characterization of the diffusion of non-electrolytes across plant
TRANSCRIPT

FOCUS PAPER
Characterization of the diffusion of non-electrolytes acrossplant cuticles: properties of the lipophilic pathway
Anke Buchholz*
Syngenta Crop Protection AG, Research Biology, CH-4332 Stein, Switzerland
Received 25 November 2005; Accepted 10 April 2006
Abstract
Systemic crop protection products are commonly
sprayed onto foliage, whereupon the active substances
must penetrate into the leaves in order to become
biologically active. Penetration of the plant cuticle is
the rate-limiting step. The diffusion of organic non-
electrolytes within cuticles is a purely physical pro-
cess that can be described and analysed in the same
way as is done for diffusion in synthetic polymer
membranes. Solute mobilities in cuticles vary consid-
erably between plant species. For a given species
they decrease with increasing solute size, and this
size selectivity holds for all of the plant species in-
vestigated so far. Wax extraction from leaf cuticles
increases the mobility of solutes tremendously, but
size selectivity is not affected. Furthermore, diffusion
within plant cuticles is extremely temperature depen-
dent. An analogous increase in solute mobility can
be achieved by using accelerators, which enhance
the fluidity of the polymer matrix and of the waxes.
The effects of temperature and plasticizers on the dif-
fusion of non-electrolytes in wax and the cutin matrix
have been used to characterize the nature of the lipo-
philic pathway. The ‘free volume’ theory can be used
to explain the influence of the size and shape of
the solute, and its dependence on temperature. The
physico-chemical nature of the diffusion pathway has
been shown, by thermodynamic analysis, to be ident-
ical for a wide range of solute lipophilicities. This ap-
proach also explains the mode of action and the
intrinsic activity of plasticizers.
Key words: Cuticular membrane, diffusion, foliar uptake, leaf
surface, penetration, solute mobility.
Introduction
Foliar uptake is a complex process determined by severalmutually affecting parameters. General characteristics—required for the optimization of foliar uptake—can only beobtained when interdependencies have been excluded. Ex-tensive work, focusing on the diffusion of non-electrolytesacross plant cuticles, has been done, which this paperattempts to review. The relevant characterization of cu-ticle penetration for polar compounds is given separatelyby Schonherr (2006). After a general introduction onthe function and character of plant cuticles, the appliedmethodology is presented. Parameters affecting plantcuticle penetration are displayed individually so that finallythe complex scenario of foliar uptake can be outlined ina simplified way.
Function of plant cuticles
All aerial surfaces of the primary parts of terrestrialhigher plants are covered with a cuticle, the function ofwhich is to prevent uncontrolled water loss (Riedererand Schreiber, 2001). The plant cuticle is not only exposedto abiotic factors like sunlight, wind, rain, etc. but italso interacts with microbes, fungi, and insects (Juniper,1995; Kolattukudy et al., 1995; Kerstiens, 1996; Schreiberet al., 2004). Furthermore, the plant cuticle is the initialcontact point between an agrochemical and the plant and,where uptake into plant tissues is required for biologicalactivity, it is the main barrier to penetration.
Morphology, chemistry, and structure ofplant cuticles
Cuticles are not simple homogeneous structures like syn-thetic polymer films. Various structural types occur in
* E-mail: [email protected]: CM, cuticular membrane; k*, solute mobility; MX, matrix membrane; UDOS, unilateral desorption of the outer surface; Vx, molar volume.
Journal of Experimental Botany, Vol. 57, No. 11, pp. 2501–2513, 2006
doi:10.1093/jxb/erl023 Advance Access publication 7 July, 2006
ª The Author [2006]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved.For Permissions, please e-mail: [email protected]
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

gymnosperms and angiosperms, which were categorizedby Holloway (1982) into six morphological types. Inves-tigators of the cuticular ultrastructure of different plantspecies have used different terms to describe these layeredstructures. Wattendorf and Holloway (1980) provided acomparative overview of the different terminologies used.The following generalization as given by Jeffree (1986)(Fig. 1) distinguishes three main zones. The cuticle properwith epicuticular and intracuticular waxes often has alamellate layer and typically extends to a thickness of50–150 nm (Jeffree, 1996). The cuticular layer is bondedto the periclinal walls of the epidermal cells by a pectin-rich layer. This pectinaceous layer is equivalent to themiddle lamella, with which it is continuous. Enzymatichydrolysis of this layer enables the cuticular membranes(Orgell, 1955), which comprise the cuticle proper and thecuticular layer, to be isolated.
The epicuticular wax film is a distinct layer on thesurface of the cutin matrix (Jetter et al., 2000). Dependingon their chemical composition, epicuticular waxes mayform an amorphous film, granules, or crystalline structuresof various shapes (Baker, 1982; Jeffree, 1986). This fine-surface structure might be modified with increasing age(Bukovac et al., 1979), by environmental stress (Hoadet al., 1992), and also by agrochemical spray solutions(Whitehouse et al., 1982; Tamura et al., 2001). Epicuticu-lar waxes have a considerable influence on the wettabilityof plant surfaces, often causing poor retention or spread-ing of spray droplets (Bukovac et al., 1979; Turunen andHuttunen, 1990; Holloway, 1994), but they rarely affecta compound’s rate of penetration into leaves (Baur, 1998).The penetration barrier (Schonherr and Schmidt, 1979)is made up of the cuticular membrane, which consists ofthe cutin polymer matrix (Holloway, 1993) and associated
waxes (Bianchi, 1995). However, the permeability of thisbarrier is independent of the overall thickness of the cuti-cle (Becker et al., 1986; Knoche et al., 2000). The intra-cuticular waxes are predominantly located in the outerlayers of the cuticle where they form the transport-limitinglayer or skin (Schonherr and Riederer, 1988). This waxlayer acts as a barrier to both diffusion and solubility(Shafer and Schonherr, 1985), and it has been establishedthat the solid and crystalline wax aggregates within thislayer determine the transport properties of the plant cuticle(Riederer and Schreiber, 1995).
Penetration across plant cuticles
The penetration of non-electrolytes across plant cuticlesrequires three steps: sorption into the cuticular lipids, dif-fusion across the cuticular membrane, and finally desorp-tion into the apoplast of the epidermal cells (Kirkwood,1999; Schonherr et al., 1999). Rates of penetration de-pend upon solute mobility in the cuticle and on the drivingforce (product of partition coefficient and concentrationgradient; see also below). Solute mobility can be meas-ured using unilateral desorption from the outer surface(UDOS). This method takes advantage of the fact thatcuticles are asymmetric membranes (see above) and distin-guishes two functional layers: the limiting skin and thesorption compartment (Fig. 2). In the cuticles studied, thethin limiting skin comprised only about 10% of the total
Fig. 1. Generalized structure of a plant cuticle (modified according toJeffree, 1986). EW, Epicuticular wax; CP, cuticle proper with lamellatestructure; CL, cuticular layer traversed by cellulose microfibrils; PL,pectinaceous layer and middle lamella; CW, cell wall; P, plasmalemma.
Fig. 2. Functional layers (not to scale) of cuticular membranes asdifferentiated in the UDOS (unilateral desorption of the outer surface) andSOFU (simulation of foliar uptake) methods: limiting skin (LS) andsorption compartment (SC). Occurrence and dimension of cuticular pegs,and protrusion of the cuticular layer into anticlinal cell walls of epidermalcells, are species dependent. The arrows denote the direction of diffusionaccording to the concentration gradient. For one time, the sorptioncompartment (UDOS) and, for the other time, the formulation residuefacing the limiting skin (SOFU) serve as donors. With UDOS rateconstants of desorption, k*, and with SOFU rate constants of penetration,k, can be determined.
2502 Buchholz
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

mass of the cuticular membrane (CM), and both sorptioncapacity and solute mobility were very small. The sorptioncompartment underneath the limiting skin amounted toabout 90% of the mass of the CM, its sorption capacitywas high, and solute mobility was much higher than inthe limiting skin (Schonherr and Baur, 1994). Model com-pounds are applied to the morphological inner surfaceof the CM and are subsequently sorbed in the sorptioncompartment. When they are homogeneously distributedwithin the sorption compartment they are unilaterally de-sorbed from the morphological outer surface.
These measured solute mobilities are first-order rateconstants of desorption (k*), which are independent ofthe partition coefficient and are directly proportional tothe diffusion coefficient in the waxy outer limiting skinsof cuticles.
D = k*3lls3lsoco ð1Þ
where lls3lsoco is the diffusion path length in the limitingskin, ls, and sorption compartment, soco. The translationto diffusion coefficients D (m2 s�1) is only restrictedsince the diffusion path length is not known precisely(Schonherr and Baur, 1994). When only the permeabilityP is determined, sorption processes are additionally in-cluded and cannot be separated.
P= ðD3KÞ=Dx ð2Þ
where K is the partition coefficient and Dx is the membranethickness. The reader is referred to Bauer and Schonherr(1992), Schonherr and Baur (1994), and Baur et al. (1996b)for further details.
The benefit of this method is that the contributionsof mobility and solubility to the rates of foliar uptakeof pesticides can be determined separately, due to the factthat the measured rate constants of desorption (k*) areindependent of the partition coefficient. When the donoris applied to the outer surface of the CM and solutes aredesorbed from the morphological inner side (SOFU=simulation of foliar uptake), rates of desorption alsofollow first-order kinetics, but they depend on both solu-bility and mobility (Baur et al., 1997b).
Plant species
When solute mobility in cuticles is studied using theUDOS method, astomatous CM must be used. Enzymaticisolation does not affect the transport properties of cuti-cles (Kirsch et al., 1997) but it is feasible only with alimited number of species, i.e. those where a continuouspectinaceous layer is present. These are most often peren-nial plants. The range of solute mobilities (e.g. for bifenox)measured for the CMs studied covered three orders ofmagnitude (Fig. 3). These different mobilities are species-specific characteristics and do not correlate with either
Decreasing solute mobility
log k*
-8.5-8-7.5-7-6.5-6-5.5-5-4.5
Prunus persica
Tilia cordata
Populus canescens
Juglans regia
Pyrus pyrifolia
P. communis cv. ConferencePrunus armeniaca
Stephanotis floribunda
Malus baccata
Pyrus communis
Pyrus pyrifolia
M. domestica cv. Golden D.M. domestica cv. Gloster
Citrus grandis
Prunus serotina
Ilex aquifolium
Prunus laurocerasus
Citrus aurantium
Strophanthus gratus
Ginkgo biloba
Melicoccus bijugatus
Hedera helix
Vanilla planifolia
Ilex paraguariensis
Fig. 3. Solute mobilities, logk*, of the model compound bifenox (logKCW, 4.44; molar volume, 216 cm3 mol�1) measured in 24 plant species (CM) at25 8C. Error bars denote 95% confidence intervals.
Lipophilic pathway 2503
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

cuticle thickness or the amount of wax embedded within,and covering, the cuticular matrix. For example, Pyruscommunis, Malus domestica, Ilex aquifolium, and Ilexparaguariensis exhibited a very similar amount of ex-tractable wax (about 24%) but its contribution to the barrierproperty of the CM varied by factors of 49 up to 4897(Table 1). No clear relationship has been found to existbetween the particular composition of the soluble lipidsand the respective cuticular permeability (Haas andSchonherr, 1979; Riederer and Schneider, 1990; Haukeand Schreiber, 1998). Histochemical methods have alsobeen used to try to understand the relationship betweenchemical composition and permeability (Leece, 1976;Kruger et al., 1996).
Cuticular waxes and cutin matrix
Cuticular wax composition is very complex and variesaccording to species, ontogenetic development, and envir-onmental conditions (Bianchi, 1995; von Wettstein-Knowles, 1995). The commonest components are n-alkanesand primary alcohols, both of which range from C16 to C36
(Holloway, 1994). Long alkyl chain compounds exhibita solid-state polymorphism (liquid!hexagonally packedsolid phase!orthorhombically packed solid phase) acrossa temperature scan (Watanabe, 1961). The physical state-of the cuticular wax may contain several phases andcrystalline forms and is likely, therefore, to be the criticalfeature that determines the transport properties of the cuti-cle. Highly ordered crystalline regions are inaccessible topermeation. The physical state of a mixture is a functionof its chemical composition and of physical variables liketemperature, pressure, etc. and is best described using aphase diagram. Phase diagrams for such complex mixturesas cuticular waxes are difficult to obtain and would notbe very informative due to their complexity. The ex-amination of a simple binary mixture of two chemically
homologous waxes (1-tetradecanol and 1-octadecanol) re-vealed very complex mixing characteristics, and suggestedfrequent phase immiscibility (Carreto et al., 2002). There-fore, a mosaic of phase domains had to be expected evenat ambient temperatures.
This coexistence of crystalline and fluid or amorphousphases was determined for the cuticular waxes extractedfrom several different plant species (Reynhardt andRiederer, 1991, 1994; Viougeas et al., 1995; Schreiberet al., 1997), and was also demonstrated in situ (Merket al., 1998). The measurement of the diffusion character-istics in reconstituted waxes is thought to be a reason-able approach since the composition, and the proportion ofcrystallites and amorphous phases is not altered duringrecrystallization (Schreiber and Schonherr, 1993). A goodcorrelation between mobilities measured in reconstitutedcuticular waxes and in CMs was found, which confirmedviscosity as an important property (Schreiber and Riederer,1996; Kirsch et al., 1997). These investigations revealedthat the degree of crystallinity and the spatial arrangementof crystals determined the permeating path through theamorphous phase, which in turn determined the effective-ness of the diffusion barrier (Schreiber et al., 1997).Extracted waxes might, however, crystallize in a differ-ent shape (spatial extension) compared with those con-fined in a polymer matrix. Due to tortuosity (the pathtaken around the crystalline obstacles) the length of thediffusion path can be much greater than the thickness ofthe barrier (Schreiber et al., 1996a; Baur et al., 1999).
The study of synthetic chemical polymer membranescan also help to elucidate the mechanism of diffusionacross CMs, even though they are, in general, thicker andless complex than plant cuticles. This enables a more de-tailed view of the functional relationships to be obtained.For instance, clay platelets are incorporated as inert fillersinto polymers to improve their mechanical and barrierproperties. To make the clay platelets compatible withhydrophobic materials, such as polyolefins and waxes,
Table 1. Comparison of solute mobilities (k*; 695% CI) for bifenox within CM and MX of 13 plant species at 25 8C
Extractable wax fraction (mg cm�2; weight percentage) and differential factors of both solute mobilities are specified.
Plant species CM log k* 95% CI MX log k* 95% CI Factor Wax (mg cm�2) Wax (%)
Populus canescens –5.00 0.07 – – – 0.053 33.9Pyrus pyrifolia –5.34 0.17 –3.29 0.05 112 0.055 25.5P. communis cv. Conference –5.43 0.11 –3.74a – 49 0.061 23.7Stephanotis floribunda –5.68 0.25 –3.73 0.02 89 0.026 10.2M. domestica cv. Golden Delicious –6.20 0.08 –3.36 0.06 692 0.044 23.7Ilex aquifolium –6.64 0.18 –4.14 0.12 316 0.162 24.6Ginkgo biloba –6.97 0.11 –5.54 0.19 27 0.054 13.4Melicoccus bijugatus –7.04 0.11 –4.81 0.06 170 0.049 8.8Hedera helix –7.16 0.14 –3.75 0.05 2570 0.022 10.2Strophanthus gratus –7.18 0.16 –5.34a – 69 0.060 19.5Vanilla planifolia –7.67 0.11 –6.93 0.06 5 0.061 10.8Ilex paraguariensis –7.89 0.11 –4.20 0.15 4897 0.114 22.3Schefflera actinophylla –8.09a – –4.57a – 3311 0.084 29.8
a Solute mobilities at 25 8C were calculated from Arrhenius graphs (Buchholz, 1998).
2504 Buchholz
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

their surfaces are treated with monolayers of specificamines. The mechanism by which these organoclays im-prove the barrier properties of the film relies on the highaspect ratio (relationship between the width and height)of the exfoliated clay platelets to impart a tortuous diffu-sion path. The tortuosity factor can be as high as several-hundred-fold for impermeable platelets with aspect ratiosof 100–500 and at modest mineral loadings of 5–10vol% (Cussler et al., 1988).
The tortuosity of nanocomposite materials, however,is rarely that high (typically 2- to 4-fold) and it has beenshown that the interphase surrounding the clay plateletsexerts a profound influence on gas permeation (Chaikoand Leyva, 2005): The polymer systems, the polyolefinstogether with the waxes, are semi-crystalline materialsconsisting of crystalline and amorphous phases. If the hydro-carbon chains are long enough, they will fold in on them-selves to form crystallites (10–20 nm). The crystallitesare held together by amorphous chain segments thatcontribute to the elastic strength of the material. If thehydrocarbon chain length is shortened sufficiently, the am-orphous chain segments become less able to bridgethe gap between the crystallites, and the material becomesbrittle. Gas diffusion can then take place at the interfacesbetween the crystallites.
Similar interactions were also demonstrated with Hor-deum wax. Longer chains bridged the amorphous zonebetween two adjacent layers of crystalline material, thuslinking the two layers (Reynhardt and Riederer, 1994).The relevance of tortuosity in cuticular waxes as an effec-tive way of reducing the permeabilities of plant cuticlesis discussed by Schreiber (2006).
A significant amount of shrinkage might accompanywax recrystallization due to the high degree of crystall-inity and the large density difference between the amorph-ous melt and the crystal phase. As the freezing points ofthe individual components in the wax differ significantly(Chaiko and Leyva, 2005), this shrinkage could lead toextensive cracking. Nanocomposite materials mixed withpolyethylene, however, displayed reduced embrittlementat high clay loadings, and the films were quite flexibleand free of gross structural defects. Within plant cuticlesthe cutin polymer can be viewed as the analogous elas-tomer ensuring flexibility and avoiding embrittlement.
The plant cuticle has to be flexible to follow deforma-tions caused either by fluctuations in turgor pressure orby wind stress. Petracek and Bukovac (1995) investigatedthe rheological properties of tomato fruit cuticles andidentified the cuticle as a viscoelastic polymer, com-prising both elastic (reversible) and plastic (irreversible)components. Furthermore, water affected the mechanicalproperties, and waxes reduced elasticity and susceptibilityto fracturing. The mechanical properties of cuticles arenot related to thickness but are likely to be altered bychemical composition (Wiedemann and Neinhuis, 1998).
Marga et al. (2001) specified that the chemical compositiondetermined both the biochemical properties as well asthe elastic or rigid behaviour.
Consequently, several structural principles describedfor synthetic polymers can be applied to the plant cuticle.The plant cuticle can be described as a matrix composedof highly ordered crystalline and disordered amorphousregions, within which the reduced dynamics in the crys-talline regions of the wax barrier make them practically in-accessible to permeants (Reynhardt and Riederer, 1991).
The cutin polymer not only serves as a matrix but alsocontributes to the transport characteristics of the cuticle(Santier and Chamel, 1998). Cutin monomers are cross-linked by ester, peroxide, and ether bridges (Kolattukudy,1984), and the presence of large amounts of di- andtri-hydroxyfatty acids may affect the degree of cross-linking. It was demonstrated that the maturation of cutinled to the addition of covalent bonds which resulted inless polarity and reduced permeability (Schmidt et al.,1981). The wax-free matrix membranes (MXs) of severalplant species showed a relatively wide range of solutemobilities (Table 1). The applicability of the UDOSmethod also provided evidence for the asymmetry of thecutin matrix. Chloroform–methanol extraction of CMto provide MX not only removes the impermeable waxcrystallites but also the amorphous waxes. Therefore, theelevated solute mobilities in MX are due to the reducedviscosity (absence of amorphous waxes) and the shorteneddiffusion path (lack of crystalline waxes causing tortuosity).
Solute size effects
Size dependence of mobility was quantified by plottingfor each plant species logk* versus molar volume (Vx)for the respective sets of model compounds. Characteristicmolar volumes were calculated according to McGowanand Mellors (1986). These molar volumes represent thevolumes at absolute zero temperature and they are closelycorrelated to van der Waals volumes (McGowan, 1969).
Solute mobilities in the CMs of numerous plant specieshave been measured using linear (Baur et al., 1996b) andcyclic organic model compounds varying in molar volumefrom 99 to 349 cm3 mol�1 (Bauer and Schonherr, 1992;Schonherr and Baur, 1994; Buchholz et al., 1998; Bauret al., 1999). Solute mobilities (k*) decreased exponent-ially with increasing molar volumes (Vx) and, with allplant species tested, good linear correlations betweenlogk* and Vx were found. This dependence could bedescribed with equations of the type
logk* = logk*0 � b9Vx ð3Þ
The y-intercepts (k*0) represent the mobility of a hypo-thetical molecule having zero molar volume and theslopes of the graphs (b9) represent the size selectivity of
Lipophilic pathway 2505
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

the barrier. The differences in size selectivity (b9) werenot significant but the y-intercepts ranged from �2.33(Populus) to �5.22 (Hedera) (Fig. 4). The latter are puremembrane parameters, which are completely indepen-dent of the properties of solutes (size and lipophilicity),and they can be related to the free volume available fordiffusion in cuticles and cuticular waxes. (The real conver-gence of this graph with the y-intercept is not known asdata describing the mobilities of non-electrolytes havingmolar volumes significantly below 100 cm3 mol�1 are stillunavailable.)
No significant differences were observed in b9 amongall the species tested, with the average value of b9 being0.0095 mol cm�3 at 25 8C. This equates to a reductionin solute mobility of 700 times if Vx is increased from100 to 400 cm3 mol�1. This size discrimination was foundto be the same when cuticular waxes were extracted andcomparable size selectivities were determined for poly-mer MXs (Baur et al., 1999). A similar correlationbetween diffusivity and solute size was also found forreconstituted waxes (Schreiber and Schonherr, 1993;Schreiber et al., 1996a; Schreiber, 2006).
Temperature
Size selectivity within plant cuticles is very temperaturesensitive. For example, in ivy cuticles, the difference insolute mobilities between IAA (Vx=130 cm3 mol�1) andtebuconazole (Vx=241 cm3 mol�1) amounts to factors of24.5, 10.5, and 2.2 at 25 8C, 35 8C, and 55 8C, respectively(Buchholz, 1998). Therefore, size selectivity diminishes atelevated temperatures and intensifies at low temperatures.
The benefit of this relationship for the estimation ofthe solute mobilities of new compounds at a given temper-ature and plant species has been impressively demonstrated(Buchholz et al., 1998). If the temperature sensitivity ofa plant species is known for only two compounds (i.e.having two respective Arrhenius graphs) then a graphof b9 versus temperature can be made. With this infor-mation, the size selectivity (b9) for any desired temperaturecan be derived and consequently the solute mobility forany third compound (Vx) is predictable.
An explanation for this (temperature-dependent) sizediscrimination can be derived from the free volumetheory (Coughlin et al., 1991; Duda and Zielinski, 1996).For a successful diffusion step within a polymer the pen-etrant needs a hole (a space between polymer chains) di-rectly adjacent to its actual position. Enough energy tomake the jump is ensured by the motion of the soluteitself, via Brownian motion. Hence it follows that onlythe movement of the adjacent polymer chains limits thesolute’s spatial motion. At a given temperature, a polymerhas a characteristic distribution of these temporary holes.Elevating the temperature causes an increase in the motionof the polymer chains which, in turn, produces a shift inthe hole-size distribution towards bigger holes. (Increasedenergy enables the formation of larger gaps between adja-cent polymer chains due to loosened bonds.) By contrast,the temporal abundance of small holes is relatively reducedat higher temperatures (Fig. 5).
An investigation of the response of solute mobility,which is independent of time, revealed a large increasewith rising temperature (Baur and Schonherr, 1995;Knoche and Bukovac, 2001; Schreiber, 2002). The tem-perature coefficients Q10 ranged from 3 (IAA in Prunus
Fig. 5. Exponential hole-size distribution of a polymer matrix asaffected by temperature (modified according to Lieb and Stein, 1972).(A) The distribution function provides the probability (P) of findinga free-volume hole of a specific size (v*) at a given temperature (T). v* isthe minimum volume hole size into which a molecule can jump. Thediffusion coefficient is considered to be proportional to the probabilityof finding a hole of volume v* or larger. (B) Increasing temperatureincreases the overall free volume available for migration (shaded area).
3
4
5
6
7
8
50 100 150 200 250 300 350 400
V x (cm
3 . mol
-1)
-lo
g k
*
Hedera helix
Malus domestica
Populus canescens
-log k * = 4.11 + 0.010 . Vx
-log k * = 5.22 + 0.009 . V x
-log k * = 2.33 + 0.011 . Vx
Fig. 4. The effect of molar volumes (Vx) of model compounds on theirmobility (k*) in cuticular membranes of three selected plant species.Averages of 16–20 cuticular membranes and 95% confidence intervalsare shown.
2506 Buchholz
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

armeniaca) to 14 (cholesterol in Hedera helix) at 20 8C(Baur et al., 1997a). The temperature effect can be quan-tified using the Arrhenius equation:
k* = k*03expð�ED=RTÞ ð4Þ
where k*0 is the pre-exponential factor, ED (kJ mol�1) isthe activation energy of diffusion, R (J mol�1 K�1) thegas constant, and T (K) the temperature. By plotting logk*(s�1) versus 1/T (K�1) the activation energy of diffusion(ED) can be calculated from the slope (b) of the graph:
ED = �13b3R32:3 ð5Þ
The Arrhenius graphs published were linear in the phy-siologically relevant temperature range of 10–45 8C, show-ing thatED was constant and that no distinct phase transitionscaused by sudden structural changes occurred. At temper-atures above 50 8C, a slight curvature of Arrhenius graphstowards lower ED occurred for some species. This was anadditional hint that cuticles could be classified as elasto-mers, since this flattening quite often occurs with rubberypolymers, as ED values reflect the onset of viscous flow(Schlotter and Furlan, 1992). Depending on species andsolute size, ED ranged from 60 to 232 kJ mol�1 (Baur et al.,1997a). This demonstrated that plant cuticles are veryeffective diffusion barriers.
The structural and functional integrity of cuticles canbe examined by measuring mobility after temperaturecycling. The comparison of subsequent experiments afterannealing can identify hysteresis effects due to re-arrangement of the wax constituents. Such experimentswere conducted at temperatures up to 70 8C (Baur et al.,1997a). Results were plant-species specific and rangedfrom superimposed graphs (Prunus laurocerasus) tothe straightening of previously curved graphs (Citrusaurantium), and in one case showed 10-fold increasedmobilities at 25 8C (Hedera helix). These different resultswere related to differences in wax composition causingdifferent melting behaviours and redistribution duringannealing.
The sonication of CMs increased solute mobility at25 8C by a factor of 4 and reduced the ED from123 kJ mol�1
to 92 kJ mol�1. Phase transitions or structural defectswere not indicated by the Arrhenius graphs (Fig. 6). Theinterpretation of these results remains speculative (e.g.loosening of chemical bonds), although a further observa-tion was that these effects diminished during storage(several months), and mobilities reverted to those of untreatedCMs. This indicated a re-equilibration between the waxcrystallites (‘healing of defects’) over time, as previously de-scribed for water transpiration (Geyer and Schonherr, 1990).
Based on the transition state theory (Glasstone et al.,1941) data can be plotted according to the thermodynamicrelationship between the pre-exponential factor (k*0) ofthe Arrhenius equation (proportional to entropy) and act-
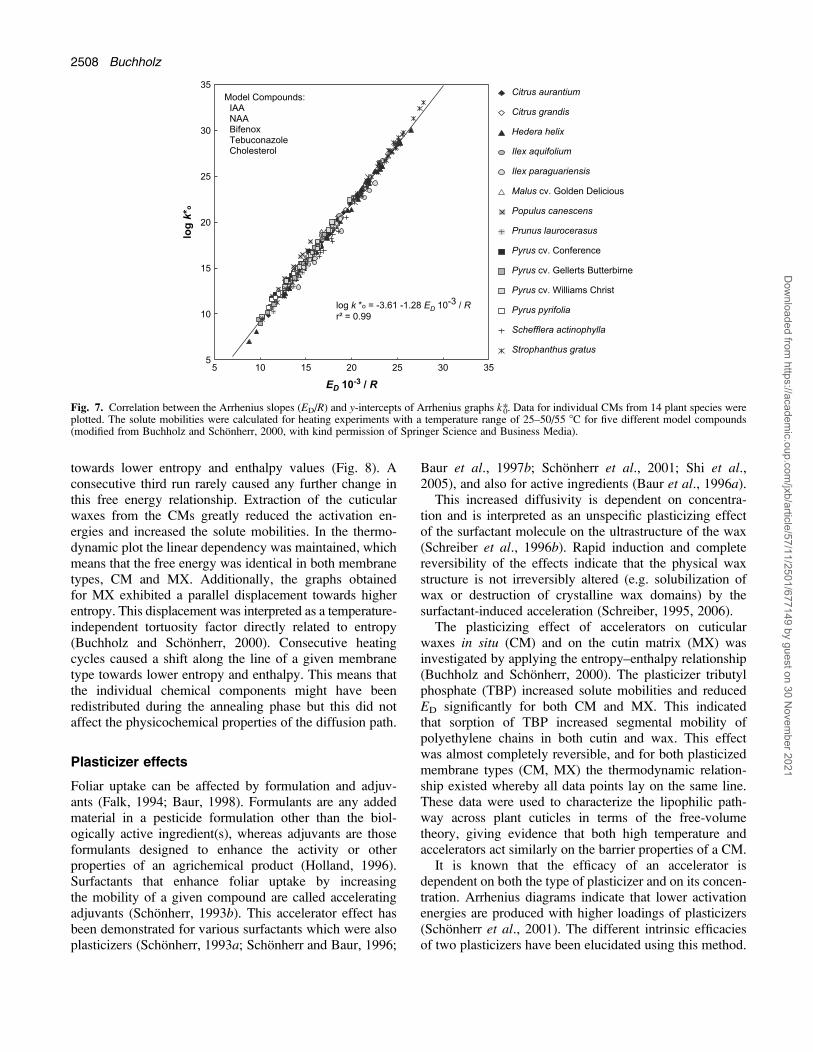
ivation energy (enthalpy) of diffusion (ED). A strict linearcorrelation was obtained for all of the CMs from 14 plantspecies and organic compounds having molar volumesranging from 130 to 349 cm3 mol�1 and cuticle/waterpartition coefficients of 18–108 (Fig. 7). This providedevidence that all of these compounds diffused—irrespec-tive of their size and lipophilicity—along the samelipophilic diffusion path. Each of the solutes must havetravelled through a microenvironment with identical phys-icochemical properties (Buchholz and Schonherr, 2000).
There are indications that water and lipophilic solutescan share the lipophilic path. This co-permeability wasalso confirmed by the simultaneous measurement of3H-labelled water and 14C-labelled organic compoundsacross plant cuticles (Niederl et al., 1998). Water can,however, take two parallel paths of diffusion: the lipo-philic path made of cutin and wax and the polar path com-posed of humidity-sensitive pores traversing the cuticles(Schreiber, 2002). Since wax extraction enhanced waterpermeability by several orders of magnitude (Schonherr,1982), whereas the effect of humidity on cuticular waterpermeability was only 2–3-fold, it is reasonable toargue that the largest fraction of water diffused acrossthe lipophilic paths in the cuticle (Schreiber et al., 2001;see also Table 2). The additional path predominated attemperatures higher than 30 8C and is likely to exhibita threshold in size and/or lipophilicity (Schreiber, 2002).
Arrhenius graphs obtained by repeat measurementson the same CM showed that, although the data pointsremained on the general line for CMs, they were shifted
60
Pyrus cv. ‚Gellerts B.‘ / Tebuconazole
T (°C)50 40 30 20
3.0 3.1 3.2 3.3 3.4
1/T.1000(K-1)
log
k*
-4.0
-3.5
-4.5
-5.0
-5.5
-6.0
-6.5
sonication
check
Fig. 6. Arrhenius graphs obtained for tebuconazole in CM of Pyrus(cv. Gellerts Butterbirne). One sample of CMs (n=10) was treatedwith sonication for 5 min in a water bath 1 d before experimental use,whereas the other sample (n=11) remained untreated.
Lipophilic pathway 2507
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021
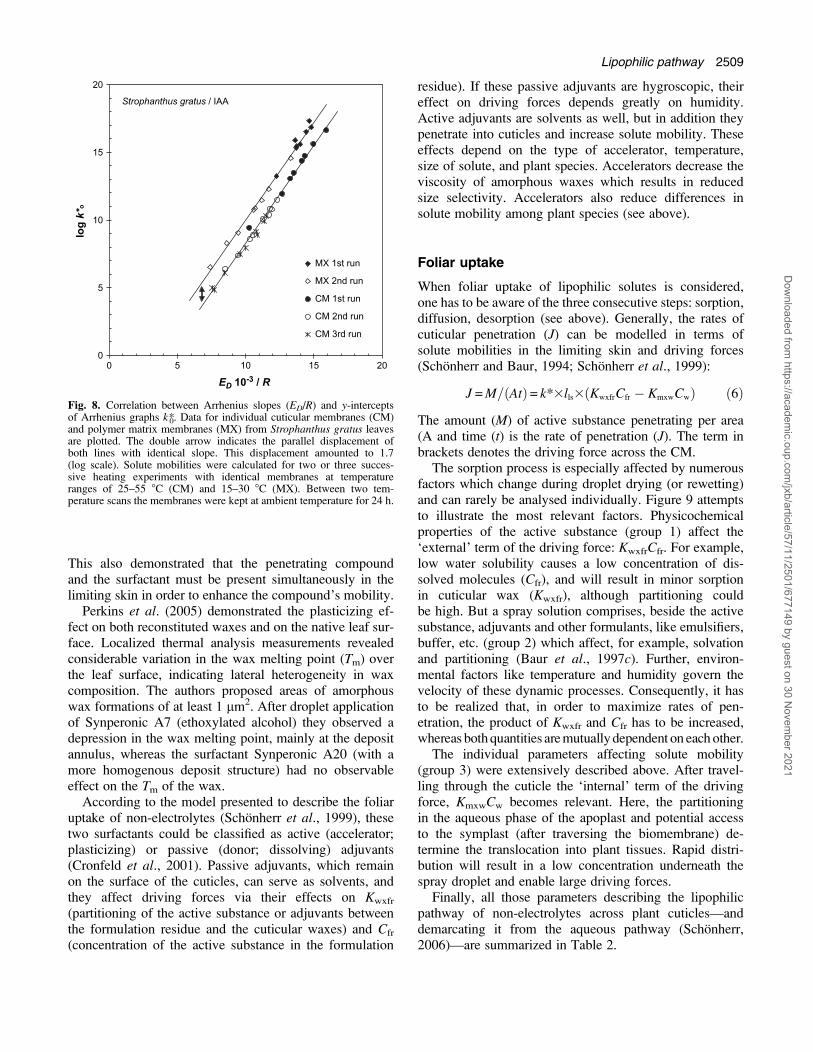
towards lower entropy and enthalpy values (Fig. 8). Aconsecutive third run rarely caused any further change inthis free energy relationship. Extraction of the cuticularwaxes from the CMs greatly reduced the activation en-ergies and increased the solute mobilities. In the thermo-dynamic plot the linear dependency was maintained, whichmeans that the free energy was identical in both membranetypes, CM and MX. Additionally, the graphs obtainedfor MX exhibited a parallel displacement towards higherentropy. This displacement was interpreted as a temperature-independent tortuosity factor directly related to entropy(Buchholz and Schonherr, 2000). Consecutive heatingcycles caused a shift along the line of a given membranetype towards lower entropy and enthalpy. This means thatthe individual chemical components might have beenredistributed during the annealing phase but this did notaffect the physicochemical properties of the diffusion path.
Plasticizer effects
Foliar uptake can be affected by formulation and adjuv-ants (Falk, 1994; Baur, 1998). Formulants are any addedmaterial in a pesticide formulation other than the biol-ogically active ingredient(s), whereas adjuvants are thoseformulants designed to enhance the activity or otherproperties of an agrichemical product (Holland, 1996).Surfactants that enhance foliar uptake by increasingthe mobility of a given compound are called acceleratingadjuvants (Schonherr, 1993b). This accelerator effect hasbeen demonstrated for various surfactants which were alsoplasticizers (Schonherr, 1993a; Schonherr and Baur, 1996;
Baur et al., 1997b; Schonherr et al., 2001; Shi et al.,2005), and also for active ingredients (Baur et al., 1996a).
This increased diffusivity is dependent on concentra-tion and is interpreted as an unspecific plasticizing effectof the surfactant molecule on the ultrastructure of the wax(Schreiber et al., 1996b). Rapid induction and completereversibility of the effects indicate that the physical waxstructure is not irreversibly altered (e.g. solubilization ofwax or destruction of crystalline wax domains) by thesurfactant-induced acceleration (Schreiber, 1995, 2006).
The plasticizing effect of accelerators on cuticularwaxes in situ (CM) and on the cutin matrix (MX) wasinvestigated by applying the entropy–enthalpy relationship(Buchholz and Schonherr, 2000). The plasticizer tributylphosphate (TBP) increased solute mobilities and reducedED significantly for both CM and MX. This indicatedthat sorption of TBP increased segmental mobility ofpolyethylene chains in both cutin and wax. This effectwas almost completely reversible, and for both plasticizedmembrane types (CM, MX) the thermodynamic relation-ship existed whereby all data points lay on the same line.These data were used to characterize the lipophilic path-way across plant cuticles in terms of the free-volumetheory, giving evidence that both high temperature andaccelerators act similarly on the barrier properties of a CM.
It is known that the efficacy of an accelerator isdependent on both the type of plasticizer and on its concen-tration. Arrhenius diagrams indicate that lower activationenergies are produced with higher loadings of plasticizers(Schonherr et al., 2001). The different intrinsic efficaciesof two plasticizers have been elucidated using this method.
5
10
15
20
25
30
35
5 10 15 20 25 30 35
log k *o = -3.61 -1.28 ED 10-3 / R
r² = 0.99
Model Compounds: IAA NAA Bifenox Tebuconazole Cholesterol
lo
g k
*o
ED 10
-3 / R
Citrus aurantium
Citrus grandis
Hedera helix
Ilex aquifolium
Ilex paraguariensis
Malus cv. Golden Delicious
Populus canescens
Prunus laurocerasus
Pyrus cv. Conference
Pyrus cv. Gellerts Butterbirne
Pyrus cv. Williams Christ
Schefflera actinophylla
Pyrus pyrifolia
Strophanthus gratus
Fig. 7. Correlation between the Arrhenius slopes (ED/R) and y-intercepts of Arrhenius graphs k*0. Data for individual CMs from 14 plant species wereplotted. The solute mobilities were calculated for heating experiments with a temperature range of 25–50/55 8C for five different model compounds(modified from Buchholz and Schonherr, 2000, with kind permission of Springer Science and Business Media).
2508 Buchholz
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021
This also demonstrated that the penetrating compoundand the surfactant must be present simultaneously in thelimiting skin in order to enhance the compound’s mobility.
Perkins et al. (2005) demonstrated the plasticizing ef-fect on both reconstituted waxes and on the native leaf sur-face. Localized thermal analysis measurements revealedconsiderable variation in the wax melting point (Tm) overthe leaf surface, indicating lateral heterogeneity in waxcomposition. The authors proposed areas of amorphouswax formations of at least 1 lm2. After droplet applicationof Synperonic A7 (ethoxylated alcohol) they observed adepression in the wax melting point, mainly at the depositannulus, whereas the surfactant Synperonic A20 (with amore homogenous deposit structure) had no observableeffect on the Tm of the wax.
According to the model presented to describe the foliaruptake of non-electrolytes (Schonherr et al., 1999), thesetwo surfactants could be classified as active (accelerator;plasticizing) or passive (donor; dissolving) adjuvants(Cronfeld et al., 2001). Passive adjuvants, which remainon the surface of the cuticles, can serve as solvents, andthey affect driving forces via their effects on Kwxfr
(partitioning of the active substance or adjuvants betweenthe formulation residue and the cuticular waxes) and Cfr
(concentration of the active substance in the formulation
residue). If these passive adjuvants are hygroscopic, theireffect on driving forces depends greatly on humidity.Active adjuvants are solvents as well, but in addition theypenetrate into cuticles and increase solute mobility. Theseeffects depend on the type of accelerator, temperature,size of solute, and plant species. Accelerators decrease theviscosity of amorphous waxes which results in reducedsize selectivity. Accelerators also reduce differences insolute mobility among plant species (see above).
Foliar uptake
When foliar uptake of lipophilic solutes is considered,one has to be aware of the three consecutive steps: sorption,diffusion, desorption (see above). Generally, the rates ofcuticular penetration (J) can be modelled in terms ofsolute mobilities in the limiting skin and driving forces(Schonherr and Baur, 1994; Schonherr et al., 1999):
J =M=ðAtÞ= k*3lls3ðKwxfrCfr � KmxwCwÞ ð6Þ
The amount (M) of active substance penetrating per area(A and time (t) is the rate of penetration (J). The term inbrackets denotes the driving force across the CM.
The sorption process is especially affected by numerousfactors which change during droplet drying (or rewetting)and can rarely be analysed individually. Figure 9 attemptsto illustrate the most relevant factors. Physicochemicalproperties of the active substance (group 1) affect the‘external’ term of the driving force: KwxfrCfr. For example,low water solubility causes a low concentration of dis-solved molecules (Cfr), and will result in minor sorptionin cuticular wax (Kwxfr), although partitioning couldbe high. But a spray solution comprises, beside the activesubstance, adjuvants and other formulants, like emulsifiers,buffer, etc. (group 2) which affect, for example, solvationand partitioning (Baur et al., 1997c). Further, environ-mental factors like temperature and humidity govern thevelocity of these dynamic processes. Consequently, it hasto be realized that, in order to maximize rates of pen-etration, the product of Kwxfr and Cfr has to be increased,whereas both quantities are mutually dependent on each other.
The individual parameters affecting solute mobility(group 3) were extensively described above. After travel-ling through the cuticle the ‘internal’ term of the drivingforce, KmxwCw becomes relevant. Here, the partitioningin the aqueous phase of the apoplast and potential accessto the symplast (after traversing the biomembrane) de-termine the translocation into plant tissues. Rapid distri-bution will result in a low concentration underneath thespray droplet and enable large driving forces.
Finally, all those parameters describing the lipophilicpathway of non-electrolytes across plant cuticles—anddemarcating it from the aqueous pathway (Schonherr,2006)—are summarized in Table 2.
0
5
10
15
20
0 5 10 15 20
MX 1st run
MX 2nd run
CM 1st run
CM 2nd run
CM 3rd run
Strophanthus gratus / IAA
ED 10
-3 / R
lo
g k*
o
Fig. 8. Correlation between Arrhenius slopes (ED/R) and y-interceptsof Arrhenius graphs k*0. Data for individual cuticular membranes (CM)and polymer matrix membranes (MX) from Strophanthus gratus leavesare plotted. The double arrow indicates the parallel displacement ofboth lines with identical slope. This displacement amounted to 1.7(log scale). Solute mobilities were calculated for two or three succes-sive heating experiments with identical membranes at temperatureranges of 25–55 8C (CM) and 15–30 8C (MX). Between two tem-perature scans the membranes were kept at ambient temperature for 24 h.
Lipophilic pathway 2509
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

Conclusions
It is the big advantage of the UDOS method that, withthis desorption measurement and the determination ofthe solute mobility k*, a pure descriptor of the diffusionprocess in CMs can be obtained. Only the feasibility ofdetermining sorption-independent mobilities in CMs(or diffusion coefficients in extracted wax; Schreiber,
2006) allowed the quantification and interpretation ofsize selectivity and tortuosity, as well as entropy/enthalpyrelationships. Therefore, the properties of the lipophilicpathway are now completely characterized. Only theinterpretation of the plant species-specific y-intercept ofa hypothetical molecule having zero molar volume, k*0,and its reference to tortuosity, remain to be explainedin more detail.
Fig. 9. Schematic drawing of a spray droplet and the underlying plant epidermis. Factors affecting cuticular penetration via the ‘lipophilic pathway’ aregrouped into (1) physicochemical properties of the active substance, (2) conditions determined by the environment and the spray solution, and (3)parameters affecting solute mobility, k*. J, Rate of cuticular penetration; lls, length of the diffusion path in the limiting skin ( 6¼ thickness); Kwxfr, partitioncoefficient wax/formulation residue; Kmxw, partition coefficient polymer matrix/water; Cfr, concentration of active substance in formulation residue;Cw, concentration of active substance in water (apoplast) (according to the model described by Schonherr et al., 1999).
Table 2. Comparison of the lipophilic and the aqueous pathways across plant cuticles
Lipophilic pathway Aqueous pathway
Plant Epicuticular waxes (Retention on leaf surface)No effect on permeability Limit access to aqueous pores
Intracuticular waxes Amorphous waxes determine viscosity Minor effect on penetrationCrystallites cause tortuosity andreduction in free volume
Cutin matrix Cross-linkage affects mobility Quality and spatial arrangement of polarconstituents determine abundance of aqueous pores
Substance(no plasticizer)
Molecular size Pronounced size selectivity Hydrate shells enforce weak size selectivity
Lipophilicity Affects driving force (sorption in waxes) –No effect on mobility
Point of deliquescence – Low POD reduces risk of crystallizedformulation residues
Concentration Effect on driving force Effect on driving forceMicroclimate Temperature Effect on fluidity, viscosity of waxes
and matrixMinor effect on penetration
Humidity Effect on driving force (solubility) Effect on driving force (solubility) andswelling of cuticle
Passive additives Surfactants (Retention; mass flow into open stomata)Effect on driving force (solvation, partitioning) Facilitation of access to aqueous pores
Minor effect on penetrationHumectants Effect on driving force (partitioning, solvation,
concentration dilution)Effect on driving force (solubility,concentration dilution)
Prevention of crystallization extends timefor penetration
Prevention of crystallization facilitatesaccess to aqueous pores and extends timefor penetration
Active additives Plasticizer Penetrate into cuticle and decrease viscosity(accelerator)
Negligible effect on penetration
Concentration dependent efficacyPlasticizer’s and substance’s mobility needa mutual fit
2510 Buchholz
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

This knowledge on factors affecting solute mobility(group 3 in Fig. 9) and the physicochemical properties ofa given non-polar active substance (group 1 in Fig. 9) arethe starting points for tailored crop protection products.
References
Baker EA. 1982. Chemistry and morphology of plant epicuticularwaxes. In: Cutler DF, Alvin KL, Price CE, eds. The plant cuticle.London: Academic Press, 139–166.
Baur P. 1998. Mechanistic aspects of foliar penetration of agro-chemicals and the effects of adjuvants. Recent Research Develop-ments in Agricultural and Food Chemistry 2, 809–837.
Baur P, Buchholz A, Schonherr J. 1997a. Diffusion in plantcuticles as affected by temperature and size of organic solutes:similarity and diversity among species. Plant, Cell and Environ-ment 20, 982–994.
BaurP,GraysonBT, Schonherr J. 1996a. Concentration-dependentmobility of chlorfenvinphos in isolated plant cuticles. PesticideScience 47, 171–180.
Baur P, Grayson BT, Schonherr J. 1997b. Polydisperse ethoxy-lated fatty alcohol surfactants as accelerators of cuticular penetra-tion. 1. Effects of ethoxy chain length and the size of thepenetrants. Pesticide Science 51, 131–152.
Baur P, Marzouk H, Schonherr J, Bauer H. 1996b. Mobilitiesof organic compounds in plant cuticles as affected by structureand molar volumes of chemicals and plant species. Planta 199,404–412.
Baur P, Marzouk H, Schonherr J, Grayson BT. 1997c. Partitioncoefficients between plant cuticle and adjuvants as related tofoliar uptake. Journal of Agricultural and Food Chemistry 45,3659–3665.
Baur P, Marzouk H, Schonherr J. 1999. Estimation of path lengthsfor diffusion of organic compounds through leaf cuticles. Plant,Cell and Environment 22, 291–299.
Bauer H, Schonherr J. 1992. Determination of mobilities of organiccompounds in plant cuticles and correlation with molar volumes.Pesticide Science 35, 1–11.
Baur P, Schonherr J. 1995. Temperature dependence of the dif-fusion of organic compounds across plant cuticles. Chemosphere30, 1331–1340.
Becker M, Kerstiens G, Schonherr J. 1986. Water permeabilityof plant cuticles: permeance, diffusion and partition coefficients.Trees 1, 54–60.
Bianchi G. 1995. Plant waxes. In: Hamilton RJ, ed. Waxes:chemistry, molecular biology and functions. Dundee: The OilyPress, 175–222.
Buchholz A. 1998. Großenselektivitat und Temperaturabhangigkeitder Diffusion organischer Verbindungen in der pflanzlichenKutikula. PhD thesis, University of Kiel, Germany.
Buchholz A, Baur P, Schonherr J. 1998. Differences among plantspecies in cuticular permeabilities and solute mobilities are notcaused by differential size selectivities. Planta 206, 322–328.
Buchholz A, Schonherr J. 2000. Thermodynamic analysis of dif-fusion of non-electrolytes across plant cuticles in the presence andabsence of the plasticiser tributyl phosphate. Planta 212, 103–111.
Bukovac MJ, Flore JA, Baker EA. 1979. Peach leaf surfaces:changes in wettability, retention, cuticular permeability, and epi-cuticular wax chemistry during expansion with special reference tospray application. Journal of the American Society for Horticul-tural Science 104, 611–617.
Carreto A, Almeida AR, Fernandes AC, Vaz WLC. 2002.Thermotropic mesotropism of a model system for the plantepicuticular wax layer. Biophysical Journal 82, 530–540.
Chaiko DJ, Leyva AA. 2005. Thermal transition and barrierproperties of olefinic nanocomposites. Chemical Materials 17,13–19.
Cronfeld P, Lader K, Baur P. 2001. Classification of adjuvantsand adjuvant blends by effects on cuticular penetration. ASTMSpecial Technical Publication STP 1400 (Pesticide Formulationsand Application Systems: 20th Volume), 81–94.
Coughlin CS, Mauritz KA, Storey RF. 1991. A general freevolume based theory for the diffusion of large molecules inamorphous polymers above Tg. 4. Polymer-penetrant interactions.Macromolecules 24, 1526–1534.
Cussler EL, Hughes SE, Ward WJ, Rutherford A. 1988. Barriermembranes. Journal of Membrane Science 38, 161–174.
Duda JL, Zielinski JM. 1996. Free-volume theory. In: Neogi, ed.Diffusion in polymers. New York, NY: Marcel Dekker, 143–171.
Falk RH. 1994. Influence of formulation and adjuvants on thefoliar location and physical form of the active ingredient. In:Holloway PJ, Rees RT, Stock D, eds. Interaction between ad-juvants, agrochemicals and target organisms. Berlin: Springer-Verlag, 53–82.
Geyer U, Schonherr J. 1990. The effect of the environment onthe permeability and composition of Citrus leaf cuticles. I.Water permeability of isolated cuticular membranes. Planta 180,147–153.
Glasstone S, Laidler K, Eyring H. 1941. The theory of rateprocesses. New York, NY: McGraw-Hill.
Haas K, Schonherr J. 1979. Composition of soluble cuticularlipids and water permeability of cuticular membranes from Citrusleaves. Planta 146, 399–403.
Hauke V, Schreiber L. 1998. Ontogenetic and seasonal develop-ment of wax composition and cuticular transpiration of ivy(Hedera helix) sun and shade leaves. Planta 207, 67–75.
Hoad SP, Jeffree CE, Grace J. 1992. Effects of wind and abrasionon cuticular integrity in Fagus sylvatica L. and consequencesfor transfer of pollutants through leaf surfaces. AgricultureEcosystems and Environment 42, 275–289.
Holland PT. 1996. Glossary of terms relating to pesticides(IUPAC Recommendations 1996). Pure and Applied Chemistry68, 1167–1193.
Holloway PJ. 1982. Structure and histochemistry of plant cuticularmembranes: an overview. In: Cutler DF, Alvin KL, Price CE, eds.The plant cuticle. London: Academic Press, 45–85.
Holloway PJ. 1993. Structure and chemistry of plant cuticles.Pesticide Science 37, 203–206.
Holloway PJ. 1994. Plant cuticles: physicochemical characteristicsand biosynthesis. In: Percy KE, Cape JN, Jagels R, Simpson CJ,eds. Air pollutants and the leaf cuticle. NATO ASI Series, SeriesG: Ecological Sciences 36, 1–13.
Jeffree CE. 1986. The cuticle, epicuticular waxes and trichomesof plants, with reference to their structure, functions and evolution.In: Juniper BE, Southwood TRE, eds. Insects and the plantsurface. London: Edward Arnold, 23–63.
Jeffree CE. 1996. Structure and ontogeny of plant cuticles. In:Kerstiens G, ed. Plant cuticles – an integrated functionalapproach. Oxford: BIOS Scientific Publishers, 33–75.
Jetter R, Schaffer S, Riederer M. 2000. Leaf cuticular waxesare arranged in chemically and mechanically distinct layers: evid-ence from Prunus laurocerasus L. Plant, Cell and Environment23, 619–628.
Juniper BE. 1995. Waxes on plant surfaces and their interactionswith insects. In: Hamilton RJ, ed. Waxes: chemistry, molecularbiology and functions. Dundee: The Oily Press, 157–176.
Kerstiens G. 1996. Signalling across the divide: a wider perspec-tive of cuticular structure: function relationships. Trends in PlantScience 1, 125–129.
Lipophilic pathway 2511
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

Kirkwood RC. 1999. Recent developments in our understandingof the plant cuticle as a barrier to the foliar uptake of pesticides.Pesticide Science 55, 69–77.
Kirsch T, Kaffarnik F, Riederer M, Schreiber L. 1997. Cuticularpermeability of the three tree speciesPrunus laurocerasusL.,Ginkgobiloba L., and Juglans regia L.: comparative investigation of thetransport properties of intact leaves, isolated cuticles, and reconstitutedcuticular waxes. Journal of Experimental Botany 48, 1035–1045.
Knoche M, Bukovac MJ. 2001. Finite dose diffusion studies. III.Effects of temperature, humidity and deposit manipulation onNAA penetration through isolated tomato fruit cuticles. PestManagement Science 57, 737–742.
Knoche M, Petracek PD, Bukovac MJ. 2000. Finite dose diffu-sion studies. I. Characterizing cuticular penetration in a modelsystem using NAA and isolated tomato fruit cuticles. Pest Man-agement Science 56, 1005–1015.
Kolattukudy PE. 1984. Biochemistry and function of cutin andsuberin. Canadian Journal of Botany 62, 2918–2933.
Kolattukudy PE, Rogers LM, Li D, Hwang C, Flaishman MA.1995. Surface signaling in pathogenesis. Proceeding of theNational Academy of Sciences, USA 92, 4080–4087.
Kruger H, van Rensburg L, Peacock J. 1996. Cuticular membranefine structure of Nicotiana tabacum L. leaves. Annals of Botany77, 11–16.
Leece DR. 1976. Composition and ultrastructure of leaf cuticlesfrom fruit trees, relative to differential foliar absorption.Australian Journal of Plant Physiology 3, 833–847.
Lieb WR, Stein WD. 1972. The molecular basis of simple diffusionwithin biological membranes. In: Bonner F, Kleinzeller A, eds.Current topics in membrane transport, Vol. 2. New York, NY:Academic Press, 1–39.
Marga F, Pesacreta TC, Hasenstein KH. 2001. Biochemcialanalysis of elastic and rigid cuticles of Cirsium horridulum. Planta213, 841–848.
McGowan JC. 1969. The effects of pressure and temperature onthe densities of liquid polymers. Polymer – the Science and Tech-nology of Polymers 10, 841–848.
McGowan JC, Mellors A. 1986. Molecular volumes in chemistryand biology: applications including partitioning and toxicity.Chichester: Ellis Horwood.
Merk S, Blume A, Riederer M. 1998. Phase behavior andcrystallinity of plant cuticular waxes studied by Fourier transforminfrared spectroscopy. Planta 204, 44–53.
Niederl S, Kirsch T, Riederer M, Schreiber L. 1998. Co-permeability of 3H-labeled water and 14C-labeled organic acidsacross isolated plant cuticles: investigating cuticular paths of dif-fusion and predicting cuticular transpiration. Plant Physiology116, 117–123.
Orgell WH. 1955. The isolation of plant cuticle with pectic enzymes.Plant Physiology 30, 78–80.
Perkins MC, Roberts CJ, Briggs D, Davies MC, Friedmann A,Hart C, Bell G. 2005. Macro- and microthermal analysis of plantwax/surfactant interactions: plasticizing effects of two alcoholethoxylated surfactants on an isolated cuticular wax and leafmodel. Applied Surface Science 243, 158–165.
Petracek PD, Bukovac MJ. 1995. Rheological properties ofenzymatically isolated tomato fruit cuticle. Plant Physiology109, 675–679.
Reynhardt EC, Riederer M. 1991. Structure and moleculardynamics of the cuticular wax from leaves of Citrus aurantiumL. Journal of Physics D: Applied Physics 24, 478–486.
Reynhardt EC, Riederer M. 1994. Structures and moleculardynamics of plant waxes. II. Cuticular waxes from the leaves ofFagus sylvatica L. and Hordeum vulgare L. European BiophysicsJournal 23, 59–70.
Riederer M, Schneider G. 1990. The effect of the environmenton the permeability and composition of Citrus leaf cuticles. II.Composition of soluble cuticular lipids and correlation withtransport properties. Planta 180, 154–165.
Riederer M, Schreiber L. 1995. Waxes: the transport barriers ofplant cuticles. In: Hamilton RJ, ed. Waxes: chemistry, molecularbiology and functions. Dundee: The Oily Press, 131–156.
Riederer M, Schreiber L. 2001. Protecting against water loss:analysis of the barrier properties of plant cuticles. Journal ofExperimental Botany 52, 2023–2032.
Santier S, Chamel A. 1998. Reassessment of the role of cuticularwaxes in the transfer of organic molecules through plant cuticles.Plant Physiology and Biochemistry 36, 225–231.
Schlotter NE, Furlan PY. 1992. A review of small moleculediffusion in polyolefins. Polymers 33, 3323–3342.
Schmidt HW, Merida T, Schonherr J. 1981. Water permeabilityand fine structure of cuticular membranes isolated enzymaticallyfrom leaves of Clivia miniata Reg. Zeitschrift fur Pflanzen-physiologie 105, 41–51.
Schonherr J. 1982. Resistance of plant surfaces to water loss:transport properties of cutin, suberin and associated lipids. In:Lange OL, Nobel PS, Osmond CB, Ziegler H, eds. Encyclopaediaof plant physiology, New Series, Vol. 12B. Berlin: Springer,153–179.
Schonherr J. 1993a. Effects of monodisperse alcohol ethoxylateson mobility of 2,4-D in isolated plant cuticles. Pesticide Science38, 155–164.
Schonherr J. 1993b. Effects of alcohols, glycols and monodisperseethoxylated alcohols on mobility of 2,4-D in isolated plant cuticles.Pesticide Science 39, 213–223.
Schonherr J. 2006. Characterization of aqueous pores in plantcuticles and permeation of ionic solutes. Journal of ExperimentalBotany 57, 2471–2491.
Schonherr J, Baur P. 1994. Modeling penetration of plant cuticlesby crop protection agents and effects of adjuvants on their ratesof penetration. Pesticide Science 42, 185–208.
Schonherr J, Baur P. 1996. Effects of temperature, surfactants andother adjuvants on rates of uptake of organic compounds. In:Kerstiens G, ed. Plant cuticles: an integrated functionalapproach. Oxford: BIOS Scientific Publishers, 134–154.
Schonherr J, Baur P, Buchholz A. 1999. Modelling foliarpenetration: its role in optimizing pesticide delivery. In: BrooksGT, Roberts TR, eds. Pesticide chemistry and bioscience, thefood-environment challenge. Cambridge: The Royal Society ofChemistry, 134–151.
Schonherr J, Riederer M. 1988. Desorption of chemicals fromplant cuticles: evidence for asymmetry. Archives of EnvironmentalContamination and Toxicology 17, 13–19.
Schonherr J, Schmidt HW. 1979. Water permeability of plantcuticles: dependence of permeability coefficients of cuticulartranspiration on vapor pressure saturation deficit. Planta 144,391–400.
Schonherr J, Schreiber L, Buchholz A. 2001. Effects of temper-ature and concentration of the accelerators ethoxylated alcohols,diethyl suberate and tributyl phosphate on the mobility of [14C]2,4-dichlorophenoxy butyric acid in plant cuticles. Pest Manage-ment Science 57, 17–24.
Schreiber L. 1995. A mechanistic approach towards surfactant/wax interactions: effects of octaethyleneglycolmonododecyletheron sorption and diffusion of organic chemicals in reconstitutedcuticular wax of barley leaves. Pesticide Science 45, 1–11.
Schreiber L. 2002. Co-permeability of 3H-labelled water and14C-labelled organic acids across isolated Prunus laurocerasuscuticles: effect of temperature on cuticular paths of diffusion.Plant, Cell and Environment 25, 1087–1094.
2512 Buchholz
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021

Schreiber L. 2006. Sorption and diffusion of lipophilic moleculesin cuticular wax and effects of accelerators on solute mobilities.Journal of Experimental Botany 57, 2515–2523.
Schreiber L, Kirsch T, Riederer M. 1996a. Transport properties ofcuticular waxes of Fagus sylvatica L. and Picea abies (L.) Karst.:estimation of size selectivity and tortuosity from diffusioncoefficients of aliphatic molecules. Planta 198, 104–109.
Schreiber L, Krimm U, Knoll D. 2004. Interactions betweenepiphyllic microorganisms and leaf cuticles. In: Varma A, AbbottL, Werner D, Hampp R, eds. Plant surface microbiology, 145–156.
Schreiber L, Riederer M. 1996. Determination of diffusioncoefficients of octadecanoic acid in isolated cuticular waxes andtheir relationship to cuticular water permeabilities. Plant, Celland Environment 19, 1075–1082.
Schreiber L, Riederer M, Schorn K. 1996b. Mobilities of organiccompounds in reconstituted cuticular wax of barley leaves: effectsof monodisperse alcohol ethoxylates on diffusion of pentachloro-phenol and tetracosanoic acid. Pesticide Science 48, 117–124.
Schreiber L, Schonherr J. 1993. Mobilities of organic compoundsin recrystallized cuticular wax of barley leaves: determination ofdiffusion coefficients. Pesticide Science 38, 353–361.
Schreiber L, Schorn K, Heimburg T. 1997. 2H NMR study ofcuticular wax isolated from Hordeum vulgare L. leaves: identifi-cation of amorphous and crystalline wax phases. European Bio-physics Journal 26, 371–380.
Schreiber L, Skrabs M, Hartmann K, Diamantopoulos P,Simanova E, Santrucek J. 2001. Effect of humidity on cuticulartranspiration of isolated cuticular membranes and leaf disks.Planta 214, 274–282.
Shafer WE, Schonherr J. 1985. Accumulation and transport ofphenol, 2-nitrophenol, and 4-nitrophenol in plant cuticles. Ecotox-icology and Environmental Safety 10, 239–252.
Shi T, Simanova E, Schonherr J, Schreiber L. 2005. Effectsof accelerators on mobility of 14C-2,4-dichlorophenoxy butyricacid in plant cuticles depends on type and concentration ofaccelerator. Journal of Agricultural and Food Chemistry 53,2207–2212.
Tamura H, Knoche M, Bukovac MJ. 2001. Evidence for surfactantsolubilization of plant epicuticular wax. Journal of Agriculturaland Food Chemistry 49, 1809–1816.
Turunen M, Huttunen S. 1990. A review of the response ofepicuticular wax of conifer needles to air pollution. Journal ofEnvironmental Quality 19, 35–45.
Viougeas MA, Rohr R, Chamel A. 1995. Structural changes andpermeability of ivy (Hedera helix L.) leaf cuticles in relation toleaf development and after selective chemical treatments. NewPhytologist 130, 337–348.
von Wettstein-Knowles P. 1995. Biosynthesis and genetics ofwaxes. In: Hamilton RJ, ed. Waxes: chemistry, molecular biologyand functions. Dundee: The Oily Press, 91–130.
Watanabe A. 1961. Synthesis and physical properties of normalhigher primary alcohols. IV. Thermal and X-ray studies on thepolymorphism of the alcohols of even carbon numbers fromdodecanol to tetratriacontanol. Bulletin of the Chemical Societyof Japan 34, 1728–1734.
Wattendorf J, Holloway PJ. 1980. Studies on the ultrastructureand histochemistry of plant cuticles: the cuticular membrane ofAgave americana L. in situ. Annals of Botany 46, 13–28.
Whitehouse P, Holloway PJ, Caseley JC. 1982. The epicuticularwax of wild oats in relation to foliar entry of the herbicidesdiclofop-methyl and difenzoquat. In: Cutler DF, Alvin KL, PriceCE, eds. The plant cuticle. Linnean Society Symposium Series 10.London: Academic Press, 315–330.
Wiedemann P, Neinhuis C. 1998. Biomechanics of isolated plantcuticles. Botanica Acta 111, 28–34.
Lipophilic pathway 2513
Dow
nloaded from https://academ
ic.oup.com/jxb/article/57/11/2501/677149 by guest on 30 N
ovember 2021