ciliate ichthyophthirius multiftlis
TRANSCRIPT

Proc. Nati. Acad. Sci. USAVol. 89, pp. 6363-6367, July 1992Developmental Biology
Developmental expression of surface antigen genes in the parasiticciliate Ichthyophthirius multiftlisTHEODORE G. CLARK*, ROYAL A. MCGRAWt, AND HARRY W. DICKERSON**Department of Medical Microbiology, and tDepartment of Physiology and Pharmacology, College of Veterinary Medicine, University of Georgia, Athens,GA 30602
Communicated by John R. Preer, Jr., April 6, 1992
ABSTRACT A 1.2-kilobase (kb) cDNA encoding a majorsurface antigen of the holotrich ciliate Ichthyophthirius multi-flUis (an obligate parasite of fish) has been isolated and used asa probe to examine the expression of immobilization antigen(i-antigen) genes in this system. The cDNA encodes a predictedprotein of 394 amino acids with a tandemiy repeated structurecharacteristic of the i-antigens of the related free-living ciliatesParamecium and Tetahymena. As shown by Northern hybrid-ization analysis with both total and poly(A)+ RNAs, the 1.2-kbcDNA recognizes distinct transcripts of 1.6 and 1.9 kb whichare differentially expressed through the parasite life cycle.During the transition from the host-associated trophozoitestage to the infective tomite stage, steady-state levels of the1.9-kb RNA undergo a marked increase of 250-fold, while the1.6-kb transcript increases only slightly. The absolute amountsofRNA encoding the i-antigen have been quantitated and werefound to reach extremely high levels equivalent to -6% of thepoly(A)+ RNA of 1. mudifihiis tomites. Southern hybridizationanalysis with I. mulkifhlis genomic DNA suggests that at leasttwo genes encode the i-antigen transcripts. In experiments toexamine the effects of temperature on the expression of I.mulifiWis i-antigen genes, levels of the 1.6- and 1.9-kb tran-scripts were found to remain relatively constant in cells main-tained at different temperature extremes. These studies indi-cate that genes encoding i-antigens of 1. multfilfis are devel-opmentally regulated, and they suggest the existence ofalternative mechanisms for the control of surface antigenexpression in ciliates.
Variation in surface antigen expression in the free-livingciliates Paramecium and Tetrahymena is a well-studied phe-nomenon (for review, see refs. 1-3). The antigens them-selves, known as immobilization antigens (or i-antigens), areencoded by a repertoire of genes that are expressed in amutually exclusive fashion; only one type of antigen is madeat any given time. While the expression ofa particular antigentype is relatively stable under a given set of conditions,antigen switching can occur in response to environmentalstimuli, most notably shifts in temperature. For example,Tetrahymena thermophila can express any of seven differentantigen types under selected culture conditions, three beingexpressed at different temperatures (1). Although the mech-anisms which control these variations are unknown, i-antigensynthesis appears to be regulated at the level of RNAtranscription and/or processing (4, 5), and experimentalalteration in i-antigen expression provides an extremelyuseful approach to the analysis of gene regulation in lowereukaryotes.The phenomenon takes on further significance in light of
the ability of parasitic protozoa (including the trypanosomes,malarial plasmodia, etc.) to alter their surface antigens as well(6-9). In these systems, antigen diversity plays an important
role in the evasion of host immunity (6, 7). Parasite surfaceantigens and the i-antigens of ciliates share an interestingstructural feature-namely, the presence of tandemly repet-itive amino acid sequence domains (10-14). Nevertheless,they are clearly the products of different genes, and themolecular mechanisms responsible for their expression ap-pear to be different. Whereas in the free-living ciliates surfaceantigen expression is readily reversible and controlled bychanges in environmental conditions, in the parasites itoccurs during the normal life cycle (either in a stage-specificmanner or sequentially during chronic infection) and is de-velopmentally regulated.
In this regard, the common fish parasite Ichthyophthiriusmultiflujis (Ich) represents a bridge between the free-livingciliates and parasitic protozoa. As a holotrich ciliate, it istaxonomically related to both Paramecium and Tetrahymena(order Hymenostomatida) and, in addition to its obviousphenotypic similarity, has i-antigens directly analogous tothose found on the free-living ciliates (15, 16). At the sametime, Ich is an obligate parasite of fish and has a life cyclereminiscent of the protozoan parasites cited above. It wastherefore of interest to examine the nature of the expressionof i-antigen genes in this system.The work presented here shows that RNA transcripts
encoding a predominant i-antigen of Icht undergo a dramaticchange in abundance during the parasite life cycle, indicatingthat, in addition to control through environmental stimuli (asin Paramecium and Tetrahymena), i-antigen genes can bedevelopmentally regulated. The fact that transcript levelsincrease in parallel with the infectivity of the organism bearson the functional role of surface antigens in this system andis consistent with previous observations suggesting that thei-antigens of Ich are involved in the development of protec-tive immunity in fish (15, 17).
MATERIALS AND METHODSParasite Culture. Ich was maintained on channel catfish,
Ictalurus punctatus, as previously described (15). For RNAisolations, trophozoites were obtained by gently scraping thesurface of infected fish and collecting the freed parasitesindividually with a glass pipette. Tomites were obtained byallowing mature trophozoites to come offfish and go throughnormal rounds of cell division to form infective cells (see Fig.2) (15).
Isolation and Characterization of i-antigen cDNA. A cus-tom-made cDNA library was prepared from trophozoitepoly(A)+ RNA in A ZAP II phage by Stratagene and screenedwith an oligonucleotide probe derived from the N-terminalamino acid sequence of a predominant 48-kDa i-antigen fromIch. The antigen itself was isolated from Ich tomites by
Abbreviations: i-antigen(s), immobilization antigen(s); Ich, Ich-thyophthirius multififfis.*The sequence reported in this paper has been deposited in theGenBank data base (accession no. M92907).
6363
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.

6364 Developmental Biology: Clark et al.
affinity chromatography with a mouse monoclonal antibody(designated 1OH3) that strongly immobilized the Ich isolateused in these studies (18). After purification by SDS/PAGE,the N-terminal amino acid sequence of the 48-kDa antigenwas determined by Edman degradation using Applied Bio-systems 470A/477A automated protein sequencers (see Fig.1). An "antisense" 24-mer oligonucleotide probe (5'-AGCAGCACCAACATCAGTCAAACC-3') correspondingto eight amino acids (Gly-Leu-Thr-Asp-Val-Gly-Ala-Ala)near the N terminus of the protein was synthesized, end-labeled with 32p, and used to screen the cDNA library (19).Among a number of positive recombinants isolated, one(designated clone 2-3) contained a 1.2-kilobase (kb) EcoRPinsert and was further analyzed. The full-length EcoRI insertand individual Pst I restriction fragments were subcloned inM13 phage, and the 1.2-kb cDNA was sequenced in itsentirety off both strands by the dideoxy method of Sanger etal. (20).Northern Hybridization Analysis. Parasites were concen-
trated by centrifugation for 2 min at 1000 x g and total RNAwas isolated from cell pellets after lysis in guanidine thiocy-anate (21). Poly(A)+ RNA was purified by two rounds ofchromatography on oligo(dT)-cellulose. Final concentrationsof total and poly(A)+ RNA were determined by absorbanceat 260 nm. RNA was fractionated by electrophoresis on 1.2%agarose gels containing 2.2 M formaldehyde and transferredto Biotrace nylon filters (Gelman) in 20x SSC (3 M NaCl/0.3M sodium citrate, pH 7.0). Filters were hybridized withalkali-denatured probe (.106 cpm/ml), then washed at 65°Cas described by Mahmoudi and Lin (22). Final wash bufferwas 40 mM Na2HPO4/1% SDS/1 mM EDTA, pH 7.2. For
- P - P D O S Q T Q A O L S DV O A a DV P C P D G T Q T Q A G L T D V G A A D
GAGCTGTTICCATGTCCTGATGGTACMTAGACTCAAGCTGGATCTGATGTAGGTGCTGCTGATI
126
246
366
486
606
726
846
966
1086
E A N G N Q P F A A N N A A RI+XCVGL I IC V P
preparation ofthe probe, the 1.2-kb cDNA insert was purifiedby agarose gel electrophoresis and labeled to >109 cpm/,ugwith [a-32P]dATP by using random oligonucleotide priming(23).QuaDitation ofRNA Transcripts. In all cases, RNA levels
were measured under hybridization conditions of probe ex-cess. Changes in transcript prevalence during developmentwere determined from densitometry scans of autoradio-graphic exposures of Northern blots in which equal amountsof RNA from different stages of the parasite were loaded.X-ray film strips with exposures in the linear range ofresponse of the film were scanned spectrophotometrically at470 nm and individual peak areas were measured and com-pared. Results were normalized to ethidium fluorescence inribosomal RNA bands or to signals generated by hybridiza-tion ofthe same filters with a heterologous actin cDNA underconditions of reduced stringency (24).
Absolute levels of RNA were determined by using asingle-stranded RNA probe as described in Fig. 3. For probesynthesis, the 1.2-kb cDNA was subcloned in pBluescript byin vivo excision from A ZAP II DNA according to the supplier(Stratagene), and the orientation of the cDNA insert wasdetermined by restriction endonuclease analysis. AntisenseRNA was then synthesized under standard conditions fromthe T7 promoter flanking the insert (25) and the template wasremoved by digestion with RQ1 DNase (Promega).
Southern Hybridization Analysis. Total genomic DNA wasisolated from Ich tomites by methods previously describedfor Tetrahymena (26). This DNA was considered to bepredominantly macronuclear in origin by analogy with otherholotrich ciliates (27). DNA was digested to completion and
L OL G T C V N C R P N F Y Y N G G A A Q G 40CTGGTACTTGTGTTAATTGCAGACCTAAIT1TACTATAATGGTGGTGCTGCTrAAGGA
C Q I N R V G S V T N A G D L A T L A T 80GMGCTM1TGGTACTTTT CGCA GDCTAGVT VCTC AAAT TCVTCKPTTACTGACTTTAGCA
O C 8 T Q C P T G T A L D D G V T D V F D R 8 A A Q C V X C K P N F Y Y N aG STMATGCAGTACITTMTGTCCTACTGGCACCACTTACTOATGAA7GNACKAG TAGCCGCATAA70 T
P Q G E A P G V Q V F A A G A Al A A G V A A V T S Q C V P C Q L N I N D 8 P A TCCTTMGGTGAAGCCTGGCTTTAC I 29M OIMG TGCAGGIECOGTACTAOZ&Glizh"TCATAQTA&qaAACgTIGCKAT
A G A Q A N L A T O C S. N Q C P T G S V L D D G V T L V F N T 8 A T L C V R C RGCAGATCAGAANLGTACCAMTATnATQCSNQCPTGACTAVLGVAATLITATLCVKCR
P N F Y Y N G G S P Q G E A P G V Q V F AA GA A A A G V A A V T S Q C V P C QCCTAAACTKANTATMTGGAACACCTTSCMGTGTAIQDG VS
L N K N D S P A T A G A Q A N L A T Q C 8 T Q C P T G T A I Q D G V T L V F S NCTMAACAAAAACGATTCTCCTGCCACTGCAGGTGCCTAGCTAATTAGCCACATAATGCAGTAiCTTAATGTCCAACIXTGGCCOATCAAGCGGACACTTGvlTTlTAGTAAT
S S T Q C S Q C I A N Y F F N G N L E A G K S Q C L K C P V S R T T P A H A P GTCATC C TACINTTGCATTIC NTAAAGKS CLKTTTAAGCCGTASTTAC ACPCAGGT
N T A T Q A T Q C I T T C P A G T V L D D G T S T N F V A S A T E C T I C S A GNTACTGCTQCATQCITTCCACAAGTACVDTAGATATCSCT VTAGC ATC SACTG
F F A S K T T G F T A G T D T C T E C T KX L T S G A T A V N I H QmlGATCAAACACTGGTTACAMvCAGTACTGATACATG TACTGAMTTACTAAAATTACTTCTGT CCACAGCTAAACCTATGAAOTTCGAAAACAT
120
160
200
240
280
320
360
394
1206
ICVPCGBIIM ?gMACE§AUATQCSTQCPTGTALDVVDUV1V ZuuuOzMV' unwv AI
AAGVAAVTSQCVPCQLNKNDSPATAGAQANLATQCS00CPTG DDGV lV AA1JCVK NFYYNGGSPQGEAPGVQVFAAGAAAAGVAAVTSQCVPCQLNKNDSPATAGAQANLATQCSTQCPTGT IDGVTLV!1SI A
I
IIIIll
FIG. 1. Nucleotide and deduced amino acid sequence ofthe Ich i-antigen cDNA. The complete nucleotide and deduced amino acid sequenceofthe 1.2-kb i-antigen cDNA is shown at the top. The N-terminal amino acid sequence ofthe 48-kDa antigen is on the first line; hyphens representamino acids that could not be identified with certainty. The areas in the sequence that are boxed and designated by Roman numerals show theregions ofextended homology within the deduced protein structure, and they are aligned relative to one another at the bottom part of the figure.Domains I and II, and II and III share 82% and 83% identity at the amino acid sequence level, respectively.
Jr rr%^T7mrT.A6,XZJ:
Proc. Naff Acad. Sci. USA 89 (1992)
I

Proc. Natl. Acad. Sci. USA 89 (1992) 6365
fractionated by electrophoresis in 0.7% agarose gels. Gelswere stained in ethidium bromide, and the DNA was trans-ferred to Zeta-Probe nylon membranes (Bio-Rad) as de-scribed by Rigaud et al. (28). Filters were prehybridized 5 hrat 650C in 6x SSC/10x Denhardt's solution (0.2% bovineserum albumin/0.2% Ficoll/0.2% polyvinylpyrrolidone)/0.1% SDS containing 10 ;kg of denatured herring sperm DNAper ml, then hybridized with alkali-denatured probe (.106cpm/ml) for 24 hr under the same conditions. Filters werewashed in two changes of 2x SSC/0.1% SDS at roomtemperature, followed by two changes of 0.2x SSC/0.1%SDS at 650C (15 min each).
RESULTSIsolation of i-antigen cDNA. i-antigens have recently been
isolated from Ich tomites and identified as a pair of integralmembrane proteins on the surface of the parasite. On affinitychromatography with immobilizing mouse monoclonal anti-bodies as the ligand, the two antigens copurify and haveapproximate molecular masses of 48 and 60 kDa on SDS/PAGE (18). Oligonucleotide probes based on the N-terminalamino acid sequence of the predominant 48-kDa protein wereused to screen an Ich cDNA library in A ZAP II, and a 1.2-kbcDNA encoding this antigen was isolated. The nucleotidesequence and corresponding amino acid sequence of the1.2-kb cDNA are shown in Fig. 1. Beginning with a proline(one residue from the N terminus ofthe protein) there is exactcorrespondence between the amino acid sequence predictedfrom the cDNA and the actual amino acid sequence deter-mined chemically from the purified i-antigen. The cDNAitself contains 1249 nucleotides and includes a long openreading frame (nucleotides 3-1187) encoding a deduced pro-tein of 394 amino acids (estimated mass = 39,429 Da). Thereare 23 UAA triplets and 1 UAG triplet interspersed through-out the correct reading frame. The presence of a glutamine(residue 8) at a position which corresponds to a UAG tripletin the cDNA indicates that Ich uses a nonstandard geneticcode shared by a number of other ciliates, in which UAA andUAG specify glutamine rather than acting as stop codons (forreview, see ref. 29).The nucleotide sequence of the 1.2-kb cDNA contains 23
repeats of 10 nucleotides or more. Not surprisingly, thistranslates into a protein with a repetitive structure thatincludes three regions of extended homology (=80 aminoacids in each) arranged in tandem within the central part ofthe deduced protein (Fig. 1). This type of repeat structure ischaracteristic of the G and the SerH3 surface antigens ofParamecium tetraurelia and Tetrahymena thermophila, re-spectively (10, 11).
Differential Expression of i-antigen Genes. To examine theexpression of i-antigen genes in this system, RNA from Ichwas subjected to Northern hybridization analysis using the1.2-kb cDNA as a probe. The results of these studies areshown in Fig. 2. Under conditions of high stringency (<10%base mismatch), the 1.2-kb cDNA recognized two transcriptsof 1.6 and 1.9 kb in Northern blots of both total and poly(A)+RNA. More importantly, when RNA transcript levels fromthe two principal life stages of the parasite (trophozoite andtomite) were compared, a striking difference was seen.During the transition from the host-associated trophozoitestage to the infective tomite stage, the 1.9-kb RNA becameextremely prominent and could be detected even in veryshort exposures of Northern blots [<1 min with 0.5 ,ug ofpoly(A)+ RNA loaded]. In densitometry scans of autoradio-grams, steady-state levels of the 1.9-kb transcript were foundto increase by at least 50-fold with poly(A)+ RNA. Evengreater changes were observed with preparations of totalRNA, and while ribosomal RNA in these samples distorts the1.9-kb region of the gel, these results suggest that some
portion of the i-antigen transcripts may be nonpolyadenyly-lated. In contrast with the dramatic change in expression ofthe 1.9-kb RNA, less than a 2-fold increase was seen with the1.6-kb transcript when poly(A)+ RNAs from trophozoitesand tomites were compared.To determine the absolute amounts of i-antigen-specific
RNA in tomites, we carried out quantitative assays using asingle-stranded (antisense) RNA probe synthesized by invitro transcription from the 1.2-kb cDNA. The results of twodifferent types of analysis with this probe are shown in Fig.3. Both methods yielded identical results and placed i-antigentranscript levels at 6% of the poly(A)+ RNA.
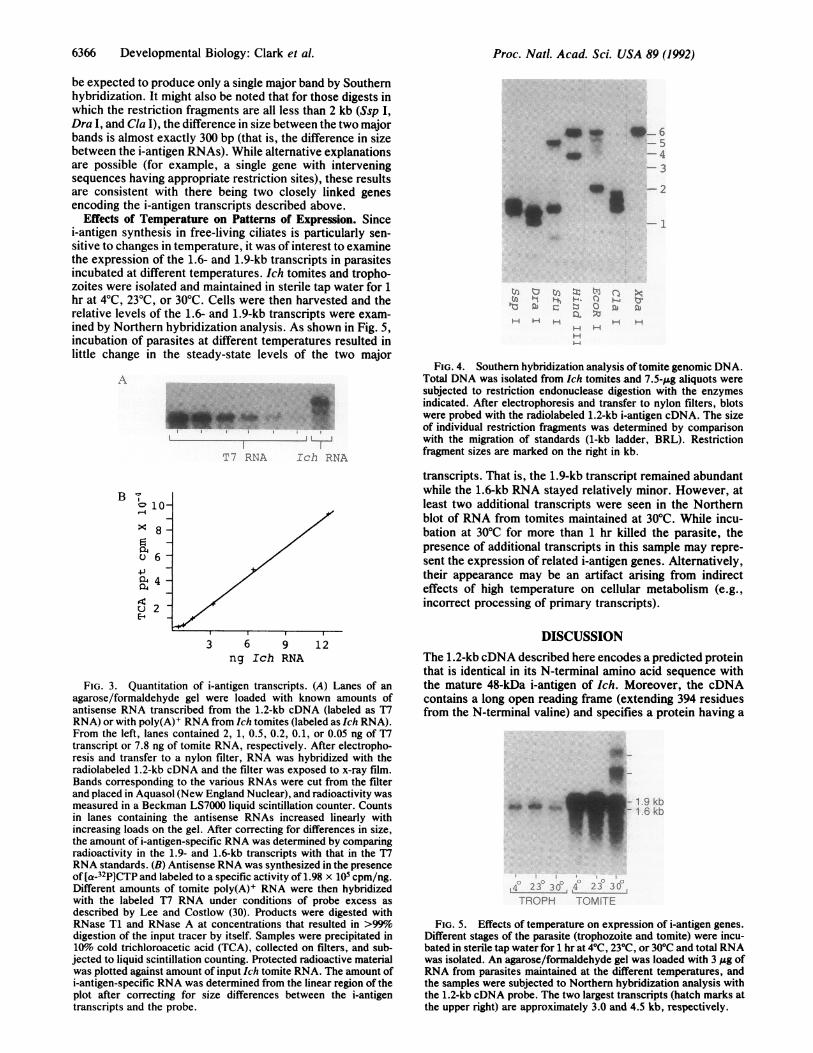
Southern Hybridization Analysis. Since the 1.2-kb cDNArecognized two distinct transcripts, an effort was made toestimate the number of different genes encoding the 48-kDaantigen. Ich genomic DNA was subjected to restrictionendonuclease digestion, blotted on filters, and hybridizedwith the 1.2-kb cDNA. The results are shown in Fig. 4.Among seven different restriction enzymes used, only one(that is, Xba I) produced a single band in the Southern blots.This was a relatively large, 6.4-kb, fragment. All otherenzymes produced two major and one or more minor bands.Four of these enzymes (Cla I, EcoRI, Ssp I, and Xba I) haveno recognition sites in the cDNA, while two (HindIII and SfuI) have single sites within 60 base pairs (bp) of the 3' terminus.If only one gene for the i-antigen exists, these enzymes would
INVASION AND
1.9kb- * -1.9kb1.6 kb- W-1.6 kb
1 2 3 4
FIG. 2. Developmental expression of i-antigen transcripts. Thelife cycle of the parasite is shown on top. Infective tomites swimthrough the water and invade the epithelium of the skin and gills ofhost fish. Once on the fish, they transform into trophozoites that feedoff host tissue. Trophozoites undergo extensive growth (reaching upto =400 gum in diameter), then leave the fish, encyst, and divide toform 200-1000 smaller tomites (=40 gm in diameter). Northernhybridization analysis of RNA from the two principal life stages ofthe parasite is shown at the bottom. Either 3 ,Ag of total RNA (lanes1 and 2), or 0.78 gg of poly(A)+ RNA (lanes 3 and 4) was electro-phoresed on agarose/formaldehyde gels, blotted on nylon filters, andprobed with the radiolabeled 1.2-kb i-antigen cDNA. Results withtrophozoite RNA (lanes 1 and 3) and tomite RNA (lanes 2 and 4) arecompared side by side. Estimates of transcript sizes were based on0.24- to 9.5-kb RNA size standards from BRL (not shown).
Developmental Biology: Clark et al.
6366 Developmental Biology: Clark et al.
be expected to produce only a single major band by Southernhybridization. It might also be noted that for those digests inwhich the restriction fragments are all less than 2 kb (Ssp I,Dra I, and Cla I), the difference in size between the two majorbands is almost exactly 300 bp (that is, the difference in sizebetween the i-antigen RNAs). While alternative explanationsare possible (for example, a single gene with interveningsequences having appropriate restriction sites), these resultsare consistent with there being two closely linked genesencoding the i-antigen transcripts described above.
Effects of Temperature on Patterns of Expression. Sincei-antigen synthesis in free-living ciliates is particularly sen-sitive to changes in temperature, it was of interest to examinethe expression of the 1.6- and 1.9-kb transcripts in parasitesincubated at different temperatures. Ich tomites and tropho-zoites were isolated and maintained in sterile tap water for 1hr at 40C, 230C, or 30'C. Cells were then harvested and therelative levels of the 1.6- and 1.9-kb transcripts were exam-ined by Northern hybridization analysis. As shown in Fig. 5,incubation of parasites at different temperatures resulted inlittle change in the steady-state levels of the two major
,i\
B v
°10
4.4
E 2
Ed
T7 RNA Ich RNA
3 6 9 12ng Ich RNA
FIG. 3. Quantitation of i-antigen transcripts. (A) Lanes of anagarose/formaldehyde gel were loaded with known amounts ofantisense RNA transcribed from the 1.2-kb cDNA (labeled as T7RNA) or with poly(A)+ RNA from Ich tomites (labeled as Ich RNA).From the left, lanes contained 2, 1, 0.5, 0.2, 0.1, or 0.05 ng of T7transcript or 7.8 ng of tomite RNA, respectively. After electropho-resis and transfer to a nylon filter, RNA was hybridized with theradiolabeled 1.2-kb cDNA and the filter was exposed to x-ray film.Bands corresponding to the various RNAs were cut from the filterand placed in Aquasol (New England Nuclear), and radioactivity wasmeasured in a Beckman LS7000 liquid scintillation counter. Countsin lanes containing the antisense RNAs increased linearly withincreasing loads on the gel. After correcting for differences in size,the amount of i-antigen-specific RNA was determined by comparingradioactivity in the 1.9- and 1.6-kb transcripts with that in the T7RNA standards. (B) Antisense RNA was synthesized in the presenceof [a-32p]CTP and labeled to a specific activity of 1.98 x 105 cpm/ng.Different amounts of tomite poly(A)+ RNA were then hybridizedwith the labeled T7 RNA under conditions of probe excess asdescribed by Lee and Costlow (30). Products were digested withRNase T1 and RNase A at concentrations that resulted in >99%odigestion of the input tracer by itself. Samples were precipitated in10%o cold trichloroacetic acid (TCA), collected on filters, and sub-jected to liquid scintillation counting. Protected radioactive materialwas plotted against amount of input Ich tomite RNA. The amount ofi-antigen-specific RNA was determined from the linear region of theplot after correcting for size differences between the i-antigentranscripts and the probe.
Adw ..
__. ._
(n rj-j(J) " t~'a 0
S.
H.A-
40-6- 5-4-3
-2
-1
C)0TuH-
FIG. 4. Southern hybridization analysis of tomite genomic DNA.Total DNA was isolated from Ich tomites and 7.5-,ug aliquots weresubjected to restriction endonuclease digestion with the enzymesindicated. After electrophoresis and transfer to nylon filters, blotswere probed with the radiolabeled 1.2-kb i-antigen cDNA. The sizeof individual restriction fragments was determined by comparisonwith the migration of standards (1-kb ladder, BRL). Restrictionfragment sizes are marked on the right in kb.
transcripts. That is, the 1.9-kb transcript remained abundantwhile the 1.6-kb RNA stayed relatively minor. However, atleast two additional transcripts were seen in the Northernblot of RNA from tomites maintained at 30TC. While incu-bation at 30TC for more than 1 hr killed the parasite, thepresence of additional transcripts in this sample may repre-sent the expression of related i-antigen genes. Alternatively,their appearance may be an artifact arising from indirecteffects of high temperature on cellular metabolism (e.g.,incorrect processing of primary transcripts).
DISCUSSIONThe 1.2-kb cDNA described here encodes a predicted proteinthat is identical in its N-terminal amino acid sequence withthe mature 48-kDa i-antigen of Ich. Moreover, the cDNAcontains a long open reading frame (extending 394 residuesfrom the N-terminal valine) and specifies a protein having a
*~-I9k
_*, _ t - ~~~19 kb1 6 kb
4 23 30 4 23 3OTROPH TOMITE
FIG. 5. Effects of temperature on expression of i-antigen genes.Different stages of the parasite (trophozoite and tomite) were incu-bated in sterile tap water for 1 hr at 40C, 230C, or 30'C and total RNAwas isolated. An agarose/formaldehyde gel was loaded with 3 ,ug ofRNA from parasites maintained at the different temperatures, andthe samples were subjected to Northern hybridization analysis withthe 1.2-kb cDNA probe. The two largest transcripts (hatch marks atthe upper right) are approximately 3.0 and 4.5 kb, respectively.
Proc. Natl. Acad. Sci. USA 89 (1992)
qwW.' wlw.-,n..4

Proc. Natl. Acad. Sci. USA 89 (1992) 6367
periodic structure that is highly reminiscent of the i-antigensof both Tetrahymena and Paramecium. These results indi-cate that the 1.2-kb cDNA used in this study encodes ani-antigen. The discrepancy in size between the predictedprotein and the i-antigen itself (that is, 39,429 Da versus-48,000 Da) may be due to secondary modifications of themature protein and/or structural anomalies leading to incor-rect estimates of size based on SDS/PAGE. Discrepancies ofthis magnitude are known to occur for other surface antigenshaving periodic structure, most notably the circumsporozoiteprotein of Plasmodium knowelsi (12). Nevertheless, the1.2-kb cDNA described here recognizes transcripts largerthan itself and is clearly not full length. Although it containsa long open reading frame extending from the N terminus ofthe mature protein, a cloning artifact (for example, an errorin synthesis, or a deletion) could account for the discrepancyas well.Under conditions of high stringency, the 1.2-kb cDNA
recognized distinct RNA transcripts of 1.6 and 1.9 kb. TheSouthern hybridization data indicate that these transcriptsmay arise from different genes. Yet, while their preciserelationship remains to be determined, the two RNAs showstrong homology at the sequence level and presumablyencode the same protein or two highly related proteins. Inthis regard, a 60-kDa membrane polypeptide copurifies withthe 48-kDa antigen on affinity chromatography with mousemonoclonal antibodies that immobilize Ich in vitro (18).Furthermore, both proteins react with monoclonal antibodieson Western blots, indicating that they have epitopes incommon (18). Since the difference in apparent molecularmass between the proteins corresponds closely to the differ-ence in size (and potential coding information) in the tran-scripts, it is tempting to speculate that the 1.6- and 1.9-kbRNAs encode the 48- and 60-kDa antigens, respectively.Definitive evidence assigning the larger transcript to the60-kDa antigen will, however, await the complete sequencingof representative full-length cDNA clones.Changes in i-antigen expression in the free-living ciliates
are known to occur in response to environmental stimuli(1-3). The results shown here provide evidence for control ata fundamentally different level. Although it might be arguedthat these changes are quantitative and that antigen switchingper se does not occur, the magnitude of variation in transcriptlevels is quite substantial and indicates that genes encodingi-antigens of Ich are developmentally regulated. Develop-mental expression of surface antigen genes occurs in a widevariety of parasitic protozoa and, while Ich is more closelyrelated to the free-living ciliates, it may have evolved thistype of control in concert with its parasitic life style. Aquestion remains, however, as to the underlying function ofthe i-antigens themselves. Do they perform similar functionsin both Ich and the free-living ciliates, or have they under-gone adaptive evolution to serve different functions in thedifferent types of organisms? Fish exposed to mild Ichinfections develop acquired immunity and produce humoralantibodies that immobilize the parasite in vitro (17, 31). Thus,the i-antigens of Ich interact with the immune system of thehost and, consistent with the role of surface antigens in otherparasitic protozoa, may be involved in infection and immu-nity. The free-living ciliates, on the other hand, live quiteapart from any host. The only occasion in which their surfaceantigens might function in such a context would be in theevasion of predators. Indeed, surface antigens of Tetrahy-mena appear to have a role in prey recognition by predatoryprotozoa (32). While this type of model could provide arationale for the environmental regulation of i-antigen ex-
pression in the free-living ciliates, direct evidence is lacking,and the precise function of the i-antigens remains in doubt.Finally, on a more applied level, because the i-antigens ofIchinteract with the immune system of fish, they have potentialas protective immunogens and may be of practical use in thetreatment of a pathogen with major impact on aquacultureworldwide.
Appreciation is extended to Debra Jackwood and Mary Smith fortheir excellent technical assistance. This work was supported bygrants from the Competitive Research Grants Program, U.S. De-partment of Agriculture (89-37266-4649), the University of GeorgiaProgram in Biological Resources and Biotechnology, and the Vet-erinary Medical Experiment Station, University of Georgia.
1. Preer, J. R., Jr. (1986) in The Molecular Biology of CiliatedProtozoa, ed. Gall, J. G. (Academic, London), pp. 301-339.
2. Schmidt, H. (1988) in Paramecium, ed. GOrtz, H.-D. (Springer,Berlin), pp. 155-166.
3. Caron, F. & Meyer, E. (1989) Annu. Rev. Microbiol. 43, 23-42.4. Preer, J. R., Preer, L. B. & Rudman, B. M. (1981) Proc. Natl.
Acad. Sci. USA 78, 6776-6778.5. Love, H. D., Jr., Allen-Nash, A., Zhao, Q. & Bannon, G. A.
(1988) Mol. Cell. Biol. 8, 427-432.6. Borst, P. & Cross, G. A. M. (1982) Cell 29, 291-303.7. Vickerman, K. (1978) Nature (London) 273, 613-617.8. Nardin, E. H. & Nussenzweig, R. S. (1978) Nature (London)
274, 55-57.9. Kasper, L. H., Bradley, M. S. & Pfefferkorn, E. R. (1984) J.
Immunol. 132, 443-449.10. Prat, A., Katinka, M., Caron, F. & Meyer, E. (1986) J. Mol.
Biol. 189, 47-60.11. Tondravi, M. M., Willis, R. L., Love, H. D., Jr., & Bannon,
G. A. (1990) Mol. Cell. Biol. 10, 6091-6096.12. Ozaki, L. S., Svec, P., Nussenzweig, R. S., Nussenzweig, V.
& Godson, G. N. (1983) Cell 34, 815-822.13. Coppel, R. L., Cowman, A. F., Lingelbach, K. R., Brown,
G. V., Saint, R. B., Kemp, D. J. & Anders, R. F. (1983)Nature (London) 306, 751-756.
14. Peterson, D. S., Wrightsman, R. A. & Manning, J. E. (1986)Nature (London) 322, 566-568.
15. Clark, T. G., Dickerson, H. W. & Findly, R. C. (1988) Dev.Comp. Immunol. 12, 581-594.
16. Dickerson, H. W., Clark, T. G. & Findly, R. C. (1989) J.Protozool. 36, 159-164.
17. Clark, T. G., Dickerson, H. W., Gratzek, J. B. & Findly, R. C.(1987) J. Fish Biol. 31, 203-208.
18. Lin, Y.-L. & Dickerson, H. W. (1992) J. Protozool. 39, 457-463.
19. Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989) MolecularCloning:A Laboratory Manual (Cold Spring Harbor Lab., ColdSpring Harbor, NY), pp. 11.20-11.55.
20. Sanger, F., Nicklen, S. & Coulson, A. R. (1977) Proc. Natl.Acad. Sci. USA 74, 5463-5467.
21. Cathala, G., Savouret, J.-F., Mendez, B., West, B. L., Karin,M., Martial, A. & Baxter, J. (1983) DNA 2, 329-335.
22. Mahmoudi, M. & Lin, V. K. (1989) Biotechniques 7, 331-333.23. Feinberg, A. P. & Vogelstein, G. (1982) Anal. Biochem. 132,
6-13.24. Schwartz, R. J., Haron, J. A., Rothblum, K. & Dugaiczyk, A.
(1980) Biochemistry 19, 5890-5898.25. Nielson, D. & Shapiro, D. (1986) Nucleic Acids Res. 14, 5936.26. Kavenoff, R. & Zimm, B. (1973) Chromosoma 41, 1-27.27. Karrer, K. M. (1986) in The Molecular Biology of Ciliated
Protozoa, ed. Gall, J. G. (Academic, London), pp. 85-110.28. Rigaud, G., Grange, T. & Pictet, R. (1987) Nucleic Acids Res.
15, 857.29. Steinbruck, G. (1990) Eur. J. Protistol. 26, 2-14.30. Lee, J. J. & Costlow, N. A. (1987) Methods Enzymol. 152,
633-648.31. Hines, R. S. & Spira, D. T. (1974) J. Fish Biol. 6, 373-378.32. Sunderman, C. A., Paulin, J. J. & Dickerson, H. W. (1986) J.
Protozool. 33, 473-475.
Developmental Biology: Clark et al.