characterization of the porphobilinogen deaminase...
TRANSCRIPT

Characterization of the Porphobilinogen Deaminase
Deficiency in Acute Intermittent Porphyria
IMMUNOLOGICEVIDENCEFORHETEROGENEITYOF THE
GENETIC DEFECT
PETERM. ANDERSON,RAMANM. REDDY,KARLE. ANDERSON,and ROBERTJ. DESNICK,Division of Medical Genetics, Mount Sinai School of Medicine,New York 10029; The Rockefeller University Hospital,New York 10021
A B S T RA C T The molecular pathology of the por-phobilinogen (PBG)-deaminase deficiency in heterozy-gotes for acute intermittent porphyria (AIP) was in-vestigated by means of biochemical and immunologictechniques. The stable enzyme-substrate intermediates(A, B, C, D, and E) of PBG-deaminase were separatedby anion-exchange chromatography of erythrocytelysates from heterozygotes for AIP and normal in-dividuals. In normal lysates, the intermediates elutedin a characteristic pattern with decreasing amounts ofactivity (A > B > C > D > E), the combined A and Bintermediates representing >75% of total recoveredactivity. In contrast, two different profiles were ob-served in lysates from heterozygotes for AIP. In mostheterozygotes, the elution profile was similar to that ofnormal individuals, but each intermediate was reduced-50%. A second profile in which the C intermediatehad disproportionately higher activity than the A or Bintermediates was observed in asymptomatic heterozy-gotes with high urinary levels of PBG(>5 ,ug/ml) aswell as in heterozygotes during acute attacks. Thesefindings suggested that the C intermediate (thedipyrrole-enzyme intermediate) may be rate limitingin the stepwise conversion of the monopyrrole, PBG,to the linear tetrapyrrole, hydroxymethylbilane. Toinvestigate further the nature of the enzymatic defectin AIP, sensitive immunotitration and immunoelectro-phoretic assays were developed with the aid of a rabbitanti-human PBG-deaminase IgG preparation producedagainst the homogeneous enzyme. Equal amounts oferythrocyte lysate activity from 32 heterozygotes forAIP from 22 unrelated families and 35 normal indivi-
Received for publication 12 December 1980 and in revisedform 17 February 1981.
duals were immunoelectrophoresed. There were nodetectable differences in the amounts of cross-reactiveimmunologic material (CRIM) in lysates from thenormal individuals and 25 heterozygotes from 21 of the22 unrelated families with AIP. In contrast, when equalenzymatic activities were coimmunoelectrophoresed,all seven heterozygotes from one family had - 1.6 timesthe amount of CRIM compared with that detected innormal lysates. Consistent with these findings, im-munotitration studies also demonstrated similar quan-tities of noncatalytic CRIM in lysates from this AIPfamily. When equal activities of the individual A, B,C, and D enzyme-substrate intermediates from normaland CRIM-positive erythrocytes were immunoelec-trophoresed, increased amounts of immunoreactiveprotein were observed for each intermediate, B > A
- C - D, from the CRIM-positive AIP variants. Onthe basis of these findings, it is hypothesized that theenzymatic defect in the CRIM-positive AIP family re-sulted from a mutation in the structural gene for PBG-deaminase which altered the catalytic as well as asubstrate binding site. These studies of the enzymaticdefect provide the first demonstration of geneticheterogeneity in AIP.
INTRODUCTION
Acute intermittent porphyria (AIP),' a dominantly in-herited inborn error of heme biosynthesis, is char-acterized by the half-normal activity of porphobilinogen(PBG)-deaminase (EC 4.3.1.8) (1, 2). The enzymaticdeficiency has been demonstrated in erythrocytes
IAbbreviations used in this paper: AIP, acute intermittentporphyria; CRIM, cross-reactive immunologic material; PBG,porphobilinogen.
J. Clin. Invest. © The American Society for Clinical Investigation, Inc. 0021-9738/81/07/0001/12 $1.00Volume 68 July 1981 1-12
1

(1-6), liver (7, 8), cultured skin fibroblasts (9), amnioticcells (10), and cultured lymphoblasts (11) from hetero-zygotes for AIP. Comparison of the physical and kineticproperties of the PBG-deaminase activity in erythro-cyte lysates from heterozygotes for AIP and normalindividuals have shown essentially identical electro-phoretic mobilities (3), heat denaturation profiles (12),and apparent Michaelis constant (Ki) values (5, 12).In addition, mitogen-stimulated lymphocytes fromheterozygotes for AIP induced only 50% of the activ-ity detected in mitogen-treated lymphocytes from nor-mal individuals (11). Although these studies are con-sistent with a structural gene mutation (1), it has beensuggested that the half-normal levels of PBG-deaminaseactivity in AIP may be due to a regulatory genedefect (11), but immunologic evidence supporting astructural mutation of the PBG-deaminase gene inAIP, i.e., the demonstration of noncatalytic cross-reactive immunologic material (CRIM), has not beenreported to date.
Increasing attention has focused on the characteriza-tion of PBG-deaminase from various sources (13-19)and the mechanism by which this monome, zc enzyme(-37,000 mol wt) converts the monopyrrole substrate,PBG, to the final tetrapyrrole product, uroporphyrinogenL (20-27). Recently, we reported the first purificationof homogeneous PBG-deaminase from human eryth-rocytes (27). Anion-exchange chromatography of thepurified enzyme resolved the activity into five forms,designated A, B, C, D, and E. Interconversion studiesand experiments with [3H]PBG demonstrated that thefive forms were stable enzyme-substrate intermediatesin the sequential condensation of four PBGmolecit esinto the tetrapyrrole, uroporphyrinogen L (27). 'i lheoccurrence of these stable PBG-deaminase inter-mediates was independently confirmed by 13C-nuclearmagnetic resonance studies that demonstrated theorder of assembly of the four pyrrole rings and es-tablished that the end-product of the PBG-deaminasereaction was the linear tetrapyrrole, hydroxymethyl-bilane, which nonenzymatically cyclized to form theuroporphyrinogen I isomer (22-25). Fig. 1 illustratesthe current concept of the stepwise enzymatic con-version of the monopyrrole, PBG, to the tetrapyrrole,hydroxymethylbilane. In the presence of uropor-phyrinogen III cosynthase, hydroxymethylbilane wasshown to be enzymatically converted into the uro-porphyrinogen III isomer, the precursor of heme,cobalamins, and the cytochromes (25, 26).
In light of the discovery of the stable PBG-deaminase enzyme-substrate intermediates, an investi-gation was undertaken to characterize the molecularpathology of the enzymatic defect in AIP biochem-ically and immunologically. We report here (a) theactivity profiles of enzyme-substrate intermediates inerythrocyte lysates from normal individuals and hetero-
Hydroxymethylbilone--Uroporphyrinogen I
ENH3 (E - +RC- NH2
NH3 ~~~~A
E-RC-RC-RC-RC-NH2 E-RC-NH2(EE) ( EB)
+RC-N
+RC-NH2
IH2 NH3
E RC-RC-RC-NH2 ERC-RC-H2
(ED ) (EC)
NH3 + RC-NH2
FIGURE 1 Current concept of tetrapyrrole biosynthesis.PBG-deaminase (E) catalyzes the sequential head-to-tail con-( isation of four molecules of PBG (RC-NH2) to form thelinear tetrapyrrole, hydroxymethylbilane. EA represents thenative enzyme, and EB, EC, ED, and EE, the mono-, di-, tri-,and tetrapyrrole enzyme-substrate intermediates, respectively.
zygotes for AIP, and (b) the quantification of PBG-deaminase immunoreactive protein in lysates fromnormal individuals and from AIP heterozygotes in 22unrelated families. These studies provide the firstdemonstration of genetic heterogeneity in this dom-inantly inherited enzymatic deficiency.
METHODS
MaterialsPBG was synthesized as described previously (28). Uro-
porphyrin standards were purchased from Porphyrin Prod-ucts, Logan, Utah. Diaminobenzidine tetrahydrochloride,8-aminolevulinic acid hydrochloride, DL-dithioerythritol DL-dithiothreitol, and human serum albumin were obtainedfrom Sigma Chemical Co., St. Louis, Mo. High purityacetonitrile was purchased from Burdick and Jackson Labo-ratories, Inc., Muskegon, Mich. Fluorescamine was ob-tained from Pierce Cheni dl Company, Rockford, Ill. En-zyme grade ammonium si late was a product of Schwartz/Mann, Div. Becton, Dickimson & Co., Orangeburg, N. Y.Female New Zealand rabbits were from the Pocono RabbitFarm, Canadensis, Pa. Goat anti-rabbit serum and peroxidase-conjugated sheep anti-rabbit IgG were obtained from N. L.Cappel Laboratories, Cochranville, Pa. DEAE-cellulose (DE-52) was a product of H. Reeve Angel & Co., Inc., Clifton,N. J. Seakem agarose was purchased from Marine Colloids,Inc., Div. FCC Corp., Rockland, Maine. High resolutionTris/barbital buffer was obtained from Gelman InstrumentCompany, Ann Arbor, Mich. The following products werepurchased from Pharmacia Fine Chemicals, Div. of PharmaciaInc., Piscataway, N. J.: Sephadex G-100, phenyl-SepharoseCL-4B, octyl-Sepharose CL-4B, Agarose IEF, Pharmalyte(pH 5-8) ampholines, and Gelbond film. All other materialsand chemicals were of the highest quality available.
2 P. M. Anderson, R. M. Reddy, K. E. Anderson, and R. J. Desnick

Humelani suibjects a ad specimen collection32 heterozygotes for AIP were studied; the diagnosis of
each was biochemically documented by demonstration ofdecreased PBG-deaminase activity in erythrocyte lysates.These stubjects were from 22 uinrelated fiamiilies representingat least 10 different ethnic backgrounds or countries oforigin. Heparinized blood was collected from each heterozy-gote and 35 age- and sex-matched normal individuals. Fol-lowing centrifugation at 2,500 g, the erythrocytes were re-moved, washed twice with 0.9% NaCl, and then either usedimmediately or stored at -20°C.
AssaysPBG-deaminase assay. Enzymatic activity was determined
by the measurement of uroporphyrin fluorescence as pre-viously described (27). One unit of enzymatic activity equals1 nmol of uroporphyrin produced per hour at 37°C.
Protein assay. Protein concentrations were determinedby the fluorescamine procedure as previously described (28).
Separation of PBG-deacminase enzyme-substrate intermlediatesAnion-exchan£ge chromatography. Washed, centrifuged
erythrocytes (2.0 ml of packed cells) were lysed by the addi-tion of 20 ml of cold distilled water and centrifuged at35,000 g for 20 min. The supernate was dialyzed against2 liters of 7 mMpotassitumii phosphate buiffer, pH 6.8, con-taining 0.1 mMDL-dithioerythritol.
A 0.9 x 30-cm column of DEAE-cellulose that had pre-viously been acid and base washed was equilibrated withthe same buffer used for dialysis (27). Each dialyzed eryth-rocyte lysate was applied to the column at a flow rate of 1.0ml/min, and then 60 ml of buffer was passed through thecolumn. PBG-deaminase enzyme-substrate intermediates wereeluted as peaks of enzymatic activity using a 400-mllinear NaCl (0 to 0.11 M) gradient. Fractions (2.0 ml) werecollected and 0.5 ml was assayed for PBG-deaminase activityat 37°C for 2 h.
Isoelectric focusing. Horizontal agarose slab gels and thePharmacia system were used for isoelectric focusing of thePBG-deaminase intermediates in accordance with the manu-facturer's instructions. Gels were prepared by heating a mix-ture of 0.3 g Agarose IEF (Pharmacia), 3.6 g sorbitol,and 27 ml distilled water in a boiling water bath. Aftercooling to 75°C, 1.9 ml of pH 5-8 Pharmalyte ampholineswas added. The mixture was then pouired into a horizontalcasting frame (11.4 x 22.5 cm with Gelbond film backing) thathad been preheated to -60°C using a portable hairdryer.Gels were either allowed to harden at least 1 h at 40C orstored overnight at 4°C in a moist chamber. Before isoelec-tric focusing, erythrocyte lysates were heated at 600C for 1 h,centrifuged at 100,000 g for 20 min, and then aliquots ofthe supernate (50-100 ,ll) were pipetted onto Whatman no.17 paper strips (Whatman, Inc., Clifton, N. J.), which wereplaced 1 cm from the cathode. The cathode contained 1 MNaOHand the anode, 0.05 M H2S04. Focusing was carriedout for 2.5 h at 10 W(constant power) at 4°C. The filter paperstrips were removed after the hemoglobin had migratedabout 1 cm from the origin. Upon completion, a 0.5-cm stripwas removed from the gel, cut into 1-cm pieces, soaked indistilled H2O, and the pH of the leached ampholytes wasdetermined. The gel was overlaid with Whatman no. 3MMfilter paper saturated with 0.5 mMPBG, covered with poly-vinylchloride film, and incubated at 37°C for 90 min. The
overlay was removed, the gel was exposed to ultravioletlight for 10 min to oxidize the uroporphyrinogen to uropor-phyrin, and the red fluorescent bands of PBG-deaminaseactivity were rapidly photographed under UV light withPolaroid type 58 film (Polaroid Corp., Cambridge, Mass.) witha Wratten no. 4 filter (Eastman Kodak Co., Rochester, N. Y.).
Crossed-immunoisoelectric focusing of the separated PBG-deaminase intermediates was performed by removing anunstained lane from the focusing gel and pouring an antibody-containing agarose gel such that the individual intermediatescould be electrophoresed directly into it. Rocket immuno-electrophoresis was carried out as described below, exceptthat a larger gel (5.0 x 7.5 cm) containing 8 ml of 1% agaroseand 40 ,ul of antiserum was used.
Immunologic studiesPurification of PBG-deaminase from human erythrocytes.
Human PBG-deaminase was purified to homogeneity as pre-viously described by Anderson and Desnick (27). The prep-aration was routinely stored after the Sephadex G-100 stepin the presence of 1.0 mM DL-dithioerythritol. Whenindividual enzyme-substrate intermediates were required,the preparation was subjected to the final DEAE-cellulosechromatographic step (27).
Production of rabbit anti-human PBG-deaminase IgG.New Zealand rabbits were injected intradermally and intra-muscularly with 150 ,ug of homogeneous human PBG-deaminase A in a 1:1 suspension of Freund's completeadjuvant. Booster injections of 150, 100, and 75 ,ug were givenat 1-mo intervals. The titers of rabbit anti-human PBG-deaminase were determined by immunotitration. The IgGfraction from the pooled rabbit sera was partially purifiedby the method of Harboe and Ingeld (29). The antibodywas precipitated by the addition of 4 vol of saturated am-monium sulfate, pH 7.0, to 6 vol of rabbit serum. Afterstirring 2 h at 4°C, the solution was centrifuged at 2,500 g for20 min and washed twice with 1.5 M ammonium sulfate,pH 7.0. Hemoglobin and albumin were in the supernateand >90% of the IgG was recovered in the pellet. The pelletwas resuspended in distilled water and extensively dialyzedagainst 5 mMpotassium phosphate buffer, pH 7.4, contain-ing 0.9% NaCl. The IgG fraction was divided into 1.5-mlaliquots and stored at -20°C.
Immunotitration of anti-human PBG-deaminase. Homo-geneous PBG-deaminase A (40 ng of enzyme diluted inbovine serum albumin to 50 ,ul having a final protein con-centration of 40 mg/ml) and 50 ,u of rabbit anti-PBG-deaminaseIgG (in serial dilutions) were mixed and incubated at 370C.After 30 min, 50 ,ul of goat anti-rabbit IgG was added andthe mixture was incubated at 37°C for 30 min. To insurequantitative precipitation of the anti-PBG-deaminase anti-bodies, the samples were left overnight at 4°C. The mix-tures were then centrifuged at 4,000 g for 15 min. Theactivity in the supemate was determined by the standardassay described above. For immunotitration of the immuno-reactive PBG-deaminase in erythrocyte lysates from normaland AIP heterozygotes, the lysate activities were equalizedand then either 50- or 25-Al aliquots were titrated againstthe antibody as described above.
Quantitation of CRIM in normal controlsand individuals with AIPSample preparation. Erythrocytes from normal individ-
uals and heterozygotes for AIP were lysed by the addi-tion of 3 vol of 1.0 mMsodium phosphate buffer, pH 7.6,
Characterization of the Enzymatic Defect in Acute Intermittent Porphyria 3

containing 1 mM DL-dithiothreitol, 1 mM MgCl2, and0.05% Triton X-100. The lysate was centrifuged at 30,000 gfor 20 min and the supemate was removed and used forassays of protein and PBG-deaminase activity. Samples werediluted and then reassayed to ensure that all lysates con-tained equal enzymatic activity before rocket immuno-electrophoresis.
Rocket immunoelectrophoresis. The rocket immunoelec-trophoretic system for human PBG-deaminase was de-veloped as a modification of standard methods (30). A 1%agarose solution was prepared in 0.06 MTris-barbital buffer,pH 8.8, and 6.0 ml was poured into a 5 x 7.5-cm plate inthe center of which a 0.4 x 2.5 x 7-cm double thicknessglass microscope slide had been placed. After the agarosehad gelled, the slide was removed and 4.0 ml of a 1%agarosesolution containing 15 ul of rabbit anti-human PBG-deaminasewas poured into the center area. The agarose had been cooledto 60-65°C before antibody was added.
For routine immunoelectrophoresis of erythrocyte lysates,11 2-mm wells were cut into the lower portion of the anti-body-containing gel and 7-,ul samples were added to eachwell. Cotton wicks were used to make contact between thegel and the 0.06 MTris-barbital electrophoresis buffer. Theplate was electrophoresed for 4 h at a constant current of30 mA. Following electrophoresis, the gel was thoroughlywashed in 0.9% NaCl for 10-12 h with several changesand then overlayed with 300 ul of sheep anti-rabbit peroxidase-conjugated IgG that previously had been diluted 1:2 with0.9% NaCl. The plate was incubated at room temperaturein a moist chamber to prevent drying. The gel was againwashed with 0.9% NaCl for at least 8 h and then stainedfor peroxidase. The staining solution was 25 mg diamino-benzidine tetrahydrochloride dissolved in 50 ml 0.1 MTris/HCI buffer, pH 7.6, containing 0.15 ml of 3% hydrogen per-oxide. After the gel and staining solution were allowed toreact for 20 min at room temperature, the gel was photo-graphed and stored in 0.9% NaCl. The peroxidase reactionmixture stained the gel brown; rockets appeared whiteagainst the brown background. Crossed immunoisoelectricgels were stained with Coomassie Brilliant Blue R-250 anddestained as previously described (31).
RESULTS
PBG-deaminase enzyme-substrate intermediates inerythrocytes. Fig. 2 shows representative DEAE-cellulose chromatographic elution profiles of the PBG-deaminase enzyme-substrate intermediates in erythro-cyte lysates from normal individuals and heterozygotesfor AIP. In 12 normal individuals the activity of thePBG-deaminase A and B forms always contained >75%of total recovered activity (Fig. 2A). Occasionally, theC intermediate partially separated into two over-lapping peaks. Two different elution patterns wereobserved in AIP heterozygotes (Fig. 2B and C). Mostheterozygotes for AIP (n = 10) had profiles similar tonormal individuals (compare 2A and B); howeverthe activities of the A, B, and C intermediates wereeach reduced by -50%. The low activities in the Dand E peaks did not permit comparison. All heterozy-gotes with this pattern were clinically asymptomaticand had urinary PBG levels <5 ,ug/ml. In contrast,eight heterozygotes who were experiencing acuteattacks of the disease and/or had elevated urinary
. Ii
UL I-
°~O-2 IA ItB Xz
_AIP-1
Z0.0
o 1 1.1
Z IAl BIB C D E
0 ~ 4 60 80 10 2 4
PBG-damin AIP-2.0 0) ax l
0. /ID
[
I I~~~~~~~~~~I
40O 60 80 100 120 140'FRACTION NUMBER
FIGURE2 DEAE-cellulose chromatographic profiles ofPBG-deaminase from normal and AiP erythrocytes. Lysates(2.0 ml) were applied to a 0.9 x 30-cm column and PBG-deaminase enzyme-substrate intermediates (A- E) wereeluted using a linear 0-0.11 M NaCl gradient as describedin Methods. Normal (top graph); AIP type 1 (AIP-i) profile(middle graph), observed in patients in remission whenurinary PBG levels were <5d,g/ml; AIP type 2 (AIP-2) pro-file (bottom graph), observed during clinical exacerbationsand/or when urinary PBG levels were >5 ug/ml.
PBG levels (>5 ug/ml) had a different pattern of en-
zyme-substrate intermediates (Fig. 2C). In these hetero-zygotes, there was a disproportionate increase in theactivity of the C intermediate, particularly when com-
4 P. M. Anderson, R. M. Reddy, K. E. Anderson, and R. J. Desnick
pared with the A and B activities. In two heterozygotes,erythrocyte samples were obtained during an acuteattack and subsequently during remission. Both hetero-zygotes had type 2 profiles (Fig. 2C) during the acuteattack and type 1 elution patterns (Fig. 2B) in remis-sion; these profiles also correlated with the level ofurinary PBGexcretion.
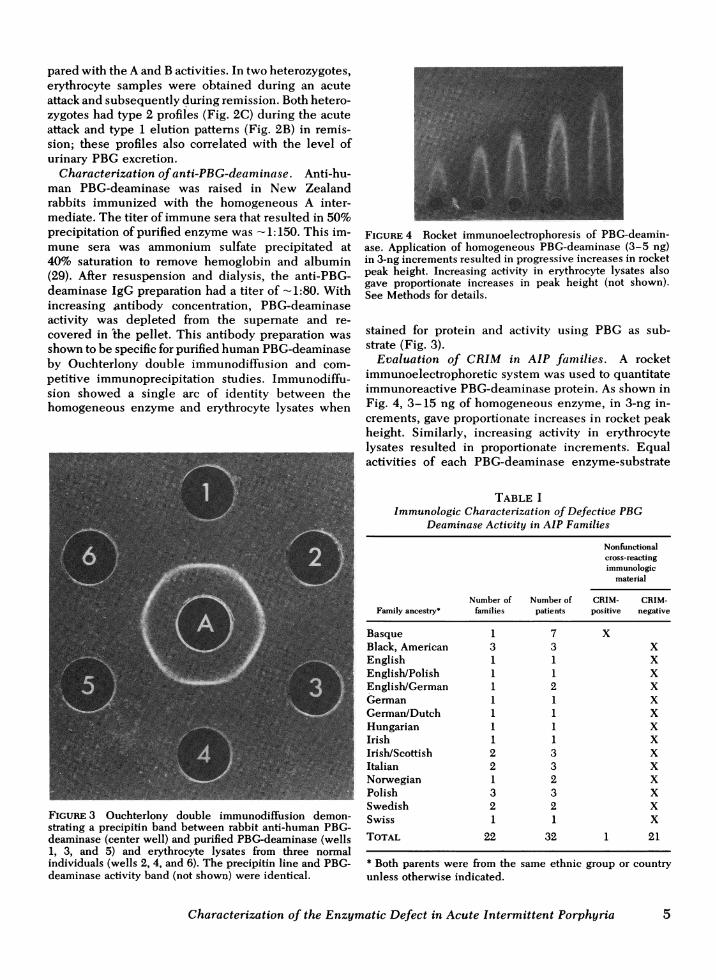
Characterization of anti-PBG-deaminase. Anti-hu-man PBG-deaminase was raised in New Zealandrabbits immunized with the homogeneous A inter-mediate. The titer of immune sera that resulted in 50%precipitation of purified enzyme was - 1:150. This im-mune sera was ammonium sulfate precipitated at40% saturation to remove hemoglobin and albumin(29). After resuspension and dialysis, the anti-PBG-deaminase IgG preparation had a titer of -1:80. Withincreasing antibody concentration, PBG-deaminaseactivity was depleted from the supernate and re-covered in 'the pellet. This antibody preparation wasshown to be specific for purified human PBG-deaminaseby Ouchterlony double immunodiffusion and com-petitive immunoprecipitation studies. Immunodiffu-sion showed a single arc of identity between thehomogeneous enzyme and erythrocyte lysates when
FIGURE 3 Ouchterlony double immunodiffusion demon-strating a precipitin band between rabbit anti-human PBG-deaminase (center well) and purified PBG-deaminase (wells1, 3, and 5) and erythrocyte lysates from three normalindividuals (wells 2, 4, and 6). The precipitin line and PBG-deaminase activity band (not shown) were identical.
FIGURE 4 Rocket immunoelectrophoresis of PBG-deamin-ase. Application of homogeneous PBG-deaminase (3-5 ng)in 3-ng increments resulted in progressive increases in rocketpeak height. Increasing activity in erythrocyte lysates alsogave proportionate increases in peak height (not shown).See Methods for details.
stained for protein and activity using PBG as sub-strate (Fig. 3).
Evaluation of CRIM in AIP families. A rocketimmunoelectrophoretic system was used to quantitateimmunoreactive PBG-deaminase protein. As shown inFig. 4, 3-15 ng of homogeneous enzyme, in 3-ng in-crements, gave proportionate increases in rocket peakheight. Similarly, increasing activity in erythrocytelysates resulted in proportionate increments. Equalactivities of each PBG-deaminase enzyme-substrate
TABLE IImmunologic Characterization of Defective PBG
Deaminase Activity in AIP Families
Nonfunctionalcross-reactingimmunologic
material
Number of Number of CRIM- CRIM-Family ancestry* families patients positive negative
Basque 1 7 XBlack, American 3 3 XEnglish 1 1 XEnglish/Polish 1 1 XEnglish/German 1 2 XGerman 1 1 XGerman/Dutch 1 1 XHungarian 1 1 XIrish 1 1 XIrish/Scottish 2 3 XItalian 2 3 XNorwegian 1 2 XPolish 3 3 XSwedish 2 2 XSwiss 1 1 X
TOTAL 22 32 1 21
* Both parents were from theunless otherwise indicated.
same ethnic group or country
Characterization of the Enzymatic Defect in Acute Intermittent Porphyria 5

intermediate isolated from normal erythrocytes re-sulted in essentially identical rocket heights. Thepresence or addition of PBGdid not affect peak height.
Table I summarizes the CRIM status in erythrocytelysates from 32 AIP heterozygotes from 22 unrelatedfamilies representing at least 10 different ethnic back-grounds. When equal amounts of erythrocyte lysateactivity from normal individuals and 25 heterozygotesfor AIP from 21 unrelated families were immunoelec-trophoresed, there were no significant differences inthe peak heights of immuno-reactive PBG-deaminase(Fig. 5). For each normal individual and each heterozy-gote from these 21 AIP families, the amount of activ-ity applied corresponded to the amount of CRIMdetected. Thus, the mutations in the AIP heterozy-gotes from these families were designated as CRIMnegative. However, in a family of Basque ancestry, theamount of CRIM was consistently greater in all sevenAIP heterozygotes studied (Fig. 6). These findingsdemonstrated the presence of noncatalytic, immuno-logically cross-reactive material (CRIM-positive) inthis AIP family of Basque ancestry.
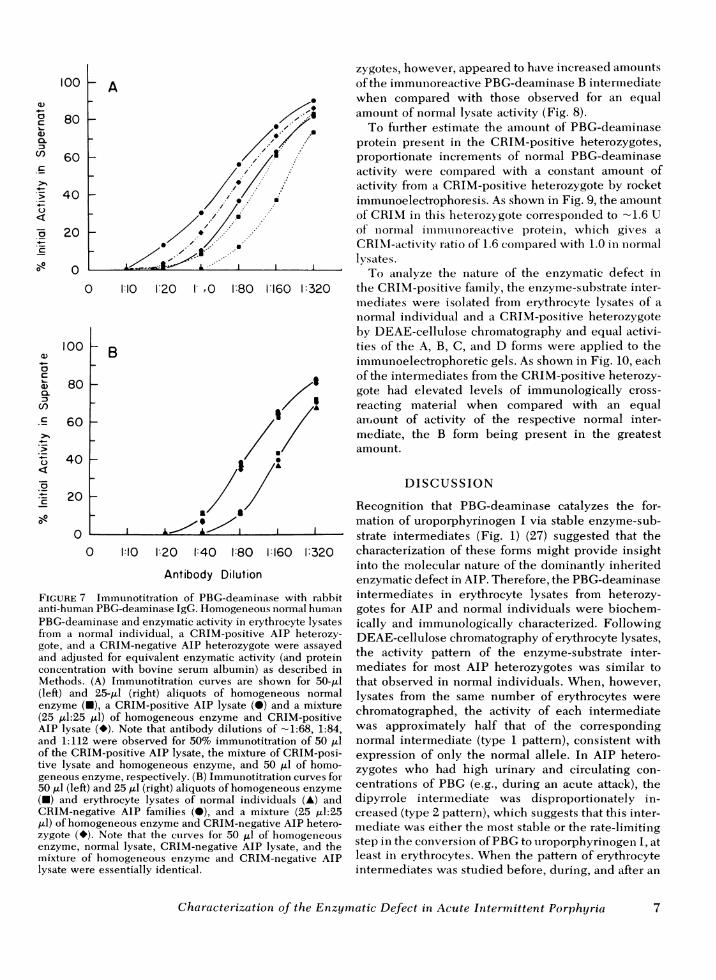
Consistent with these results, competitive immuno-titration studies (Fig. 7) revealed the presence of non-catalytic PBG-deaminase protein in the AIP hetero-zygotes from the Basque family (Fig. 7A) and the ab-sence of noncatalytic CRIM in the other AIP familiesstudied (Fig. 7B). When samples containing equalactivities of homogeneous PBG-deaminase or PBG-deaminase in erythrocyte lysates from normal individ-uals or the CRIM-positive family were immuno-titrated, the antibody dilutions required to precipitate50% of the activity in the supernates were -1:112,1:112, and 1:68, respectively. A mixture containingequal activities from a CRIM-positive and a normallysate required on antibody dilution of - 1:86 for 50%immunoprecipitation. On the basis of these findings,the estimated amount of immunoreactive protein in theCRIM-positive variant was 1.65-fold that in normal
FIGURE 5 Rocket immunoelectrophoresis of PBG-deamin-ase in erythrocytes from CRIM-negative AIP heterozygotesand normal individuals. Equal enzymatic activities wereapplied to the gel. No differences in rocket peak heights ofimmunoreactive enzyme were observed for equal activitiesof erythrocyte PBG-deaminase from normals (odd channels)and the (CRIM-negative) AIP heterozygotes (even channels).
A-1 A-2 A-3 A-4 A-5FIGURE 6 Rocket immunoelectrophoresis of PBG-deaminasein erythrocytes from CRIM-positive AIP heterozygotes.Equal enzymatic activity in erythrocyte lysates from hetero-zygotes of an AIP kindred of Basque ancestry (Al-A5, evenlanes) and normal individuals (odd lanes) were applied. Therocket peak heights of immunoreactive enzyme from eachAIP heterozygote was increased when compared with thosein lysates from normal individuals or unaffected membersof this kindred (not shown).
lysates, assuming identical antibody avidities. In con-trast, when equal amounts of erythrocyte lysate activityfrom a CRIM-negative AIP heterozygote and a normalindividual as well as purified enzyme were immuno-titrated, essentially the same antibody dilution was re-quired to precipitate 50%of activity from each (Fig. 7B),which indicates a direct correspondence betweenactivity and immunotitratable enzyme protein.
Characterization of the noncatalytic PBG-deaminasein the CRIM-positive family. The Km and stabilityof the PBG-deaminase activity in erythrocyte lysatesfrom the CRIM-positive AIP heterozygotes were com-pared with those of GRIM-negative AIP heterozygotesand normal individuals. The Kmvalues in all three sourceswere essentially the same (-10 ,tM). However, theimmunoreactive PBG-deaminase in the CRIM-positiveheterozygotes remained stable after heating the lysatefor 30 min at 60°C, whereas the immunoreactiveenzyme protein in normal and CRIM-negative AIPlysates was in each case decreased -15% after 30 minof heat treatment. Agarose isoelectric focusing demon-strated no significant differences in the isoelectricpoints of homogeneous human PBG-deaminase andPBG-deaminase in erythrocyte lysates from normal,CRIM-positive, or CRIM-negative individuals. Rou-tinely, five bands of enzymatic activity were visualizedwith pI of -6.4, 6.1, 5.8, 5.5, and 5.3. When sampleswere subjected to isoelectric focusing in the firstdimension and then to crossed-immunoelectrophoresisin the second dimension, no evidence for a mutantprotein with different isoelectric properties was ob-tained; a precipitin line of identity, which corre-sponded to each of the five PBG-deaminase inter-mediates, was observed in lysates from normal in-dividuals and CRIM-negative and CRIM-positive AIPheterozygotes (Fig. 8; CRIM-negative AIP lysate notshown). Lysates from the CRIM-positive AIP hetero-
6 P. M. Anderson, R. M. Reddy, K. E. Anderson, and R. J. Desnick
80
60
40
20
0
100
80
60
40
20
0
loKt A
0 1: 10 :20 I' 0 1:80 1:160 1:320
) B
'I0 I~~~~
I L.. ki
0 1:10 1:20 1:40 1:80 1:160 1:320
Antibody Dilution
FIGURE 7 Immunotitration of PBG-deaminase with rabbitanti-human PBG-deaminase IgG. Homogeneous normal humanPBG-deaminase and enzymatic activity in erythrocyte lysatesfrom a normal individual, a CRIM-positive AIP heterozy-gote, and a CRIM-negative AIP heterozygote were assayedand adjusted for equivalent enzymatic activity (and proteinconcentration with bovine serum albumin) as described inMethods. (A) Immunotitration curves are shown for 50-Al(left) and 25-Al (right) aliquots of homogeneous normalenzyme (U), a CRIM-positive AIP lysate (*) and a mixture(25 ,ul:25 Al) of homogeneous enzyme and CRIM-positiveAIP lysate (*). Note that antibody dilutions of -1:68, 1:84,and 1:112 were observed for 50% immunotitration of 50 ,lof the CRIM-positive AIP lysate, the mixture of CRIM-posi-tive lysate and homogeneous enzyme, and 50 Al of homo-geneous enzyme, respectively. (B) Immunotitration curves for50 Al (left) and 25 ,tl (right) aliquots of homogeneous enzyme
(-) and erythrocyte lysates of normal individuals (A) andCRIM-negative AIP families (@), and a mixture (25 ,ul:25,l) of homogeneous enzyme and CRIM-negative AIP hetero-zygote (*). Note that the curves for 50 Al of homogeneousenzyme, normal lysate, CRIM-negative AIP lysate, and themixture of homogeneous enzyme and CRIM-negative AIPlysate were essentially identical.
zygotes, however, appeared to have increased amountsof the immunoreactive PBG-deaminase B intermediatewhen compared with those observed for an equalamount of normal lysate activity (Fig. 8).
To fu-rther estimate the amount of PBG-deaminaseprotein present in the CRIM-positive heterozygotes,proportionate increments of normal PBG-deaminaseactivity were compared with a constant amount ofactivity from a CRIM-positive heterozygote by rocketimmunoelectrophoresis. As shown in Fig. 9, the amountof CRIM in this heterozygote corresponded to -1.6 Uof normlal imimunliioreactive protein, which gives aCRIM-activity ratio of 1.6 compared with 1.0 in normallvIsates.
To analyze the nature of the enzymatic defect inthe CRIM-positive family, the enzyme-substrate inter-mediates were isolated from erythrocyte lysates of anormal individual and a CRIM-positive heterozygoteby DEAE-cellulose chromatography and equal activi-ties of the A, B, C, and D forms were applied to theimmunoelectrophoretic gels. As shown in Fig. 10, eachof the intermediates from the CRIM-positive heterozy-gote had elevated levels of immunologically cross-reacting material when compared with an equalairmount of activity of the respective normal inter-mediate, the B form being present in the greatestamount.
DISCUSSION
Recognition that PBG-deaminase catalyzes the for-mation of uroporphyrinogen I via stable enzyme-sub-strate intermediates (Fig. 1) (27) suggested that thecharacterization of these forms might provide insightinto the molecular nature of the dominantly inheritedenzymatic defect in AIP. Therefore, the PBG-deaminaseintermediates in erythrocyte lysates from heterozy-gotes for AIP and normal individuals were biochem-ically and immunologically characterized. FollowingDEAE-cellulose chromatography of erythrocyte lysates,the activity pattern of the enzyme-substrate inter-mediates for most AIP heterozygotes was similar tothat observed in normal individuals. When, however,lysates from the same number of erythrocytes werechromatographed, the activity of each intermediatewas approximately half that of the correspondingnormal intermediate (type 1 pattern), consistent withexpression of only the normal allele. In AIP hetero-zygotes who had high urinary and circulating con-centrations of PBG (e.g., during an acute attack), thedipyrrole intermediate was disproportionately in-creased (type 2 pattern), which suggests that this inter-mediate was either the most stable or the rate-limitingstep in the conversion of PBGto uroporphyrinogen I, atleast in erythrocytes. When the pattern of erythrocyteintermediates was studied before, during, and after an
Characterization of the Enzymatic Defect in Acute Intermittent Porphyria7
a)c
Q._C
0-0.C/
0
'I)4-0c
0L)(a.
Uf)c
-
C.)
C-
0Il
7

N.i.,
e
FIGURE 8phoresis ofmal indivicelectric foactivity inshown) lys,for each balowing crosproteins, oractivity baipositive (Agotes. See I
acute attack in the same heterozygote, the type 2pattern (increased C intermediate) occurred during theattack concomitant with high urinary PBG levels... '. 'This would suggest that the induction of hepatic 8-aminolevulinate synthase during an acute attack re-sulted in the increased production of 8-aminolevulinicacid and PBG (32) that gained access to the eryth-rocyte where the PBG became bound to the PBG-deaminase (33). Thus, the patterns of the PBG-deaminase intermediates in AIP heterozygotes wereconsistent with the expression of the normal alleleand were dependent, in part, on the substrate con-centration in erythrocytes.
Immunologic characterization of the enzymatic de-fect revealed genetic heterogeneity in AIP. Mono-specific rabbit anti-human PBG-deaminase was pro-duced for the first time, permitting immunotitrationand immunoelectrophoretic studies of the enzyme.Erythrocytes proved to be the enzyme source of choice;the use of cultured lymphocytes or cultured fibro-blasts was precluded since 1 ml of packed cultured
i^6 F D E cells had -0.5 and 0.18%, respectively, of the totalactivity in 1 ml of packed normal erythrocytes. Whenthe amount of CRIM was determined by rocket im-
_o~~~~~~~munoelectrophoresis, erythrocyte lysates from hetero-RN-: .;0 00; zygous members of 21 unrelated AIP families were
:s iX> found to have GRIM-activity ratios of 1.0, i.e., the ratio'sgv-"~ [ > lofimmunoreactive protein (estimated by rocket peak,~~~~,~~height) corresponded to that observed for an equal
amount of enzymatic activity from normal lysates(Fig. 5). Similarly, when equal amounts of PBG-deaminase activity in erythrocytes from these AIPheterozygotes and normal individuals were immuno-titrated against the monospecific antibody, the 50%immunotitration values were essentially identical,which indicates the absence of noncatalytic immuno-reactive protein in the erythrocytes from the heterozy-gotes in these families (Fig. 7). Thus, the mutationin these 21 unrelated AIP families was designatedas CRIM negative.
In contrast, when erythrocyte lysates from all sevenheterozygous members (whose clinical manifestationswere indistinguishable from the above CRIM-nega-tive heterozygotes) of an AIP kindred of Basqueancestry were subjected to rocket immunoelectro-
Isoelectric focusing and crossed-immunoelectro- phoresis (Fig. 6) and immunotitration (Fig. 7), non-PBG-deaminase in erythrocyte lysates from nor- catalytic immunoreactive protein was observed. The
luals and CRIM-positive AIP heterozygotes. Iso- . .cusing revealed five bands when stained fornormal, CRIM-positive, and CRIM-negative (not positive AIP heterozygotes, as determined by immuno-ates. No significant differences in the pl values electrophoresis and immunotitration, were both -1.6,and in the different lysates were observed. Fol- compared with 1.0 obtained for normal or CRIM-nega-ssed-immunoelectrophoresis of the focused lysate tive AIP heterozygotes. Isoelectric focusing followednly immunoreactive proteins corresponding to thends were observed in the normal (N), CRIM- by crossed-immunoelectrophoress of the focused pro-.), or CRIM-negative (not shown) AIP heterozy- teins did not reveal differences in the pI values or im-Methods for details. munoelectrophoretic profiles of the PBG-deaminase
8 P. M. Anderson, R. M. Reddy, K. E. Anderson, and R. J. Desnick
A

Activity: /tOS'ource: N
.- a.-l
I I1.0 125A N
I I1.0 /50A N
I I1.0 /.75A N
I1.0 2.0A Nl
I I1.0 /0A N
FIGURE 9 Estimation of the CRIM-activity ratio (CRIM/enzymatic activity) in a CRIM-positiveAIP heterozygote. Every other channel contained increments of normal (N) lysate activity (1.0,1.25 . . . and 2.0 U). Whencompared with a constant amount of activity (1.0 U) from the CRIM-positive AIP heterozygote (A), the amount of immunoreactive PBG-deaminase in the AIPheterozygote was about equal to 1.6 units of normal activity. Thus, the ratio of CRIM tospecific activity for the CRIM-positive heterozygote was -1.6. See Methods for details.
intermediates in lysates from normal individuals orfrom CRIM-negative or CRIM-positive AIP heterozy-gotes. Thus, immunoreactive protein was only de-tected in the regions corresponding to the activitybands for the A, B, C, and D intermediates, whichwere resolved by isoelectric focusing (Fig. 8). In addi-tion, the immunoreactive PBG-deaminase from theCRIM-positive AIP heterozygotes was thermal stable(60°C for 30 min). These findings suggested that thenonfunctional, immunoreactive protein occurred asstable substrate-bound intermediates that did not havedetectably altered isoelectric points. If the nonfunc-tional immunoreactive protein had an altered charge(e.g., owing to an amino acid substitution), an addi-tional peak(s) of immunoreactive protein should be ob-served after isoelectric focusing followed by crossedimmunoelectrophoresis.
The fact that the noncatalytic, immunoreactiveprotein focused with the normal catalytic PBG-deaminase intermediates permitted the considerationof several different models for the further characteriza-tion of the nature of the enzymatic defect in theCRIM-positive AIP heterozygotes in light of severalhypothetical models. Fig. 11 illustrates several dif-ferent types of mutations that might be revealed bycharacterizing the CRIM status of the PBG-deaminaseintermediates in the CRIM-positive heterozygotes. Onthe assumption that the normal enzyme monomer(A) has two binding sites for PBGand an active site(34), the stepwise formation of the mono-, di-, tri-, andtetrapyrroles by the enzyme-substrate intermediates,B through E, can be depicted as shown. If substratebinding site-i was defective, no PBGcould bind andonly the free enzyme or A intermediate would ac-cumulate. If the second substrate binding was de-fective, the CRIM-positive B intermediate would ac-cumulate, presumably in an amount greater than the
A form. A defective catalytic site would result in theaccumulation of the first three intermediates, presum-ably the amount of immunoreactive C >B, and B > A.If a mutation resulted in altered binding and/orcatalysis, a kinetic mutation would result (markedlyhigher K.), and all of the enzyme-substrate inter-mediates would accumulate; the amount of immuno-reactive B or C intermediate would depend on whetherthe binding or catalytic site was more defective.Finally, if a mutation altered the enzyme such thatthe hydroxymethylbilane could not be released, allforms might accumulate, presumably the E inter-mediate in the greatest amount. To discriminateamong these possible mutations, erythrocyte A, B, C,and D intermediates from the CRIM-positive AIPheterozygote were separated and immunologicallyquantified (Fig. 10). Each of the intermediates fromthe CRIM-positive AIP heterozygotes had elevated
FIGURE 10 Rocket immunoelectrophoresis of PBG-deaminaseintermediates. Equal activities of each intermediate, isolatedby DEAE-cellulose chromatography from normal andCRIM-positive AIP erythrocytes, were immunoelectro-phoresed. From left to right: lanes 1, 2, 4, 6, 8, 10, and 11,normal erythrocyte PBG-deaminase intermediates A, A, B, C,D, A, A, respectively; lanes 3, 5, 7, and 9, PBG-deaminaseA, B, C, and D, respectively, from AIP. Note that each inter-mediate from the CRIM-positive AIP heterozygote had an in-creased amount of immunoreactive material, the greatestamount in the B or monopyrrole-enzyme intermediate.
Characterization of the Enzyi'natic Defect in Acute Intermittent Porphyria 9

ActiveNORMAL ste
SubstroteBinding Sites
MUTATIONS
1. DefectiveSubstrateBindingSite-I
2. DefectiveSubstrateBinding Site-2
Hydroxymethylbilone
Km = 6,uM
A AccumulationA
A
3. Defective fCatalytic Site
B4
CSB0
B > A Accumulation
C > B > A Accumulation
Mutation ( _ -KmMarkedly Increased
A B >C= D- E Accumulation
5. Defective B\ (P NoRelease tJD?) 0 +- Product
E > D> C > B > A Accumulation
FIGURE 11 Possible alterations in PBG-deaminase resulting from different point mutations inthe structural gene. The normal enzyme (1) is assumed to have one active site and two substratebinding sites; the stepwise formation of the mono-, di-, tri-, and tetrapyrrole-enzyme intermediates(B, C, D, and E, respectively) is shown. Different mutations altering substrate binding (1 and 2),the catalytic site (3), the Km(4) or product release (5), and the expected formation andlor accumula-tion of the enzyme intermediates for each are depicted. See Discussion for details.
levels of immunologically cross-reacting material whencompared with an equal amount of activity for the re-spective normal intermediate. Notably, the im-munoreactive B form was present in the greatestamount. These findings suggested that the structuralgene defect in this family best fits the model for aKm mutation with a markedly altered, but functional,active site and an altered substrate binding site-2.
An unexpected result was the fact that in 21 of the22 unrelated AIP families, the PBG-deaminase muta-tion was CRIM negative. Several possible explana-tions may be advanced to account for these findings.First, the mutation(s) in these families may havealtered the enzyme conformation such that its anti-genic site or sites was not recognized by our anti-PBG-deaminase. It is, however, unlikely that a rabbitpolyvalent antibody preparation (35, 36) would notrecognize many, if not most, structurally altered pro-teins analogous to the experience with other enzyme
deficiency disorders (37). A more likely explana-tion would be a point mutation that resulted in amarkedly unstable PBG-deaminase that would berapidly degraded. Preliminary attempts to identifya highly unstable enzyme included studies of reticu-locyte-enriched preparations and cultured lympho-cytes and fibroblasts. Peripheral erythrocytes from twounrelated CRIM-negative AIP heterozygotes and twonormal individuals were separated into age cohorts bydiscontinuous density gradient centrifugation (38). Al-though the reticulocyte-rich fractions contained in-creased PBG-deaminase activity when compared withunfractionated circulating erythrocytes, the CRIM-activity ratio for the CRIM-negative AIP and normalreticulocytes were both essentially 1.0. In addition,rocket immunoelectrophoretic studies of culturedlymphocytes and fibroblasts (109 cells/sample) fromCRIM-negative AIP heterozygotes and normal in-dividuals did not reveal the presence of nonfunc-
10 P. M. Anderson, R. M. Reddy, K. E. Anderson, and R. J. Desnick

tional immunoreactive material, whereas cultured cellsfrom a CRIM-positive AIP heterozygote had non-catalytic immunoreactive enzyme. Further investiga-tion of the molecular nature of the CRIM-negativemutations may require the use of recombinant DNAtechniques to obtain appropriate DNA probes toevaluate the possibility that the genetic defectsin these AIP kindreds results from chain-terminatingdefects. mRNAprocessing defects, or partial or com-plete gene deletions analogous to those recentlyidentified in the a- and,8p-thalassemias (39-43).
In summary, these investigations have, for the firsttime, identified molecular genetic heterogeneity inAIP. The finding of stable enzyme-substrate inter-mediates for PBG-deaminase is unique among humanenzymes, especially for a monomeric protein whosefunction is necessary for the production of suchmetabolically essential human molecules as the hemo-globins, cytochromes, and cobalamins. The findingof a genetically defective PBG-deaminase which canbe detected immunologically, both as the free proteinand as substrate-bound intermediates, provides thefirst molecular evidence for a structural gene muta-tion as the primary defect in AIP. Further investiga-tion of the enzymatic defect in other AIP kindreds,using the immunologic techniques reported here,should provide additional insight into the molecularpathology and genetic heterogeneity of this dominantlyinherited disorder of human heme biosynthesis.
ACKNOWLEDGMENTSThe authors wish to thank Dr. Bertram Felsher, Long Beach,Calif., for generously providing erythrocytes from a num-ber of AIP heterozygotes; Mrs. Gundula LaBadie and Dr.Eugene Ainbender, Mount Sinai School of Medicine, for theirhelpful suggestions for the immunologic studies; Dr. SergioPiomelli, New York, for assistance with the separation oferythrocytes by age; and Mrs. Linda Lugo for her expertclerical assistance.
This research was supported in part by a research grant(1-535) from the March of Dimes Birth Defects Foundationand a grant (AM 26824) from the National Institutes ofHealth. Dr. Anderson was the recipient of a National In-stitutes of Health predoctoral fellowship (HD 07105) inmedical genetics.
REFERENCES1. Tschudy, D. P. 1974. Porphyrin metabolism and the
porphyrias. In Diseases of Metabolism. G. C. Duncan,editor. W. B. Saunders Company, Philadelphia, Pa.775-824.
2. Meyer, U., and R. Schmid. 1978. The porphyrias. In TheMetabolic Basis of Inherited Disease. J. B. Stanbury,J. B. Wyngaarden, and D. S. Fredrickson, editors.McGraw-Hill Book Company, New York. 1166-1220.
3. Strand, L. J., U. A. Meyer, B. F. Felsher, A. G.Redeker, and H. S. Marver. 1972. Decreased red celluroporphyrinogen I synthetase activity in intermittentacute porphyria.J. Clin. Invest. 51: 2430-2536.
4. Meyer, V. A., L. J. Strand, M. Doss, A. C. Rees, and H. S.Marver. 1972. Intermittent acute porphyria: demonstra-tion of a genetic defect in porphobilinogen metabolism.N. Engl. J. Med. 286: 1277-1282.
5. Magnussen, C. R., J. B. Levine, J. M. Doherty, J. 0.Cheesman, and D. P. Tschudy. 1974. A red cell enzymemethod for diagnoses of acute intermittent porphyria.Blood. 44: 857-868.
6. Brodie, M. J., M. R. Moore, and A. Goldberg. 1977. En-zyme abnormalities in the porphyrias. Lancet. II: 699-701.
7. Strand, L. J., B. F. Felsher, A. G. Redeker, and H. S.Marver. 1970. Heme biosynthesis in intermittent acuteporphyria: decreased hepatic conversion of porpho-bilinogen to porphyrins and increased delta amino-levulinic acid synthetase activity. Proc. Natl. Acad. Sci.U. S. A. 67: 1315-1320.
8. Miyagi, K., R. Cardinal, I. Bossenmaier, and C. J.Watson. 1971. The serum porphobilinogen and hepaticporphobilinogen deaminase in normal and porphyricindividuals.J. Lab. Clin. Med. 78: 683-695.
9. Bonkowski, H. L., D. P. Tschudy, E. C. Weinbach, P. S.Ebert, and J. M. Doherty. 1975. Porphyrin synthesisand mitochondrial respiration in acute intermittentporphyria: studies using cultured fibroblasts. J. Lab.Clin. Med. 85: 93-100.
10. Sassa, S., G. Solish, R. D. Levere, and A. Kappas. 1975.Studies in porphyria. IV. Expression of the gene defect ofacute intermittent porphyria in cultured human skinfibroblasts and amniotic cells. Prenatal diagnosis ofthe porphyric trait. J. Exp. Med. 142: 722-731.
11. Sassa, S., G. L. Zalar, and A. Kappas. 1978. Studies inporphyria. VII. Induction of uroporphyrinogen I syn-thase and expression of the gene defect in acute inter-mittent porphyria in mitogen-stimulated human lym-phocytes.J. Clin. Invest. 61: 499-508.
12. Sassa, S., S. Granick, D. R. Bickers, H. L. Bradlow,and A. Kappas. 1974. Studies in porphyria. III. A micro-assay for uroporphyrinogen I synthetase, one of threeabnormal enzyme activities in acute intermittent por-phyria and its application to the study of the geneticsof this disease. Proc. Natl. Acad. Sci. U. S. A. 71:732-736.
13. Stevens, E. S., R. B. Frydman, and B. Frydman. 1968.Separation of porphobilinogen deaminase and uropor-phyrinogen III cosynthase from human erythrocytes.Biochim. Biophys. Acta. 158: 496-498.
14. Sancovich, H. A., A. M. C. Battle, and M. Grinstein.1969. Porphyrin biosynthesis. VI. Separation and purifica-tion of porphobilinogen deaminase and uroporphyrino-gen isomerase from cow liver: porphobilinogen deamin-ase an allosteric enzyme. Biochim. Biophys. Acta. 191:130-143.
15. Frydman, R. B., and B. Frydman. 1970. Purification andproperties of porphobilinogen deaminase from wheatgerm. Arch. Biochem. Biophys. 136: 193-202.
16. Davies, R. C., and A. Neuberger. 1973. Polypyrrolesformed from porphobilinogen and amines by uropor-phyrinogen synthetase of Rhodopseudomonas spheroides.Biochem. J. 133: 471-492.
17. Jordan, P. M., and D. Shemin. 1973. Purification andproperties of uroporphyrinogen I synthetase from Rho-dopseudomonas spheroides. J. Biol. Chem. 248: 1019-1024.
18. Higuchi, M., and L. Bogorad. 1975. The purification andproperties of uroporphyrinogen I synthetase and uro-porphyrinogen III cosynthetase: interactions between theenzymes. Ann. N. Y. Acad. Sci. 244: 401-418.
Characterization of the Enzymatic Defect in Acute Intermittent Porphyria 11

19. Miyagi, K., M. Kanishima, J. Kawakami, F. Nakada, Z. J.Petryka, and C. J. Watson. 1979. Uroporphyrinogen Isynthase from human erythrocytes: separation, purifica-tion and properties of isozymes. Proc. Natl. Acad. Sci.U. S. A. 76: 6172-6176.
20. Frydman, B., R. Frydman, A. Valasinas, S. Levy, and G.Feinstein. 1975. The mechanism of uroporphyrinogenbiosynthesis. Ann. N. Y. Acad. Sci. 244: 371-395.
21. Scott, I. A., K. S. Ho, and M. Kajiwaiz. 1976. Biosyn-thesis of uroporphyrinogen III from porphobilinogen:resolution of the enigmatic "switch" mechanism. J. Am.Chern. Soc. 98: 1589-1591.
22. Battersby, A. R. 1978. The discovery of nature's bio-synthetic pathways. Exp)erien tia (Basel). 34: 1-13.
23. Battersby, A. R., C. J. R. Fookes, G. W. J. Matcham,and E. McDonald. 1979. Order of assembly of the fourpyrrole rings during biosynthesis of natural porphyrins.
J. Chem. Soc. Chem. Commun. 12: 539-541.24. Jordan, P. M., and J. S. Seehra. 1979. The biosynthesis
of uroporphyrinogen III: order of assembly of the fourporphobilinogen molecules in the formation of thetetrapyrrole ring. FEBS (Fed. Eur. Biochem. Soc.) Lett.104: 364-366.
25. Jordan, P. M., and A. Berry. 1980. Preuroporphyrinogen,a universal intermediate in the biosynthesis of uro-porphyrinogen III. FEBS (Fed. Eur. Biochem. Soc.)Lett. 112: 86-88.
26. Battersby, A. R., C. J. R. Fookes, G. W. J. Matcham,and E. McDonald. 1980. Biosynthesis of the pigments oflife: formation of the macrocycle. Nature (Lond.). 285:17-21.
27. Anderson, P. M., and R. J. Desnick. 1980. Purificationand properties of uroporphyrinogen I synthase fromhuman erythrocytes: identification of stable enzyme-substrate intermediates.J. Biol. Chem. 255: 1993-1999.
28. Anderson, P. M., and R. J. Desnick. 1979. Purificationand properties of 8-aminolevulinate dehydrase fromhuman erythrocytes.J. Biol. Chem. 254: 6924-6930.
29. Harboe, N., and A. Ingeld. 1973. Antibodies: produc-tion, purification, and titre determination. Scand. J. Im-munol. 2(Suppl. 1): 161-162.
30. Laurell, C. B. 1972. Electroimmunoassay. Scand. J. Clin.Lab. Itnvest. 29(Suppl. 24): 21-37.
31. Beratis, N. G., G. U. LaBadie, and K. Hirschhorn. 1978.Characterization of the molecular defect in infantile andadult acid a-glucosidase deficiency fibroblasts. J. Clin.Invest. 62: 1264-1274.
32. Tschudy, D. P., M. G. Perlroth, H. S. Marver, A. Collins,G. Hunter, Jr., and M. Rechcigl, Jr. 1965. Acute inter-mittent porphyria: the first "overproduction disease"localized to a specific enzyme. Proc. Natl. Acad. Sci.U. S. A. 53: 841-846.
33. Viljoen, D., D. Becker, and S. Kramer. 1975. Porphyrinsynthesis and movement across human and rabbit redcell membrane with delta aminolevulinic acid as sub-strate. Comp. Biochemn. Physiol. A Comp. Physiol. 50:587-593.
34. Russell, C. S., and P. Rockwell. 1980. The effects ofsulfhydryl agents on the activity of wheat germ uro-porphyrinogen I synthase. FEBS (Fed. Eur. Biochem.Soc.) Lett. 116: 199-202.
35. Slaughter, C. A., M. C. Coseo, C. Abrams, M. R. Cancro,and H. Harris. 1980. The use of hybridomas in en-zyme genetics. In Monoclonal Antibodies. R. H. Kennett,T. J. McKearn, and K. B. Bechtel, editors. PlenumPublishing Corp., New York. 103-120.
36. Prager, E. M., and A. C. Wilson. 1971. The dependenceof immunochemical cross-reactivity upon sequence re-semblance among lysozymes. 1. Micro-complement fixa-tion studies. J. Biol. Chem. 246: 5978-5989.
37. Sutton, H. E., and R. P. Wagner. 1975. Mutation and en-zyme function in humans. Annu. Rev. Genet. 9: 187-212.
38. Corash, L. M., S. Piomelli, H. 0. Chen, C. Seaman,and E. Gross. 1974. Separation of erythrocytes accord-ing to age on a simplified density gradient. J. Lab.Clin. Med. 84: 147-151.
39. Kan, Y. W., J. Holland, A. Dozy, and H. Varmus. 1975.Demonstration of non-functional fl-globin mRNA inhomozygous 3-thalassemia. Proc. Natl. Acad. Sci. U. S. A.72: 5140-5144.
40. Kan, Y. W., J. P. Holland, A. Dozy, S. Charache, and H.Kazazian. 1975. Deletion of the /8-globin structure genein hereditary persistence of foetal haemoglobin. Nature(Lond.). 258: 162-163.
41. Kantor, J. A., P. H. Turner, and A. W. Nienhuis. 1980.Beta thalassemia: mutations which affect processing ofthe ,/-globin mRNAprecursor. Cell. 21: 149-157.
42. Proudfoot, N. J., M. H. M. Shander, J. L. Manley, M. L.Gefler, and T. Maniatis. 1980. Structure and in vitro tran-scription of human globin genes. Science (Wash. D. C.).209: 1329-1336.
43. Wetherall, D. J., and J. B. Clegg. 1979. Recent de-velopments in the molecular genetics of human hemo-globin. Cell. 16: 467-479.
12 P. M. Anderson, R. M. Reddy, K. E. Anderson, and R. J. Desnick