012). .................................................................................................................. 172
Figure 5.13. Workflow diagram outlining the required functions to reconstruct the 3D FE
burr-hole model. .............................................................................................................. 173
Figure 5.14. Overview of the processes used to create the burr-hole model. ............................. 173

xvi
Figure 5.15. The decay length models were subjected to a 10 N load to select the one that had
the closest mechanical environment compared to the full-length model. The red arrow
shows where the load was applied. .................................................................................. 174
Figure 5.16. Distribution of von Mises stress: (a) full-length model, (b) decay length model
(tibial crest, medial view). ............................................................................................... 175
Figure 5.17. Distribution of principal strains: (a) full-length model, (b) decay length model
(tibial crest, medial view). ............................................................................................... 176
Figure 5.18. Distribution of principal strains: (a) full-length model, (b) decay length model
(tibial crest, lateral view). ................................................................................................ 177
Figure 5.19 Distribution of von Mises stress within the scaffold: (a) full-length model, (b)
decay length model (tibial crest, medial view). ................................................................ 177
Figure 5.20 Distribution of principal strains within the scaffold: (a) full-length model, (b)
decay length model (tibial crest, medial view). ................................................................ 178
Figure 5.21. (a) Origins of the progenitor cells, (b) sample elements in the middle section and
top surface. ...................................................................................................................... 183
Figure 5.22. Mechanical stimuli of an outer radial sample element at the mid-section of the
scaffold (at peak load). Tibia was subjected to axial compression loads of 2, 1, and 0.5
N (1 Hz). ......................................................................................................................... 183
Figure 5.23. Mechanical stimuli of two sample elements at outer radial side of the scaffold
(at peak load): (1) mid-section, and (2) proximal surface. The tibia was subjected to a 2
N (1 Hz) axial compression load. ..................................................................................... 184
Figure 5.24. Mechanical stimuli of three sample elements located on the proximal surface of
the scaffold (at peak load): (1) outer radial, (2) middle, and (3) inner radial zone. The
tibia was subjected to a 2 N (1 Hz) axial compression load. ............................................. 184
Figure 5.25. The predicted interfragmentary strain, at peak load, under the cortical shaft for
the three loading cases (2, 1, 0.5 N axial compression, 1 Hz). .......................................... 185
Figure 5.26. Predicted fracture healing patterns under the 2, 1 and 0.5 N (1 Hz) axial
compression load. ............................................................................................................ 186
Figure 5.27. Cross-sectional view of the scaffold showing the accelerated healing of the core
compared to the outer layers. The tibia was subjected to a 2 N (1 Hz) axial compression
load. ................................................................................................................................ 187

xvii
Figure 5.28. The prediction of octahedral shear strain for different cell diffusion rates (0.025
and 0.01 s
mm2
). .............................................................................................................. 189
Figure 5.29. Predicted fracture healing patterns under the 1 N (1 Hz) axial compression load
for different rates of cell diffusion (0.025 and 0.01 s
mm2
)............................................... 189
Figure 5.30. Axial positions of the burr-hole fracture: (a) 2.55 mm and (b) 3.13 mm from the
proximal end. Both fractures are located in the trabecular bone and the tibia was
subjected to a 1 N (1 Hz) axial compression load. Bar = 0.7 mm. .................................... 191
Figure 5.31. Predicted mechanical stimuli in the outer radial location on the proximal surface
of the scaffold for two different hole positions: 3.13 mm versus 2.55 mm from the
proximal end. The tibia was subjected to a 1 N (1 Hz) axial compression force................ 192
Figure 5.32. Predicted fracture healing patterns for the two positions of burr-hole fractures.
The tibia was subjected to a 1 N (1 Hz) compression load................................................ 192
Figure 5.33. Cross sections of the 3.13 mm and 2.55 mm fracture cases at day 13. The 3.13
mm fracture case had slightly accelerated healing. Both tibiae were subjected to a 1 N
axial compression load (1 Hz).......................................................................................... 192
Figure 5.34. The origins of progenitor cells when the fracture is located in the bone marrow. .. 193
Figure 5.35. Predicted mechanical stimuli for two locations of the fracture (outer radial
sample element on the proximal surface): (1) in trabecular bone and (2) in bone marrow.
Tibia was subjected to a 2.5 N (1 Hz) compression load. ................................................. 194
Figure 5.36. The predicted tissue pattern for two locations of the fracture with different cell
origins: in trabecular bone, and in bone marrow. The proximal end of tibia was
subjected to a 2.5 N (1 Hz) compression load. ................................................................. 195
Table 5.7. Mechanical properties used for an osteoporotic murine tibia (Li and Aspden
1997b, Lacroix 2000, Macdonald et al. 2011, Sun et al. 2008, Isaksson et al. 2006). The
values can be contrasted to those for healthy bone in Table 5.6. ....................................... 197
Figure 5.37. The predicted mechanical stimuli at peak load for the osteoporotic and normal
bone. Tibia was subjected to a 2.5 N (1 Hz) axial compression load. ............................... 198
Figure 5.38. The predicted interfragmentary strain under the cortical shaft, at peak load, for
the osteoporotic and normal bone. Tibia was subjected to a 2.5 N (1 Hz) axial
compression load. ............................................................................................................ 198
Figure 5.39. The predicted tissue patterns for fracture repair within osteoporotic versus
normal bone. Tibia was subjected to a 2.5 N (1Hz) axial compression load. ..................... 199

xviii
Figure 5.40. Red arrow shows the bending load that was applied to the tibia. The distal end
was fixed and the hole size was 0.7 mm. Bar = 0.7 mm. .................................................. 200
Figure 5.41. Predicted mechanical stimuli for the outer radial sample element at located at the
proximal surface of the scaffold. The tibia was subjected to 0.04 and 0.02 Nm (1 Hz)
bending loads. ................................................................................................................. 201
Figure 5.42. Predicted tissue pattern for the murine tibia subjected to bending loads of 0.04
and 0.02 Nm (1 Hz). ........................................................................................................ 201
Figure 5.43. The gradual change of tissue type during the healing period within a young
(D=0.025 s
mm 2
), and an old murine tibia (D=0.01 s
mm 2
). ............................................ 204
Figure 5.44. The overall stiffness of the scaffold during the healing process for young and old
murine tibia with different diffusion rates (D=0.025 s
mm 2
and D=0.01
s
mm 2
). ............. 204
Figure 5.45. Histological slides (Day 7): (a) a large and (b) a small amount of cartilage were
present in the fracture site of young and elder murine tibia, respectively. In the
histological slides, cartilage is shown in red (Lu et al. 2005). In the computational study
(Day 7) similarly, more cartilaginous tissue differentiated in the younger murine tibia (c,
d). .................................................................................................................................... 205
Figure 5.46. The gradual change of tissue type during the healing period within a normal and
an osteoporotic murine tibia. ............................................................................................ 206
Figure 5.47. The overall stiffness of the scaffold during the healing process for normal and
osteoporotic bone. ........................................................................................................... 207
Figure 5.48. The healing progression in a normal bone and an osteoporotic bone from CT
images show that the normal bone had more mineralisation (Taiani 2012). ...................... 207
Figure 5.49. CT images showing the healing process within the burr-hole fracture model of
the (a) unloaded murine tibia, (b) tibia under bending load of 0.02 Nm (Zhang et al.
2007). .............................................................................................................................. 208

xix
List of Symbols, Abbreviations and Nomenclature
Roman letters
B Body force per unit mass
Cp Specific heat coefficient
D Diffusion rate,
E
Hydrostatic stress
Young‟s modulus
e Void ratio
GS Greyscale value
I Osteogenic index
J Deformation gradient
K Diffusive drag coefficient
k Fluid permeability,
Thermal conductivity
kc
n
Contact permeability
Porosity
p Fluid pore pressure
Ra Source of mass
S Octahedral shear stress,
Biphasic mechanical stimulus,
T
Specific matrix surface
Temperature
t Time
V
v
Volume
Fluid velocity
W Strain energy
Matrices and vectors
I Identity matrix
k Permeability tensor
q Fluid flow
T Deviatoric shear tensor
u Displacement
v Velocity
w Relative fluid displacement
Strain
Local body force
Stress

xx
Greek letters
Thermal diffusivity
Octahedral shear strain,
Specific weight
Stretch ratio
, Lamè material constants
Micro
oisson‟s ratio
Density
τ Fluid shear stress
Superscripts
f Fluid
s Solid

1
Chapter One: Introduction
1.1 Background and motivation
Bone is a complex, composite tissue composed of a mineralized component, and soft
extracellular matrix proteins, with dynamic interactions between these components across
multiple scales (Carter et al. 1998). This dynamic tissue has an exceptional ability to self-heal
throughout life. In healthy bone, there is a consistent balance between bone resorption and bone
formation. However, bone diseases or aging may affect the factors regulating this process, and
result in an altered bone micro-architecture, bone loss and reduced structural strength, making
bone more prone to fracture (McDonnell, McHugh and O'Mahoney 2007, Currey 2012, Jepsen
and Andarawis-Puri 2012).
The treatment and prevention of bone fracture is a global health problem for difficult-to-heal
fractures that can occur in osteoporotic elderly individuals. It is generally accepted that the rate
of fractures increases substantially with age (Knopp et al. 2005, Egermann, Goldhahn and
Schneider 2005). It has been estimated that at least 40 % of women and 15 % of men suffer from
a bone fracture after age 65 (Riggs, Khosla and Melton 2002). According to Gullberg et al.
(1997) hip fractures are set to increase by 310 % and 240 % for men and women by 2050,
respectively.
To better preserve the quality of life of afflicted individuals, research has focused on
understanding the mechanisms involved in the maintenance and repair of the musculoskeletal
system in order to improve treatment of broken bones. It is well known that bone is able to
optimise its mechanical properties to different physical conditions, including during fracture
repair, yet the mechanisms initiating and regulating bone maintenance and repair are not clearly
known (McMahon, O'Brien and Prendergast 2008, Morgan et al. 2010). Non-ideal mechanical

2
conditions and cell-matrix interactions may disrupt the process of healing and lead to non-unions
and cell apoptosis (Kwong and Harris 2008, Geris et al. 2009, Einhorn 2005). Therefore,
effective positive mechanical factors are encompassed by the range and frequency of the applied
loads.
The effect of frequency on bone regeneration, ranging from 1 to 20 Hz, has been investigated
computationally at the bone-implant interface (Geris et al. 2009). A bone chamber was used to
allow for controlled implant axial displacement of up to 90 µm. Bone tissue formation was
predicted using a biphasic mechanoregulatory algorithms (Lacroix et al. 2002, Prendergast,
Huiskes and Soballe 1997) and was compared with histology (Vandamme et al. 2007, Duyck et
al. 2006). In contrast to findings in orthopaedic implant osseointegration (Rubin and McLeod
1994), lower frequencies (in the order of 1 Hz) were predicted to promote bone formation.
According to Geris et al. (2009), frequencies above 1 Hz did not promote bone formation, the
bone chamber was mostly filled with soft tissue, and less bone-to-implant contact was observed.
Rubin and McLeod (1994) compared loading frequencies of 1 and 20 Hz to investigate the effect
of loading rate on bone growth and reported that the 20 Hz loading case promoted bone growth
into the implant pores whereas the 1 Hz loading case delayed the differentiation process.
Goodship et al. (1998) showed that a higher rate of loading promoted healing in the primary
stages, but delayed the process in the later healing stages. This may be due to the viscoelastic
nature of the tissue. During the initial stages, the higher rates of movement increased the fluid
velocity and promoted fibrous tissue differentiation. However, the high loading rates inhibited
bone formation later in the healing process (Goodship et al. 1998). Further experimental and
computational studies are essential to verify the real effect of frequencies and loading rates on
bone stimulation and growth.

3
New bone regeneration is also related to the magnitude and direction of the loads, however, the
mechanisms of transduction of the mechanical stimuli into a biological response remains unclear
(Isaksson et al. 2008, van der Meulen and Huiskes 2002). High magnitudes of load were shown
to result in delayed healing caused by the persistence of cartilage not differentiating into bony
tissue (Claes and Heigele 1999, Einhorn 2005). Isaksson et al. (2006) showed that increasing the
load delayed healing and a further increase led to: (1) a non-union, (2) having fibrous tissue in
the interfragmentary gap and (3) cartilaginous tissue in the external callus. A comprehensive
understanding of the mechanobiology of tissue formation, could improve treatment techniques to
better prevent non-unions and accelerate the healing process.
The bioengineering of tissue-engineered constructs offers a promising treatment of fractures
where stem cells and scaffolds are combined to enhance bone repair. Central to the success of
these initiatives is a detailed understanding of how stem cells respond to and interact with
mechanical cues from the bone and the biomaterial scaffold as they differentiate into mature
tissue. A number of empirically-based mechanobiology algorithms have been proposed to predict
tissue differentiation that incorporates mechanical factors. Experiments to evaluate the effect of
mechanical regimes on tissue regeneration are expensive and time consuming. To more
comprehensively understand tissue regeneration, computational models have been developed to
determine the mechanical and physical conditions within healing tissues. Empirically-based
algorithms have been implemented into computational models to predict bone tissue formation.
A number of different algorithms have been proposed and are based on different mechanical
factors, e.g., fluid hydrostatic pressure, interstitial fluid flow, and tissue strain (Prendergast et al.
1997, Sandino and Lacroix 2011, Isaksson et al. 2008). These algorithms are correlated with
experimental in vivo studies to determine which factors involved to simulate the healing process.

4
Improved validation of these algorithms can enhance our understanding of tissue
mechanobiology and lead to development of more sophisticated algorithms. The computational
predictions can then be used to design better clinical treatment options for bone fracture
treatment. These algorithms have been used to understand the mechanobiology of a number of
clinical applications including (1) prediction of tissue differentiation during fracture healing in
long bones (Lacroix et al. 2002, Isaksson et al. 2008), (2) modelling the bony in-growth on the
surface of bone implants (Geris et al. 2004, Moreo, Garcia-Aznar and Doblare 2009a, Moreo,
Garcia-Aznar and Doblare 2009b), (3) prediction of the tissue regeneration pattern in a multi-
scale model of a lumbar vertebral fracture (Boccaccio, Kelly and Pappalettere 2011), (4)
understanding the process of bone healing in trabecular bone (Shefelbine et al. 2005), and (5)
improving the design of scaffolds for bone tissue engineering (Kelly and Prendergast 2005,
Milan, Planell and Lacroix 2010, Byrne et al. 2007).
The underlying mechanisms that are responsible for a promoted bone healing are still unknown.
A better understanding of the factors that affect tissue regeneration is essential to explore how
the mechanical stimuli are sensed and transmitted to the tissues and cells. These studies can then
be used to further develop the mechanoregulation algorithms to include simulation of
osteoporotic bone and treatment with pharmacological approaches. These algorithms can then be
used to design bone scaffold models, which best transfer the mechanical signals and better
promote the healing process.
1.2 Bone
The adult skeleton is composed of 213 bones, excluding the sesamoid bones (Mitchell et al.
2005). The skeleton serves several mechanical and physiological functions in the human body:
(1) supports the body and protects softer internal organs (e.g. brain, heart and lung), (2) provides

5
locomotion with the help of muscles and joints, (3) ensures a balance of minerals that are
essential for the body and forms blood cells in the bone marrow. The first two functions are
mechanical and the latter is physiological (Clarke 2008, Einhorn 1998, Herzog and Nigg 1999).
Bone is inherently a hierarchical structure. The hierarchy levels of bone structure are nano-scale
(organic and inorganic phases and water), micro-scale (the visible structure under microscope),
meso-scale (cortical and trabecular bone) and macro-scale (the whole bone) (Currey 2012).
From a meso-scale point of view, bone can be divided into cortical (or compact) and trabecular
(or cancellous) bone. The adult skeleton is composed of 80 % cortical and 20 % trabecular bone
(Figure 1.1). Cortical bone forms the outer surface of the bones, whereas trabecular bone is
rarely found on the outer surface of the bone and is usually covered by a thin layer of cortical
bone. Different bones have different ratios of trabecular to cortical bone. The amount of
trabecular bone is three times greater than cortical in vertebrae, whereas in the femoral head it is
50:50 and for the radial diaphysis it is 5:95 (Clarke 2008).
Figure 1.1. Cortical and trabecular bone shown in a µCT cross-sectional view of a murine
tibia.

6
Cortical bone which improves the ability to resist bending and torsional loads due to its high
density and geometric configuration, can be found in the shaft, and proximal and distal ends of
the long bones. It Cortical bone is mainly composed of cylindrical lamellar-shaped elements,
termed osteons. The vertical alignment of the osteons gives strength to the cortical bone to bear
mechanical loads. Two connective tissues cover the inner and outer surfaces of the cortical bone:
the endosteum and the periosteum. The endosteum covers the inner surface of the cortical bone
facing the marrow cavity, whereas the periosteum is a layer of fibrous tissue on the external
surface of the cortical bone that isolates bone from the surrounding tissues. These connective
tissues are involved in fracture repair and are filled with cells needed to maintain the bone
formation/resorption balance. At the periosteal surface, bone formation exceeds bone resorption,
and thus the cortical diameter increases with aging. On the other hand, at the endosteal surface,
bone resorption is greater than bone formation and thus the marrow cavity expands with aging.
The endosteal surface is usually subjected to higher mechanical strains with greater remodelling
activities compared to the outer regions (Clarke 2008).
Unlike cortical bones, trabecular bone is less dense formed by honeycomb like structures with
less rigidity overall. Trabecular bone can be found at the ends of the long bones, throughout the
length of short bones, in large flat bones and where muscle is attached to bone. (Yaszemski et al.
1996, Clarke 2008). The porosity of trabecular bones ranges from 75 %-95 %, in contrast to
cortical bone where the porosity ranges from 5 %-10 % (Burr, Sharkey and Martin 1998).
Although trabecular bone has less strength, it has greater metabolic activity compared to compact
bone. The interconnected pores in trabecular bone are filled with bone marrow, which is
composed of blood vessels, nerves, and different types of cells. Bone marrow, like the outer

7
periosteal surface of the cortical bone, is responsible for bone remodelling activities during
fracture repair.
At the microscopic level, two types of bone can be distinguished: (1) woven or primary bone,
and (2) lamellar or secondary bone. Lamellar bone has an organised structure with aligned
collagen fibres and forms gradually (Burr et al. 1998). In contrast, woven bone forms rapidly and
has a randomly arranged structure with less strength. Woven bones form when there is an
immediate need of bone remodelling; such as during fracture healing, joint development or
osteochondral defects. The woven bone will be mostly replaced with the lamellar bone during
the repair process (Buckwalter et al. 1996).
Bone can be synthesised in two different pathways: (1) intramembranous ossification, and (2)
endochondral ossification. Intramembranous ossification is the process by which woven bones
differentiate from osteoblasts and mesenchymal cells, and then begin to secrete osteons. In this
type of ossification cartilage does not form. This process may occur during the formation of flat
bones, the growth of short bones and thickening of long bones (Buckwalter et al. 1996). During
endochondral ossification, bone forms from a cartilaginous template. During the healing process,
the cartilaginous tissue will be gradually replaced by bone. This process occurs during formation
and remodelling of long bones (Buckwalter et al. 1996).
There are two different processes involved in the dynamic adaptive behaviour of bone: (1) bone
modelling, and (2) bone remodelling. The process of bone growth, in which the bone mass
increases, is called bone modelling. Normal physiological activities may lead to micro-cracks at
the surface of the bones. Bone, as an active tissue, repairs the cracks by replacing the old bone
with new bone. The process of coupled bone formation and resorption is called bone
remodelling. In this process, both mass formation and resorption may occur. During the

8
remodelling process, just a fraction of the bone surface is active; whereas during bone growth
almost the entire tissue is involved (Herzog and Nigg 1999).
There are four important cell types within the bone extracellular matrix that are responsible for
bone modelling and remodelling: (1) osteoprogenitor cells that are composed of mesenchymal
stem cells and are able to differentiate into osteoblasts, (2) osteoblasts that are bone-forming
cells derived from bone marrow progenitors, and whose main function is to secrete osteons, (3)
osteoclasts that dissolve bone matrix by secreting acids and enzymes, and (4) osteocytes, the
most abundant cell in bone, that differentiate from osteoblasts and are directly integrated with the
bone matrix (Burr et al. 1998, Heino and Hentunen 2008).
When bone is mechanically loaded, the resulting deformation induces the flow of interstitial fluid
within the tissue. Osteocytes are believed to be able to sense the fluid flow and the local
deformation (Apostolopoulos and Deligianni 2009, Cowin 2002, Deligianni and Apostolopoulos
2008). Osteocytes are sensitive to the loading rate due to viscoelastic interactions between the
cells and the extracellular matrix (i.e. the matrix that provides structural support to the cells).
Immediately after the deformation of the extra cellular matrix, signalling molecules are produced
by the cells and bone generation is initiated. Osteocytes preserve the material properties of the
bone through the remodelling process (Galli, Passeri and Macaluso 2010, Apostolopoulos and
Deligianni 2009).
In response to mechanical perturbation, osteocytes can sense the mechanical environment
through shear sensitive membrane receptors, and communicate to surrounding osteoblasts and
osteoclasts to regulate bone formation and resorption (Turner and Pavalko 1998, Galli et al.
2010). Mechanical loads activate the cellular processes required for bone regeneration: energy
metabolism, gene activation, production of growth factors and matrix synthesis (Rangaswami et

9
al. 2009). It has also been suggested that osteocytes are ideally located to sense and detect the
local mechanical environment (Bacabac et al. 2008, Verborgt, Gibson and Schaffler 2000).
Apoptosis (i.e. programmed cell death) of osteocytes can result in loss of communication
between cells and a delayed healing process (Noble 2003).
1.2.1 Bone repair
The treatment of bone fractures is a major challenge and a global health issue. Bone fractures can
reduce the quality of life significantly. It is generally accepted that elderly patients are more
prone to bone fractures (Knopp et al. 2005) than bones of young individuals which are more
resistant to fracture. Furthermore, fractures heal more quickly in youth than in adults, which may
be the result of the reduced ability to recruit mesenchymal stem cells (Bailon-Plaza and van der
Meulen 2001, Bailón-Plaza and van der Meulen 2003, Geris et al. 2009). It has been shown that
by age 80 the bone mineral density (BMD) of the spine, hip and forearms decreases by 13-18 %
in men (Schulmerich et al. 2006) and 15-54 % in women (Ahlborg et al. 2003).
Mesenchymal stem cells are found in the red bone marrow and are able to differentiate into
skeletal tissues such as bone, cartilage, fibrocartilage, and fibrous tissues (Bielby, Jones and
McGonagle 2007, Loboa et al. 2003). Mesenchymal stem cells are essential to repair and replace
damaged tissues (Claes and Heigele 1999), regulate the balance between osteoblast and
osteoclast cells, and play a key role in the bone repair process (Bielby et al. 2007). Due to the
lower concentration of mesenchymal stem cells in human adult bones, they are less able to repair
the fracture. Furthermore, the healing process takes longer (Postacchini et al. 1995). In healthy
bone, there is a controlled balance between the activities of bone forming cells (i.e. osteoblast)
and bone resorbing cells (i.e. osteoclast). However, bone diseases such as osteoporosis, reduces
the bone mass, changes the bone architecture and its mechanical properties. Moreover, the

10
balance between bone formation and bone resorption is reduced (Figure 1.2). Therefore, bone
mass begins to decrease as bone absorption outpaces bone formation. The loss of mass leads to
lower bone density and increases the risk of fracture (Knopp et al. 2005, Geris et al. 2009). A
comparison between two different females, one with normal bone and the other with osteoporotic
bone, has shown that in addition to the loss of bone mass, the bone strength is also reduced
significantly in osteoporotic bones. For osteoporotic bone, a 20 % reduction in mass density
resulted in a 40 % reduction in stiffness (Cole, Meulen and Adler 2010, Cole and van der Meulen
2011).
Figure 1.2. µCT images of vertebrae from two patients: (a) Normal bone in a 74 year old
woman (b) osteoporotic bone in a 94 year old woman (Cole et al. 2010).
1.2.1.1 Biological stages of fracture healing
Fracture healing can occur in one of two ways: (1) primary/direct healing that occurs without a
callus (i.e. a mass of undifferentiated cells) formation, and (2) secondary/indirect fracture repair
that involves the regeneration of both the original geometry and cellular events. Primary healing
is divided into a series of four sequential stages (Bailon-Plaza and van der Meulen 2001,
Gerstenfeld et al. 2003): (1) inflammation, (2) callus differentiation, (3) ossification, and (4)
remodelling.

11
A fracture disrupts bone tissues and blood vessels (Figure 1.3a) and the first stage of fracture
healing initiates. Immediately after a bone ruptures, a hematoma forms, i.e., blood emanates
from the damaged vessels and fills the fracture gap. The hematoma is a source of signalling
molecules that initiates the healing process (Marsell and Einhorn 2011). Progenitor cells produce
inflammatory cells that are responsible for making new blood vessels, fibrous tissues, and
supporting cells (Frost 1989). The formation of new blood vessels (i.e. angiogenesis) accelerates
the differentiation of granulation tissues and the transport of progenitor cells to the fracture zone.
Initial formation of granulation tissues enables the migration of stem cells throughout the site of
injury and increases the concentration of mesenchymal stem cells. By this stage, a connective
tissue matrix has been formed. This matrix acts as a scaffold that accelerates the migration of
mesenchymal stem cells and the initial external callus formation.
The second stage of fracture healing (i.e. callus differentiation) consists of bone and cartilage
formation in different regions of the callus (Figure 1.3b). During the first 24 hours, mesenchymal
stem cells differentiate into fibroblasts, osteoblasts, and chondrocytes. Growth factors (e.g.
growth factor beta (TGF-β), fibroblast growth factors (FSFs)), and bone morphogenetic proteins
(BMPs) can hasten the repair process. TGF-β increases the number of progenitor cells and leads
to rapid callus formation. FSFs and BMPs are necessary for the differentiation of fibrous tissue.
Osteoblasts strengthen the bone by producing the collagen fibrils and minerals. Osteoblasts
secrete collagen fibrils in a random direction and start converting to the intramembranous woven
bone along the bone (Bailon-Plaza and van der Meulen 2001, Shapiro 2008). The differentiation
initiates from the first day of fracture and continues until the lamellar structure is formed and the
matrix strengthens. The weak structure of the woven bone strengthens by the development of
well-organised lamellar bone. When an adequate amount of woven bone has been differentiated

12
and woven bone is able to act as a scaffold, the formation of lamellar bone initiates. Unlike the
osteoblasts that were differentiated along the bone; mesenchymal stem cells begin to differentiate
into chondrocytes in the interior of the callus and close to the fracture surface, after about seven
days. The soft callus increases the mechanical strength of the fracture. After seven days, cartilage
will have formed from the differentiated chondrocytes. Cartilage has an avascular matrix with a
single cell type (chondrocyte) (Bailon-Plaza and van der Meulen 2001). The key event in the
second stage is the formation of cartilaginous tissue (Buckwalter et al. 1996). The stiffness of the
bone tissue in the second stage has increased from the first stage.
After ten to twelve days, the third stage (i.e., ossification) is initiated (Figure 1.3c). At this stage,
the strength of the callus is close to normal bone. Although the material properties of the callus
are lower than those of bone, its dense composition and large diameter provides enough strength
to resist mechanical loads close to physiological levels. As the cartilage develops and increases
in size, it begins to differentiate into woven bone. This process continues until all of the cartilage
is replaced by bone (i.e. ossification), and at this point, a bony bridge closes the fracture gap.
During the ossification stage, the extracellular matrix, which was formed in the first stage, is
calcified and strengthens the bone structure.
The last stage is the remodelling process (Figure 1.3d), in which bone starts to remodel and
restore its original shape (Buckwalter et al. 1996). The woven bones are gradually resorbed by
osteoclasts and then replaced with lamellar bone. During the remodelling process the callus size
decreases, but the strength of bone increases due to formation of stronger lamellar bone.

13
Figure 1.3. Fracture healing stages: (a) inflammation, (b) callus differentiation, (c)
ossification, (d) remodelling (Bailon-Plaza and van der Meulen 2001).
1.2.1.2 Source of progenitor cells for fracture healing
As mentioned in the previous section, bone regeneration during fracture repair involves several
types of cells. At the fracture site, four major tissue types exist: cortical bone, periosteum,
undifferentiated fibrous tissue and bone marrow (Figure 1.4). The cells essential for fracture
healing are recruited from these tissues (Einhorn 2005, Lee et al. 2008). Although it is known
that the presence of progenitor cells is vital for initiation of bone regeneration, the origin of these
cells is still a subject of debate (Yoo and Johnstone 1998, Ball et al. 2011, Allen, Hock and Burr
2004). There is now good evidence proving that the bone marrow and the inner layer of
periosteum are the two main sources of progenitor cells (Allen et al. 2004, Henrotin 2011, Aubin
1999, Iwaki et al. 1997). Furthermore, the endosteal cells lining the inner cortex are reported to
be the source of repair cells (Shapiro 2008). Moreover, it has been shown that the progenitor
cells can also be found in non-osseous tissues (e.g. surrounding muscle tissues) which are crucial
for bone healing (Liu et al. 2011, Einhorn 2005, Yoo and Johnstone 1998). The progenitor cells
initiating from the surrounding soft tissues play a key role in stabilising the fracture gap. These
progenitor cells duplicate and differentiate into fibrous tissues during the initial stages of healing.

14
Next, fibrous tissues form cartilaginous tissues (i.e. endochondral ossification) which lead to the
development of an early callus and a stiffer bridging.
If there is poor migration, infiltration and differentiation of these progenitor cells, the formation
of bony tissue may be delayed and the fracture gap will be poorly bridged (Yoo and Johnstone
1998, Li et al. 2005, Lang 2011, Allen et al. 2004). The mechanosensitivity of bone may change
with aging and disease (e.g. osteoporosis), and thus bone may not be able to detect the loading
environment as well (Lang 2011). Long-term mechanical load has also been shown to make the
cells less sensitive (Saxon et al. 2005). Consequently, there can be increased resorption with not
enough bone formation, leading to eventual osteoporosis (Li et al. 2005, Allen et al. 2004). The
healing process may be promoted by improving the migration of progenitor cells into the fracture
gap.
Figure 1.4 The essential tissues involved in fracture healing (Einhorn 2005).
1.3 Problem statement and rationale
Although bone is a complex natural material with outstanding mechanical properties and
remarkable self-healing capabilities, in some cases healing is impaired. If the fracture gap
exceeds a critical size, delayed healing and non-union of bones occur (Einhorn 1998, Egermann

15
et al. 2005). Fractures due to osteoporosis result in large health care costs every year, particularly
for elderly people (Egermann et al. 2005, Reginster and Burlet 2006). The bones of
postmenopausal women have increased fragility due to the reduced bone quality (Reginster and
Burlet 2006). The majority of bone reconstructive surgeries are based on non-cell-based
therapies. For example, the current method of treatment is to fill the fracture gap with a graft.
Bone grafts can be obtained from healthy bone in the patient, from donors, or can be created
from synthetic materials with a chemical composition similar to the mineral component of the
human bone. Using grafts has several disadvantages, such as the risk of pain, infection and
rejection, the small amount of bone available for grafting, the possibility of transferring diseases
to the patient and the longer healing time. Immobilization methods have not shown satisfactory
results to treat fractures in osteoporotic bones (Egermann et al. 2005, Chen, Chen and Hsu 2007).
In osteoporotic bones, there is a risk of failure at the implant fixation due to poor bone quality,
bone brittleness and decreased fixation strength (Moroni et al. 2006). The grafts prevent soft
tissue from growing into the fracture site, which delays the healing process. Due to these
drawbacks, the necessity of a rapid, less-invasive fracture repair technique has been highlighted
for difficult-to-heal fractures (Egermann et al. 2005, Chen et al. 2007, Wade and Richardson
2001).
It has been well proven that bone is able to optimise its mechanical properties to different
physical conditions. Defining how mechanical stimuli modulate tissue differentiation during
skeletal healing can improve the treatment of orthopaedic injuries and determine the effect of the
mechanical environment on skeletal repair. There is inadequate evidence of how mechanical
loads are transferred to the tissues or are sensed by the cells. An understanding of bone
mechanobiology could help developing new clinical therapies for different applications, such as

16
improving the design of implants and scaffolds, and developing cell-based treatments with
pharmacological approaches to repair fractures in osteoporotic bones.
Several animal models have been used to study the influence of mechanics on bone healing. In
vivo models have been used for many years to investigate the effects of mechanical conditions
on bone healing. The focus of these models was to explore the effect of different factors on the
healing outcome, and not during the repair. Factors such as mechanical stability,
interfragmentary movement, frequency of the loads and the number of loading cycles and
fracture size were investigated using in vivo models (McKibbin 1978, Goodship et al. 1998,
Claes, Eckert-Hubner and Augat 2003, Claes et al. 1997).
In order to explore the effect of mechanical loads on the healing process, special techniques are
required to measure the mechanical stimuli within the tissues. These measurement techniques
should not disrupt the healing process and be able to give accurate data about the fracture site.
Although several methods have been developed to find the mechanical conditions in the healing
tissue indirectly (e.g. measuring interfragmentary movement and load sharing between implant
and bone), these methods were unable to measure the mechanical behaviour within the whole
tissue (Goodship and Kenwright 1985, Augat et al. 2008). Recently, imaging techniques (e.g.
µCT) have been used to determine the change in the architecture, density, and local mechanical
environment within the tissue during the healing process (Taiani et al. 2010, Zhang and Yokota
2011, Bishop et al. 2006).
It has been reported that axial torsion leads to a delayed healing in several studies with transverse
and oblique fractures, e.g., transverse fracture in long bones of rabbits and sheep, and oblique
fracture in dogs (Augat et al. 2003, Yamagishi and Yoshimura 1955, Aro, Wahner and Chao
1991). However, Bishop et al. (2006) used a sheep model with a 2.4 mm transverse tibia

17
osteotomy to determine the effect of axial torsion on the bone repair process. It was hypothesised
that the torsional load does not necessarily lead to delay in fracture healing. A 7.2 degree rotation
(0.5 Hz) was applied to sheep tibiae with a maximum interfragmentary principal strain of 25 %.
Another group was subjected to axial compression load of 360 N (0.5 Hz) with a maximum
principal strain of 25 %. After eight weeks of healing, the histological results of the control
group (no motion), axial compression group and the torsional group were compared. The axial
torsion was found to stimulate differentiation of cartilaginous tissue in the interfragmentary gap
and bony tissue was observed in the internal and external callus sites, whereas less cartilaginous
tissue was observed in the no motion group (Figure 1.5). The four-point bending stiffnesses
were calculated: the group subjected to the axial torsion were stiffer compared to the axial
compression group and had a slightly higher stiffness compared to the control group (Bishop et
al. 2006).
Figure 1.5. (a) Sheep tibia subjected to torsion. (b) Histological section showing the
cartilaginous formation in the interfragmentary gap and the bony tissue in the internal and
external callus (Bishop et al. 2006).
A finite element model of an ovine tibia with the same fracture gap (2.4 mm) subjected to axial
torsion (Isaksson et al. 2006) was created using the biphasic mechanoregulatory algorithm

18
proposed by Prendergast et al. (1997). The model's predictions were compared to the
experimental observations. The algorithm was able to predict the gap bridging, and the healing
patterns were close to the in vivo results (Bishop et al. 2006). In another in vivo study, the effect
of lateral loading on the healing rate of the open wounds was investigated. It was hypothesised
that the cyclic lateral loading may lead to accelerated wound closure. The 0.5 mm diameter
wounds were generated surgically in the left and right femur necks. The load was applied for
three minutes per day for three consecutive days to the left knee, while the surgical holes in the
right femurs were used as control (no motion). Comparing the results, the healing process was
accelerated throughout the femur, especially in the femoral midshaft and neck compared to the
control group (Figure 1.6). According to peripheral quantitative computed tomography (pQCT)
images, the mineral density was increased in the loading group (Zhang and Yokota 2011). The
mechanical testing revealed that the stiffness of the femur increased by mechanical loading. It
was concluded that knee loading could lead to faster healing in surgical wounds in the femur
neck and midshaft. Developing a loading device for human use to load the knee laterally may
help to reduce the risk of fracture and lead to enhanced healing. However, the load magnitudes
and proper loading conditions should be investigated for human use.
Taiani et al. (2010) investigated a burr-hole fracture murine model treated with embryonic stem
cells implanted in a collagen scaffold at the fracture site. A 0.7 mm burr-hole fracture, with a
depth of 1.07±0.21 mm, was drilled through the medial cortex and the medullary cavity of the
proximal tibia. The centre of the hole below the top of the epiphysis was 2.49±0.26 mm (Taiani
et al. 2010). The burr-hole fracture was filled with collagenous scaffolds seeded with embryonic
stem cells, and the mice were allowed to move immediately after surgery. During 4 weeks of
healing µCT imaging was used to track the healing progress. There was enhanced recovery

19
observed at the fracture site with stem cell seeded scaffolds compared with controls, and an
increase in trabecular number and trabecular bone volume (Figure 1.7) (Taiani 2012).
1 week 2 weeks 3 weeks
(c)
(a) (b)
Knee loading
Figure 1.6. (a) Lateral loading of the left knee, (b) µCT images of burr-holes in control
group with delayed healing, and (c) loaded group with promoted healing. Bar = 1 mm
(Zhang and Yokota 2011).
Day 0 1 week 2 weeks 4 weeks
(b)
(a)
Figure 1.7. µCT images demonstrating the healing process and bone regeneration during 4
weeks: (a) in the control group, (b) collagenous scaffold seeded with ES cell-derived
osteoblasts implanted into the fracture site (Taiani 2012). Bar = 1 mm.
There are a tremendous number of variables involved in experimental investigations of bone
mechanobiology (e.g., the type of mechanical load, frequency, material properties of the scaffold

20
and pore size) that make them expensive, complicated and time consuming. As a result,
computational algorithms of mechanobiological processes have become an essential tool to
determine the mechanical environment within the tissues without disturbing the healing process
(e.g. stress, strain, fluid velocity) and develop a more complete understanding of the mechanical
factors involved in fracture healing. Mechanoregulatory algorithms quantitatively relate the
mechanical stimuli with the tissue behaviour using in vivo and in vitro data (Prendergast et al.
1997, Sandino and Lacroix 2011, Claes and Heigele 1999). Hence, both experimental and
computational models are essential to develop more realistic and sophisticated computational
models.
The following steps are required to develop computational models: (1) geometric reconstruction
of the in vivo model, (2) application of the in vivo loads and boundary conditions, (3)
determination of the material properties of the tissues using histological techniques, fluorescent
labelling and µCT imaging, and (4) model validation against in vivo models of fracture healing
using µCT imaging, mechanical testing and histological slides (Epari et al. 2006, Lienau et al.
2005, Claes and Heigele 1999, Smith-Adaline et al. 2004). Figure 1.8 shows how computational
and experimental studies can be used to develop a mechanoregulatory model.
Some of the proposed algorithms considered soft tissues as single-phase (solid) linear elastic
materials and neglected the effect of the interstitial fluid. These algorithms were regulated by
strain and hydrostatic pressure or just the deviatoric strain (Carter and Wong 1988, Carter,
Blenman and Beaupre 1988, Claes and Heigele 1999). These algorithms could not predict the
bone healing pattern when bone was subjected to axial torsion (Isaksson et al. 2006). Due to the
biphasic nature of soft tissues, considering the effect of fluid flow makes the models more

21
realistic. Using poroelastic analysis, the mechanical stimuli such as fluid velocity, fluid
pressurization, and tissue stress and strain can be explored over time within the tissue.
Several biphasic algorithms were later proposed. As an example, the algorithm proposed by
Prendergast et al. (1997) was based on the interstitial fluid velocity and octahedral shear strain.
Unlike the single phase algorithms, this theory predicted the bone healing patterns under axial
torsion (Isaksson et al. 2006). This theory has two constants that weigh the relative sensitivities
for octahedral shear strain and fluid velocity. These empirical constants do not have a physical
meaning and were obtained from trial and error comparisons of model predictions for fracture
healing patterns with in vivo observations. The complexity of biological events makes the use of
trial-and-error methods qualitative at best without a rigorous basis to constants used in the
models.
In addition to the mechanical behaviour, some cellular activities, such as cell proliferation,
apoptosis, callus growth and angiogenesis have been modelled (Sandino and Lacroix 2011,
Isaksson et al. 2008, Prendergast et al. 1997). However, in biological models there are various
simplifying assumptions which should be further developed. For example, the cell migration
rates are not well known and are estimations or obtained from experimental data from other
species. Further, the computational studies need validation against experimental data. Most of
the time, the experiments have not been performed by the same group and the boundary
conditions and mechanical loads applied to the model are not exactly known.
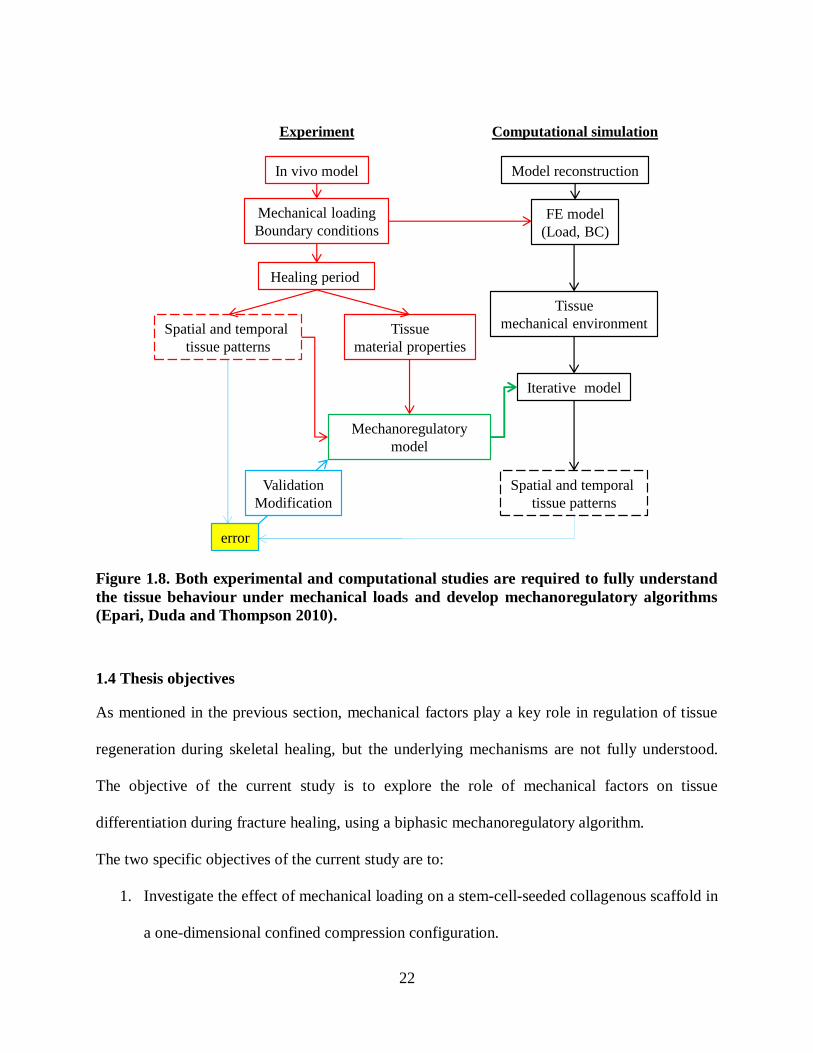
22
In vivo model
Mechanical loading
Boundary conditions
Healing period
Spatial and temporal
tissue patterns
Tissue
material properties
Model reconstruction
Computational simulationExperiment
FE model
(Load, BC)
Tissue
mechanical environment
Mechanoregulatory
model
Iterative model
Spatial and temporal
tissue patterns
error
Validation
Modification
Figure 1.8. Both experimental and computational studies are required to fully understand
the tissue behaviour under mechanical loads and develop mechanoregulatory algorithms
(Epari, Duda and Thompson 2010).
1.4 Thesis objectives
As mentioned in the previous section, mechanical factors play a key role in regulation of tissue
regeneration during skeletal healing, but the underlying mechanisms are not fully understood.
The objective of the current study is to explore the role of mechanical factors on tissue
differentiation during fracture healing, using a biphasic mechanoregulatory algorithm.
The two specific objectives of the current study are to:
1. Investigate the effect of mechanical loading on a stem-cell-seeded collagenous scaffold in
a one-dimensional confined compression configuration.

23
2. Predict the development of differentiated tissues in a tibia burr-hole fracture murine
model with computational mechanoregulatory algorithms.
The long-term goal of our research is to understand the mechanical factors involved in a tibia
burr-hole fracture murine model that is seeded with stem cells for a tissue-engineered repair, as
described by Taiani et al. (2010). The key findings from their study showed that the embryonic
stem cells were effectively differentiated into osteoblasts in a collagen I matrix in vitro (Taiani
2012). Furthermore, the embryonic stem-cell-derived osteoblasts were implanted into the
fracture site and contributed to new bone generation without tumour formation (Figure 1.9),
(Taiani 2012).
Experimental study
(Taiani et al., 2010)
1. Tissue differentiation in a collagenous scaffold seeded with ESCs.
2. Implantation of the scaffold into a burr-hole fracture model of a murine tibia.
(1) Stem cell seeded collagenous
scaffold under confined compression.
Objectives
(2) 3D µCT-based FE model of a
murine tibia with a burr-hole fracture.
Figure 1.9. Objectives of the current study based on the in vitro collagenous scaffold and in
vivo burr-hole murine model.
The tissue differentiation in a collagenous scaffold seeded with stem cells was investigated in a
1D computational model. An axisymmetric model of the loading device in our experimental
studies was created to model the confined compression test. Due to symmetry, an axisymmetric
finite element model was created to investigate the mechanical stimulus transferred to the stem

24
cell seeded gel. A poroelastic analysis was performed to consider the effect of interstitial fluid
within the scaffold. Since the boundary and loading conditions acting on the scaffold may also
play a crucial role in the differentiation process, the collagenous scaffold was subjected to
different magnitudes of pressure (20 kPa, 5 kPa, 1 Hz). The load transfer from the system to the
collagenous gel was investigated, and the distribution of mechanical stimuli was determined
within the solid and fluid phases of the gel. The predicted values of strain and fluid velocity
within the scaffold were found to be negligible in the 5 kPa (1Hz) case compared to the 20 kPa
(1Hz) case. Therefore, 20 kPa pressure was expected to stimulate the cells better and induce
appropriate biophysical stimuli to the scaffold compared to the 5 kPa (1Hz) case. Thus, a
biphasic mechanoregulatory algorithm was used to predict the tissue differentiation within the
scaffold (20 kPa, 5 kPa, 1 Hz). It was hypothesised that the 5 kPa predict less activity within the
scaffold. As expected within the 20 kPa loading case the cell proliferation and differentiation
were augmented. Hence, the use of appropriate mechanical loading led to promoted
differentiation within the scaffold. The loaded collagenous gel may then be implanted into the
fracture site and contribute to an enhanced healing.
In the second part of the thesis, a stem-cell-seeded collagenous scaffold was implanted into the
fracture site of a murine tibia, to explore the effect of mechanical loading on tissue
differentiation. To determine more realistic knowledge about the mechanical environment within
the fracture gap, the actual geometry of the murine tibia was reconstructed from the µCT images.
The reconstructed FE model was based on the in vivo murine model used by Taiani et al. (2010).
The burr-hole defect and the constructed computational model are shown in Figure 1.10.

25
(c) (d)
2.49±0.26 mm
0.71±0.04 mm
(a) (b)
2.55 mm
0.70 mm
1.04±0.21 mm1. mm
Figure 1.10. Location of the burr-hole in the medial aspect of the tibia: (a)
reconstructed FE model based on the in vivo study, (b) µCT image of the in vivo model
(Taiani 2012). A section through the long axis of the burr-hole looking proximally: (c)
reconstructed FE model based on the in vivo study and (d) µCT image (Taiani 2012).
The transfer of mechanical load to a fracture site depends on the quality and structure of the
surrounding bone. Tissue formation in an osteoporotic bone with less strength and increased
porosity is compromised compared to healthy bone. Moreover, the sensitivity of the bone to
mechanical perturbations may have been reduced. However, it is not clear whether the reduced
bone sensitivity is due to a reduced cell number or cell sensitivity. Computational models can be
used to determine the mechanical environment within the fracture site and investigate the factors
that can best promote fracture healing.

26
In the current study, mechanoregulatory algorithms were used and the tissue formation was
predicted under different loading conditions. To simulate both biological and mechanical factors
during bone regeneration, an existing biphasic mechanoregulatory algorithm regulated by
octahedral shear strain and interstitial fluid velocity was implemented into our simulations
(Prendergast et al. 1997, Isaksson et al. 2006). To simulate the recruitment of progenitor cells, a
diffusion process coupled to the poroelastic stress analysis was developed and verified against
the work by Isaksson et al. (2006).
1.5 Thesis overview
The second chapter of the thesis reviews research on mechanoregulatory mechanisms of bone
repair. The general goal of these studies was to investigate the effect of mechanical loading
driving stem cells to differentiate in a fracture. Chapter 2 of the thesis presents a general
description of the principal bone mechanoregulatory theories. The computational simulations
that have been developed and their applications in bone tissue engineering are discussed. Chapter
3 opens with an overview of bone mechanics and soft tissue biphasic theory that is used in
computational mechanoregulatory models. Next, the development and validation of the
implemented mechanoregulatory algorithm are described. The validated algorithm was
implemented into two idealised murine models and the effects of mechanical loading on the
healing process were investigated. In chapter 4, the simulation of the loading device used to load
the stem cell seeded scaffold is explained, and the effect of confined compression was explored
on tissue differentiation. Chapter 5, the reconstruction of murine tibia from µCT images is
discussed, and then a collagenous scaffold seeded with stem cells were implanted into the
fracture site. Using a biphasic mechanoregulatory algorithm the effect of mechanical stimulation

27
was determined in the burr-hole fracture murine model. Chapter 6, summarises the conclusions
and discuss the limitations of the current study and future prospects.

28
Chapter Two: Mechanoregulation Algorithms of Tissue Differentiation in Bone
It is known that mechanical factors can influence the biological synthetic processes in a variety
of tissues, which commonly referred to as mechanobiology. Biological tissues and particularly
those with a mechanical function, e.g. bone, are mechanosensitive, and can optimise their
material properties to different physical conditions (Sandino and Lacroix 2011, Palomares et al.
2009). Mechanical loads significantly affect all stages of tissue regeneration during skeletal
healing, however, it is not fully understood how tissue regeneration can be affected by
mechanical conditions. A number of different mechanoregulation algorithms have been
developed based on empirical comparisons of tissue differentiation patterns observed under
different loading regimes. In the first two sections, the first proposed mechanoregulatory
algorithms are outlined. In the third section, the mechanostat theory and the relationship between
tissue strain levels, and bone modelling and remodelling is presented. Finally, the algorithms
used in the computational simulations to model bone repair are discussed (Figure 2.1). These
algorithms can be broken into: single-phase models, biphasic models, and models with biological
factors.

29
Figure 2.1. Types of the mechanoregulatory algorithms.
2.1 Pauwels theory
The theory proposed by Pauwels (1960) was the first attempt to understand the effect of
mechanical factors on tissue differentiation. He proposed that mechanical forces affect tissue
regeneration through mechanical deformation. The first and second invariants of the stress tensor
are considered as the mechanical stimuli, and used to predict the tissue regeneration pathway.
Two stress invariants, (1) octahedral shear (or distortional) stress S, and (2) hydrostatic (or
dilatational) stress D, represent the change in the tissue shape and volume, respectively
(Timoshenko and Gere 1972). These invariants are defined as follows:
2
31
2
32
2
213
1S 2.1
3213
1D 2.2
where 321 ,, are the principal stresses.

30
The octahedral shear stress deforms the material without inducing any change in its volume (i.e.
it causes octahedral shear strain). On the other hand, when the material is subjected to
hydrostatic stress, there will be a volumetric change without causing any distortion (i.e. it causes
pure volumetric strain).
Combining histological findings and mechanical theories, Pauwels (1960) concluded that the
elongation of extra cellular matrix without any volume change, caused by octahedral shear stress,
promotes collagenous formation. However, a volumetric change in the extracellular matrix
(caused by a hydrostatic stress) will enhance differentiation of cartilaginous tissue. A
combination of volumetric change and distortion within the extracellular matrix will lead to
regeneration of fibrous tissue. Pauwels (1960) believed that bone formation would not be
initiated unless there is a stable mechanical environment (Figure 2.2). The damaged bone
stabilises the environment by formation of fibrous and cartilaginous tissue. Therefore, after
formation of fibrous and cartilaginous tissue, the bony tissue starts to differentiate (Pauwels
1960, Lacroix 2000).
Figure 2.2. A schematic of Pauwels' theory, octahedral shear stress and hydrostatic
pressure used as biophysical stimuli (Pauwels 1960).

31
2.2 Interfragmentary strain theory
The interfragmentary strain theory was developed by (Perren 1979, Tubbs 1981). They suggested
that each tissue type has a specific strength and strain tolerance. In other words, different types of
tissues can absorb different levels of strain. A detailed knowledge of the strain within the callus
would enable the prediction of tissue differentiation. For instance, granulation tissue can stretch
approximately 100 % before rupture, whereas cartilage tolerates strains up to 10 % only, and
bone would fail under strains greater than 2 % (Figure 2.3). Hence, if the strain within the tissue
were more than 2 %, differentiation and maintenance of a bony tissue would not be possible
according to this theory. The healing of bone initiates from the differentiation of tissues with the
highest tolerance to strain (granulation tissue) and continues to cartilage and bony tissue due to
the differences in strain that can be tolerated by these tissues (Figure 2.3).
The interfragmentary strain is the interfragmentary motion divided by the fracture gap size when
bone is considered as a one dimensional (1D) object. Although the movement of the fracture gap
can easily be measured, the interfragmentary theory has two main drawbacks. First, bone fracture
is three-dimensional (3D) and is only being modelled as a 1D problem. Second, the
interfragmentary theory predicts enhanced healing in the fractures with larger gap size. However,
this is in contrast with experimental observations (Claes et al. 1997, Augat et al. 1998).
Figure 2.3. Strain tolerance of different types of tissues (Tubbs 1981).

32
2.3 Mechanostat theory, (Frost, 1987)
The changes in bone such as three longitudinal growth, bone modelling and bone remodelling
are always controlled by a mechanism called mechanostat (Frost, 1987). The mechanism should
check the efficiency of the bone itself, the mechanism that transforms the mechanical signal to
the bone and the sensors that are responsible to sense the signal resulting from the applied
mechanical loads. The strains applied to the bone, depending on their magnitude, may lead to
bone modelling or remodelling. During the modelling process the mass density of the cortical
bone increases; whereas during the bone remodelling the existing cortical endosteal and
trabecular bone is absorbed in some regions and will be replaced by new bony tissue. The bone
modelling and remodelling process will automatically start when strains deviate from certain
setpoints. These strain setpoints change if the responsible agents for sensing or transmitting the
mechanical load to the bone are damaged (e.g. postmenopausal, osteoporosis and osteogenesis
imperfecta). Diseases affect the sensitivity of the cells and make them insensitive or over-active
(Frost, 1987).
A number of set points have been identified to describe bone behaviour: (1) minimum effective
strain (MES), (2) the strain range that lead to bone loss due to an imbalance in the remodelling
process and bone resorption outpaces bone formation ([0,MESr]), (3) the strain higher than this
setpoint lead to bone modelling and bone will be added to surfaces (MESm), (4) the strains
higher than a special setpoint that damaged bone and bone repair should initiate (MESp), and (5)
the bone fracture strength (Fx), (Figure 2.4). When bone is subjected to strains lower than MESm
it is known to be in a homoeostatic state in which bone remodelling is in equilibrium (AW,
adapted window). There is a balance between bone resorption and formation (dead zone or lazy
zone), and there is no net change in bone mass. If bone is under a mild over load (MOW, mild

33
over load window), the bone formation outpaces bone resorption. In case of higher rates of
strain, where micro-damage occurs woven bone formation initiates (POW, pathological overload
window), (Figure2.4).
When the bone is not in the lazy zone, the bone cells will be activated and the bone formation
and absorption will continue until they reach to the homeostatic state. Frost compared this
mechanism with a thermostat, in which the preset temperatures are like the setpoints in a bone. In
case of an increase or a decrease in room temperature, the system will work to raise the
temperature back to its target value.
MES MESm MESp Fx
AW MOW POW
remodelling
lamellar drifts
woven bone drifts
Form
atio
nR
esorp
tion
Figure 2.4. Graph shows the mechanostat theory proposed by Frost (Frost, 2003). The
relationship between the tissue strain level and change in bone mass is presented. AW,
MOW and POW refer to the dead zone, mild overload and excessive load, respectively.
MESm and MESp are the bone modelling and the micro-damage thresholds, respectively.
Fx is the bone fracture strength. These setpoints vary between individuals and are
hypothesised to be genetically determined.

34
2.4 Computational simulations of tissue differentiation
2.4.1 Single solid phase model (Carter’s theory, 1988)
Carter et al. (1988) proposed a mechanoregulation theory based on the octahedral shear stress
and hydrostatic (or dilatational) stress (Carter et al. 1988, Carter and Wong 1988). It was the first
time that finite element analysis (FEA) was used to make quantitative predictions of tissue
differentiation. In this algorithm, bone was considered as a linear elastic single phase material
(solid). Equation 2.3 was used to predict tissue differentiation:
kDSI 2.3
where I was defined as the “osteogeni index” representing the tendency for ossification, S was
the octahedral shear stress, D was the hydrostatic stress, and k was an empirical constant to be
determined.
High values for I represented bony tissue differentiation and accelerated ossification, whereas
low values predicted artilaginous tissue differentiation. ased on Carter‟s theory, fi rous tissue
differentiation occurred in areas subjected to high levels of shear stress. On the other hand,
cartilage differentiation occurred in areas under high hydrostatic compressive stress and low
amount of shear stress. Good vascularity, low shear and high hydrostatic stress stimulate bone
formation. It was found that in areas under low shear and hydrostatic stress levels along with
poor vascularity, there would be no bone formation and only cartilage would differentiate
(Figure 2.5).
The model was further modified to consider the number of load cycles (n) to simulate tissue
differentiation and gradual change of tissue strength (Figure 2.6). The obtained equation is
shown as follows (Carter and Wong 1988):

35
c
i 1
iii kDSnI , c1,2,...,i . 2.4
where in showed the number of loading cycles, and i was the discrete loading conditions, k was
an experimental constant.
A two-dimensional FE model of a femoral midshaft rabbit osteotomy with an external callus was
created. The model was subjected to axial and bending loads. The octahedral shear stress and the
hydrostatic stress were calculated from the computational analysis. Different values of k (0, 0.5.
1, 2) were used to find the osteogenic index (I). Depending on the obtained values of I, contour
plots of the osteogenic index were used to predict areas of endochondral ossification (higher
values of I) and cartilaginous formation (lower I values). The contours were then compared with
the available histological slides. The best match was found using the value of 2 for k. Another
computational study was performed to predict the tissue patterns in an idealised long bone and
the proximal femur. However, comparing the numerical results with the available histological
slides, it was concluded that k could vary between 0.3 and 1. Therefore, for each simulation
different values of k were used to predict tissue patterns (Carter, Wong and Orr 1991).
Carter et al. (1998) proposed another theory which was based on tensile strain and hydrostatic
stress, (Figure 2.6). According to the modified algorithm: (1) low hydrostatic stress and tensile
strain promote direct intramembranous ossification, (2) high tensile strain and hydrostatic
compressive stress stimulate the formation of fibrocartilage, (3) high tensile strain promotes the
development of fibrous tissue, and (4) hydrostatic compressive stress is the stimulus for
cartilaginous tissue differentiation.
An axisymmetric FE model of a long bone fracture was created. The cortical bone was modelled
as a rigid body and other tissues (e.g. fibrous tissue, cartilage and bone) were assumed to be

36
linearly elastic and isotropic. The axial compressive load was applied to the cortical shaft. The
patterns of tensile strain and hydrostatic stress were determined at the fracture site. It was
concluded that bone formation occurs in the areas that cartilage tissue existed. Bone formation
might be delayed by intermittent hydrostatic compressive stress and can be promoted by tensile
strain (Carter and Wong 1988).
Bone is a biphasic material consisting of a solid and fluid phase with viscoelastic properties.
However, both soft and bony tissues were modelled as a single phase, linearly elastic material in
Carter‟s algorithm. Hen e, this model may not e a le to fully apture and explain the tissue
differentiation processes.
Figure 2.5. The effect of mechanical loading and vascularity on bone differentiation (Carter
et al. 1988).

37
Figure 2.6. Carter's mechanobiology theory. Principal tensile strain and hydrostatic stress
history are the key biophysical stimuli n p 2001).
2.4.2 Single solid phase model (Claes and Heigele, 1999)
Claes and Heigele introduced a new hypothesis, which relates the local tissue formation in a
fracture gap to the local stress and strain (Claes and Heigele 1999). An axisymmetric FE model
of a sheep fracture with external callus was used to calculate stresses and strains in the callus.
The initial connective tissue was modelled as a hyperelastic material and other parts (e.g. soft,
intermediate, and stiff callus, ossification and cortex) were considered linear elastic.
It was believed that cartilage forms in regions experiencing excessive hydrostatic compressive
stress; fi rous tissue differentiates under high tensile strain levels and fi ro artilage forms from a
combination of hydrostatic pressure and tensile strain (Claes and Heigele 1999). The proposed
algorithm by Claes et al. (1999) is similar to the one proposed by Carter et al. (1988) are similar.
It was the first time that the thresholds were quantified to predict what tissue would form. Figure
2.7 illustrates the rule of tissue differentiation relating mechanical stimuli to the tissue types as
hypothesized by Claes and Heigele (1999).

38
Figure 2.7. Single phase model introduced by Claes et al. (1999) in which hydrostatic
pressure and strain are the key biophysical stimuli.
2.4.3 Single solid phase model (Gardner et al., 2000)
A 2D FE model based on orthogonal radiographs of a diaphyseal tibial fracture was created to
find the maximum principal stress at four critical stages of the bone healing process (Gardner et
al. 2000). The callus was divided into different regions representing different types of tissues.
The material properties were defined based on the tissues observed in histological slides and
updated before each healing stage. The tissue patterns were correlated with the stress and strain
distributions. It was hypothesized that high levels of principal stress may decrease the tissue
stability and delay the healing process. The maximum strain in the interfragmentary gap was
computed during the healing process. According to the results, the strain decreased from 70 % to
2 % during two months of repair. Since the material properties used during the analysis were
based on tissues observed in the histological slides, the predicted stress and strain patterns were
more accurate when compared to previous algorithms. However, the data was obtained from
only one patient and no comparisons were done using other patients. Furthermore, the complex
3D fracture was modelled in 2D, which may decrease the quantitative predictions of the
simulation.

39
2.4.4 Biphasic model (Kuiper et al., 1996-2000)
In 1996, Kuiper et al. created an axisymmetric model of human bone. The tissues were
considered as biphasic materials, composed of solid and fluid phases, to incorporate the effect of
the interstitial fluid movement. The fluid shear strain and shear stress were used as biophysical
stimuli to drive tissue differentiation. Moreover, bone resorption was also predicted using the
strain energy (Kuiper et al. 1996, Kuiper et al. 2000a, Kuiper et al. 2000b). The mass turnover
was determined as follows:
refS
ρ
Uρθa
dt
dρ 2.5
where θ is a time constant, ρa is the internal free surface per unit volume, U denotes the
tissue level strain energy, ρ is the apparent density and refS is a reference value of ρ
U . The
strain energy density would be:
σε 2
1U 2.6
where ε is the strain tensor and σ is the stress tensor. Resorption ( 0dt
dρ ) would occur if
refref Ss1ρ
USs1 and 0.35s (Van Rietbergen et al. 1993).
Kuiper et al. (2000) deduced that in a fracture callus, strain plays an important role to stimulate
tissue differentiation. Fluid shear stress (τ) an e found using the Carman-Kozeny equation:
3
2
int ndl.dA
dAkGμ
2.7

40
where dl.dA
dAint is known as the specific surface (s), dA is the cross sectional area, intdA is the
internal surface, dl is the infinitesimal length, k is the tissue permeability; n is the porosity, µ is
the fluid viscosity, and G is a constant. Solution of this equation determines the specific surface s
and is then used to find the fluid shear stress τ:
s
pη
2.8
where
z
p,
y
p,
x
pp is the pressure gradient. A schematic of the algorithm is illustrated in
Figure 2.8. An axisymmetric model of a fracture callus under different compressive loads was
modelled to investigate the healing patterns. The cyclic loads were applied to the cortical bone.
According to the numerical study, the amount of fibrous tissue was increased in case of
increasing the magnitude or duration of movements. However, the healing was delayed by
increasing the magnitude of movements (Matsushita and Kurokawa 1998, Goodship and
Kenwright 1985).
Figure 2.8. Biphasic model introduced by Kuiper et al. (2000a), using fluid shear strain and
stress as the biophysical stimuli.

41
2.4.5 Biphasic model (Prendergast et al., 1997).
Prendergast et al. (1997) used a biphasic FE model to understand the differentiation of tissues at
the implant-bone interfaces. Octahedral shear strain and interstitial fluid flow were considered as
the mechanical stimuli and have been successfully used to predict the key events during fracture
healing. The poroelastic theory was used to investigate the fluid flow and the tissue strain (Mow
et al. 1980, Prendergast et al. 1997). The combination of biophysical stimuli of tissue shear strain
and fluid flow act in concert as the mechanical stimulus (S):
b
v
a
γS oct 2.9
where S is the mechanical stimulus, octγ is the octahedral shear strain, v is the magnitude of the
interstitial fluid flow velocity; and a and b are empirical constants ( 3.75%a and
sμm
3b )
(Lacroix et al. 2002). The octahedral shear strain can be determined as follows:
2
23
2
31
2
21oct εεεεεε3
2γ 2.10
where are the principal strains (Timoshenko and Gere 1972).
In Figure 2.9, the curved solid line shows that, with a state of high shear strain, mesenchymal
stem ells would differentiate into fi rous onne tive tissue. The red dashed line shows that
when the tissue is subjected to a lower mechanical stimuli bone formation is promoted. The
interfragmentary motion within the gap reduces and fibrous and cartilaginous tissue will then
differentiate into bone. According to the algorithm, negligible magnitudes of mechanical stimuli
lead to tissue resorption.

42
Figure 2.9. Proposed biphasic algorithm by Lacroix et al. (2002) in which octahedral shear
strain and fluid velocity are the biophysical stimuli.
Table 2.1. Mechanoregulatory stimulus for tissue differentiation (Prendergast et al. 1997).
Mechanoregulatory stimulus (S) Predicted tissue differentiation
S>3 fibrous connective tissue
3>S>1 chondrocytes and cartilage
1>S>0.267 osteoblasts & immature bone
0.267>S>0.01 mature bone & remodelling
S<0.011 Resorption
Depending on the loading conditions, various magnitudes for the mechanical stimulus, S, are
possible which can lead to fibrous tissue, immature and mature cartilage, and immature,
intermediate and mature bone differentiation (Lacroix et al. 2002). High values for S promote the
differentiation of mesenchymal stem cells into fibrous tissues 6S3 , intermediate values
stimulate cartilage differentiation 3S1 , and low levels lead to formation of immature

43
1S0.267 and mature bony tissue 0.267S0.011 . Different values for S representing
different types of tissues are shown in Table 2.1.
2.4.6 Biphasic model (Sandino and Lacroix, 2011)
Sandino and Lacroix (2011) made further modifications to the model introduced by Prendergast
et al. (1997). The purpose of their study was to explore the effect of mechanical perturbation and
perfusion flow on tissue differentiation in a calcium phosphate-based glass porous scaffold. The
octahedral shear stress within the tissue and the shear stress within the fluid (instead of fluid flow
velocity) were used as mechanical stimuli. The fluid shear stress was used instead of fluid
velocity to investigate the effect of fluid perfusion into a scaffold. The solid phase was modelled
as a linear elastic solid and the fluid phase was considered as a Newtonian fluid. In the fluid
model different levels of viscosity simulated tissue regeneration. In addition to the prediction of
tissue differentiation and resorption (e.g. fibrous, cartilaginous and bony tissue), the prediction of
cell death was also added to the algorithm proposed by Prendergast et al. (1997). The variation of
fluid shear stress was observed during the differentiation process. Sandino and Lacroix (2011)
concluded that the variations of fluid shear stress should be controlled in vivo to avoid a delay in
the formation of new tissues. The modified graph is illustrated in Figure 2.10. The
mechanoregulation index M can be represented by the equation:
b
η
a
γM oct 2.11
where octγ is the o tahedral shear strain, τ is the fluid shear stress, and a and b are the empirical
constants (a=0.0375, b=10 [MPa]). Different intervals for M representing different types of
tissues are shown in Table 2.2.
Fluid shear stresses were found using Newtonian fluid formulation:

44
2
0
0
02
0
0
02
vμη
21
31
32
2.12
where µ is the fluid viscosity,
zv,
yv,
xvv is the velocity gradient, and
1ζ and 3ζ
are the minimum and maximum principal fluid stresses, respectively.
Figure 2.10. Biphasic model introduced by Sandino and Lacroix (2011) in which fluid shear
stress and octahedral shear strain were used as biophysical stimuli.
Table 2.2. Biphasic model prediction of tissue differentiation by Sandino and Lacroix
(2011).
Mechanoregulatory stimulus (M) Predicted tissue differentiation
M>6 o τ<0.01 no tissue formation
3>M>6 fibrous tissue
1>M>3 Cartilage
1>M>0.001 Bone

45
2.4.7 Models with biological factors
There are many biological factors affecting the fracture healing process: proliferation, migration,
growth factors, tissue growth and cell modelling. Bailon-Plaza and van der Meulen (2001)
presented a purely biological algorithm to study the effect of osteogenic and chondrogenic
growth factors on fracture healing. Mechanical factors were not taken into consideration in this
model. Tissue differentiation was controlled by seven biological variables: concentration of
mesenchymal stem cells, chondrocytes, osteoblasts, osteogenic and chondrogenic growth factors
and the density of a om ined fi rous/ artilaginous extra ellular matrix or a one extra ellular
matrix. A finite difference method was used to solve a system of 2D partial differential equations
based on the presence of osteogenic and chondrogenic growth factors. The temporal tissue
regulation and cellular events were modelled using the finite difference method (Bailon-Plaza
and van der Meulen 2001). The numerical model predicted the distribution of different tissues
over time, the rate of osteogenic growth factor production by osteoblasts, and determined that the
duration of initial release of growth factors are important to have a successful healing and
ossification. The computational prediction of the time points of osteogenic growth factor
concentrations in callus were close to empirical evidence in the rat fracture callus (Joyce et al.
1990). Bailon-Plaza and van der Meulen (2003) further modified this algorithm by adding
mechanical factors to the model. In addition, Geris et al. (2006) used the bioregulatory model to
simulate a fracture healing process in a murine tibia and the computational results were
compared to experimental observations. The concentration of the growth factor over time was
compared with an experimental study done on murine fracture healing (Cho, Gerstenfeld and
Einhorn 2002). The chondrogenic growth factors were mainly observed in the first week of the
fracture and the osteogenic growth factors were observed in the second week of the healing

46
process (Cho et al. 2002, Geris et al. 2006). The major drawback with these models is that there
was no differen e etween the fi rous and artilaginous tissue components, and no distinction
etween fi roblasts and chondrocytes. Geris et al. (2008) further developed the model by adding
the angiogenesis aspect in bone fracture healing.
Another algorithm which models cell migration was proposed by Lacroix et al. (2002). In
general, cells have a random translational motion like the diffusion of the particles in a fluid or a
gas. They modelled cell proliferation and migration using a diffusion equation as an
approximation, which can be expressed as follows:
2
2
2
2
2
2
z
ρ
y
ρ
x
ρD
t
ρ 2.13
where ρ is the ell density and D is a diffusivity constant and is related to the cell proliferation
and migration rate. The migration, proliferation and cell differentiation is dependent on the
obtained cell density. The material properties were updated based on the average of computed
mechanical stimuli (fluid velocity and octahedral shear strain) in the previous 10 days and on the
temporal and spatial cell concentration:
updated
max
cellgran.
max
cellmaxfinal E
ρ
ρE
ρ
ρρE
2.14
where cell is the cell density obtained from equation 2.17, max is the maximum cell density,
updatedE is the Young‟s modulus o tained from equation 2.9 (Prendergast et al. 1997), updatedE is
the Young‟s modulus of the granulation tissue. The same equation was used to al ulate
permea ility and oisson‟s ratio.

47
The fracture healing stages in a human tibia were modelled by combining both cell migration,
and the biphasic algorithm proposed by Prendergast et al. (1997). A 2D axisymmetric FE of a
human tibia was modelled with a biphasic material description. The results were compared with
the patterns of the healing observed in vivo. The model successfully showed that the healing
process would be longer with larger fracture gaps. Moreover, the differentiation of fibrous tissue
was accelerated by applying higher levels of strain to the tibia in the early stages of fracture
repair (Lacroix et al. 2002). Later, this algorithm was used by Isakson et al. (2006) to model the
fracture healing process in an axisymmetric model of an ovine tibia. It was shown that the high
rates of mechanical stimuli increased the interfragmentary movement and delayed the bone
formation or lead to a non-union in the later stages of healing.
Garcia et al. (2007) developed the first mathematical model to explore the mechanical effects on
the callus size and geometry, and tissue differentiation. The model simulated different cellular
events during fracture healing: (1) mesenchymal stem cell migration, (2) proliferation, death and
differentiation, matrix synthesis, degradation, damage of mesenchymal, chondrocyte, fibroblast
and osteoblast, (3) calcification, and (4) remodelling over time (Garcia-Aznar et al. 2007).
Doblare et al. (2003) examined the tissue composition (collagen types, proteoglycans, mineral
and water) during the healing stages to determine the material properties and permeability of
different tissues (Doblare and Garcia 2003). The second invariant of the deviatoric strain tensor
( 2J ) was used as the mechanical stimulus:
22
2 tracetrace2
1J εε 2.15
where ε is the strain tensor.

48
The rate of change of the matrix volume was a linear function of cell density in each tissue. For
granulation, fibrous and cartilaginous tissue, and woven bone the formulation used is:
ii
i
matrix Qρt
V
2.16
where i indi ated the ell type, ρ is the ell density, Q is the matrix production per cell and unit
time, and the matrix production for lamellar bone was calculated as follows:
vrem
i
matrix Srkt
V
2.17
where
r was the formation/resorption of bone matrix volume per available bone surface per
time, vS was the specific bone surface and remk was the percentage of bone surface active for
remodelling.
According to their study, increases in gap movement increase the callus size (Doblare and Garcia
2003). The computational simulation for an axisymmetric sheep model was compared to the
interfragmentary motion in a sheep experiment (Claes et al. 1997). The callus shapes predicted
by their algorithm had a radial thickness approximately equal to the cortical thickness and had an
axial extension in both distal and proximal directions close to the radial thickness. These results
were in agreement with previous empirical studies that observed for healing osteotomies in
larger mammals (Claes et al. 1998, Goodship et al. 1998).
Another study in orporated a “random walk” or sto hasti model of ell migration with a latti e
modelling approach (Perez and Prendergast 2007). In this model, the cell migration was
modelled in two different ways: with and without preferred direction. An implant-bone interface
was investigated by this method and the results were compared with the diffusion model

49
proposed by Lacroix et al. (2002). Both models had very close predictions for the temporal
change of stiffness in the tissue differentiated within the gap. The only difference was that the
“random walk” ased model in the work by Perez and Prendergast (2007) showed greater
variation in the patterns of the differentiated tissues compared to the model proposed by Lacroix
et al. (2002). The diffusion process predicted continuous patterns of tissue differentiation,
whereas the random walk model had more heterogeneous tissue pattern. There were no
histological slides available to compare the diffusion and random walk models and show, which
best matched. However, the stochastic model had a more rapid reduction of the relative
displacement between the bone and implant (Perez and Prendergast 2007). Using the same
parameters resulted in different predictions of cell distribution, due to the random nature of the
stochastic model. This variation is comparable with experimental studies in cell histology.
Checa and Prendergast (2010) developed a model to incorporate both the mechanical
environment and an oxygen supply as regulators of cell differentiation. A mechanoregulatory
algorithm based on the fluid velocity and octahedral shear strain was used to predict tissue
differentiation inside the bone chamber inserted into a rat tibia using a poroelastic analysis
(Prendergast et al. 1997). The effect of angiogenesis in a scaffold with regular morphology was
also simulated. The computational model was able to mimic the growth and remodelling process
of the capillary network formation and investigate its effect on tissue differentiation. A lattice
approach was used to model cell migration, differentiation, proliferation, apoptosis and
angiogenesis were simulated. Each lattice point was either empty or occupied by a cell. Cell
activities were simulated by moving a cell from a lattice point to another (migration), dividing a
cell so that the daughter cell filled the neighbouring lattice point (proliferation), deleting a cell at
a lattice point (apoptosis), and the endothelial cells were linked to form capillaries within the

50
lattice (angiogenesis) (Prendergast, Checa and Lacroix 2010). The vessel growth was controlled
by a mechanoregulatory stimulus. The random walk theory (Perez and Prendergast 2007) was
implemented in the lattice approach to model cell migration. The mechanobiological algorithm
proposed by Prendergast et al. (1997) was used to consider the effect of mechanical loading on
MSCs differentiation and the effect of vascularity was added. As in the original Prendergast
algorithm, bone formation occurred when the mechanical stimuli was low. However, in the new
model (Checa and Prendergast 2009), the cell differentiation depended on the existence of a
nearby blood vessel. In the presence of oxygen, bone could differentiate, however, if blood
vessels were not available only cartilaginous tissues could differentiate (Figure 2.12). Probability
equations were used to simulate the growth direction and length of the blood vessels. The
formation of the vascular network was based on three events: (1) formation of vessel sprouts
from existing vessels or sprouts, (2) the growth of sprouts, and (3) merging a sprout tip to
another sprout tip or sprout. The path of the endothelial cell at the capillary tip determined the
direction of capillary. The endothelial path was itself determined using the random walk theory.
The sprout branching was a function of sprout length, and the rate of sprout growth was a
function of mechanical stimuli (fluid velocity and octahedral shear strain) (Figure 2.11). The
governing equations depend on both mechanical factors and the existence of vascular endothelial
growth factor (VEGF). VEGF stimulates the development of new blood vessels. In the
computational study by Checa and Prendergast (2009), the vessels reached the core of the
scaffold in six days, and after the first week, the rate of vessel growth decreased which is in
agreement with experimental data of Mikos et al. (1993). Predictions based on the model
indicated that higher mechanical loads caused slower formation of a capillary network and led to
a slower bony tissue differentiation (Checa and Prendergast 2010, Checa and Prendergast 2009).

51
Pro
bab
ilit
y b
ran
chin
g
Sprout lengthL(min) L(max)
maxΔL
Rat
e o
f gro
wth
Mechanical stimulus (S)
maxS
1
(a) (b)
Figure 2.11. (a) The relationship between the sprout branching and the sprout length, (b)
the rate of sprout growth as a function of mechanical stimulus (Checa and Prendergast
2009).
Figure 2.12. Mechanoregulatory algorithm proposed by Checa and Prendergast (2009)
simulating tissue differentiation by both the local mechanical environment and the
presence of oxygen from nearby blood vessels.
Isaksson et al. (2008) developed an algorithm that directly coupled mechanical stimuli to cellular
events. Two separate FE models of the mechanical environment in the tissues and a cell model
ran in parallel and data from one model was used to provide key driving factors to the other. The
cellular activities and tissue differentiation depended on the mechanical stimuli predicted
(deviatoric shear strain and fluid velocity). The mechanical stimuli were determined using the

52
Prendergast et al. (1997) algorithm. The cells acted as transducers during tissue differentiation.
In other words, the activities of each cell (mesenchymal stem cells, fibroblasts, chondrocytes or
osteoblasts) were based on the obtained mechanical stimuli and the activities of other cell types.
Each cell type either migrated, proliferated, differentiated or apoptosed. The effects of growth
factors were also implicitly modelled in the cell activities. The cell proliferation was assumed to
decrease linearly with cell density as the space was occupied and there was lack of nutritional
resources. The rate of cell migration was different for each cell type: mesenchymal stem cells
and fibroblasts had high rates, whereas chondrocyte and osteoblasts had low rates. The migration
was modelled using a diffusion analysis, where the migration was controlled by temporal cell
concentration.
The potential of the model was evaluated in a 2-D FE study of a human long bone osteotomy.
Comparing the computational results with the empirical observations showed that the model
predicted several aspects of bone healing such as cell and tissue distributions during fracture
repair, the effects of excessive mechanical perturbations on the healing process, e.g., periosteal
stripping and impaired effects of cartilage remodelling (Figure 2.13).

53
Figure 2.13. The mechanobiological algorithm proposed by Isaksson et al. (2008)
simulating tissue differentiation according to local mechanical environment, cellular events
and matrix production.
Sandino and Lacroix (2011) proposed a model including blood vessel network formation which
was evaluated by simulating a porous scaffold of calcium phosphate based glass with
heterogeneous geometry. The scaffold was subjected to an unconfined compression simulated by
applying displacement at the top nodes and by fixing the bottom nodes. To allow fluid flow
across the boundaries, null pressure was imposed at the nodes on the boundaries. The
angiogenesis and cell differentiation were modelled in the interconnected pores of the scaffold.
In contrast with previous studies, which studied the effect of angiogenesis in a scaffold with
constant pore size and pore distribution, an irregular geometry was used in this simulation.
Micro-CT images were used to determine the real scaffold and bone geometry. Mechanical

54
stimuli values were determined using the algorithm proposed by Prendergast et al. (1997).
Cellular activities including proliferation, migration, differentiation and MSCs and endothelial
cell initialization were modelled using a lattice approach. The MSCs attached to the wall of the
scaffold at the beginning of the simulation, and then started to move randomly. Each lattice point
had a probability, p, for locating a cell. The endothelial cells were assumed to be outside of the
interconnected pores and the empty lattice points had the probability of 0.1 to locate an
endothelial cell. An event could occur with the probability of p, and could not occur with the
probability of (1-p). The probability was based on the mechanical stimulus, S in the lattice point.
Blood vessels were represented by linking the endothelial cell within the neighbouring lattice
points. The angiogenesis simulation was based on two main factors: (1) modelling the direction
and the rate of growth of each vessel, and (2) its probability of branching. Three approaches were
used to study the growth direction: (1) a random direction, (2) the primary direction used in the
whole analysis, and (3) the direction was set toward regions with higher VEGF concentration.
The vessel growth rate depended on the mechanical stimulus at the beginning of the blood
vessel. Vessel branching was determined according to its length. Tissue differentiation was
predicted based on the combination of both mechanical stimulus and the vessel network
formation. Different types of tissues were predicted within the scaffold subjected to unconfined
strain; however, osteogenesis was not predicted in most of the pores at the center of the scaffold.
Such understanding of tissue differentiation based on angiogenesis and mechanical stimulus
might lead to better scaffold developments. A schematic of the proposed algorithm is shown in
Figure 2.14.

55
Figure 2.14. Mechanoregulatory algorithm proposed by Sandino and Lacroix (2011)
simulating tissue differentiation based on mechanical stimuli, cellular events and
angiogenesis.
Nagel and Kelly (2010) extended the algorithm proposed by Prendergast et al. (1997) to
investigate the effects of mechanical loading on collagen fibre directions and tissue
differentiation. An anisotropic biphasic material model was used to simulate an idealised rat
femur fracture model subjected to cyclic bending. The mechanical stimulus was based on fluid
velocity and octahedral shear strain. Cell migration and proliferation were modelled using a mass
diffusion equation. Angiogenesis and the effect of growth factors, or any other biological factors,
were not modelled. The material properties were updated based on the average of computed
mechanical stimuli in the previous 10 steps (days) and on the temporal, cell concentration as well
as the fibre orientations.
Fibre directions were updated according to the predicted unit vectors ( ia ,0 , ig ,0 ) from the last ten
days. The unit vectors represented the preferred fibre directions:

56
n
ni i
n
ni i
n
n
ni i
n
ni i
n
g
gg
a
aa
9 0
9 0
10
9 0
9 0
10
,
,
,
,
,
,
. 2.18
The mechanical stimulus tensor (S):
3
1j js jj vvS 2.19
where S was the stimulus tensor, js were the eigenvalues of S ( 321 sss ), and jj vv was
the basis of its eigenvectors.
Finally, the directions of the fibres were found using the following equations:
2
3
2
2
2
1
321i0,
sss
sssa
321 vvv
2.20
2
3
2
2
2
1
321i0,
sss
sssg
321 vvv
2.21
where is are the positive eigenvalues of the stimulus tensor (S), and i0,a and i0,g are the
averages of the predicted unit vectors in the preferred fibre directions from the last 10 iterations.
The role of mechanical stimuli in regulating the architecture of the tissue during tissue
differentiation was investigated. The fracture model was subjected to cyclic bending loads, and
the non-union predicted by the computational analysis was compared to the histological slides
from developing neoarthroses (Cullinane et al. 2002). The patterns of differentiated tissues were
also compared with other available experimental and computational studies (Cullinane et al.
2002, Cullinane et al. 2003, Hayward and Morgan 2009). The predicted results were compared

57
with empirical and computational studies and were found to be close to the in vivo observations
(Nagel and Kelly 2010). Figure 2.15 is a schematic of the algorithm used by Nagel and Kelly
(2010).
Figure 2.15. Schematic of the algorithm used by Nagel and Kelly (2010) to incorporate
collagen fibre orientations.
The computational simulation of bone healing started by Carter et al. (1988), using a linear
elastic model; and then continued by Claes et al. (1999) with a hyperelastic model. Biphasic
algorithms were then developed and tissues were subsequently modelled as biphasic materials.
The algorithms got more sophisticated over time by introducing aspects such as cell migration,
proliferation, and angiogenesis which were simulated and added to the proposed algorithms.
The first objective of the present study is to explore the mechanical stimuli transmitted to the
cells within a collagenous scaffold under a confined compression test. Since tissues are biphasic
materials, a poroelastic analysis was required to explore both solid and fluid phase behaviour.
Hence, a poroelastic analysis was performed to investigate the temporal change of pore pressure,
fluid velocity and octahedral shear strain within the scaffold. The cell differentiation within a
stem cell seeded collagenous scaffold was also modelled. The biphasic algorithm proposed by

58
Prendergast et al. (1997) was implemented into our FE model to predict the temporal tissue
differentiation based on the octahedral shear strain and fluid velocity. In addition to the
mechanical behaviour, cell migration and proliferation were also approximated using a diffusion
analysis (Lacroix et al. 2002). This approach and the random walk model predicted a close
stiffness during tissue differentiation. However, after each analysis, the random walk gives
different tissue distributions even using the same parameters, whereas the mass diffusion method
always predicts the same tissue pattern. The mass diffusion method was a good approximation of
the differentiated tissue patterns and the overall stiffness. The tissue prediction was modelled
based on the mechanical stimuli and cell concentration using a coupled poroelastic and mass
diffusion analysis (ABAQUS v6.11).
In the second objective of the current study, the biphasic model was also implemented into a
burr-hole fracture model of a murine tibia to determine the effects of mechanical stimuli on
tissue differentiation. The implemented model yielded a good approximation of the cell
mechanobiology within the scaffold and the burr-hole murine fracture model. To have a more
realistic model, the long-term goal is to develop the implemented model by adding more
biological aspects such as using the random walk model, simulating random differentiation of
bony tissues, angiogenesis, effects of growth factors and pharmacological treatments. Various
mechanoregulatory algorithms are summarised in Table 2.3. The algorithm used in the present
study is shown in the last row.

59
Table 2.3. Summary of the mechanoregulatory algorithms of musculoskeletal tissue
differentiation.
Application Materia
l model
Mechanical
stimuli and
Formulation
Biological factors
Migration
differentiatio
n
proliferation
Growth
factor
/ Callus
growth
Vascularisation
(Carter et
al. 1988,
Carter and
Wong 1988)
initial fracture healing
single solid linear elastic
octahedral shear stress(S),
hydrostatic stress (D)
kDSI
observed as a major local factor for bone
differentiation
The theory was used by: (1) diarthrodial Joints (Carter and Wong 1988), (2) development fracture callus (Blenman, Carter and Beaupre 1989).
(Carter et
al. 1998)
fracture
healing, distraction
osteogenesis
single
solid linear elastic
octahedral shear strain,
hydrostatic stress
The theory was used by: oblique fracture callus (Loboa, Beaupre and Carter 2001).
(Gardner et
al. 2003-6)
fracture healing
single solid linear elastic
maximum principal stress
(Claes and
Heigele
1999)
fracture healing
single solid
hyperelastic
principal strain hydrostatic pressure
Kuiper et al.
1996, 2000
fracture healing
biphasic linear
poroelastic
fluid shear stress, shear strain
sp
η
,
3n
2SkGμ
(Prendergast
et al. 1997)
implant interface
biphasic linear
poroelastic
octahedral shear strain,
fluid velocity
bv
aoctγ
S
(Bailon-
Plaza and
van der
Meulen
2001)
fracture healing
MSC diffusion, chondrocyte,
osteoblast
osteogenic, chondrogenic
(Lacroix et
al. 2002)
fracture healing
biphasic linear
poroelastic
octahedral shear strain,
fluid velocity
bv
aoctγ
S
MSC diffusion
(Geris
et al.
2006,
Geris et
al.
2004)
loaded implants
biphasic linear
poroelastic
octahedral shear strain,
fluid velocity
bv
aoct
γS
MSC diffusion
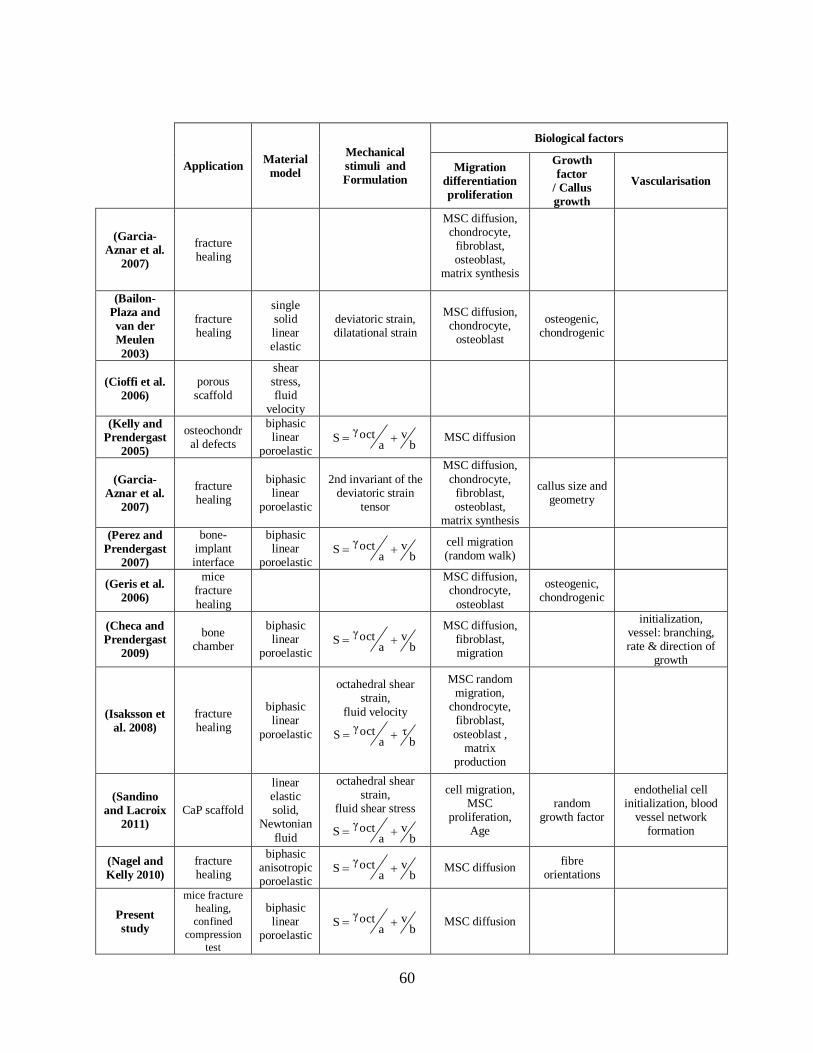
60
Application Material
model
Mechanical
stimuli and
Formulation
Biological factors
Migration
differentiation
proliferation
Growth
factor
/ Callus
growth
Vascularisation
(Garcia-
Aznar et al.
2007)
fracture healing
MSC diffusion, chondrocyte,
fibroblast, osteoblast,
matrix synthesis
(Bailon-
Plaza and
van der
Meulen
2003)
fracture healing
single solid linear elastic
deviatoric strain, dilatational strain
MSC diffusion, chondrocyte,
osteoblast
osteogenic, chondrogenic
(Cioffi et al.
2006)
porous scaffold
shear stress, fluid
velocity
(Kelly and
Prendergast
2005)
osteochondral defects
biphasic linear
poroelastic b
va
octγS MSC diffusion
(Garcia-
Aznar et al.
2007)
fracture healing
biphasic linear
poroelastic
2nd invariant of the deviatoric strain
tensor
MSC diffusion,
chondrocyte, fibroblast, osteoblast,
matrix synthesis
callus size and geometry
(Perez and
Prendergast
2007)
bone-implant interface
biphasic linear
poroelastic b
va
octγS
cell migration (random walk)
(Geris et al.
2006)
mice fracture
healing
MSC diffusion, chondrocyte,
osteoblast
osteogenic, chondrogenic
(Checa and
Prendergast
2009)
bone chamber
biphasic linear
poroelastic b
va
octγS
MSC diffusion, fibroblast, migration
initialization, vessel: branching, rate & direction of
growth
(Isaksson et
al. 2008)
fracture healing
biphasic linear
poroelastic
octahedral shear strain,
fluid velocity
bη
aoctγ
S
MSC random migration,
chondrocyte, fibroblast,
osteoblast , matrix
production
(Sandino
and Lacroix
2011)
CaP scaffold
linear elastic solid,
Newtonian
fluid
octahedral shear strain,
fluid shear stress
bv
aoctγ
S
cell migration, MSC
proliferation, Age
random growth factor
endothelial cell initialization, blood
vessel network formation
(Nagel and
Kelly 2010)
fracture healing
biphasic anisotropic poroelastic
bv
aoctγ
S MSC diffusion fibre
orientations
Present
study
mice fracture
healing,
confined
compression
test
biphasic linear
poroelastic b
va
octγS MSC diffusion

61
Chapter Three: Development and Verification of the Finite Element Model
In this chapter, the development and validation of the finite element model are presented. Section
1 reviews bone mechanics and its behaviour under different mechanical conditions. In section 2,
a brief review of biphasic theory in orthopaedic applications is presented. Section 3 outlines the
theory and algorithms that have been developed to predict tissue distribution during fracture
repair. Next, the finite element model implemented in this study was qualitatively validated
against a previously published numerical-experimental study of Isaksson et al. (2006). Finally,
two case studies simulating experimental tissue differentiation (Bishop et al. 2006, Gardner et al.
2006) are presented as further validation of the differentiation algorithms. A summary of the
overall model development and verification is shown in Figure 3.1.
Figure 3.1. Summary of model development and verification.
3.1 Bone mechanics
The mechanical properties of bone are related to its ability to absorb shocks and resist static or
cyclic compression, tension, torsion and shearing stress loads. A comprehensive description of
the mechanical properties of bone is needed to understand the functional adaptation of the
skeletal system. Uniaxial testing (tension or compression) or three-point bending methods have

62
been used to measure mechanical properties such as bone strength and stiffness. The measured
properties can be compared in diseased versus healthy bone to diagnose musculoskeletal diseases
(e.g. osteoporosis) or estimate fracture risk. The relationship between the applied load and the
deflection of the bone can be found from the load-displacement curve (Figure 3.2). The slope of
the linear region represents the stiffness. In addition to the stiffness, the ultimate load and
displacement, and the fracture load can be determined from the load-deflection curve. The area
under the curve represents the amount of work required to rupture the bone.
Figure 3.2. The force-displacement plot representing bone behaviour (Cole et al. 2010).
The corresponding stress-strain curve can be obtained knowing the cross sectional area and the
original length. The slope of the stress-strain urve within the elasti se tion denotes Young‟s
modulus (stiffness) of the bone and the ultimate stress represents the strength. The area under the
curve represents the bone toughness. Toughness is the amount of energy that bone can withstand
before fracture takes place.

63
The ability of bone to bear mechanical loads depends on three factors: (1) the mass and size of
the bone, (2) the spatial distribution of mass, and (3) the inherent material properties of bone.
Improvement in bone strength may result from an increase in the overall mass, structural
modification and redistribution of bone mass in the regions that experience high loads, or
enhancing the material properties of bone (Cole and van der Meulen 2011, Bouxsein 2006).
Bone is a composite material composed of collagen fibres and mineral particles. The material
properties of bone tissue depend on the mechanical characteristics of the two phases and their
interactions. Bone deformation is a combination of the elastoplastic behaviour of the mineral
particles, and the elastic behaviour of the collagen fibres (Cole and van der Meulen 2011). The
stiffness and the strength of the bone are primarily due to the mineral particles, whereas the
collagen fibres have negligible effect on bone strength (Rivadeneira et al. 2003, Battaglia et al.
2003, Boskey and Coleman 2010). The key role of the collagen fibres in the extracellular matrix
is to augment the toughness and reduce the post yield deformation of the bone (Wang et al. 2002,
Burr 2002, Boskey, Wright and Blank 1999). As its structure provides flexibility to the bone,
collagen defects can increase the fracture risk (Mann et al. 2001).
It has been shown that the elastic behaviour of bone tissue can be altered significantly by
decalcification (Burstein, Reilly and Martens 1976, Boskey and Coleman 2010, Cole and van der
Meulen 2011). In an experimental study, Burstein et al. (1976) decalcified 45 bone specimens
and measured their mechanical properties. The average yield stress decreased from the initial
value of 160 MPa to 76.5 MPa in the decalcified specimens. On the other hand, the yield strain
had marginal change and the slope of the plastic curve did not differ significantly (Burstein et al.
1976).

64
Disease, injury and aging can change the mechanical behaviour of the bone. For example,
mineralization is higher and the size of mineral crystals is larger in older bones than in younger
bones. As shown in Figure 3.3, higher mineralized bones are stiffer and more brittle (lower
deflection) (Currey 2012, Jepsen and Andarawis-Puri 2012, Boskey and Coleman 2010). Hence,
less work is required to fracture bones with higher mineral contents. A bone from a child has
been shown to have larger ultimate deflection and be less stiff compared to an adult bone.
35
30
25
20
15
0
Young‟s m
odulus [M
a]
10
5
200 220 240 260 280 300
Mineral [mg ]
200 220 240 260 280 300
Mineral [mg ]
Wo
rk [
MJ
]20
10
5
2
0.2
1
0.5
(b)(a)
Figure 3.3. Relationship between the mineral content and bone mechanical properties: (a)
by increasing the mineral density, the stiffness increases, (b) while an increase in the bone
mineral content leads to more brittleness (Currey 2012).
During growth, bone mass increases in the first 30 years, then there is no significant change in
the bone mass for approximately 10 years. Around age 40, there is a gradual loss in bone mass.
Bone loss initiates earlier in life for women and increases at a higher rate, especially after
menopause than for males. Due to the accelerated bone loss in women, the risk of fracture is
higher: 40 % for women and 13-25 % for men (Figure 3.4) (McDonnell et al. 2007). Moreover,

65
more importantly the high risk of fracture may be result of bone micro-architecture and the
relative hange in smaller women‟s ones.
Peak bone massAge-related
bone loss
Menopausal
bone loss
Women
Men
0 20 40 60 80
Bone
mas
s
Age [years]
Figure 3.4. Variation of bone mass in men and women across the lifespan (McDonnell et al.
2007).
3.1.1.1 Osteoporotic bone mechanics
As bone ages, there can be an increase in bone fragility, and bone can be more prone to
osteoporosis. The bone fracture risk increases with a decrease in bone mineral density, and
geometric/structural changes. For example, in the post-menopausal period, endosteal bone
resorption increases whereas periosteal apposition slows down; the decrease and thinning of the
cortical wall results which is less resistant to bending (Sharir, Barak and Shahar 2008,
McDonnell et al. 2007). The material properties of cancellous bone from osteoporotic femoral
heads were determined through an unconstrained compression test by Li and Aspden (1997a),
(Figure 3.5). Osteoporotic bones were reported to have less stiffness, Young‟s modulus, mass
density and post yield deformation compared to normal bones (Li and Aspden 1997a). The low
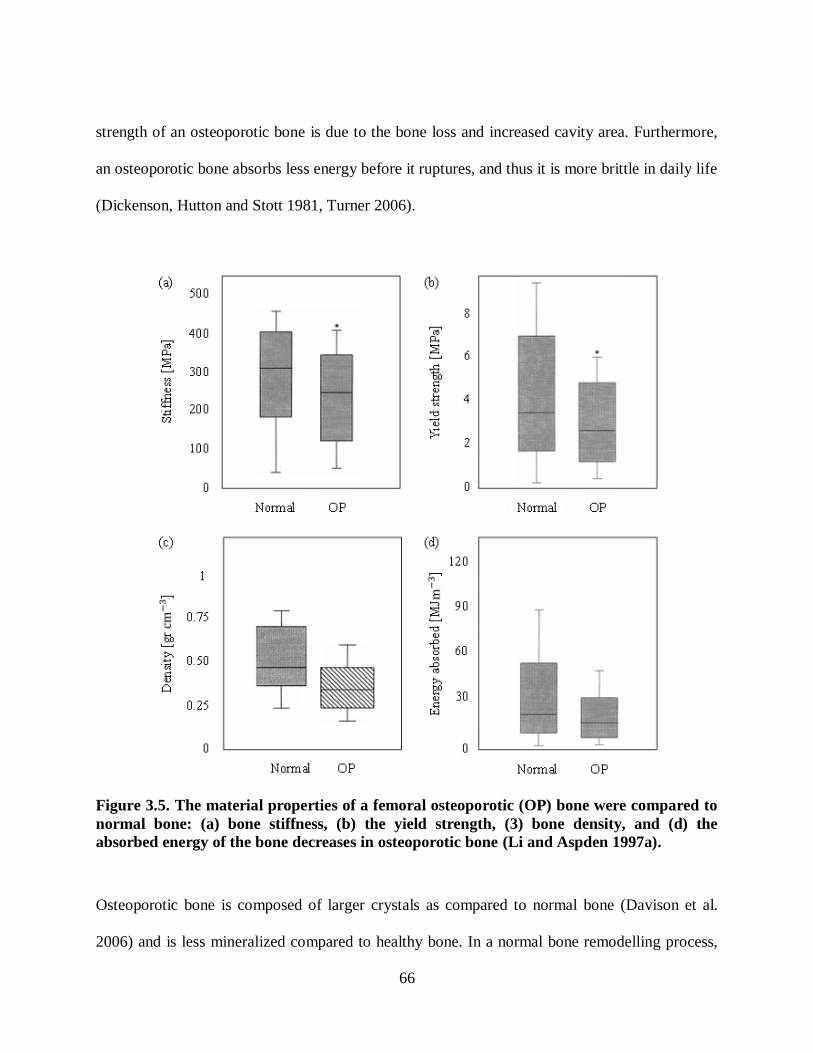
66
strength of an osteoporotic bone is due to the bone loss and increased cavity area. Furthermore,
an osteoporotic bone absorbs less energy before it ruptures, and thus it is more brittle in daily life
(Dickenson, Hutton and Stott 1981, Turner 2006).
Figure 3.5. The material properties of a femoral osteoporotic (OP) bone were compared to
normal bone: (a) bone stiffness, (b) the yield strength, (3) bone density, and (d) the
absorbed energy of the bone decreases in osteoporotic bone (Li and Aspden 1997a).
Osteoporotic bone is composed of larger crystals as compared to normal bone (Davison et al.
2006) and is less mineralized compared to healthy bone. In a normal bone remodelling process,

67
the osteoclasts resorb bone and the cavity is immediately filled with a collagenous osteoid.
Primary mineralization usually begins after a few days and continues until the remodelling cavity
has been 50 to 60 % filled. Next, the secondary mineralization begins at a slower pace and the
cavity will be filled to the maximum level of 90 to 95 % over years. In an osteoporotic bone,
with the increased levels of bone resorption, there is less time for secondary mineralization,
resulting in a lower stiffness (Roschger et al. 2001). Tingart et al. (2003) found that the mineral
density in a 87-year-old man is 0.17 2cm
gr in humeral head and is 0.23
2cm
gr at the surgical neck,
whereas these numbers are 0.5 and 0.55 2cm
gr for a 65-year-old man, respectively (Figure 3.6).
Humeral
head
Surgical
neck
(b)(a)
Figure 3.6. The radiographs of the proximal humerus in (a) a 87-year-old man with low
bone mineral density and, (b) a 65-year-old man with higher mineral density (Tingart et al.
2003).
In osteoporotic patients, the vertical trabeculae are the ones left standing and bear the greater
loads, compared to the horizontal trabeculae that have been absorbed to a greater extent. This
makes the trabecular structure anisotropic and weaker in the horizontal direction (Thomsen,
Ebbesen and Mosekilde 2002). In a computational study, the strain in the head of an osteoporotic

68
femur was 70 % higher and less uniformly distributed compared to the no osteoporotic femur
(Van Rietbergen et al. 2003). The bone porosity increases with age (Figure 3.7) leading to bone
loss and lower bone strength. McCalden et al. (1993) reported that bone porosity is higher for
women compared with men after age 70. The relationship etween the density and the Young‟s
modulus of the subchondral bone plate from the femoral head has been determined in
osteoporotic and normal bones (Figure 3.8) (Li and Aspden 1997b).
30
20
10
020 40 60 80 100 1200
Poro
sity
[%
]
Age [years]
female
0.29827x5.6662y
male
0.13010x2.1774y
male
female
Figure 3.7. The graph demonstrates the relationship between porosity and age for men and
women (McCalden et al. 1993).
0 20 40 60 80 60 80
5
10
15
20
25
normal
10.516.6ρE
icosteoporot
21.422.1ρE
Sti
ffn
ess
[GP
a]
Density [gr ]
Figure 3.8. The relationship between stiffness and density of the subchondral bone plate (Li
and Aspden 1997b).

69
The changes in the structure and the mechanical behaviour of 11 osteoporotic bones were
monitored for 18 months of teriparatide treatment (Macdonald et al. 2011). The high-resolution
peripheral quantitative computed topography used to perform a three dimensional morphological
analysis in osteoporotic women with the mean age of 68.7 ± 12.7 years. The results indicate that
the: (1) quality and the structure of bone declined, (2) total mineral density decreased, (3)
cortical porosity increased, (4) trabeculae became thinner at proximal and distal sites, and (5)
bone strength did not have significant changes during this period (Macdonald et al. 2011). The
values of total mineral density, cortical area, trabecular thickness and the void ratio of the
trabecular and cortical osteoporotic tibia are shown in Table 3.1 (Macdonald et al. 2011).
Table 3.1. The quantitative commutative tomography outcomes at the distal woman tibia
(Macdonald et al. 2011).
Total mineral density Cortical area Trabecular
thickness
Cortical void
ratio
Trabecular void
ratio
180.5 [mg HA/ 3cm ] 689.4 [2mm ] 690 [mm] 0.222 8.1
3.1.2 Bone structure and optimisation
In long bones, the cortical shaft is a thick-walled tube which surrounds and protects the bone
marrow. Moreover, the hollow shape decreases its weight, bulk and metabolic cost to maintain
and has an optimised geometry with enough stiffness to bear high load magnitudes in its day-to-
day activities. During daily activities, bones must endure three common types of loading (e.g.
axial loading, bending, and torsion). In the case of combined loading, the superposition method
can be used to measure the stress within the bone (Taylor et al. 1996, Sverdlova and Witzel
2010). Fracture patterns can reflect how the loads were applied to the bone. Transverse fractures
are produced by excessive tensile forces, spiral fractures can result from torsion, and oblique

70
fractures can be produced by compressive forces (Figure 3.9) (Giotakis and Narayan 2007). If
bone bends due to a lateral impact loading, the side contacted by the load is in compression,
whereas the other side is in tension. Since, bone has higher strength in compression compared to
tension, the fracture will initiate from the side in tension. The fracture gradually advances to the
middle of the bone which has higher resistance (compressed zone). When the crack reaches the
compressive zone, it runs along the directions of the maximal shear stress (at 45 degree to the
longitudinal axis) and creates a butterfly fragment (Sharir et al. 2008, Giotakis and Narayan
2007). The apex of the butterfly fragment denotes the tension side, whereas the base indicates the
compression side (Figure 3.9d).
Figure 3.9. Excessive torque may cause a spiral fracture (a), tension causes transverse
fracture (b), oblique fracture may be created by compression (c), and butterfly fragments
may result from bending (d) (Giotakis and Narayan 2007).
Although bone is an anisotropic, heterogeneous tissue with viscoelastic properties, it is often
considered as a homogeneous and isotropic material. The magnitude of stresses within the bone
can be estimated using the equations in Table 3.2. In the following equations: ζ is the normal

71
stress, η is the shear stress, M is the bending moment, T is the applied torsion, xxI is the areal
moment of inertia about x-axis, J is the polar moment of inertia about the torsional axis, pr and
er are the periosteal (outer) radius and endosteal (inner) radius of the bone. The geometry of
cortical bone alters with age: the periosteal radius increases as a result of periosteum apposition,
whereas the endosteal radius decreases due to bone resorption. The outer diameter of the cortical
section gets larger and the total thickness becomes thinner (less cross sectional area).
Using the simplified bending equation in Table 3.2, three cases were compared: (1) a reference
bone with a solid circular cross section, (2) a young bone, and (3) an adult bone (Figure 3.10).
The material properties of the young bone, with hollow cross section and same periosteal
diameter as the reference bone were compared with the reference bone. Although the young bone
had 25 % less bone mass, it was only 6 % weaker than the reference bone. On the other hand, an
increase of 76 % was observed in the bending stiffness of an adult bone that had a larger outer
diameter (a 25 % increase) but same bone mass as the reference bone due to mass redistribution
(Figure 3.10, Cole and van der Meulen 2011). The bone mass was distributed further from the
bending plane compared to the young bone. Since the moment of inertia ( xxI ) is proportional to
the difference of the radius to the 4th power (
4
e
4
p rr ), the stiffness increased from 94% to 170%.
Table 3.2. Estimation of normal stress under axial, bending and torsional loads.
Cross sectional area Areal moment of inertia Axial Bending Torsion
2
e
2
p rrπA
xx
4
e
4
pxx
2IJ
rrπ4
1I
A
Fζ
xx
px
maxI
rMζ
J
Trp
max

72
periosteumperiosteum
apposition
endosteum
resorptionendosteum
stiffening
young bone adult bone
100% 100% 125%
A 100% 75% 100%
100% 94% 170%xxI
pr
Figure 3.10. Variation in the size and mass distribution affects the bending stiffness of
bone. The middle figure represents a young bone with thicker cortical shell, whereas the
bone with thinner cortical shaft, at the right, represents an adult bone. In the older bone,
resorption of the inner surface and apposition of the outer surface decreased the bone
thickness. The bone mass was distributed further from the bending plane compared to the
young bone.
3.1.3 Mechanical behaviour of cortical bone
Cortical bone is composed of mineral particles embedded in collagen fibres. Due to the
anisotropic distribution of collagen fibres (Fartzel et al. 1992), bone mechanical behaviour is
direction dependant and should be treated as an anisotropic material. The strength and stiffness
of cortical bone are higher when bone is loaded along the longitudinal axis compared with
directions normal to the diaphysis axis (Reilly and Burstein, 1975; Lipson and Katz, 1984). Bone
is an active tissue and can adapt itself to the mechanical behaviour. Hence, daily activities can
lead to bone turnover and micro-damage repair. It has also been shown that it is stronger in
compression than in tension (Reilly and Burstein, 1975). Cortical bone has almost the same
mechanical behaviour in the radial and circumferential directions, and thus can be considered
transversely isotropi . The longitudinal and transverse Young‟s moduli are reported to e a out
17900 and 10900 MPa for human femoral cortical bone, respectively. The ultimate tensile and

73
compressive stresses in the longitudinal direction are about 135 and 205 MPa, whereas for the
transverse direction these values are about 53 and 131 MPa, respectively. The ultimate shear
stress is reported to be 65 MPa. These values demonstrate the relatively low strength of cortical
bone in shear, and the stiffer behaviour in the longitudinal direction (Reilly and Burstein, 1975).
Cortical bone is quite brittle and cannot bear large strains beyond the yield point. The stress-
strain behaviour of human cortical bone is presented in Figure 3.11.
Figure 3.11. The stress-strain curve illustrates that bone is stiffer in the longitudinal
i c ion. Yo ng’s mo l s is h s m in nsion n comp ssion, whereas in
compression bone has more strength (Kutz 2003).
The creep response of cortical bone has been evaluated under low, intermediate and high stress
levels. In low and moderate loading levels, bone returns to its original state, whereas at higher
stress levels, close to the yield point, permanent residual strain will develop (Fondrk, 1988).
Several factors can affect the mechanical behaviour of the cortical bone such as porosity,
mineralization and aging. A marginal increase in the bone mineralization may lead to a higher
Young‟s modulus and strength (Boskey and Coleman 2010, Cole and van der Meulen 2011). In

74
contrast, aging of bone tissue can have a negative effect on the tissue strength. The ultimate
tensile stress and Young‟s modulus of ti ial orti al one were determined in ompression tests
across the lifespan (Burstein et al. 1976). The results indicate a 2 % and 4.7 % decrease per
decade for the ultimate stress and modulus, respectively (Figure 3.12).
Figure 3.12. Yo ng’s mo l s n l im stress of tibial cortical bone decrease with
increasing age. The rate of decrease is higher for ultimate stress compared to the modulus
(Burstein et al. 1976).
Cortical bone is a viscoelastic material exhibiting both viscous and elastic properties. The
loading rate moderately affects the mechanical behaviour of the tissue. The strain rate sensitivity
has been evaluated in a uniaxial tension test. The stress-strain curves were obtained while strain
rate was increasing by six orders of magnitude from 0.001 to 1500 % per second. Daily activities
occur in a range of strain rates between 0.01 to 1.0 % per second and the higher strain rates can
be the result of car accidents or gunshots. The results indicate that Young`s modulus and ultimate
stress change by a factor of 2 and 3, respectively. As shown in Figure 3.13, the strain rate in the
range of daily activities has only a minor effect on the stiffness of the tissue (Kutz 2003).

75
Figure 3.13. The response of cortical bone under different strain rates (Kutz 2003).
3.1.4 Mechanical behaviour of cancellous bone
Trabecular bone is a heterogeneous material, i.e., its material properties vary spatially. In
addition to the inhomogeneity, cancellous bone is anisotropic (i.e. the physical behaviour of the
tissue differs in each direction).
At the end of long bones, cancellous struts and plates transfer large forces to the strong midshaft.
The irregular shape and small size of the struts make the measurement of trabecular bone
diffi ult. The reported Young‟s modulus for tra e ular one ranges from 1 to 19 GPa (Ashman
and Rho 1988, Rho, Hobatho and Ashman 1995). According to Neibur et al. (2000), trabecular
bone ultimate stress is higher in compression than in tension. Both stiffness and strength of the
tissue decrease 10 % per decade (Mosekilde and Danielsen 1987, Lakes and Saha 1979). When
trabecular bone is compressed greater than the ultimate load, the stiffness will be reduced and if
reloaded will develop higher strains (Figure 3.14).

76
Figure 3.14. The stress-strain plot for a load-unload-reload trabecular sample. The loading
s s poin 1, i is nlo poin 2 n lo poin 3. Th ini i l Yo ng’s
modulus from the linear section of the reloading (3-4) is the same as the original Young’s
modulus (1-2) at the beginning, but reduces quickly to residualE . A permanent residual strain
residualε will be developed (Keaveny, Wachtel and Kopperdahl 1999).
Keaveny et al. (1999) concluded that excessive loads cause residual strain and geometric
changes which increase the risk of fracture by significantly decreasing the mechanical
properties of the underlying trabecular bone (Figure 3.14).
3.2 Soft tissue biphasic theory
Soft tissues such as fibrous tendons and ligaments, cartilage, bone and granulation matrix are
composed of a solid matrix and significant amounts of interstitial fluid (van der Meulen and
Huiskes 2002, Van der Voet 1992). To determine the mechanical response of the tissues, the
behaviour of each phase should be considered as well as the interactions between the phases (the
solid displacements and fluid velocities).
The fluid related time dependent structural analysis of soft tissues can be described in two ways:
(1) poroelastic theory, or (2) mixture theory. In the poroelastic theory, material is considered as a
porous solid which is fully saturated with fluid. This theory has been developed to describe the

77
behaviour of soil (Biot, 1941). The biphasic theory considers the material as a more general
continuum mixture of solid and fluid phases. Mow et al. (1980) employed iot‟s (1941)
biphasic theory to describe the two-phase behaviour of soft tissues, used by Ateshian et al.
(1994) to determine the importance of interstitial fluid in the load support mechanism of articular
cartilage (Mow et al. 1980, Ateshian et al. 1994). It has been proven that the poroelastic theory is
equivalent to the biphasic theory when the fluid phase is considered inviscid (Mow et al. 1980,
Simon 1992). Due to the complex structure of soft tissues, complexity of the applied loads and
material properties, and thus limited availability of analytical solutions, the use of numerical
methods and finite element models (FEM) is essential. It has been shown that FEM packages,
e.g. ABAQUS, can be used to analyse the biphasic behaviour of soft tissues with reasonable
accuracy (Prendergast, van Driel and Kuiper 1996, Van der Voet 1992, Wu, Herzog and Epstein
1998). In the following section, the biphasic theory for soft tissues is briefly summarized
(Ateshian et al. 1994, Mow et al. 1980).
The behaviour of tissues in a free draining confined compression test can be explained by the
movement of the interstitial fluid. As shown in Figure 3.15, a ramp displacement was applied to
the articular cartilage (OAB) and kept constant for the rest of the test (BCDE). Immediately after
application of an axial compression load, the stress increases due to the pressurization of the
fluid phase. The fluid is forced out of the tissue as the load increases, and continues to flow even
when the displacement is kept constant. The redistribution of the fluid leads to stress relaxation
and a steady state condition is reached. The stress at the equilibrium state represents the stress
within the solid phase (Mow et al. 1980).

78
Figure 3.15. (a) Movement of fluid when tissue is under a free draining confined
compression test, (b) ramp deformation is applied (increased with a linear ramp to B and
remained constant from B to E, (b) the flow exudes immediately after the deformation is
applied and the stress reaches its maximum amount (B). The fluid continues to flow. The
tissue reaches an equilibrium point and the stress decreases and reaches a plateau (E)
(Mow et al. 1980).
3.2.1 Kinematics
Bone tissue can be modelled as a two-phase material: a porous, linear elastic,
isotropic/anisotropic, homogeneous solid matrix with constant isotropic permeability and an
incompressible, inviscid fluid phase. To define quantities associated with solid and fluid phases,
the superscripts s and f are used, respectively. The tissue volume element (dV) consists of a
sufficient number of solid grains sdV , and volume of interstitial fluid fdV . The medium is
assumed to be fully saturated and the voids completely filled with the fluid. Hence, we get:

79
fs dVdVdV . 3.1
In order to describe the two-phase material, the volume fractions are defined as:
1θθ
n1dV
dVθ
ndV
dVθ
fs
ss
ff
. 3.2
where (n) is the porosity of the medium, equal to the fluid volume fraction, and measures the
void spaces. The void ratio (e) relates the fluid and solid volume fractions:
n1
ne
dV
dVe
s
f
. 3.3
Two measures of densities can be defined for each point: the true and apparent densities. The
true densities (i.e. the constituent mass divided by the constituent volume) are:
f
ff
T
s
ss
T
dV
dmρ
dV
dmρ
. 3.4
On the other hand, the apparent densities (i.e. the constituent mass divided by the tissue volume)
are:
dV
dmρ
dV
dmρ
ff
ss
. 3.5

80
The total apparent density ( ) of the mixture is the sum of solid sρ and fluid fρ apparent
densities, which can be expressed as follows:
fs nρρn1ρ . 3.6
According to the continuum mixture theory, each component has its own motion. Every spatial
position x in the mixture is simultaneously filled by the material solid sX and fluid f
X
constituents at any time t. The fluid particles and solid grains in the solid skeleton are moving
with different velocities in different directions and at different positions. However, these details
are ignored and an average motion for each material point is considered (Truesdell 1966). The
motion of the solid and fluid constituents can be described by a mapping between reference
and current configurations, where s
iX and f
iX are arbitrary material points in the reference
configuration, and s
ix and f
ix in the current configuration:
t,Xx
t,Xx
f
i
f
i
f
i
s
i
s
i
s
i
. 3.7
The average displacement vector can be defined for solid and fluid phases separately as follows:
f
i
f
i
f
i
f
i
s
i
s
i
s
i
s
i
Xt,Xu
Xt,Xu
. 3.8
Then the relative fluid displacement is then given by (Biot, 1941):
s
i
f
i
f
i uuθw . 3.9
where s
iu and f
iu are the primary unknown fields.
3.2.2 Conservation of mass
The balance of mass for the solid and fluid particles can be expressed as:

81
0vtρdiv
t
tρ sss
. 3.10
0vtρdiv
t
tρ fff
. 3.11
where div is the divergence operator.
The solid and fluid constituents are assumed to be intrinsically incompressible. Hence, the
continuity equation for the whole mixture can be obtained by adding equations 3.10 and 3.11 to
give:
0vvdiv ffss . 3.12
3.2.3 Conservation of linear momentum
For the biphasic theory, the solid sσ and fluid f
σ stress tensors can be expressed as follows:
v
ijij
ff
ij
e
ijij
ss
ij
ζpδθζ
ζpδθζ
. 3.13
where e
σ is the elastic stress of the solid, v
σ is the viscous stress of the fluid, and p is the fluid
pore pressure. The elastic effective stress is taken to be positive in tension, whereas pore pressure
in the fluid is assumed to be positive in compression. The fluid is considered inviscid in our
study. Therefore, the viscous shear stress is considered null within the fluid 0vσ . This is an
acceptable assumption because when a fluid flows through a porous material a viscous drag force
much greater than the fluid shear stress is applied to the solid skeleton.
According to the principle of effective stress, the total stress σ of the mixture can be
decomposed in two parts: (1) the pore pressure (p) in the fluid acting on the solid skeleton and in

82
the fluid in every direction (Terzaghi, 1923), and (2) the total stress minus pore pressure stress
that represents the effective elastic stress which is applied to the solid skeleton ( ):
ij
e
ijij pδζζ . 3.14
If the material is assumed to be isotropic with infinitesimal deformations, the linear elastic stress
tensor of the solid phase can be expressed by the Lamè material constants and as:
ij
s
ijkk
se
ij ε2μδελζ . 3.15
where Ɛ is the Green strain tensor for the solid phase. The Lamè material constants are related to
Young‟s modulus sE and oisson‟s ratio s of the solid phase:
ss
ssss
λμ
2μ3λμE
. 3.16
ss
ss
μλ2
λν
. 3.17
The conservation of linear momentum for solid and fluid phases can be expressed as:
sssπσB
divρ
Dt
vDρ s
sss
. 3.18
fπ
fffff
f ζdivBρDt
vDρ . 3.19
where Dt
vD ss
and Dt
D f fv
are the material time derivatives, s
B and f
B are the body forces per unit
mass,s
σ andf
σ are solid and fluid stresses, and s
π and f
π are the local body forces. The local

83
body forces account for the interactions between the solid and fluid constituents that must
satisfy:
s
i
f
i
s
i
f
i vvKvvK
fsππ
. 3.20
where K is the diffusive drag coefficient and is related to the permeability (k) of the tissue, which
is considered constant in our study (Ysart and Mason 1994, Lai, Mow and Roth 1981):
k
θK
2f
. 3.21
Assuming the external body forces and the fluid and solid accelerations are negligible, the
momentum equation for the constituents and the medium is given by:
0πσ
0πσss
ffdiv
div. 3.22
0σσσfs divdiv . 3.23
Substituting the obtained solid and fluid stress equations into the solid and fluid momentum
equations leads to:
pk
t
div
divgradμλpgrad
2
2ss
s
ss
u
0uus
. 3.24
The FE solver ABAQUS v6.11 was used in the current study, in which a porous medium is
modelled as a multiphase material: solid particles and one or more fluids. The fluid phase is
composed of a liquid phase and the other is often gas. The wetting liquid is assumed to be
relatively incompressible, whereas gas is relatively compressible. When the porous medium is

84
considered fully saturated, the voids are completely filled with the wetting liquid (e.g. in the
present study). The elementary volume (dV) is the sum of the grains of solid phase gdV and the
volume of fluid phase fdV . ABAQUS calculates the effective stress, *
σ , as follows:
Iσσ*
auX1Xuw . 3.25
where σ is the total stress, wu and au are pressure stresses in the wetting liquid and the other
fluid, respectively. When the medium is fully saturated, X is equal to 1.0.
The porous medium is modelled by attaching the finite element mesh to the solid phase and the
wetting liquid can flow through this mesh. The mechanical part of the model is based on the
effective stress principal. For the fluid phase, the rate of increase in the fluid mass at a point
should be equal to the rate of mass of fluid flowing to that point (i.e. continuity equation). The
fluid flow is defined y the Dar y‟s law. The mass ontinuity equation is relative to the fluid
pore pressure. A negative pore pressure represents a suction condition. In a transient analysis, a
backward difference operator is used to integrate the continuity equation; the accuracy of the
analysis is controlled by the selected time integration and the maximum pore pressure.
The Young‟s modulus and oisson ratio are defined in A AQUS and then the Lamè material
constants of the solid phase calculated to find the stress in the solid grains. The void ratio (e) and
permeability are also defined in ABAQUS. Since ABAQUS uses the poroelasticity theory, the
introduced permeability in the biphasic model (k) (Mow et al. 1980) needs to be converted to the
poroelastic permeability ( k ):
γkk . 3.26
where
3mm
N0681.9 e is the specific weight of the interstitial fluid (Wu et al. 1998).

85
3.3 Finite element model of mechanoregulation
The following sections describe how mechanoregulatory models were used to simulate cellular
processes and to predict tissue differentiation during fracture repair. Thereafter, the model has
been verified using a previously published numerical-experimental study (Isaksson et al. 2006).
3.3.1 Adaptive mechanoregulation algorithm
In the previous chapter, several mechanoregulation algorithms were reviewed. These algorithms
are based on different mechanical factors and were implemented into different computational
models. Therefore, it was difficult to identify the best one to simulate the process of healing.
Isaksson et al. (2006) developed the FE model of ovine tibia and implemented all of the
proposed algorithms into the model to identify the critical features that should be included,
however, the only algorithm able to predict the healing pattern under torsion was the one
proposed by Prendergast et al. (1997). Other algorithms (Carter et al. 1988, Carter et al. 1998,
Carter and Wong 1988) could not predict healing and callus bridging and the tissue distributions
did not agree with in vivo empirical results (Isaksson et al. 2006). Therefore, the biphasic theory
that is regulated by the octahedral shear strain within the tissue and the velocity of the interstitial
fluid was used for the current study (Figure 3.16) (Prendergast et al. 1997). As described in
Section 2.4.5 the key mechanical stimulus is obtained using the following equation:
b
v
a
γS
foct . 3.27
where S is the biophysical stimulus, octγ is the octahedral shear strain, fv is the fluid velocity,
03750a . and s
μm3b are empirical constants determined from formation of interfacial tissue
at implant surfaces in the condyle of dogs. The mechanoregulatory pathway obtained from the

86
variation of biophysical stimuli within the interfacial tissue during the healing period
(Prendergast et al. 1997). The cell deformation of the solid and fluid phase was investigated by
calculating the octahedral shear strain and fluid flow, which were calculated from a poroelastic
analysis using ABAQUS.
The bone tissues (e.g. cortical bone and bone marrow) and the fracture site were defined as
biphasic materials. At the beginning of the analysis, the fracture site (i.e. callus) was assumed to
e filled with stem ell seeded granulation tissue. The Young‟s modulus, oisson‟s ratio,
permeability and void ratio were used to define the poroelastic properties of the tissues as listed
in Table 3.3. The continuum elements with pore pressure properties were used to mesh the
fracture models (e.g. CAX8P, C3D8P). The boundary conditions and mechanical loads were
applied to the FE model. The poroelastic analysis was conducted and the fluid velocity and
octahedral shear strain were computed at each integration point throughout the fracture site.
According to the predicted mechanical stimuli and based on the algorithm proposed by
Prendergast, the distribution of the differentiated tissue was determined (Lacroix et al. 2002,
Prendergast et al. 1997). High values for S promote the differentiation of mesenchymal stem
cells into fibrous tissues 6S3 , intermediate values stimulate cartilage differentiation
3S1 , and low levels lead to formation of immature 1S0.267 and mature bony tissue
2670S0110 .. (Figure 3.16). The material properties of the intact zone were kept constant
during the analysis.

87
Figure 3.16. Proposed biphasic algorithm by Prendergast et al. (1997); strain and fluid
velocity are the biophysical stimuli.
As bone heals, new tissues with new material properties differentiate at the fracture site. Hence,
in the computational analysis, the gradual change of material properties simulates the tissue
regeneration and the bone repair process. Changing the material properties temporally could be
achieved using an iterative process. Firstly, the poroelastic analysis calculated the mechanical
stimuli based on the bone geometry, applied loads, defined boundary conditions and material
properties (i.e. the unfractured bone, and the fracture site initially composed of granulation
tissue). Secondly, the differentiated tissues were predicted by the algorithm and the properties
were updated (Lacroix et al. 2002, Prendergast et al. 1997). Thirdly, the analysis was continued
until the mechanical stimuli reached a steady state and no further differentiation occured (i.e.
bone healed). The differentiated tissues at the fracture site were also modelled as linear
poroelastic materials. Table 3.3 illustrates the tissue material properties used (Isaksson et al.
2006). The required stages to model the iterative process are summarized in Figure 3.17.

88
Table 3.3. Poroelastic tissue material properties (Isaksson et al. 2006).
Constant (intact site) Variable (fracture site)
cortical
bone marrow granulation
tissue
fibrous
tissue cartilage
immature
bone
mature
trabecular
bone ovine tibia
E [MPa] 15750 2 0.2 2 10 1000 6000
0.325 0.167 0.167 0.167 0.167 0.325 0.325
k
Ns
mm 4
510 0.010 0.010 0.010 0.005 0.100 0.370
e 0.041 4.00 4.00 4.00 4.00 4.00 4.00
Fracture zone is initially filled with
granulation tissue. (step=1)
FE poroelastic analysis
(ABAQUS)
Fluid velocity & octahedral shear strain
in each element
Prediction of tissue phenotype using
the biphasic algorithm
Updating the material properties
Δstepstepstep
Figure 3.17. The iterative model used to simulate fracture repair.
3.3.1.1 User defined subroutine: USDFLD
During the bone healing progression, the material properties of the fractured zone change over
time. This temporal change is a function of the fluid velocity and octahedral shear strain
(Prendergast et al. 1997). A user defined subroutine (USDFLD) was written in FORTRAN, to

89
link the mechanical properties to the field variables (i.e. fluid velocity and octahedral shear
strain). The mechanical stimulus, S, was calculated at each integration point based on the
predicted fluid velocity and strain from the previous step. Therefore, by treating the mechanical
stimulus as the field variable to be computed in USDFLD, the material properties were updated
based on the fluid velocity and strain. USDFLD updated the solution-dependant material
properties of the fra ture zone (e.g. Young‟s modulus, oisson‟s ratio and permea ility) and
imported them into ABAQUS.
The USER DEFINED FIELD comment was used in ABAQUS input file to indicate the
USDFLD subroutine should be called during the analysis. One field variable, with the values
changing from 1 to 5, was chosen to define the tissue type (Figure 3.18). Table 3.4 shows how
the field variable (FV) was defined in the USDFLD. In the table, S is the mechanical stimulus, E
is the Young‟s modulus, ν is the oisson‟s ratio, k is the permea ility.
The field variable was set to be a function of the magnitude of fluid velocity (FLVEL) and
octahedral shear strain. To access the material point data the utility routine GETVRM was used,
and the values of fluid velocity and strain were stored during the analysis. The values defined in
the subroutine were not automatically stored by ABAQUS. In order to have access to the
calculated values, they must have been saved as solution dependant variables (SDV). The value
for fluid velocity was stored as SDV(1), the value of octahedral shear strain was stored as
SDV(2) and the mechanical stimulus, S, was saved as SDV(3). The stored SDVs were then used
in the following step to predict the updated material properties.
The magnitude of fluid velocity and octahedral shear strain were computed within the subroutine
using the following equations:

90
2
3
2
2
2
1 FLVELFLVELFLVELFLVEL , 3.28
2
23
2
31
2
21 EEPEEPEEPEEPEEPEEP3
2oct , 3.29
where FLVEL is the fluid velocity and 1FLVEL ,
2FLVEL and 3FLVEL are the components
of fluid velocity in directions x, y and z; and EEP1, EEP2 and EEP3 are the principal strains. The
velocity, strain and S values were calculated using a poroelastic analysis, at the maximum
loading.
Since the field variable (FV) was history dependent, the obtained FV in step n, was saved as a
new state variable (SDV(n)) and used in the following step (n+1). The number of field variables
(FVs) and state variables (SDVs) were set using DEPENDENCIES and DEPVAR comments.
The number of state variables (DEPVAR) depends on the number of steps in the analysis. The
partial input data is shown in Figure 3.18.
Table 3.4. Dependence of material properties on field variable (FV) in the fracture site.
Material status Material properties
FV Tissue type S E [MPa] ν k
Ns
mm 4
Mature bony tissue 0.267S0.011 6000. 0.300 0.370 1
Immature bony tissue 1S0.267 1000. 0.300 0.100 2
Cartilaginous tissue 3S1 10. 0.167 0.005 3
Fibrous tissue 6S3 2.0 0.167 0.010 4
Granulation tissue Step one 0.2 0.167 0.010 5

91
*Elastic, DEPENDENCIES=1
**_Young‟s modulus and oisson‟s ratio hanging as a fun tion of field variable_**
*Material, name=Scaffold
*Depvar
105,
Elastic, dependencies=1
6000., 0.3, , 1.
1000. 0.3, , 2.
10.,0.167, , 3.
2.,0.167, , 4.
0.2,0.167, , 5.
*User Defined Field
Number of state variable
Number of field variables
Values of field variable
Figure 3.18. A partial input file showing how the field variable was defined in the input file.
3.3.1.2 Smoothing process
As seen in Table 3.4 the Young‟s modulus of tissues an differ y orders of magnitude. Hen e,
while updating the material properties a sudden change might occur from one step to the next.
These rapid changes may lead to convergence problems in computational analysis. To enable
slower changes, a homogenization procedure was used to prevent numerical instabilities (Lacroix
et al. 2002).
The field variable was calculated in each step based on its values in the previous 10 steps (at the
maximum loading). For the first 10 steps, the values of the field variable were averaged with the
FV related to the granulation tissue. Therefore, the most dominant mechanical environment in
these 10 steps determined the tissue type. Table 3.5 illustrates how the field variables were
smoothed through the analysis.

92
Table 3.5. Applying the smoothing process to the algorithm.
Step number Field variable
1 step1step1
gran.
S FV
FVFV
2
step2step2
gran.step1
S FV
10
FV9FVFV
3
step3step3
gran.step2step1
S FV
10
FV8FVFVFV
11
step11step11
step10.step2step1
S FV
10
FV...FVFVFV
n
step(n)step(n)
1).-step(n9)-step(n10)-step(n
S FV
10
FV...FVFVFV
3.3.1.3 Diffusion of progenitor cells
It has been shown that the migration of progenitor cells has a significant effect on the
regeneration process and should be considered in the fracture healing simulation (Lacroix et al.
2002). After bone fracture, progenitor cells can originate from the surrounding tissues such as the
bone marrow, the inner layer of periosteum and surrounding muscle tissues (Frost 1989,
McKibbin 1978, Simmons 1985, Henrotin 2011).
The cells were able to move through callus over time, independent of the local differentiated
tissue types, from different origins. To incorporate cell migration, a diffusion process coupled to
the poroelastic stress analysis could be included for calculating both mechanical stimuli and cell
concentration over time. ABAQUS v6.10 does not support elements having both poroelastic and
mass diffusion properties. However, an element type that has temperature-pore pressure
properties is supported by ABAQUS v6.10. The diffusion of mass and heat conduction obey

93
similar equations with different variables (density and temperature). Due to the mathematical
similarities, a heat transfer analysis with the temperature (T) as variable can be used to simulate
the cell diffusion process with cell concentration, [cells], as a density variable. The temperature
was allowed to diffuse (instead of mass) within the callus during the analysis. The main
difference between the heat conduction and mass diffusion is that heat conduction propagates by
particle contact, whereas mass diffusion propagates by mass movement through the medium,
however, the fundamental governing equations are similar.
Mass diffusion is expressed using the Fi k‟s se ond law equation, whi h an e written for ell
diffusion as follows (Crank 1956, Cussler 2009):
a2
2
2
2
2
2
Rz
cells
y
cells
x
cellsD
t
cells
. 3.30
where cells is the current cell concentration,
t
cells
shows the change in cell concentration
over time, D time
length 2
is the diffusion coefficient and aR is the source of mass which is zero in
our case (Cussler 2009, Crank 1956).
The heat flux, t
T
, should be proportional to the temperature gradient. If there is no thermal
heat generation, the heat equation an e expressed as Fourier‟s law:
k
g
z
T
y
T
x
Tα
t
T2
2
2
2
2
2
. 3.31
where T is the temperature, is a positive constant (i.e. thermal diffusivity) and g is the energy
generation per unit volume (Wylie and Barrett 1995, Cussler 2009). Since there is no source of

94
energy (mass, in the case of mass diffussion), g is omitted from the equation. Knowing that
pρC
k, the equation can be rewritten as:
2
2
2
2
2
2
p z
T
y
T
x
T
ρC
k
t
T. 3.32
where k is the thermal conductivity coefficient, ρ is the density of the material, and pC is the
specific heat coefficient (Wylie and Barrett 1995, Cussler 2009).
The conduction heat transfer in equation 3.31 can be applied directly to mass transfer problems
equation 3.30 to represent a fluid diffusing through a solid. The movement of cells through a
solid can thus be modelled by heat conduction (Cussler 2009). If temperature (T) is replaced with
the cell concentration ([cells]), and the thermal diffusivity ( ) is replaced by the diffusion
coefficient (D) then the heat conduction equation is identical to the mass diffusion equation.
Equations 3.30 and 3.31 have the same mathematical form and under the same initial conditions
and boundary conditions, both will have identical results. Figure 3.19 presents the profiles for
concentration and temperature with the same boundary conditions. It can be seen that the profiles
of concentration and temperature are identical.
Therefore, a conduction heat transfer analysis was carried in ABAQUS to simulate cell
recruitment. According to Table 3.6, by setting the density and specific heat coefficient equal to
one in the heat transfer analysis in ABAQUS, the conductivity coefficient becomes equivalent to
the mass diffusion coefficient D. Hence, the conduction heat transfer analysis was used to find
the mass concentration during the analysis. The conductivity coefficient (diffusion coefficient)
should be defined such that the progenitor cells would diffuse through the entire callus after the
estimated healing period (Gardner et al. 2006, Geris et al. 2004, Lacroix et al. 2002).

95
Figure 3.19. The profiles of (a) temperature and (b) fluid velocity are shown under the
same boundary condition show the same pattern (Cussler 2009).
Table 3.6. Similarity between mass diffusion and heat transfer equations.
Mass
diffusion
2
2
2
2
2
2
z
T
y
T
x
Tα
t
T Setting Dα ,
DρC
k
p
Heat transfer
2
2
2
2
2
2
z
cells
y
cells
x
cellsD
t
cells
To evaluate and validate the approach, the fluid velocity, octahedral shear strain and the
temperature (cell concentration) were predicted from the coupled diffusion-linear poroelastic
stress analysis for an axisymmetric model of a human tibia. There was no tissue differentiation
considered at this stage. The geometry, boundary conditions and mechanical load were chosen
based on a previous study and are shown in Figure 3.20a (Lacroix et al. 2002). The element type,
poroelastic and thermal material properties used in the simulation are illustrated in Table 3.7.
Fixed temperature (concentration) was imposed for each of the three origins (Figure 3.20b). The
mechanical stimuli as well as the temperature were calculated over time. The cell proliferation
was illustrated by plotting the temperature change, over time for three sample elements. The
regions closer to the temperature source (cells) had its maximum value and then diffused through
the callus over time. As an example, two plots having the bone marrow as cell origin, illustrating
the cell proliferation vs. time and cell concentration vs. distance are shown in Figure 3.21. (a)

96
Cell concentration within the callus at two time points: 0.5s and 1 s, (b) cell concentration over
time in three sample elements, (c) the cell concentration at two time points through the callus.
Figure 3.20. (a) Axisymmetric model of a human tibia, the radius of the cortical and bone
marrow are 15 and 9 mm (at the left). The cortical, bone marrow and callus are shown in
red, grey and green, respectively, (b) three origins for progenitor cells are shown. Arrows
indicate the cell origins (at the right).
Table 3.7. Element type, poroelastic and thermal properties of the tissues of a human.
tissue Element type
Poroelastic material properties Thermal properties
E [MPa] tyypermeabilik
Ns
mm4
e tyconductivik pC ρ
Cortical CAX4P 20000 0.3 0.00001 0.01 - - -
Marrow CAX4P 2 0.17 0.01 4 - - -
Callus CAX4PT 1 0.17 0.01 4 2.37 1 1

97
Figure 3.21. (a) Cell concentration within the callus at two time points: 0.5s and 1 s, (b) cell
concentration over time in three sample elements, (c) the cell concentration at two time
points through the callus.
The developed user-defined subroutine USDFLD was modified to consider the effect of
temperature (cell concentration). The material properties were updated based on the average of
computed mechanical stimuli in the previous 10 steps (days) and on the temporal and spatial
temperature (cell concentration). The utility routine GETVRM was used to call the material point
data of temperature (TEMP). The field variable as a function of temperature (cell concentration)
was obtained as follows:
10
FV...FVFVFV
1).-step(n9)-step(n10)-step(n
n ,mechanical
. 3.33

98
granmaxn ,mechanicalmax1n total, FVTEMP
TEMP1FV
TEMP
TEMPFV
. 3.34
In equation 3.33, FV is the field variable, which is a function of mechanical stimulus S explained
in Table 3.4. To account for the results from the mass diffusion analysis as well as the
poroelastic analysis, the rule of mixtures was used to update the material properties of the
elements over time (equation 3.34).
mechanicalFV is the field variable obtained from the poroelastic analysis at step n, granFV is the
field variable representing the material properties of the granulation tissue, maxTEMP is the
maximum temperature (cell concentration) obtained from the diffusion analysis, and totalFV is
the predicted field variable for the following step (n+1). Hence, the updated field variable in the
next iteration is relative to the cell concentration as well as the mechanical stimuli. A schematic
of the healing process is summarised in Figure 3.22.

99
Beginning of the differentiation(Day 1)
FEA ( ABAQUS)LoadingMaterial Properties
Biomechanical stimuli
n+1 Step (Day)
Smoothing process,New material properties(coupled poroelastic/mass diffusion analysis)
(Prendergast et al., 1997)
Figure 3.22. Schematic of the implemented algorithm to predict tissue distribution. The
coupled diffusive-poroelastic analysis for the mechanical stimulus and cell concentration
were obtained for each element (ABAQUS).
3.4 Verification of the implemented model
The FE method implemented in this study was initially verified by comparing the results to an
axisymmetric ovine tibia fracture model of Isaksson et al. (2006) with the same geometry and
loading regimes. The predicted patterns of the regenerated tissue during the repair process were
found to be very similar.
An axisymmetric FEM of an ovine tibia with a 3 mm fracture gap and external callus was
created (Figure 3.23b). An axial compression ramp load of 300 N was applied to the cortical
bone at the top of the model with the frequency of 1 Hz. The bone marrow, cortical and callus
were modelled with dimensions shown in Table 3.8. The nodes on the centre line were
constrained radially, whereas the bottom nodes were fixed vertically. Fluid was allowed to move
through and between different tissues. Based on experimental observations, the external callus is

100
impermeable. Therefore, there was no fluid exudation from external boundaries (Isaksson et al.
2006).
As explained in the previous section, the biphasic theory regulated by shear strain and fluid flow
was implemented into our simulation to investigate the tissue differentiation over healing time
(Prendergast et al. 1997). It was assumed that the callus consisted of stem cell seeded granulation
tissue initially, and cells could differentiate into fibrous tissue, immature and mature cartilage,
immature and mature bone. The tissue material properties can be found in Table 3.3. Progenitor
cells from three origins were able to move into the callus (Figure 3.20b). Cell migration and
proliferation were modelled as a diffusion process, coupled to the tissue-deformation stress
analysis. The diffusion coefficients were set such that after 6 weeks, the progenitor cells could
spread throughout the entire callus (Lacroix et al. 2002). A coupled diffusion-poroelastic analysis
was modelled. The model was meshed and the number of elements in the callus, bone marrow
and orti al one were ompared with Isaksson‟s model (Table 3.8).
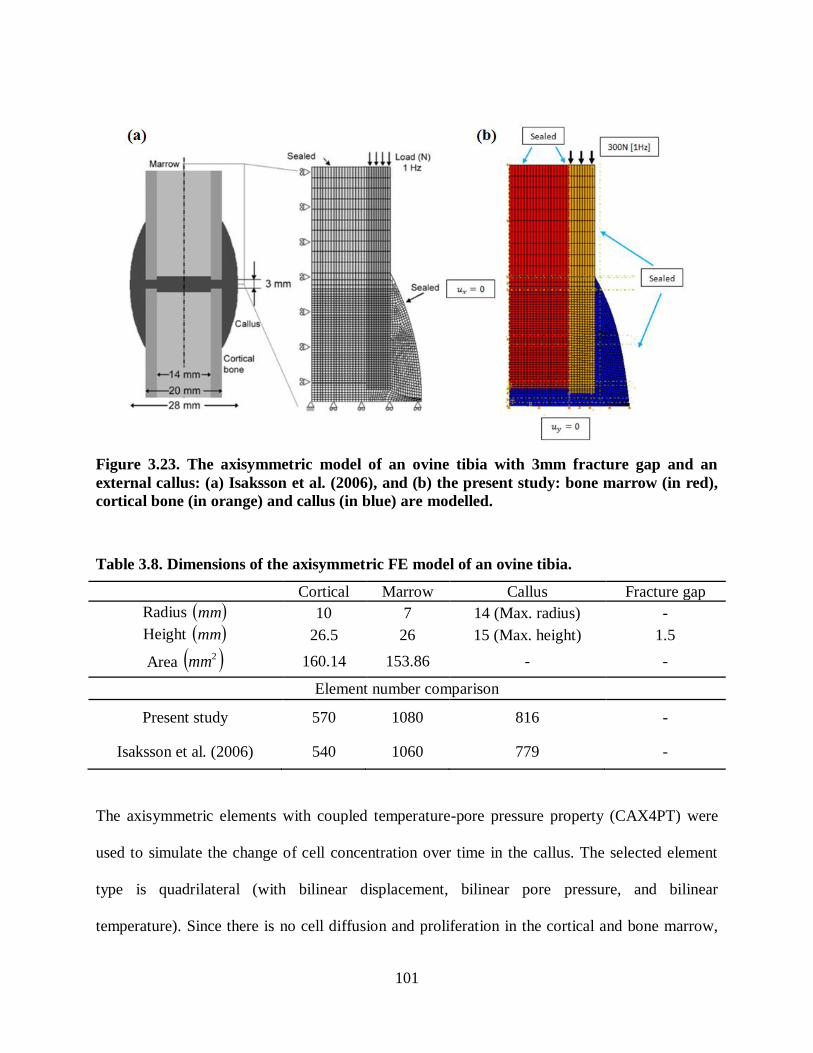
101
Figure 3.23. The axisymmetric model of an ovine tibia with 3mm fracture gap and an
external callus: (a) Isaksson et al. (2006), and (b) the present study: bone marrow (in red),
cortical bone (in orange) and callus (in blue) are modelled.
Table 3.8. Dimensions of the axisymmetric FE model of an ovine tibia.
Cortical Marrow Callus Fracture gap
Radius mm 10 7 14 (Max. radius) -
Height mm 26.5 26 15 (Max. height) 1.5
Area 2mm 160.14 153.86 - -
Element number comparison
Present study 570 1080 816 -
Isaksson et al. (2006) 540 1060 779 -
The axisymmetric elements with coupled temperature-pore pressure property (CAX4PT) were
used to simulate the change of cell concentration over time in the callus. The selected element
type is quadrilateral (with bilinear displacement, bilinear pore pressure, and bilinear
temperature). Since there is no cell diffusion and proliferation in the cortical and bone marrow,

102
simple axisymmetric pore pressure elements (CAX4P) were used for tissues other than callus.
The material properties were updated in the callus based on the cell concentration and
mechanical stimuli at peak loads using the user defined subroutine USDFLD. The change of
mechanical stimuli in the computational analyses is faster than in vivo, and FE analysis could not
exactly mimic what happens during the bone repair. To determine the mechanical stimuli, the
load history was calculated over a time period. According to the study by Isaksson et al. (2006),
the applied load in each step represented the average load that had been applied to the bone
during one day of healing. However, this assumption needs more discussion due to the time
dependent and viscoelastic behaviour of tissues. Even a marginal change in the load frequency
may have considerable effect on the fluid behaviour. According to the literature, each step in our
analysis represented one day of healing (Isaksson et al. 2006). The analysis was continued until
tissue differentiation had reached a steady-state. In other words, there was no change in the
mechanical stimuli and cell concentration.
3.4.1 Results
The predicted sequence of tissue regeneration in the simulated fracture model had the same
pattern as observed in Isaksson et al. (2006). Figure 3.24 shows the healing pattern in the present
study (steps 6-50). Bone formation initiated in the internal and external callus, independently.
Intramembranous ossification started from the callus tip and along the periosteal surface,
followed by cartilaginous tissue differentiation within the rest of the external callus and
medullary. Cartilaginous tissue then differentiated into bony tissue and then gradually spread
throughout the callus (endochondral ossification). According to the results, even after 30 steps
(days of healing), a considerable amount of fibrous tissue still existed in the interfragmentary
gap. Comparing the change of fluid velocity in different regions, it can be observed that the

103
maximum fluid velocity is 0.23 [ m/s] at the tip of external callus, however it is around 12 times
larger under the cortical shaft (2.84 [ m/s]) during healing (Figure 3.25). Therefore, high
magnitudes of fluid velocity may be the result of delayed tissue differentiation. Since axial
compressive load had been applied to the cortical shaft, the zones in the interfragmentary gap
(under the cortical shaft) experienced higher load magnitudes and thus fluid velocity had
increased.
The predicted tissue distributions were compared with those obtained by Isaksson et al. (2006)
(Figure 3.26-28). As presented in Figure 3.26, fibrous tissue was surrounded by cartilage under
the cortical shaft. In addition, intramembranous ossification occurred in the external callus at the
callus tip and along the periosteum at the beginning of the analysis in both models. After 50
steps, a marginable amount of fibrous tissue, surrounded by mature cartilaginous tissue, exists at
the fracture gap. However, the rest of callus was filled with intermediate and/or mature bony
tissue. Mature bone was observed in most parts of the external callus. Intermediate bone exists at
the bottom right side of the external callus and at left corner of internal callus (Figure 3.26 and
Figure 3.27).

104
Figure 3.24. Prediction of fracture healing in the present study during 50 steps (days).
Cortical bone was subjected to a 300 [N] axial compression load (1 Hz).

105
Figure 3.25. The change of fluid flow [ m/s] over time under the cortical shaft and callus
tip during fracture healing. Cortical bone was subjected to a 300 N axial compression
loading (1 Hz).
Figure 3.26. Comparison of the two simulations during fracture healing in the first steps.
Cortical bone was subjected to a 300 N axial compression loading (1 Hz): (a) the present
study, and (b) Isaksson et al. (2006). In both models, intramembranous ossification
occurred at the callus tip and periosteum. Also, fibrous tissue and cartilaginous tissue can
be found in the same zones.

106
Figure 3.27. Comparison of the two simulations during fracture healing at step 50, cortical
bone was under 300 N axial compression loading (1 Hz): (a) the present study, (b) Isaksson
et al. (2006). The amount of fibrous tissue decreased significantly under the cortical shaft.
The distribution of mature and intermediate bone distribution is almost identical.

107
Step 9
Step 13
Step 23
Step 33
Step 50
(a) (b)
(a)
(b)
Figure 3.28. Overall similar healing patterns were observed over time under a 300 [N] axial
compression load in (a) the present study, and (b) Isaksson et al. (2006).
3.4.2 Discussion
The temporal and spatial tissue differentiation in Isaksson et al. (2006) and the present study
were quite similar, but not exactly the same (Figure 3.28). The minor differences may be due to
the different method of implementing the mass diffusion analysis. In the previous study done by
Isaksson et al. (2006), MATLAB was linked to ABAQUS in order to update the element material

108
properties (Lacroix et al. 2002, Sandino and Lacroix 2011) whereas in this study a user defined
subroutine was written in FORTRAN. In Isaksson et al. (2006), the values of Young‟s modulus,
oisson‟s ratio and permea ility were averaged separately in MATLAB to predict the tissue
type. However, in this study, all of the material properties were obtained through the field
variable and by determining the FV in each step, the material properties were updated
automatically, slightly differently, through FV.
Isaksson et al. (2006) used a diffusion process uncoupled to the tissue poroelastic analysis.
Moreover, the eight-node element with 9 integration points was used (CAX8P). However, in this
study a fully coupled diffusion-poroelastic analysis was performed. Due to the available coupled
temperature-pore pressure elements in ABAQUS v6.10, a four-node axisymmetric quadrilateral
elements with 4 integration points (CAX4PT) was used for the callus, and a four-node
axisymmetric element with 4 integration points was used for bone marrow and cortical
(CAX4P).
Despite these differences in model implementation, the predicted tissue differentiation for both
models was very similar. Hence, the current model can be considered verified against Isaksson et
al. (2006) and we were confident in implementing the method into our FE models of murine tibia
and investigating the effect of mechanical loading on tissue differentiation and bone repair.
3.5 Fracture healing case studies
The mechanoregulatory algorithm was implemented into an idealised models of a murine tibia.
First, an axisymmetric model was simulated to investigate the effect of load magnitude on
fracture healing. Second, a three dimensional (3D) model was created to determine the effect of
torsional load as well as combined loading on the temporal and spatial differentiation patterns.

109
3.5.1 An axisymmetric idealized murine model
An axisymmetric FE model of a proximal section of a murine tibia with a 0.4 mm fracture gap
and external callus were created based on the described method in the previous sections (Figure
3.29a). The mid-diaphyseal dimensions of the modelled murine tibia are shown in Table 3.9
(Windahl et al. 1999, Geris et al. 2004). To investigate the influence of load magnitude on
fracture repair, axial compression loads of 0.5, 1, 2 N (1 Hz) were applied to the top of the
cortical shaft (Figure 3.29b). Verification of the results was done by comparing to CT images
of a previously published experimental study (Gardner et al. 2006).
Figure 3.29. (a) Axisymmetric FE model of a murine tibia (bone marrow in red, cortical
bone in grey and callus in blue), (b) the amplitude of the applied cyclic axial compression
loads.
Tissues were assumed to follow the biphasic algorithm proposed by Prendergast et al. (1997) and
the material properties were set as described in Table 3.10. The Young‟s modulus, oisson‟s
ratio, permeability and the void ratio were defined for each tissue. Cortical bone has the highest

110
stiffness. However, it has the lowest permeability due to a low void ratio of 0.0416. Immature,
intermediate and mature woven bones were modelled to represent the tissue differentiation over
time. The Young‟s modulus of immature, intermediate and mature woven one are 500<E<1000
MPa, 1000<E<2000 MPa, and from 2000<E<6000 MPa, respectively. Mature and immature
cartilage were also modelled with the Young‟s modulus of 10<E<500 and 5<E<10, respe tively.
Finally, the Young‟s modulus of fi rous tissue ranged between 1 and 5 MPa. The material
properties used in this study can be found in Table 3.10.
Table 3.9. Geometry of the murine tibia proximal section (Windahl et al. 1999, Geris et al.
2004).
Cortical periosteal circumference mm 4.51
Cortical endosteal circumference mm 3.12
Maximum external callus radius mm 1.2
Fracture gap mm 0.4
The boundary conditions were set such that the nodes located on the symmetry axis are
constrained in the x-direction and the bottom of the model in the y-direction. The external
boundaries were assumed to be impermeable. All tissues were modelled using pore pressure
elements. The bone marrow and cortical bone contained 444 and 240 simple axisymmetric pore
pressure (CAX4P) elements, and the callus was meshed using 480 axisymmetric temperature-
pore pressure elements (CAX4PT).
It was assumed that a fixed number of progenitor cells originated from the bone marrow,
periosteum and surrounding muscle tissues and were able to move within the callus. The
diffusion coefficient set such that after 3 weeks, the progenitor cells would have spread
throughout the callus (Gardner et al. 2006).

111
The mechanical stimuli (fluid velocity and tissue octahedral shear strain) and the cell
concentration were computed at the callus elements (at maximum loading) by implementing a
coupled diffusive-poroelastic analysis. The geometric nonlinearity (NLGEOM) option in
ABAQUS was switched on in order to account for large deformations in the tissues.
Table 3.10. Material properties used for the study (Rho, Ashman and Turner 1993,
Isaksson et al. 2006).
Constant Variable
Cortical Marrow Granulation
tissue
Fibrous
tissue Cartilage
Immature
bone
Mature
bone ovine tibia
E [GPa] 6 0.002 0.0002 310- 3105
3105 -
0.5 0.5-1 2-6
0.325 0.167 0.167 0.167 0.167 0.325 0.325
k
Ns
mm 4
510 0.010 0.010 0.010 0.005 0.100 0.370
e 0.041 4.00 4.00 4.00 4.00 4.00 4.00
According to the obtained mechanical stimuli and cell concentration, the material properties of
the cells within the callus elements were updated using a rule of mixtures (equation 3.34). The
callus cells within an element differentiated into fibrous tissue, mature and immature cartilage
and mature, intermediate and immature woven bone. There was no tissue differentiation within
the marrow and cortical bone. Therefore, the material properties of marrow and cortical bone
were kept constant over time. The analysis continued until steady-state was reached.
3.5.1.1 Results
Bone formation was successfully simulated over time with the sequential prediction of fibrous,
cartilage and mature bony tissue. The predicted sequence of tissue regeneration in the murine

112
fracture model occurred in the same pattern observed in vivo (Figure 3.30). In all cases, the bone
formation started from the internal and external callus, independently. For the external callus, the
bone formation started at the tip of the callus and gradually spread throughout the callus. First,
cartilaginous tissue formed and then differentiated into bony tissue. The interfragmentary
movement was computed for all cases. Under the 0.5 N loading case the interfragmentary
movement was negligible after 10 steps. However, for 1 N and 2 N the movement decreased to
0.04 mm and 0.07 mm. The fluid velocity and strain were reduced over time, which illustrates
that the callus became stiffer over time. For the 0.5 N loading case, the mechanical stimuli had
less magnitude and reached a steady state faster than higher load magnitudes (Figure 3.31). After
14 steps (days), the interfragmentary gap still contained fibrous tissue for the load amplitudes of
1 or 2 N whereas cartilage was predicted for the 0.5 N loading case (Figure 3.30). According to
the obtained results, the bone healing rate was the highest under 0.5 N, intermediate under 1 N
and lowest under 2 N axial compression, which corresponded to in vivo observations.
Figure 3.30. Predicted tissue differentiation in the present study under (a) 0.5 N, (b) 1 N
and (c) 2 N axial compression load (1 Hz).

113
0
10
20
30
40
50
0 5 10 15 20
Oct
ahed
ral s
hea
r st
rain
[%
]
Step (day)
0.5 N 1 N 2 N
0
1
2
3
4
5
6
0 5 10 15 20
Flu
id v
elo
ciy
[µm
/s]
Step (day)
0.5 N 1 N 2 N
Figure 3.31. Predicted mechanical stimuli for a sample element under the cortical shaft
during the healing process for three axial compression loading magnitudes.
3.5.1.2 Discussion
The bone under higher mechanical conditions had higher interfragmentary movement and higher
fluid velocity. The higher magnitude of movement led to instability and a delay in bone healing.
The computational results were compared with a previous experimental study (Gardner et al.
2006). The murine bones had a similar fracture size and were under identical mechanical
conditions in both cases. In agreement with the micro-CT images from Gardner et al. (2010), the
computational simulations predicted bone repair enhancement under low magnitudes of load
(Figure 3.32c). As seen in Figure 3.30 the bone under 0.5 N had stiffer bridging. Octahedral
shear strain magnitudes were higher in the 2 N case, which led to less consolidation and fibrous
tissue was still observed after 2 weeks (Figure 3.32b). At higher loads the callus was poorly
bridged as seen in the CT images (Figure 3.32c).

114
Figure 3.32. a) Axisymmetric FE model of a murine tibia. The inner and outer diameter of
cortical bone (gray) and external callus (blue) are 1, 1.5 and 2.4 mm, respectively, b)
Predicted tissue differentiation in the present study, c) CT images from Gardner et al.
(2006) for different load magnitudes (1 Hz).
3.5.2 A 3D idealised model of murine tibia
Since axial torsion could not been applied to the 2D model, a poroelastic three-dimensional (3D)
finite element model of a murine ti ia with a 0.4 mm gap and external allus was reated
(ABAQUS v6.10). In order to be able to apply torsional load, a 3D model was created (Figure
3.35a). To decrease the computational cost, a 22.5 degree wedge was modelled. The boundary
conditions were chosen so that the bottom surface was fixed in axial and angular directions and
the vertical axis at the left was restricted radially. The bone marrow and cortical bone contained
759 and 432 simple three dimensional 8-node trilinear displacement and pore pressure (C3D8P)
elements, and the callus was meshed using 949 three dimensional 8-node trilinear displacement,
pore pressure, and temperature elements (C3D8PT). The external surface of the callus and
external surfaces of the bone were assumed to be impermeable. The NLGEOM option in
ABAQUS was switched on to consider geometric nonlinearity in the tissues. The diffusion

115
coefficient was set such that, after 3 weeks, the mesenchymal stem cells would spread
throughout the callus (Gardner et al. 2006). It was assumed that initially the callus consisted of
stem cell seeded granulation tissue. Murine embryonic stem cells were transplanted due to their
regenerative capacity. The elements within the callus were able to differentiate into fibrous
tissue, immature and mature cartilage, and immature, intermediate and mature bony tissue.
Progenitor cells migrated into the callus from surrounding tissues, marrow, periosteum and
external soft tissues. To investigate the influence of mechanical environments, axial torsion (8
degrees, 1 Hz) alone, and then combined, axial compression (0.4 N, 1 Hz) and torsion (8 degrees,
1 Hz) were applied to the cortical shaft with a maximum octahedral shear strain magnitude of 25
%, based on a previous in vivo study (Bishop et al. 2006). In the experimental study, cyclic axial
torsion was applied over a fractured sheep tibia with a maximum principal strain of 25 %
(Bishop et al. 2006).
3.5.2.1 Results
The predicted sequence of tissue differentiation in the 3D murine fracture model occurred as
expected in both cases. First, the granulation tissue differentiated into cartilaginous tissue which
was initiated from the callus tip at a faster pace. The bony tissue, formed from cartilage
(endochondral ossification), was first observed in the external callus and then spread throughout
the callus gradually. As expected, fibrous tissue existed in the medullary cavity even after 15
days of healing. The interfragmentary movement decreased with healing in both cases.
For the axial torsion case, the magnitude of octahedral shear strain and fluid velocity were
plotted over time for three sample elements located in the external callus, interfragmentary
region and internal callus (Figure 3.34). As seen in Figure 3.34, the element under the cortical
shaft has the highest mechanical stimuli. The strain reaches 6 % under the cortical bone, whereas
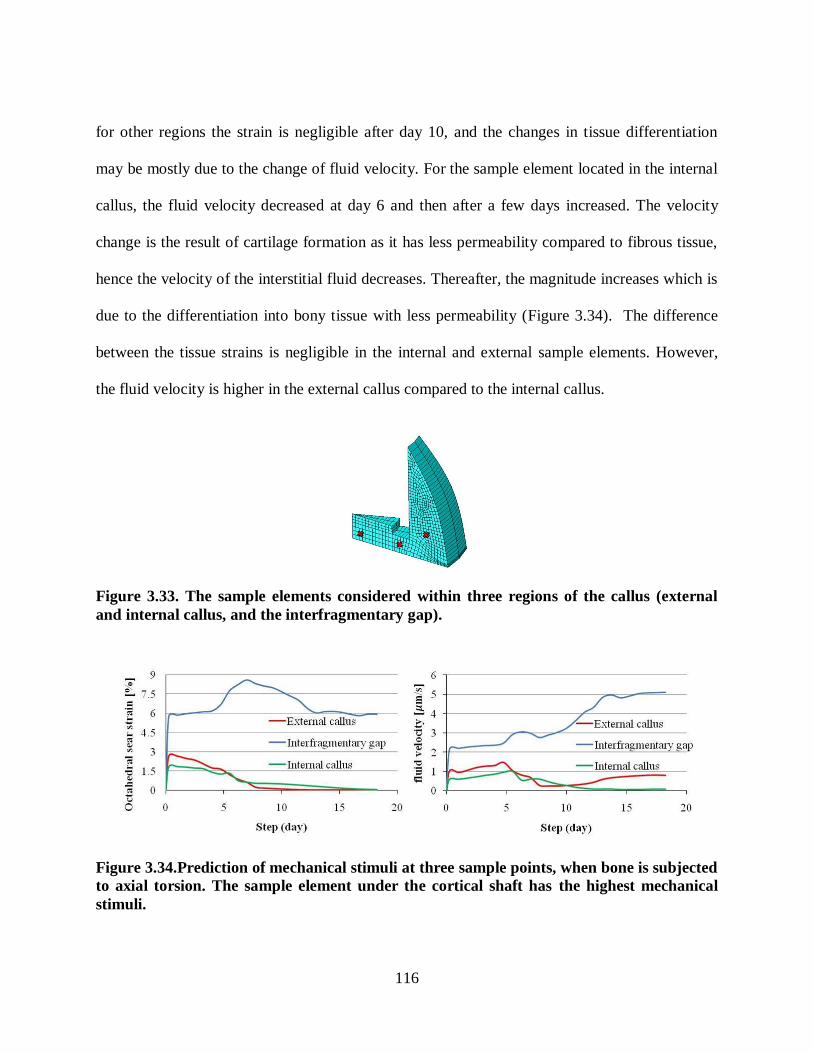
116
for other regions the strain is negligible after day 10, and the changes in tissue differentiation
may be mostly due to the change of fluid velocity. For the sample element located in the internal
callus, the fluid velocity decreased at day 6 and then after a few days increased. The velocity
change is the result of cartilage formation as it has less permeability compared to fibrous tissue,
hence the velocity of the interstitial fluid decreases. Thereafter, the magnitude increases which is
due to the differentiation into bony tissue with less permeability (Figure 3.34). The difference
between the tissue strains is negligible in the internal and external sample elements. However,
the fluid velocity is higher in the external callus compared to the internal callus.
Figure 3.33. The sample elements considered within three regions of the callus (external
and internal callus, and the interfragmentary gap).
Figure 3.34.Prediction of mechanical stimuli at three sample points, when bone is subjected
to axial torsion. The sample element under the cortical shaft has the highest mechanical
stimuli.

117
The mechanical stimuli had higher magnitudes when the bone was under combined loading
compared to the torsion alone case. Under pure axial torsion, bony tissue was also formed in the
internal callus. In addition, there were greater bony areas and density. However, under combined
loading, only cartilaginous tissue was observed in the internal region and no bone had
regenerated. The algorithm predicted stiffer bridging and promoted healing in pure torsion
(Figure 3.35b). Whereas, poorly bridged callus and promoted healing were predicted with added
axial compression (Figure 3.35c).
Figure 3.35. (a) A 3D FEM of a murine tibia with 0.4 mm gap, predicted tissue
differentiation in the model under: (b) Axial torsion (8 degree, 1 Hz), (c) Axial torsion &
compression (8 degrees, 0.45 MPa, 1 Hz).
3.5.2.2 Discussion
Fibrous tissue was still present under the cortical shaft after 2 weeks of healing, which may be
related to the higher magnitudes of fluid velocity at the interfragmentary gap compared to other
regions. The algorithm predicted stiffer bridging and promoted better healing in torsion

118
compared to the combined loading. Under combined loading the bone was probably overloaded
and had more movement, thus did not have as a stable condition as pure axial torsion. Under
axial torsion, by formation of bony tissue in the external callus, the interfragmentary strain
decreased. This may explain why the bone under axial torsion had stiffer bridging and bony
tissue had differentiated in the internal callus. On the other hand, due to higher mechanical
stimuli, no bony tissue was observed in the internal callus for the combined loading regime.
The predictions are in agreement with the experimental study of (Bishop et al. 2006) that applied
axial torsion with the maximum strain of 25 % to the tibial osteotomy in sheep. As seen in Figure
3.36 bony tissue exists in the internal and external callus for both the empirical and
computational studies (orange arrows). Furthermore, the computational model predicted
cartilaginous tissue under the cortical shaft (green arrow), which also exists in the experimental
study (Bishop et al. 2006).
Figure 3.36. (a) Sheep tibial section subjected to torsion (histological slide) (Bishop et al.
2006), (b) predicted tissue differentiation subjected to torsion (day 15, in the present study).

119
3.6 Summary of the computational analyses
The verified algorithm was implemented into two idealised murine models and the effects of
mechanical loading on the healing process were investigated:
1. A poroelastic two dimensional (2D) FE model of a murine tibia with a 0.4 mm gap and
external callus was created (ABAQUS v6.11). To explore the influence of load
magnitude on fracture repair, axial compression loads of 0.5, 1, 2 N (1 Hz) were applied
to the top of the cortical shaft, and the results were compared to reconstructed µCT
images of a previously published experimental study (Gardner et al. 2006).
2. A poroelastic idealised three-dimensional (3D) FE model of a murine tibia with a 0.4 mm
gap and external callus was created (ABAQUS v6.11). To investigate the influence of
mechanical environments, axial compression (0.4 N, 1 Hz) and torsional rotation (8
degrees, 1 Hz), were applied to the cortical shaft with a maximum octahedral shear strain
magnitude of 25 %. The predictions agreed with the experimental study of Bishop et al.
(2006) that applied axial torsion with the maximum strain of 25 % to the tibial osteotomy
in sheep.
The first specific objective of the study was to investigate the mechanical environments within a
stem cell collagenous scaffold (Chapter 4):
1. An axisymmetric FE model of the modified Flexcell system was created to apply
confined compression test, using ABAQUS v6.11.
2. The load-deflection FE results for the rate of 1 N/s were validated against the preliminary
experimental results from our group (Olesja Hazenbiller, University of Calgary, M.Sc.
student).

120
3. The biphasic algorithm was implemented into the computational model to predict tissue
differentiation under confined compression (5 and 20 kPa, 1 Hz). In agreement with our
experimental results, gel became stiffer by differentiating into cartilaginous tissues
(Olesja Hazenbiller, University of Calgary, M.Sc. student).
The second specific objective was to simulate tissue differentiation within a µCT-based FE
model of a murine tibia (Chapter 5):
1. The µCT based FE model of a burr-hole murine tibia was reconstructed using
Simpleware software. The non-homogeneous grey-scale based mechanical properties
were used.
2. The mechanical behaviour of the reconstructed model was verified against the published
experimental study of Stadelmann et al. (2009). Octahedral shear strain was estimated
numerically in three different zones of the murine tibia and compared with the available
empirical results. The computational and empirical results were in good agreement
qualitatively.
3. The biphasic algorithm was implemented into the computational model to predict the
development of differentiated tissues in a closed fracture model treated with a stem cell
seeded soft collagenous scaffold: (1) under different loading conditions (e.g. axial
compression and bending), (2) for different locations of the scaffold, and (3) different
rates of diffusion.
The following figure summarizes the computational simulations conducted in this thesis (3.37).

121
Figure 3.37. Summary of the computational simulations in the current study.

122
Chapter Four: Collagenous scaffold under confined compression
Biomechanical stimuli have been shown to modulate stem cell differentiation and fracture
healing progression. The effect of mechanical loading on a collagenous scaffold under confined
compression was investigated in this chapter. The first section describes the computational
simulation developed to characterize the mechanical behaviour of a confined compression
experiment developed to load very soft gels. In the second section, the computational model was
used to investigate the experimental studies performed by our group on gene expression analysis
and tissue differentiation within stem cell seeded collagenous gels. Finally, the biphasic
mechanoregulatory algorithm that is regulated by shear strain and interstitial fluid velocity
(Lacroix et al. 2002, Prendergast et al. 1997) was implemented into the FE model to determine
the tissue differentiation in confined compression (cyclic loading).
4.1 Confined compression loading device description
Biomechanical stimuli can affect fracture healing and remodelling progression, however, a
complete understanding of the role of mechanical factors and cell-matrix interactions is needed.
To investigate the influence of mechanical stimuli on cellular responses and cell differentiation
the FX-4000 ™ Flex ell ® ompression plus ™ system was modified to apply one-dimensional
(1D) loads to a collagenous scaffold used in tissue engineering (Figure 4.1).
The Flexcell system is a computer-controlled instrument designed to apply mechanical loads to
cell cultures and tissue explants (Flexcell International, Hillsborough, NC, USA). An axial
compression load is applied to the cells or gels using a range of air pressure (0-44 kPa) to deform
a flexi le sili one mem rane lo ated at the ottom of iopress ™ ulture plates (Flex ell
International). The system is restrained at the top by a rigid lid. During loading the cells are

123
restrained to the central region of each plate by foam rings, however, the rings can expand in the
radial direction (Figure 4.2).
Figure 4.1. FX-4000 ™ Fl xc ll ® comp ssion pl s ™ sys m.
The system can apply static, cyclic and controlled load regimes with various frequency ranges
from 0.1 to 5 Hz to the cultured cells or tissues. It also allows the selection of different
waveforms such as static, sinusoidal, heart, triangular, square and custom waves.
In our study, the Flexcell system was modified to compress very soft cell seeded collagenous
gels. The Flexcell system mimics an unconfined compression test condition (Figure 4.2).
However, in order to apply 1D loads to very soft gels, modifications to the device were made to
enable a confined compression test on the gels. The BioPress® compression culture plates and
the loading cell base were modified (Olesja Hazenbiller, M.Sc. student, University of Calgary).
The loading cell base was remodelled and replaced. In addition to the cell base, other parts were
added to the system: a porous permeable plug, and silicone rings with square and round cross
sections.

124
The modified system contains seven parts: (1) the flexible membrane, (2) modified cell base, (3)
collagen gel, (4) square ring, (5) porous plug, (6) round ring, and (7) the top lid. The parts added
to the system are presented in Figure 4.3.
Figure 4.2. Schematic of the Flexcell system cross-section in the uncompressed and
compressed configurations.
A 316L stainless steel (SS) porous plug with 10 µm pore size is used to permit nutrient and
oxygen exchange as well as provide a rigid surface to transfer loads (MOTT Corporation,
Farmington, CT, USA). It has been shown that the 316L SS plug has appropriate
biocompatibility for short term and long term cell cultures (Jacobs and Oloff 1985, Puleo et al.
1991). The plug allows for fluid exudation from the top surface, and also enables the exchange of
oxygen needed for cell culture in an incubator. Moreover, the plug has similar physical
properties to (mature) trabecular bone with approximately similar porosity properties (Wen
2009).

125
Figure 4.3. Parts of the modified system: (1) cell base, (2) square ring, (3) porous plug, (4)
round ring, (5) lid, and (6) fixed lid.
Silicone rings possess good biocompatibility and thus are used in implants and prostheses,
gaskets, seals and O-rings (Schulmerich et al. 2006). The silicone rings used in our experiments
have a good resistance to chemical and biological degradation. The silicone ring with a circular
cross section was used to bridge the gap between the plug and the top lid, and maintain contact
between the porous plug and the fixed lid. There were three main reasons for adding the square
ring to the system:
(1) The collagen gel had very low stiffness and could not maintain its shape; therefore, the
silicone square ring acted as a support for the soft gel in a 1D configuration; (2) it was used to
ensure the loading conditions were predominantly 1D, and (3) during the loading and unloading
period, it was used to maintain the gel within the loading device. The schematic of the system is
presented in Figure 4.4 and 4.5.

126
Figure 4.4. The modified system to conduct confined compression test.
4.2 Computational modelling of the system
A finite element model was developed (ABAQUS v.6.11) to determine the stress-strain response
of the collagen gels as a function of applied pressure in the modified Flexcell system. A non-
linear axisymmetric analysis was performed to simulate the modified system and estimate the
strains transmitted to the cells. Six parts were considered in the model: (1) the cell base, (2) the
collagen gel, (3) the square ring (4) the porous plug (5) the round ring, and (6) the lid. The
flexible membrane was not considered in our simulation because of its negligible stiffness with
respect to the other parts.

127
Figure 4.5. The parts added to the modified Flexcell system.
4.2.1 The cell base and the top lid
The cell base and the top lid were modelled as analytical rigid shells due to their high stiffness
compared to the other parts. Analytical rigid surfaces are geometric surfaces that can be
described by straight and curved line segments, and their motion is defined through a single node
with six degrees of freedom (i.e., a rigid body reference node). The reference points were
positioned at the centre of mass for both parts. An axial compression force was applied to the
reference node of the cell base at the bottom while a fixed boundary condition was implemented
at the top lid. Assuming the base and the lid to be rigid bodies improved computational time
without affecting the overall accuracy of the analysis.
4.2.2 The collagen gel and the porous plug
The material properties of the collagenous soft gel were considered to be similar to granulation
tissue with a Young‟s modulus of 0.2 M a and a oisson‟s ratio of 0.167 and a permea ility of

128
0.01 Ns
mm 4
. The Young‟s modulus of the gel was also determined (approximately 0.15 MPa)
from the stress-strain curve obtained experimentally (Olesja Hazenbiller, M.Sc. student,
University of Calgary). The collagen gel was modelled as a biphasic material (80 % fluid and 20
% solid). The void ratio (e) was set to 4 and meshed using continuum eight-node axisymmetric
pore pressure elements (CAX8P).
A 3.3175 mm thick 316L stainless steel porous plug with a 12.7-mm-diameter was created. It
was also modelled as a biphasic material. The permeability of a 316L stainless steel plug with a
0.5 µm pore size was reported to be close to the permeability of the cortical bone (510
Ns
mm 4
)
(Wen 2009). Since the porous plug that we used had much higher pore size (10 µm), the
permeability of the plug was set close to the permeability of mature trabecular bone with the
value of 0.37 Ns
mm 4
. The void ratio was calculated from the geometry and the density. Knowing
the mass and density of the plug
3cm
gr7.99ρ1.031gr,m the volume would be:
3
solid cm0.254ρ
mV . 4.1
The volume could also be determined from the radius and the height of the plug. For a height
(h=0.125 in) and a radius (r=0.25 in) the volume would be:
32
total cm0.402hπrV . 4.2
The difference between these volumes established the void volume ( 3
f 0.148cmV ). Therefore,
the void ratio of the porous plug was computed and used in the computational simulation:

129
0.582V
Ve
s
f
. 4.3
The void ratio of the plug (e = 0.582) was 7 times less than the trabecular bone (e = 4.0). The
values for Young‟s modulus, oisson‟s ratio, permea ility and void ratio of the ollagen gel and
the porous plug used in the model can be found in Table 4.1.
Table 4.1. Poroelastic properties of the porous plug and the collagen gel (Isaksson et al.
2006, Tromas et al. 2012).
E MPa k
Ns
mm 4
e
Porous plug 193000 0.30 0.37 0.582
Collagenous gel 0.2 0.167 0.01 4.0
4.2.3 The silicone rings
The silicone rings were modelled as isotropic hyperelastic materials and meshed using eight-
node axisymmetric quadrilateral elements with hybrid formulation (CAX8H). The hybrid
elements were used to consider the incompressibility of the silicone rings. The geometry of the
rings is provided in Table 4.2.
Hyperelastic materials are described in terms of the strain energy stored in the material per unit
volume (i.e. strain energy potential). The derivative of the strain energy function with respect to
a strain component determines the corresponding stress component. The stress-strain data
obtained from a uniaxial compression test (Olesja Hazenbiller, M.Sc. student, University of
Calgary) was used as material input into ABAQUS v6.11
The strain energy potential (W) is defined as follows in the Marlow form, which is suited for
quasi-incompressible materials. Also, for the cases in which only one set of data is available

130
(e.g., only uniaxial), the Marlow potential fits experiments better than more complex potentials,
such as the Moonly-Rivlin or Ogden types (Mazurkiewicz 2009). The Marlow potential take the
form:
JWIWW el1dis
. 4.4
where disW is the distortional component obtained from uniaxial or biaxial test data, elW is the
volumetri omponent defined from volumetri data, oisson‟s ratio or the lateral strains
obtained from the test data, and J is the volume ratio. 1
I is the first distortional strain defined
as:
i3
1
i
32
22
12
1
λJλ
λλλI
, 4.5
where J is the volume ratio and i are principal stretch ratios. For a strictly incompressible
material 3I is equal to one and the strain energy potential is a function of
1I (Darwish 2004).
Table 4.2. The geometry of the silicone rings.
Thickness [mm] Inner diameter [mm] Outer diameter [mm]
Round ring 2 6 10
Square ring 1 10.7 12.7
Uniaxial stress-strain testing was conducted to define the material properties of the rings (Olesja
Hazenbiller, M.Sc. student, University of Calgary). The force-deflection behaviour of the rings
obtained from an experiment is shown in Figure 4.6. Since we had one set of uniaxial test data,
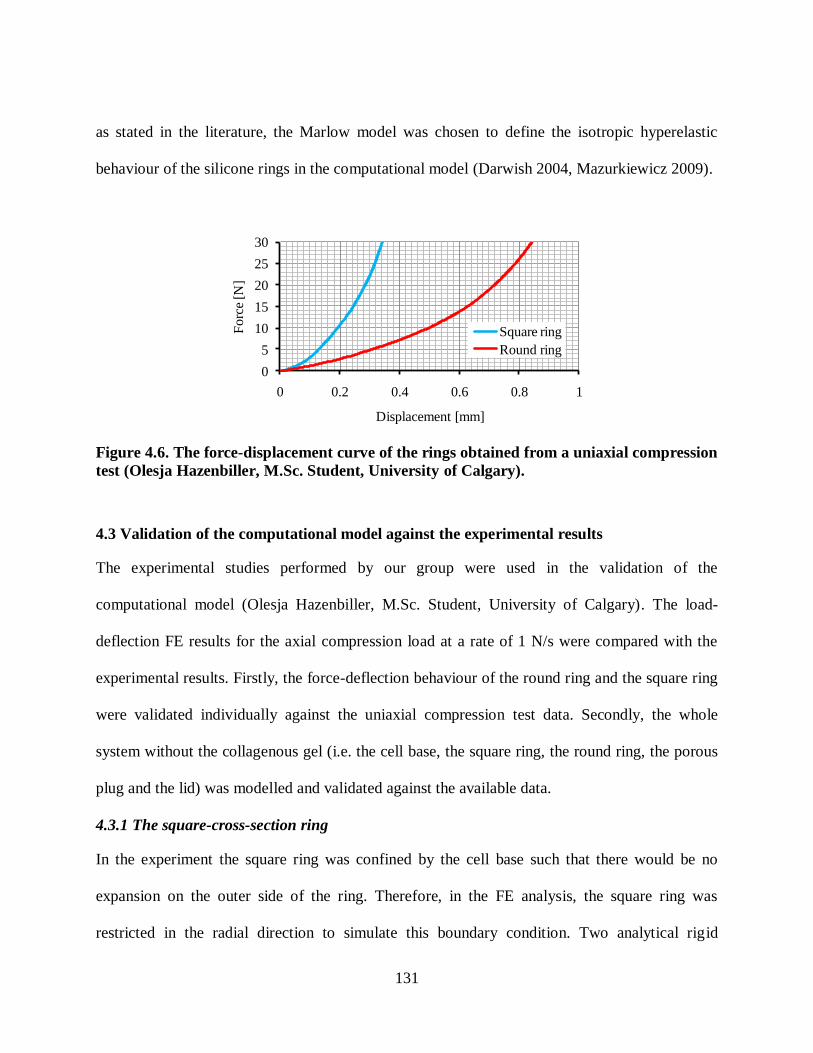
131
as stated in the literature, the Marlow model was chosen to define the isotropic hyperelastic
behaviour of the silicone rings in the computational model (Darwish 2004, Mazurkiewicz 2009).
0
5
10
15
20
25
30
0 0.2 0.4 0.6 0.8 1
Fo
rce
[N]
Displacement [mm]
Square ring
Round ring
Figure 4.6. The force-displacement curve of the rings obtained from a uniaxial compression
test (Olesja Hazenbiller, M.Sc. Student, University of Calgary).
4.3 Validation of the computational model against the experimental results
The experimental studies performed by our group were used in the validation of the
computational model (Olesja Hazenbiller, M.Sc. Student, University of Calgary). The load-
deflection FE results for the axial compression load at a rate of 1 N/s were compared with the
experimental results. Firstly, the force-deflection behaviour of the round ring and the square ring
were validated individually against the uniaxial compression test data. Secondly, the whole
system without the collagenous gel (i.e. the cell base, the square ring, the round ring, the porous
plug and the lid) was modelled and validated against the available data.
4.3.1 The square-cross-section ring
In the experiment the square ring was confined by the cell base such that there would be no
expansion on the outer side of the ring. Therefore, in the FE analysis, the square ring was
restricted in the radial direction to simulate this boundary condition. Two analytical rigid

132
surfaces were created at the top and the bottom of the square ring (Figure 4.7). The rigid body at
the bottom was completely fixed. An axial compression load at a rate of 1 N/s was applied to the
top rigid body for 30 seconds. Two frictionless surface-to-surface contacts were defined
between: (1) the upper surface of the square ring (slave) and the top rigid body (master), and (2)
the lower surface of the square ring (slave) and the rigid body at the bottom (master).
Interactions between the master surfaces and the slave surface were established to allow for
direct frictionless surface-to-surface contact. Rigid bodies should always be defined as the
master surfaces whereas the body with less stiffness should be the slave surface. Figure 4.77
illustrates the axial displacement profile of the square ring under axial compression after 30 s.
The force-displacement curve was obtained and compared to the experimental (Figure 4.100).
Top rigid surface
Bottom rigid surface (fixed)
0xu
1 [N/s]
t= 30 s
Symmetry line
Figure 4.7. The axial displacement [mm] of the axisymmetric model of the square ring
under 30 N axial compression load at t=30 s. The inner and outer diameters are 10.7 mm
and 12.7 mm, respectively, and the height is 1 mm.
4.3.2 The circular-cross-section ring
An axisymmetric FE model of the ring with a circular cross sectional area was created. The
material was defined as an isotropic hyperelastic material using the Marlow form. The material

133
properties obtained from the uniaxial compression test were imported into ABAQUS to define
the hyperelastic properties of the material. The circular-cross-section ring, meshed using CAX8H
elements, was sandwiched between two analytical rigid bodies and, as in the experiments, it was
allowed to expand radially. Frictionless surface-to-surface contact was defined between the ring
and the rigid bodies: (1) the upper portion of the round ring (slave) and the top rigid body
(master), and (2) the lower portion of the round ring and the bottom rigid body (master). The
force was applied at a rate of 1 N/s to the ring for 30 seconds while the bottom rigid body was
completely fixed. The prediction of axial displacement after 30 seconds is presented in Figure
4.8. The force-displacement graph over 30 seconds of the ramp load was compared to the
experimental data (Figure 4.10).
Top rigid surface
Bottom rigid surface (fixed)
1 [N/s]
t= 30 s Symmetric line
Free to expand radially.
Figure 4.8. The axial displacement [mm] of the axisymmetric model of the round ring
under 30 N axial compression load at t=30 s. The inner and outer diameters are 6 mm and
10 mm, respectively, and the radius of the cross section is 2 mm.

134
4.3.3 The system excluding the collagenous gel
An axisymmetric model of the whole system excluding the soft collagenous gel was created: (1)
the cell base, (2) the square ring, (3) the porous plug, (4) the square ring, and (5) the top lid. The
base and the top lid were defined as analytical rigid bodies. The porous plug was represented as
an elasti material with the Young‟s modulus of 193000 [M a] and oisson‟s ratio of 0.3. The
plug was meshed using 8-node axisymmetric continuum elements (CAX8). The rings were
considered as isotropic hyperelastic materials using the Marlow formulation. The silicone rings
were meshed using 8-node axisymmetric continuum elements with hybrid formulation
(CAX8H).
Frictionless surface-to-surface contacts were defined between surfaces, while one surface was
the master surface and the other was the slave surface. Four groups of master and slave surfaces
were defined as follows:
1. The top lid (master) and the upper surface of the round ring (slave).
2. The top surface of the porous plug (master) and the lower surface of the round ring
(slave).
3. The bottom surface of the porous plug (master) and the top surface of the square ring
(slave).
4. The cell base (master) and bottom surface of the square ring (slave).
The boundary conditions were defined as follows: (1) the reference point of the top lid was fixed,
(2) the outer edge of the porous plug and square ring were confined in the radial direction, and
(3) the round ring was free to expand radially.

135
The axial compression force at a rate of 1 N/s was applied to the reference point of the cell base
in 30 seconds. The prediction of the axial displacement within the system, excluding the gel, is
illustrated in Figure 4.9. Under an axial compression load of 30 N, the deflections for the round
ring and the square ring were 0.874 mm and 0.325 mm, respectively. The round ring carried
most of the reaction force and had greater deflection compared to the square ring under the same
loading condition. The load-deflection results of the system at a load rate of 1 N/s were
compared and validated against experimental results. It can be observed that the empirical and
FE results are in good agreement (Figure 4.10).
Top lid (rigid surface, fixed)
Cell base (rigid surface) 1 [N/s]
t= 30 s
Symmetry line
Free to expand radially.
0xu
0xu
Figure 4.9. The axial displacement [mm] of the axisymmetric FE model of the system
without gel under 30 N axial compression load at t=30 s. The diameter of the porous plug is
12.7 mm and the height is 3.175 mm.

136
0
5
10
15
20
25
30
0 0.2 0.4 0.6 0.8 1 1.2
Forc
e [N
]
Displacement [mm]
FE-Square
ring
Exp-Round
ring
Exp-Square
ring
FE-Round
ringFE-Whole
model
Exp-Whole
model
Figure 4.10. Comparison of load-displacement curves between the FE and experimental
results under axial compression load applied at a rate of 1 N/s.
4.3.4 The system including the collagenous gel
The collagenous gel and the porous plug were modelled as poroelastic materials, and were
meshed using the 8-node, axisymmetric continuum elements with pore pressure degree of
freedom (CAX8P). The pore pressure was set to zero at the top edge of the porous plug to allow
fluid exudation in the vertical direction. The material properties for gel and the plug can be found
in Table 4.1. The other parts were modelled as explained in the previous sections: (1) the top lid
and cell base modelled as analytical rigid bodies, (2) the silicone rings were modelled as
hyperelastic materials using CAX8H elements with hybrid formulation. The axial compression
load at a rate of 1 N/s was applied to the system. The axisymmetric FE model is presented in
Figure 4.11.

137
Figure 4.11. The axisymmetric FE model of the modified Flexcell system.
Contact formulation: The surface-to-surface contact relationships with small sliding
formulation were established between all surfaces. The surface-to-surface formulation was
selected for our situation because the normal directions of the contacting surfaces were opposite
to each other. Small sliding was used to allow slight sliding of surfaces along each other and
assure that the nodes were always in contact with the same local surface. To overcome contact
convergence problems, an adjustment zone depth of 0.2 mm was defined for the upper and lower
side of the round ring in contact with the top lid and the porous plug. This adjustment was
established to eliminate small gaps caused by numerical errors without creating any strain in the
model. The adjustment zone extends from the master surface. Any nodes on the slave surface
within the adjusting zone were moved precisely onto the master surface. The adjustment zone
was defined at the beginning of the analysis. The motion of these nodes onto the master surface

138
does not create any strain in the model. Figure 4.12 shows how the nodes moved into contact if
the adjustment zone is used or if „a‟ was set equal to zero.
The established surface-to-surface contacts are as follows:
1. The top lid (master) and the upper side of the round ring (slave).
2. The top surface of the porous plug (master) and the lower side of the round ring (slave).
3. The bottom surface of the porous plug (master) and the top surface of the square ring
(slave).
4. The bottom surface of the porous plug (master) and the top surface of the collagen gel
(slave).
5. The inner edge of the square ring (master) and the outer edge of the collagen gel (slave).
6. The cell base (master) and the bottom surface of the collagen gel (slave).
7. The cell base (master) and the bottom surface of the square ring (slave).
Figure 4.12. (a) The contact surface at the beginning of the analysis, (b) if no
adjustment zone was used, (c) the j s m n ‘a’ w s fin n h no s wi hin h
zone were moved onto the master surface (ABAQUS v6.11 user manual).

139
The meshes were matched between the contact surfaces at the beginning of the computational
analysis to obtain a better convergence rate and thus more accuracy (Figure 4.11).
A pore pressure degree of freedom existed on both sides of the contact interfaces, as the collagen
gel and the porous plug were defined as poroelastic materials. The contact surface between the
upper side of the collagen gel and the bottom surface of the porous plug should allow for the free
movement of fluid and a biphasi “jump ondition” should need to be satisfied (Hou et al. 1989):
0nqnvv sff , 4.6
where q is the fluid flow, f is the fluid phase volumetric fraction, sv and fv are the solid and
fluid phase fluid velocities, respectively, and n is the outward normal to the contact interface.
The fluid flowed in the direction normal to the contact interface and did not flow tangentially
along the surface (ABAQUS v6.11 neglected the tangential flow). The flow normal to the
interface was defined as follows (Federico et al. 2004, Pawaskar, Fisher and Jin 2010)
(ABAQUS v6.11 user manual):
BAcacross ppkq . 4.7
where acrossq was the fluid flow normal to the interface, Ap and Bp were pore pressures at points
on opposite sides of the interface (master and slave); and ck was the contact permeability with
units of Ns
mm3
.
A positive value of pressure gradient corresponded to fluid flow out from the master surface or
flow into the slave surface, while a negative pressure gradient represented that no flow could
exude from the master surface and the fluid from the slave surface flows into the master surface.

140
The contact permeability was set equal 1 Ns
mm3
and thus the fluid could flow freely between the
contact interfaces (Pawaskar et al. 2010, Federico et al. 2004).
The axial compression load was applied to the cell base at a constant rate of 1 N/s for 20
seconds. The collagen gel was compressed between the base and the permeable porous plug, and
was confined by the sides of the square ring and the base. Therefore, the deformation was
predominantly in the axial direction. The compressive stress in the gel increased due to
pressurization of the fluid phase. The solid matrix was compressed as the interstitial fluid was
forced out of the scaffold. The square ring, due to its greater stiffness, controlled the overall
strain of the collagen. The total displacement of the cell base was the sum of the deformations of
the square ring and round ring (Figure 4.13).
0
4
8
12
16
20
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8
Forc
e [N
]
Displacement [mm]
Whole system including the gel
Base Round Ring Square Ring or Collagen
Figure 4.13. Model predictions of the force-displacement curve for the whole system
including the collagen gel under confined compression: ramp load with the rate of 1 N/s.
The distribution of axial strain demonstrated that the elements at the superficial layers of the
collagenous scaffold experienced higher peak strains and fluid velocities (Figure 4.14). Four
sample elements were selected in the axial direction to examine the effect of confined

141
compression under the applied ramp load. During the initial ramp phase the elements closer to
the plug experienced higher magnitudes of compression. For instance, the element at the top
under a 20 N compression load experienced 40 % compressive strain and a fluid velocity of 6
µm/s, whereas the mechanical stimuli for the middle or bottom elements were 3 % compressive
strain and a fluid velocity of 4105 µm/s (Figure 4.15, Figure 4.16).
Axial strain, collagenous scaffold
Symmetry line
t=20 s
Symmetry line
Fluid velocity [mm/s], collagenous scaffoldt=20 s
Figure 4.14. Distribution of axial strain (EE2) and fluid velocity (FLVEL, [mm/s]), within
the collagenous scaffold under 20 N axial compression load at t=20 s.

142
Figure 4.15. Change of fluid velocity over time within four sample elements of the
collagenous scaffold. The scaffold was loaded at a rate of 1 N/s. Four sample elements are
shown through the depth of the collagen (at the right).
Figure 4.16. Change of axial strain over time within four sample elements of the
collagenous scaffold The scaffold was loaded at a rate of 1 N/s. Four sample elements are
shown through the depth of the collagen (at the right).
The displacement of the rings and the cell base were compared in two models: (1) the
collagenous gel not included in the simulation (Figure 4.9); (2) the collagenous gel included in
the model (Figure 4.17). The total displacement was greater for the model without the gel
(Figure 4.17). The solid phase transferred a significant portion of the load to the interstitial fluid
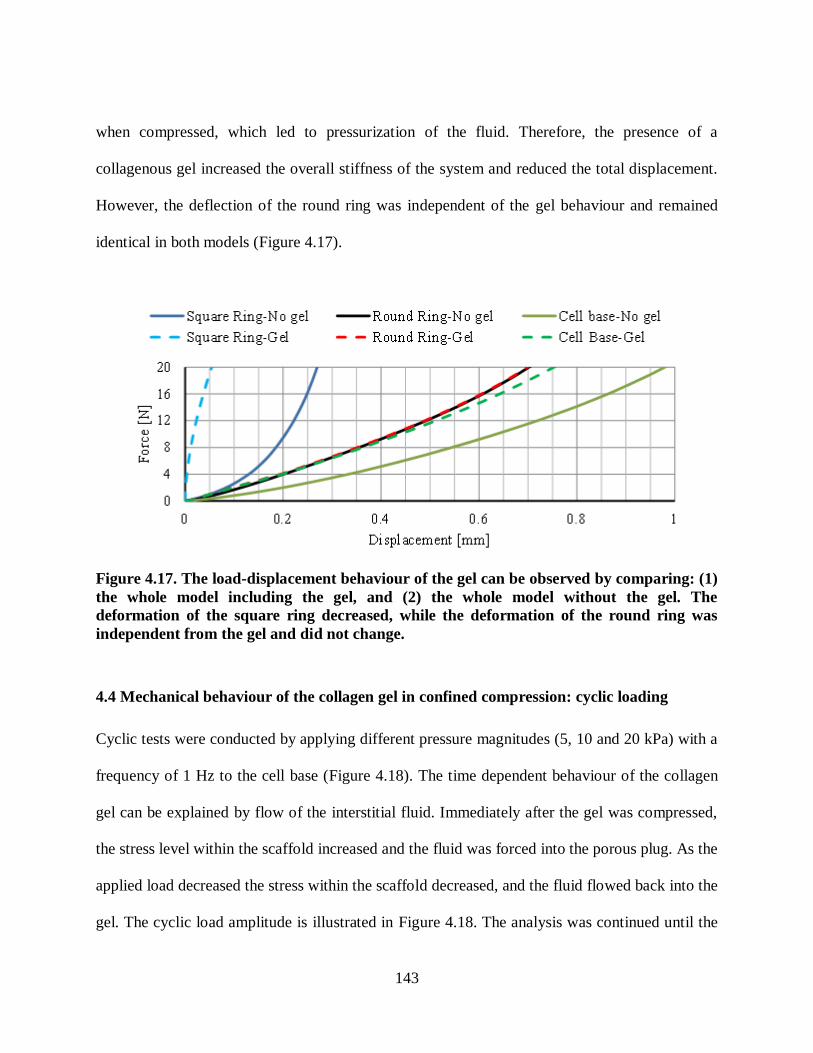
143
when compressed, which led to pressurization of the fluid. Therefore, the presence of a
collagenous gel increased the overall stiffness of the system and reduced the total displacement.
However, the deflection of the round ring was independent of the gel behaviour and remained
identical in both models (Figure 4.17).
Figure 4.17. The load-displacement behaviour of the gel can be observed by comparing: (1)
the whole model including the gel, and (2) the whole model without the gel. The
deformation of the square ring decreased, while the deformation of the round ring was
independent from the gel and did not change.
4.4 Mechanical behaviour of the collagen gel in confined compression: cyclic loading
Cyclic tests were conducted by applying different pressure magnitudes (5, 10 and 20 kPa) with a
frequency of 1 Hz to the cell base (Figure 4.18). The time dependent behaviour of the collagen
gel can be explained by flow of the interstitial fluid. Immediately after the gel was compressed,
the stress level within the scaffold increased and the fluid was forced into the porous plug. As the
applied load decreased the stress within the scaffold decreased, and the fluid flowed back into the
gel. The cyclic load amplitude is illustrated in Figure 4.18. The analysis was continued until the

144
tissue subsided to a steady state to explore the influence of mechanical loading on temporal and
spatial cellular responses in the Flexcell experiments with time in culture. The stress at the
equilibrium point represented the stress within the solid phase.
Figure 4.18. The amplitude of the applied cyclic axial compression loads. The pressure,
ranging from 5-20 [kPa], was applied to the cell base with a frequency of 1 Hz.
A 5 kPa (1 Hz) was applied at the bottom until the pore pressure subsided to a steady state (300
s) to mimic experimental investigations. Three sample elements were selected through the axial
depth of the gel: (1) top element at the top surface of the gel, (2) the middle element, and (3) the
bottom element at the bottom of the gel. The strain curve of the top sample element over the first
40 seconds is presented in Figure 4.19. The superficial layers experienced higher peak strains
and pore pressure compared to the deeper layers (Figure 4.20, Figure 4.21 and Figure 4.23). The
scaffold was strained non-uniformly due to a relatively quick compression. The superficial layers
experienced 3.3 % axial compressive strain, while an internal tension was generated at the deeper
layers. Although initially the magnitudes of strain, fluid velocity and the pore pressure were
higher at the superficial layers of the scaffold, after approximately 300 s the system reached an
equilibrium state. The nonhomogeneous distribution of the peak strain relaxed to an

145
approximately constant value of 1.6 % axial compression and 1.2 % octahedral shear strain
(Figure 4.20, Figure 4.21). The fluid velocity increased to 1.3 [µm/s] and at the equilibrium stage
had subsided to 0.2 µm/s (Figure 4.22). The fluid pore pressure at the equilibrium state went to
zero. Therefore, the pore pressure at peak loading represented the stress of 0.02 [MPa] in the
solid phase at the equilibrium state (Figure 4.23).
Figure 4.19. Model prediction for the axial strain during confined compression: cyclic
loading (P=5 kPa, 1 Hz). Compressive strain in top element under axial compression.
Figure 4.20. Prediction of axial strain at the peak loading in three selected sample elements
in the collagenous scaffold in confined compression under cyclic loading of P=5 kPa (1 Hz).
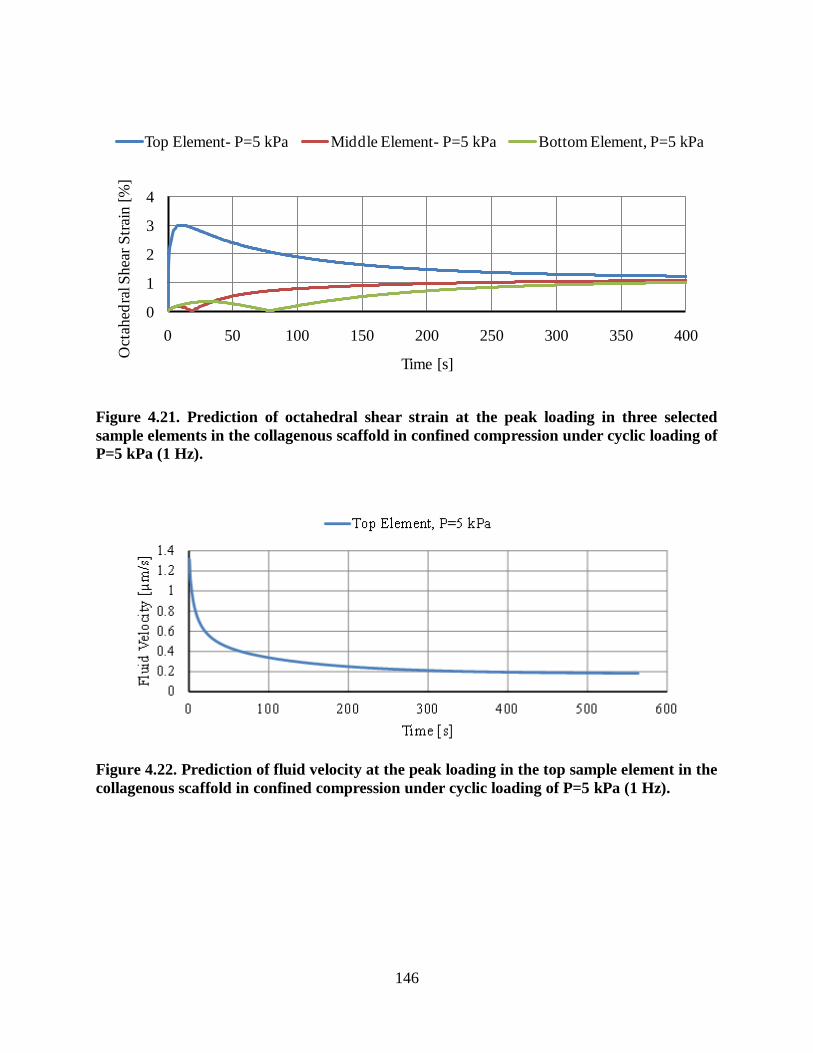
146
0
1
2
3
4
0 50 100 150 200 250 300 350 400
Oct
ahed
ral S
hea
r S
trai
n [
%]
Time [s]
Top Element- P=5 kPa Middle Element- P=5 kPa Bottom Element, P=5 kPa
Figure 4.21. Prediction of octahedral shear strain at the peak loading in three selected
sample elements in the collagenous scaffold in confined compression under cyclic loading of
P=5 kPa (1 Hz).
Figure 4.22. Prediction of fluid velocity at the peak loading in the top sample element in the
collagenous scaffold in confined compression under cyclic loading of P=5 kPa (1 Hz).

147
0
0.02
0.04
0.06
0.08
0 50 100 150 200 250 300 350 400 450pore
pre
ssure
[M
Pa]
Time [s]
Top Element, P=10 kPa Middle Element, P=10 kPa Bottom Element, P=10 kPa
Figure 4.23. Prediction of pore pressure at the peak loading in three selected sample
elements in the collagenous scaffold in confined compression under cyclic loading of P=5
kPa (1 Hz).
Three different pressure levels with the same loading rate (5, 10 and 20 kPa, 1 Hz) were applied
to the cell base to investigate the effect of load magnitude on the mechanical stimuli, temporally
and spatially. A longer time was required to reach a steady state and the strain value increased in
the collagen matrix with increased applied pressure (Figure 4.24). The predicted equilibrium
strains were 7.7, 3.4 and 1.6 % within the solid section (top sample element), when the cell base
was under 20, 10 and 5 kPa pressure, respectively.
The stress-strain response of the collagenous scaffold is shown for the 10 kPa (1 Hz) case
(Figure 4.25). As the gel was compressed, the solid matrix deformed and the interstitial fluid was
forced out of the gel, which led to an increase in the deformation of the collagen matrix and an
increase in fluid pressure. However, by decreasing the pressure to zero, the fluid flowed back
into the gel, which led to a decrease in the strain level within the solid phase. The time dependant
nature of the gel could be observed by the stress-strain curve during the loading and unloading
(hysteresis). The width of the ellipses in Figure 4.25 illustrates the energy loss due to viscous

148
dissipation and fluid exudation from the gel. The initial loading cycles were associated with a
stiffer response and greater energy loss.
Figure 4.24. Comparison of axial strains at the peak loading in the top sample element in
the collagenous scaffold in confined compression under different cyclic applied loads of
P=5, 10, 20 kPa (1 Hz).
Figure 4.25. The gel was subjected to 10 kPa pressure (1 Hz). The hysteresis of the stress-
strain curve shows the effects of the interstitial flow and viscous dissipation (the graph
shows the first 33 seconds of analysis).

149
The mechanical behaviour of the 1D FE model of the modified Flexcell system was validated
against experimental results (Figure 4.10) (Olesja Hazenbiller, M.Sc. student, University of
Calgary). The influence of mechanical loading on temporal and spatial cellular responses was
explored. The response of the collagen gel in confined compression under cyclic loads up to 20
kPa was evaluated. The magnitudes of axial strain, fluid velocity and pore pressure were
predicted at the peak loading in the top, middle and bottom sample elements. As the applied
pressure increased, the equilibrium strain, fluid velocity and pore pressure increased (Table 4.3).
This model provides a tool to understand the internal stress-strain response within the modified
Flexcell system for mechanobiological experiments with very soft gels.
Table 4.3. The predicted equilibrium strains, fluid velocities and pore pressure at the peak
loading in the top, middle and bottom sample elements.
Pressure [kPa] Axial strain [%] Fluid velocity [µm/s] Pore pressure [MPa]
Top sample element
5 (1 Hz) 1.6 0.18 0.01
10 (1 Hz) 3.4 0.43 0.02
20 (1 Hz) 7.7 1.06 0.05
Middle sample element
5 (1 Hz) 1.4 0.001 0.01
10 (1 Hz) 3.1 0.010 0.02
20 (1 Hz) 6.3 0.040 0.05
Bottom sample element
5 (1 Hz) 1.3 0.00007 0.01
10 (1 Hz) 3.0 0.00070 0.02
20 (1 Hz) 5.8 0.00200 0.05
4.5 Prediction of tissue differentiation in confined compression
The aim of this study was to investigate the effect of mechanical loading on a stem cell seeded
collagenous scaffold under confined compression. FE modelling and mechanoregulatory
algorithms were used to explore the influence of mechanical loading on temporal and spatial
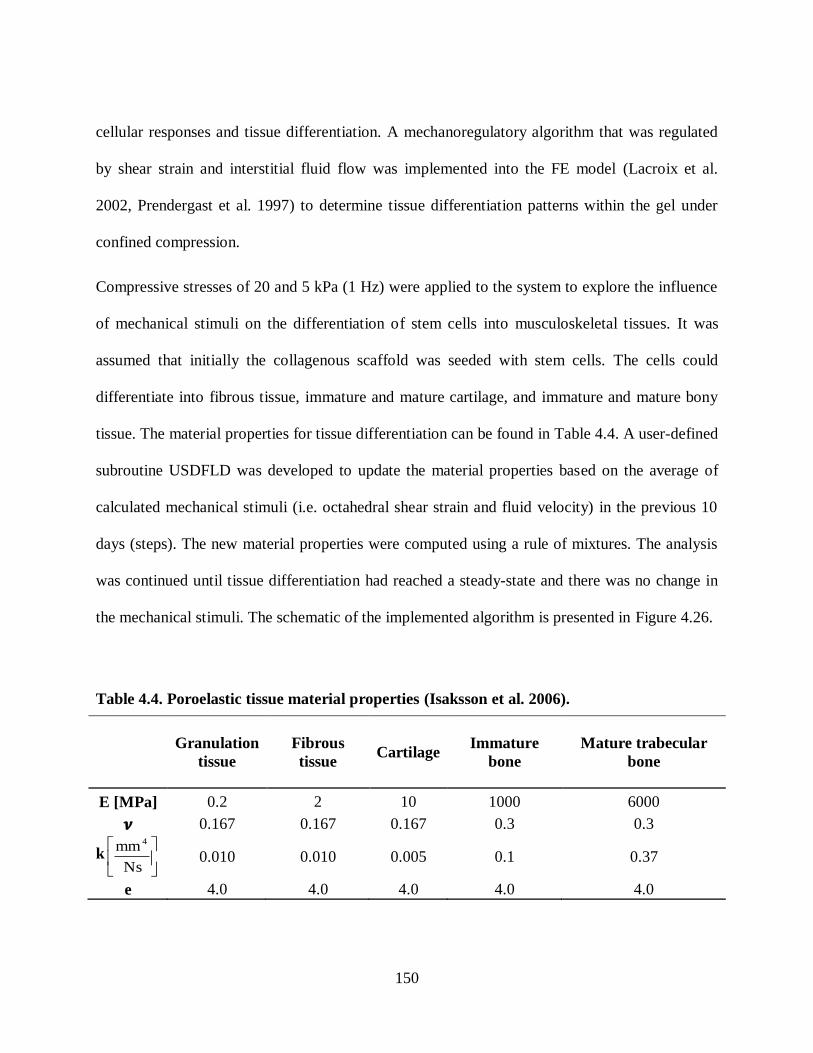
150
cellular responses and tissue differentiation. A mechanoregulatory algorithm that was regulated
by shear strain and interstitial fluid flow was implemented into the FE model (Lacroix et al.
2002, Prendergast et al. 1997) to determine tissue differentiation patterns within the gel under
confined compression.
Compressive stresses of 20 and 5 kPa (1 Hz) were applied to the system to explore the influence
of mechanical stimuli on the differentiation of stem cells into musculoskeletal tissues. It was
assumed that initially the collagenous scaffold was seeded with stem cells. The cells could
differentiate into fibrous tissue, immature and mature cartilage, and immature and mature bony
tissue. The material properties for tissue differentiation can be found in Table 4.4. A user-defined
subroutine USDFLD was developed to update the material properties based on the average of
calculated mechanical stimuli (i.e. octahedral shear strain and fluid velocity) in the previous 10
days (steps). The new material properties were computed using a rule of mixtures. The analysis
was continued until tissue differentiation had reached a steady-state and there was no change in
the mechanical stimuli. The schematic of the implemented algorithm is presented in Figure 4.26.
Table 4.4. Poroelastic tissue material properties (Isaksson et al. 2006).
Granulation
tissue
Fibrous
tissue Cartilage
Immature
bone
Mature trabecular
bone
E [MPa] 0.2 2 10 1000 6000
0.167 0.167 0.167 0.3 0.3
k
Ns
mm 4
0.010 0.010 0.005 0.1 0.37
e 4.0 4.0 4.0 4.0 4.0

151
Figure 4.26. Schematic of tissue differentiation algorithm.
Three sample elements were selected through the axial depth of the gel. Model predictions from
the differentiation algorithm for fluid flow and octahedral shear strain indicated a reduction over
time within the scaffold that gradually reached a steady state (5 and 20 kPa, 1 Hz). Since the
fluid was forced into the porous plug, the elements at the top surface of the gel experienced
higher magnitudes of fluid velocity and strain (20 and 5 kPa, 1 Hz). The fluid velocities of the
three sample elements were compared during tissue differentiation for the gel subjected to a 20
kPa (1Hz) compressive stress (Figure 4.27).
The temporal change of the mechanical stimuli showed that the magnitudes of fluid velocity and
octahedral shear strain were smaller at 5 kPa of applied pressure compared to the 10 kPa case
(Figure 4.28). Under a 5 kPa pressure (1 Hz), the magnitude of fluid velocity and octahedral
shear strain within the gel were negligible and were not of sufficient magnitude to induce tissue

152
differentiation. According to the biphasic algorithm small magnitudes of strain and fluid velocity
lead to bony tissue formation (Lacroix et al. 2002). When the scaffold was subjected to a 5 kPa
pressure the mechanical stimuli were very small, and thus the algorithm predicted cartilage and
bone tissue formation. The predicted sequence of tissue differentiation in the gel subjected to a 5
kPa pressure did not occur in the same pattern observed in vitro (not shown), which shows that
the mechanoregulatory algorithm could not properly model the actual tissue differentiation
process.
Figure 4.27. Prediction of fluid velocity at the peak loading in three sample elements in the
collagenous scaffold during tissue differentiation (P = 20 kPa, 1 Hz).
When the system was subjected to a 20 kPa compressive stress (1 Hz) tissue differentiation was
promoted in the zones closer to the loading but the superficial layers had delayed tissue
differentiation. The fibrous tissue differentiation initiated from the bottom regions, followed by
differentiation into a cartilaginous tissue, which then spread throughout the scaffold gradually.
At day 6 (step 6), small amounts of undifferentiated tissue still existed at the top surface of the

153
gel (Figure 4.29b). In the superficial regions the higher values of mechanical stimuli resulted in
increased fluid flow, which delayed the differentiation process into cartilaginous tissues for the
20 kPa load case (Figure 4.28, Figure 4.29b).
Figure 4.28. Mechanical stimuli in a sample element at the superficial layer for a 5 kPa and
a 20 kPa (1 Hz) compressive pressure applied to the system.
Gene expression analysis within the stem cell seeded collagenous scaffold from our experimental
studies indicated that the 3D collagen1 gels at day 15 had promoted chondrogenesis (Figure 4.29,
Figure 4.30) (Olesja Hazenbiller, M.Sc. student, University of Calgary). In agreement with the
experimental results the gel became stiffer by differentiating into cartilaginous tissues as
predicted by the finite element analysis (Figure 4.29a, Figure 4.29b, Figure 4.30).
154
Figure 4.29. (a) Gel before and after loading, (b) FE prediction of tissue differentiation (a
20 kPa compressive stress, 1 Hz).
Figure 4.30. Compressive load significantly influenced chondrogenesis (Col 2-day 15),
(Olesja Hazenbiller, M.Sc. student, University of Calgary).
4.6 Summary
A computational model was developed to characterize the mechanical environment within the
modified Flexcell system. Next, a biphasic mechanoregulatory algorithm was implemented into
the 1D model to help develop an understanding of the biosynthetic responses within the cell
seeded gels when subjected to mechanical perturbations (Lacroix et al. 2002). The model

155
predictions suggested that loading the stem cell-seeded soft gel led to differentiation of stem cells
into cartilaginous tissue. Hence, the mechanical stimuli provided a suitable environment for cell
biosynthesis and differentiation. Both empirical and computational data suggest that mechanical
stimulation of the scaffold may be an effective way to initiate differentiation pathways prior to
implantation for tissue engineering applications. The computational and experimental studies
(e.g. gene expression, µCT, mechanical testing and the mineralization patterns) will be used
simultaneously in future studies to further develop mechanoregulatory models with a more
robust quantitative base.

156
Chapter Five: Tissue Differentiation in a Burr-hole Fracture Model in a Murine Tibia
This chapter presents a computational analysis of tissue differentiation within a burr-hole
fracture model of a murine tibia. A mechanoregulatory algorithm is implemented in a finite
element model of the in-vivo burr-hole fracture model developed by Taiani et al. (2010). The
tissue differentiation patterns to predict bone formation within the fracture are investigated for a
number of variables. An introduction to the software used to develop the finite element mesh is
first given to understand the required stages for the FE model regeneration and volumetric
meshing. Then, the steps to generate the FE models of the intact murine tibia are presented. Next,
the mechanical behaviour of the reconstructed model was validated against a previous
experimental-numerical study (Stadelmann et al. 2009). Subsequently, the development of the
burr-hole fracture model in the murine tibia are outlined. Finally, the biphasic
mechanoregulatory algorithm was implemented into the developed model to predict tissue
differentiation in the burr-hole model (Isaksson et al. 2006, Prendergast et al. 1997). The burr-
hole model was then filled with a stem cell seeded soft collagenous scaffold and tissue
differentiation was predicted for a variety of case studies:
1. tibia was subjected to axial compression loads ( 2, 1 and 0.5 N, 1 Hz),
2. different rate of cell diffusion (0.025 and 0.01 s
mm 2
),
3. different axial positions of the fracture (4.95, 3.13 and 2.55 mm),
4. different cell origins,
5. osteoporotic bone with poor mechanical behaviour, and
6. the tibia was also subjected to bending load (0.02, 0.04 Nm).
The overview of this chapter and computational investigations conducted is shown in Figure 5.1.

157
Reconstruction of a 3D murine tibia using
µCT images.
Mechanoregulatory algorithm.
Loading
conditions. Location of
the burr-hole.
Rate of
diffusion.
Verification against a previous experimental
study (Stadelmann et al., 2009).
Osteoporotic
bone.
Figure 5.1. Overview of this chapter.
5.1 Introduction
Micro computed tomography (µCT) is known as an imaging technique for medical purposes in a
minimally invasive way. Recently, this technique has been used to create precise FE models
from complex geometries (Silva, Brodt and Hucker 2005, Stadelmann et al. 2009). A three-
dimensional finite element model (3D FEM) of a murine tibia structure was recovered from
(µCT) scans and were meshed, using Simpleware v3.5.3 image processing software (Simpleware
Ltd., Exeter, UK). This software consists of three modules:
(1) ScanIP is the core image-processing platform. Medical file formats, such as CT, µCT,
and magnetic resonance imaging (MRI) data can be imported into the software for
processing and manipulation.
(2) ScanFE converts the segmented 3D images into volumetric or surface models. Mesh
generation is performed within this module for a variety of commercial FE platforms.

158
(3) ScanCADcan be used to modify models with Boolean operations on masks of the multi-
part models to add parts or create layers (e.g. union, subtraction, invertion and
intersection).
5.2 Reconstruction of a murine tibia
In this study, the FE reconstruction of an intact murine tibia was performed in three sequential
stages explained in the following sections.
5.2.1 Importing and preparing the data (ScanIP module)
The right tibia from a female CD1 mouse (8 weeks old, the skin and muscle were cleaned) was
scanned at a 7 µm resolution using a SCANCO Medical µCT 35 at fMcCaig Institute (University
of Calgary). Full-length scans were obtained consisting of 2883 slices. Tagged image file format
(TIFF) data were created with 0.007 mm interval and imported into the ScanIP module. The
histogram, i.e. a graph showing the number of pixels at different intensity values, was computed
to find the frequency of the greyscale values for the various tissues. The histogram provided
information about the threshold values and noise reduction of the images. The memory required
for image analysis was estimated at 1900 Mb. To reduce the memory requirement, the empty
space around the object was cropped. The data were rescaled by assigning the pixel spacing of
0.03 mm in all directions using linear interpolation. Moreover, the resolution of the datasets was
down-sampled (i.e. increase the pixel spacing) to make the segmentation step faster and easier.
These steps reduced memory usage to a more manageable 97 Mb.
5.2.2 Image processing (ScanIP module)
When the data was imported into the ScanIP module, some noise (unnecessary points around the
object) was detected. The noise could result from the heterogeneous x-ray emission or the
inappropriate position of sample holder with respect to the x-ray source (e.g. the sample holder

159
did not fit in the diameter of the camera field of view, Figure 5.2). A noise-reducing filter was
used to reduce noise while preserving features as much as possible. The noise reduction
decreased the number of unnecessary voxels and made the segmentation process easier
(identifying tissue type).
sample holder
detecto
r
x-r
ay s
ourc
e
sample holder
(a) (b)
Figure 5.2. Top view of sample holder is located fitted within the diameter of the field of
view (orange lines), (b) schematic of the sample holder used for scanning the murine tibia
(µCT 35, User Manual).
Segmentation is the basic step of FE model generation. In this step, the volumes of interest are
identified and classified into appropriate groups of tissues (e.g. tibia: cortical bone, bone marrow,
and top and bottom trabecular bone). The greyscale value of each pixel represented the amount
of x-ray absorbed by that tissue. These values were based on the relative signal observed
throughout the s anned volume, and vary from “ la k” to “white”. A voxel of righter intensity
indicates one with greater stiffness. Hence, each tissue, depending on its intensity, could be
distinguished from other tissues once defined thresholds are applied. The greyscale values
belonging to the particular tissue types were replaced by a mask. The mask described how a
tissue filled the space. All masks were defined by a binary component (0 or 1), where a value of

160
zero meant that the pixel did not belong to the tissue whereas a value of one meant that the pixel
belonged to the tissue.
In our case, four masks were needed: cortical bone, bone marrow, and proximal and distal
trabecular bone. To render our sample in three dimensions, a threshold-based method was used
to segment the raw data. Each tissue was distinguished by the lower and upper threshold bounds
depending on the range of the tissue greyscale values. In ScanIP, the greyscale values were
represented by integers in the range of 0-255. The appropriate threshold level was determined for
each tissue. The cortical bone could be distinguished visually from the trabecular bone (Figure
5.3). Since the cortical bone was stiffer compared to the trabecular bone, the upper threshold
bound of 255 was used for the cortical. In order to find the optimum lower threshold parameter
for the cortical bone, different masks were created using different lower and upper threshold
bounds: [50, 255], [100, 255], [200, 255] and [225, 255]. In the first three bounds, the trabecular
bone was also included in the mask. Hence, the lower threshold bound for cortical bone was
higher than 200. Different bounds were tested and finally the lower and upper threshold bounds
of 202 and 255 were selected for cortical bone. The upper bound for trabecular bone was set 201
and a similar set of steps were conducted to distinguish the lower threshold bound for trabecular
bone ([1, 201], [50, 201], [100, 201], [150,201]). Values less than 190 were determined to
represent the bone marrow. Therefore, 190 and 201 for trabecular bone, and the values lower
than 190 were used to construct the bone marrow.

161
Figure 5.3. Cortical and trabecular bone could be visually distinguished in the µCT cross-
sectional view of a murine tibia.
The threshold-based method was used to create the mask for the cortical bone. Erroneous holes
and isolated small areas may appear after segmentation due to noise absorption in the scanning
stage. First, the morphological close filter was used to add connectivity to the cortical mask, and
then the cavity fill filter was performed to fill the remaining small holes in the cortical mask. The
avity fill filter filled “internal” holes while preserving the outer boundaries of the marrow
cavity. Thereafter, the island removal filtration was carried out to have a continuum mask and
get rid of the redundant islands (detached segmented pixels in the cortical mask). To facilitate the
meshing process minor editing was necessary to smooth out the sharp edges near the ends of the
tibia. The sharp edges at the ends of the tibia were erased using the paint segmentation tool.
Finally, the re ursive Gaussian filter with the value of σ = 0.045 mm (1.5 times the spacing) was
performed to increase the smoothness of the segmented cortical surfaces (Figure 5.4). The larger
values of σ resulted in smoother surfaces, and it was suggested by the software to use the values
of 1 times to 3 times the spacing of the image data (0.03 mm) for smoothing the model.

162
(a)
(b)
Figure 5.4. Bone geometry before and after applying the recursive Gaussian filter.
After the cortical mask had been created, the trabecular mask was segmented using again the
threshold-based method (lower and upper threshold values, [190, 201]). To create a continuous
mask, initially the morphological close filter was used to merge fine structures of the trabecular
bone. Next, the cavity fill filter was internal holes of the trabecular mask, which are neither
connected to the cortical bone or bone marrow. However, due to the spongy structure of the
trabecular bone, the spaces between the trabeculae were very large and only some of them were
filled using the cavity fill filter. Then, the floodfill filter was used to fill the remaining gaps
between the trabeculae. In case of an overlap (between the cortical and trabecular mask), the
Boolean subtraction was performed and the trabecular mask subtracted from the cortical mask. A
similar set of steps was used to establish the mask for the bone marrow. The created masks are
shown in Figure 5.5. The masks in different locations of the tibia can be seen in Figure 5.5, with
the cortical mask shown in red, trabecular masks in blue and marrow mask in yellow.
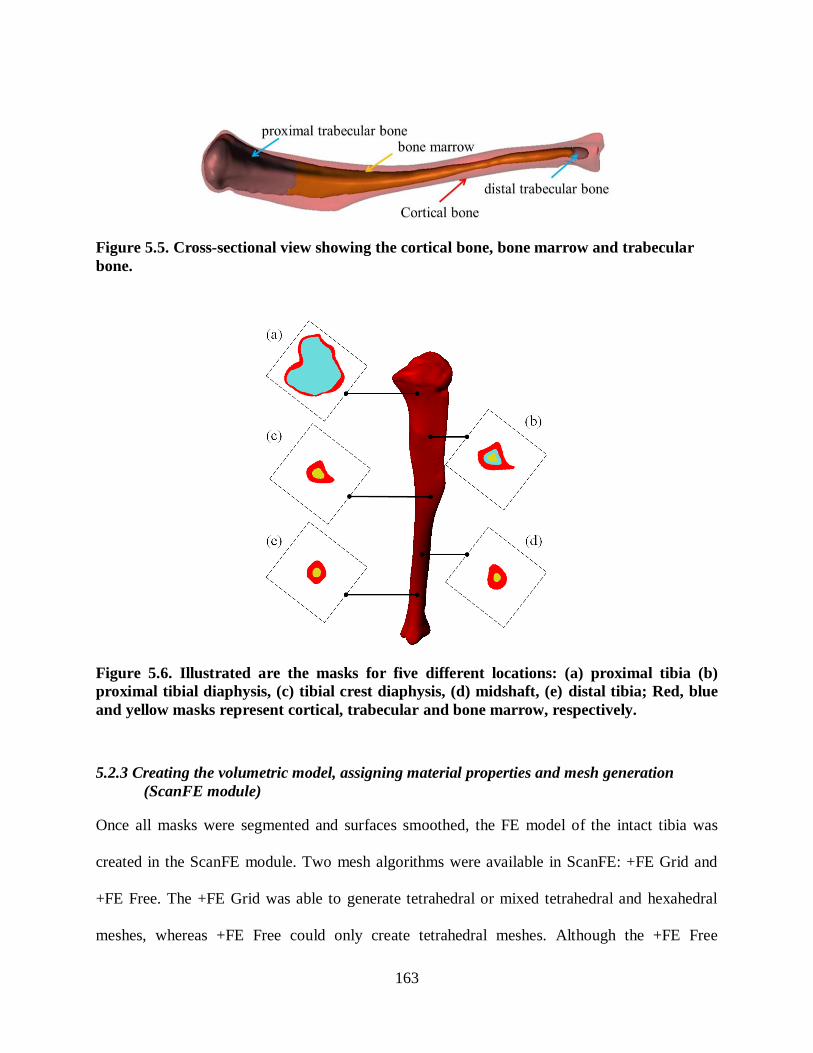
163
Figure 5.5. Cross-sectional view showing the cortical bone, bone marrow and trabecular
bone.
Figure 5.6. Illustrated are the masks for five different locations: (a) proximal tibia (b)
proximal tibial diaphysis, (c) tibial crest diaphysis, (d) midshaft, (e) distal tibia; Red, blue
and yellow masks represent cortical, trabecular and bone marrow, respectively.
5.2.3 Creating the volumetric model, assigning material properties and mesh generation
(ScanFE module)
Once all masks were segmented and surfaces smoothed, the FE model of the intact tibia was
created in the ScanFE module. Two mesh algorithms were available in ScanFE: +FE Grid and
+FE Free. The +FE Grid was able to generate tetrahedral or mixed tetrahedral and hexahedral
meshes, whereas +FE Free could only create tetrahedral meshes. Although the +FE Free

164
approach had slower mesh generation, it had more flexibility in the element creation process and
offered more control over the element size and mesh density. The minimum and maximum edge
length, the target maximum error and the surface change rate could be defined in the advanced
parameter settings. The target maximum error is the distance that the remeshed surface can move
from the original mesh. Surface change rate defines how fast the size of the elements can alter
during meshing; higher rates reduce the mesh quality.
In the present study, the +FE Free meshing algorithm was applied to have more control over the
element size, number and quality. The maximum and minimum target edge length (0.4 and 0.6
mm) and other options were defined in the advanced parameter settings. Three mesh densities
were generated for mesh convergence and sensitivity analysis. The selection of the element size
was made on the basis of a convergence study.
The material properties, e.g. the Young‟s modulus and oisson‟s ratio, were defined in the
material tab of the ScanFE module. Three material type options were available: placeholder,
homogeneous and greyscale based. The material properties could be defined later in the FE
software by choosing the placeholder option. The homogeneous option defined the material
model as linear elasti . A Young‟s modulus and oisson‟s ratio of 2.0 [MPa] and 0.167,
respectively, were specified for the bone marrow (Isaksson et al. 2006). The greyscale based
option automated the material properties for each tissue based on the image density and was used
for the cortical bone and trabecular bone within the present study. The greyscale values were
mapped to a mass density, and the mass density was mapped to a Young‟s modulus and
oisson‟s ratio. The mapping fun tion was assumed to be linear. The greyscale values of each
voxel were mapped to the mass density ( ) using the following equation:
GSβαρ 5.1

165
where GS was the element greyscale value and α and β were the mapping oeffi ients. Young‟s
modulus (E) and oisson‟s ratio (ν) were computed from the mass density for each element
(Equations 5.2, 5.3).
cbρaρE 5.2
dρν 5.3
where a, b and c were defined based on the published equations by Rho et al. (1995) for a
longitudinal compressed human tibia bone (Table 5.1) (Rho et al. 1995, Stadelmann et al. 2009).
The constants (a, b, c and d) defined the mapping function. The range of greyscale based
Young‟s modulus values that were assigned to the masks are shown in Table 5.2. The
Simpleware software uses a formulation derived for humans (Rho et al, 1995). Since there was
no available data that related the murine density to its Young‟s modulus, equation (5.2) was
used. However, experimental studies should be performed in future to scale the formula for the
murine tibia. After assigning the meshing parameters and defining the material properties,
volume meshes were generated in the ScanFE module. Figure 5.7 presents anterior, posterior,
medial and lateral views of the meshed murine tibia. An input file was then generated and
imported into ABAQUS v6.11. Figure 5.8 summarizes the required steps from greyscale data to
generation of the volumetric mesh.
Table 5.1. The relationship between the mechanical properties and mass density (Rho et al.
1995).
Dependent variable Tibial cortical bone Tibial trabecular bone
Density [3mkg ] GS0.916114ρ GS0.916114ρ
Yo ng’s mo l s [MPa] 13ρ-3842E 5.54ρ-326E
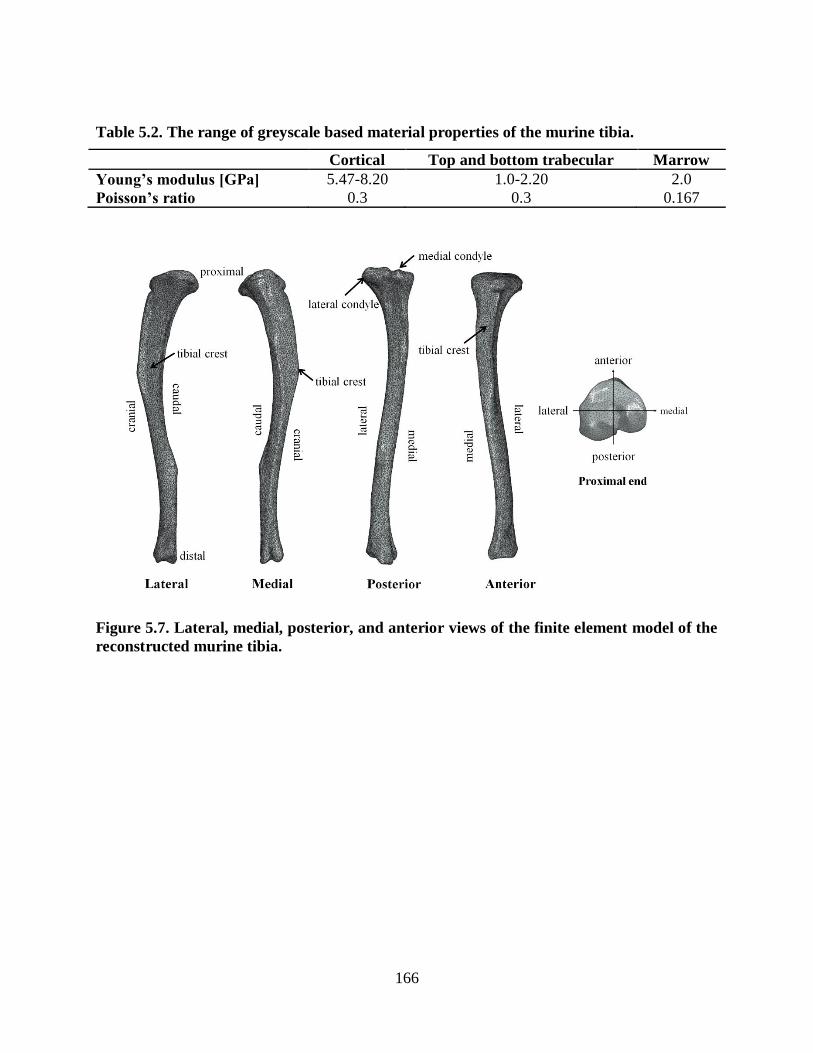
166
Table 5.2. The range of greyscale based material properties of the murine tibia.
Cortical Top and bottom trabecular Marrow
Yo ng’s mo l s [GP ] 5.47-8.20 1.0-2.20 2.0
Poisson’s io 0.3 0.3 0.167
Figure 5.7. Lateral, medial, posterior, and anterior views of the finite element model of the
reconstructed murine tibia.

167
Figure 5.8. (a) Volume image, (b) greyscale data, (c) segmented mask, (d) isolated
segmented mask, (e) smoothed mask (recursive Gaussian filter), (f) mesh generation of the
extracted volume.
5.2.4 Convergence study
A convergence test was carried out to ensure the selected number of elements was sufficient to
predict the mechanical behaviour of the tibia. A linear elastic material model was applied to the
regenerated model. Three meshes with increasing density (74665, 84580 and 96563 elements)
were created to asses mesh convergence. The higher density meshes were obtained using an
minimum element size of 0.4 mm and a maximum element size of 0.6 mm. The meshing with the
medium density was composed of elements in the range of 0.25 to 0.4 mm, and the highest
density meshes were meshed using the element size in the range of 0.2 to 0.4 mm.
Both linear and nonlinear geometric models were simulated to evaluate their effects on the
results. The tissues were composed of 10-node quadratic tetrahedron elements (C3D10). The
convergence study was performed for an applied axial compressive load of 2 N to the proximal
point of the tibia. All the nodes at the distal end were restricted in all directions. To evaluate the

168
mesh sensitivity, the maximum axial displacement in the model, and the summation of the axial
reaction forces at the distal nodes were considered for convergence. The increase in the element
number resulted in less than 0.1 % difference for the maximum displacement, and 0.2 % for the
axial reaction force at the distal end (Table 5.3). Hence, it was concluded that 74665 elements
were adequate for an accurate estimation of the mechanical behaviour within the model.
Table 5.3. Convergence study for three mesh densities.
Total element number 74665 84580 96563
Axial reaction force [N] 1.8439 1.8510 1.8472
Max. displacement [mm] 0.7688 0.7683 0.7691
Error in the axial reaction force [%] 0.177 0.208 -
Error in the max. displacement [%] 0.039 0.104 -
5.3 Verification of the generated FE model of the intact tibia
The mechanical behaviour of the reconstructed tibia was verified against a previously published
numerical-experimental study (Stadelmann et al. 2009). To match the experimental study, the
distal zone of the tibia was completely fixed. Axial compression loads were applied to the
proximal end increasing from zero to 10 N. The force-strain behaviour in a murine tibia was
determined at three different locations (proximal, midshaft and distal tibia) (Figure 5.9). In
Figure 5.9, the flat surface with the widest area at the proximal region represented the proximal
tibia (zone1), the region between the tibial crest and fibula junction was defined as the midshaft
tibia (zone 2), and the region between the fibula junction to the end of the tibia was defined as
the distal tibia (zone 3). The tissues were considered linear elastic and were meshed with 10-
node quadratic tetrahedron elements (C3D10). The material properties used are shown in Table

169
5.2. Principal strain values were estimated numerically at the three different regions of the tibia
(zone 1, zone 2 and zone 3) for comparison with strain-gauge measurements (Stadelmann et al.
2009). Figure 5.10 shows that the strain magnitudes were in good agreement with the
experimental values (maximum 10 % deviation) of Stadelmann et al. (2009). The maximum,
minimum and medium principal strains are shown when the bone was subjected to a 10 N axial
compression load (Figure 5.11).
Figure 5.9. Three zones that were compared: zone 1 (proximal tibia), zone 2 (tibial crest),
zone 3 (distal tibia).
(a) (b)
Figure 5.10. Force-strain relations measured by (a) Stadelmann et al. (2009), and (b)
predicted in the present study.

170
Figure 5.11. The distribution of principal strains when the tibia was subjected to a 10 N
axial compression load.
5.4 Development of the burr-hole fracture model
The geometry and position of the burr-hole fracture were selected based on a previous
experimental study (Taiani et al. 2010). The burr-hole was bored from one side of the cortical
bone into the medullary cavity. Immediately after the injury, the holes were filled with the
collagen gel seeded with stem cells (Figure 5.12) (Taiani 2012). A 0.7 mm burr-hole fracture,
with the depth of 1.07±0.21 mm, was created through the medial cortex and the medullary cavity
of the proximal tibia. The centre of the hole to the top of the epiphysis was 2.49±0.26 mm.
To generate the burr-hole model, the segmented data of the intact mouse tibia were imported into
the ScanCAD module. The CAD primitive generation tool was used to create a cylindrical
scaffold with a diameter of 0.7 mm. The scaffold was positioned within the tibia based on the
burr-hole geometry and location using the CAD manipulation tool. The CAD model of the
cylindrical scaffold was first converted into an image-based mask and then all of the masks
including the cylindrical scaffold mask were exported into ScanIP module. Several Boolean
operations were performed on the masks to create the hole in the proximal tibia (e.g. subtract).

171
The multipart volumetric model was meshed in ScanFE module using the +FE Free meshing
algorithm. The model was meshed using different target edge lengths, and finally the best mesh
was selected based on the information obtained from the element qualities: (1) the mesh statistics
in Simpleware, and (2) performing a mesh sensitivity analysis. In Simpleware, the mesh statistics
was in the log toolbox and gave a summary of the number of elements, nodes, average and the
worst element qualities. The maximum and minimum lengths of the elements for the cortical,
bone marrow and trabecular bone masks were set as 0.4 and 0.6 mm. To ensure accurate
mechanical behaviour within the fracture site (e.g. strain and fluid velocity) a finer discretization
was used to mesh the region of interest (scaffold). An element size of 0.2 mm was set in the
mesh refinement volume tool for the scaffold. After setting the mesh parameters and material
properties, the model was meshed with the total of 79790 elements and the input file was
generated and imported into ABAQUS v6.11 for computational analysis. The stages required to
generate the volumetric mesh are summarized in Table 5.4, and Figure 5.13 and 5.14.
Table 5.4. Elements numbers for different parts of the fracture model.
Cortical Proximal
trabecular
Distal
trabecular Scaffold Marrow Total
Number of
elements 43911 16462 1341 1890 16186 79790

172
Figure 5.12. Location of the burr-hole in the medial aspect of the tibia: (a) FE model, (b)
experimental fracture model (Taiani 2012). A section through the long axis of the burr-
hole: (c) FE model, (d) experimental fracture model (Taiani 2012). A section through the
frontal plane of the fractured tibia: (e) FE model, (f) experimental fracture model (Taiani
2012).

173
µCTImage processing tool (ScanIP):
segmentation of the intact tibia.
CAD integration module (ScanCAD):
creating the scaffold.
converting the CAD model into mask.
creating hole using Boolean operations.
Mesh generation module (ScanFE):
creating the FE model.
the volumetric mesh generation.
Export to ScanIP
Export the meshed model
(input file) to
FE model ABAQUS
Figure 5.13. Workflow diagram outlining the required functions to reconstruct the 3D FE
burr-hole model.
Figure 5.14. Overview of the processes used to create the burr-hole model.
5.4.1 Selection of the decay length model
Saint-Venant‟s prin iple states that the fixed boundaries only have an effect on the stress-strain
distribution within nearby regions which are relatively far away from the indenter. The linear
elastic model of the intact full-length tibia was compared to three different decay length models.
To create the decay length models sections were created in the following regions: (1) the
proximal tibia, (2) the tibial crest, and (3) the distal tibia (Figure 5.15). The models were
subjected to a 10 N axial compression load at the proximal end while the distal end was

174
completely fixed in all models. The distributions of stress and strain in the models were
compared to the full-length model. According to the results the tibial crest model was the one
that had the closest stress-strain distribution to the full-length tibia.
Figure 5.15. The decay length models were subjected to a 10 N load to select the one that
had the closest mechanical environment compared to the full-length model. The red arrow
shows where the load was applied.
The stress-strain distributions within the fractured models of the tibial crest and the full-length
model were also compared. The fractured tibiae were subjected to an axial compression load 10
N at the proximal end while the bottom end was totally fixed. The mechanical environments

175
were also similar in the burr-hole models (Figure 5.16, 5.17, 5.18), in particular in the region of
interest around the burr-hole (Figure 5.19, Figure 5.20).
Figure 5.16. Distribution of von Mises stress: (a) full-length model, (b) decay length model
(tibial crest, medial view).

176
Figure 5.17. Distribution of principal strains: (a) full-length model, (b) decay length model
(tibial crest, medial view).

177
Figure 5.18. Distribution of principal strains: (a) full-length model, (b) decay length model
(tibial crest, lateral view).
Figure 5.19 Distribution of von Mises stress within the scaffold: (a) full-length model, (b)
decay length model (tibial crest, medial view).

178
Figure 5.20 Distribution of principal strains within the scaffold: (a) full-length model, (b)
decay length model (tibial crest, medial view).
5.5 Tissue differentiation predictions within the burr-hole fracture
Several parametric studies were performed with the FE model of the fractured tibia to investigate
the effect of different loading regimes (axial compression, bending), rate of cell diffusion, origin
of the progenitor cells, position of the burr-hole fracture and bone quality (osteoporotic bone)
(Table 5.5).
In the current study, the loads (0.5-2.5 N) were selected based on the previous in vivo studies
(0.5-13 N), (Stadelmann et al. 2009, Zhang and Yokota 2011, Gardner et al. 2006, De Souza et
al. 2005). The peak force acting on the tibia during normal walking was determined to be 1.2 N
(Prasad et al. 2010). In another study, the peak strains at the tibial midshaft were reported for
normal walking (200-300 µƐ) and jumping (400-600 µƐ), (De Souza et al. 2005). According to
the study by Stadelmann et al. the loads between ~1.2-2.1 N and ~2.8-4.2 N produced strains of

179
200-300 µƐ (normal walk) and 400-600 µƐ (jumping) in the murine ti ial rest (Stadelmann et al.
2009, De Souza et al. 2005). Hence, 0.5 N represented a slow walk, 1-2 N represented a normal
walk and 2.5 N represented running.
Load magnitudes play a critical role in fracture healing, lower axial compression loads improved
healing (0.5 N, 1 Hz) compared to higher load magnitudes (2, 1 N, 1 Hz) in a previous in vivo
study (Gardner et al. 2006). The same load magnitudes were used in the current study to
investigate the effect of load magnitudes on fracture healing (2, 1, 0.5 N, 1 Hz).
Mice fractures generally heal by 21 days (Gardner et al, 2006), therefore, the optimal duration of
the progenitor cells to spread throughout the callus was 21 days (21 steps). To find the
appropriate diffusive rate, the mass diffusion analysis was performed using different values of
diffusive rate, and finally using the diffusive rate of 0.025 s
mm 2
, the progenitor cells could
spread throughout the scaffold in selected in 21 days (steps). The diffusion coefficient was
hypothesized to decrease with aging (Chen et al. 2005, Park et al. 2005). A lower diffusive rate
of 0.01 s
mm2
was selected to represent a lower diffusion (elder murine) in which the cells were
partially spread through the scaffold after 21 days. To investigate the effect of diffusion rate on
healing process, the diffusive rates of 0.025 (younger bone) and 0.01 (elder bone) s
mm2
were
used.
The effect of mechanical stimuli on bone healing was explored by comparing two fracture
positions (3.13 mm and 2.55 mm from the proximal end of the tibia). Moreover, the change in
the number of cell origins were investigated: a fracture bored in the trabecular bone with 2 cell
origins and another fracture in the bone marrow with three cell origins (Lacroix 2000).

180
Furthermore, the osteoporotic bones are difficult to heal due to reduced mechanical properties
(Macdonald et al. 2011, Sharir et al. 2008). Osteoporotic bones were reported to have less
stiffness, Young‟s modulus and higher void ratio and permeability compared to the normal bones
(Li and Aspden 1997a, McDonnell et al. 2007, Sun et al. 2008). To investigate the effect of
reduced mechanical properties, a model of an osteoporotic murine tibia was used and its healing
rate compared to a normal bone. Moreover, bending load was reported an effective mean to
enhance bone formation in a murine tibia (Zhang et al. 2006). Hence, lateral loading were given
to the proximal end of the tibia to stimulate tissue differentiation (0.04, 0.02 Nm).
Table 5.5. Summary of the variables for the parametric studies.
Parametric studies Load
Diffusivity
[s
mm2
]
Number of
cell origins
Distance
[mm]
Axial compression 0.5, 1, 2 N 0.025 2 2.55
Cell diffusion rate 1 N 0.01, 0.025 2 2.55
Fracture
position
Trabecular
bone 1 N 0.025 2
2.55
3.13
Trabecular
bone and
bone marrow
2.5 N 0.025 2 2.55
3 4.95
Osteoporotic bone 2.5 N 0.025 2 2.55
Normal bone
Bending 0.02, 0.04
Nm 0.025 2 2.55
For all of the models, a stem cell seeded collagenous scaffold was initially implanted into the
gap. The cortical bone, bone marrow and trabecular bone tissues were defined as poroelastic
materials using 10-node modified quadratic tetrahedron pore pressure elements (C3D10MP). The
modified element used, had three additional displacement variables and one additional pore
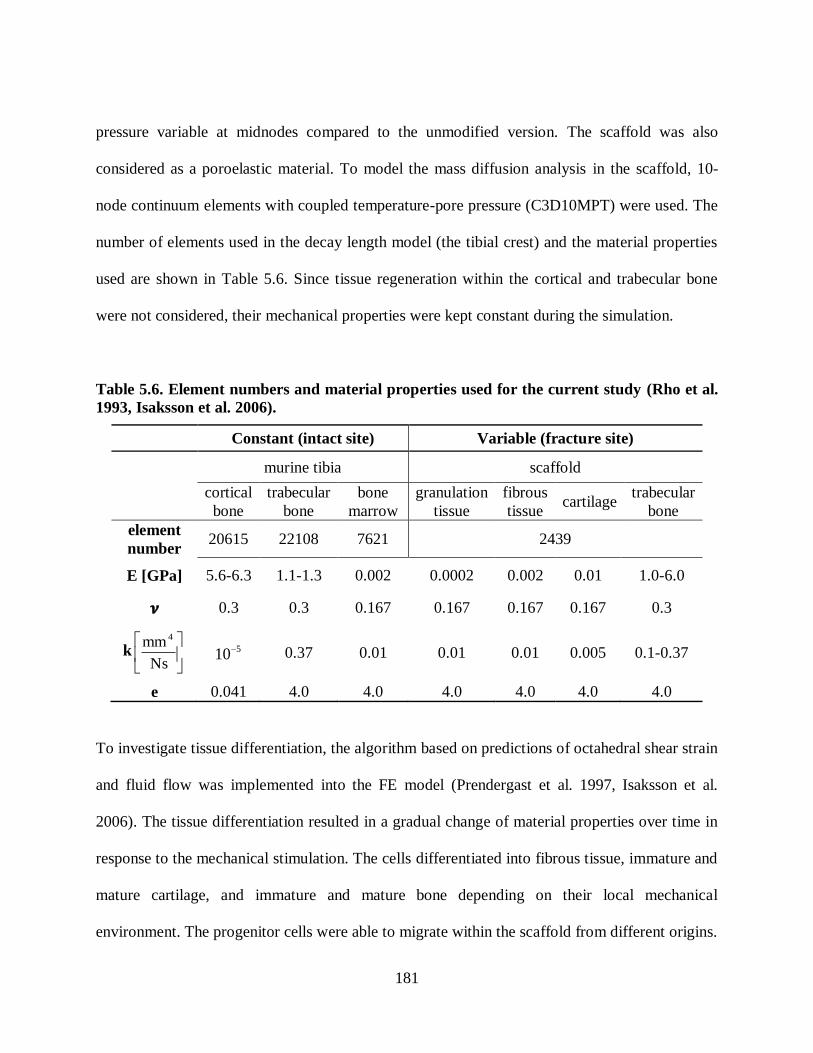
181
pressure variable at midnodes compared to the unmodified version. The scaffold was also
considered as a poroelastic material. To model the mass diffusion analysis in the scaffold, 10-
node continuum elements with coupled temperature-pore pressure (C3D10MPT) were used. The
number of elements used in the decay length model (the tibial crest) and the material properties
used are shown in Table 5.6. Since tissue regeneration within the cortical and trabecular bone
were not considered, their mechanical properties were kept constant during the simulation.
Table 5.6. Element numbers and material properties used for the current study (Rho et al.
1993, Isaksson et al. 2006).
Constant (intact site) Variable (fracture site)
murine tibia scaffold
cortical
bone
trabecular
bone
bone
marrow
granulation
tissue
fibrous
tissue cartilage
trabecular
bone
element
number 20615 22108 7621 2439
E [GPa] 5.6-6.3 1.1-1.3 0.002 0.0002 0.002 0.01 1.0-6.0
0.3 0.3 0.167 0.167 0.167 0.167 0.3
k
Ns
mm 4
510 0.37 0.01 0.01 0.01 0.005 0.1-0.37
e 0.041 4.0 4.0 4.0 4.0 4.0 4.0
To investigate tissue differentiation, the algorithm based on predictions of octahedral shear strain
and fluid flow was implemented into the FE model (Prendergast et al. 1997, Isaksson et al.
2006). The tissue differentiation resulted in a gradual change of material properties over time in
response to the mechanical stimulation. The cells differentiated into fibrous tissue, immature and
mature cartilage, and immature and mature bone depending on their local mechanical
environment. The progenitor cells were able to migrate within the scaffold from different origins.

182
A user-defined subroutine USDFLD was developed to update the material properties based on
the average of computed mechanical stimuli in the previous 10 days. The rule of mixtures was
used to al ulate the new material properties (Young‟s modulus, oisson‟s ratio and
permeability). In each iteration, the mechanical stimuli were calculated at the maximum loading.
The simulation continued until the tissue distribution in the scaffold reached a steady state.
5.5.1 Investigation of axial compression load
The decay length model (the tibial crest) of the reconstructed murine tibia with a 0.7 mm fracture
gap was used to represent a closed burr-hole fracture model (Figure 5.12) (Taiani 2012). Tissue
differentiation patterns were investigated for 2, 1 and 0.5 N (1 Hz) compression loads applied to
the proximal end of the tibia. The distributed loading was selected to avoid stress concentration.
Since the hole was created in the trabecular bone (2.55 mm from the proximal site), the
progenitor cells could migrate into the fracture site from two origins: (1) the surrounding muscle
tissues, and (2) the endosteum (Figure 5.21a). The diffusion coefficients were set such that after
21 days (steps) the progenitor cells would spread throughout the entire fracture zone (Gardner et
al. 2006). Each step represented the average load that had been applied to the bone during one
day of healing (Isaksson et al. 2006).
The granulation tissue gradually differentiated into stiffer tissues over time. Therefore, by
stiffening the scaffold the mechanical stimuli decreased and the interfragmentary gap movement
reached a steady state (Figure 5.23). The tissue distribution changed over time due to the change
of mechanical stimuli and cell concentration. The model predicted an increase in the mechanical
stimuli in cell seeded scaffold in the first few days (days 1-3). Thereafter, the octahedral shear
strain started to reduce while the fluid velocity decreased more gradually (days 4-8) and then
reached a steady state (days 9-21) (Figure 5.22). The predicted magnitudes of the mechanical
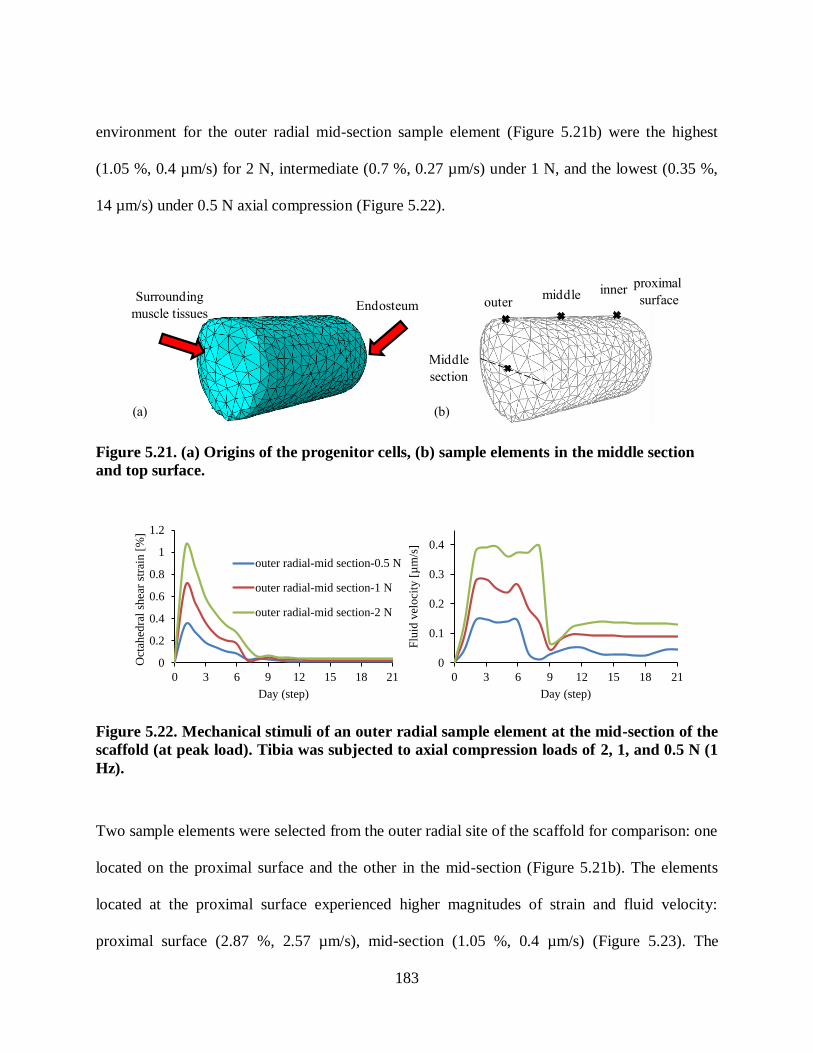
183
environment for the outer radial mid-section sample element (Figure 5.21b) were the highest
(1.05 %, 0.4 µm/s) for 2 N, intermediate (0.7 %, 0.27 µm/s) under 1 N, and the lowest (0.35 %,
14 µm/s) under 0.5 N axial compression (Figure 5.22).
Surrounding
muscle tissuesEndosteum
proximal
surface
Middle
section
outermiddle inner
(a) (b)
Figure 5.21. (a) Origins of the progenitor cells, (b) sample elements in the middle section
and top surface.
0
0.2
0.4
0.6
0.8
1
1.2
0 3 6 9 12 15 18 21
Oct
ahed
ral
shea
r st
rain
[%
]
Day (step)
outer radial-mid section-0.5 N
outer radial-mid section-1 N
outer radial-mid section-2 N
0
0.1
0.2
0.3
0.4
0 3 6 9 12 15 18 21
Flu
id v
eloci
ty [
µm
/s]
Day (step)
Figure 5.22. Mechanical stimuli of an outer radial sample element at the mid-section of the
scaffold (at peak load). Tibia was subjected to axial compression loads of 2, 1, and 0.5 N (1
Hz).
Two sample elements were selected from the outer radial site of the scaffold for comparison: one
located on the proximal surface and the other in the mid-section (Figure 5.21b). The elements
located at the proximal surface experienced higher magnitudes of strain and fluid velocity:
proximal surface (2.87 %, 2.57 µm/s), mid-section (1.05 %, 0.4 µm/s) (Figure 5.23). The
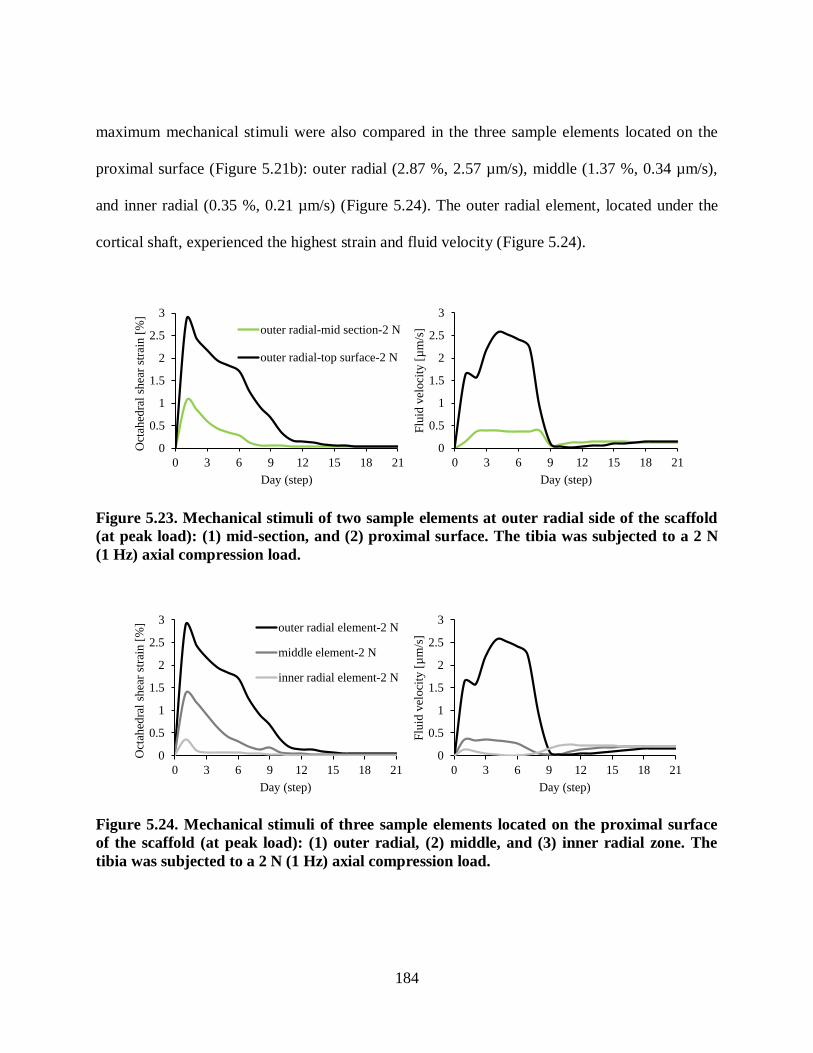
184
maximum mechanical stimuli were also compared in the three sample elements located on the
proximal surface (Figure 5.21b): outer radial (2.87 %, 2.57 µm/s), middle (1.37 %, 0.34 µm/s),
and inner radial (0.35 %, 0.21 µm/s) (Figure 5.24). The outer radial element, located under the
cortical shaft, experienced the highest strain and fluid velocity (Figure 5.24).
0
0.5
1
1.5
2
2.5
3
0 3 6 9 12 15 18 21
Oct
ahed
ral
shea
r st
rain
[%
]
Day (step)
outer radial-mid section-2 N
outer radial-top surface-2 N
0
0.5
1
1.5
2
2.5
3
0 3 6 9 12 15 18 21
Flu
id v
eloci
ty [
µm
/s]
Day (step)
Figure 5.23. Mechanical stimuli of two sample elements at outer radial side of the scaffold
(at peak load): (1) mid-section, and (2) proximal surface. The tibia was subjected to a 2 N
(1 Hz) axial compression load.
0
0.5
1
1.5
2
2.5
3
0 3 6 9 12 15 18 21
Oct
ahed
ral
shea
r st
rain
[%
]
Day (step)
outer radial element-2 N
middle element-2 N
inner radial element-2 N
0
0.5
1
1.5
2
2.5
3
0 3 6 9 12 15 18 21
Flu
id v
elo
city
[µ
m/s
]
Day (step)
Figure 5.24. Mechanical stimuli of three sample elements located on the proximal surface
of the scaffold (at peak load): (1) outer radial, (2) middle, and (3) inner radial zone. The
tibia was subjected to a 2 N (1 Hz) axial compression load.

185
During the fracture healing period, the interfragmentary strain was calculated for the three
loading cases under the cortical shaft as follows:
gap
3
entaryinterfragml
Δuε . 5.4
where entaryinterfragmε was the interfragmentary strain, 3Δu was the interfragmentary motion in axial
direction, and gapl was the gap size of 0.7 mm. The 2 N loading case had the maximum
interfragmentary strain (0.3 %) whereas the 1 and 0.2 N loading cases had the strains of 0.2 and
0.1, respectively (Figure 5.25).
0
0.05
0.1
0.15
0.2
0.25
0.3
0 3 6 9 12 15 18 21
Inte
rfra
gm
enta
ry s
trai
n [
%]
Day (step)
0.5 N 1 N 2 N
Figure 5.25. The predicted interfragmentary strain, at peak load, under the cortical shaft
for the three loading cases (2, 1, 0.5 N axial compression, 1 Hz).
The predicted healing patterns are shown in Figure 5.26. In all cases, the bone formation initiated
from the outer edges of the scaffold and gradually proceeded the inner regions. The 0.5 N
loading case had accelerated healing compared to others (e.g., see day 11 in Figure 5.26), which
was due to the lower mechanical stimuli that the elements were experiencing in the 0.5 N loading
case. However, under 0.5 N axial compression load, the mechanical stimuli had only a marginal

186
amount of bone formation at the inner radial location of the scaffold and fibrous tissue was still
present (0.5 N, Day 14, Figure 5.26), while for the two other cases bony tissue was formed (2
and 1 N, Day 14, Figure 5.26).
fibrous tissue immature cartilage mature cartilage immature bone mature bone
0.5 N
1 N
2 N
Day 6 Day 9 Day 12
Day 6 Day 11
Day 14
Day 9
Day 9Day 6 Day 11
Day 11
Day 12
Day 12
Day 14
Day 14
Figure 5.26. Predicted fracture healing patterns under the 2, 1 and 0.5 N (1 Hz) axial
compression load.
For all of the axial loading cases, bony tissue started to form from both proximal and distal sides,
which may be due to the initially higher concentration of progenitor cells at those regions
compared to other zones (i.e. outer and inner radial sides of the scaffold were the origins of the
progenitor cells). In the 2 N loading case, the inner radial side had increased healing compared to
the middle and outer radial sides. This may be due to the lower mechanical stimuli at the inner
radial (e.g. strain on day 11, inner: 0.0096 %, middle: 0.05 %, outer: 0.18 %, Figure 5.24), (2 N,
Days 11-14, Figure 5.26). On day 12 (2 N) the zone under the cortical shaft had differentiated
into cartilaginous tissue and bony tissue did not form. This showed the zone under the cortical

187
shaft was subjected to higher magnitudes of displacement, strain and fluid velocity (e.g. day 12
strain, outer: 0.14 %, middle: 0.03 %, inner: 0.0094 %, Figure 5.24).
The core of the scaffold had an increased rate of healing compared to the outer layers (Figure
5.27). For the 2 N loading case the inner core had differentiated into mature cartilage at day 10
(Day 10, Figure 5.27) whereas the outer layer was still immature cartilage. On day 11, the inner
tissue had differentiated into immature bone whereas the outer layer was still cartilaginous (Day
11, Figure 5.27). The same sequence could be observed for differentiation into mature and
immature bone for the inner core compared to the outer layers of the scaffold (Days 12-14,
Figure 5.27). This may be due to higher mechanical stimuli at the outer layers compared to the
core (e.g. on day 13 strain, outer layer: 0.12 %, core: 0.04 %) (Figure 5.23).
immature cartilage mature cartilage immature bone mature bone
Day 11Day 10 Day 12 Day 13 Day 14
Figure 5.27. Cross-sectional view of the scaffold showing the accelerated healing of the core
compared to the outer layers. The tibia was subjected to a 2 N (1 Hz) axial compression
load.
5.5.2 Influence of cell diffusivity rate
To investigate the effect of cell diffusion rate, two magnitudes of mass diffusivity were
investigated and the healing patterns compared. The burr-hole fracture model with a 0.7 mm gap
located in the trabecular bone (2.55 mm from the proximal end) was used (Figure 5.12). The
proximal end of the tibia was subjected to a cyclic 1 N (1 Hz) axial compression load and the cell

188
diffusivities were set 0.025 and 0.01 s
mm 2
. The higher diffusion constant (0.025 s
mm2
)
represented a younger bone compared to the 0.01 s
mm2
case (Geris et al. 2009). Mice fractures
generally heal by 21 days (Gardner et al, 2006). Using the diffusive rate of 0.025 s
mm2
, the
progenitor cells could spread throughout the entire scaffold after 3 weeks (21 steps). On the other
hand, the diffusive rate of 0.01 s
mm2
, represented a lower diffusion in which cells were partially
spread through the scaffold after 21 days. The progenitor cells were able to migrate faster in the
model with a higher mass diffusion constant. The octahedral shear strain was computed for the
middle sample element at the proximal surface in both models (Figure 5.21b, Figure 5.28). The
octahedral shear strain was similar in the initial stages indicating that the cell concentration did
not differ greatly at the beginning of the analysis (Figure 5.28). However, the cells migrated
quicker to the middle zone in the model with a higher diffusivity (0.025 s
mm2
) and the rate of
differentiation was accelerated (Figure 5.29). The model with a high diffusion rate led to a stiffer
matrix in the scaffold and thus lower octahedral shear strain (Figure 5.28). The healing pattern
showed that both models had the same tissue patterns in the initial stages (Day 6, Figure 5.29).
However, in the higher diffusion case, bone had formed by day 14 whereas the fracture site was
still cartilaginous in the lower diffusion case (Figure 5.29).
During aging, the ability of bone to recruit progenitor cells is reduced (Bailón-Plaza and van der
Meulen 2003, Geris et al. 2009). This partly explains why fractures heal quicker in youth than in
adults (Bailón-Plaza and van der Meulen 2003, Geris et al. 2009). The sensitivity of the cells to

189
mechanical perturbations is reduced (Lang 2011). In agreement with the literature, the fracture
healed quicker in the young bone (with higher diffusion rate) compared to the older bone with
lower diffusion rate (Figure 5.29).
0
0.2
0.4
0.6
0.8
1
0 3 6 9 12 15 18
Oct
ahh
edra
l sh
ear
stra
in [
%]
Day (step)
D = 0.01 D= 0.025
Figure 5.28. The prediction of octahedral shear strain for different cell diffusion rates
(0.025 and 0.01 s
mm2
).
granulation tissue fibrous tissue immature cartilage mature cartilage immature bone mature bone
D = 0.025
Day 9Day 6 Day 11 Day 12 Day 14
Day 9Day 6 Day 11 Day 12 Day 14
D = 0.01
Day 7
Day 7 Day 9
Day 9 Day 11
Day 11 Day 12
Day 12 Day 14
Day 14
Figure 5.29. Predicted fracture healing patterns under the 1 N (1 Hz) axial compression
load for different rates of cell diffusion (0.025 and 0.01 s
mm2
).

190
5.5.3 Influence of fracture position
To determine the effect of fracture position on healing two parametric studies were performed:
1. The position of fractures differed axially, but both were located in the trabecular bone to
explore the effect of changes in mechanical stimuli on tissue differentiation.
2. The position of fractures differed axially, with one located in the trabecular bone and the
other located in the bone marrow. In these cases, both the number of progenitor cells and
the mechanical stimuli differed.
5.5.3.1 Different mechanical stimuli with the same cell origins
To investigate the effect of fracture position on the healing rate two different burr-hole fractures
with the same diameter (0.7 mm) were created in the trabecular bone (both had the same cell
origins and diffusivity). The holes were located at different axial positions with respect to the
proximal end of the tibiae: 2.55 mm versus 3.13 mm (Figure 5.30). An axial compression load of
1 N (1 Hz) was applied to the proximal end of the tibia and the predicted healing patterns were
contrasted for the two models.

191
2.55 mm3.13 mm
(a) (b)
Figure 5.30. Axial positions of the burr-hole fracture: (a) 2.55 mm and (b) 3.13 mm from
the proximal end. Both fractures are located in the trabecular bone and the tibia was
subjected to a 1 N (1 Hz) axial compression load. Bar = 0.7 mm.
The predicted mechanical stimuli were smaller in the 3.13 mm fracture case (Figure 5.31). For
the 2.55 mm fracture case the maximum octahedral shear strain and fluid velocity were 1.91 %
and 1.65 µm/s, whereas 0.76 % and 0.65 µm/s for the 3.13 mm fracture case, respectively. The
lower mechanical stimuli better promote bone formation and as expected the fracture farther
from the proximal end exhibited accelerated healing (Figure 5.32). In the initial days, the
increased mechanical stimuli delayed fibrous tissue differentiation in the 2.55 mm fracture case,
whereas most of the scaffold was composed of immature cartilage in the 3.13 mm fracture case
(Day 6, Figure 5.32). By days 10 and 13 the immature bone (in yellow) was more widely
distributed in the 3.13 mm case compared to the 2.55 mm case (Figure 5.32). In the 3.13 mm
fracture case most of the cross section was composed of the mature bone (orange), however, for
the 2.55 mm fracture case, the mature bone at the core of the scaffold was surrounded by the
immature bone at the outer layer (Figure 5.33).
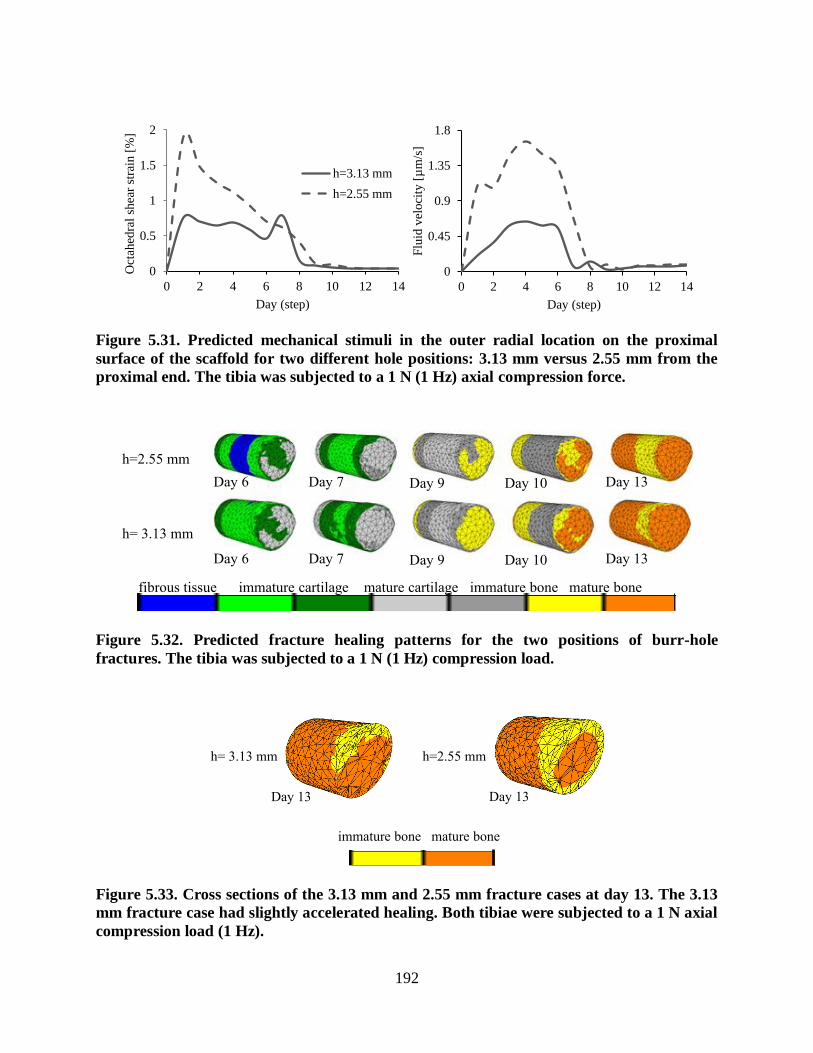
192
0
0.5
1
1.5
2
0 2 4 6 8 10 12 14
Oct
ahed
ral
shea
r st
rain
[%
]
Day (step)
h=3.13 mm
h=2.55 mm
0
0.45
0.9
1.35
1.8
0 2 4 6 8 10 12 14
Flu
id v
eloci
ty [
µm
/s]
Day (step)
Figure 5.31. Predicted mechanical stimuli in the outer radial location on the proximal
surface of the scaffold for two different hole positions: 3.13 mm versus 2.55 mm from the
proximal end. The tibia was subjected to a 1 N (1 Hz) axial compression force.
fibrous tissue immature cartilage mature cartilage immature bone mature bone
h=2.55 mm
Day 7Day 6 Day 9 Day 10 Day 13
Day 7Day 6 Day 9 Day 10 Day 13
h= 3.13 mm
Figure 5.32. Predicted fracture healing patterns for the two positions of burr-hole
fractures. The tibia was subjected to a 1 N (1 Hz) compression load.
immature bone mature bone
h=2.55 mm
Day 13Day 13
h= 3.13 mm
Figure 5.33. Cross sections of the 3.13 mm and 2.55 mm fracture cases at day 13. The 3.13
mm fracture case had slightly accelerated healing. Both tibiae were subjected to a 1 N axial
compression load (1 Hz).

193
5.5.3.2 Different mechanical stimuli and cell origins
To investigate the effect of both fracture position and origin of progenitor cells on the healing
rate, two different burr-hole fractures with the same diameter (0.7 mm) were created in the
trabecular bone (case A) and bone marrow (case B). The fracture located in the trabecular bone
had 2 cell origins (Figure 5.21a) whereas the one within the bone marrow had three cell origins
(Figure 5.34). The holes were located at different axial positions with respect to the proximal end
of the tibiae: 2.55 mm versus 4.95 mm. An axial compression load of 2.5 N (1 Hz) was applied
to the proximal end of the tibia. A higher load magnitude (2.5 N instead of 1 N) was used to
ensure the mechanical stimuli within the 4.95 mm fracture case was not negligible and was
adequate to stimulate the tissue differentiation. The octahedral shear strain and fluid velocity
were negligible in the initial days and increased over time in case B. Then with stiffening of the
scaffold, the mechanical stimuli decreased gradually and reached a steady state (Figure 5.35).
Surrounding
muscle tissuesEndosteum
Bone marrow
Figure 5.34. The origins of progenitor cells when the fracture is located in the bone
marrow.

194
0
1
2
3
4
5
0 3 6 9 12 15 18
Oct
ahed
ral
shea
r st
rain
[%
]
Day (step)
Trabecular bone
Bone marrow
0
0.5
1
1.5
2
2.5
3
3.5
4
4.5
0 3 6 9 12 15 18
Flu
id v
eloci
ty [
µm
/s]
Day (step)
Figure 5.35. Predicted mechanical stimuli for two locations of the fracture (outer radial
sample element on the proximal surface): (1) in trabecular bone and (2) in bone marrow.
Tibia was subjected to a 2.5 N (1 Hz) compression load.
The predicted patterns of tissue differentiation showed that healing accelerated in Case B (Figure
5.36). By day 6 considerable amounts of fibrous tissue existed in the scaffold for case A,
however, in case B immature and mature cartilage were already observed (Day 6, Figure 5.36).
After 13 days, cartilaginous tissue still existed under the cortical shaft of case A, whereas tissue
had differentiated into bone in case B. This may be due to the lower mechanical stimuli,
particularly fluid velocity, and the higher concentration of progenitor cells in case B. Since the
progenitor cells originated from three origins in case B, the cells migrated through the fracture
site faster than case A (with only two cell origins). Therefore, a greater source of progenitor cells
accelerated the healing period (Figure 5.36).
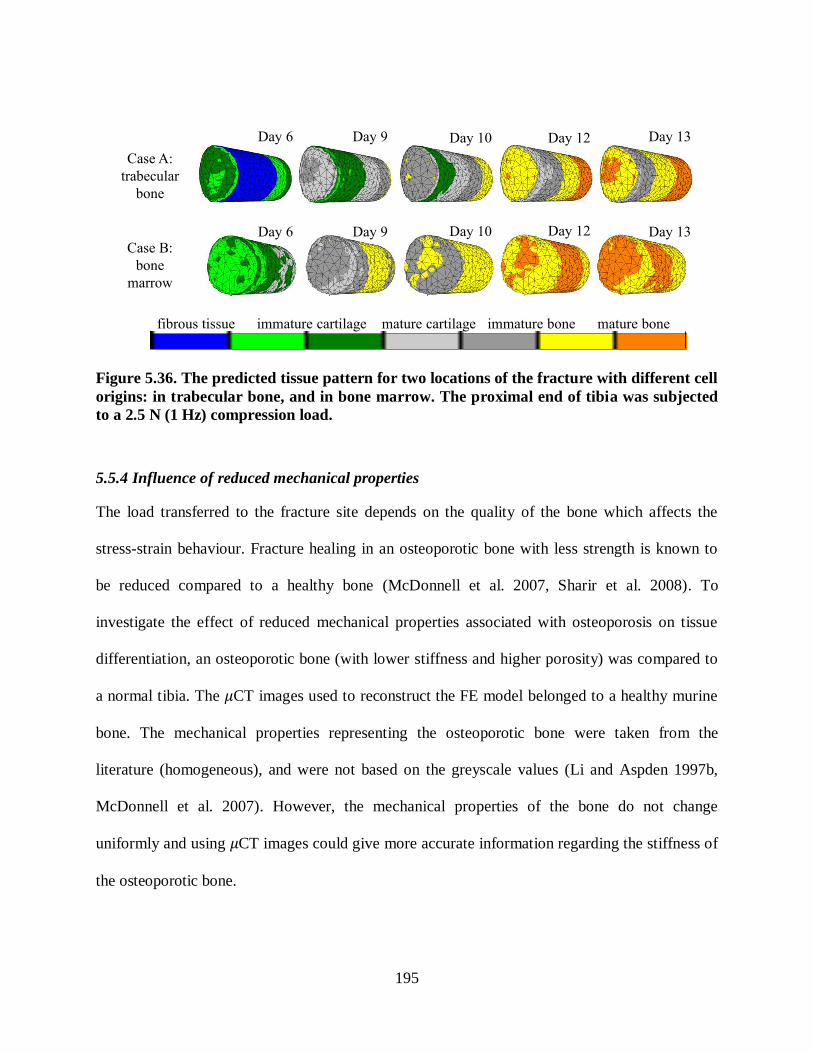
195
fibrous tissue immature cartilage mature cartilage immature bone mature bone
Case A:
trabecular
bone
Day 9Day 6 Day 10 Day 12 Day 13
Day 9Day 6 Day 10 Day 12 Day 13
Case B:
bone
marrow
Figure 5.36. The predicted tissue pattern for two locations of the fracture with different cell
origins: in trabecular bone, and in bone marrow. The proximal end of tibia was subjected
to a 2.5 N (1 Hz) compression load.
5.5.4 Influence of reduced mechanical properties
The load transferred to the fracture site depends on the quality of the bone which affects the
stress-strain behaviour. Fracture healing in an osteoporotic bone with less strength is known to
be reduced compared to a healthy bone (McDonnell et al. 2007, Sharir et al. 2008). To
investigate the effect of reduced mechanical properties associated with osteoporosis on tissue
differentiation, an osteoporotic bone (with lower stiffness and higher porosity) was compared to
a normal tibia. The CT images used to reconstruct the FE model belonged to a healthy murine
bone. The mechanical properties representing the osteoporotic bone were taken from the
literature (homogeneous), and were not based on the greyscale values (Li and Aspden 1997b,
McDonnell et al. 2007). However, the mechanical properties of the bone do not change
uniformly and using CT images could give more accurate information regarding the stiffness of
the osteoporotic bone.

196
The Young‟s modulus of the orti al one was al ulated using a relation etween the stiffness
and density of the osteoporotic bone as follows (Li and Aspden 1997a):
22.1ρ21.4E . 5.5
where ρ = 1.02 is the density 3cmgr
and E is the Young‟s modulus [G a].
The Young‟s modulus and void ratio of the orti al one were assumed to e 1.23 G a and 0.22
for osteoporotic bone (Li and Aspden 1997b, McDonnell et al. 2007). A value of 0.4 GPa and 11
were used for the Young‟s modulus and void ratio of the tra e ular one (Sun et al. 2008). The
permeability of the osteoporotic trabecular and cancellous bone were selected based on a
parametric study performed by Lacroix (2000). In this parametric study, three different
magnitudes of permeability were used (high, middle and low) to find the best to represent normal
bone. The middle value was selected to represent the permeability of the normal bone (Lacroix
2000). Since the osteoporotic bone had greater porosity the highest values were selected for
cortical (0.001 Ns
mm 4
) and trabecular bone (0.5 Ns
mm 4
) (McDonnell et al. 2007). The oisson‟s
ratio of an osteoporotic murine tibia was not available in the literature; a oisson‟s ratio similar
to that of a healthy bone was used. However, further research is needed to obtain more realistic
values for the oisson‟s ratio. The me hani al properties used for the osteoporoti one are
shown in Table 5.7.
A model of the murine tibia with a 0.7 mm fracture gap at an axial distance of 2.55 mm from the
proximal end of the tibia was used (Figure 5.12) (Taiani 2012). An axial compression load of 2.5
N (1 Hz) was applied to the proximal end of the tibia to simulate the running (De Souza et al.
2005, Stadelmann et al. 2009, Prasad et al. 2010). The fracture was located in the trabecular

197
bone, and surrounding muscle tissues and endosteum were considered as the two origins of
progenitor cells in both cases. The mechanical stimuli, interfragmentary strain and healing
patterns were compared between normal and osteoporotic bone (Figure 5.39-5.39).
Table 5.7. Mechanical properties used for an osteoporotic murine tibia (Li and Aspden
1997b, Lacroix 2000, Macdonald et al. 2011, Sun et al. 2008, Isaksson et al. 2006). The
values can be contrasted to those for healthy bone in Table 5.6.
Constant (intact site) Variable (fracture site)
murine tibia scaffold
cortical
bone
trabecular
bone
bone
marrow
granulation
tissue
fibrous
tissue cartilage
trabecular
bone
element
number 20615 22108 7621 2439
E [GPa] 1.23 0.4 0.002 0.0002 0.002 0.01 1.0-6.0
0.3 0.3 0.167 0.167 0.167 0.167 0.3
k
Ns
mm 4
310 0.5 0.01 0.01 0.01 0.005 0.1-0.37
e 0.22 8.1 4.0 4.0 4.0 4.0 4.0
The maximum octahedral shear strain and fluid velocity within the fracture were much higher in
the osteoporotic bone compared to the normal bone (Figure 5.37). This may be explained by
much lower stiffness, higher permeability and void ratio of the surrounding osteoporotic bone.
The octahedral shear strain and fluid velocity were 14 % and 8.5 µm/s for the osteoporotic bone
and only 4.8 % and 4.3 µm/s for the normal bone, respectively (Figure 5.37). The
interfragmentary strain was also calculated for the fracture gap (under the cortical shaft). The
interfragmentary strain within the osteoporotic bone was 4.6 times higher than in the normal
bone (Figure 5.38). The higher values of mechanical stimuli increased the motion within the
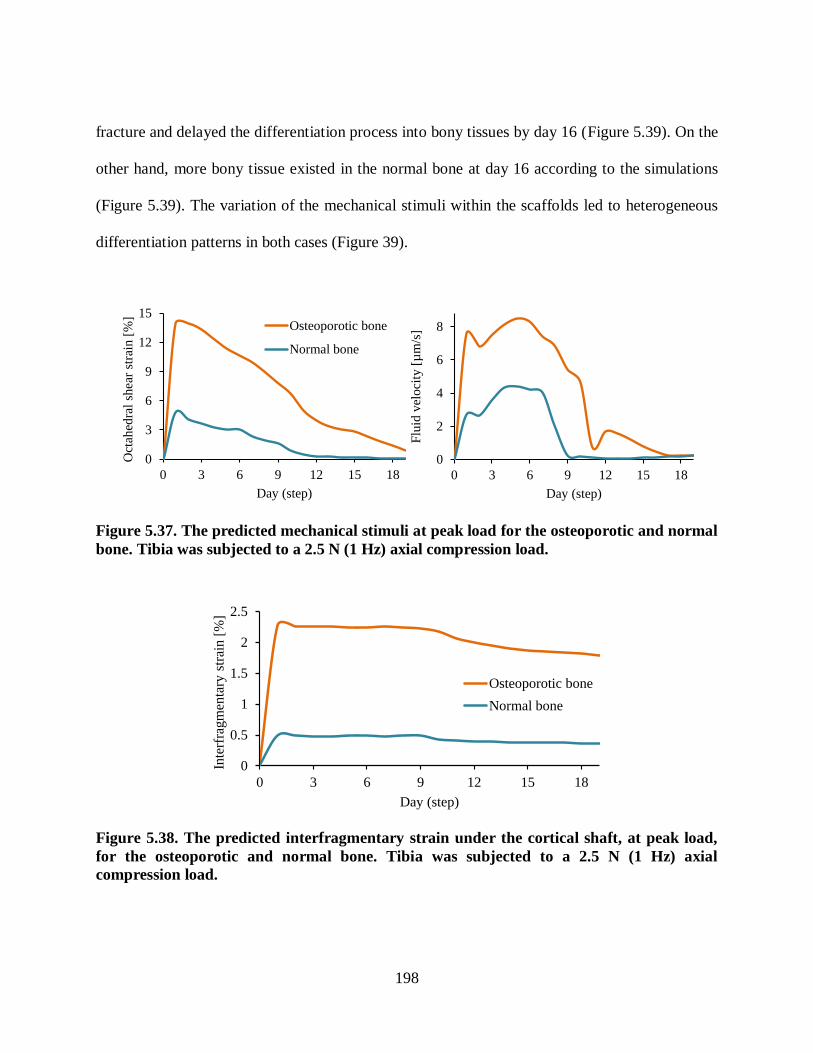
198
fracture and delayed the differentiation process into bony tissues by day 16 (Figure 5.39). On the
other hand, more bony tissue existed in the normal bone at day 16 according to the simulations
(Figure 5.39). The variation of the mechanical stimuli within the scaffolds led to heterogeneous
differentiation patterns in both cases (Figure 39).
0
3
6
9
12
15
0 3 6 9 12 15 18
Oct
ahed
ral
shea
r st
rain
[%
]
Day (step)
Osteoporotic bone
Normal bone
0
2
4
6
8
0 3 6 9 12 15 18
Flu
id v
eloci
ty [
µm
/s]
Day (step)
Figure 5.37. The predicted mechanical stimuli at peak load for the osteoporotic and normal
bone. Tibia was subjected to a 2.5 N (1 Hz) axial compression load.
0
0.5
1
1.5
2
2.5
0 3 6 9 12 15 18
Inte
rfra
gm
enta
ry s
trai
n [
%]
Day (step)
Osteoporotic bone
Normal bone
Figure 5.38. The predicted interfragmentary strain under the cortical shaft, at peak load,
for the osteoporotic and normal bone. Tibia was subjected to a 2.5 N (1 Hz) axial
compression load.

199
The fracture site (scaffold) of the osteoporotic bone was filled mainly with the fibrous tissue in
the initial stages of healing (day 7), whereas the fibrous tissue had differentiated into
cartilaginous tissue in the normal bone at day 7 (Figure 5.39). The fracture model of the normal
tibia had accelerated healing compared to the osteoporotic bone. The delayed healing of the
osteoporotic bone may be due to higher magnitudes of mechanical stimuli at fracture site (Figure
5.37).
granulation tissue fibrous tissue immature cartilage mature cartilage immature bone mature bone
Normal
Day 9Day 7 Day 11 Day 14 Day 16
Day 9Day 7 Day 11 Day 14 Day 16
Osteoporotic
Figure 5.39. The predicted tissue patterns for fracture repair within osteoporotic versus
normal bone. Tibia was subjected to a 2.5 N (1Hz) axial compression load.
5.5.5 Influence of bending load
To explore the effect of bending load, 2 (0.04 Nm, 1 Hz) and 1 N (0.02 Nm, 1 Hz) posteriorly
directed loads were applied to the proximal tibia to cause bending (0.04, 0.02 Nm). The model of
a burr-hole fracture with a 0.7 mm gap located in the trabecular bone (2.55 mm from the
proximal point) was used (Figure 5.40). The mechanical stimuli were compared for both loading
cases. Since the bone was subjected to a lower bending load in the 0.02 Nm, the case mechanical
stimuli were lower (Figure 5.41). The fracture stiffened in both cases over time and the
mechanical stimuli reached a steady state by day 11 (Figure 5.41, Figure 5.42). The fracture

200
region in the bone for the 0.02 Nm case had differentiated into immature bone by day 10 whereas
the 0.04 Nm loading still contained cartilaginous tissue. By day 11 most of the fracture site had
differentiated into mature bone in the 0.02 Nm case, whereas for the 0.04 Nm case cartilaginous
tissue as well as immature bone were existed (Figure 5.41). This indicated that the higher
mechanical bending load might lead to a delay in tissue differentiation and fracture healing.
Figure 5.40. Red arrow shows the bending load that was applied to the tibia. The distal end
was fixed and the hole size was 0.7 mm. Bar = 0.7 mm.

201
0
0.1
0.2
0.3
0.4
0.5
0.6
0 3 6 9 12 15 18 21 24
Oct
ahed
ral s
hea
r st
rain
[%
]
Day (step)
0.02 Nm
0.04 Nm
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0 3 6 9 12 15 18 21 24
Flu
id v
elo
city
[µ
m/s
]
Day (step)
Figure 5.41. Predicted mechanical stimuli for the outer radial sample element at located at
the proximal surface of the scaffold. The tibia was subjected to 0.04 and 0.02 Nm (1 Hz)
bending loads.
granulation tissue fibrous tissue immature cartilage mature cartilage immature bone mature bone
0.02 N
Day 7Day 5 Day 9 Day 10 Day 11
Day 7Day 5 Day 9 Day 10 Day 11
0.04 N
Figure 5.42. Predicted tissue pattern for the murine tibia subjected to bending loads of 0.04
and 0.02 Nm (1 Hz).
5.6 Summary and discussion
A µCT based FE model of a murine tibia was developed to investigate the healing patterns in a
burr-hole fracture model. Greyscale-based material properties were used for the cortical and
trabecular tissues. Initially, the mechanical behaviour of the intact tibia was verified against a
previous experimental-numerical study (Stadelmann et al. 2009). Thereafter, burr-hole fracture
models were created to investigate the tissue differentiation within the fracture site under

202
different conditions (axial compression load, diffusivity rate, fracture position, osteoporotic bone
and bending load) using the biphasic mechanoregulatory algorithm (Lacroix et al. 2002).
Accelerated fracture healing was predicted in conditions where the tibia was subjected to lower
magnitude of axial compression load (0.5 N, 1 Hz), which was in agreement with the previous
experimental study of Gardner et al. (2006). When tibia was subjected to higher loads (2, 1 N, 1
Hz), the bone formation was delayed and the fracture site was composed of less bony tissue.
Hence, the stiffness of the scaffold under the 0.5 N axial compression load was higher than other
cases. The axial compression loads of 2 and 1 N (1 Hz) produced strains at the fracture site,
which were higher than the threshold values proposed by Prendergast et al. (1997) and were not
appropriate for bone differentiation. The elements located at the outer layer of the scaffold were
exposed to higher range of mechanical stimuli (Figure 5.23), and therefore tissue differentiation
was delayed (Figure 5.27). The magnitudes of mechanical stimuli regulated healing pathways
within different regions of the scaffold and the differentiation of the bony tissue initiated from
regions that experienced less mechanical stimuli (the core of the scaffold). There were
inadequate histological slides or CT images to show where the bony tissue was initially formed
at the fracture. He et al. (2011) investigated the fracture repair in a burr-hole murine femur (in
vivo), and was shown that in the formation of bone initiated from outer layers. Unlike our case
that the fracture site was filled with a stem cell seeded scaffold, the burr-hole in the murine
femur had not been filled with a scaffold and the fracture was supposed to heal on its own (He et
al. 2011). The existence of the scaffold and different loading condition in our model may
resulted the difference in the regions that bony tissue was initially formed.

203
To investigate the effect of cell diffusion rate, the healing progression was compared using
different rates of diffusivity (young murine: D=0.025 s
mm 2
and old murine: D=0.01
s
mm 2
).
Initially all of the elements were assumed to be composed of granulation tissue (E=0.2 MPa).
During the healing period, the stiffness of the elements was gradually increased by
differentiating into fibrous tissue, cartilaginous tissue or bony tissue (Figure 5.43). The stiffness
of the scaffold was computed using the rule of mixtures for each day (step) of healing (Figure
5.44). In both cases, a gradual increase in the stiffness of the scaffold was observed during the
healing period; however, the model with a higher rate of cell migration had accelerated healing
compared to the model with a slower rate of cell migration (Figures 5.43-44). The computational
predictions were compared qualitatively with the experimental results from Lu et al. (2005),
which investigated the healing progression of closed fractures in young and adult murine tibia. In
the experimental study, the mice were able to move freely after the injury. The histological slides
and molecular analyses showed that the elder mice had a delayed endochondral ossification and
required a longer healing period (Figure 5.45a-b, Lu et al. 2005). However, younger mice with
higher concentration of progenitor cells were more metabolically active and had a faster fracture
healing. At day 7, Lu et al. (2005) observed a large amount of cartilaginous tissue in the
histological slide of the younger mice. The computational simulation also predicted a small
amount of cartilaginous tissue presented in the older murine by Day 7 (Figure 5.45). Both
computational and in vivo studies, predicted an accelerated healing for the juvenile tibiae. The
experimental and computational data suggested that the rate of cell diffusion affected the healing
process. Therefore, increasing the rate of diffusion in elderly patients may promote the process of
mineralisation and fracture repair and may lead to development of novel treatments.

204
Mature bone
Fibrous tissue
Cartilage
Immature bone
Granulation tissue0
0.5
1
1.5
2
2.5
3
3.5
4
0 2 4 6 8 10 12 14 16 18
Day (step)
D=0.025 D=0.01
Figure 5.43. The gradual change of tissue type during the healing period within a young
(D=0.025 s
mm 2
), and an old murine tibia (D=0.01 s
mm 2
).
Figure 5.44. The overall stiffness of the scaffold during the healing process for young and
old murine tibia with different diffusion rates (D=0.025 s
mm 2
and D=0.01
s
mm 2
).

205
(c) (d)
granulation fibrous tissue immature /mature cartilage
Young Old
Day 7 Day 7 (Lu et al., 2005)
Young Old
Day 7 Day 7 (Lu et al., 2005)
OldYoung
(a) (b)
Young Old
Figure 5.45. Histological slides (Day 7): (a) a large and (b) a small amount of cartilage were
present in the fracture site of young and elder murine tibia, respectively. In the histological
slides, cartilage is shown in red (Lu et al. 2005). In the computational study (Day 7)
similarly, more cartilaginous tissue differentiated in the younger murine tibia (c, d).
Two models with different fracture positions but same number of cell origins were created to
explore the effect of fluid velocity and octahedral shear strain on the healing process. The
fracture models were located 2.55 mm and 3.13 mm from the proximal tibia and the cells
migrated into the fracture site from endosteum and surrounding muscle tissues. The fracture
located further from the proximal tibia (3.13 mm) was predicted to have an accelerated healing
compared to the 2.55 mm case. Since the cell rate and cell origin were the same in both models,
higher magnitudes of strain and fluid velocity within the scaffold might resulted in a delayed
fracture healing process in the 2.55 mm case (Figures 5.31-32). Another fracture model was
created with three cell origins (endosteum, surrounding muscle tissues and bone marrow) in the
4.95 mm distance from the proximal tibia. The predictions showed that the healing was
accelerated in the model located in bone marrow cavity with higher cell concentration and lower
mechanical stimuli through the healing process (Figure 5.35). There were no histological slides
available to compare to computational results.
The osteoporotic bone, with a lower bone stiffness surrounding the fracture, plus a higher
permeability and void ratio, had delayed healing compared to the normal tibia. The rate of
diffusivity was kept constant, in both normal and osteoporotic cases, to solely investigate the

206
effect of mechanical properties on tissue differentiation. The FE model of the closed burr-hole
fracture was created based on a in vivo study (Taiani et al. 2010). The stiffness of the scaffolds
were calculated during the healing process using the computational simulations. The scaffold
stiffness gradually increased over time in both normal and osteoporotic murine tibia (Figure
5.46); however, the overall stiffness of the fracture site within the normal tibia was higher
(Figures 5.39, 5.47). The computational predictions showed that the normal tibia had promoted
fracture healing compared to the osteoporotic case. This was in agreement with experimental
observations that the osteoporotic bones are difficult to heal (Taiani 2012, He et al. 2011). The
CT images from Taiani et al. (2012) and the computational predictions both indicate that the
osteoporotic tibia had less mineralisation compared to the normal case (Figures 5.39, 5.48). He et
al. (2011) created burr-hole fracture models in mice femur to analyse the healing process in
normal and osteoporotic bones, and the CT images from the osteoporotic bone showed a
delayed healing.
Figure 5.46. The gradual change of tissue type during the healing period within a normal
and an osteoporotic murine tibia.

207
Figure 5.47. The overall stiffness of the scaffold during the healing process for normal and
osteoporotic bone.
Normal
Osteoporotic (Taiani, 2012)
Day 0 Day 7 Day 14 Day 28
1 mm
Figure 5.48. The healing progression in a normal bone and an osteoporotic bone from CT
images show that the normal bone had more mineralisation (Taiani 2012).
Zhang et al. (2007) showed that bending load accelerated healing in burr-hole fracture models of
murine tibiae (Figure 5.49). The left knees were subjected to a 0.02 Nm bending for three
minutes per day for three consecutive days, whereas the surgical holes in the right tibiae were

208
used as control (no motion). According to the CT images of Zhang et al. (2007), the mineral
density was increased, and the healing process was accelerated in the loaded tibiae (Figure
5.49b). Moreover, the mechanical testing revealed that the stiffness of the left tibiae increased by
mechanical loading. Since Zhang et al. (2007) found that the bending of 0.02 Nm was an
appropriate magnitude to stimulate the healing process, the same bending load was used in the
present study. Moreover, a higher load of 0.04 Nm (1 Hz) was also applied to the burr-hole
fracture model of a murine tibia to explore the effect of load magnitudes on tissue differentiation.
In both models, the stiffness of the scaffolds increased during healing which led to a decrease in
the magnitudes of mechanical stimuli. However, more bony tissue was formed under the lower
bending load (0.02 Nm, 1 Hz) which resulted a stiffer bridging at the fracture site. The results
once more highlight the importance of mechanical load magnitudes on the healing rate and
patterns of differentiated tissues. The mechanical loading, fracture position and the rate of the
cell diffusion showed to have a significant effect on the mechanical environment at the fracture
site and the healing process. Using a well-defined closed murine fracture model (e.g. Taiani et al.
2010) may help to further investigate the effect of different factors on fracture repair.
(a)
(b)
Day 7 Day 14 Day 21
Figure 5.49. CT images showing the healing process within the burr-hole fracture model
of the (a) unloaded murine tibia, (b) tibia under bending load of 0.02 Nm (Zhang et al.
2007).

209
Chapter Six: Conclusions, limitations and future directions
Computational models of mechanoregulatory algorithms have been shown to be helpful for
research in implant design and fracture repair. A variety of biological, physiological and
mechanical factors have a major influence on the bone healing process. Furthermore, the
mechanical properties and the geometry of the bone can change over time in health and disease
as well as vary in different bones. This complex behaviour makes it challenging to achieve a
comprehensive understanding of bone mechanobiology. Computational models of
mechanoregulatory algorithms can complement experimental investigations to provide a broader
understanding of underlying mechanical and biological factors involved in fracture repair. The
effect of mechanical stimuli (e.g. fluid velocity and fluid pressurization, and the tissue stress and
strain) and their effect on fracture healing can be explored temporally and spatially using
numerical models.
6.1 Summary and conclusions
A biphasic mechanoregulatory algorithm based on mechanical stimuli (octahedral shear strain
and fluid velocity) was used in the current work to model the fracture healing in a murine tibia
(Lacroix et al. 2002, Prendergast et al. 1997). The mechanical stimuli applied to the stem cells
over time resulted in a gradual change of the tissue material properties within the fracture site.
The cells could differentiate into fibrous tissue, immature and mature cartilage, and immature
and mature bone. To simulate the migration and proliferation of the cells through the regenerated
tissue, a diffusion process coupled to the poroelastic stress analysis was developed. Generally,
murine fractures heal by 21 days in a healthy tibia (Gardner et al. 2006). Therefore, the diffusion
coefficients were set such that after 3 weeks, the progenitor cells could spread throughout the
entire scaffold. A user defined subroutine, USDFLD, was written in Fortran and then linked with

210
ABAQUS v6.11 to update the material properties (the Young‟s modulus, oisson‟s ratio,
permeability and void ratio), depending on the mechanical stimuli and cell concentration
(Chapter 3).
To evaluate the mechanoregulatory algorithm, a 2D and a 3D idealised murine fracture models
were created based on previous experimental studies (Gardner et al. 2006, Bishop et al. 2006).
The boundary conditions and loading conditions were matched with the in vivo studies and the
healing process was predicted (Chapter 3).
1. A murine tibia was subjected to different values of axial compression loads (2, 1, 0.5 N, 1
Hz) (Gardner et al. 2006):
The predicted sequence of tissue regeneration in the murine fracture model occurred in
the same pattern observed in vivo. The magnitude of axial compressive load had effect on
the healing rate and patterns. In agreement with the CT images from Gardner et al.
(2006), the computational simulations predicted bone repair enhancement under low
magnitudes of load.
2. A murine tibia was subjected to axial torsion (8 degrees, 1 Hz) and a combined loading of
axial torsion and axial compressive load (8 degrees,0.4 N, 1 Hz), with a maximum
octahedral shear strain magnitude of 25 % (Bishop et al. 2006):
The computational results were able to predict the bridging of the fracture gap and tissue
distribution was similar to the histological slide from the in vivo study (Bishop et al.
2006).
It was concluded that the axial torsional load led to promoted healing compared to the
combined loading (axial compression and torsion). Although it was reported that axial
torsion delayed the healing process and caused non-unions (Augat et al. 2003, Aro et al.

211
1991, Yamagishi and Yoshimura 1955), it did not inhibit the healing process (Bishop et
al. 2006).
From both studies, it was concluded that the mechanoregulatory algorithm was reliable and could
predict realistic healing patterns during healing process compared to the experimental studies.
In Chapter 4, a computational model was developed to characterize the mechanical environment
within the modified Flexcell system. A 1D FE model of the modified Flexcell system was
created to load very soft gels (confined compression). Initially, the load-deflection FE was
validated against the preliminary experimental results from our group (Olesja Hazenbiller,
University of Calgary, M.Sc. student). The bottom of the system was subjected to 20, 10 and 5
kPa (1Hz) compressive stresses. The model predictions suggested that the elements at the
superficial layers of the collagenous scaffold experienced higher peak strains and fluid velocities
whereas the mechanical stimuli were lower in the deeper layers (middle and bottom, closer to the
loading). The biphasic algorithm was implemented into the computational model and the tissue
differentiation under confined compression was investigated. The mechanical stimuli provided
an appropriate mechanical environment for cell differentiation. Both experimental and
computational results suggest that mechanical perturbation of the gel may be an effective way to
initiate tissue differentiation pathways prior to implantation for tissue engineering applications.
The aim of chapter 5 was to generate the intact and damaged murine tibia FE models and
implement the biphasic model to predict the development of differentiated tissue differentiation
within the closed fracture model (treated with a stem cell seeded soft collagenous scaffold) under
load regimes. The µCT based FE model of a burr-hole murine tibia was reconstructed using
Simpleware software. The mechanical behaviour of the reconstructed model was verified against
a previously published numerical-experimental study (Stadelmann et al. 2009). The biphasic

212
algorithm was then implemented into the fracture model to predict the development of
differentiated tissues for a variety of case studies:
Axial compression. The proximal end of the tibia was subjected to different magnitudes of axial
compression load (2, 1, 0.5 N, 1 Hz). In agreement with the 2D idealized murine model, the
model predicted enhanced healing under 0.5 N (1Hz) loading condition. This indicated that the
magnitude of the applied load might have a significant effect on bone healing (Gardner et al.
2006).
Rate of cell diffusion. The diffusion coefficient may decrease with aging and delay the healing
process (Chen et al. 2005, Park et al. 2005). Two diffusive rates (0.025 and 0.01 s
mm 2
) were
selected to represent a normal and slow diffusion. Since murine tibia fractures generally heal by
21 days (Gardner et al. 2006), the normal diffusion rate was set such that the cells spread
throughout the scaffold in 21 days (steps). The lower diffusive rate represented an elder murine,
in which the cells were partially spread (after 21 days). The computational results suggest that
the rate of cell diffusion plays an important role on the healing process. In agreement with the
literature (Li et al. 2005), the higher diffusive rates led to accelerated healing in the
computational study.
Fracture position. The fracture located further from the proximal tibia had promoted healing
compared to the fracture model with a hole closer to the proximal end of the tibia. The higher
mechanical stimuli delayed the healing process (Lacroix et al. 2002, Isaksson et al. 2006).
Therefore, the magnitudes of mechanical stimuli are key factors to influence the differentiation
pathways.

213
Reduced mechanical properties. The effect of reduced mechanical properties was investigated
using an osteoporotic bone. The bone healing was delayed in the osteoporotic tibia compared to
the healthy bone. The mechanical stimuli had larger magnitudes in the osteoporotic bone due to
higher void ratio and permea ility (higher fluid velo ity) and lower Young‟s modulus (higher
octahedral shear strain). This was in agreement with in vivo observations that the osteoporotic
bones are difficult to heal (Li and Aspden 1997b, Roschger et al. 2001, McDonnell et al. 2007,
Taiani 2012).
Bending load. Bending load (e.g. 0.02 Nm) was reported to enhance bone healing process in a
murine tibia (Zhang et al. 2006). The tibia was subjected to bending loads (0.02, 0.04 Nm) and
the healing process was predicted. The stiffer bridging was observed when the bone was
subjected to a lower bending load (0.02 Nm). This highlights the influence of load magnitude on
tissue differentiation and bone repair process.
6.2 Limitations
In developing computational mechanoregulatory models there are a number of simplifying
assumptions which are here discussed.
Bone geometry. The axisymmetric or 3D idealised models do not fully represent the exact
geometry and the loading on the bone. For example, due to the natural curvature of the tibia, the
axial compression load applied to the cortical shaft induces combined compression and bending
strains. Therefore, a 3D CT based FE model was reconstructed in the current study to ensure
more accurate distributions of the tissue strain and stress. Although all tissues were modelled as
poroelastic materials, in order to reduce the computational cost only ~1/3 of the tibia length was
used. A comparison of the mechanical environment in the full-length model and the 1/3 decay
length model showed that the distributions were very similar within 0.5 %.

214
Mechanical properties of the tissues. The mechanical properties for the various tissues were
taken from a variety of literature sources. Parametric studies have also been used to determine
appropriate parameters in the published mechanoregulatory algorithms (Lacroix et al. 2002,
Isaksson et al. 2006). However, further research is needed to confirm the material properties in
the current application.
The tissues were described as poroelastic materials in this study and their properties were taken
from published literature (Isaksson et al. 2006, Sandino and Lacroix 2011). The material
properties of the tissues were updated in the user-define subroutine USDFLD based on a linear
interpolation between the tabular data of the mechanical properties. Moreover, whenever the
obtained field variable was outside of the range specified (last: mature bone or first: granulation
tissue), the last or first available material data (last: mature bone or first: granulation tissue) was
used and there was no extrapolation. However, the obtained values based on the linear
assumption might not represent the real values of the mechanical properties (e.g. the change in
the material properties might be nonlinear).
The cortical and trabecular tissues were considered homogenous for the idealised models
(Chapter 3). To make the model more realistic in the CT based FE model of the murine tibia,
grey-scale based material properties (nonhomogeneous Young‟s modulus) related to density
distribution were chosen for the cortical and trabecular bone (Chapter 5). However, the material
properties were only considered to be isotropic. Anisotropic material properties would help to
distinguish lamellar bone from woven bone in the final stages of healing process.
Boundary conditions and mechanical loading. Only constant loads were applied to the cortical
bone during the analysis. The loads used in the current study (0.5-10 N) were selected based on
the previous in vivo studies. Applied loads to the murine tibiae were between 0.5 N and

215
maximum 13 N in vivo studies (Stadelmann et al. 2009, Zhang and Yokota 2011, Gardner et al.
2006, De Souza et al. 2005). The peak force acting on the tibia during normal walking was
determined to be 1.2 N (Prasad et al. 2010). In another study, the peak strains at the tibial
midshaft were reported for normal walking (200-300 µƐ) and jumping (400-600 µƐ), (De Souza
et al. 2005). According to the study by Stadelmann et al. (2009), the loads between 1.2-2.1 N
(normal walking) and 2.8-4.2 N (jumping) produced strains of 200-300 µƐ and 400-600 µƐ in the
murine tibial crest. The axial compression load in the range of 0-10 N used to verify the
mechanical behaviour of the reconstructed murine model against the numerical-experimental
study (Stadelmann et al. 2009). In our parametric studies (Chapter 5), the axial compressive
loads in the range of 0.5-2.5 N was used to simulate slow walk (0.5 and 1 N), normal walk (2 N)
and running (2.5 N). However, the boundary conditions and loading conditions used in the FE
models likely do not reflect in vivo conditions and further studies is needed to obtain more
realistic data.
Biological processes. The biological processes were greatly simplified in the computational
mechanoregulatory models. The cell migration and proliferation were modelled using a diffusion
equation in the present study. The model simulated cell migration as well as cell proliferation
independent from the mechanical stimuli. However, cell migration and proliferation might be
coupled to the mechanical perturbations. In the study by Perez and Prendergast (2007), cell
migration and proliferation were modelled with two methods (diffusion equation and random
walk) and the healing pattern was predicted and compared. Both models had similar predictions
for the temporal change of stiffness in the tissue differentiated within the gap. The only
differen e was that the “random walk” ased model showed greater variation in the patterns of
the differentiated tissues compared to the model proposed by Lacroix et al. (2002). The diffusion

216
process predicted continuous patterns of tissue differentiation, whereas the random walk model
had more heterogeneous tissue pattern. However, there was no histological slide to validate the
computational slides and it is not really known which one could predict more realistic patterns
(Perez and Prendergast 2007). According to the fact that both models had predicted similar
patterns of tissue differentiation, the diffusion equation was used in this study. However, the
diffusion equation always predicts the same tissue pathways, whereas the predictions of the
“random walk” ased model differ in ea h analysis ( loser to what happens in vivo). The
migrations of the cells were in a preferred direction based on a diffusion gradient. Stochastic
models without a preferred migration, such as random walk, may be more appropriate (Perez and
Prendergast 2007). Angiogenesis is known to play a key role in fracture healing (Sandino and
Lacroix 2011), but it was neglected in this study. In our simulations the callus size was kept
constant during the analysis and tissue volumetric growth was not explicitly modelled (Garcia-
Aznar et al. 2007). However, during the healing process, the size of callus might change
depending on the direction and magnitude of the load that was applied to the fracture site. In case
of having very low or extreme mechanical stimuli, cell apoptosis or cell death might happen,
(excluded from our simulations).
Single phase models cannot predict a realistic healing process, as the influence of the interstitial
fluid is neglected (Isaksson et al. 2006). However, the mechanoregulatory algorithm used in our
studies is based on the interstitial fluid velocity and the octahedral shear strain. Cell migration
and proliferation were modelled using a mass diffusion analysis coupled with the poroelastic
analysis. The implemented algorithm has been shown to predict the key events during fracture
healing (Perez and Prendergast 2007, Isaksson et al. 2006). We also qualitatively validated the
model against previous in vivo studies and the predicted tissue distributions were similar to the

217
available histological slides and CT images (Gardner et al. 2006, Bishop et al. 2006). The
differentiation algorithms will be further developed and validated using gene expression patterns
and high resolution CT images of mineralization patterns in well-defined fracture models.
6.3 Future directions
Our future research will focus on further development of the mechanoregulation algorithms and
their validation with in vivo and ex vivo models of fracture healing within the closed murine
fracture model. The developments can be broken down into two main aspects: (1) mechanical
behaviour and (2) cellular and biological system.
Mechanical behaviour of the tissue. Incorporating the nonhomogeneous, non-linear material
properties for soft tissues would improve the accuracy of the predictions of the mechanical (e.g.
fluid velocity, and tissue stress and strain). The callus size was kept constant during this study;
however, incorporating a tissue volumetric growth model may help achieve a more realistic
repair model. The mechanical stimuli were only explored at a macroscopic level (trabecular and
cortical bone were defined as continuum materials). Multi-scale models can be created using the
high-resolution CT images to investigate the actual stimuli acting at the cellular level and a
micro-structural level. Since the mechanical properties of the osteoporotic bone do not change
uniformly, using CT images of an osteoporotic murine tibia might increase the accuracy of the
model.
Cellular and biological system. The biological factors driving stem cell differentiation can be
further developed using a stochastic representation for cell migration, or considering the effect of
vascularisation. Moreover, cellular synthesis of bone nodules can be randomly added to the
fracture model based on our experimental research with collagen gels and stem cells. The
process of cell absorption and cellular death or apoptosis should also be considered in our future

218
studies. Furthermore, the effectiveness of pharmacological treatments and growth factors for
osteoporotic fractures and mechanisms involved will be explored.
The prediction of stem cell differentiation can be correlated with tissue culture investigations,
high-resolution CT images of mineralization patterns and gene expression analysis to develop a
more quantitative understanding of the role of mechanical factors in tissue formation. This
knowledge is essential to optimise the design of scaffold systems to best transfer mechanical
signals between cells and the extracellular matrix within the bone-healing environment and better
promote the healing process.

219
References
ABAQUS Theory Manual, Version 6.11-1, Dassault Systemes Simulia Corp., Providence, RI.
Ahlborg, H. G., O. Johnell, C. H. Turner, G. Rannevik & M. K. Karlsson (2003) Bone loss and
bone size after menopause. N Engl J Med, 349, 327-34.
Allen, M. R., J. M. Hock & D. B. Burr. 2004. Periosteum: biology, regulation, and response to
osteoporosis therapies. In Bone, 1003-12. United States.
Apostolopoulos, C. A. & D. D. Deligianni (2009) Prediction of local cellular deformation in
bone--influence of microstructure dimensions. J Musculoskelet Neuronal Interact, 9, 99-
108.
Aro, H. T., H. T. Wahner & E. Y. Chao (1991) Healing patterns of transverse and oblique
osteotomies in the canine tibia under external fixation. J Orthop Trauma, 5, 351-64.
Ashman, R. B. & J. Y. Rho (1988) Elastic modulus of trabecular bone material. J Biomech, 21,
177-81.
Ateshian, G. A., W. M. Lai, W. B. Zhu & V. C. Mow (1994) An asymptotic solution for the
contact of two biphasic cartilage layers. J Biomech, 27, 1347-60.
Aubin, J. E. 1999. Osteoprogenitor cell frequency in rat bone marrow stromal populations: role
for heterotypic cell-cell interactions in osteoblast differentiation. In J Cell Biochem, 396-
410. United States.
Augat, P., J. Burger, S. Schorlemmer, T. Henke, M. Peraus & L. Claes. 2003. Shear movement at
the fracture site delays healing in a diaphyseal fracture model. In J Orthop Res, 1011-7.
United States.
Augat, P., K. Margevicius, J. Simon, S. Wolf, G. Suger & L. Claes (1998) Local tissue properties
in bone healing: influence of size and stability of the osteotomy gap. Journal of
orthopaedic research : official publication of the Orthopaedic Research Society, 16, 475-
481.
Augat, P., R. Penzkofer, A. Nolte, M. Maier, S. Panzer, G. v Oldenburg, K. Pueschl, U. Simon &
V. Buhren. 2008. Interfragmentary movement in diaphyseal tibia fractures fixed with
locked intramedullary nails. In J Orthop Trauma, 30-6. United States.
Bacabac, R. G., D. Mizuno, C. F. Schmidt, F. C. MacKintosh, J. J. Van Loon, J. Klein-Nulend &
T. H. Smit. 2008. Round versus flat: bone cell morphology, elasticity, and
mechanosensing. In J Biomech, 1590-8. United States.
Bailon-Plaza, A. & M. C. van der Meulen (2001) A mathematical framework to study the effects
of growth factor influences on fracture healing. J Theor Biol, 212, 191-209.
Bailón-Plaza, A. & M. C. H. van der Meulen (2003) Beneficial effects of moderate, early loading
and adverse effects of delayed or excessive loading on bone healing. Journal of
biomechanics, 36, 1069-1077.
Ball, M. D., I. C. Bonzani, M. J. Bovis, A. Williams & M. M. Stevens (2011) Human periosteum
is a source of cells for orthopaedic tissue engineering: a pilot study. Clin Orthop Relat
Res, 469, 3085-93.
Battaglia, T. C., A. C. Tsou, E. A. Taylor & B. Mikic (2003) Ash content modulation of
torsionally derived effective material properties in cortical mouse bone. J Biomech Eng,
125, 615-9.
Bielby, R., E. Jones & D. McGonagle. 2007. The role of mesenchymal stem cells in maintenance
and repair of bone. In Injury, S26-32. Netherlands.

220
Biot, M. A. (1941) General Theory of Three‐Dimensional Consolidation. J Appl Phys, 12, 155-
64.
Bishop, N. E., M. van Rhijn, I. Tami, R. Corveleijn, E. Schneider & K. Ito. 2006. Shear does not
necessarily inhibit bone healing. In Clin Orthop Relat Res, 307-14. United States.
Blenman, P. R., D. R. Carter & G. S. Beaupre (1989) Role of mechanical loading in the
progressive ossification of a fracture callus. J Orthop Res, 7, 398-407.
Boccaccio, A., D. J. Kelly & C. Pappalettere (2011) A mechano-regulation model of fracture
repair in vertebral bodies. J Orthop Res, 29, 433-43.
Boskey, A. L. & R. Coleman (2010) Aging and bone. Journal of dental research, 89, 1333-1348.
Boskey, A. L., T. M. Wright & R. D. Blank (1999) Collagen and bone strength. Journal of bone
and mineral research : the official journal of the American Society for Bone and Mineral
Research, 14, 330-335.
Bouxsein, M. L. (2006) Biomechanics of osteoporotic fractures. Clinical Reviews in Bone and
Mineral Metabolism, 4, 143-153.
Buckwalter, J. A., M. J. Glimcher, R. R. Cooper & R. Recker (1996) Bone biology. II:
Formation, form, modeling, remodeling, and regulation of cell function. Instr Course
Lect, 45, 387-99.
Burr, D. B. 2002. The contribution of the organic matrix to bone's material properties. In Bone,
8-11. United States.
Burr, D. B., N. A. Sharkey & R. B. Martin. 1998. Skeletal tissue mechanics. New York:
Springer.
Burstein, A. H., D. T. Reilly & M. Martens (1976) Aging of bone tissue: mechanical properties.
J Bone Joint Surg Am, 58, 82-6.
Byrne, D. P., D. Lacroix, J. A. Planell, D. J. Kelly & P. J. Prendergast. 2007. Simulation of tissue
differentiation in a scaffold as a function of porosity, Young's modulus and dissolution
rate: application of mechanobiological models in tissue engineering. In Biomaterials,
5544-54. England.
Carter, D. R., G. S. Beaupre, N. J. Giori & J. A. Helms (1998) Mechanobiology of skeletal
regeneration. Clin Orthop Relat Res, S41-55.
Carter, D. . G. S. eaupr . 2001. Skeletal function and form: mechanobiology of skeletal
development, aging, and regeneration. Cambridge: Cambridge University Press.
Carter, D. R., P. R. Blenman & G. S. Beaupre (1988) Correlations between mechanical stress
history and tissue differentiation in initial fracture healing. J Orthop Res, 6, 736-48.
Carter, D. R. & M. Wong (1988) The role of mechanical loading histories in the development of
diarthrodial joints. J Orthop Res, 6, 804-16.
Carter, D. R., M. Wong & T. E. Orr (1991) Musculoskeletal ontogeny, phylogeny, and
functional adaptation. J Biomech, 24 Suppl 1, 3-16.
Checa, S. & P. J. Prendergast (2009) A mechanobiological model for tissue differentiation that
includes angiogenesis: a lattice-based modeling approach. Ann Biomed Eng, 37, 129-45.
Checa, S. & P. J. Prendergast 2010. Effect of cell seeding and mechanical loading on
vascularization and tissue formation inside a scaffold: a mechano-biological model using
a lattice approach to simulate cell activity. In J Biomech, 961-8. United States: 2009
Elsevier Ltd.

221
Chen, J., S. Sotome, J. Wang, H. Orii, T. Uemura & K. Shinomiya (2005) Correlation of in vivo
bone formation capability and in vitro differentiation of human bone marrow stromal
cells. J Med Dent Sci, 52, 27-34.
Chen, Y.-J., H.-T. Chen & H.-C. Hsu (2007) P140. Transpedicular Bone Graft with
Instrumentation in Painful Osteoporotic Compression Fractures. The Spine Journal, 7,
147S-148S.
Cho, T. J., L. C. Gerstenfeld & T. A. Einhorn (2002) Differential temporal expression of
members of the transforming growth factor beta superfamily during murine fracture
healing. J Bone Miner Res, 17, 513-20.
Cioffi, M., F. Boschetti, M. T. Raimondi & G. Dubini (2006) Modeling evaluation of the fluid-
dynamic microenvironment in tissue-engineered constructs: a micro-CT based model.
Biotechnol Bioeng, 93, 500-10.
Claes, L., P. Augat, G. Suger & H. J. Wilke (1997) Influence of size and stability of the
osteotomy gap on the success of fracture healing. J Orthop Res, 15, 577-84.
Claes, L., K. Eckert-Hubner & P. Augat (2003) The fracture gap size influences the local
vascularization and tissue differentiation in callus healing. Langenbecks Arch Surg, 388,
316-22.
Claes, L. E. & C. A. Heigele (1999) Magnitudes of local stress and strain along bony surfaces
predict the course and type of fracture healing. J Biomech, 32, 255-66.
Claes, L. E., C. A. Heigele, C. Neidlinger-Wilke, D. Kaspar, W. Seidl, K. J. Margevicius & P.
Augat (1998) Effects of mechanical factors on the fracture healing process. Clin Orthop
Relat Res, S132-47.
Clarke, B. (2008) Normal bone anatomy and physiology. Clin J Am Soc Nephrol, 3 Suppl 3,
S131-9.
Cole, J. H., M. C. H. Meulen & R. A. Adler. 2010. Biomechanics of Bone Osteoporosis. ed. P.
M. Conn, 157-179. Humana Press.
Cole, J. H. & M. C. van der Meulen (2011) Whole bone mechanics and bone quality. Clin
Orthop Relat Res, 469, 2139-49.
Cowin, S. C. (2002) Mechanosensation and fluid transport in living bone. Journal of
musculoskeletal & neuronal interactions, 2, 256.
Crank, J. 1956. The mathematics of diffusion. Oxford: Clarendon Press.
Cullinane, D. M., A. Fredrick, S. R. Eisenberg, D. Pacicca, M. V. Elman, C. Lee, K. Salisbury,
L. C. Gerstenfeld & T. A. Einhorn (2002) Induction of a neoarthrosis by precisely
controlled motion in an experimental mid-femoral defect. J Orthop Res, 20, 579-86.
Cullinane, D. M., K. T. Salisbury, Y. Alkhiary, S. Eisenberg, L. Gerstenfeld & T. A. Einhorn
(2003) Effects of the local mechanical environment on vertebrate tissue differentiation
during repair: does repair recapitulate development? J Exp Biol, 206, 2459-71.
Currey, J. D. (2012) The structure and mechanics of bone. Journal of Materials Science, 47, 41-
54.
Cussler, E. L. 2009. Diffusion: mass transfer in fluid systems. Cambridge: Cambridge University
Press.
Darwish, S. M. (2004) Analysis of weld-bonded dissimilar materials. International Journal of
Adhesion and Adhesives, 24, 347-354.
Davison, K. S., K. Siminoski, J. D. Adachi, D. A. Hanley, D. Goltzman, A. B. Hodsman, R.
Josse, S. Kaiser, W. P. Olszynski, A. Papaioannou, L. G. Ste-Marie, D. L. Kendler, A.

222
Tenenhouse & J. P. Brown. 2006. Bone strength: the whole is greater than the sum of its
parts. In Semin Arthritis Rheum, 22-31. United States.
De Souza, R. L., M. Matsuura, F. Eckstein, S. C. Rawlinson, L. E. Lanyon & A. A. Pitsillides.
2005. Non-invasive axial loading of mouse tibiae increases cortical bone formation and
modifies trabecular organization: a new model to study cortical and cancellous
compartments in a single loaded element. In Bone, 810-8. United States.
Deligianni, D. D. & C. A. Apostolopoulos (2008) Multilevel finite element modeling for the
prediction of local cellular deformation in bone. Biomechanics and modeling in
mechanobiology, 7, 151-159.
DeWolf, J. T., F. P. Beer & E. R. Johnston. 2006. Mechanics of materials. Boston: McGraw-Hill
Higher Education.
Dickenson, R. P., W. C. Hutton & J. R. Stott (1981) The mechanical properties of bone in
osteoporosis. The Journal of bone and joint surgery. British volume, 63-B, 233.
Doblare, M. & J. M. Garcia (2003) On the modelling bone tissue fracture and healing of the bone
tissue. Acta Cient Venez, 54, 58-75.
Duyck, J., K. Vandamme, L. Geris, H. Van Oosterwyck, M. De Cooman, J. Vandersloten, R.
Puers & I. Naert. 2006. The influence of micro-motion on the tissue differentiation
around immediately loaded cylindrical turned titanium implants. In Arch Oral Biol, 1-9.
England.
Egermann, M., J. Goldhahn & E. Schneider (2005) Animal models for fracture treatment in
osteoporosis. Osteoporos Int, 16 Suppl 2, S129-38.
Einhorn, T. A. (1998) One of nature's best kept secrets. J Bone Miner Res, 13, 10-2.
Einhorn, T. A. (2005) The science of fracture healing. J Orthop Trauma, 19, S4-6.
Epari, D. R., G. N. Duda & M. S. Thompson (2010) Mechanobiology of bone healing and
regeneration: in vivo models. Proc Inst Mech Eng H, 224, 1543-53.
Epari, D. R., H. Schell, H. J. Bail & G. N. Duda. 2006. Instability prolongs the chondral phase
during bone healing in sheep. In Bone, 864-70. United States.
Fratzl-Zelman, N., K. Koller & K. Klaushofer (1992) Mineral crystals in al ified tissues: a
comparative study by SAXS. Journal of Bone and Mineral Research, 7(3), 329–34.
Federico, S., G. La Rosa, W. Herzog & J. Z. Wu (2004) Effect of fluid boundary conditions on
joint contact mechanics and applications to the modeling of osteoarthritic joints. J
Biomech Eng, 126, 220-5.
Frost, H. M. (1989) The biology of fracture healing. An overview for clinicians. Part I. Clin
Orthop Relat Res, 283-93.
Frost, H. M. (2003) Bone's mechanostat: A 2003 update. Anat Rec A Discov Mol Cell Evol Biol,
275(2), 1081-101.
Galli, C., G. Passeri & G. M. Macaluso. 2010. Osteocytes and WNT: the mechanical control of
bone formation. In J Dent Res, 331-43. United States.
Garcia-Aznar, J. M., J. H. Kuiper, M. J. Gomez-Benito, M. Doblare & J. B. Richardson. 2007.
Computational simulation of fracture healing: influence of interfragmentary movement
on the callus growth. In J Biomech, 1467-76. United States.
Gardner, M. J., M. C. van der Meulen, D. Demetrakopoulos, T. M. Wright, E. R. Myers & M. P.
Bostrom (2006) In vivo cyclic axial compression affects bone healing in the mouse tibia.
J Orthop Res, 24, 1679-86.

223
Gardner, T. N., T. Stoll, L. Marks, S. Mishra & M. Knothe Tate. 2000. The influence of
mechanical stimulus on the pattern of tissue differentiation in a long bone fracture--an
FEM study. In J Biomech, 415-25. United States.
Geris, L., A. Andreykiv, H. Van Oosterwyck, J. V. Sloten, F. van Keulen, J. Duyck & I. Naert
(2004) Numerical simulation of tissue differentiation around loaded titanium implants in
a bone chamber. J Biomech, 37, 763-9.
Geris, L., A. Gerisch, C. Maes, G. Carmeliet, R. Weiner, J. Vander Sloten & H. Van Oosterwyck
(2006) Mathematical modeling of fracture healing in mice: comparison between
experimental data and numerical simulation results. Med Biol Eng Comput, 44, 280-9.
Geris, L., A. Gerisch, J. V. Sloten, R. Weiner & H. V. Oosterwyck. 2008. Angiogenesis in bone
fracture healing: a bioregulatory model. In J Theor Biol, 137-58. Netherlands.
Geris, L., K. Vandamme, I. Naert, J. Vander Sloten, J. Duyck & H. Van Oosterwyck. 2009.
Numerical simulation of bone regeneration in a bone chamber. In J Dent Res, 158-63.
United States.
Gerstenfeld, L. C., D. M. Cullinane, G. L. Barnes, D. T. Graves & T. A. Einhorn (2003) Fracture
healing as a post-natal developmental process: molecular, spatial, and temporal aspects of
its regulation. J Cell Biochem, 88, 873-84.
Giotakis, N. & B. Narayan (2007) Stability with unilateral external fixation in the tibia.
Strategies Trauma Limb Reconstr, 2, 13-20.
Goodship, A. E., J. L. Cunningham & J. Kenwright (1998) Strain rate and timing of stimulation
in mechanical modulation of fracture healing. Clin Orthop Relat Res, S105-15.
Goodship, A. E. & J. Kenwright (1985) The influence of induced micromovement upon the
healing of experimental tibial fractures. J Bone Joint Surg Br, 67, 650-5.
Gullberg, B., O. Johnell & J. A. Kanis (1997) World-wide projections for hip fracture.
Osteoporos Int, 7, 407-13.
Hayward, L. N. & E. F. Morgan (2009) Assessment of a mechano-regulation theory of skeletal
tissue differentiation in an in vivo model of mechanically induced cartilage formation.
Biomech Model Mechanobiol.
He, Y, G. Zhang, X. Pan, Z. Liu, L. Zheng, C. Chan, K. Lee, Y. Cao, G. Li, L. Wei, L. Hung K.
Leung & L. Qin (2011) Impaired bone healing pattern in mice with ovariectomy-induced
osteoporosis: A drill-hole defect model. J Bone, 48, 1388-1400.
Heino, T. J. & T. A. Hentunen (2008) Differentiation of osteoblasts and osteocytes from
mesenchymal stem cells. Curr Stem Cell Res Ther, 3, 131-45.
Henrotin, Y. 2011. Muscle: a source of progenitor cells for bone fracture healing. In BMC Med,
136. England.
Herzog, W. & B. M. Nigg. 1999. Biomechanics of the musculo-skeletal system. Chichester, U.K:
Wiley.
Hou, J. S., M. H. Holmes, W. M. Lai & V. C. Mow (1989) Boundary conditions at the cartilage-
synovial fluid interface for joint lubrication and theoretical verifications. J Biomech Eng,
111, 78-87.
Isaksson, H., C. C. van Donkelaar, R. Huiskes & K. Ito. 2008. A mechano-regulatory bone-
healing model incorporating cell-phenotype specific activity. In J Theor Biol, 230-46.
Netherlands.

224
Isaksson, H., W. Wilson, C. C. van Donkelaar, R. Huiskes & K. Ito (2006) Comparison of
biophysical stimuli for mechano-regulation of tissue differentiation during fracture
healing. J Biomech, 39, 1507-16.
Iwaki, A., S. Jingushi, Y. Oda, T. Izumi, J. I. Shida, M. Tsuneyoshi & Y. Sugioka (1997)
Localization and quantification of proliferating cells during rat fracture repair: detection
of proliferating cell nuclear antigen by immunohistochemistry. J Bone Miner Res, 12, 96-
102.
Jacobs, A. M. & L. M. Oloff (1985) Podiatric metallurgy and the effects of implanted metals on
living tissues. Clin Podiatry, 2, 121-41.
Jepsen, K. J. & N. Andarawis-Puri (2012) The amount of periosteal apposition required to
maintain bone strength during aging depends on adult bone morphology and tissue-
modulus degradation rate. J Bone Miner Res.
Joyce, M. E., A. B. Roberts, M. B. Sporn & M. E. Bolander (1990) Transforming growth factor-
beta and the initiation of chondrogenesis and osteogenesis in the rat femur. J Cell Biol,
110, 2195-207.
Keaveny, T. M., E. F. Wachtel & D. L. Kopperdahl (1999) Mechanical behavior of human
trabecular bone after overloading. J Orthop Res, 17, 346-53.
Kelly, D. J. & P. J. Prendergast. 2005. Mechano-regulation of stem cell differentiation and tissue
regeneration in osteochondral defects. In J Biomech, 1413-22. United States.
Kim, W. D., W. S. Kim, C. S. Woo & H. J. Lee (2004) Some considerations on mechanical
testing methods of rubbery materials using nonlinear finite element analysis. Polymer
International, 53, 850-856.
Knopp, E., N. Troiano, M. Bouxsein, B. H. Sun, K. Lostritto, C. Gundberg, J. Dziura & K.
Insogna. 2005. The effect of aging on the skeletal response to intermittent treatment with
parathyroid hormone. In Endocrinology, 1983-90. United States.
Kuiper. JH, Richardson JB and Ashton BA (1996) Mechanical signals in early fracture caull. In:
………Vander Sloten J, Lowet G, Van Audekercke R, Van der Perre G, editors. Proc 10th
Europ
………Soc Biomech, 154.
Kuiper. J. H., B. A. Ashton & J. B. Richardson (2000a) Computer simulation of fracture callus
………formation and stiffness resorption. In Prendergast PJ, Lee TC, Carr AJ, editors. Proc 12th
………Europ Soc Biomech, 61.
Kuiper JH, Richardson JB and Ashton BA (2000b) Computer simulation to study the effect of
………fracture site movement on tissue formation and fracture stiffness resorption. In European
……….Congress on Computational Methods in Applied Sciences and Engineering ECCOMAS.
Kutz, M. 2003. Standard handbook of biomedical engineering and design. New York: McGraw-
Hill.
Kwong, F. N. & M. B. Harris. 2008. Recent developments in the biology of fracture repair. In J
Am Acad Orthop Surg, 619-25. United States.
Lacroix, D. 2000. Simulation of tissue differentiation during fracture healing. ProQuest, UMI
Dissertations Publishing.
Lacroix, D., P. J. Prendergast, G. Li & D. Marsh (2002) Biomechanical model to simulate tissue
differentiation and bone regeneration: application to fracture healing. Med Biol Eng
Comput, 40, 14-21.

225
Lai, W. M., V. C. Mow & V. Roth (1981) Effects of nonlinear strain-dependent permeability and
rate of compression on the stress behavior of articular cartilage. J Biomech Eng, 103, 61-
6.
Lakes, R. & S. Saha (1979) Cement line motion in bone. Science, 204, 501-3.
Lang, T. F. (2011) The bone-muscle relationship in men and women. J Osteoporos, 2011,
702735.
Lee, D. Y., T. J. Cho, J. A. Kim, H. R. Lee, W. J. Yoo, C. Y. Chung & I. H. Choi (2008)
Mobilization of endothelial progenitor cells in fracture healing and distraction
osteogenesis. Bone, 42, 932-41.
Li, B. & R. M. Aspden (1997a) Composition and mechanical properties of cancellous bone from
the femoral head of patients with osteoporosis or osteoarthritis. J Bone Miner Res, 12,
641-51.
Li, B. & R. M. Aspden (1997b) Mechanical and material properties of the subchondral bone
plate from the femoral head of patients with osteoarthritis or osteoporosis. Ann Rheum
Dis, 56, 247-54.
Li, C. Y., M. B. Schaffler, H. T. Wolde-Semait, C. J. Hernandez & K. J. Jepsen (2005) Genetic
background influences cortical bone response to ovariectomy. J Bone and mineral
research, 20, 2150-2158.
Lienau, J., H. Schell, G. N. Duda, P. Seebeck, S. Muchow & H. J. Bail. 2005. Initial
vascularization and tissue differentiation are influenced by fixation stability. In J Orthop
Res, 639-45. United States.
Lipson, S. F. & J. L. Katz (1984) The relationship between elastic properties and micro-structure
………of bovine cortical bone. J Biomechanics, 17, 231–40.
Liu, R., O. Birke, A. Morse, L. Peacock, K. Mikulec, D. G. Little & A. Schindeler (2011)
Myogenic progenitors contribute to open but not closed fracture repair. BMC
Musculoskelet Disord, 12, 288.
Loboa, E. G., G. S. Beaupre & D. R. Carter (2001) Mechanobiology of initial pseudarthrosis
formation with oblique fractures. J Orthop Res, 19, 1067-72.
Loboa, E. G., T. A. Wren, G. S. Beaupre & D. R. Carter (2003) Mechanobiology of soft skeletal
tissue differentiation--a computational approach of a fiber-reinforced poroelastic model
based on homogeneous and isotropic simplifications. Biomech Model Mechanobiol, 2,
83-96.
Lu, C., T. Miclau, D. Hu, E. Hansen, K. Tsui, C. Puttlitz & R. S. Marcucio (2005) Cellular basis
for age-related changes in fracture repair. J Orthop Res, 23, 1300-07.
Macdonald, H. M., K. K. Nishiyama, D. A. Hanley & S. K. Boyd (2011) Changes in trabecular
and cortical bone microarchitecture at peripheral sites associated with 18 months of
teriparatide therapy in postmenopausal women with osteoporosis. Osteoporos Int, 22,
357-62.
Mann, V., E. E. Hobson, B. Li, T. L. Stewart, S. F. Grant, S. P. Robins, R. M. Aspden & S. H.
Ralston (2001) A COL1A1 Sp1 binding site polymorphism predisposes to osteoporotic
fracture by affecting bone density and quality. J Clin Invest, 107, 899-907.
Marsell, R. & T. A. Einhorn. 2011. The biology of fracture healing. In Injury, 551-5.
Netherlands: 2011 Elsevier Ltd.
Matsushita, T. & T. Kurokawa (1998) Comparison of cyclic compression, cyclic distraction and
rigid fixation. Bone healing in rabbits. Acta Orthop Scand, 69, 95-8.

226
Mazurkiewicz, D. (2009) Problems of numerical simulation of stress and strain in the area of the
adhesive-bonded joint of a conveyor belt. ARCHIVES OF CIVIL AND MECHANICAL
ENGINEERING, 9, 75-91.
McCalden, R. W., J. A. McGeough, M. B. Barker & C. M. Court-Brown (1993) Age-related
changes in the tensile properties of cortical bone. The relative importance of changes in
porosity, mineralization, and microstructure. J Bone Joint Surg Am, 75, 1193-205.
McDonnell, P., P. E. McHugh & D. O'Mahoney (2007) Vertebral osteoporosis and trabecular
bone quality. Ann Biomed Eng, 35, 170-89.
McKibbin, B. (1978) The biology of fracture healing in long bones. J Bone Joint Surg Br, 60-B,
150-62.
McMahon, L. A., F. J. O'Brien & P. J. Prendergast (2008) Biomechanics and mechanobiology in
osteochondral tissues. Regen Med, 3, 743-59.
Mikos, A. G., G. Sarakinos, M. D. Lyman, D. E. Ingber, J. P. Vacanti & R. Langer (1993)
Prevascularization of porous biodegradable polymers. Biotechnol Bioeng, 42, 716-23.
Milan, J. L., J. A. Planell & D. Lacroix (2010) Simulation of bone tissue formation within a
porous scaffold under dynamic compression. Biomech Model Mechanobiol, 9, 583-96.
Mitchell, A. W. M., R. L. Drake, H. Gray & W. Vogl. 2005. Gray's anatomy for students.
Philadelphia: Elsevier/Churchill Livingstone.
Moreo, P., J. M. Garcia-Aznar & M. Doblare. 2009a. Bone ingrowth on the surface of
endosseous implants. Part 1: Mathematical model. In J Theor Biol, 1-12. England.
Moreo, P., J. M. Garcia-Aznar & M. Doblare. 2009b. Bone ingrowth on the surface of
endosseous implants. Part 2: Theoretical and numerical analysis. In J Theor Biol, 13-26.
England.
Morgan, E. F., K. T. Salisbury Palomares, R. E. Gleason, D. L. Bellin, K. B. Chien, G. U.
Unnikrishnan & P. L. Leong. 2010. Correlations between local strains and tissue
phenotypes in an experimental model of skeletal healing. In J Biomech, 2418-24. United
States: 2010 Elsevier Ltd.
Moroni, A., A. Hoang-Kim, V. Lio & S. Giannini (2006) Current augmentation fixation
techniques for the osteoporotic patient. Scand J Surg, 95, 103-9.
Mosekilde, L. & C. C. Danielsen (1987) Biomechanical competence of vertebral trabecular bone
in relation to ash density and age in normal individuals. Bone, 8, 79-85.
Mow, V. C., S. C. Kuei, W. M. Lai & C. G. Armstrong (1980) Biphasic creep and stress
relaxation of articular cartilage in compression? Theory and experiments. J Biomech Eng,
102, 73-84.
Nagel, T. & D. J. Kelly (2010) Mechano-regulation of mesenchymal stem cell differentiation and
collagen organisation during skeletal tissue repair. Biomech Model Mechanobiol, 9, 359-
72.
Niebur, G. L., M. J. Feldstein, J. C. Yuen, T. J. Chen, T. M. Keaveny (2000) High-resolution
finite element models with tissue strength asymmetry accurately predict failure of
trabecular bone. J Biomech Eng, 33(12), 1575-83.
Noble, B. (2003) Bone microdamage and apoptosis. J European Cells and Materials, 6, 46-56.
Palomares, K. T., R. E. Gleason, Z. D. Mason, D. M. Cullinane, T. A. Einhorn, L. C. Gerstenfeld
& E. F. Morgan (2009) Mechanical stimulation alters tissue differentiation and molecular
expression during bone healing. J Orthop Res, 27, 1123-32.

227
Park, J. S., H. Y. Kim, H. W. Kim, G. N. Chae, H. T. Oh, J. Y. Park, H. Shim, M. Seo, E. Y.
Shin, E. G. Kim, S. C. Park & S. J. Kwak. 2005. Increased caveolin-1, a cause for the
declined adipogenic potential of senescent human mesenchymal stem cells. In Mech
Ageing Dev, 551-9. Ireland.
Pauwels, F. (1960) Eine neue Theorie über den Einfluß mechanischer Reize auf die
Differenzierung der Stützgewebe. Zeitschrift für Anatomie und Entwicklungsgeschichte,
121, 478-515.
Pawaskar, S. S., J. Fisher & Z. Jin (2010) Robust and general method for determining surface
fluid flow boundary conditions in articular cartilage contact mechanics modeling. J
Biomech Eng, 132, 031001.
Perez, M. A. & P. J. Prendergast. 2007. Random-walk models of cell dispersal included in
mechanobiological simulations of tissue differentiation. In J Biomech, 2244-53. United
States.
Perren, S. M. (1979) Physical and biological aspects of fracture healing with special reference to
internal fixation. Clinical orthopaedics and related research, 175.
Postacchini, F., S. Gumina, D. Perugia & C. De Martino (1995) Early fracture callus in the
diaphysis of human long bones. Histologic and ultrastructural study. Clin Orthop Relat
Res, 218-28.
Prasad, J., B. P. Wiater, S. E. Nork, S. D. Bain & T. S. Gross. 2010. Characterizing gait induced
normal strains in a murine tibia cortical bone defect model. In J Biomech, 2765-70.
United States: 2010 Elsevier Ltd.
Prendergast, P. J., S. Checa & D. Lacroix. 2010. Computational Models of Tissue
Differentiation. 353-372. Dordrecht: Springer Netherlands.
Prendergast, P. J., R. Huiskes & K. Soballe (1997) ESB Research Award 1996. Biophysical
stimuli on cells during tissue differentiation at implant interfaces. J Biomech, 30, 539-48.
Prendergast, P. J., W. D. van Driel & J. H. Kuiper (1996) A comparison of finite element codes
for the solution of biphasic poroelastic problems. Proc Inst Mech Eng H, 210, 131-6.
Puleo, D. A., L. A. Holleran, R. H. Doremus & R. Bizios (1991) Osteoblast responses to
orthopedic implant materials in vitro. J Biomed Mater Res, 25, 711-23.
Rangaswami, H., N. Marathe, S. Zhuang, Y. Chen, J. C. Yeh, J. A. Frangos, G. R. Boss & R. B.
Pilz. 2009. Type II cGMP-dependent protein kinase mediates osteoblast
mechanotransduction. In J Biol Chem, 14796-808. United States.
Reginster, J.-Y. & N. Burlet (2006) Osteoporosis: a still increasing prevalence. Bone, 38, S4-9.
Reilly, D. T. & Burstein A. H. (1975) The elastic and ultimate properties of compact bone tissue.
J Biomechun, 8, 393-405.
Rho, J. Y., R. B. Ashman & C. H. Turner (1993) Young's modulus of trabecular and cortical
bone material: ultrasonic and microtensile measurements. J Biomech, 26, 111-9.
Rho, J. Y., M. C. Hobatho & R. B. Ashman (1995) Relations of mechanical properties to density
and CT numbers in human bone. Med Eng Phys, 17, 347-55.
Riggs, B. L., S. Khosla & L. J. Melton, 3rd (2002) Sex steroids and the construction and
conservation of the adult skeleton. Endocr Rev, 23, 279-302.
Rivadeneira, F., J. J. Houwing-Duistermaat, N. Vaessen, J. M. Vergeer-Drop, A. Hofman, H. A.
Pols, C. M. Van Duijn & A. G. Uitterlinden (2003) Association between an insulin-like
growth factor I gene promoter polymorphism and bone mineral density in the elderly: the
Rotterdam Study. J Clin Endocrinol Metab, 88, 3878-84.

228
Roschger, P., S. Rinnerthaler, J. Yates, G. A. Rodan, P. Fratzl & K. Klaushofer. 2001.
Alendronate increases degree and uniformity of mineralization in cancellous bone and
decreases the porosity in cortical bone of osteoporotic women. In Bone, 185-91. United
States.
Rubin, C. T. & K. J. McLeod (1994) Promotion of bony ingrowth by frequency-specific, low-
amplitude mechanical strain. Clin Orthop Relat Res, 165-74.
Sandino, C. & D. Lacroix (2011) A dynamical study of the mechanical stimuli and tissue
differentiation within a CaP scaffold based on micro-CT finite element models. Biomech
Model Mechanobiol, 10, 565-76.
Saxon, L. K., A. G. Robling, I. Alam & C. H. Turner. 2005. Mechanosensitivity of the rat
skeleton decreases after a long period of loading, but is improved with time off. In Bone,
454-64. United States.
Schulmerich, M. V., W. F. Finney, R. A. Fredricks & M. D. Morris (2006) Subsurface Raman
spectroscopy and mapping using a globally illuminated non-confocal fiber-optic array
probe in the presence of Raman photon migration. Appl Spectrosc, 60, 109-14.
Shapiro, F. 2008. Bone development and its relation to fracture repair. The role of mesenchymal
osteoblasts and surface osteoblasts. In Eur Cell Mater, 53-76. Scotland.
Sharir, A., M. M. Barak & R. Shahar. 2008. Whole bone mechanics and mechanical testing. In
Vet J, 8-17. England.
Shefelbine, S. J., P. Augat, L. Claes & U. Simon. 2005. Trabecular bone fracture healing
simulation with finite element analysis and fuzzy logic. In J Biomech, 2440-50. United
States.
Silva, M. J., M. D. Brodt & W. J. Hucker (2005) Finite element analysis of the mouse tibia:
estimating endocortical strain during three-point bending in SAMP6 osteoporotic mice.
Anat Rec A Discov Mol Cell Evol Biol, 283, 380-90.
Simon, . . (1992) Multiphasi poroelasti finite element models forsoft tissue stru tures. J
Applied Mechanics Reviews, 45, 191-218.
Simmons, D. J. (1985) Fracture healing perspectives. Clin Orthop Relat Res, 100-13.
Smith-Adaline, E. A., S. K. Volkman, M. A. Ignelzi, Jr., J. Slade, S. Platte & S. A. Goldstein.
2004. Mechanical environment alters tissue formation patterns during fracture repair. In J
Orthop Res, 1079-85. United States.
Stadelmann, V. A., J. Hocke, J. Verhelle, V. Forster, F. Merlini, A. Terrier & D. P. Pioletti
(2009) 3D strain map of axially loaded mouse tibia: a numerical analysis validated by
experimental measurements. Comput Methods Biomech Biomed Engin, 12, 95-100.
Sun, S. S., H. L. Ma, C. L. Liu, C. H. Huang, C. K. Cheng & H. W. Wei. 2008. Difference in
femoral head and neck material properties between osteoarthritis and osteoporosis. In
Clin Biomech (Bristol, Avon), S39-47. England.
Sverdlova, N. S. & U. Witzel (2010) Principles of determination and verification of muscle
forces in the human musculoskeletal system: Muscle forces to minimise bending stress. J
Biomech, 43, 387-96.
Taiani, J. T., R. J. Krawetz, N. I. Zur Nieden, Y. Elizabeth Wu, M. S. Kallos, J. R. Matyas & D.
E. Rancourt (2010) Reduced differentiation efficiency of murine embryonic stem cells in
stirred suspension bioreactors. Stem Cells Dev, 19, 989-98.
Taiani, J. T. 2012, 'Enhanced bone repair using embryonic stem cells in a mouse model of
impaired fracture healing', PhD thesis, University of Calgary.

229
Taylor, M. E., K. E. Tanner, M. A. Freeman & A. L. Yettram (1996) Stress and strain
distribution within the intact femur: compression or bending? Med Eng Phys, 18, 122-31.
Thomsen, J. S., E. N. Ebbesen & L. I. Mosekilde. 2002. Age-related differences between
thinning of horizontal and vertical trabeculae in human lumbar bone as assessed by a new
computerized method. In Bone, 136-42. United States.
Tingart, M. J., M. Apreleva, D. von Stechow, D. Zurakowski & J. J. Warner (2003) The cortical
thickness of the proximal humeral diaphysis predicts bone mineral density of the
proximal humerus. J Bone Joint Surg Br, 85, 611-7.
Tromas, C., J. C. Stinville, C. Templier & P. Villechaise (2012) Hardness and elastic modulus
gradients in plasma-nitrided 316L polycrystalline stainless steel investigated by
nanoindentation tomography. Acta Materialia, 60, 1965-1973.
Truesdell, C. 1966. The Elements of continuum mechanics.
Tubbs, N. (1981) Current concepts of internal fixation of fractures. Injury, 13, 179-179.
Turner, C. H. (2006) Bone strength: current concepts. Ann N Y Acad Sci, 1068, 429-46.
Turner, C. H. & F. M. Pavalko (1998) Mechanotransduction and functional response of the
skeleton to physical stress: the mechanisms and mechanics of bone adaptation. J Orthop
Sci, 3, 346-55.
van der Meulen, M. C. & R. Huiskes. 2002. Why mechanobiology? A survey article. In J
Biomech, 401-14. United States.
Van der Voet, A. F. 1992. Finite element modelling of load transfer through articular cartilage.
Ottawa: National Library of Canada.
Van Rietbergen, B., R. Huiskes, F. Eckstein & P. Ruegsegger (2003) Trabecular bone tissue
strains in the healthy and osteoporotic human femur. J Bone Miner Res, 18, 1781-8.
Van Rietbergen, B., R. Huiskes, H. Weinans, D. R. Sumner, T. M. Turner & J. O. Galante (1993)
ESB Research Award 1992. The mechanism of bone remodeling and resorption around
press-fitted THA stems. J Biomech, 26, 369-82.
Vandamme, K., I. Naert, L. Geris, J. Vander Sloten, R. Puers & J. Duyck. 2007. Influence of
controlled immediate loading and implant design on peri-implant bone formation. In J
Clin Periodontol, 172-81. Denmark.
Verborgt, O., G. J. Gibson & M. B. Schaffler (2000) Loss of osteocyte integrity in association
with microdamage and bone remodeling after fatigue in vivo. J Bone Miner Res, 15, 60-
7.
Wade, R. & J. Richardson (2001) Outcome in fracture healing: a review. Injury, 32, 109-114.
Wang, X., X. Shen, X. Li & C. M. Agrawal (2002) Age-related changes in the collagen network
and toughness of bone. Bone, 31, 1-7.
Wen, D. 2009. Cortical Bone Engineering: Scaffold Design And Cell Selection. Cleveland State
University.
Windahl, S. H., O. Vidal, G. Andersson, J. A. Gustafsson & C. Ohlsson (1999) Increased cortical
bone mineral content but unchanged trabecular bone mineral density in female ERbeta(-/-
) mice. J Clin Invest, 104, 895-901.
Wu, J. Z., W. Herzog & M. Epstein. 1998. Evaluation of the finite element software ABAQUS
for biomechanical modelling of biphasic tissues. In J Biomech, 165-9. United States.
Wylie, C. R. & L. C. Barrett. 1995. Advanced engineering mathematics. New York: McGraw-
Hill.

230
Xu, P. & J. E. Mark (1995) Strain-induced crystallization in elongated polyisobutylene
elastomers. Polymer Gels and Networks, 3, 255-266.
Yamagishi, M. & Y. Yoshimura (1955) The biomechanics of fracture healing. The Journal of
bone and joint surgery. American volume, 37-A, 1035.
Yaszemski, M. J., R. G. Payne, W. C. Hayes, R. Langer & A. G. Mikos (1996) Evolution of
bone transplantation: molecular, cellular and tissue strategies to engineer human bone.
Biomaterials, 17, 175-85.
Yoo, J. U. & B. Johnstone (1998) The role of osteochondral progenitor cells in fracture repair.
Clin Orthop Relat Res, S73-81.
Ysart, G. E. & R. M. Mason. 1994. Responses of articular cartilage explant cultures to different
oxygen tensions. In Biochim Biophys Acta, 15-20. Netherlands.
Zhang, P., S. M. Tanaka, H. Jiang, M. Su & H. Yokota. 2006. Diaphyseal bone formation in
murine tibiae in response to knee loading. In J Appl Physiol, 1452-9. United States.
Zhang, P., Q. Sun, C. H. Turner & H. Yokota (2007) Knee loading accelerates bone healing in
mice. J Bone Miner Res, 22, 1979-87.
Zhang, P. & H. Yokota. 2011. Knee loading stimulates healing of mouse bone wounds in a
femur neck. In Bone, 867-72. United States: 2011 Elsevier Inc.

231
Appendix A: User defined subroutine: USDFLD
A user-defined subroutine USDFLD was developed in FORTRAN to update the material
properties based on the average of computed mechanical stimuli in the previous 10 days, and on
the cell concentration.
SUBROUTINE USDFLD(FIELD,STATEV,PNEWDT,DIRECT,T,CELENT,
1 TIME,DTIME,CMNAME,ORNAME,NFIELD,NSTATV,NOEL,NPT,LAYER,
2 KSPT,KSTEP,KINC,NDI,NSHR,COORD,JMAC,JMATYP,MATLAYO,
3 LACCFLA)
C
INCLUDE 'ABA_PARAM.INC'
C
CHARACTER*80 CMNAME,ORNAME
CHARACTER*3 FLGRAY(15)
DIMENSION FIELD(NFIELD),STATEV(NSTATV),DIRECT(3,3),
1 T(3,3),TIME(2)
DIMENSION ARRAY(15),ARRAY1(15),JARRAY(15),JMAC(*),JMATYP(*)
1 ,COORD(*)
C
REAL SC_ST,EEP1,EEP2,EEP3,FLVEL1,FLVEL2,Oct_ShearStrain,
1 Oct_sum,Oct_1,Oct_2,Oct_3,VelMax,Vel_sum,FLVEL3,TEMP
SC_ST=STATEV(1)
CALL GETVRM('TEMP',ARRAY,JARRAY,FLGRAY,JRCD,
1 JMAC,JMATYP,MATLAYO,LACCFLA)
TEMP= ABS(ARRAY(1))
C
CALL GETVRM('EEP',ARRAY,JARRAY,FLGRAY,JRCD,
1 JMAC,JMATYP,MATLAYO,LACCFLA)
EEP1 = ARRAY(1)
EEP2 = ARRAY(2)
EEP3 = ARRAY(3)
C
CALL GETVRM('FLVEL',ARRAY1,JARRAY,FLGRAY,JRCD,
1 JMAC,JMATYP,MATLAYO,LACCFLA)
FLVEL1 = ARRAY1(1)
FLVEL2 = ARRAY1(2)
FLVEL3 = ARRAY1(3)
C
Oct_1 = ((EEP1-EEP2)*(EEP1-EEP2))
Oct_2 = ((EEP1-EEP3)*(EEP1-EEP3))
Oct_3 = ((EEP2-EEP3)*(EEP2-EEP3))
Oct_sum = (Oct_1)+(Oct_2)+(Oct_3)
Oct_ShearStrain=(2./3.)*SQRT(Oct_sum)
STATEV(3)= Oct_ShearStrain
C
Vel_sum=((FLVEL1)*(FLVEL1))+((FLVEL2)*(FLVEL2))
1 +((FLVEL3)*(FLVEL3))
VelMax= SQRT(Vel_sum)
STATEV(4)= VelMax

232
SC_ST = (STATEV(3)/0.0375)+((STATEV(4)*1000.)/(3.))
STATEV(1)=SC_ST
C
C INITIAL CONDITIONS FOR THE FIRST STEP
C
IF (KSTEP.EQ.1 .OR. KSTEP.EQ.2 ) THEN
FIELD(1) = 5
CMNAME='GRANULATION TISSUE'
IF (KSTEP.EQ.2 .AND. TIME(1).EQ.0) THEN
STATEV(29)=TEMP
IF ((STATEV(1).LT.6).AND.(STATEV(1).GE.3)) THEN
STATEV(2) = 4
CMNAME='FIBROUS TISSUE'
ELSE IF ((STATEV(1).LT.3) .AND. (STATEV(1).GE.1)) THEN
STATEV(2) = 3
CMNAME='CARTILAGE'
ELSE IF ((STATEV(1).LT.1.) .AND. (STATEV(1).GE.0.267)) THEN
STATEV(2) = 2
CMNAME='IMMATURE BONE'
ELSE IF ((STATEV(1).LT.0.267) .AND. (STATEV(1).GE.0.011)) THEN
STATEV(2) = 1
CMNAME='MATURE BONE'
ELSE
STATEV(2) = 5
CMNAME='NO TISSUE FORMATION'
ENDIF
END IF
END IF
C
IF (KSTEP.EQ.3 .OR. KSTEP.EQ.4 ) THEN
FIELD(1) = (STATEV(29)/500000.)*((STATEV(2)+(9.*5.))/(10.D0))
1 +((1-(STATEV(29)/500000.))*5.)
IF (KSTEP.EQ.4 .AND. TIME(1).EQ.0) THEN
STATEV(30)=TEMP
IF ((STATEV(1).LT.6).AND.(STATEV(1).GE.3)) THEN
STATEV(5) = 4
CMNAME='FIBROUS TISSUE'
ELSE IF ((STATEV(1).LT.3) .AND. (STATEV(1).GE.1)) THEN
STATEV(5) = 3
CMNAME='CARTILAGE'
ELSE IF ((STATEV(1).LT.1.) .AND. (STATEV(1).GE.0.267)) THEN
STATEV(5) = 2
CMNAME='IMMATURE BONE'
ELSE IF ((STATEV(1).LT.0.267) .AND. (STATEV(1).GE.0.011)) THEN
STATEV(5) = 1
CMNAME='MATURE BONE'
ELSE
STATEV(5) = 5
CMNAME='NO TISSUE FORMATION'
ENDIF
END IF
END IF
C
CCC...

233
C
IF (KSTEP.EQ.99 .OR. KSTEP.EQ.100) THEN
FIELD(1)=(STATEV(104)/500000.)*
1 ((STATEV(77)+STATEV(76)+STATEV(75)+STATEV(74)+
2 STATEV(73)+STATEV(72)+STATEV(71)+
3 STATEV(70)+STATEV(69)+(STATEV(68)))/(10.D0))
4 +((1-(STATEV(104)/500000.))*5.)
IF (KSTEP.EQ.100 .AND. TIME(1).EQ.0) THEN
STATEV(105)=TEMP
IF ((STATEV(1).LT.6).AND.(STATEV(1).GE.3)) THEN
STATEV(78) = 4
CMNAME='FIBROUS TISSUE'
ELSE IF ((STATEV(1).LT.3) .AND. (STATEV(1).GE.1)) THEN
STATEV(78) = 3
CMNAME='CARTILAGE'
ELSE IF ((STATEV(1).LT.1.) .AND. (STATEV(1).GE.0.267)) THEN
STATEV(78) = 2
CMNAME='IMMATURE BONE'
ELSE IF ((STATEV(1).LT.0.267) .AND. (STATEV(1).GE.0.011)) THEN
STATEV(78) = 1
CMNAME='MATURE BONE'
ELSE
STATEV(78) = 5
CMNAME='NO TISSUE FORMATION'
ENDIF
END IF
END IF
IF (JRCD.NE.0)THEN
write(6,*) 'REQUEST ERROR IN USDFLD FOR ELEMENT NUMBER ',
1 NOEL,'INTEGRATION POINT NUMBER ',NPT
ENDIF
RETURN
END