characterization of interaction between brevetoxin and its
TRANSCRIPT

Florida International UniversityFIU Digital Commons
FIU Electronic Theses and Dissertations University Graduate School
11-7-2016
Characterization of Interaction Between Brevetoxinand Its Native Receptor and Identification of theRole of Brevetoxin in Karenia brevisWei ChenFlorida International University, [email protected]
DOI: 10.25148/etd.FIDC001236Follow this and additional works at: https://digitalcommons.fiu.edu/etd
Part of the Biochemistry Commons, and the Marine Biology Commons
This work is brought to you for free and open access by the University Graduate School at FIU Digital Commons. It has been accepted for inclusion inFIU Electronic Theses and Dissertations by an authorized administrator of FIU Digital Commons. For more information, please contact [email protected].
Recommended CitationChen, Wei, "Characterization of Interaction Between Brevetoxin and Its Native Receptor and Identification of the Role of Brevetoxinin Karenia brevis" (2016). FIU Electronic Theses and Dissertations. 2988.https://digitalcommons.fiu.edu/etd/2988

FLORIDA INTERNATIONAL UNIVERSITY
Miami, Florida
CHARACTERIZATION OF INTERACTION BETWEEN BREVETOXIN AND ITS
NATIVE RECEPTOR AND IDENTIFICATION OF THE ROLE OF BREVETOXIN
IN KARENIA BREVIS
A dissertation submitted in partial fulfillment of
the requirements for the degree of
DOCTOR OF PHILOSOPHY
in
CHEMISTRY
by
Wei Chen
2016

ii
To: Dean Michael R. Heithaus College of Arts, Sciences and Education This dissertation, written by Wei Chen, and entitled Characterization of Interaction Between Brevetoxin and Its Native Receptor and Identification of the Role of Brevetoxin in Karenia brevis, having been approved in respect to style and intellectual content, is referred to you for judgment. We have read this dissertation and recommend that it be approved.
_______________________________________
Yuan Liu
_______________________________________ Xiaotang Wang
_______________________________________
Jose M. Eirin-Lopez
_______________________________________ Rudolf Jaffe
_______________________________________
Kathleen Rein, Major Professor
Date of Defense: November 7, 2016 The dissertation of Wei Chen is approved.
_______________________________________ Dean Michael R. Heithaus
College of Arts, Sciences and Education
_______________________________________ Andrés G. Gil
Vice President for Research and Economic Development and Dean of the University Graduate School
Florida International University, 2016

iii
© Copyright 2016 by Wei Chen
All rights reserved.

iv
DEDICATION
I dedicate this work to my parents, my grandparents, my boyfriend and his family for
their endless love and support. Without their understanding and encouragement, the
completion of this work would not have been possible.

v
ACKNOWLEDGMENTS
I am really grateful to the numerous people who help me a lot in the pursuit of this
Ph.D degree. Without their guidance and support the accomplishment of this
dissertation would not have been possible. It is my great pleasure to express my
gratitude to all of them in my humble acknowledgment
First and foremost, I would like to thank my major professor Dr. Kathleen Rein for
the opportunity to study and work in her group to pursue a doctoral degree. Without
her endless support, patience and supervision, my skill of writing and presenting
could not be improved during entire degree process. I do appreciate everything she
has done for encouraging me to overcome lots of obstructions during the past five
years.
I would also like to thank my other committee members, Dr. Yuan Liu, Dr. Xiaotang
Wang, Dr. Rudolf Jaffe and Dr. Jose M. Eirin-Lopez for their suggestion, guidance
and valuable time througout the years.
I would like to acknowledge all the faculty and staff members of the chemistry
department for the support, help and coordination. Big thanks to the dean’s office of
the College of Arts, Sciences and Education for travel funding.
I would like to thank my lab mates for their support and friendship. Because of you,
my five years are full with love and happiness. Special thanks to Li Liu, Ryan Cassell
Pengfei Sun and Freddy Rodriguez for their help in my study.
Finally, I wish to express my appreciation to those who have helped me in any respect
during my study at FIU, as well as expressing my apology that I could not mention
personally one by one.

vi
ABSTRACT OF THE DISSERTATION
CHARACTERIZATION OF INTERACTION BETWEEN BREVETOXIN AND ITS
NATIVE RECEPTOR AND IDENTIFICATION THE ROLE OF BREVETOXIN IN
KARENIA BREVIS by
Wei Chen
Florida International University, 2016
Miami, Florida
Professor Kathleen Rein, Major Professor
Algae are important to marine and fresh-water ecosystems. However, some species
of algae are harmful or even toxic. They can consume oxygen or block sunlight that
is essential for other organisms to live. Indeed, some algae blooms can produce
toxins that damage the health of the environment, plants, animals, and humans.
Harmful algal blooms (HABs) which are often more green, brown, or dark-colored
than red have spread along the coastlines and in the surface waters of the United
States. Therefore, scientists are making great efforts to study HABs in order to
maintain human and ecosystem health.
Karenia brevis, the major harmful algal bloom dinoflagellate of the Gulf of Mexico,
plays a destructive role in the region. Karenia brevis, responsible for Florida red tide,
is the principle HAB dinoflagellate in the Gulf of Mexico. K. brevis blooms can
produce brevetoxin: ladder-shaped polyether (LSP) compounds, which can lead to
adverse human health effects, like reduced respiratory function through inhalation
exposure, or neurotoxic shellfish poisoning through consumption of contaminated
shellfish. The poisoning has been attributed to their affinity for voltage-sensitive
sodium ion channels causing channel opening and depolarization of excitable cell

vii
membranes. Conservative estimate suggests that the economic impact from all
harmful algal bloom events in the United States is at least $82 million/year. The
public health costs occupy $37 million alone.
The study presented herein utilized fluorescent and photolabile brevetoxin probes to
demonstrate that brevetoxin localizes in the chloroplast of K. brevis where it binds
to light harvest complex II (LHC II) and thioredoxin (Trx). It had been discovered
that the TrxR/Trx system was inhibited by brevetoxin-2 (PbTx-2) with an IC50 of 25
µM. The mechanism of the inhibition was discussed in this work. The research also
revealed that the K. brevis high-toxic and low-toxic strains have a significant
difference in their ability, not only to produce brevetoxin, but also to perform NPQ
and in the production of ROS. I compared and contrasted various metabolic and
biochemical parameters in two strains of K. brevis which had a ten-fold difference in
toxin content. The work could shed light on the physiological role that brevetoxin
fills for K. brevis and may contribute to understanding the effect of ladder-shaped
polyether compounds on both marine animals and exposed humans and shall inform
improved treatments for brevetoxicosis.

viii
TABLE OF CONTENTS
CHAPTER PAGE
Chapter 1 Introduction ................................................................................................... 1 1.1 Harmful algal blooms (HABs) ................................................................................ 1 1.2 Karenia brevis ( K. brevis ) ..................................................................................... 4
1.2.1 The historical records of K. brevis ............................................................... 5 1.2.2 The historical effects of K. brevis ................................................................ 6
1.3 Brevetoxins and related ladder-shaped polyether (LSP) compounds ...................... 7 1.4 Biological activities of LSPs ................................................................................. 12
1.4.1 Biological activities of brevetoxin on animal ............................................ 12 1.4.2 Biological effects of brevetoxin on human ................................................ 15 1.4.3 The effect of brevetoxin on cellular process .............................................. 16
1.5 LSP compounds bind to transmembrane protein .................................................. 18 1.6 The interaction between PbTxs VGSC at the molecular level .............................. 20 1.7 The effect of brevetoxin on economy .................................................................... 23 1.8 Brevetoxins play an important role in K. brevis, but it is still elusive .................. 24 1.9 The significance of studying the dinoflagellates ................................................... 26 1.10 Research objectives ............................................................................................. 26
Chapter 2 Localization of PbTxs to a subcellular organelle and identification of a target protein from K. brevis ........................................................................................ 28
2.1 Localize PbTxs to one or more organelles or cellular structures in K.brevis ........ 28 2.1.1 Objective .................................................................................................... 28 2.1.2 Introduction ................................................................................................ 28 2.1.3 Results and discussion ............................................................................... 31
2.2 Isolation of a target protein from K. brevis as PbTxs natural receptor .................. 33 2.2.1 Objective .................................................................................................... 33 2.2.2 Introduction ................................................................................................ 33 2.2.3 Results and Discussions ............................................................................. 36 2.2.4 Conclusion ................................................................................................. 42
2.3 Materials and Method ............................................................................................ 44 2.3.1 Fluorescent probe experiment .................................................................... 44 2.3.2 Photoaffinity probe experiment ................................................................. 45
Chapter 3 The effects of brevetoxins on the thioredoxin reductase/thioredoxin system and related enzymes ..................................................................................................... 50
3.1 Objective ............................................................................................................... 50 3.2 Introduction ........................................................................................................... 50 3.3 Results ................................................................................................................... 59
3.3.1 Modified papain assay ............................................................................... 59 3.3.2 Papain assay ............................................................................................... 60 3.3.3 Fluorescent assay for Trx/TrxR system ..................................................... 64 3.3.4 Ellman’s assay ........................................................................................... 66 3.3.5 Adduct formation between PbTx-2 and selenocysteine ............................. 72 3.3.6 Adduct formation between PbTx-2 and TrxR ........................................... 73 3.3.7 Reaction of TrxR with Sel-green probe ..................................................... 75 3.3.8 Grx, GPx fluorescent assay ........................................................................ 76

ix
3.4 Conclusion ............................................................................................................. 78 3.5 Materials and Methods .......................................................................................... 80
3.5.1 Modified papain assay ............................................................................... 80 3.5.2 Papain assay ............................................................................................... 81 3.5.3 Fluorescent assay for Trx/TrxR system ..................................................... 82 3.5.4 Ellman’s assay ........................................................................................... 83 3.5.5 The preparation of selenocysteine -PbTx-2 adduct ................................... 84 3.5.6 The preparation of TrxR-PbTx-2 adduct. .................................................. 84 3.5.7 Reaction of Sel-green probe with TrxR ..................................................... 85 3.5.8 Fluorescent assay for Grx and GPx ........................................................... 86
Chapter 4 Comparison of biochemical parameters in high toxin and low toxin producing strains of K. brevis ....................................................................................... 89
4.1 Objective ................................................................................................................. 89 4.2 Introduction ............................................................................................................. 89 4.3 Results and discussion ............................................................................................. 98 4.3.1 The antioxidants in two strains of K. brevis .............................................. 99
4.3.2 Redox state of protein and non-protein thiols in low vs high toxin cultures of K. brevis .......................................................................................... 102
4.3.3 The reaction of Sel-green probe in two K. brevis strains. ........................ 105 4.3.4 The activity of Trx and Grx in two K. brevis strains ............................... 106
4.4 Conclusion ............................................................................................................. 107 4.5 Materials and Methods .......................................................................................... 110
4.5.1 Culture methods ....................................................................................... 110 4.5.2 Preparation of K. brevis homogenate ....................................................... 110 4.5.3 ABTS assay .............................................................................................. 110 4.5.4 Modified papain assay ............................................................................. 111 4.5.5 Ellman’s assay ......................................................................................... 112 4.5.6 Sel- green reacts with homogenate of tow K. brevis strains .................... 113 4.5.7 Thioredoxin activity in homogenate of two strains K. brevis .................. 114 4.5.8 Grx activity in homogenate of two strains K. brevis ............................... 115
Chapter 5 Summary and future work ........................................................................... 116
5.1 Summary ............................................................................................................... 116 5.2 Future work ........................................................................................................... 117
5.2.1 Identify all biological receptors for brevetoxin in K. brevis and localize the binding site on native receptors ................................................................. 117 5.2.2 Analysis of the essential proteins, carotenoids and lipids for NPQ in low vs high toxin K. brevis ............................................................................... 119 5.2.3 Determination the role of brevetoxin in two K. brevis strains ................ 120
References .................................................................................................................... 121
VITA ............................................................................................................................ 144

x
LIST OF TABLE
TABLE PAGE
Table 1 Human poisoning syndromes caused by HAB ................................................. 4
Table 2 The peptide sequence of 27 kD band and the proteins with matching sequences. .................................................................................................................... 37 Table 3 The intensity of all bands on SDS-PAGE gel ................................................. 41
Table 4 The peptide sequence of the 12 kDa band the proteins with matching sequences ..................................................................................................................... 41 Table 5 The motifs in thioredoxin , glutaredoxin, PDI and DsbA .............................. 51
Table 6 Relative V0 for the assays shown above in Figure 24,26 and 27 .................... 63
Table 7 The inhibition % for the assays shown above in Fig 24,26 and 27 ................ 64
Table 8 The predicted Se isotopic ratio ....................................................................... 73
Table 9 The major ROS molecules and their metabolism ........................................... 90
Table 10 The alignments of TrxR in K. brevis, Homo sapiens and Emiliania huxleyi ......................................................................................................... 99 Table 11 Comparison of NPQ and ROS in two K. brevis strains .............................. 108
Table 12 Comparison of various biochemical parameters in two K. brevis strains at 19th days ................................................................................................................. 108

xi
LIST OF FIGURES
FIGURE PAGE
Figure 1 Karenia brevis ................................................................................................. 5
Figure 2 The structure of brevetoxin A .......................................................................... 8
Figure 3 The structure of brevetoxin B .......................................................................... 8
Figure 4 The structure of ciguatoxin B .......................................................................... 8
Figure 5 The structure of yessotoxin ............................................................................. 8
Figure 6 Structures of small polyether ladder toxins produced by ................................ 9
Figure 7 Synthesis of fluorescent brevetoxin probe .................................................... 31
Figure 8 The photograph of localization of brevetoxin in K. brevis live cells ............ 32
Figure 9 The structure of okadaic acid ........................................................................ 33
Figure 10 The most reliable and common high efficiency labeling of target proteins in photoaffinity probe ..................................................................................... 34 Figure 11 The synthesis of brevetoxin photoaffinity probe ......................................... 36
Figure 12 SDS-PAGE of protein fractions from photoaffinity experiments ............... 37
Figure 13 Xanthophyll cycle ........................................................................................ 39
Figure 14 Spinach homogenate was incubated with the photoaffinity probe in different pH buffer ....................................................................................................... 41 Figure 15 Trx catalyzes the reaction as a nucleophilic substitution reaction (SN2) ..... 52
Figure 16 Electron flow of E. coli TrxR/Trx system ................................................... 53
Figure 17 Reaction mechanism of TGR ...................................................................... 55
Figure 18 Electron flow of mammalian TrxR/Trx system ........................................... 56
Figure 19 The Trx/TrxR system plays a key role in biochemical processes associated with many human diseases ......................................................................... 57 Figure 20 Electron flow of mammalian GSH/GSSG system ....................................... 58
Figure 21 The redox cycle of GPx coupled with mammalian GSH/GSSG system ..... 59

xii
Figure 22 The equation of modified papain assay ....................................................... 59
Figure 23 The principle of modified papain assay to determine Trx activity .............. 60
Figure 24 The modified papain assay with / without PbTx-2 ...................................... 61
Figure 25 The modified papain assay with / without PbTx-2 but without cystamine. 62
Figure 26 The modified papain assay with / without PbTx-3 with cystamine ............ 62
Figure 27 The papain assay with / without brevetoxin ................................................ 63
Figure 28 The reaction in sensitive fluorescent assay for Trx/TrxR system ............... 64
Figure 29 The effect of PbTx-2 on Trx/TrxR system .................................................. 65
Figure 30 The effect of PbTx-3 on Trx/TrxR system .................................................. 66
Figure 31 The reduction of Ellman’s reagent .............................................................. 66
Figure 32 The effect of PbTx-2 on the Ellman’s assay ............................................... 67
Figure 33 The electron flow in C-terminal reaction center in mammalian TrxR ........ 68
Figure 34 The Michael acceptors demonstrated to inhibit TrxR ................................. 69
Figure 35 The hypothesized explanation for the observation in fluorescent Trx/TrxR assay with PbTx-2 ....................................................................................... 70 Figure 36 The hypothesized explanation for the observation in DTNB assay with TrxR and PbTx-2 ......................................................................................................... 70 Figure 37 The effect of truncated recombinant human ................................................ 71
Figure 38 The effect of PbTx-3 on DTNB reduction by mammalian TrxR ................ 72
Figure 39 The mass spectrum of adduct formation between PbTx-2 with selenocysteine .............................................................................................................. 73 Figure 40 MS/MS analysis of the peptides from rat liver TrxR .................................. 74
Figure 41 MS/MS analysis of the peptides from the adduct between PbTx-2 and TrxR ...................................................................................................................... 74 Figure 42 Reaction of Sel-Green probe with selenocysteine ....................................... 75
Figure 43 Release of fluorescent reporter from Sel-green probe upon incubation with TrxR in the presence and absence of PbTx-2 compared to standards of L-selenocysteine .............................................................................................................. 76

xiii
Figure 44 The principle of sensitive fluorescent assay for activity of Grx .................. 77
Figure 45 The reaction in sensitive fluorescent assay for GPx system ........................ 77
Figure 46 The effect of Grx activity by PbTx-2. ......................................................... 77
Figure 47 The effect of GPx activity by PbTx-2 ......................................................... 78
Figure 48 The common pathways to produce ROS ..................................................... 93
Figure 49 Major physiological function of the mammalian thioredoxin system ......... 95
Figure 50 The structure of carboxy-H2DCFDA ........................................................... 96
Figure 51 The ROS production in two strains of K. brevis ......................................... 96
Figure 52 NPQs for the low and high toxic algal cultures to the pulses of four different excitation wavelengths .................................................................................. 97 Figure 53 The reaction in ABTS assay ...................................................................... 100
Figure 54 The antioxidant capacity in two strains of K. brevis ................................. 101
Figure 55 The total amount of ascorbic acid in two stains of K. brevis .................... 101
Figure 56 The detection of thiols in K. brevis homogenate by modified papain assay ............................................................................................................... 103 Figure 57 The concentration of cells over the lifetime of high toxin and low toxic strains ......................................................................................................... 103 Figure 58 The redox state of high toxic and low toxic strains. .................................. 105
Figure 59 The totally glutathione in two strains of K. brevis and the ratio of native reduced glutathione to total glutathione in two strains of K. brevis .......................... 105 Figure 60 The fluorescence is released by the reaction of sel-green probe with thiol groups and selenol groups in K. brevis homogenate ......................................... 106 Figure 61 Using the fluorescent Trx / TrxR system kit detects the Trx activity in K. brevis homogenate ................................................................................................. 107 Figure 62 Using the fluorescent Grx kit detects the Grx activity in K. brevis homogenate ................................................................................................................ 107 Figure 63 Spinach homogenate was incubated with photoaffinity probe in PBS for 1 hour .................................................................................................................... 118

xiv
Figure 64 A-ring derivative from PbTx-2 or PbTx-3 A-ring diol ............................. 118

xv
LIST OF ABBREVIATIONS
AAO ascorbic acid oxidase
ABTS 2,2'-azino-bis(3-ethylbenzothiazoline-6-sulphonic acid
Abs absorbance
ASP amnesic shellfish poisoning
ATP adenosine triphosphate
AZ tetrafluorophenyl azides
AZP azaspiracid poisoning
BP benzophenone
BSA bovine serum albumin
CFP ciguatera fish poisoning
CSD Cambridge Structural Database
Cys cysteine
Chl chlorophyll
1Chl* singlet chlorophyll
3Chl* triplet chlorophyll
DAPI 4, 6-diamidino-2-phenylin-dole)
D dinoxanthin
DD diadinoxanthin
DIA trifluoromethylphenyl diazirines
DMF dimethylfomamide
DNase deoxyribonuclease
DNA deoxyribonucleic acid
DSP diarrhefic shellfish poisoning
DTNB 5, 5’-dithio-bis-(2-nitrobenzoic acid)

xvi
DTT dithiothreitol
E. coli Emilliania huxlei
EDTA ethylenediaminetetraacetic acid
FAD flavin adenine dinucleotide
FITC fluorescein isothiocyanate
FT-ICR fourier transform ion cyclotron resonance mass spectrometer
FTR ferredoxin thioredoxin reductase
GpA glycophorin A
Gpx glutathione peroxidase
GR glutathione reductase
Grx glutaredoxin
GSH glutathione
GSSG glutathione disulfide
H2O2 hydrogen peroxide
hTrxR human thioredoxin reductase
IAC iodoacetamide
ICW Intracoastal Waterway
IC50 half maximal inhibitory concentration
IPCC Intergovernmental Panel on Climate Change
K. brevis K. brevis
kDa kilodalton
Ki the concentration to produce half maximum inhibition
Km the substrate concentration to produce half maxium
velocity

xvii
LipDH lipoarmide dehydrogenase
LSP ladder shape polyether
mRNA messenger ribonucleic acid
MW molecular weight
NADPH nicotinamide adenine dinucleotide phosphate
NCBI National Center for Biotechnology information
NMR nuclear magentic resonance spectroscopy
NO nitric oxide
NPQ non-photochemical quenching
NSP neurotoxic shellfish poisoning
NTRC NADPH dependent thioredoxin reductase
OA okadaic acid
O2- superoxide anion
. OH hydroxyl radical
PAL photoaffinity labeling
PAM pulsed amplitude modulation
PBS phosphate buffered saline
PbTx brevetoxin
PDI protein disulfide isomerase
PE polyether
Prx peroxiredoxins
PSP paralytic shellfish poisoning
ROS reactive oxygen species
SDS-PAGE sodium dodecyl sulfate polyacrylamide gel electrophoresis

xviii
Sec selenocysteine
SN2 nucleophilic substitution reaction
SNAr nucleophilic aromatic substitution
SOD superoxide dismutase
SPR surface plasmon resonance
TCEP tris (2-carboxyethyl) phosphine hydrochloride
TEAC trolox equivalent antioxidant capacity
TGR thioredoxin glutathione reductase
Tris Tris(hydroxymethyl)aminomethane
TNB 2-nitro-5-thiobenzoic acid
Trx thioredoxin
TrxR thioredoxin reductase
VDE violaxanthin de-epoxidase
VGSC voltage-gated sodium channel
YTX yessotoxin
ZE zeaxanthin epoxidase
2-Cys Prxs 2-cysteine peroxiredoxins
β bata
α alpha
0C degress Celsius
g gram
hr hour
l liter
µ micro
λ lamdba

xix
m milli
mol mole
min minute
% percentage

1
Chapter 1 Introduction
1.1 Harmful algal blooms (HABs)
Microscopic algae, or phytoplankton are common in coastal waters. Although
phytoplankton are small, they can grow explosively under certain conditions that are
not yet well understood, creating a phenomenon commonly known as algae blooms.
Algal blooms may occur in freshwater as well as in marine environments. During a
bloom, algae may become concentrated enough to turn the water red, brown or black.
These blooms are often referred to as red tides.
The algae reproduce rapidly when the nutrient levels increase above the normal
conditions. An intense bloom can cause harmful impacts on marine ecosystems. It can
consume oxygen or block sunlight that is essential for the survival of other organisms.
As a result, the concentrated algal blooms, which spread across the surface of the
ocean or freshwater, can inhibit the growth of other species (Baden, Bourdelais et al.
2005). Algal blooms may or may not occur seasonally, are often localized to specific
geographic regions, and may be triggered or exacerbated by environmental factors
(Baden, Bourdelais et al. 2005). These may be related to changes in species of algae’s
composition or other environmental cues. It has been hypothesized that eutrophication,
nutrient sources and some natural processes contribute to the occurrence of harmful
algae bloom, such as river runoff, pollution, changes in the temperature of the oceans
and so on. These all have been associated with blooms (Baden, Bourdelais et al. 2005).
Because of the nutrient rich environments, coastal regions are most severely impacted
by marine blooms. It has been suggested that blooms have increased in frequency and
duration along the coastlines and in the surface waters of the United States as a result

2
of human activities (Fleming, Backer et al. 2005). Many of the nutrients which
support algal growth may be found in certain fertilizers which are discharged into the
water as agricultural waste (Stumpf, Culver et al. 2003). Increased nitrogen (N) and
phosphorus (P) input resulting from human activities can stimulate bloom growth
along the coast (Harrison, Caraco et al. 2005, Seitzinger, Harrison et al. 2005,
Burkholder, Dickey et al. 2006, Bricker, Longstaff et al. 2008). In some cases, the
connections between algal blooms and eutrophication are clear and well documented.
For example, in the 1970s a reduction in red tide, which was proportional to mandated
pollution reductions, was observed in the Inland Sea of Japan for a decade (Takahashi,
Kawamura et al. 2009). Also, an increase in nutrient input in the northwestern Black
Sea in the 1980s, is correlated with an increase in algal blooms during the subsequent
two decades (Bodeanu, Moncheva et al. 1998). Over the past several decades, red tide
has occurred almost annually along the Gulf Coast of Florida, has spread along the
Texas and eastern Florida coasts, and occasionally along the coasts of Louisiana,
Mississippi, and Alabama (Thyng, Hetland et al. 2013). It is indicated that
eutrophication is responsible for the increasing of the red tide in Gulf of Florida
(Heisler, Glibert et al. 2008). However, the mechanisms of the stimulation of algal
bloom by nutrients are not always clear and remain controversial.
In addition to eutrophication, climate change is among the suspected factors for the
increased frequency of blooms. Global warming has led to the temperature rising on
the surface of ocean. Recently, the Intergovernmental Panel on Climate Change
(IPCC) verified that over the past 40 years, the ocean absorbed 90 percent of earth’s
net energy, resulting in nearly 4 oC increase in the ocean (Collins, Knutti et al. 2013).
Warmer water provides blooms a favorable environment to grow. As a result, a range

3
expansion of algal bloom has been observed under warm weather conditions.
However, debate on the ultimate cause of range expansions is ongoing (Smayda 2007,
Hallegraeff 2010).
Although algae are essential to marine and fresh-water ecosystems, some species of
algae are harmful or even toxic. Some algae can produce toxins that damage the
health of the environment, plants, animals, and humans. These algal blooms, which
can cause environmental or public health threats, are referred to as harmful algal
blooms (HABs) (Hallegraeff 1993). Some HABs are associated with the production
of algal toxins. Toxins are a form of secondary metabolite; or organic compounds that
are not essential to the normal growth or development of an organism. Secondary
metabolites are typically produced by one species or a few closely related species and
are generally believed to convey a selective advantage to the producing organism.
Toxins pose a serious threat to other species sharing the aquatic environment with the
algae. Animals, who consume contaminated seafood, sea grass, and contaminated
seawater or inhale contaminated aerosols can suffer severe poisoning often resulting
in mass mortalities of marine animals. For example, the historical records have shown
that Florida red tide bloom can cause massive mortalities of at least 100 species of
marine animals, such as shellfish, sea birds, manatees and dolphins (Martin and
Martin 1976). A number of human poisoning syndromes are caused by ingestion of
HAB related toxins in contaminated seafood (Table 1) (Anderson, Moore et al. 2015),
such as paralytic shellfish poisoning (PSP), neurotoxic shellfish poisoning (NSP),
amnesic shellfish poisoning (ASP), ciguatera fish poisoning (CFP) and azaspiracid
poisoning (AZP), which are caused by HAB related toxins (Anderson, Moore et al.
2015). An alternate exposure route resulting from human activities or wave action can
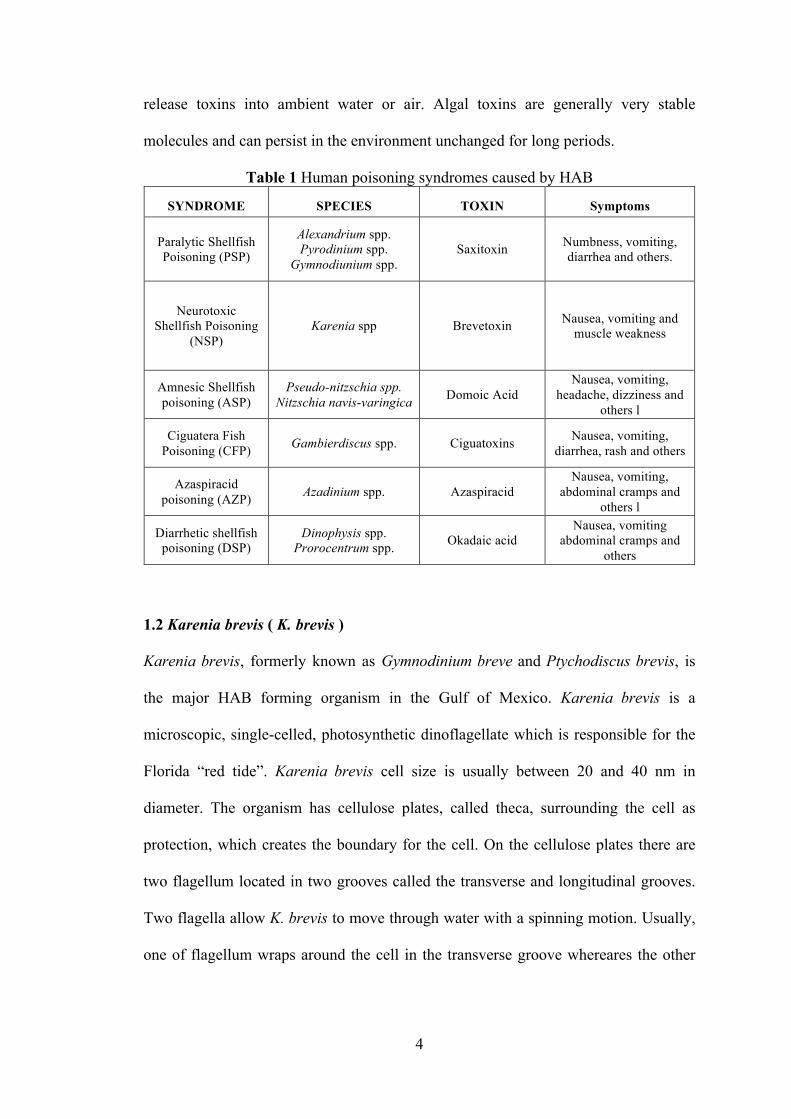
4
release toxins into ambient water or air. Algal toxins are generally very stable
molecules and can persist in the environment unchanged for long periods.
Table 1 Human poisoning syndromes caused by HAB
SYNDROME SPECIES TOXIN Symptoms
Paralytic Shellfish Poisoning (PSP)
Alexandrium spp. Pyrodinium spp.
Gymnodiunium spp. Saxitoxin Numbness, vomiting,
diarrhea and others.
Neurotoxic Shellfish Poisoning
(NSP) Karenia spp Brevetoxin Nausea, vomiting and
muscle weakness
Amnesic Shellfish poisoning (ASP)
Pseudo-nitzschia spp. Nitzschia navis-varingica Domoic Acid
Nausea, vomiting, headache, dizziness and
others l
Ciguatera Fish Poisoning (CFP) Gambierdiscus spp. Ciguatoxins Nausea, vomiting,
diarrhea, rash and others
Azaspiracid poisoning (AZP) Azadinium spp. Azaspiracid
Nausea, vomiting, abdominal cramps and
others l
Diarrhetic shellfish poisoning (DSP)
Dinophysis spp. Prorocentrum spp. Okadaic acid
Nausea, vomiting abdominal cramps and
others
1.2 Karenia brevis ( K. brevis )
Karenia brevis, formerly known as Gymnodinium breve and Ptychodiscus brevis, is
the major HAB forming organism in the Gulf of Mexico. Karenia brevis is a
microscopic, single-celled, photosynthetic dinoflagellate which is responsible for the
Florida “red tide”. Karenia brevis cell size is usually between 20 and 40 nm in
diameter. The organism has cellulose plates, called theca, surrounding the cell as
protection, which creates the boundary for the cell. On the cellulose plates there are
two flagellum located in two grooves called the transverse and longitudinal grooves.
Two flagella allow K. brevis to move through water with a spinning motion. Usually,
one of flagellum wraps around the cell in the transverse groove whereares the other

5
one extends on the longitudinal groove. Both the theca and flagella are visible in
Figure. 1. The nucleus is located in the lower part of the cell. Usually, becuase of the
presence of chloroplasts, K. brevis is yellow-green in color. Karenia brevis is an
obligate autotroph with facultative heterotrophy. It can gain energy from some
organic molecules and utilizes inorganic molecules such as nitrogen and phosphorus.
It also can produce energy from photosynthesis (Hitchcock, Kirkpatrick et al. 2010).
Figure 1 Karenia brevis. Image courtesy of Florida Fish and Wildlife Conservation Commission
Light is an essential requirement for K. breivs, thus it cannot survive at depths below
200 feet. Normally K. brevis is found on the surface of the water. The optimal
temperature range for K. brevis is 22-28 degrees Celsius. In addition, K. breivs can
live in a salinity range of 25-45 ppt. Under the appropriate conditions including
sufficient light and nutrients, K. brevis can concentrate from 100,000 to 250,000 cells/
liter within 2 months (Tester and Steidinger 1997). Blooms of K. brevis may also
provide a significant source of fixed carbon thorough photosynthesis (Vargo, Heil et
al. 2002).
1.2.1 The historical records of K. brevis
Karenia brevis blooms were first recorded centuries ago. According to historical
records, a red tide was first mentioned and documented in Gulf of Mexico in 1658
(Magaña, Contreras et al. 2003). It has been documented on the Florida west coast

6
since the 1800s. In the past decade, the Florida red tide blooms have been widespread
in the Gulf of Mexico, especially from Clearwater to Sanibel Island (Backer,
Kirkpatrick et al. 2005). Most commonly, K. brevis blooms can last more than two
months in regions of the Gulf of Mexico including: the west Florida shelf, the
Campeche Bay between Rio Ciatzacoalcos and Rio Grijalva, and Texas coast between
Port Arthur and Galveston Bay (Tester and Steidinger 1997). A bloom which
occurred in 2005 persisted for nearly one year (Hoagland, Jin et al. 2014).
Blooms of K. breveis result in detrimental effects to marine life and local economies
and as people living, working, and visiting the affected coastal areas (Backer,
Kirkpatrick et al. 2005). It is documented that cell densities as low as 103 cell/L will
cause respiratory irritation, whereas a concentration of more than 5 x 103 cells/L can
induce neurotoxic effects. Massive fish kills can be caused by densities over 105
cell/L, and cell densities over 106 cell/L result in water discoloration (Backer, Fleming
et al. 2003). An offical government record described the first reported fish kill in the
Gulf of Mexico in 1792 in the city of Veracruz, Mexico (Magaña, Contreras et al.
2003). It was reported that multitudes of dead fish washed onto the beaches of
Veracruz. The document also recored that same phenomenon occurred repeatedly in
that port.
1.2.2 The historical effects of K. brevis
It has been proposed that there are different effects which led to the massive fish and
marine animals killed by Florida red tide such as oxygen deprivation and air-born
aerosols.

7
Oxygen deprivation: Biochemical decomposition begins when the red tide organisms
die, which can cause oxygen level to decrease rapidly. In July 1971, a large spread of
red tide in Tampa Bay completely devastated the benthic ecosystem in the Bay.
Oxygen deprivation, rather than toxin, was one of the considerable contributors to the
death of creatures such as turtles, barnacles, birds and sponges (Simon and Dauer
1972).
Air-born aerosols: In the late 1800s massive fish kills along with human respiratory
irritation and dry cough in Veracruz, Mexico were reported when a K. brevis bloom
reached high concentration (Tester and Steidinger 1997, Magaña, Contreras et al.
2003). It is believed that highly concentrated K. brevis blooms played an essential role
in the poisoning of marine life. In 1996, at least 149 manatees died along the
southwest coast of Florida. According to lung pathology, a K. brevis produced toxin
was detected in the lung (Flewelling, Naar et al. 2005). It is highly likely that the
toxin had been inhaled by the manatees.
1.3 Brevetoxins and related ladder-shaped polyether (LSP) compounds
Brevetoxin are the parent of a small class of compounds called polyether ladders
which are classified as polyketides according to their biogenic origin (Lee, Qin et al.
1989). The best known LSP are brevetoxin A (PbTx-A) (Figure 2), brevetoxin B
(PbTx-B) (Figure 3), ciguatoxin (Figure 4) and yessotoxin (Figure 5).

8
Figure 2 The structure of brevetoxin A
Figure 3 The structure of brevetoxin B
Figure 4 The structure of ciguatoxin B
Figure 5 The structure of yessotoxin

9
Inspection of these structures reveals numerous common features: these compounds
consist of trans fused polyether rings with oxygen atoms that alternate from the
bottom to top of the molecule (Rein and Borrone 1999). In the past decades, there are
not only numerous polycyclic ethers were isolated from blooms or laboratory culture
of K. brevis, such as brevisamide, hemibrevetoxin and brevisin (Figure 6) (Satake,
Campbell et al. 2009) and brevenal (Bourdelais, Campbell et al. 2004).
Figure 6 Structures of small polyether ladder toxins produced by dinoflagellates Brevisin is a polycyclic ether that is consisted of two separate fused polyether rings
linked with a methylene group. Brevisin and brevenal are both natural competitors of
brevetoxin for its binding site (Bourdelais, Campbell et al. 2004, Satake, Campbell et
al. 2009). Those polyether compounds hold great interest among many scientific
groups because of their unique structure and enigmatic biosynthetic origin. However,
the first challenge was characterization of polyether compounds. The molecular
structure of brevetoxin B was first determined by Nakanishi’s group in 1981 (Lin,
Risk et al. 1981). The absolute stereochemistry of brevetoxin B was elucidated by X-

10
ray crystallography (Shimizu, Bando et al. 1986). The determination of the first
polyether ladder structure by X-ray crystallography simplified the subsequent
structure elucidation of other polyether ladders. The structure of brevisin was
determined by MS and 2D NMR (Satake, Campbell et al. 2009). The structure of the
even more complex polyether ladder, ciguatoxinn, which is isolated from
dinoflagellate G. toxicus was elucidated by NMR (Murata, Legrand et al. 1989,
Murata, Legrand et al. 1990).
The biosynthetic pathways for these natural polyether ladder products remain a puzzle.
As a result of the lack of enzymes or gene sequence for biosynthetic pathways, stable
isotope labeling experiments and structural features were used to develop the
biosynthetic principles. In 1985, Nakanishi proposed the currently held hypothesis for
the biosynthetic pathway of brevetoxin. It was hypothesized that these polyether
ladder compounds arise from the cyclization of a polyepoxide precursor (Nakanishi
1985).
Numerous individual brevetoxins (PbTxs) have been isolated and characterized from
K. brevis bloom as well as laboratory culture. Taken together, PbTxs are lipid-soluble
polyether compounds with molecular weights around 1000 D (Backer, Fleming et al.
2003; Backer, Kirkpatrick et al. 2005). There are two principal types of PbTxs that
differ according to the carbon skeleton; PbTx-A and PbTx-B. A flexible backbone of
10 fused polyether rings makes up PbTx-A which is the minor, but most toxic
congener. PbTx-B, the most abundant of the two, is comprised of a rigid backbone of
11 polyether rings (Nakanishi 1985, Baden 1989). Almost a dozen derivatives of
PbTx-A and PbTx-B occur naturally. Principal A-type brevetoxin include PbTx-1,

11
PbTx-7 and PbTx-10, while PbTx-2, PbTx-3, PbTx-8 and PbTx-9 belong to B-types
brevetoxins. Others, such as PbTx-8, a chloromethyl ketone derivative of PbTx-2,
appears to be an artifact of chloroform extraction of PbTx-2 (Nakanishi 1985, Matile,
Berova et al. 1996, Baden, Bourdelais et al. 2005, Murrell and Gibson 2009); In the
Florida red tide PbTx-2 is the major brevetoxin congener in bloom water, while
PbTx-3 is contained as greatest amount in aerosols (Pierce, Henry et al. 2005).
Furthermore, some polar brevetoxin derivatives were isolated from K. brevis cultures
and natural blooms as well. PbTx-1 and PbTx-2 both possess a lactone functionality
in the A- ring and share a common, rigid region in the terminal four rings with a side
chain allowing modification (Rein, Baden et al. 1994, Rein, Lynn et al. 1994). All
natural brevetoxins and derivatives have these features. However, in fact no derivative
is more toxic than PbTx-1 or PbTx-2 (Baden, Bourdelais et al. 2005). As identified by
LC/MS, some brevetoxin derivatives were confirmed as open A-ring of brevetoxin
PbTx-1, PbTx-7, PbTx-2 and PbTx-3. Some are oxidized from PbTx-1 and PbTx-2
(Abraham, Plakas et al. 2006).
The synthesis of LSPs remains a significant challenge. After twelve years of effort,
the first total synthesis of brevetoxin B was reported in 1995 (Nicolaou 1996,
Nicolaou, Yang et al. 1998). In the first generation strategy, hydroxyl epoxide
cyclization and the intramolecular conjugate addition were used as key reactions to
form the fused tetrahydropyran ring systems (Ring ABC, ring FG and ring IJK).
However, the step to thionate and bridge the last remaining bond required for
completion of the brevetoxin B skeleton was unsuccessful (Nicolaou, Hwang et al.
1995). It failed to construct the fused oxepane rings in brevetoxin B simultaneously.
Thus, stepwise bis (oxepane) synthesis approach was used to display the first strategy.

12
However, the goal to fuse additional rings onto the DEFG ring system is still elusive
(Nicolaou, Theodorakis et al. 1995). Finally, in his third strategy he successfully
formed the ring ABCDEF and ring IJK, leaving the didehydrooxocane ring H as final
ring to construct. In total, this process consists of 123 steps (Nicolaou, Rutjes et al.
1995). Nowadays, several groups have synthesized brevetoxin successfully. In 2004,
brevetoxin B was synthesized in 90 steps (Stephen and Kettle 1997, Matsuo,
Kawamura et al. 2004).
1.4 Biological activities of LSPs
The LSP compounds are responsible for a variety of some poisonings. Maitotoxin and
ciguatoxin both isolated from G. toxicus are associated with the CFP symptoms such
as chills, sweating, neurologic dysfunction and generalized weakness lasting from 10
min to 30 hours from consuming contained fish or food (Levine 1995). Yessotoxin
(YTX) which is produced by the dinoflagellate Protoceratium reticulatum,
accumulates in the gland of Japanese scallops (Murata, Kumagai et al. 1987, Satake,
MacKenzie et al. 1997). Yessotoxin associated lethality occurs through acute heart
disease. According to histopathology, mice exposed to yessotoxin, exhibited
intercytoplasmic edema (Satake, MacKenzie et al. 1997). Thus, cardiac muscle cells
are affected by yessotoxin.
1.4.1 Biological activities of brevetoxin on animal
Brevetoxins can persist in the ecosystem for up to eight months after the absence of
K. brevis blooms. For an instance, in 2007 K. brevis was first observed in early
September near Jacksonville, FL. Subsequently, it expands through the east coast of

13
Florida, including the Indian River Lagoon and the Atlantic Intracoastal Waterway
(ICW) (Walsh, Weisberg et al. 2009). The maximum concentration of K. brevis was
up to 106 cell / L in October. The bloom terminated in December 2007. However,
brevetoxin had been detected in sediments and seagrass epiphytes in July and August
2008 in ICW. The highest concentration of brevetoxin in sediments has been found
near Patrick Air Force Base at 89 ng per gram of dry sediment (Hitchcock,
Fourqurean et al. 2012). Average concentration is < 20 ng per gram of dry sediment
during non-bloom periods (Flewelling 2008). At the same time, brevetoxins were
detected in six of nine seagrass beds sampled at the south of the Mosquito Lagoon at
concentration of 6-18 ng per gram of dry epiphytes (Hitchcock, Fourqurean et al.
2012). In general, brevetoxins can persist almost one year in either seagrass epiphytes
or sediments after the termination of a bloom.
Since brevetoxins remain in seagrass and sediments for several months following
bloom termination, it seems highly likely that marine life can consume the brevetoxin
from contaminated seagrass or sediments. Subsequently, brevetoxins may be
transported through the food chain. In the other words, blooms of brevetoxin-
producing K. brevis have been associated with the mortality of fishery resources as
well as the death of marine mammals such as the endangered Florida manatees,
dolphins, turtles and shorebirds. The major route of exposure related to the deaths of
dolphins, manatees, turtles and seabirds is ingestion of PbTxs. For instance, some
shorebirds such as sanderlings (Calidris alba) and ruddy turnstones (Arenaria
interpres) scavenged dead fish deposited onshore during a K. brevis bloom. It is
reported that those shorebirds picked the eye orbit, gills and abdominal flesh or other
tissues that contained lethal levels of PbTxs for these small shorebirds (van Deventer,

14
Atwood et al. 2012). In 2006, more than 300 sea turtles stranded along the west coast
of Florida during prolonged blooms along the coast. Some strange behavios of sea
turtles had been observed, such as swimming in circles, lack of coordination, head
bobbing, muscle twitching and jerky body movement. High densities of brevetoxins
were detected in the organs or immune cells of sea turtles, which contributed to the
stranding or death (Walsh, Leggett et al. 2010). Also, in 1996, Florida red tide caused
a significant loss in the population of the highly endangered Florida manatee (Walsh,
Leggett et al. 2010). Thus fish and sea grass can be vectors in which high
concentrations of brevetoxins accumulate. Analysis of seagrass collected in manatee’s
stomach revealed the presence of brevetoxin (Flewelling, Naar et al. 2005). In both
bloom and non-bloom periods, suspected red tide-related deaths of manatee occurred
in southwest Florida in 2002, 2003, 2005, 2007, and most recently in 2013.
Representatively, a major dolphin’s death, which occurred near the Florida panhandle
in the early 2000s, was associated with exposure to brevetoxin via food. High
concentrations of brevetoxins were detected from fish within the stomach of the
dolphin (Flewelling, Naar et al. 2005, Hoagland, Jin et al. 2009).
Over the last decade, animal studies had demonstrated the possible health effects of
both long and short terms of brevetoxin exposure. After short-term treatment, rapid
systemic distribution of brevetoxin occurred on mice after exposure to aerosolized
brevetoxin. Particularly it is concentrated in the neurologic system (Benson, Stagner
et al. 2005, Tibbetts, Baden et al. 2006). Long-term exposure of rodents (rats and
mice) to aerosols including brevetoxin showed that viral clearance in the immune
system was dysfunctional. Low level exposures (~10 pg/ml of PbTx-2 and PbTx-3) of
both asthmatic and non- asthmatic sheep resulted in decreased respiratory function.

15
However, the asthmatic sheep suffered more intense and longer lasting effects
(Abraham, Bourdelais et al. 2005, Abraham, Bourdelais et al. 2005, Abraham and
Baden 2006, Fleming, Kirkpatrick et al. 2011).
1.4.2 Biological effects of brevetoxin on human
Brevetoxin poisoning of humans can arise through food, water and air (Backer,
Fleming et al. 2003, Sikorski 2006, Fleming, Kirkpatrick et al. 2007, Grimes 2009,
Fleming, Kirkpatrick et al. 2011). NSP is associated with exposure of brevetoxin
through the consumption of contaminated shellfish. The other most common route of
exposure to the brevtoxins is inhalation (Backer, Fleming et al. 2003, Backer,
Kirkpatrick et al. 2005). The particle size of brevetoxin aerosol formed by wave
action during a red tide has a geometric mean of about 8-9 µm. In comparison,
particles that are less than 5 µm can enter the lower airway. That means only about
10-20% of these particles can enter the human lung (Pierce, Henry et al. 2003, Cheng,
McDonald et al. 2005, Cheng, Villareal et al. 2005, Cheng, Zhou et al. 2005, Pierce,
Henry et al. 2005, Fleming, Kirkpatrick et al. 2011). Nonetheless, during an active
Florida red tide season many respiratory symptoms are reported by people who spend
time on or near the west coast beaches, such as lifeguards and tourists (Cheng,
Villareal et al. 2005, Cheng, Zhou et al. 2005, Fleming, Backer et al. 2005, Fleming,
Jerez et al. 2007, Milian, Nierenberg et al. 2007). On the basis of local health care
data from Florida, a significantly higher number of hospital admission for acute and
subchronic respiratory effects such as asthma, bronchitis and pneumonia occur during
active Florida red tides period, when compared to non-active periods, particularly for
coastal residents. A comprehensive review of emergency room admissions in Florida,

16
suggested that patients require emergency care during the first hour to prevent
respiratory failure (Abraham, Plakas et al. 2008, Watkins, Reich et al. 2008). Humans
who consume contaminated shellfish can experience NSP which is characterized by
acute gastrointestinal and neurologic symptoms (Kirkpatrick, Fleming et al. 2004).
Asthma-like symptoms are related to inhalation of PbTxs by marine mammals or
people near the beach (Fleming, Backer et al. 2005, Fleming, Kirkpatrick et al. 2005).
Thus, PbTxs are responsible for a variety of negative economic, environmental, and
human health effects (Fleming, Backer et al. 2005, Errera, Bourdelais et al. 2010).
1.4.3 The effect of brevetoxin on cellular process
Multiple cellular effects are associated with brevetoxin exposure. Adverse effects
have been observed in the immune system of many species, including manatee, rat
and loggerhead sea turtle. The mechanism of brevetoxin immunotoxicity has not been
understood completely. However, several potential mechanisms have been suggested.
For example, brevetoxin inhibits the active sites of an important protease called
cathepsin (Sudarsanam, Virca et al. 1992, Kirkpatrick, Fleming et al. 2004,
Kirkpatrick, Fleming et al. 2006), which can affect the degradation of polypeptides. In
order to understand the method of the inhibition, a group developed an approach via
3D structure to search a molecule as an inhibitor for this known homologous protein
(Sudarsanam, Virca et al. 1992). They utilized the 3D structure of papain as model,
which has similar tertiary structure to cathepsins L to search for inhibitors which
filled the binding pocket using a comparative modeling technique. After searching in
the Cambridge Structural Database (CSD) of small molecules, brevetoxin B was
shown to fit in the binding pocket of papain and cathepsin L. Subsequently, both

17
papain and cathepsin L were shown to be inhibited by brevetoxin with Kis of 25 µM
and 0.6 µM, respectively.
Other effects of brevetoxin exposure include DNA damage (Murrell and Gibson
2009), reduced phagocytosis (Benson, Tischler et al. 1999), decreased plaque forming
ability (Benson, Hahn et al. 2004, Benson, Stagner et al. 2005) and chromosomal
aberration (Sayer, Hu et al. 2006). Several studies have indicated that oxidative stress
may occur as the cellular response to brevetoxin exposure. For instance, After the
human U-937 cells were treated with PbTx-2, the amount of glutathione was
decreased while oxidative stress occurred (Walsh, Leggett et al. 2009). Other studies
on marine organisms revealed a link between brevetoxin exposure and oxidative
stress. The activity of the oxidative stress marker superoxide dismutase (SOD) was
detected and correlated to the plasma brevetoxin levels and reactive oxygen in
rescued Florida manatees after exposure to a red tide bloom (Walsh, Stuckey et al.
2007, Walsh, Butawan et al. 2015). When compared to healthy turtles, a two-fold
increase in SOD was observed in rescued loggerhead turtles which correlated with
plasma brevetoxin levels at 500 ng / ml (Walsh, Leggett et al. 2010). Brevetoxin
exposure at 5.4-15.0 µg/L increased catalase activity in coral larvae and lipid
peroxidation in coral larvae and fish gills (Woo, Liu et al. 2006, Ross, Ritson-
Williams et al. 2010). Furthermore, PbTx-2 was shown to induce apoptosis in human
Jurkat cells (Walsh, Leggett et al. 2008). When Jurkat cells exposed to 5-10 µg/ml
PbTx-2 or PbTx-6, the decrease of cellular metabolic activity was observed. After 24h,
the viability of cells that were treated with PbTx-2 decreased. In contrast, apoptosis
was increased under PbTx-2 or PbTx-6 exposure. Unfortunately, the mechanism for
brevetoxin induced oxidative stress is still unknown.

18
1.5 LSP compounds bind to transmembrane protein
The best-known biological effect of brevetoxins is the activation of voltage-gated
sodium channel (VGSC), a transmembrane protein, which induces the ion influx
resulting in numerous physiological effects.
The VGSC is composed in large part of transmembrane α-helices, and membrane-
integral α-helix peptides are considered as common interacting motifs of LSPs.
Voltage-gated sodium channel is essential for the initiation and propagation of action
potentials in excitable cells. Voltage-gated sodium channel has long been believed to
be the principle mammalian target of the brevetoxins. In addition, because of the
similarity of yessotoxin to brevetoxin, it has been suggested that yessotoxin increased
the permeability of sodium ions in cell membrane, resulting in the symptoms of CFP.
For example, the α-helices of glycophorin A (GpA), a membrane protein found in
erythrocytes, have been used as a model system for evaluating the interaction between
LSPs and α-helices. The association between α-helical glycine and valine residues of
GpA result in dimer formation in membrane environments (Bormann, Knowles et al.
1989). The association of LSPs such as YTX, desulfated YTX, PbTxs and artificial
polyethers with GpA results in disruption of the oligomers or dimers. The disruption
can be evaluated by SDS–PAGE. The results demonstrated that the LSPs interact with
the transmembrane portion of glycophorin A to induce the dissociation of oligomers
of the protein. In order to quantify the interaction of LSPs with GpA, surface plasmon
resonance (SPR) and saturation transfer difference NMR were applied. The
dissociation constants (Kd) of YTX and desulfated YTX with a transmembrane
domain peptide of GpA were determined to be in the submillimolar range (Ujihara,

19
Oishi et al. 2008). All results suggested that hydrophobic matching is important for
LSP binding to transmembrane α-helices. Moreover, taken together, it indicated that
hydrophobic interactions play a key role in molecular recognition of the α-helical
peptides by LSPs.
Both brevetoxins and ciguatoxin bind to and activate the VGSC at site 5. These site 5
toxins induce channel opening at normal resting potentials leading to a change in the
biophysical properties of the VGSCs. Like brevetoxins, VGSC opening at normal cell
resting membrane potentials results in an influx of Na+ ions and cell depolarization.
Although ciguatoxin acts at the same receptor site of the Na+ channel as brevetoxin,
the affinity of brevetoxin for sodium channel is around 30 times lower than that of
CTX-1. In addition, CTX-1 competitively inhibits the binding of brevetoxin to the
voltage-dependent Na+ channel of rat membranes. As a consequence, ciguatoxin
cause the an abnormally prolonged Na+ channel opening in nerve membranes,
slowing the nerve conduction velocity (Lehane and Lewis 2000, Kumar-Roiné,
Matsui et al. 2011). In addition, compared to the short-term effects on nerve and
muscle tissue by brevetoxin, ciguatoxin induces long-term nerve damage. The plasma
membrane is unable to maintain the internal environment of cells and volume control,
resulting in cell and mitochondrial swelling and bleb formation. Thus, ciguatoxin is
responsible for the disruption of important ion exchange system (Lehane and Lewis
2000). Fortunately, intravenous (IV) mannitol infusion is the most common treatment
for CFP (Friedman, Fleming et al. 2008). Mannitol is given at a dose of 0.5 to 1.0
g/kg body weight over 30-45 minutes. However, it must be administered within 24
hours of exposure to be effective. Also, the osmotic reduction of neuronal edema was

20
indicated to mediate the effect of mannitol infusion (Pearn 2001, Friedman, Fleming
et al. 2008).
Last but not least, maitotoxin, a water soluble toxin, acts on calcium channels
(Taglialatela, Amoroso et al. 1986). Treatment of cells with low concentrations of
maitotoxin results in activation of voltage sensitive calcium channels demonstrating
that maitotoxin affects the uptake of calcium ion in cells. However, no affinity was
found by maitotoxin for voltage-gated sodium channel through binding experiment on
rat brain cortical synaptosomes. Furthermore, it is also reported maitotoxin is
associated with neurotransmitter, hormone release, phospholipid metabolism and
smooth muscle contractions (Schettini, Koike et al. 1984, Berta, Phaneuf et al. 1988,
Gusovsky and Daly 1990).
1.6 The interaction between PbTxs VGSC at the molecular level
1.6.1 PbTxs exert their effects through specific binding of PbTx to the VGSC
Previous work has shown that there are multiple distinct receptor sites for neurotoxins
in sodium channels. For example, receptor site 1 binds the inhibitors tetrodotoxin and
saxitoxin, which block ion transport through the sodium channel. Grayanotoxin and
the alkaloids veratridine, batrachotoxin and aconitine bind to receptor site 2, which
cause repetitive firing and persistent activation of sodium channels (Catterall 1980,
Catterall, Morrow et al. 1981, Catterall 1986). Receptor site 3 binds the β-scorpion
toxins and sea anemone toxins, which can inhibit sodium channel inactivation and
enhance persistent activation of sodium channels by veratridine and other toxins
acting at receptor site 2 (Tejedor and Catterall 1988, Rogers, Qu et al. 1996).
Receptor site 4 binds ß-scorpion toxins from American scorpions, which can shift the

21
voltage dependence of sodium channel activation (Catterall 1977). These other
neurotoxins that bind to the sodium-voltage gate channel failed to displace 3H labeled
PbTx from its binding site. The previous experiments led to the conclusion that the
PbTxs bind to a unique site in the VGSC that was termed site 5 (Poli, Mende et al.
1986). The specific binding is reversible and temperature-dependent. Through
analysis, PbTx-3 binding was determined have a Kd of 2.9 nM (at 4°C). Equilibrium
binding was also observed after incubation of 3H labeled PbTx-3 with Tilapia brain
synaptosomes with a Kd of 6.1 nM (Stuart and Baden 1988). It was also determined
that PbTx-3 can be displaced by PbTx-1 and PbTx-2 at specific binding site.
Electrophysiological studies revealed that specific binding of PbTx to the VGSC
results in membrane depolarization in a dose dependent manner (Huang, Wu et al.
1984). When PbTx is incubated with crayfish giant axons, the extent of maximum
depolarization is about 40 mV, with an EC50 of 1.7 nM (Huang, Wu et al. 1984). Thus,
PbTxs cause depolarization of excitable membranes, facilitating the inward flow of
sodium ions into the cells (Trainer and Baden 1991, Dechraoui, Naar et al. 1999).
Binding of PbTxs to the sodium channel causes a shift the channel activation potential
to more negative values, inhibits channel inactivation and prolongs mean open times
of the channel (Jeglitsch, Rein et al. 1998). Thus, PbTxs disrupt the normal
transmission of electrical signals within the neurological system. A massive release of
neurotransmitter is stimulated by the depolarization of nerve terminals, which results
in a wide range of responses in the organs. Therefore the consumption of seafood
contaminated with PbTxs has the potential to disrupt normal neurological systems
causing the illness clinically described as NSP (Twiner, Bottein Dechraoui et al.
2007).

22
1.6.2 Specific interactions between brevetoxin and the VGSC
The VGSC is composed of an α-subunit and two β-subunits. The α-subunit is the
major subunit and made up of four domains. The four domains of the channel
containing six transmembrane α-helices with the connecting amino acid sequence
between helices 5 and 6 coming together to line the pore through which the ions travel
(Heinemann, Terlau et al. 1992, Hille 2001). Early studies, which helped to define the
interaction between the PbTxs and the VGSC at the molecular level included structure
activity studies, conformational analysis and photoaffinity labeling of the receptor
(Poli, Mende et al. 1986, Trainer and Baden 1991, Trainer, Thomsen et al. 1991, Rein,
Baden et al. 1994).
Brevetoxin-A and brevetoxin-B bind to the same unique site of the VGSC at site 5
which is located on domain IV of the α-subunit of the channel. Using photoaffinity
labeled PbTx in rat brain synaptosomes the PbTx high affinity receptor site was
localized to the α-subunit. After binding of the photoaffinity probe, a p-azidobenzoate
tritium-labeled brevetoxin derivative, to the VGSC and photoloysis, it was shown that
the VGSC was covalently labeled with the PbTx-3 derivative. After digestion with
proteolytic enzymes, antibody mapping of the proteolytic fragments localized the
covalent incorporation of this neurotoxin to the sodium channel α-subunit on the
extracellular side of S5 in domain IV and on the extracellular end of the S6 in domain
I (Rein, Baden et al. 1994, Trainer, Baden et al. 1994).
Neuronal tissues in marine mammal have similar sodium channel density as terrestrial
mammals. Brevetoxin (PbTx-3) binds to manatee brain synaptosomes with nanomolar

23
affinity at 37 oC. (Trainer and Baden 1999). The dissociation constant (Kd) is 7.5±2.5
nM for manatee synaptosomes, which is a similar affinity to terrestrial mammals
(Trainer and Baden 1999). Since 1982 to 2005, hundreds of manatees were found
dead or dying on the beaches of the Florida west coast when high densities of K.
brevis were present, including 39 deaths in 1982, 149 deaths in 1996 and 34 deaths in
2002. Unbelievably, in 2005 it is reported that 85 manatees died because of K. brevis
(O'Shea, Rathbun et al. 1991, Bossart, Baden et al. 1998). In Assuming 11 pg
brevetoxin was produced from a single cell (Stuart and Baden 1988). If the affinity of
manatee for brevetoxin is 7.5 nM, approximately, 3 x 105 cell/L brevetoxins are
required. Whereas in fact concentration of brevetoxin cells at that period persisted at
100,000 cells/L. This discovery indicated that manatees were exposed to a lethal dose
of brevetoxin in 1996 (Trainer and Baden 1999). Neuronal cells, skeletal muscle cells
and cardiac cells are all excitable tissues, which contain VGSC to mediate the rapidly
activation and inactivation of Na+ influx. A comparative study examined the binding
properties of brevetoxins with skeletal muscle and cardiac sodium channel α-subunit
isoforms of human embryonic cells. Brevetoxin B was found to have isoform
selectivity with cardiac channel and skeletal muscle channel (Dechraoui and Ramsdell
2003). It is determined that the inhibitory constants Ki for PbTx-3 were 4.7 nM for
skeletal muscle channels, while it is 20.5 nM for cardiac sodium channels.
1.7 The effect of brevetoxin on economy
As a consequence of a bloom along coastal waters, local economies suffer not only
direct losses to fishing and tourism but also economic effects associated with animal
death, clean-up, rescue and rehabilitation, and public health also cause additional

24
losses. A single toxic bloom can result in as much as $22 million in lost revenue from
tourism and related business (Dyson and Huppert 2010, Anderson, Moore et al. 2015).
In 2000, a large bloom of K. brevis occurred in Galveston County, Texas. It required
$18 million for direct cleanup and subsequent losses to fishing and tourism (Evans
and Jones 2001). The economic costs caused by the harmful effect of K. brevis are
estimated to exceed $38 million annually (Twiner, Bottein Dechraoui et al. 2007). A
cost model using the data on emergency room visits for respiratory damage, the K.
brevis density, measurement of pollutant, annual population change and tourism
estimated that the cost of illness associated with K. brevis in Sarasota county alone
can range from $0.5-$4 million (Hoagland, Anderson et al. 2002, Hoagland, Jin et al.
2009, Kirkpatrick, Bean et al. 2010). Furthermore, during an active Florida red tide
period the costs resulting from lifeguard absenteeism can reach up to $100,000 in
Sarasota Beach alone. In 2014, Hoagland estimated that annual costs of illness caused
by Florida red tide ranged from $60,000 and $700,000 per year. However, it is
assumed that this cost could be increased to $ 1 million per year by several long-last
Florida red tide. Furthermore, in future the cost of illness would range between $2 and
$24 million under a worst case scenario (Hoagland, Jin et al. 2014).
1.8 Brevetoxins play an important role in K. brevis, but it is still elusive
Because PbTxs are retained in K. brevis cells, it has been presumed that they play a
functional role for K. brevis. Although a primary cellular function for PbTxs has
remained elusive, it is reported that the responses to environmental conditions, such
as salinity, influence the growth potential and toxin production of harmful
dinoflagellates (Grzebyk, Béchemin et al. 2003, Magaña, Contreras et al. 2003). In
2011, it was proposed that PbTxs play an important role in osmoregulation by K.

25
brevis. It was found that osmotic stress triggered PbTx production by K. brevis in
three K. brevis clones under variable salinity conditions that result in changing
osmotic stress. A rapid decrease in salinity from typical oceanic conditions of 35 ppt
to approximate coastal conditions of 27 ppt resulted the concentration of brevetoxin
per cell increasing within 5 days under hypoosmotic stress and provided evidence to
suggest PbTxs play a role in osmoregulation. Furthermore, in order to understand the
response of K. brevis to osmotic stress, a time course experiment to measure changes
in cell volume and PbTxs production was performed. <3h, 24h, 6d, 67d, and 111d
after the rapid salinity shift PbTxs content was examined. Brevetoxin increased within
3 h after the hypoosmotic stress and continued to increase through day 6. After 6 days
the concentration of PbTxs reached the highest level (Errera and Campbell 2011).
However, in 2012 this conclusion was challenge by several other research groups who
attempted to repeat Errera and Campbell’s experiments. Two separate laboratories
reported that a sudden decrease in salinity from 35 ppt to 27 ppt, did not trigger
brevetoxin production. These results were consistent across each of eight K. brevis
strains (Sunda, Burleson et al. 2013). Consequently, this debate generated much
attention from researchers. Still, the cellular function of PbTxs is under debate.
Chemical and predatory defense has been proposed as the reasons for brevetoxin
production by K. brevis. Feeding experiment experiments using PbTxs have been
conducted to better understand how K. brevis escapes predation pressure. Rotifers are
common microzooplankton grazers which co-exist in coastal water with K. brevis.
Compared to a known palatable food for rotifers, when K. brevis cells were offered as
a sole diet for rotifers, all rotifers stopped grazing and breeding. However, it is not
clear that the deterrence of grazing was a result of the presence of PbTxs. Rotifers fed

26
with yeast containing pure PbTx-2, PbTx-3 and PbTx-9 were not deterred from
grazing (Kubanek, Snell et al. 2007). Thus, defending K. brevis against grazing by
may or may not be a role for brevetoxins.
1.9 The significance of studying the dinoflagellates
In the past decades, scientists have made significant progress in determining the
effects and the underlying mechanisms of dinoflagellate toxins on higher organisms.
Because of the extensive damage and pharmacological effects caused by K. brevis,
there is considerable interest in understanding the function of PbTxs. If a metabolic or
biochemical role for brevetoxins can be identified, this may provide for more accurate
prediction of bloom dynamics and toxicity. The role of brevetoxin in cells can lead to
more accurate predictions, mitigation and management of toxic blooms. While
numerous hypotheses have been proposed and several studies performed, the intrinsic
role of dinoflagellate toxins remains unknown In addition; the discovery of the native
role of brevetoxin in K. brevis may provide insight into understanding the role of
other polyether compounds in dinoflagellates such as ciguatoxin, maitotoxin and
yessotoxin.
1.10 Research objectives
The research will focus on identifying the intrinsic biochemical function of PbTxs and
identification of a molecular receptor for brevetoxin in K. brevis. Localization of
PbTxs to a subcellular organelle and identification of a target protein from K. brevis,
which interacts with PbTxs as well as comparison of biochemical and metabolic
indicators in K. brevis strains with vastly different toxin content, may point towards a
native function for PbTxs.

27
Objectives
1. To localize brevetoxin in K. brevis using a fluorescently labeled brevetoxin probe.
2. To identify one or more target protein(s) of brevetoxins from K. brevis.
3. To characterize interaction between brevetoxin and its native receptor protein
4. Compare and contrast various metabolic and biochemical parameters in two strains of
K. brevis which have a tenfold difference in toxin content.

28
Chapter 2 Localization of PbTxs to a subcellular organelle and identification of a
target protein from K. brevis
2.1 Localize PbTxs to one or more organelles or cellular structures in K. brevis
2.1.1 Objective
The localization of brevetoxin to one or more organelles in K. brevis cells should
provide clues as to the intrinsic role of brevetoxin in K. brevis. In this section, we
describe the use of a fluorescently labeled brevetoxin probe along with fluorescence
microscopy to identify the site of localization of brevetoxin.
2.1.2 Introduction
Labeling of biological molecules is one of the most common imaging methods used in
bioanalytical fields, such as medicine, pharmacy, cellular biology and environmental
science. Labels can be radioactive or molecules with absorption or fluorescence.
Fluorescence techniques have been used for biological imaging for many decades.
Thousands of fluorescence probes are used to stain the organelles in living cells and
many fluorescence stains are commercially available. For instances, DAPI (4’, 6-
diamidino-2-phenylin-dole) is a fluorescent dye which is specific for DNA. In the
1970s DAPI was discovered to stain the DNA of yeast cells. It was demonstrated that
DAPI fluorescence was sensitive to DNase (Williamson and Fennell 1975). DAPI
forms a fluorescent complex through its high affinity binding of DAPI with the minor
grove of A-T rich sequence of DNA (Williamson and Fennell 1975). Thus, DAPI can
be a biological stain for nucleic acids and chromosomes (Lin and Alfi 1976,
Schweizer 1976, Schweizer 1976, Lin, Comings et al. 1977). Blue-fluorescent

29
Hoechst 33342 dye also stains the nuclei and hematoxylin is a stain for nuclei in
living cells (Fischer, Jacobson et al. 2008). Rhodamine is one of numerous
histological stains, which have been utilized for many years as a specific probe to
localize the mitochondria in living cells. Rhodamine 123 also can be used in detection
of the energy state of isolated mitochondria (Azzone 1972, Johnson, Walsh et al.
1980). MitoFluor™ Far Red 680 accumulates in mitochondria, and its fluorescence
(emission maximum 700 nm) was red. Green-fluorescent LysoTracker® Green dye
accumulates in the lysosomes.
The dyes described above are general dyes for staining organelles. Reactive reporter
dyes may be used to detect specific classes of molecules within a cell. In 2008, a type
of fluorescent probe called MitoPY1 for imaging hydrogen peroxides within
mitochondria of live cells was determined. MitoPY1 was synthesized with a peroxide-
responsive element and mitochondrial targeting phosphonium moiety to localize
hydrogen peroxide to cellular mitochondrial (Dickinson and Chang 2008). A
rhodamine modified as an electron donor (4-amino- or 4- hydroxyphenyl ether), is
used for detection of highly reactive oxygen species (ROS) in mitochondria of live
cells. thereby ROS should cause cleavage of the ether moiety resulting in fluorescence
in mitochondria (Koide, Urano et al. 2007). Thus, these indirect and modified
fluorescent probes are utilized to selectively detect biomolecules in live cells.
The fluorescent tagging of specific molecules is a promising technique for localizing
molecules in live cells. For example, immunofluorescent staining is able to detect
biomolecules in living cells as long as the specific antibody is provided. The
fluorochrome-labeled antibody is used to stain intracellular biomolecules as a

30
common and direct method. Permeabilization of the plasma membrane is an essential
and often problematic, step in this method (Suzuki, Matsuzaki et al. 2007). For
additional examples, labeled oligonucleotide probes were used to image individual
mRNA molecules in fixed cells, which made each mRNA molecule visible as
fluorescent microscopy (Raj, Van Den Bogaard et al. 2008). Using a designed and
synthesized fluorescent probe included a small receptor domain and ligand label some
recombinant proteins which contains four cysteines at 5 positions of an α helix. The
small receptor domain is composed of six natural amino acids that could be
genetically mutated-bind to proteins of interest. The ligand is membrane-permeant
and nonfluorescent until it binds specifically with high affinity to the tetracysteine
domain (Griffin, Adams et al. 1998).
In order to understand the biochemical function for brevetoxins, our first goal is to
localize brevetoxins to one or more organelles or cellular structures in K. brevis.
Consequently, using a fluorescently tagged brevetoxin probe is an effective and direct
approach to localize PbTxs in K. brevis. The site of localization can be visualized via
fluorescence microscopy
The synthesis of a fluorescently labeled brevetoxin probe has been reported
previously (Cassell, Chen et al. 2015). Briefly, Alexa Fluor® 488 alkyne (4) is used as
fluorescent dye linked to brevetoxins, which are highly lipophilic molecules (Figure7).
The fluorescently labeled PbTx-2 probe (5) was prepared by coupling brevetoxin with
an Alexafluor alkyne derivative (4). A thiol-Michael reaction between PbTx-2 (1) and
an azidothiol (2) provided a azide functionalized thiazepine linker (3), which was
subjected to a 1, 3-dipolar cycloaddition reaction with Alexa Fluor 488 alkyne (4).

31
The Alexa Fluor dye was chosen specifically because it is cell-impermeable. Thus, if
the brevetoxin - Alexa Fluor® 488 Alkyne conjugate can penetrate the membranes, it
is only because of brevetoxin. The thiol-Michael reaction has been described as a
“click” reaction (Nair, Podgorski et al. 2013). “Click chemistry” is commonly used to
combine an azide labeled molecule and an alkyne labeled molecule. A “click
“reaction is a fast reaction which can be performed under mild conditions (Nair,
Podgórski et al. 2013). Subsequently, the probe was incubated with two K. brevis
strains, a high toxin producing strain and a low toxin producing strain. Stained cells
emitting light at 500-540 nm were observed using fluorescence microscopy. Thus
using live cell imaging, the location of brevetoxins in K. brevis cells can be identified.
Figure 7 Synthesis of fluorescent brevetoxin probe. For brevity, only the K-ring of PbTx-2 is shown. a) MeOH, b) tBuOH/H2O/DMF, CuSO4·-5H2O, sodium ascorbate
2.1.3 Results and discussion
In order to determine the location of brevetoxin in K. brevis, the fluorescent probe (5)
and Alexa Fluor® 488 Alkyne (4) were each incubated for 1 hr with K. brevis living
cells. After washing the cells with culture medium, the cells were visualized by using

32
a fluorescent microscope equipped with a 450–490 nm band pass/515 nm long pass
EX/EM filter set. Chlorophyll auto fluorescence (red) could be clearly seen in both
treatments (Figure 8). However, incubation with the fluorescent PbTx-2 probe also
demonstrated localization of the fluorescent PbTx conjugate (yellow) to the
chloroplasts (Figure 8 A and B). Additive color is produced by combining a number
of primary colors such as green, red and blue in equal proportions. Thus, although
Alexa Fluor® 488 alkyne is a green dye, based on the additive color mixing, the green
color combines with red color resulting in yellow. However, the control experiment,
with Alexa Fluor® 488 alkyne alone, failed to label K. brevis cells (Fig 8 C). This
experiment demonstrated that PbTxs localize to the chloroplast of K. brevis
Figure 8 The photograph of localization of brevetoxin in K. brevis live cells (A) Fluorescent brevetoxin derivative (0.4 mg / ml) was incubated with live K. brevis culture for 30 mins in dark. The cells were observed through 40X lenses of a fluorescent microscope equipped with a 450–490 nm band pass/515 nm long pass EX/EM filter set. (B): The zoom in image of labeled cells. (C): The zoom in image of cells in control experiment: Alexa Fluor® 488 Alkyne (0.06 mg / ml) was incubated with live K. brevis culture for 30 mins in dark
The finding is consistent with the localization of okadaic acid (OA) (Fig 9), a marine
toxin produced by some dinoflagellates of the genera Prorocentrum and Dinophysis
(Zhou and Fritz 1994). The location of OA has been identified first with
immunofluorescence. Toxin-producer cells were incubated with anti-OA fluorescein
isothiocyanate (FITC) - conjugated antibody. Fluorescence was observed at the

33
plastids. In order to obtain even more specific localization, immunogold labelling of
anti-OA antibody was used. Using the immunofluorescence micrographs through
transmission electron microscope operating at 80 kV, the gold particles were visible
with the pyrenoid region of the chloroplast. Thus, it had been proposed that OA was
localized primarily within chloroplasts of toxin-producing Prorocentrum species
(Zhou and Fritz 1994).
Figure 9 The structure of okadaic acid
2.2 Isolation of a target protein from K. brevis as PbTxs natural receptor
2.2.1 Objective
Identification of a brevetoxin binding protein in K. brevis may provide some clues to
the intracellular function of PbTxs in K. brevis cells.
2.2.2 Introduction
Photoaffinity labeling (PAL) provides a direct and general method to identify binding
proteins. Photoaffinity labeling can directly identify the interacting site at the
molecular level. In photoaffinity labeling, a new covalent linkage is created between a
light-sensitive, detectable ligand and a biopolymer upon irradiation, in a reversibly
bound state (Dormán and Prestwich 2000). A typical photoaffinity probe incorporates
three groups, a ligand, a photoreactive group, and a tag for purification. The
photoreactive groups should have some essential features: the photoreactive groups (1)
must be stable under ambient light with an activation wavelength longer than 300 nm

34
(Dormán and Prestwich 2000). (2) should preferably provide a single covalent adduct
between the probe and target protein avoiding release of an activated form that results
in non-specific labeling (3) have an activated form that reacts with C-H groups and X-
H bonds. The most reliable and common high efficiency labeling of target proteins
are tetrafluorophenyl azides (AZ), trifluoromethylphenyl diazirines (DIA) and
benzophenone (BP) (Fleming 1995, Weber and Beck-Sickinger 1997)(Figure10 ).
These photoactiveatable groups have been used extensively to identify the target of
drugs, to determine the affinity and selectivity of the drug-target interaction, and to
identify the binding site on the target.
Figure 10 The most reliable and common high efficiency labeling of target proteins in photoaffinity probe
Although less common, photoaffinity labeling has also been used in non-
pharmaceutical applications as well, with several studies using DIA as the photophore.
In 2003, using DIA modified photoaffinity probe that attached to 2’ -position of the
sugar moiety, a potential binding site to the receptor molecule, successfully detected
the potential membrane receptor proteins for leaf movement of legumes (Sugimoto,
Fujii et al. 2004). DIA that linked to maitotoxinin with a biotin detectable group was
used to identify a binding protein for maitotoxin in red blood cells (Konoki,
Hashimoto et al. 2009). Although the target protein has not been characterized, based
on the SDS-PAGE, a 23 kD protein was specifically labeled. Competition binding
experiments with PbTx-2 demonstrated that both toxins bound to the same site on this

35
protein. Thus, the use of a PbTx-2 photoaffinity probe could be a direct and effective
method to discover the target protein of PbTxs in K. brevis.
The synthesis of the photoaffinity probe has been previously reported (Cassell, Chen
et al. 2015). The photoaffinity probe (10) was consisted of a photolabile diazirine and
a biotin tag. In order to isolate a target protein, the biotin-streptavidin interaction was
used to separate the target protein from others. Briefly, the known aldehyde (6)
(Hashimoto, Hatanaka et al. 2001) was alkylated with 1-azido-6-bromohexane to
provide azide (7). “Click chemistry” was used to link the azide functionalized
diazirine with an alkyne functionalized biotin to provide (8). Reductive alkylation of
the aldehyde moiety with cystamine, followed by reduction of the intermediate
disulfide, provided the thiol (9). Thiol (9) then underwent a thiol-Michael reaction
with PbTx-2 (1), followed by cyclization to the thiazapeine provided photoaffinity
probe (10) (Figure 11).

36
Figure 11 The synthesis of brevetoxin photoaffinity probe (10). For brevity, only the K-ring of PbTx-2 is shown. a) K2CO3, KI, DMF; b) tBuOH/H2O/DMF, CuSO4·5H2O, sodium ascorbate; c) (NH2CH2CH2S)2, NaBH4, EtOH; d) DTT, MeOH; e) MeOH; 2.2.3 Results and Discussions
A homogenate of K. brevis cells was separated into soluble and membrane fractions
by centrifugation. These fractions were each incubated either with the photoaffinity
probe (10) or the photoaffinity label (8), followed by photolysis and protein isolation
using streptavidin immobilized on magnetic beads. After incubation of the brevetoxin
photoaffinity probe (10) with the soluble protein fraction and photolysis no proteins
were observed in SDS PAGE after purification on streptavidin beads. However,
incubation and photolysis of the membrane fraction with the brevetoxin photoaffinity
probe (10) consistently revealed a protein with an apparent mass of 27 kD when
analyzed by SDS-PAGE (12). A control experiment with photoaffinity label (8) alone
did not yield any bands by SDS-PAGE.

37
The 27 kD bands were subjected to digestion with trypsin. After analysis of the
excised band by ESI LC-MS/MS followed by BLAST (basic local alignment search
tools) analysis of the amino acid sequence against NCBI non-redundant database, it
was determined that there were two thylakoid proteins which interacted with
brevetoxin: the light harvesting complex II (LHC II) of the photosynthetic apparatus
and thioredoxin (Trx). The identified peptide fragments and their closest match in the
BLAST analysis are shown in Table 2. Subsequently, we determined that our
photoaffinity probe also binds to the LHC II of spinach (Figure13 lane 6).
Figure 12 SDS-PAGE of protein fractions from photoaffinity experiments. K. brevis membrane fractions incubated with photoaffinity probe (10) or (8). Lane 1: molecular weight markers. Lane 2: K. brevis membrane fraction incubated with (10) (7 mg protein). Lane 3: pre-incubation membrane fraction (7 mg protein). Lane 4: K. brevis homogenate incubated with (8) (18 mg protein). Lane 5: pre-incubation membrane fraction (6 mg protein). Lane 6: spinach homogenate incubated with (10) (16 mg protein)
Table 2 The peptide sequence of 27 kD band and the proteins with matching sequences.
Protein name Organism Peptide sequence
LHCII Emiliania huxlei GGKPGYFPTFK LHCII Emiliania huxlei PGYFPTFK LHCII Emiliania huxlei LAMLGLFSLISEAK LHCII Emiliania huxlei VPGAVPALAGK
Thioredoxin Emiliania huxlei IIHLTDDSFDTDVLK Thioredoxin Emiliania huxlei MIAPILDEIADEYQGK Thioredoxin Emiliania huxlei GIPTLLLFK
Thioredoxin 1 Aliivibrio salmonicida LNIDQNAGTPPK

38
Thylakoids are membrane-bound structures that are located in chloroplast stroma. The
light-dependent reactions of photosynthesis occur in the thylakoids. The thylakoid is
composed of a thylakoid membrane surrounding a thylakoid lumen. In the thylakoid
membrane the photosynthetic pigments are bound to proteins which are embedded in
the membrane. There are many integral membrane proteins that play important roles
in light harvesting and the light dependent reactions of photosynthesis take place in
thylakoid membranes. Photosystems I and II, cytochrome b6f complex and ATP
synthase are four major protein complexes in the thylakoid membrane. LHC is a
complex of subunit protein that is a part of a larger supercomplex of a photosystem.
LHC II surrounds the photosynthetic reaction center in thylakoid membranes to both
harvest and transfer light energy to photosystem II and to protect it from oxidative
damage that is caused by excess sunlight. The molecular weight of LHC II is 27 kDa.
It is highly conserved across species. It is essential for both light harvesting and non-
photochemical quenching (NPQ). NPQ is a mechanism to quench chlorophyll
fluorescence and dissipate the excess chemical energy as heat rather than chlorophyll
fluorescence which results in the formation of ROS. Thus, an important function of
LHCII is to prevent oxidative damage to photosystem II. LHCII binds to 14
chlorophyll and 4 carotenoid molecules. These pigments are used for light harvesting
and energy dissipation. In terrestrial plants, the four carotenoids are two luteins,
neoxanthin and zeaxanthin or violaxanthin. In dinoflagellates the zeaxanthin and
violaxanthin are replaced by dinoxanthin (D) and diadinoxanthin (DD) respectively.
Lutein and neoxanthin are replaced by fucoxanthin in K. brevis (Demers, Roy et al. ,
Arsalane, Rousseau et al. 1994, Moisan, Olaizola et al. 1998, Vijayan and Somayajula
2014). The zeaxanthin/violaxanthin interconversion facilitates light energy

39
dissipation through the xanthophyll cycle (Figure 13). Under high light conditions, the
violaxanthin is converted to zeaxanthin in a process catalyzed by the enzyme
violaxanthin de-epoxidase (VDE). It has been assumed that zeaxanthin binds to LHC
II which undergoes a conformational change moving zeaxanthin into close proximity
to the chlorophyll molecule permitting the quenching of chlorophyll fluorescence.
Under low-light conditions zeaxanthin is converted back to violaxanthin by the
enzyme zeaxanthin epoxidase (ZE) (Fuciman, Enriquez et al. 2012). However, it is
not clear if violaxanthin is released from LHC II or converted within LHC II.
Figure 13 Xanthophyll cycle
Under excess light condition, photosynthesis begins; the lumenal side of the thylakoid
membrane experiences a significant drop in pH (from 8 to 5). NPQ is initiated by this
drop in pH on the lumenal side of the thylakoid membranes in response to light. In
oreder to determine if brevetoxin binding to LHCII is pH dependent, the binding of
brevetoxin photoaffinity probe to LHCII in spinach membranes was examined by
using a pH range from pH 2 (simulating high light conditions) to pH 8 (simulating
low light conditions). After incubation of the crude spinach homogenate with the
brevetoxin photoaffinity probe, photolysis at 312 nm for 20 min and affinity

40
purification on magnetic streptavidin beads, SDS PAGE analysis was performed on
the eluant. Inspection of the SDS PAGE gel revealed a pH dependence on binding
(Figure14). In the Figure 14, the red box is actin added as an internal standard for
normalization. The green box is the 27 kD band contained LHCII. The orange box is
the smaller band which has numerous photosystem I and II membrane proteins,
Rubisco small subunit, thioredoxin and a serine/threonine protein kinase. Brevetoxin
bind more strongly to 27 kD protein at low pH (or conditions which simulate high
light). Nevertheless, at high pH (or conditions which simulate low light) brevetoxin
binds with several lower molecular weight proteins (below 20 kDa, Figure 14) The
lower molecular weight band was also observed in every experiment. In order to
determine the relative intensities of each band, a constant amount of actin was mixed
with the eluant as an internal standard (Table 3). The lower molecular weight band
was also subjected to digestion with trypsin followed by MS/MS analysis. After
analysis of the digested band by ESI LC-MS/MS followed by BLAST analysis of the
amino acid sequence against NCBI non-redundant database, five different
photosystem I and six different photosystem II membrane proteins were identified., as
well as three proteins which are located in the stroma: thioredoxin, a protein kinase,
and Rubisco (Table 4). Thus, it appears that at high pH or high light, brevetoxin may
be simply released from LHC II into the thylakoid membrane where it might interact
with many proteins.
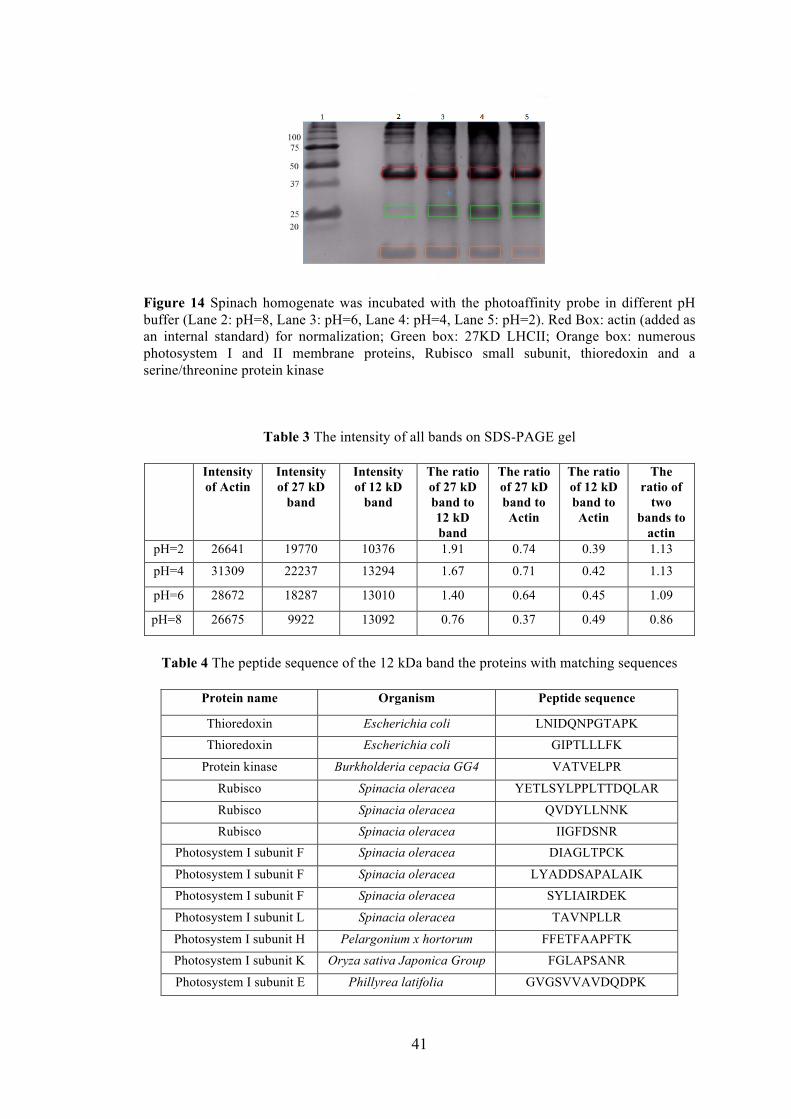
41
Figure 14 Spinach homogenate was incubated with the photoaffinity probe in different pH buffer (Lane 2: pH=8, Lane 3: pH=6, Lane 4: pH=4, Lane 5: pH=2). Red Box: actin (added as an internal standard) for normalization; Green box: 27KD LHCII; Orange box: numerous photosystem I and II membrane proteins, Rubisco small subunit, thioredoxin and a serine/threonine protein kinase
Table 3 The intensity of all bands on SDS-PAGE gel
Intensity of Actin
Intensity of 27 kD
band
Intensity of 12 kD
band
The ratio of 27 kD band to 12 kD band
The ratio of 27 kD band to Actin
The ratio of 12 kD band to Actin
The ratio of
two bands to
actin pH=2 26641 19770 10376 1.91 0.74 0.39 1.13
pH=4 31309 22237 13294 1.67 0.71 0.42 1.13
pH=6 28672 18287 13010 1.40 0.64 0.45 1.09
pH=8 26675 9922 13092 0.76 0.37 0.49 0.86
Table 4 The peptide sequence of the 12 kDa band the proteins with matching sequences
Protein name Organism Peptide sequence
Thioredoxin Escherichia coli LNIDQNPGTAPK Thioredoxin Escherichia coli GIPTLLLFK
Protein kinase Burkholderia cepacia GG4 VATVELPR
Rubisco Spinacia oleracea YETLSYLPPLTTDQLAR
Rubisco Spinacia oleracea QVDYLLNNK Rubisco Spinacia oleracea IIGFDSNR
Photosystem I subunit F Spinacia oleracea DIAGLTPCK
Photosystem I subunit F Spinacia oleracea LYADDSAPALAIK Photosystem I subunit F Spinacia oleracea SYLIAIRDEK
Photosystem I subunit L Spinacia oleracea TAVNPLLR
Photosystem I subunit H Pelargonium x hortorum FFETFAAPFTK Photosystem I subunit K Oryza sativa Japonica Group FGLAPSANR
Photosystem I subunit E
Phillyrea latifolia
GVGSVVAVDQDPK

42
Photosystem II protein D2 Acutodesmus obliquus AYDFVSQEIR
Photosystem II protein D2 Acutodesmus obliquus AAEDPEFETFYTK
Photosystem I subunit S Spinacia oleracea ENELFVGR Photosystem I subunit S Spinacia oleracea FVDEPTTGLEK
Photosystem I subunit Q Spinacia oleracea VSASEILNVK
Photosystem I subunit R Agave tequilana GVYQFVDK Photosystem II protein V Spinacia oleracea SFADIITSIR
Photosystem II protein V Spinacia oleracea PNEYFTESR
Photosystem II protein V Spinacia oleracea QGIPLITGR Photosystem II protein V Spinacia oleracea FDSLEQLDEFSR
Cytochrome f Spinacia oleracea EVVDIIPR
LHC II Oryza sativa Japonica Group VGGGPLGEGLDK
LHC II Spinacia oleracea VAGGPLGEVIDPLYPGGSFDPLGLADDPEAFAELK
Many of the proteins identified were considerably larger than the 12–15 kD indicated
by the markers. Thus, it was surprising to find them in this low MW band. It is
believed that because protease inhibitors were not present in our homogenization
buffer, high MW proteins might have been cleaved during the incubation period.
2.2.4 Conclusion
According to the result of the experiment using both the fluorescent PbTx-2
derivative (5) and the brevetoxin photoaffinity probe (10), it was demonstrated that
the brevetoxin is located in the chloroplast. It could be argued that the labels influence
the physiochemical properties of brevetoxin. However, these two labels have distinct
properties, yet each of these derivatives migrated to the chloroplast. The Alexa Fluor
dye, in particular, is not membrane-permeable, and the Alexa Fluor brevetoxin
conjugate would seem even less likely to enter the thylakoid membrane than native
brevetoxin. In addition, it is very clear that fluorescent PbTx-2 derivative did not label
any other organelles in the living cells. The finding that brevetoxins localize to the
lipophilic thylakoid membranes should come as no surprise. PE ladder molecules are

43
known to migrate to the cell membranes where they interact with membrane proteins.
Brevetoxins have also been demonstrated to aggregate into artificial lipid bilayers
(Woo, Liu et al.). Our findings are consistent with the localization of another
dinoflagellate toxin, okadaic acid (Zhou and Fritz 1994), to the chloroplast and the
cyanobacterial toxin, microcystin, to thylakoid area of cyanobacteria (Young,
Thomson et al. 2005)
In addition, it was determined using photoaffinity probe (10) that LHC II and Trx are
native proteins to which brevetoxin binds in K. brevis. The association with LHCII
was reduced at neutral pH or low light, conditions which were used for the initial
photoaffinity experiments. However, LHC II is highly concentrated in the thylakoids.
It is possible that our photoaffinity probe linked to LHCII simply as a result of its
high density in the thylakoids and that the true brevetoxin target in chloroplasts is a
less abundant membrane protein, such as an ion channel. Ion channels selective for
chloride, potassium, or divalent cations such as calcium and magnesium have been
found in outer- and inner-envelope membranes of chloroplasts, as well as from
thylakoid membranes, by using the patch clamp technique and have been
hypothesized to regulate photosynthesis by modulating the proton motive force
(Checchetto, Teardo et al. 2013).
In addition to LHC II, Trx is identified from both 25 kDa and 12 kDa band. It was
identified in every photoaffinity binding experiment performed. Trx is a ubiquitous
antioxidant enzyme that can be found in all living organisms from archaea to
mammals. The role of Trx is the regulation of redox homeostasis via thiol-disulfide
exchange reactions (Collet and Messens 2010) with its protein targets. It acts as a

44
reductase in redox control and protects proteins from oxidative damage (Holmgren
1985). In addition Trx helps the cells tolerate various environmental stresses, such as
ROS (Messens and Silver 2006). Numerous thioredoxin-dependent regulatory
networks have been identified in photosynthetic organisms. Thus, a better
understanding of the interaction between Trx with brevetoxin may contribute to
understanding the role of brevetoxin in K. brevis and may also have implications for
human and animal toxicity.
2.3 Materials and Method
2.3.1 Fluorescent probe experiment
Materials for fluorescent probe experiment Alexa Fluor 488; Fluorescent PbTx-2
derivative; Live K. brevis culture, a fluorescent microscope (Leica Leitz, Model
DMRB)
Method for fluorescent probe experiment
Culture growth K. brevis culture (Wilson strains) were obtained from Mote Marine
Laboratory (Sarasota, Florida) and maintained in RE medium (Sinigalliano, Winshell
et al. 2009), in a growth chamber at ~20ºC under 35 µmol.photons.m-2.s-1. Growth
was monitored by counting a 1:10 dilution of culture in Z pak reagent using a
Beckman Z-series Coulter Counter with aperture size between 10~30 µm according to
the manufacturer’s instructions.
Incubation of K. brevis with the fluorescent PbTx-2 derivative or the Alexa Fluor
488 alkyne: Live K. brevis culture (3 ml) was incubated for 1h in the dark with either
the fluorescent PbTx-2 derivative (5) or Alexa Fluor 488 alkyne (4). The final

45
concentration of the Alexa Fluor 488 alkyne was 4 µm. Because the exact
concentration of the PbTx-2 derivative was not known, the concentration of the
solution of the fluorescent PbTx-2 derivative was adjusted such that the fluorescence
was equal to that of the Alexa Fluor 488 alkyne solution. After incubation, the cells
were centrifuged for 8 min at 130 X g. The supernatant was discarded, and the cells
were resuspended in RE medium (1 ml) (Sinigalliano, Winshell et al. 2009) and
centrifuged again for 8 min at 130 X g. The supernatant was discarded, and the cells
were resuspended in RE medium (1 ml). The live cells were visualized by using a
fluorescent microscope (Leica Leitz, Model DMRB) with a 450–490 nm band
pass/515 nm long pass EX/EM filter.
2.3.2 Photoaffinity probe experiment
Materials for brevetoxin photoaffinity experiment K. brevis culture;Phosphate
buffered saline (PBS) buffer; Bovine serum albumin (BSA); Coomassie Protein
Assay Reagent (BioRad); Spinach; Photoaffinity probe (10); photoaffinity label (8);
Streptavidin beads (streptavidin immobilized on magnetic beads, Thermo Scientific
Pierce); Triton X100;
Method for brevetoxin photoaffinity experiment
Preparation of protein homogenates and membrane suspensions: The
homogenate of K. brevis cells was separated into soluble and membrane fractions. K.
brevis culture (200–400 ml) was concentrated by centrifugation for 5 min at 466 X g,
and the supernatant was discarded. The cells were resuspended in phosphate-buffered
saline (PBS, 400 µl) and vortexed for 1 min. This suspension was centrifuged for 10

46
min at 14,000 X g, and the supernatant (soluble fractions) and the pellet (membrane
fractions) were separated. The pellet was resuspended in PBS (500 µl). An aliquot (50
µl) of each fraction was analyzed by Bradford protein assay. Final protein
concentrations ranged from 0.5–0.9 µg / µl.
Spinach (5 g) was crushed in PBS (20 ml) and the biomass concentrated by
centrifugation (5 min at 14000 X g). The supernatant was discarded. The pellet was
resuspended in PBS buffer (5 ml, and an aliquot (50 µl) was reserved for Bradford
assay. The final protein concentration was 1.0–2.5 µg / µl.
Bradford assay: BSA stock slolution (30 µl, 10 mg / ml) was diluted to 300 µl (1 mg
/ ml) in PBS. Six concentrations of BSA (0 mg / ml, 1 mg / ml, 0.5 mg / ml 0.25 mg /
ml 0.125 mg / ml, 0.0625mg / ml) were prepared as standards. Commassie Protein
Assay Reagent (250 µl) was added to microplate wells. 20 µl each of standard and
unknown sample were mixed with Commassie Protein Assay Reagent in each well.
Each protein solution was assayed in duplicate. After incubation at room temperature
for 5 min, absorbance was measured at 595 nm (SynergyTM 2, Bioteck Instrument,
Inc).
Incubation of protein homogenates with photoaffinity probe or photoaffinity
label: The PbTx-2 photoaffinity probe (10) or label (8) were incubated at room
temperature with the soluble protein or membrane fractions at a probe/protein ratio of
0.2 µg probe per mg protein for 1 h. After incubation, the mixture was photolyzed
(Vilber Lourmat transilluminator, Model FLX-20M) for 20 min at 312 nm. The
soluble fraction was applied directly to a suspension of streptavidin beads (200 µl,

47
Dynabeads, 2.8 mm or 1.0 mm, Life Technologies). Triton X-100 (1 %, 200 µl) was
added to the membrane fractions (200 µl). The mixture was vortexed and centrifuged
for 5 min at 14 000 X g and applied to a suspension of streptavidin beads (200 µl).
After incubation for 30 min at room temperature, the beads were separated from the
solution by using a strong magnet and washed with PBS buffer (4 x 200 µl). Each
wash was analyzed for protein content, and the final wash contained no detectable
protein. Protein was eluted from the beads by using 0.1% SDS solution (3 x 250 µl) at
100◦C for 5 min. An aliquot of each elution was analyzed by Bradford protein assay,
concentrated to dryness by lyophilization (FreeZone, Model 2.5, 86 L/min), and
resuspended in water (10 µl) and loading buffer (2 µl, 0.25M Tris-HCl, pH 6.8, 15%
SDS, 20% glycerol, 25% mercaptoethanol, 0.01% Bromophenol Blue) for analysis by
SDS-PAGE (12% acryalamide) with Tris·glycine SDS, pH 8.3 running buffer.
Acrylamide gels were stained with PAGE Blue (Fermentas), according to the
manufacturer’s instructions. The 25 kD band was excised from the SDS-PAGE gel.
In-gel digestion: In-gel digestion and LC-ESI MS/MS analysis was performed by the
Proteomics and Mass Spectrometry Facility at the UMass Medical School. Gel slices
were cut into 1x1 mm pieces and placed in 1.5 ml Eppendorf tubes with water (1 ml)
for 30 min. The water was removed, and ammonium bicarbonate (50 µl, 250 mM)
was added. For reduction, a solution of 1,4-dithiothreitol (DTT; 5 µl, 45 mM) was
added, and the samples were incubated at 50ºC for 30 min. The samples were cooled
to room temperature and, for alkylation, iodoacetamide solution (5 µl, 100 mM) was
added and allowed to react for 30 min. The gel slices were washed with water (20 X 1
ml). The water was removed, ammonium bicarbonate/acetonitrile (50:50, 1 ml, 50
mM) was placed in each tube, and the samples were incubated at room temperature

48
for 1 h. The solution was removed, and acetonitrile (200 µl) was added to each tube,
at which point the gels slices turned opaque white. The acetonitrile was removed, and
the gel slices were further dried in a Speed Vac. Gel slices were rehydrated in trypsin
(50 µl, 2 ng / µl, Sigma) in 0.01% ProteaseMAX Surfactant (Promega) / 50 mM
ammonium bicarbonate. Additional bicarbonate buffer was added to ensure complete
submersion of the gel slices. Samples were incubated at 37 ºC for 21 h. The
supernatant of each sample was removed and placed in a separate 1.5 ml Eppendorf
tube. Gel slices were further dehydrated with acetonitrile/1% formic acid (80:20, 100
µl). The extract was combined with the supernatants of each sample. The samples
were dried in a Speed Vac. Samples were dissolved in 25 µl of 5% acetonitrile in 0.1%
trifluoroacetic acid prior to injection on LC-MS/MS.
LC-MS/MS on Q Exactive: For peptide sequencing, a 3.0 µl aliquot was directly
injected onto a custom-packed 2 cm X 100 mm C18 Magic 5 m particle trap column.
Labeled peptides were eluted and sprayed from a custom-packed emitter (75 µm X 25
cm C18 Magic 3 µm particle) with a linear gradient from 95% solvent A (0.1% formic
acid in water) to 35% solvent B (0.1% formic acid in acetonitrile) over 40 min at a
flow rate of 300 nL/min on a Waters Nano Acquity UPLC system. Data dependent
acquisitions were performed on a Q Exactive mass spectrometer (Thermo Scientific),
according to an experiment in which full MS scans from 300–1750 m/z were acquired
at a resolution of 70,000, followed by 12 MS/MS scans acquired under higher-energy
collisional dissociation (HCD) fragmentation at a resolution of 35 000 with an
isolation width of 1.2 Da.

49
Database searching: Raw data files were processed with Proteome Discoverer (v.
1.3). Charge state deconvolution and deisotoping were not performed. All MS/MS
samples were analyzed by using Mascot (Matrix Science, London, UK; v. 2.4.1).
Mascot was set up to search the NCBInr_20130907 database (32265160 entries),
assuming the digestion enzyme strict trypsin. Mascot was searched with a fragment
ion mass tolerance of 0.050 Da and a parent ion tolerance of 10.0 ppm. The
carbamidomethyl of cysteine was specified in Mascot as a fixed modification.
Gln >pyro-Glu of the N terminus, oxidation of methionine, and acetylation of the N
terminus were specified in Mascot as variable modifications.
Criteria for protein identification: Scaffold (v. 4.1.1, Proteome Software Inc.,
Portland, OR) was used to validate MS/MS-based peptide and protein identifications.
Peptide identifications were accepted if they could be established at greater than 80.0 %
probability by the Peptide Prophet algorithm (Keller, Nesvizhskii et al. 2002) with
Scaffold delta-mass correction. Protein identifications were accepted if they could be
established at greater than 90.0% probability and contained at least two identified
peptides. Protein probabilities were assigned by the Protein Prophet algorithm
(Nesvizhskii, Keller et al. 2003). Proteins that contained similar peptides and could
not be differentiated based on MS/MS analysis alone were grouped to satisfy the
principles of parsimony. Proteins sharing significant peptide evidence were grouped
into clusters.

50
Chapter 3 The effects of brevetoxins on the thioredoxin reductase/thioredoxin
system and related enzymes.
3.1 Objective
The findings described in the previous chapter indicated that brevetoxin binds to Trx.
This finding promoted us to examine how brevetoxin affects the TrxR/Trx system.
Because Trx is present in all living organisms and is key to regulation of the activity
of numerous enzymes and enzyme systems, this work can contribute to our
understanding of the key role of berevetoxin in K. brevis cells and mechanism of
toxicity to other organisms.
3.2 Introduction
Trx is the parent of a family of enzymes which share structural (but not necessarily
sequence) homology and function through thiol-disulfide exchange. The thioredoxin
superfamily is responsible for maintenance of the redox state within cells through the
reduction of thiol disulfide bridges. The thioredoxin superfamily includes the Trx 1
and Trx 2 (Trx A and Trx C) in the cytoplasm (Miranda-Vizuete, Damdimopoulos et
al. 1997, Stewart, Åslund et al. 1998) and glutaredoxins 1, 2, 3 (GrxA, GrxB, GrxC)
(Aslund, Ehn et al. 1994). A number of eukarytoic proteins such as protein disulfide
isomerase (PDI) also belong to the thioredoxin superfamily (Carvalho, Fernandes et
al. 2006). Some bacterial proteins like E.coli DsbA (a bacterial thiol disulfide
oxidoreductase) also contains thioredoxin domains (Carvalho, Fernandes et al. 2006).
Although those proteins do not all share much similarity in sequence, all proteins
have a common sequence CXXC motif in the active site (Table 5). This motif is a
highly conserved region which presents at the end of an α–helix in all proteins. The
amino acids between these two cysteines vary within the thioredoxin superfamily

51
(Carvalho, Fernandes et al. 2006). In thioredoxin the sequence is CGPC, CPYC in
glutaredoxins, CPHC in PDIs, and CGHC in DsbA.
Table 5 The motifs in thioredoxin , glutaredoxin, PDI and DsbA (Holmgren, Söderberg et al. 1975)
Enzyme CXXC motif Function Thioredoxin CGPC Reductant Glutaredoxin CPYC Reductant
PDI CHGC Oxidant Isomerase DsbA CPHC Oxidant
Trx catalyzes the disulfide reduction as a bimolecular nucleophilic substitution
reaction (SN2). During this process, the disulfide bond of the target is transferred to
Trx. In other words, the substrate protein acquires the electrons from Trx and Trx
becomes oxidized to the disulfide. The N-terminal cysteine of this CXXC motif is
deprotonated under physiological conditions and exposed to the aqueous
surroundings. This N-terminal cysteine of the motif serves as the nucleophilic
cysteine. The other cysteine in the motif is usually buried in the enzyme and
protonated (Carvalho, Fernandes et al. 2006). In all enzymes, the nucleophilic
cysteine must be deprotonated to participate in thiol-disulfide exchange. Typically, a
disulfide bond of the substrate is attacked by the N-terminal cysteine thiolate to form
a mixed disulfide bond (Holmgren 1995, Jeng, Reymond et al. 1998). The thiolate on
the C-terminal cysteine is stabilized with two backbone amide hydrogen bonds,
lowering the pKa (Roos, Foloppe et al. 2009). N-terminal cysteine is attacked by C-
terminal cysteine thiolate to form the Trx disulfide. A reduced substrate protein and
oxidized Trx are the products of this exchange reaction (Figure 15).

52
Figure 15 Trx catalyzes the reaction as a bimolecular nucleophilic substitution reaction (SN2)
The reaction between Trx and its substrate is stoichiometric. Oxidized Trx is more
stable than reduced Trx (Collet and Messens 2010). It has been proposed that the rate
of the thiol- disulfide exchange is affected by several factors, such as the pKa of the
nucleophilic cysteine (Shaked, Szajewski et al. 1980), the local electrostatic
environment of the nearby amino acid (Bulaj, Kortemme et al. 1998, Hansen,
Østergaard et al. 2005), the molecular strain (Østergaard, Henriksen et al. 2001), the
pH of the surrounding solvent (Setterdahl, Chivers et al. 2003) and the pKa values of
the leaving thiol groups of the oxidized substrate (Shaked, Szajewski et al. 1980,
Jensen, Hansen et al. 2009).
Over the past 50 years, Trx has been studied extensively. The first Trx was discovered
in 1964 in E. coli as an electron donor to ribonucleotide reductase, an essential
enzyme for DNA synthesis (Laurent, Moore et al. 1964, Moore, Reichard et al. 1964).
Since then, Trx has been determined to play multiple cellular roles, such as reductase
in redox control (Holmgren 1985), protecting proteins from oxidative damage and
inactivation (Holmgren 1985, Holmgren 1989, Holmgren 1995) and helping the cells
to combat various environmental stresses (reactive oxygen species (ROS),
peroxynitrite, arsenate) (Landino, Skreslet et al. 2004). Some Trxs can also regulate
cell death via denitrosylation and modulate the inflammatory response (Rahman and
Adcock 2006). In addition, it has been shown that Trx plays an essential role in the
life cycle of viruses. Trxs are located in various cellular compartments. For instance, a

53
number of Trxs are found in the cytosol (Arnér and Holmgren 2000), while others are
abundant in nucleus or mitochondria (Hirota, Matsui et al. 1997, Hirota, Murata et al.
1999). Still others are associated with the cell membrane (Martin and Dean 1991).
In 1964 the Trx/TrxR system was determined to be comprised of Trx, together with
thioredoxin reductase (TrxR) and NADPH (Laurent, Moore et al. 1964). In general,
the electrons which are provided by NADPH are transferred to the active site of Trx
via TrxR. The disulfide of oxidized Trx must be reduced back to a dithiol to regain its
active form. The enzyme TrxR, with reducing equivalents ultimately derived from
NADPH (Figure16) performs this reduction.
Figure 16 Electron flow of E. coli TrxR/Trx system
TrxR is a member of the pyridine nucleotide disulfide oxidoreductase family that
includes glutathione reductase (GR) and lipoamide dehydrogenase (LipDH)
(Mustacich and Powis 2000). Both TrxR and GR use NADPH as a cofactor via FAD
to transfer electrons to the redox centers, while LipDH uses NADH instead of
NADPH (Williams Jr 1992). In addition, two types of TrxR have been identified.
Although they both have same functions, the amino acid sequence and catalytic
mechanisms are distinct (Gromer, Schirmer et al. 1999, Mustacich and Powis 2000).
The type of TrxR with a low molecular weight (~35 kDa) has been characterized in

54
Archaea, bacteria and Eukarya (Becker, Gromer et al. 2000). There are two binding
domains in E. coli TrxR: FAD binding domain and NADPH binding domain. The
connection of these two domains is not close (Williams 1995). In E. coli TrxR the
FAD domain is rotated 66 degrees with NADPH domain remaining, so that the two
prosthetic groups move into close contact. The conformational change allows
electrons to pass from NADPH to FAD then to reduce the disulfide bond (Lennon and
Williams 1997, Mustacich and Powis 2000). The conserved active site residues are –
Cys-Ala-Thr-Cys- in the redox center of E. coli TrxR.
A high molecular weight (~55 kDa) TrxR is often referred to as mammalian as it was
first identified in Homo sapiens. Subsequently, this high molecular weight TrxR has
been found to be widespread in the animal kingdom (Mustacich and Powis 2000,
Bauer, Gromer et al. 2003). Mammalian TrxR is distinct from bacterial TrxR by
virtue of a third redox center in a C-terminal extension of the enzyme consisting of a
cysteine and a selenocysteine (Sec) (Cys497-Sec498-Gly- in human TrxR). The
purification of rat liver Trx and TrxR demonstrated that mammalian TrxR is larger
than mammalian Trx. Mammalian TrxR has broader substrate specificity than
prokaryotic forms (Luthman and Holmgren 1982, Arnér 2009). Mammalian TrxR is a
large selenoenzyme, which has three isoforms: TrxR1 in the cytosol (Arnér 2009),
TrxR2 in mitochondria and TrxR3 or thioredoxin glutathione reductase (TGR)
(testis)(Lillig and Holmgren 2007). TGR has common properties to other flavoprotein
oxidoreductase. However, TGR is the unique enzyme that connects thioredoxin and
glutathione system. TGR has an extra reaction sites (CVNVGC) at the N-terminal.
The reducing equivalents are transferred from NADPH to the FAD. Then, the
adjacent cysteine in the reaction site (CVNVGC) at C-terminal. The reduced cysteine
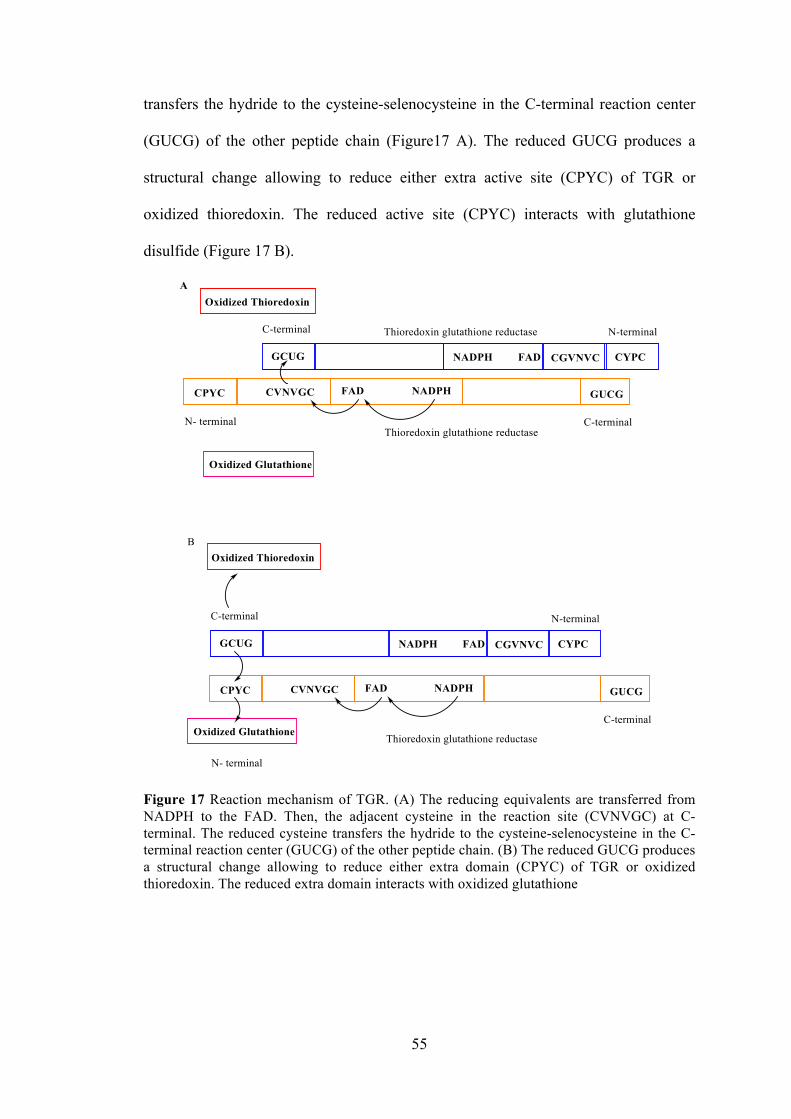
55
transfers the hydride to the cysteine-selenocysteine in the C-terminal reaction center
(GUCG) of the other peptide chain (Figure17 A). The reduced GUCG produces a
structural change allowing to reduce either extra active site (CPYC) of TGR or
oxidized thioredoxin. The reduced active site (CPYC) interacts with glutathione
disulfide (Figure 17 B).
Figure 17 Reaction mechanism of TGR. (A) The reducing equivalents are transferred from NADPH to the FAD. Then, the adjacent cysteine in the reaction site (CVNVGC) at C-terminal. The reduced cysteine transfers the hydride to the cysteine-selenocysteine in the C-terminal reaction center (GUCG) of the other peptide chain. (B) The reduced GUCG produces a structural change allowing to reduce either extra domain (CPYC) of TGR or oxidized thioredoxin. The reduced extra domain interacts with oxidized glutathione

56
It has been found that the selenium is the essential active site residue in mammalian
TrxR. Blocking or mutation at the selenium active site results in a loss of ability to
reduce Trx (Figure18).
Figure 18 Electron flow of mammalian TrxR/Trx system The Trx/ TrxR system plays a role as a general protein disulfide reductase. It is
involved in regulating DNA synthesis, gene transcription, cell growth and metabolism
(Krauth-Siegel and Coombs 1999, Arnér and Holmgren 2000, Mustacich and Powis
2000, Williams 2000). The Trx/TrxR system also plays a key role in biochemical
mechanisms associated with many human diseases (Figure19). For instance, Trx and
TrxR overexpression is observed in many cancer cells, which has been hypothesized
to be a protection against apoptosis (Baker, Payne et al. 1997, Powis, Mustacich et al.
2000, Lincoln, Ali et al. 2002, Urig and Becker 2006, Anestål, Prast-Nielsen et al.
2008). TrxR is an important target for many cancer treatments. Cisplatin and arsenic
trioxide as clinical drugs are used to inhibit TrxR resulting in blocking or mediating
the activity of DNA synthesis and defense against oxidative stress (Lu, Chew et al.
2007, Arnér 2009).

57
Figure 19 The Trx/TrxR system plays a key role in biochemical processes associated with many human diseases
In most organisms in addition to Trx/TrxR system, glutathione/ glutathione disulfide
(GSH/GSSG) is a major pathway for regulating cellular redox homeostasis and
antioxidant defense. The mammalian GSH/GSSG system involved GSH recycling of
oxidized GSSG catalyzed by GR using NADPH as electron donor The N-terminal
domains of GR resemble those of TrxR , including the FAD prosthetic group, the
NADPH binding domain and the N-terminal redox-active CVNVGC motif (Arscott,
Gromer et al. 1997). However, GR lacks the C-terminal redox center (GCUG) of
TrxR (Lu and Holmgren 2014).
In addition to Trx, Grx and glutathione peroxidase (GPx) also belong to the family of
oxidoreductases which function through thiol-disulfide exchange. These proteins have
similar three-dimensional structures and contain an active site dithiol/disulfide group
which protrudes from a hydrophobic protein surface The disulfide bond in Grx is
reduced continually by GSH in the mammalian GSH/GSSG system (Figure 20).
While Trx and Grx show low sequence homology, they exhibit similar three-

58
dimensional topology including the so-called thioredoxin fold, consisting of an N-
terminal βαβ motif and a C-terminal ββα motif with the central β-sheets surrounded
by the α-helices and an active site CXXC motif, which protrudes from the
hydrophobic surface of the protein.(Martin 1995, Netto, de Oliveira et al. 2016)
. Figure 20 Electron flow of mammalian GSH/GSSG system
GPx catalyzes the reduction of ROS in cells, including hydrogen peroxide, by reduced
glutathione to protect the cells from oxidative damage. Moreover, most noteworthy is
that GPx is a selenoprotein. GPx has four identical subunits. Each subunit contains a
selenocysteine in the active site, which participates directly within the catalytic cycle.
The selenium undergoes a redox cycle involving the selenol (GPx-SeH) as an active
site to reduce hydrogen peroxides and other harmful peroxide, which oxidizes GPx-
SeH to selenenic acid (GPx-SeOH). A reduced GSH reacts with GPx-SeOH to form
selenenyl sulfide adduct (GPx-SeSG). A second GSH is the ultimate electron donor to
regenerate the reduced form of the selenocysteine by attacking the GPx-SeSG to form
the oxidized GSSG (Figure21) (Morgenstern 2005, Toppo, Vanin et al. 2008).

59
Figure 21 The redox cycle of GPx coupled with mammalian GSH/GSSG system
3.3 Results
3.3.1 Modified papain assay
The modified papain assay is a sensitive spectrophotometric assay that is commonly
used for the quantitation of thiol groups. The assay involves the reaction of a thiol
with an inactive mixed disulfide of papain, a cysteine protease, resulting in the
stoichiometric formation of active papain (Figure 22) (Singh, Blattler et al. 1993). The
active site cysteine of papain is converted to an inactive mixed disulfide which may
be reduced by low MW thiols to the active form. The active papain then catalyzes the
hydrolysis of N-benzoyl-L-arginine-p-nitroanilide to yield p-nitroanilide.
Figure 22 The equation of modified papain assay
The chromogenic product can be detected at 412 nm. Furthermore, the addition of the
disulfide cystamine to the assay allows for the detection of inaccessible protein thiols.
Cystamine is reduced to cysteamine by protein thiols and the resulting cysteamine
acts as a shuttle between high MW inaccessible protein thiols and modified papain.

60
Thus the difference in papain activity with and without cystamine is a reflection of
high MW inaccessible thiols. The initial amount of thiol can be obtained from the
proportion of absorbance compared to the concentration of thiol standards. Here, this
method can also be used to determine the content of reduced Trx. Because the
reaction of Trx with it’s substrates is stoichiometric, the modified papain assay can be
used to amplify the signal of Trx. Trx is used to reduce the small disulfide cystamine
to cysteamine, which will in turn reduce the papain disulfide stoichiometrically
(Figure23). Thus the modified papain assay may be used to assay for Trx activity by
amplification of the Trx signal.
Figure 23 The principle of modified papain assay to determine Trx activity
3.3.2 Papain assay
The effect of PbTx-2 on Trx was examined using the modified papain assay.
Inhibition of this two enzyme (Trx/papain) system was observed in the presence of
PbTx-2 (15 µM) when compared to the positive control reaction (no PbTx) whether
the PbTx-2 was pre-incubated with Trx or not pre-incubated (Figure24).
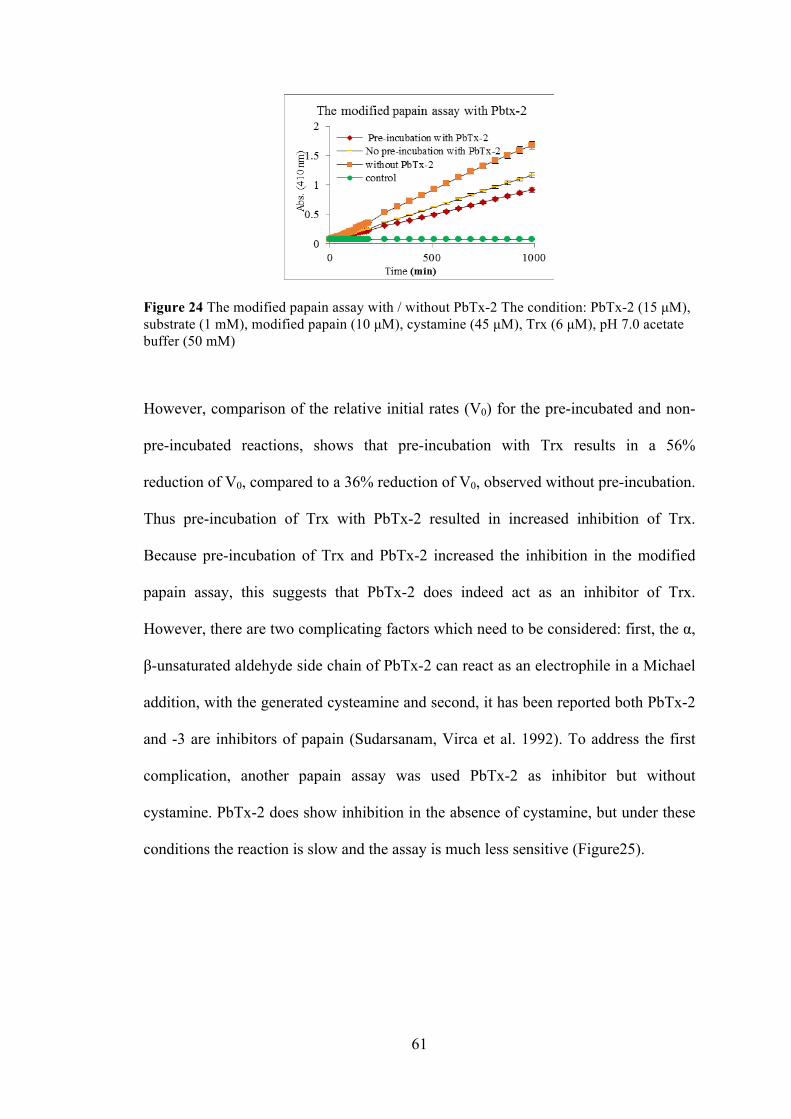
61
Figure 24 The modified papain assay with / without PbTx-2 The condition: PbTx-2 (15 µM), substrate (1 mM), modified papain (10 µM), cystamine (45 µM), Trx (6 µM), pH 7.0 acetate buffer (50 mM)
However, comparison of the relative initial rates (V0) for the pre-incubated and non-
pre-incubated reactions, shows that pre-incubation with Trx results in a 56%
reduction of V0, compared to a 36% reduction of V0, observed without pre-incubation.
Thus pre-incubation of Trx with PbTx-2 resulted in increased inhibition of Trx.
Because pre-incubation of Trx and PbTx-2 increased the inhibition in the modified
papain assay, this suggests that PbTx-2 does indeed act as an inhibitor of Trx.
However, there are two complicating factors which need to be considered: first, the α,
β-unsaturated aldehyde side chain of PbTx-2 can react as an electrophile in a Michael
addition, with the generated cysteamine and second, it has been reported both PbTx-2
and -3 are inhibitors of papain (Sudarsanam, Virca et al. 1992). To address the first
complication, another papain assay was used PbTx-2 as inhibitor but without
cystamine. PbTx-2 does show inhibition in the absence of cystamine, but under these
conditions the reaction is slow and the assay is much less sensitive (Figure25).

62
Figure 25 The modified papain assay with / without PbTx-2 but without cystamine. The condition: PbTx-2 (15 µM), substrate (1 mM), modified papain (10 µM), Trx (6 µM), pH 7.0 acetate buffer (50 mM)
Thus, the rate of the reaction in the presence of Pbtx-3 (which does not react with
cysteamine) was determined to see if the effect of brevetoxin is due to reaction with
cysteamine. It is clear that the Pbtx-3 impacts the rate (Figure 26).
Figure 26 The modified papain assay with / without PbTx-3 with cystamine. The condition: Pbtx-3 (15 µM), substrate (1 mM), modified papain (10 µM), cystamine (45 µM), Trx (6 µM), pH 7.0 acetate buffer (50 mM)
In order to understand the inhibition of papain by Pbtx-2 and Pbtx-3, a standard
papain assay which didn’t contain the cystamine was performed in the presence of
PbTx-2 and PbTx-3 (Figure27). Figure 27 shows that both brevetoxins inhibit papain.
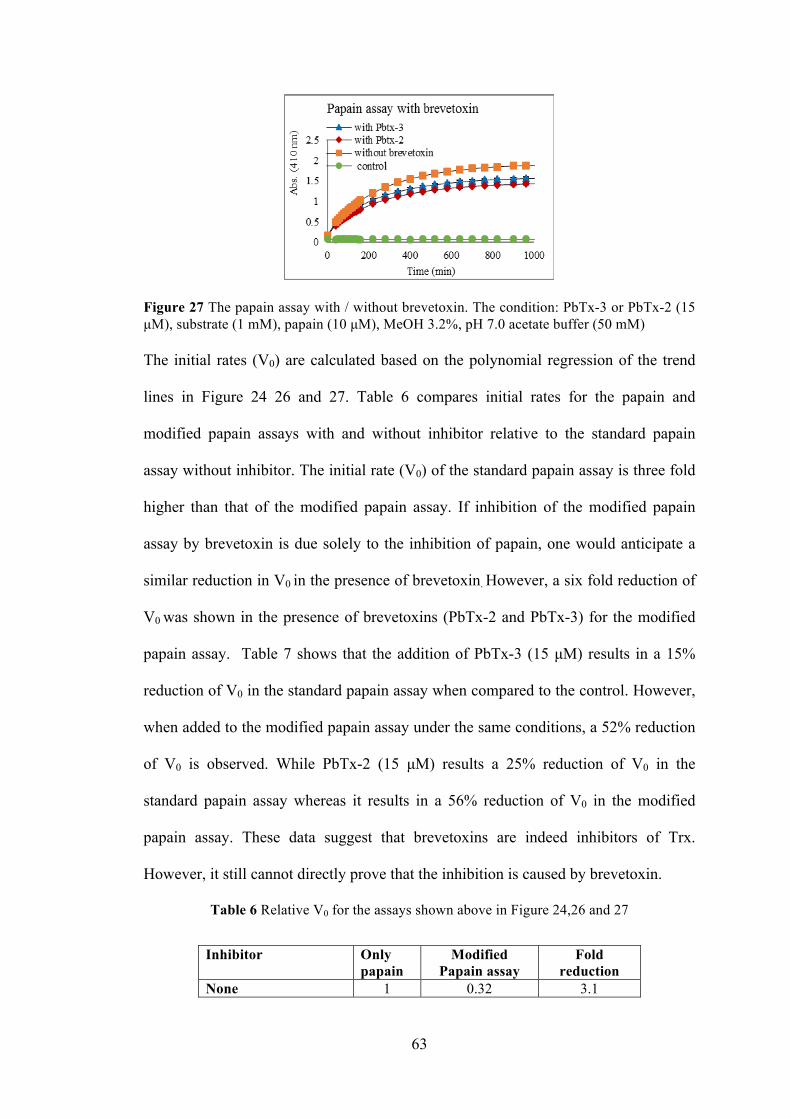
63
Figure 27 The papain assay with / without brevetoxin. The condition: PbTx-3 or PbTx-2 (15 µM), substrate (1 mM), papain (10 µM), MeOH 3.2%, pH 7.0 acetate buffer (50 mM) The initial rates (V0) are calculated based on the polynomial regression of the trend
lines in Figure 24 26 and 27. Table 6 compares initial rates for the papain and
modified papain assays with and without inhibitor relative to the standard papain
assay without inhibitor. The initial rate (V0) of the standard papain assay is three fold
higher than that of the modified papain assay. If inhibition of the modified papain
assay by brevetoxin is due solely to the inhibition of papain, one would anticipate a
similar reduction in V0 in the presence of brevetoxin. However, a six fold reduction of
V0 was shown in the presence of brevetoxins (PbTx-2 and PbTx-3) for the modified
papain assay. Table 7 shows that the addition of PbTx-3 (15 µM) results in a 15%
reduction of V0 in the standard papain assay when compared to the control. However,
when added to the modified papain assay under the same conditions, a 52% reduction
of V0 is observed. While PbTx-2 (15 µM) results a 25% reduction of V0 in the
standard papain assay whereas it results in a 56% reduction of V0 in the modified
papain assay. These data suggest that brevetoxins are indeed inhibitors of Trx.
However, it still cannot directly prove that the inhibition is caused by brevetoxin.
Table 6 Relative V0 for the assays shown above in Figure 24,26 and 27
Inhibitor Only papain
Modified Papain assay
Fold reduction
None 1 0.32 3.1

64
Brevetoxin-2 (15 µM) 0.75 0.12 6.3 Brevetoxin-3 (15 µM) 0.85 0.15 5.6
Table 7 The inhibition % for the assays shown above in Fig 24,26 and 27
Only papain Modified papain assay PbTx-2 25% 56% PbTx-3 15% 52%
3.3.3 Fluorescent assay for Trx/TrxR system
As an alternative, the activity of Trx can be amplified at the front end by including
TrxR and NADPH in the reaction to continuously reduce the Trx. The method used is
based on the reduction of insulin by reduced Trx with NADPH as ultimate electron
donor via TrxR (Montano, Lu et al. 2014). The insulin is labeled with fluorescein
isothiocyanate-labeled (FiTC). These FiTC-modified insulin servers as the substrate.
Insulin is composed of two peptide chains which are the chain A and chain B. It is
found that the fluorescence is quenched by the disulfides between chain A and B.
However, the fluorescence is increased on reduced form of insulin (Heuck and
Wolosiuk 1997). Here, the disulfides are able to be reduced by Trx /TrxR system, the
fluorescence of FiTC is record after 30 min incubation with the mixture of Trx, TrxR
and NADPH (Figure 28) (Montano, Lu et al. 2014).
Figure 28 The reaction in sensitive fluorescent assay for Trx/TrxR system
To determine if brevetoxin acts as inhibitor of Trx, the inhibition of Trx by PbTx-2
and -3 was determined using a commercially available assay kit (Montano, Lu et al.
2014). PbTx-2 and PbTx-3 at concentrations ranging from 10 to 35 µM were
incubated with a mixture of pre-reduced TrxR and Trx for one hour. After adding
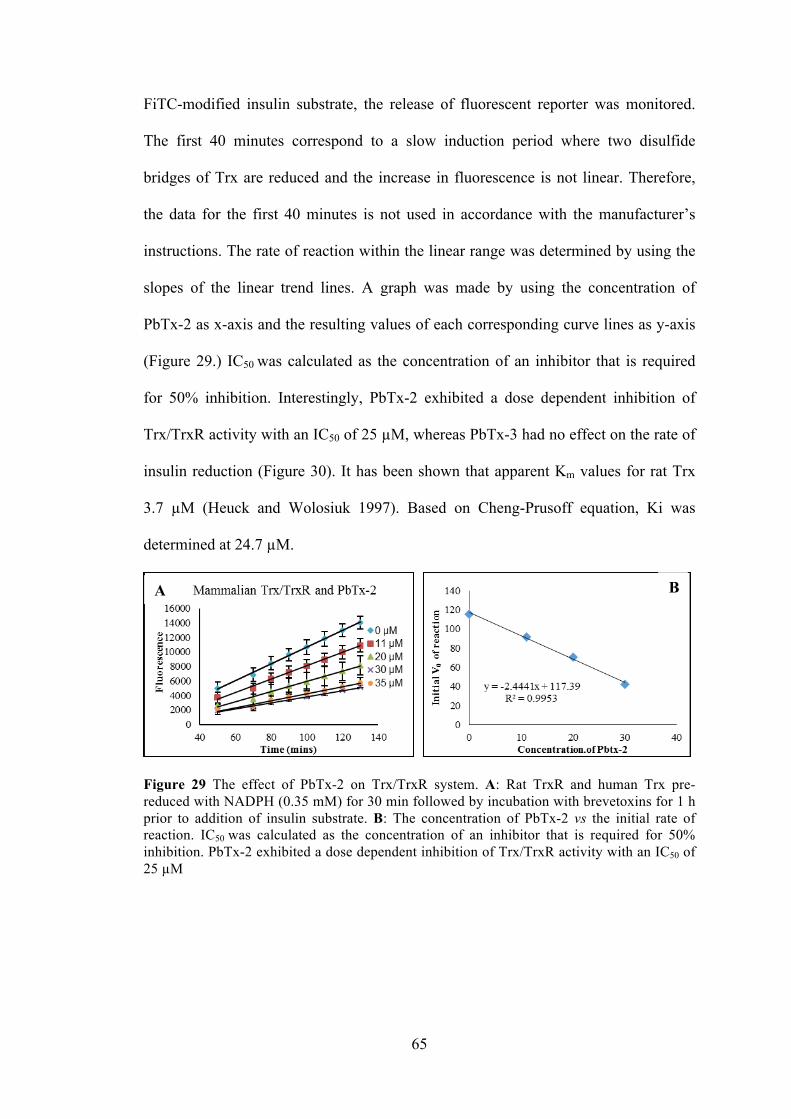
65
FiTC-modified insulin substrate, the release of fluorescent reporter was monitored.
The first 40 minutes correspond to a slow induction period where two disulfide
bridges of Trx are reduced and the increase in fluorescence is not linear. Therefore,
the data for the first 40 minutes is not used in accordance with the manufacturer’s
instructions. The rate of reaction within the linear range was determined by using the
slopes of the linear trend lines. A graph was made by using the concentration of
PbTx-2 as x-axis and the resulting values of each corresponding curve lines as y-axis
(Figure 29.) IC50 was calculated as the concentration of an inhibitor that is required
for 50% inhibition. Interestingly, PbTx-2 exhibited a dose dependent inhibition of
Trx/TrxR activity with an IC50 of 25 µM, whereas PbTx-3 had no effect on the rate of
insulin reduction (Figure 30). It has been shown that apparent Km values for rat Trx
3.7 µM (Heuck and Wolosiuk 1997). Based on Cheng-Prusoff equation, Ki was
determined at 24.7 µM.
Figure 29 The effect of PbTx-2 on Trx/TrxR system. A: Rat TrxR and human Trx pre-reduced with NADPH (0.35 mM) for 30 min followed by incubation with brevetoxins for 1 h prior to addition of insulin substrate. B: The concentration of PbTx-2 vs the initial rate of reaction. IC50 was calculated as the concentration of an inhibitor that is required for 50% inhibition. PbTx-2 exhibited a dose dependent inhibition of Trx/TrxR activity with an IC50 of 25 µM
A
B

66
Figure 30 The effect of PbTx-3 on Trx/TrxR system. A: Rat TrxR and human Trx pre-reduced with NADPH (0.35 mM) for 30 min followed by incubation with brevetoxins for 1 h prior to addition of insulin substrate. B: The concentration of PbTx-2 vs the initial rate of reaction. However, PbTx-3 had no effect on the rate of insulin reduction
3.3.4 Ellman’s assay
Ellman’s assay utilizes 5, 5’-dithio-bis-(2-nitrobenzoic acid), also known as DTNB, to
react with free sulfhydryl group to yield a mixed disulfide and 2-nitro-5-thiobenzoic
acid (TNB). TNB is a measurable yellow-colored product (Figure 31) which can be
detected at 412 nm (Riener, Kada et al. 2002). To detect the Trx or TrxR activity,
reduced Trx or TrxR can be used to reduce DTNB producing the yellow TNB which
can be detected at 412 nm.
Figure 31 The reduction of Ellman’s reagent
Because the Trx/TrxR assay is a two enzyme system (Trx/TrxR), it was still not clear
if the inhibition observed in the presence of PbTx-2 is due to the inhibition of Trx, or
TrxR or both. Unlike bacterial TrxR, which shows high substrate selectivity for Trx,
mammalian TrxR reduces a broad range of substrates, including small disulfides such
as DTNB, making it possible to assay for TrxR activity in the absence of Trx. Thus,
A
B

67
the activity of TrxR alone in the presence and absence of PbTx-2 was determined by
using Ellman’s reagent.
PbTx-2 (22 µM) was incubated with the rat TrxR in the presence of NADPH. After
30 min, DTNB was added to the reaction mixture and the absorbance of TNB- was
monitored for 60 mins. Surprisingly, the Vo of DTNB reduction by TrxR was
enhanced 2.5 fold in the presence of PbTx-2 when compared to the control reaction
which lacked PbTx-2. (Figure32 A). The same result was obtained when PbTx-2 was
added simultaneously with DTNB. In other words, without pre-incubation of TrxR by
NADPH and PbTx-2. (Figure32 B)
Figure 32 The effect of PbTx-2 on the Ellman’s assay. (A) The effect of PbTx-2 on the DTNB reduction by pre-incubated TrxR. Rat TrxR was pre-reduced with NADPH (0.10 mM) for 30 min with PbTx-2 (18 µM), followed by addition of DTNB (1.8 mM). (B) The effect of PbTx-2 on the DTNB reduction by non-incubated TrxR. Rat TrxR was pre-reduced with NADPH (0.10 mM) for 30 min without PbTx-2 (18 µM), followed by addition of DTNB (1.8 mM)
That surprising result was reproducible and results triggered our curiosity: Why does
PbTx-2 enhance the activity of TrxR?
As mentioned earlier, mammalian TrxR is distinct from bacterial TrxR by virtue of a
third C-terminal redox center consisting of a cysteine and a selenocysteine (Sec or U).
The TrxRs which were used in all experiments to this point were mammalian. In the
A
B

68
enzymatic mechanism of Trx reduction by TrxR, NADPH provides the reducing
equivalents and initiates this cascade of redox reactions across three redox active sites:
FAD which is first reduced by NADPH, next FADH2 reduces an N-terminal disulfide
(in human TrxR:- Cys59-Val-Asn-Val-Gly-Cys64) which undergoes thiol disulfide
exchange with a C-terminal disulfide or seleno-sulfide (in human TrxR:-Gly-Cys497-
Sec498-Gly). The C-terminal redox center of TrxR undergoes thiol-disulfide exchange
with oxidized Trx (Figure 33).
Figure 33 The electron flow in C-terminal reaction center in mammalian TrxR
Furthermore, it is reported the activity of Sec is essential for mammalian TrxR to
catalyze the reduction of Trx (Lee, Bar-Noy et al. 2000, Zhong and Holmgren 2000).
When the Sec site is blocked or modified, the mammalian TrxR is unable to reduce
the oxidized Trx disulfide to activated Trx-(SH)2. Sec (U) is a selenium containing
analog of cysteine but with a stronger nucleophile, which makes Sec a highly reactive
amino acid. Thus many compounds such as quinol (Bradshaw, Matthews et al. 2005),
curcumin (Fang, Lu et al. 2005) and 4-hydroxy-2-nonenal (Fang and Holmgren
2006)(Figure34) inhibit TrxR as electrophilic compounds. These molecules can

69
function as electrophiles, or Michael acceptors, for the nucleophilic Cys or Sec
residues, resulting in irreversible inhibition of TrxR. Curcumin was found to alkylate
both the C-terminal cysteine and selenocysteine of TrxR. On the other hand, 4-
hydroxy-2-nonenal and quinols have been demonstrated to label the C-terminal
cysteine and selenocysteine of TrxR as well as the redox active cysteines of Trx.
Figure 34 The Michael acceptors demonstrated to inhibit TrxR Comparison of these compounds to PbTx-2, reveals that they have a common
functional group: an α, β-unsaturated carbonyl. Thus, it is assumed that the α, β-
unsaturated aldehyde K-ring side chain of PbTx-2 may function as an efficient
electrophile, reacting with Sec. Thus, if the TrxR and PbTx-2 form an adduct, this
would result in the irreversible inhibition of Trx reduction, which can explain the
reason PbTx-2 shows inhibition of the TrxR/Trx system towards insulin reduction
while the allylic alcohol, PbTx-3 does not (Figure35). This explanation is also
consistent with the observation that DTNB reduction by TrxR is actually enhanced in
the presence of PbTx-2. It has been previously demonstrated that small molecules
such as DTNB can be reduced by the N-terminal redox center while the C-terminal
center is alkylated (Cenas, Nivinskas et al. 2004, Eckenroth, Lacey et al. 2007). In the
reduction of Trx by TrxR, is has been shown that the reduction of the C-terminal
seleno-sulfide is the rate-determining step. Reduction of DTNB by the N-terminal
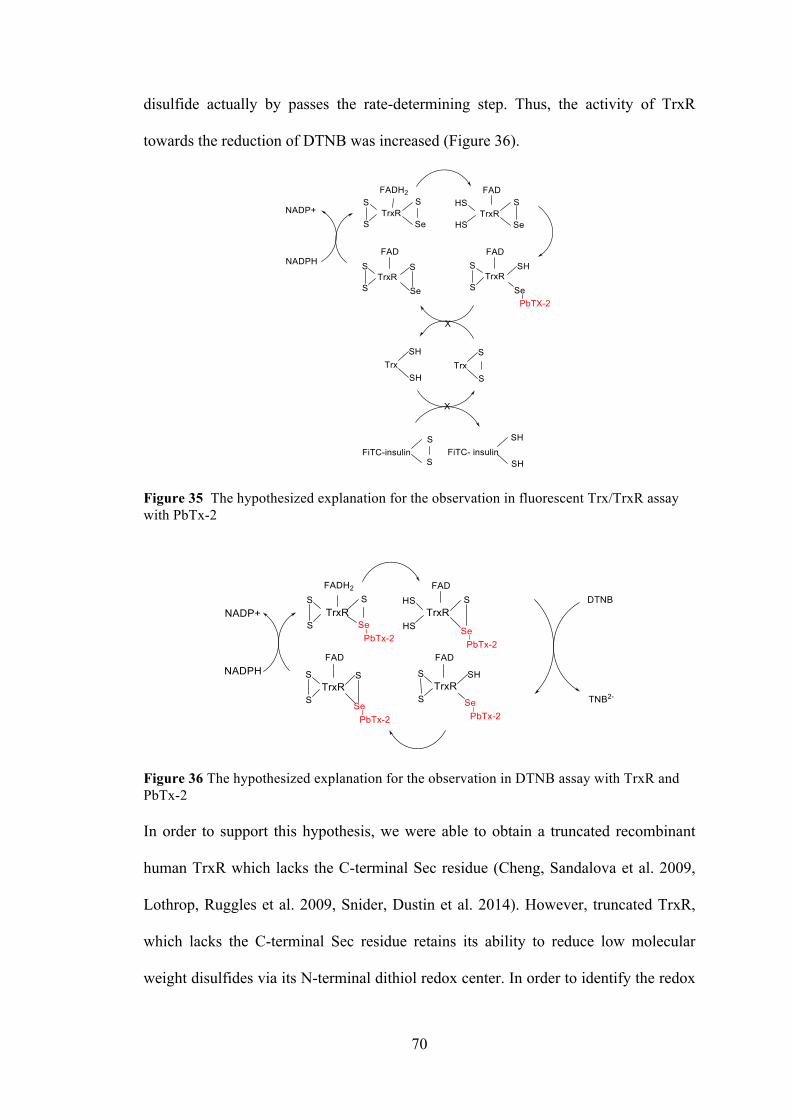
70
disulfide actually by passes the rate-determining step. Thus, the activity of TrxR
towards the reduction of DTNB was increased (Figure 36).
Figure 35 The hypothesized explanation for the observation in fluorescent Trx/TrxR assay with PbTx-2
Figure 36 The hypothesized explanation for the observation in DTNB assay with TrxR and PbTx-2 In order to support this hypothesis, we were able to obtain a truncated recombinant
human TrxR which lacks the C-terminal Sec residue (Cheng, Sandalova et al. 2009,
Lothrop, Ruggles et al. 2009, Snider, Dustin et al. 2014). However, truncated TrxR,
which lacks the C-terminal Sec residue retains its ability to reduce low molecular
weight disulfides via its N-terminal dithiol redox center. In order to identify the redox
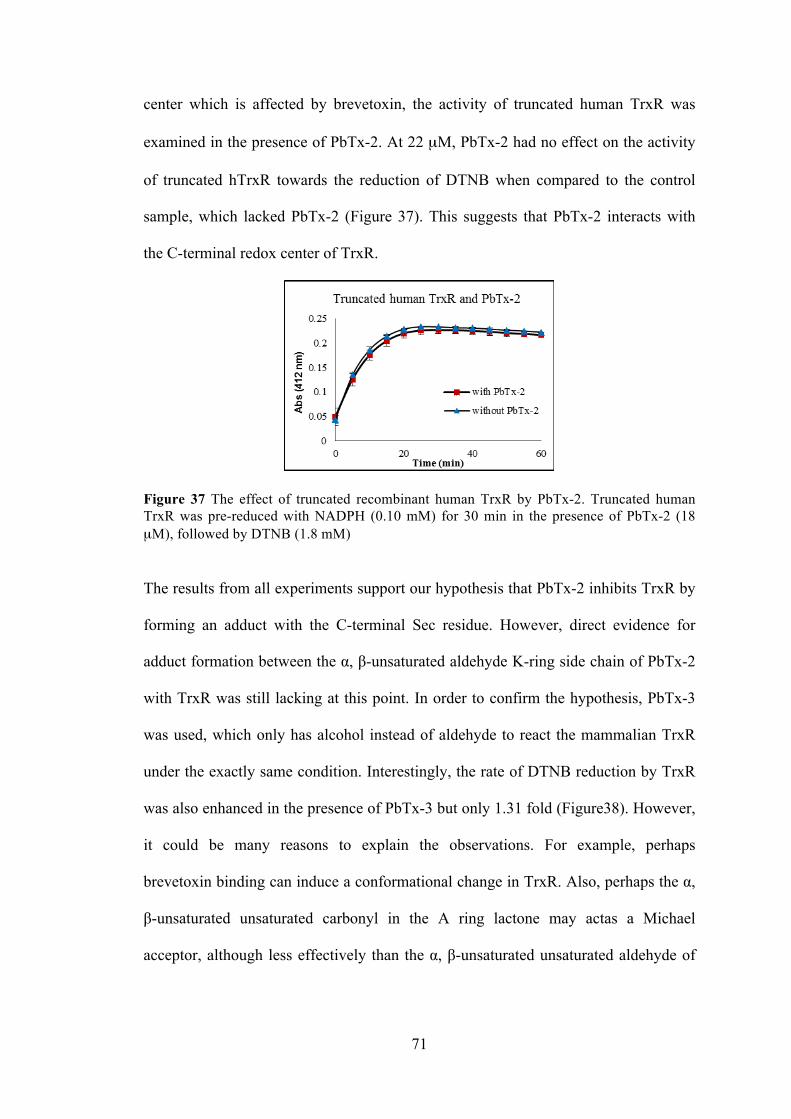
71
center which is affected by brevetoxin, the activity of truncated human TrxR was
examined in the presence of PbTx-2. At 22 µM, PbTx-2 had no effect on the activity
of truncated hTrxR towards the reduction of DTNB when compared to the control
sample, which lacked PbTx-2 (Figure 37). This suggests that PbTx-2 interacts with
the C-terminal redox center of TrxR.
Figure 37 The effect of truncated recombinant human TrxR by PbTx-2. Truncated human TrxR was pre-reduced with NADPH (0.10 mM) for 30 min in the presence of PbTx-2 (18 µM), followed by DTNB (1.8 mM)
The results from all experiments support our hypothesis that PbTx-2 inhibits TrxR by
forming an adduct with the C-terminal Sec residue. However, direct evidence for
adduct formation between the α, β-unsaturated aldehyde K-ring side chain of PbTx-2
with TrxR was still lacking at this point. In order to confirm the hypothesis, PbTx-3
was used, which only has alcohol instead of aldehyde to react the mammalian TrxR
under the exactly same condition. Interestingly, the rate of DTNB reduction by TrxR
was also enhanced in the presence of PbTx-3 but only 1.31 fold (Figure38). However,
it could be many reasons to explain the observations. For example, perhaps
brevetoxin binding can induce a conformational change in TrxR. Also, perhaps the α,
β-unsaturated unsaturated carbonyl in the A ring lactone may actas a Michael
acceptor, although less effectively than the α, β-unsaturated unsaturated aldehyde of
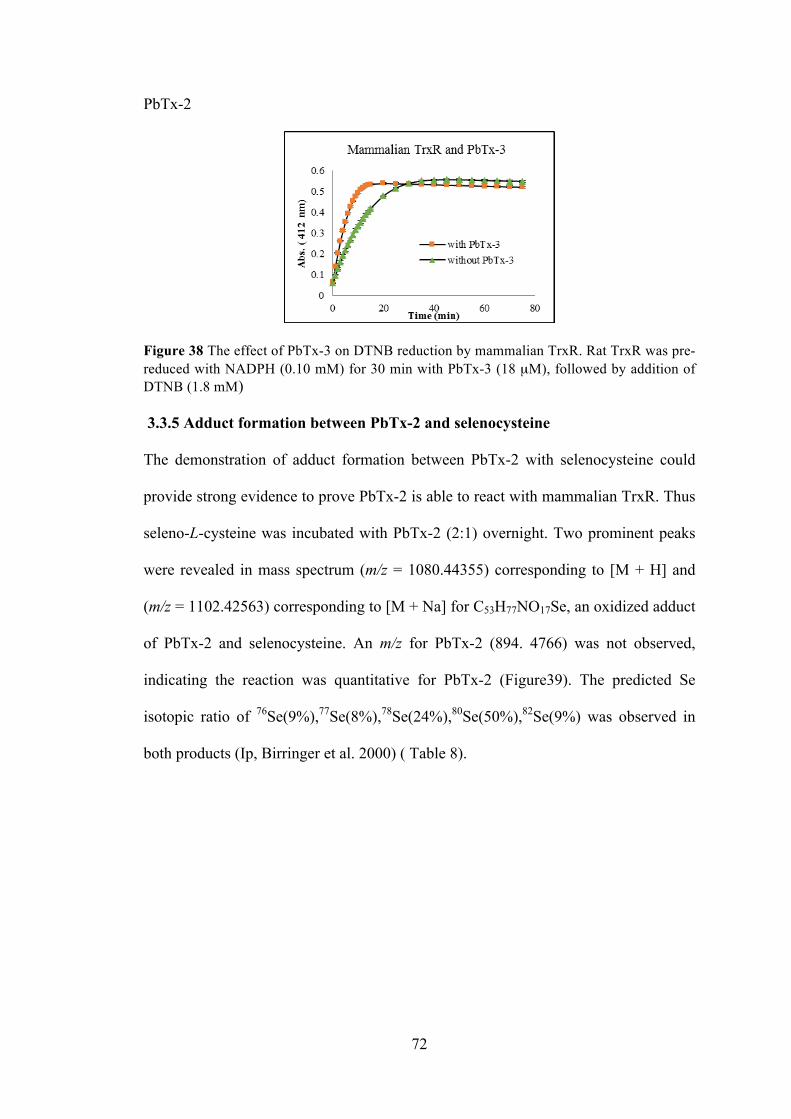
72
PbTx-2
Figure 38 The effect of PbTx-3 on DTNB reduction by mammalian TrxR. Rat TrxR was pre-reduced with NADPH (0.10 mM) for 30 min with PbTx-3 (18 µM), followed by addition of DTNB (1.8 mM) 3.3.5 Adduct formation between PbTx-2 and selenocysteine
The demonstration of adduct formation between PbTx-2 with selenocysteine could
provide strong evidence to prove PbTx-2 is able to react with mammalian TrxR. Thus
seleno-L-cysteine was incubated with PbTx-2 (2:1) overnight. Two prominent peaks
were revealed in mass spectrum (m/z = 1080.44355) corresponding to [M + H] and
(m/z = 1102.42563) corresponding to [M + Na] for C53H77NO17Se, an oxidized adduct
of PbTx-2 and selenocysteine. An m/z for PbTx-2 (894. 4766) was not observed,
indicating the reaction was quantitative for PbTx-2 (Figure39). The predicted Se
isotopic ratio of 76Se(9%),77Se(8%),78Se(24%),80Se(50%),82Se(9%) was observed in
both products (Ip, Birringer et al. 2000) ( Table 8).
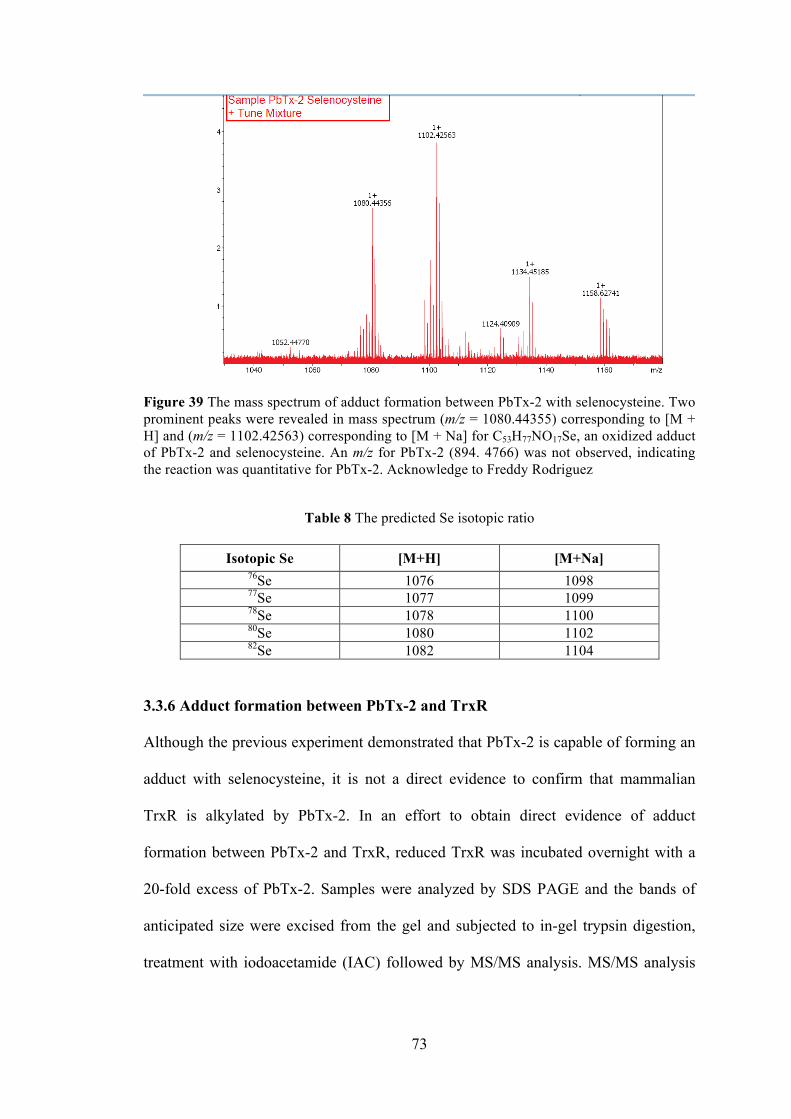
73
Figure 39 The mass spectrum of adduct formation between PbTx-2 with selenocysteine. Two prominent peaks were revealed in mass spectrum (m/z = 1080.44355) corresponding to [M + H] and (m/z = 1102.42563) corresponding to [M + Na] for C53H77NO17Se, an oxidized adduct of PbTx-2 and selenocysteine. An m/z for PbTx-2 (894. 4766) was not observed, indicating the reaction was quantitative for PbTx-2. Acknowledge to Freddy Rodriguez
Table 8 The predicted Se isotopic ratio
Isotopic Se [M+H] [M+Na]
76Se 1076 1098 77Se 1077 1099 78Se 1078 1100 80Se 1080 1102 82Se 1082 1104
3.3.6 Adduct formation between PbTx-2 and TrxR
Although the previous experiment demonstrated that PbTx-2 is capable of forming an
adduct with selenocysteine, it is not a direct evidence to confirm that mammalian
TrxR is alkylated by PbTx-2. In an effort to obtain direct evidence of adduct
formation between PbTx-2 and TrxR, reduced TrxR was incubated overnight with a
20-fold excess of PbTx-2. Samples were analyzed by SDS PAGE and the bands of
anticipated size were excised from the gel and subjected to in-gel trypsin digestion,
treatment with iodoacetamide (IAC) followed by MS/MS analysis. MS/MS analysis

74
of the peptides from the control sample (TrxR only and no PbTx-2) revealed the
presence of numerous peptides of predicted size according to the amino acid sequence
and the anticipated IAC modified fragments (Fig 40). These included the predicted C-
terminal peptide SGGDILQSGCUG, doubly charged and dialkylated with IAC at
both the cysteine and selenocysteine residues. From the PbTx-2 incubated TrxR
sample, the anticipated PbTx-2 modified C-terminal peptide SGGDILQSGCU(PbTx-
2)G was not observed (Figure41). Nonetheless, in the PbTx-2 treated TrxR, the
peptide sequence SGGDILQSGCUG either modified with IAC or unmodified was
conspicuously absent. Because it has been published that when the enzyme is
oxidized, it is resistant to carboxypeptidase or trypsin treatment (Zhong, Arnér et al.
1998). In our case, DTT was used to reduce all disulfide bonds before trypsin
treatment. However, the peptide sequence SGGDILQSGCUG was still un-observable.
It is possible that PbTx-2 alkylated TrxR is resistant to trypsin digestion.
Figure 40 MS/MS analysis of the peptides from rat liver TrxR. Samples were analyzed by SDS PAGE and the bands of anticipated size were excised from the gel and subjected to in-gel trypsin digestion, treatment with iodoacetamide (IAC) followed by MS/MS analysis. The peptides of predicted size according to the amino acid sequence were highlighted by yellow. The anticipated IAC modified fragments were highlighted by green. The predicted C-terminal peptide SGGDILQSGCUG, doubly charged and dialkylated with IAC at both the cysteine and selenocysteine residues (last row)
Figure 41 MS/MS analysis of the peptides from the adduct formation between PbTx-2 and TrxR. Samples were analyzed by SDS PAGE and the bands of anticipated size were excised from the gel and subjected to in-gel trypsin digestion, treatment with iodoacetamide (IAC) followed by MS/MS analysis. The peptides of predicted size according to the amino acid sequence were highlighted by yellow. The anticipated PbTx-2 modified C-terminal peptide SGGDILQSGCU(PbTx-2)G was not observed. Nonetheless, in the PbTx-2 treated TrxR, the

75
peptide sequence SGGDILQSGCUG either modified with IAC or unmodified was conspicuously absent (last row) 3.3.7 Reaction of TrxR with Sel-green probe
The selenol selective probe, Sel-green (11) was prepared as previously reported
(Zhang, Ge et al. 2015).The Sel-green probe undergoes a nucleophilic aromatic
substitution (SNAr) reaction selectively with selenols to release a fluorescent
coumarin reporter (Figure 42). After incubation of reduced TrxR with an 8 fold
excess of PbTx-2, Sel-green was added and the fluorescence increase was monitored
(λex/em = 370/502 nm). The addition of the Sel-green probe, to reduced mammalian
TrxR resulted in release of the fluorescent reporter. If Sec is alkylated by PbTx-2,
then the fluorescence reporter should not be released in the presence of PbTx-2 and
fluorescence should not be observed. We anticipated a 1:1 stoichiometry the reaction
between TrxR and the Sel-green probe and while the concentration of TrxR (5.3 µM
based on FAD absorbance) is below the limit for accurate quantitation for the
reporter, the fluorescence in the absence of PbTx-2 is comparable to the fluorescence
released in the presence of selenocysteine (6 µM) as a control. Fluorescence is clearly
reduced in the presence of PbTx-2 to a level that is only slightly higher than the
control sample which doesn’t contains PbTx-2 (Figure 43).
Figure 42 Reaction of Sel-Green probe with selenocysteine
NO2O2N
SHN
O OO
O
N
ON
NH2
O NO2
O2N
SeCO2H
H2NSec
SO2
Fluorescent ReporterSel-green 11

76
Figure 43 Release of fluorescent reporter from Sel-green probe upon incubation with TrxR in the presence and absence of PbTx-2 compared to standards of L- selenocysteine (3 µM and 6 µM). Rat TrxR (5.3 µM ) was incubated with sel-green probe (43 µM ) with presence and absence of PbTx-2 (48 µM ). Control experiment: Seleno-L-cysteine (3 µM and 6 µM ) was incubated with sel-green probe ( 43 µM ) 3.3.8 Grx, GPx fluorescent assay
The activities of other reductases which are similar to Trx or TrxR was determined in
the presence of PbTx-2. As mentioned above, Grx and GPx are both related to the
GSH reduction system to protect cells from oxidative damage by thiol-disulfide
exchange. Thus, it is anticipated that PbTx-2 may affect their activity as well. The
activity of Grx and GPx in the presence and absence of PbTx-2 was examined using
commercially available assay kits.
In Grx assay, the Grx activity was measured by a coupled reaction of GSH /GSSH
system. NADPH was added as electron donor to reduce GSSG to GSH via GR. The
oxidized Grx was reduced by GSH. In this kit, an eosin- modified GSH was used as
its substrate (Figure 44). An increase in fluorescence was recorded at 545 nm
emission and 520 nm excitation over 120 mins. The rate of reaction was treated as
activity of Grx.
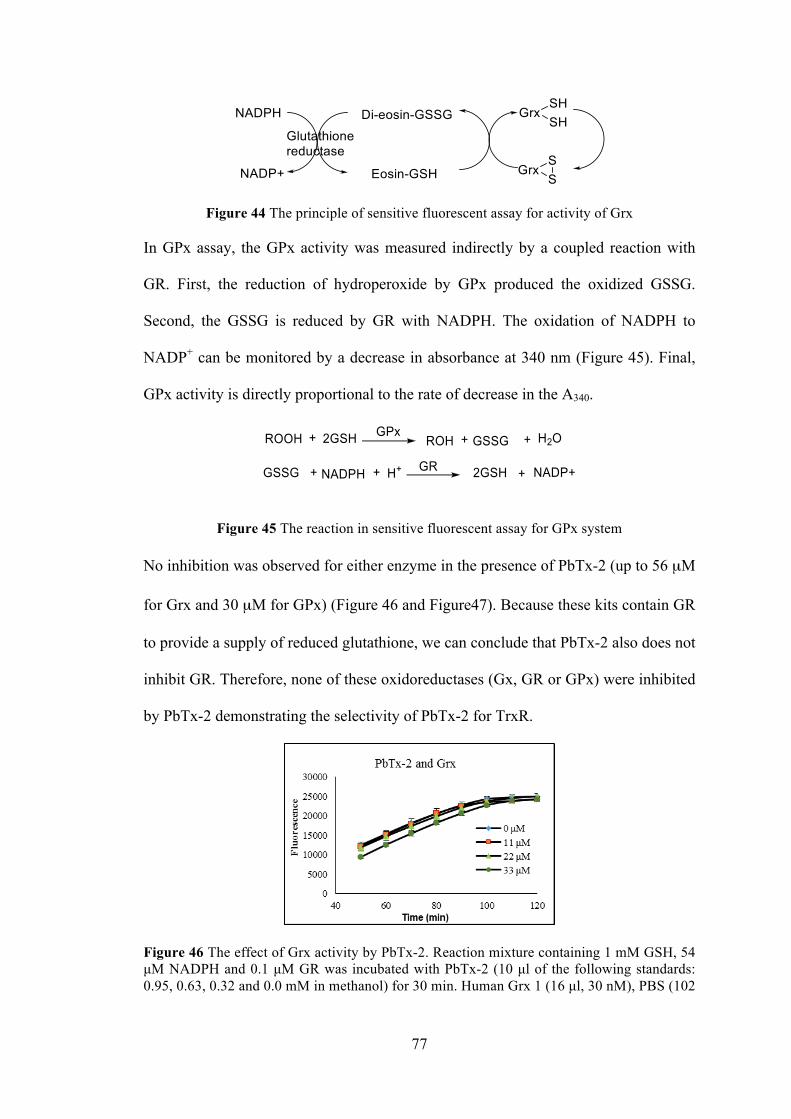
77
Figure 44 The principle of sensitive fluorescent assay for activity of Grx
In GPx assay, the GPx activity was measured indirectly by a coupled reaction with
GR. First, the reduction of hydroperoxide by GPx produced the oxidized GSSG.
Second, the GSSG is reduced by GR with NADPH. The oxidation of NADPH to
NADP+ can be monitored by a decrease in absorbance at 340 nm (Figure 45). Final,
GPx activity is directly proportional to the rate of decrease in the A340.
Figure 45 The reaction in sensitive fluorescent assay for GPx system No inhibition was observed for either enzyme in the presence of PbTx-2 (up to 56 µM
for Grx and 30 µM for GPx) (Figure 46 and Figure47). Because these kits contain GR
to provide a supply of reduced glutathione, we can conclude that PbTx-2 also does not
inhibit GR. Therefore, none of these oxidoreductases (Gx, GR or GPx) were inhibited
by PbTx-2 demonstrating the selectivity of PbTx-2 for TrxR.
Figure 46 The effect of Grx activity by PbTx-2. Reaction mixture containing 1 mM GSH, 54 µM NADPH and 0.1 µM GR was incubated with PbTx-2 (10 µl of the following standards: 0.95, 0.63, 0.32 and 0.0 mM in methanol) for 30 min. Human Grx 1 (16 µl, 30 nM), PBS (102
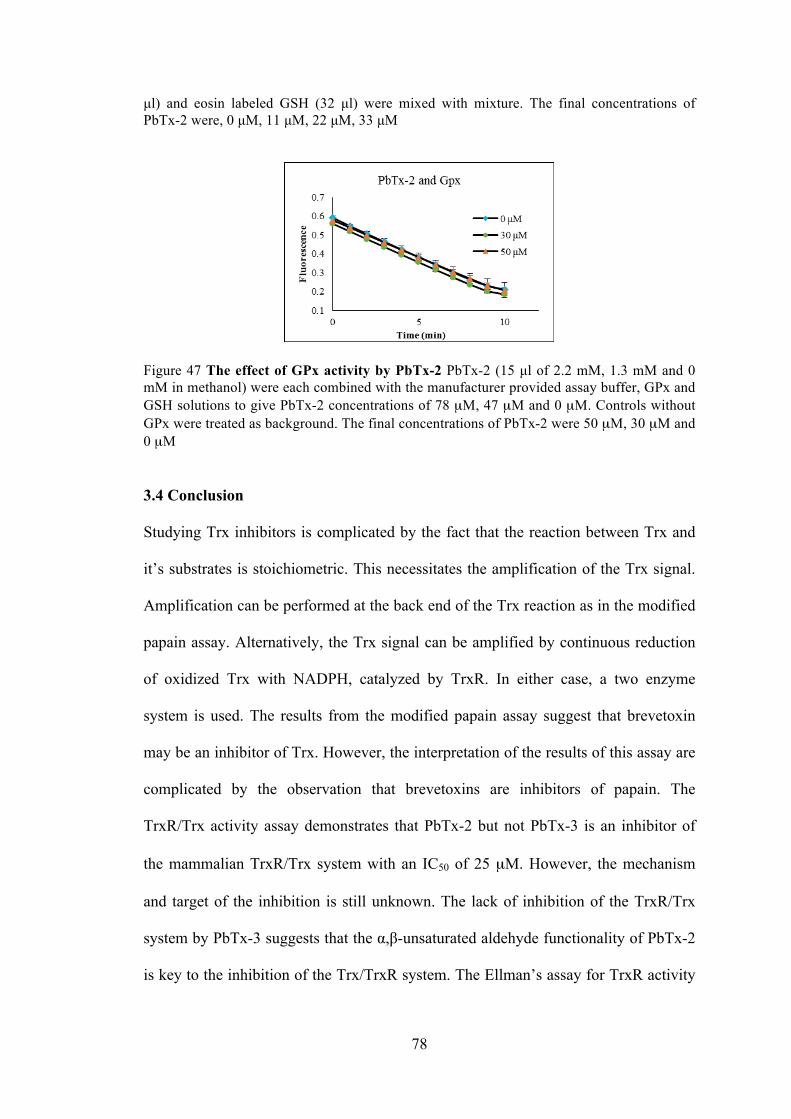
78
µl) and eosin labeled GSH (32 µl) were mixed with mixture. The final concentrations of PbTx-2 were, 0 µM, 11 µM, 22 µM, 33 µM
Figure 47 The effect of GPx activity by PbTx-2 PbTx-2 (15 µl of 2.2 mM, 1.3 mM and 0 mM in methanol) were each combined with the manufacturer provided assay buffer, GPx and GSH solutions to give PbTx-2 concentrations of 78 µM, 47 µM and 0 µM. Controls without GPx were treated as background. The final concentrations of PbTx-2 were 50 µM, 30 µM and 0 µM
3.4 Conclusion
Studying Trx inhibitors is complicated by the fact that the reaction between Trx and
it’s substrates is stoichiometric. This necessitates the amplification of the Trx signal.
Amplification can be performed at the back end of the Trx reaction as in the modified
papain assay. Alternatively, the Trx signal can be amplified by continuous reduction
of oxidized Trx with NADPH, catalyzed by TrxR. In either case, a two enzyme
system is used. The results from the modified papain assay suggest that brevetoxin
may be an inhibitor of Trx. However, the interpretation of the results of this assay are
complicated by the observation that brevetoxins are inhibitors of papain. The
TrxR/Trx activity assay demonstrates that PbTx-2 but not PbTx-3 is an inhibitor of
the mammalian TrxR/Trx system with an IC50 of 25 µM. However, the mechanism
and target of the inhibition is still unknown. The lack of inhibition of the TrxR/Trx
system by PbTx-3 suggests that the α,β-unsaturated aldehyde functionality of PbTx-2
is key to the inhibition of the Trx/TrxR system. The Ellman’s assay for TrxR activity

79
shows that both PbTx-2 and to a lesser extent, PbTx-3 enhance the activity of
mammalian TrxR. The enhancement of DTNB reduction by TrxR observed in the
presence of PbTx-2 was unexpected. According to previous reports, small molecules
such as DTNB can be reduced by the N-terminal reaction center of TrxR when the C-
terminal center is either alkylated with electrophiles or absent as in the case of
truncated TrxR (Eckenroth, Lacey et al. 2007, Cheng, Sandalova et al. 2009, Lothrop,
Ruggles et al. 2009) It can be assumed that the modification of TrxR by PbTx-2
occurs at the C-terminal redox center. The observation that truncated TrxR which
lacks the C-terminal redox center, is unaffected by PbTx-2 also supports the
hypothesis that PbTx-2 affects mammalian TrxR at the C-terminal redox center.
Furthermore, the rate of DTNB reduction was not affected by pre-incubation of
reduced TrxR with PbTx-2 or addition of PbTx-2 and DTNB to pre-reduced TrxR. It
indicates that the alkylation of the C-terminal redox residue(s) of TrxR by PbTx-2
occurs rapidly relative to the reduction of DTNB. Crystal structures of TrxR with and
without Trx have shown that the C-terminal (Cys496/Sec497) residues of TrxR are
located at the end of a mobile arm. Prior to docking with Trx, the mobile arm is in
what is known as the “reduced waiting position” and the C-terminal residues are
buried in the interior of the enzyme. Upon docking with Trx, the mobile arm swings
out of this position to the surface of the enzyme (Fritz-Wolf, Kehr et al. 2011). It is
possible that alkylation with the large brevetoxin molecule also causes the mobile are
to move away from the N-terminal disulfide making it more accessible by DTNB,
bypassing the slow reduction of the C-terminal selenosulfide resulting in a rate
enhancement.

80
Based on the adduct formed between TrxR and selenocysteine, it seems most likely
that modification of TrxR takes place at the Sec residue as selenols are better
nucleophiles than thiols: selenols have a lower pKa than thiols and would be
deprotonated to the selenide under physiological conditions. While our MS
experiments using TrxR and PbTx-2 did not provide direct evidence for Sec
alkylation, the conspicuous absence of the IAC modified C-terminal peptide,
SGGDILQSGCUG, which was identified in the control sample, suggests modification
at this site. Alkylation of this peptide by PbTx-2 would prevent IAC alkylation, and
the absence of this peptide suggests that the PbTx-2 modified peptide may be resistant
to trypsin digestion or may not ionize efficiently. Because the MS analysis of PbTx-2
treated TrxR failed to provide direct evidence of alkylation of Sec, the selenol
selective Sel-green probe was prepared to monitor for the presence of the reduced
selenol of TrxR in the presence and absence of PbTx-2. As anticipated, in the absence
of PbTx-2 the release of an equivalent of fluorescent coumarin reporter was observed
upon the addition of Sel-green to reduced TrxR. However, in the presence of PbTx-2
the observed fluorescence was significant lower which indicates that Sec had been
alkylated with PbTx-2.
3.5 Materials and Methods
3.5.1 Modified papain assay
The materials for the modified papain assay Papain, methyl methanethiosulfonate,
N-benzoyl-L-arginine-p-nitroanilide, immobilized TCEP (Tris [2-carboxyethyl]
phosphine hydrochloride), cystamine and thioredoxin 1 recombinant protein were
purchased from Thermo Scientific Co.. Brevetoxins (PbTx-2 and -3) were isolated
from K. brevis cultures according to published methods (Baden, Mende et al. 1981).

81
UV/Visible measurements were performed in 96 or 384 well microplates using a
Synergy® 2 (Biotek Instrument, Inc.)
Preparation of papain-S-SCH3 (~1.2 mg / ml) Papain stock solution (1.5 ml, 26.4
mg / ml) was added to a solution of cysteine (0.6 mM, 28.5 ml of 20 mM sodium
phosphate – 1 mM EDTA buffer). The solution was kept at room temperature for 30
min. The methyl methanethiosulfonate (117 µmol) was added in mixture. The
reaction mixture was kept in ice for 4 hr. The mixture was dialyzed (Mr cutoff 6000-
8000) against 5 mM sodium acetate buffer, containing 50 mM NaCl, pH 4.7 at 4oC 12
hr.
The method for the modified papain assay The thioredoxin solution (1 mg / ml, 1
mM Tris – HCl pH.7) was incubated with immobilized TCEP for 1 hr. The reduced
thioredoxin solution (90 µl,1 µg / µl) was reacted with 20 µg brevetoxin (1 µg / µl,
PbTx-2 or PbTx-3) for 1 hr in one clean test tube. The solution of papain-S-SCH3
was diluted with an equal volume of 40 mM sodium phosphate buffer. The resulting
solution is ~0.6 mg / ml papain-S-SCH3 at pH 7.5, containing 1 mM EDTA. Papain-
S-SCH3 solution (0.5 ml), substrate (0.7 ml) and cystamine (15 µl, 26.4 mg / ml) were
added in the test tube with mixture. A tube containing papain-S-SCH3, without any
thiol, serves as the control. The mixture is carried out in triplicate in plate. Each well
was added 300 µl mixture. Record the absorbance at 410 nm every 60 min up to 15 hr.
3.5.2 Papain assay
The materials for the papain assay Papain, N-benzoyl-L-arginine-p-nitroanilide
and TCEP were purchased from Thermo Scientific Co. Brevetoxins (PbTx-2 and -3)
were isolated from K. brevis cultures according to published methods (Baden, Mende
et al. 1981). UV/Visible measurements were performed in 96 or 384 well microplates
using a Synergy® 2 (Biotek Instrument, Inc.)

82
The method for the papain assay The solution of papain (304 µl, 26.4 mg / ml ), N-
benzoyl-L-arginine-p-nitroanilide (0.7 ml) were added in the test tube with mixture.
PbTx-2 and PbTx-3 (20 µl, 1 mg / ml) was added to the tube respectively. Using the
Tris buffer filled the finial volume to 625 µl. A tube without any brevetoxin, served as
the control. The mixture is carried out in triplicate in plate. Each well was added 300
µl mixture. Record the absorbance at 410 nm every 60 min up to 15 hr.
3.5.3 Fluorescent assay for Trx/TrxR system
The materials for the fluorescent assay for Trx/TrxR system: Unless otherwise
noted, reagents were purchased from Sigma-Aldrich or Acros chemical companies
and used without further purification. Brevetoxins (PbTx-2 and -3) were isolated from
K. brevis cultures according to published methods (Baden, Mende et al. 1981).
Enzyme assay kit was purchased from Cayman Chemical. Assay kit was used
according to the manufacturer’s instructions with noted exceptions. Fluorescence
measurements were performed in 96 or 384 well microplates using an Infinite®
M1000 PRO (Tecan Group Ltd.) microplate readers.
The methods for the fluorescent assay for Trx/TrxR system. Experiments were
performed in 96-well plates with 270 µl of assay buffer (50 mM Tris-HCl, 1 mM
EDTA, 0.14 mg / ml BSA, pH 7.5) containing 0.13 µM rat TrxR, 0.35 mM NADPH
and 0.026 µM Trx. After incubation for 30 min at 37 0C to reduce the TrxR, stock
solutions of PbTx-2 or PbTx-3 (11 µl each of 1.1, 0.96, 0.64, 0.35 and 0 mM in
methanol) were added to give final toxin concentrations of 35 µM, 30 µM, 20 µM, 11
µM, 0 µM. This mixture was incubated at 37 0C for 1 h. Fluorescent substrate (70 µl,

83
0.4 mg / ml, FiTC labeled bovine insulin) was added to each aliquot (final
concentration of 0.016 mg / ml). Samples were analyzed in triplicate by recording
fluorescence at 545 nm (λex = 520 nm) every 10 min for 3 h.
3.5.4 Ellman’s assay
The materials for the DTNB assay: Unless otherwise noted, reagents were
purchased from Sigma-Aldrich or Acros chemical companies and used without
further purification. Brevetoxins (PbTx-2 and -3) were isolated from K. brevis
cultures according to published methods (Baden, Mende et al. 1981). Truncated
human TrxR (Novus Biologicals) was purchased from Fisher Scientific. Rat liver
TrxR was purchased from Cayman Chemical. DTNB was purchased from Thermo
Scientific Co.. NADPH was purchased from Molecular Toxicology, Inc. UV/Visible
measurements were performed in 96 or 384 well microplates using a Synergy® 2
(Biotek Instrument, Inc.)
The method for the DTNB assay for TrxR or truncated TrxR. Method A (pre-
incubation with PbTx-2): Experiments were performed in 96-well plates with 160 µl
of PBS containing 0.1 mM NADPH. 22 µM PbTx-2 and 0.56 µM rat TrxR or 0.68
µM truncated human TrxR. After incubation for 30 min at 37 0C to reduce the TrxR,
40 µl of a DTNB solution (Ellman’s reagent: 5,5’-dithio-bis-(2-nitrobenzoic acid) (10
mM in 0.1 M sodium phosphate and 1 mM EDTA, pH 8.0) was added for final
concentrations of 18 µM PbTx-2 and 2 µM DTNB. The absorbance at 410 nm was
measured every 5 min for up to 60 min. Method B (no pre-incubation with PbTx-2):
The assay was performed as described above except that PbTx-2 and DTNB solutions
were mixed and added simultaneously after NADPH reduction of TrxR.

84
3.5.5 The preparation of selenocysteine -PbTx-2 adduct
The materials for making selenocysteine-PbTx-2 adduct PbTx-2 isolated from K.
brevis cultures according to published methods (Baden, Mende et al. 1981). Seleno-L-
cysteine was borrowed from Dr. Lees lab. Immobilized TCEP was purchased from
Thermo Scientific Co.. The formation was detected from a Bruker Solarix 7.0 T using
ultrahigh resolution Fourier Transform Ion Cyclotron Resonance Mass Spectrometer
(FT-ICR).
Preparation and analysis of selenocysteine-PbTx-2 adduct seleno-L-cysteine(75
µL, 1 mg /mL) was incubated with an aqueous suspension of immobilized TCEP
((tris(2-carboxyethyl) phosphine, 30 µL, >8 µmol/mL, immobilized onto 4%
crosslinked beaded agarose) for 1 h to reduce the diselenide. The reaction vial was
flushed with nitrogen and centrifuged for 10 min at 14,000 x g. The supernatant was
transferred via syringe to a solution of PbTx-2 (200 µL, 1 µg/µL in methanol) and
stirred under nitrogen for 24 h. Accurate mass spectra was acquired using a Bruker
Solarix 7.0 T using ultrahigh resolution Fourier Transform Ion Cyclotron Resonance
Mass Spectrometer (FT-ICR) confirmed the formation of an oxidized (+15.999 amu)
selenocysteine-PbTx-2 adduct of formula C53H77NO17Se. HRMS calc’d for [M+H+]:
1080.44355. Found: 1080.44356 (Δ = -0.56 ppm). HRMS calc’d for [M+Na+]:
1102.42545. Found: 1102.42550 (Δ = -0.12 ppm).
3.5.6 The preparation of TrxR-PbTx-2 adduct.
The materials for making TrxR-PbTx-2 adduct Rat liver TrxR was purchased from
Cayman Chemical. PbTx-2 isolated from K. brevis cultures according to published

85
methods (Baden, Mende et al. 1981). NADPH was purchased from Molecular
Toxicology, Inc..
Preparation and analysis of TrxR-PbTx-2 adduct. Rat TrxR (6 µM) and NADPH
(0.16 mM) were incubated for 30 min in 44 mM Tris base, 0.9 mM EDTA. 0.4 M
NaCl, 0.1 % glycerol, pH 7. PbTx-2 (5.6 mM in Me2SO) was added to give a final
PbTx-2 concentration of 0.24 mM and the reaction was incubated for 12 h at room
temperature. The sample was concentrated 5 fold and analyzed by SDS PAGE (12%
acrylamide, 10% SDS). The band was excised and stored at -80 °C. Gel digestion and
LC-Electrospray Ionization MS/MS (LC-ESI MS/MS) analysis was performed by the
Proteomics and Mass Spectrometry Facility at the UMass Medical School as
previously described.
3.5.7 Reaction of Sel-green probe with TrxR
The materials for reaction of sel-green with TrxR Sel-Green was synthesized by
Freddy Rodriguez. Rat liver TrxR was purchased from Cayman Chemical. PbTx-2
isolated from K. brevis cultures according to published methods (Baden, Mende et al.
1981). NADPH was purchased from Molecular Toxicology, Inc. Seleno-L-cysteine
was borrowed from Dr. Lees lab. Fluorescence measurements were performed in 96
or 384 well microplates using a an Infinite® M1000 PRO (Tecan Group Ltd.)
microplate readers.
Reaction of Sel-green probe with TrxR. The selenol selective probe, Sel-green, was
synthesized according to published methods (Zhang, Ge et al. 2015). Rat TrxR (6

86
µM) and NADPH (0.1 mM) were incubated for 30 min in 45 mM Tris base, 0.9 mM
EDTA. 0.5 M NaCl, 0.1 % glycerol, pH 7. PbTx-2 (1.1 mM in methanol) was added
for a final toxin concentration of 48 µM. Samples were prepared in triplicate. A
control sample was prepared by the addition of an equal volume of methanol without
PbTx-2. A blank sample was prepared without TrxR. The solutions were incubated
for 1 h. Sel-green probe (9 µl, 481 µM in 20% acetone in PBS) was added to each
reaction mixture (82 µl). After 55 min, fluorescence at 502 (λex = 370 nm) was
measured. Calibration standards were prepared as follows: A stock solution of seleno-
L-cysteine (1.0 ml, 27 µM, in PBS) was reduced using immobilized TCEP (1.0 ml, >
8 µmol/ml) for 1 h. After centrifugation (14000 x g, 2 min), the supernatant was
transferred to a clean tube to provide a stock solution of L-selenocysteine (54 µM).
Aliquots of the stock solution of L-selenocysteine was diluted with Sel-green probe in
PBS for final concentrations of L-selenocysteine of 0 µM, 3 µM, 6 µM, 12 µM, 25
µM and 43 µM Sel-green probe, incubated for 55 min, and fluorescence was
measured at 502 nm (λex = 370 nm). Calibration standards were prepared in
triplicate. For this experiment, the concentration of FAD (flavin adenine dinucleotide)
was calculated to be 5.3 µM by measuring the absorbance of at 450 nm and using an
extinction coefficient ε450 = 11,300 M–1cm–1 for FAD (Zhong and Holmgren 2000).
3.5.8 Fluorescent assay for Grx and GPx
The materials for the activity assay for Grx Fluorescent Grx kit was purchase from
Cayman Chemical. PbTx-2 isolated from K. brevis cultures according to published
methods (Baden, Mende et al. 1981). Fluorescence measurements were performed in

87
96 or 384 well microplates using an Infinite® M1000 PRO (Tecan Group Ltd.)
microplate readers.
The method for the activity assay for Grx. Experiments were performed in 96-well
plates with 160 µl of a reaction mixture containing 0.17 M potassium phosphate, 1.7
mM EDTA, 1 mM GSH, 0.1 mg / ml BSA, 54 µM NADPH, 0.1 µM GR, pH 7.5
which was prepared according to the manufacturer’s instructions. To aliquots (160 µl)
of the reaction mixture was added PbTx-2 (10 µl of the following standards: 0.95,
0.63, 0.32 and 0.0 mM in methanol), 16 µl of human Grx 1 (30 nM in 0.83 M
potassium phosphate and 8.3 mM EDTA, pH 7.5, final concentration 1.5 nM), 102 µl
of PBS and 32 µl of eosin labeled GSH bound to BSA (0.2 mM in distilled water).
Samples were analyzed in triplicate by recording fluorescence at 545 nm (λex = 520
nm) every 10 min for up to 2 h. The final concentrations of PbTx-2 were, 0 µM, 11
µM, 22 µM, 33 µM.
The materials for the activity assay for GPx Fluorescent GPx kit was purchase
from Cayman Chemical. PbTx-2 isolated from K. brevis cultures according to
published methods (Baden, Mende et al. 1981)Fluorescence measurements were
performed in 96 or 384 well microplates using an Infinite® M1000 PRO (Tecan
Group Ltd.) microplate readers
The method for the activity assay for GPx The assay was carried out according to
the manufacturer’s instructions except that stock solutions of PbTx-2 (15 µl of 2.2
mM, 1.3 mM and 0 mM in methanol) were each combined with the manufacturer
provided assay buffer, GPx and GSH solutions to give PbTx-2 concentrations of 78
µM, 47 µM and 0 µM. This mixture was incubated for 1 h at room temperature, after

88
which the assay was completed according to the manufacturer’s instructions. The
absorbance was read once every minute at 340 nm for 10 min. Samples were analyzed
in triplicate. Controls without GPx were treated as background. The final
concentrations of PbTx-2 were 50 µM, 30 µM and 0 µM.

89
Chapter 4 Comparison of biochemical parameters in high toxin and low toxin
producing strains of K. brevis.
4.1 Objective
Compare and contrast various metabolic and biochemical parameters in two strains of
K. brevis which have a tenfold difference in toxin content
4.2 Introduction
In chapter 2 and 3, by using fluorescent and photoaffinity derivatives of brevetoxin it
was concluded that brevetoxin localizes to the chloroplast of K. brevis where it
interacts with two cholorplast proteins: LHCII and Trx (Cassell, Chen et al. 2015). It
was also shown that brevetoxin inhibits the Trx/TrxR system within an IC50 of 25 µM.
In addition to inhibition of the Trx/TrxR system, brevetoxin affects mammalian TrxR
leading to the enhancement of the activity of this enzyme. The evidence indicates that
brevetoxin affects mammalian TrxR at the C-terminal selenocysteine active site
residue. These discoveries lead us back to our original question: what is the
biochemical role of brevetoxin in K. brevis? Or more specifically: what are the
implications of TrxR inhibition by brevetoxin for K. brevis? In fact, LHC II, Trx and
TrxR are all common antioxidants related to reactive oxygen species (ROS) in cells.
When the excess ROS accumulate in cells, the state is called oxidative stress. Various
biomolecules such as proteins (Stadtman 1992, Nordberg and Arnér 2001), lipids,
lipoprotein S (Yla‐Herttuala 1999), and DNA (Marnett 2000, Nordberg and Arnér
2001), are threatened by the reactive oxygen intermediates. Oxidative stress is
proposed to be involved in DNA damage in the mitochondria (Cadenas and Davies
2000, Finkel and Holbrook 2000). However, a small concentration of ROS is required
for normal cell function as well. It has been demonstrated that ROS are involved in

90
intracellular signaling and redox regulation. The term ROS includes a number of
chemically reactive molecules which are derived from oxygen (Galter, Mihm et al.
1994, Halliwell 1996, Fridovich 1999, Betteridge 2000) (Table 9). Some are
extremely reactive like hydroxyl radical, in contrast some are less reactive, such as
superoxide anion (O2-) and hydrogen peroxide (Nordberg and Arnér 2001) Table 9
also shows the main sources of production and the enzymatic defense system for
various ROS.
Table 9 The major ROS molecules and their metabolism
ROS molecule Enzymatic defense systems Products Superoxide (O2
-) Superoxide dismutase (SOD), Superoxide reductase
H2O2+O2 H2O2
Hydrogen peroxide (H2O2)
Glutathione peroxidase, Catalases,
Peroxiredoxins (Prx)
H2O+GSSG H2O+O2
H2O
Hydroxyl radical (.OH)
Nitric oxide (NO) Glutathione/TrxR GSNO
Superoxide anion (O2-) is formed from molecular oxygen. Although it is a free
radical, it is not highly reactive. Because it cannot penetrate lipid membranes, which
means superoxide anion can only remain where it is produced (Nordberg and Arnér
2001). Usually, superoxide anion is formed in the electron-rich aerobic environment
such as the inner mitochondrial membrane with the respiratory chain (Derick,
Williams et al. 2001). Lipoxygenase and cyclooxygenase are also generated
superoxide (Kontos, Wei et al. 1985, Mattei, Virdis et al. 1996). In addition to
superoxide, hydrogen peroxide has similar intracellular effects (Sundaresan, Yu et al.
1995, Finkel 1998, Kamata and Hirata 1999, Rhee 1999, Patel, Moellering et al.

91
2000). Hydrogen peroxide is a highly important ROS molecule because H2O2 is able
to penetrate biological membranes. Many antioxidant enzyme systems, such as
catalase, glutathione peroxidase and peroxiredoxins, are able to remove H2O2 (Chae,
Kang et al. 1999, Chae, Kim et al. 1999, MatÉs, Pérez-Gómez et al. 1999). Hydroxyl
radical (.OH) is the most reactive ROS molecule, causing more damage to biological
systems than other ROS. Usually .OH is obtained from H2O2 (Neyens and Baeyens
2003). Last but not least is nitric oxide (NO), a well-known as a signaling molecule.
Additionally, NO has been identified as an inhibitor of lipid peroxidation in cell
membranes (Hogg and Kalyanaraman 1998, Rubbo, Radi et al. 2000). NO is able to
regulate gene transcription and expression as well (Bogdan 2001).
Due to the high reactivity of ROS, it causes damage to numerous biomolecules. For
example, ROS is treated as a mutagen. ROS causes a number of alterations such as
DNA cleavage, DNA protein cross-linking and oxidation of purines when the cells are
exposed to oxidative stress. It is determined that ROS are related to DNA mutation,
cancer and apoptosis (Halliwell, Zentella et al. 1997, Kamata and Hirata 1999). Lipids
are also an excellent target for ROS because of the multiple double bonds in
polyunsaturated fatty acids (Steinberg 1997, Kamata and Hirata 1999). Lipid
peroxidation is essential for the generation of atherosclerotic plaques (Halliwell 1993,
Frei 1999). The incidences of cardiovascular disease are increasing in western
countries. The most important prevention and medical treatments are to decrease lipid
peroxidation, which is associated with excessive amounts of ROS. Furthermore, ROS
has been shown to react with several amino acids in vitro, resulting in modification or
inhibition of the activity of enzymes (Stadtman and Berlett 1997, Butterfield, Koppal
et al. 1998).

92
ROS are affected by antioxidants such as the Trx and GSH systems (Lowther, Brot et
al. 2000, Nordberg and Arnér 2001). LHCII also affects ROS production. In fact,
plants and algae love light but also hate light. Oxygenic photoautotrophic organisms
require light for life. However, when the input of excess high light exceeds the
saturation limit of the photosynthetic apparatus this leads to an increase the
production of ROS. Chloroplasts have evolved a number of mechanisms to mitigate
the damaging effects of ROS, which include both the scavenging of ROS and the
suppression of ROS production through NPQ (Figure 48 A). When a Chl absorbs
light energy, it is excited from ground state to singlet molecule (1Chl*). The singlet
1Chl* can be re-emitted as Chl fluorescence that is used as photochemistry. Also, it
can be de-excited by NPQ. However, singlet 1Chl* is able to turn to triplet state (3Chl*)
which can transfer energy to ground state O2 to form singlet oxygen (1O2*). Singlet
oxygen (1O2*) is an extremely damaging ROS. In normal light, the energy is
transferred to photochemistry. The yield of Chl fluorescence is generally low (Krause
and Weis 1991). However, under excess light, the increase of 1Chl* leads to yield high
level of 3Chl* which generates more 1O2*. 1O2
* causes oxcide damage to the cells
(Figure48 B). However, NPQ can either dissipate the 1Chl* as heat or quench Chl
fluorescence to prevent cells from oxygen damage (Müller, Li et al. 2001). Thus,
NPQ is one of mechanisms to suppress to ROS production. If the cells cannot perform
NPQ to dissipate light energy, more ROS will be produced. As mentioned above,
LHCII is an integral part of the photosynthetic apparatus where it harvests and
distributes light energy using four LHCII bound carotenoid molecules. It is also
central to the process of NPQ or the dissipation of light energy as heat under
conditions of high light.

93
Figure 48 The common pathways to produce ROS. First, high light energy is transfered to photochemistry. (A) The common pathway to distribute high light energy in leaves. Once the photochemistry reaches saturation, the excess energy can either be dissipated to heat by NPQ or switch to chlorophyII fluorescence. (B) The possible way to release excited Chl. When Chl absorbs light it is excited from the ground state to the singlet excited state. The singlet excited form, 1Chl* can go back to the ground state by three ways: emitting light as fluorescence, turning to heat and being used in photosynthetic reactions (Müller, Li et al. 2001) In addition to LHC II, in photosynthetic organisms, the Trx/TrxR system is located
within the chloroplast lumen and numerous thioredoxin-dependent regulatory
networks have been identified within the chloroplasts of photosynthetic organisms
(Nikkanen and Rintamäki 2014), including VDE activity (Hall, Mata‐Cabana et al.
2010). In plants, there are highly versatile Trx systems which include two different
TrxRs: ferredoxin thioredoxin reductase (FTR) and NADPH dependent thioredoxin
reductase (NTRC). These TrxRs are localized to plant chloroplast. They contribute to
the regulation of proteins in chloroplast by Trx system. Additionally, it is believed
that the two reductases are selective for different Trx targets. FTR is active only in the
light and is believed to be responsible for regulation of light reactions. While NTRC
enzyme is unique for oxygenic photosynthetic organisms (Nikkanen and Rintamäki
2014).
In mammal, Trxs are also critical to the scavenging of excess ROS by reduction of 2-
cysteine peroxiredoxins (2-Cys Prxs), enzymes whose sole function is the

94
detoxification of peroxides (König, Baier et al. 2002) It is reported that the expression
of Trx and TrxR are induced by oxidative stress in adult T cells (Nakamura, Matsuda
et al. 1994, Taniguchi, Taniguchi-Ueda et al. 1996). When the cells are exposed to
oxidative stress, Trx/TrxR system is the sophisticated antioxidant regulatory system to
maintain the balance of ROS. In Figure49, the major physiological functions of the
mammalian thioredoxin system are shown. In particular, mammalian TrxR, which
reduces more substrates than mammalian Trx, reduces not only the disulfide in
oxidized Trx, but also different protein disulfides, PDI, low molecular weight
disulfides such as DTNB, and nondisulfide compounds, such as hydrogen peroxides,
and nitroblue tetrazolium (NBT)(Arnér, Zhong et al. 1999, Nordberg and Arnér
2001). Mammalian TrxR reduces some selenium-containing enzymes and compounds
either directly or by way of Trx (Kumar, Bjornstedt et al. 1992). For examples,
selenite and selenocysteineand sec-containing GPx are reduced by mammalian TrxR
as well. The active site of GPx is regenerated directly by TrxR (Nordberg and Arnér
2001). It has been determined that sulfur and selenium compounds are able to reduce
the ROS generation through their antioxidant properties. For example, patients with
pulmonary tuberculosis show reduced oxidative stress after taking selenium
supplementation (Seyedrezazadeh, Ostadrahimi et al. 2008). Commonly, sulfur and
selenium compounds protect against disease through ROS radical scavenging and
enzymatic decomposition of oxygen metabolites (Mugesh and Singh 2000, Valko,
Rhodes et al. 2006).

95
Figure 49 Major physiological function of the mammalian thioredoxin system
Furthermore, there are other cellular antioxidant enzyme systems protecting cells and
organisms from excessive ROS. Superoxide dismutase (SOD) catalyzes the reaction
of O2*- (Yost and Fridovich 1973) to produce molecular oxygen and hydrogen
peroxide. Catalases are mainly heme-containg enzymes. Catalases catalyze the
conversion of hydrogen peroxide to water and molecular oxygen (Aebi 1984). All
GPx catalyze the reduction of H2O2 through the assistance of the GSH system. The
catalytic mechanism was proposed in Chapter 3. Last but not least, the GSH system
is the most abundant intracellular thiol-based antioxidant. GSH can be present in
millimolar concentration in living cells (Armstrong 1997, Van Bladeren 2000). GSH
serves as a sulfhydryl buffer to protect against oxidative damage in cells. The
oxidized GSSG can also be reduced by Trx system(Kanzok, Schirmer et al. 2000).
Both LHCII and Trx regulate the oxidative state of the cell and redox signaling. Both
affect the amount of ROS production. In addition to these brevetoxin targeted
proteins, the GSH system and many antioxidants are involved in preventing oxidative
stress in cells. The finding that brevetoxin binds to LHCII and Trx raises an important
question: Does brevetoxin influence the ROS level and the antioxidant content in K.
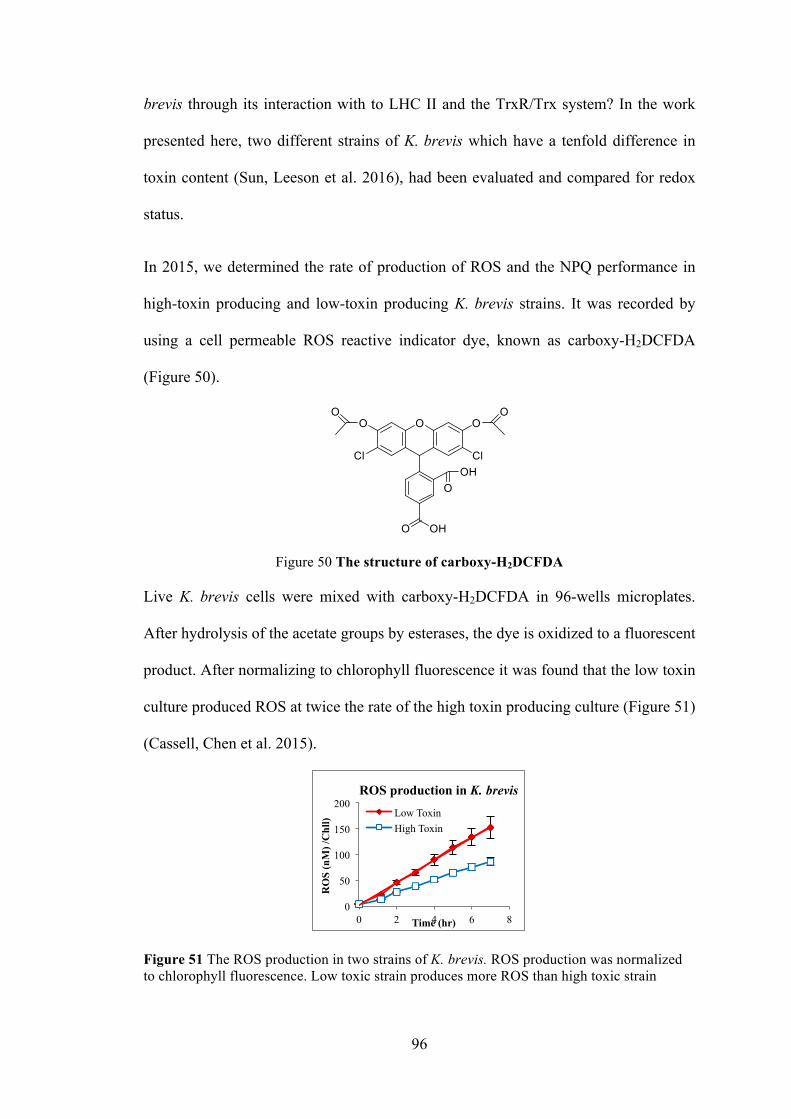
96
brevis through its interaction with to LHC II and the TrxR/Trx system? In the work
presented here, two different strains of K. brevis which have a tenfold difference in
toxin content (Sun, Leeson et al. 2016), had been evaluated and compared for redox
status.
In 2015, we determined the rate of production of ROS and the NPQ performance in
high-toxin producing and low-toxin producing K. brevis strains. It was recorded by
using a cell permeable ROS reactive indicator dye, known as carboxy-H2DCFDA
(Figure 50).
Figure 50 The structure of carboxy-H2DCFDA
Live K. brevis cells were mixed with carboxy-H2DCFDA in 96-wells microplates.
After hydrolysis of the acetate groups by esterases, the dye is oxidized to a fluorescent
product. After normalizing to chlorophyll fluorescence it was found that the low toxin
culture produced ROS at twice the rate of the high toxin producing culture (Figure 51)
(Cassell, Chen et al. 2015).
Figure 51 The ROS production in two strains of K. brevis. ROS production was normalized to chlorophyll fluorescence. Low toxic strain produces more ROS than high toxic strain
0
50
100
150
200
0 2 4 6 8
RO
S (n
M) /
Chl
l)
Time (hr)
ROS production in K. brevis Low Toxin High Toxin
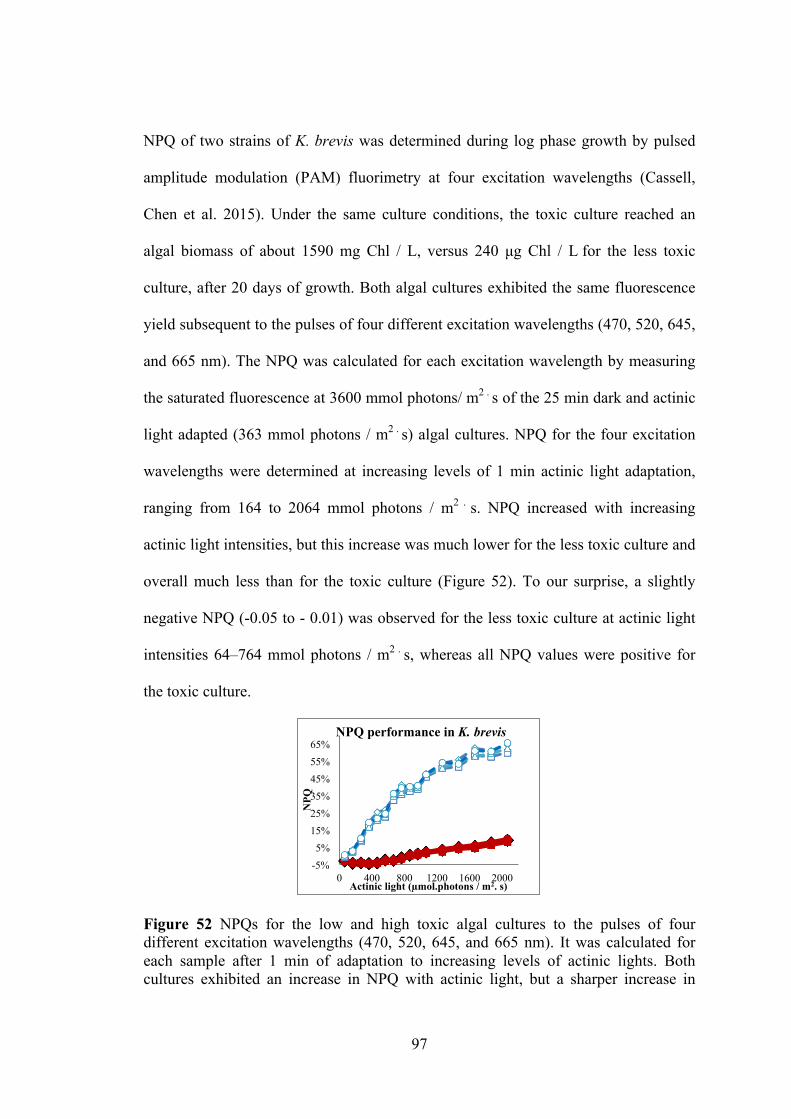
97
NPQ of two strains of K. brevis was determined during log phase growth by pulsed
amplitude modulation (PAM) fluorimetry at four excitation wavelengths (Cassell,
Chen et al. 2015). Under the same culture conditions, the toxic culture reached an
algal biomass of about 1590 mg Chl / L, versus 240 µg Chl / L for the less toxic
culture, after 20 days of growth. Both algal cultures exhibited the same fluorescence
yield subsequent to the pulses of four different excitation wavelengths (470, 520, 645,
and 665 nm). The NPQ was calculated for each excitation wavelength by measuring
the saturated fluorescence at 3600 mmol photons/ m2 . s of the 25 min dark and actinic
light adapted (363 mmol photons / m2 . s) algal cultures. NPQ for the four excitation
wavelengths were determined at increasing levels of 1 min actinic light adaptation,
ranging from 164 to 2064 mmol photons / m2 . s. NPQ increased with increasing
actinic light intensities, but this increase was much lower for the less toxic culture and
overall much less than for the toxic culture (Figure 52). To our surprise, a slightly
negative NPQ (-0.05 to - 0.01) was observed for the less toxic culture at actinic light
intensities 64–764 mmol photons / m2 . s, whereas all NPQ values were positive for
the toxic culture.
Figure 52 NPQs for the low and high toxic algal cultures to the pulses of four different excitation wavelengths (470, 520, 645, and 665 nm). It was calculated for each sample after 1 min of adaptation to increasing levels of actinic lights. Both cultures exhibited an increase in NPQ with actinic light, but a sharper increase in
-5% 5%
15% 25% 35% 45% 55% 65%
0 400 800 1200 1600 2000
NPQ
Actinic light (µmol.photons / m2. s)
NPQ performance in K. brevis

98
magnitude of the NPQs were found for the toxic culture. (Light toxic strain in 470 nm, 520 nm, 645 nm, 665 nm, high toxic strain in 470 nm, 520 nm, 645 nm, 665 nm) The NPQ and ROS experiments revealed that the high toxic strain can perform NPQ
to prevent cells from damage by ROS. In fact, the low-toxic strain produces ROS at
about twice the rate of the high toxic strain, and does not perform NPQ as efficiently
as the high toxic strain. It is unclear if the lack of toxin causes the NPQ deficiency or,
if the NPQ deficiency results in lower toxin production.
4.3 Results and discussion
Initially we had assumed that K.brevis TrxR would be more like that of a plant. Plant
TrxR is more similar to prokaryotic TrxR which does not have the Cys-Sec. However,
it is reported a “mammalian like” TrxR was found in Emeliania huxleii which has the
C-terminal selenosulfide by researching the gene sequence (Araie, Suzuki et al. 2008).
Thus, it is very likely that K. brevis does has “mammalian like TrxR. Using the
sequence from Emeliania huxleii as a query in a BLAST search of the K. brevis
transcriptome library it is revealed that two “mammalian like” TrxR sequence in
which the C-terminal selenosulfide is present (Darcie, Alan et al. 2014).
UGA codon is normally a termination codon, whereas UGA codon is able to code Sec
in a special way. A selenocysteine insertion sequence (SECIS) presents in the mRNA,
which is responsible to make UGA codon to encode Sec (U). SECIS is defined as a
nucleotide sequence which typically located following the UGA codon. In Archaea
and eukaryotes, the SECIS element is located in the 3’ untranslated region of the
mRNA. The SECIS element was found in both DNA sequence of TrxR in K. brevis.
In addition, a selenocysteine residue was discovered in one amino acids sequence of

99
TrxR in K. brevis, which was aligned with human TrxR and “mammalian like ” TrxR
sequence in Emeliania huxleii (Table 10)
Table 10 The alignments of TrxR in K. brevis, Homo sapiens and Emiliania huxleyi
4.3.1 The antioxidants in two strains of K. brevis
Antioxidants play an important role in protecting cells from oxidative stress. GSH,
ascorbic acid, Trx, carotenoids and vitamin E are central to the antioxidant defense in
most of species. Thus, it is suspected antioxidant capacity may not be same between
high toxic and low toxic strain. Examining the antioxidant capacity and ascorbic acid
content may contribute to understanding the role of brevetoxin in K. brevis.
Here, the 2,2'-azino-bis (3-ethylbenzothiazoline-6-sulphonic acid (ABTS) assay was
used to detect all antioxidants such as lipophilic carotenoids, GSH, Trx and ascorbic

100
acid (Figure53). The ABTS assay measures the decolorization (reduction) of the
ABTS radical cation which has a long wavelengthabsorption (Re, Pellegrini et al.
1999).
Figure 53 The reaction in ABTS assay
Total antioxidant capacity of K. brevis homogenates was determined using ABTS
radical cation (ABTS.+) decolorization (Re, Pellegrini et al. 1999) and expressed as
trolox equivalents. Trolox is similar to vitamin E which is used in biological
applications to reduce oxidative damage. Trolox equivalent antioxidant capacity
(TEAC) is commonly used as a standard in the ABTS assay for measuring the
concentration of antioxidant in unknown sample. Here, the homogenate of two K.
brevis strains or trolox standard was used to decolorizw the ABTS!+ in the assay. The
concentration of total antioxidant was estimated based on the trolox concentration
standard curve. Furthermore, ascorbic acid concentration (as trolox equivalents) in K.
brevis homogenates was estimated by incubating with ascorbic acid oxidase (AAO).
AAO catalyzes the oxidation of ascorbic acid to dehydroascorbic acid. Thus, the
difference of absorbance in the presence and absence of AAO provides an estimate of
ascorbic acid concentration in the homogenate. Total antioxidant or ascorbic acid
content were normalized to protein concentration. It was shown that the high toxic

101
strain has a 1.2-fold antioxidant capacity than the low toxic strain on three of the four
days tested (Figure 54). In contrast, the low toxic strain consistently had higher levels
of ascorbic acid (Figure55). It showed that the low toxic strain has maximum 4.5-fold
of ascorbic acid at 19 growing days, when it compared to high toxic strain.
Figure 54 The antioxidant capacity in two strains of K. brevis. ABTS�+ was produced by oxidation of aqueous ABTS (1.3 mL, 7 mM) with ammonium persulfate (21.2 µl, 2.45 mM). The Either 50 µl K. brevis homogenate or trolox standards (0 µM, 25 µM, 50 µM, 100 µM, 200 µM) were added to the ABTS.+ solution (100 µl). The absorbance was read at 734 nM after incubation for 5 min
Figure 55 The total amount of ascorbic acid in two strains of K. brevis. Ascorbic acid concentration (as trolox equivalents) of K. brevis homogenate was estimated by incubating with of ascorbic acid oxidase (2 µl, 0.01 U/ µL) for 30 min prior to the assay and comparing absorbance to parallel samples without AAO
500.00
1000.00
1500.00
2000.00
12 Days 19 Days 20 Days 21 Days Trol
ox e
quiv
alen
t ant
ioxi
dant
ca
paci
ty (n
mol
/mg)
Antioxidant capacity in high toxic and low toxic strain
High toxic strain Low toxic strain
0.00
200.00
400.00
600.00
800.00
12 Days 19 Days 20 Days 21 Days Trol
ox e
quiv
alen
t am
ount
of
asco
rbic
aci
d (µ
mol
/ m
g)
Total amount of ascorbic acid in high toxic and low toxic strain
High toxic strain Low toxic strain

102
4.3.2 Redox state of protein and non-protein thiols in low vs high toxin cultures
of K. brevis
Because brevetoxin is an inhibitor of the Trx/ TrxR system, it follows that the
presence of brevetoxin may impact the amount of free thiols in K. brevis. Therefore,
we examined the thiol content in high and low toxin producing strains of K. brevis.
Modified papain assay As mentioned in Chapter 3, this assay is more sensitive to
thiols than the Ellman’s assay as the signal is amplified by the enzymatic activity of
papain. Thus, the thiol content, normalized to protein concentration, was determined
first by using the modified papain assay in the high toxin and low toxin K. brevis
strains. Because the free thiol groups may be buried in proteins and not readily
accessible by modified papain, cystamine was added into the reaction to access not
only low MW thiols, but also free thiols in proteins. This experiment revealed a
higher thiol content is in the low toxic homogenate compared to toxic strain either
under light or dark conditions (Figure56). No matter under dark adapter or sun light, a
two-fold reduction of V0 was shown in the low toxic strain compared to high toxic
strains for the modified papain assay. Again, this assay is complicated by the fact that
brevetoxin is an inhibitor of papain. Thus, Ellman’s assay has to be used to detect the
thiol content in K. brevis homogenate.
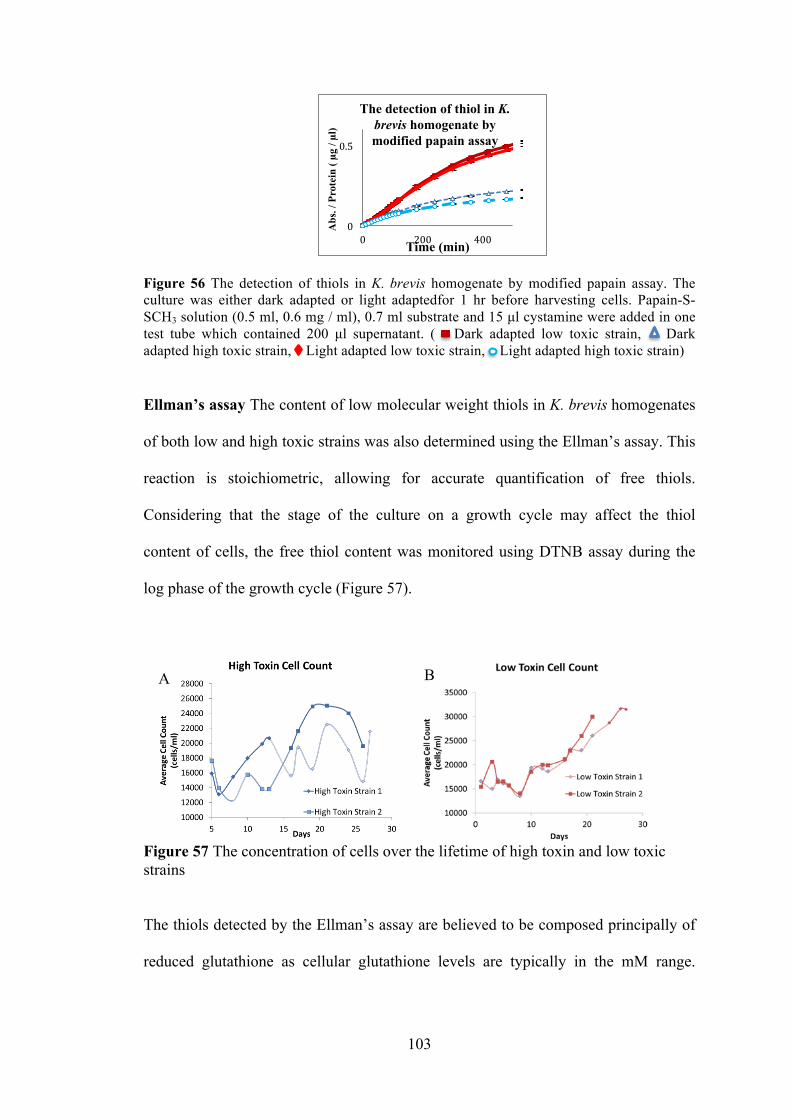
103
Figure 56 The detection of thiols in K. brevis homogenate by modified papain assay. The culture was either dark adapted or light adaptedfor 1 hr before harvesting cells. Papain-S-SCH3 solution (0.5 ml, 0.6 mg / ml), 0.7 ml substrate and 15 µl cystamine were added in one test tube which contained 200 µl supernatant. ( Dark adapted low toxic strain, Dark adapted high toxic strain, Light adapted low toxic strain, Light adapted high toxic strain)
Ellman’s assay The content of low molecular weight thiols in K. brevis homogenates
of both low and high toxic strains was also determined using the Ellman’s assay. This
reaction is stoichiometric, allowing for accurate quantification of free thiols.
Considering that the stage of the culture on a growth cycle may affect the thiol
content of cells, the free thiol content was monitored using DTNB assay during the
log phase of the growth cycle (Figure 57).
Figure 57 The concentration of cells over the lifetime of high toxin and low toxic strains
The thiols detected by the Ellman’s assay are believed to be composed principally of
reduced glutathione as cellular glutathione levels are typically in the mM range.
0
0.5
0 200 400
Abs
. / P
rote
in ( µg
/ µl
)
Time (min)
The detection of thiol in K. brevis homogenate by modified papain assay
A
B
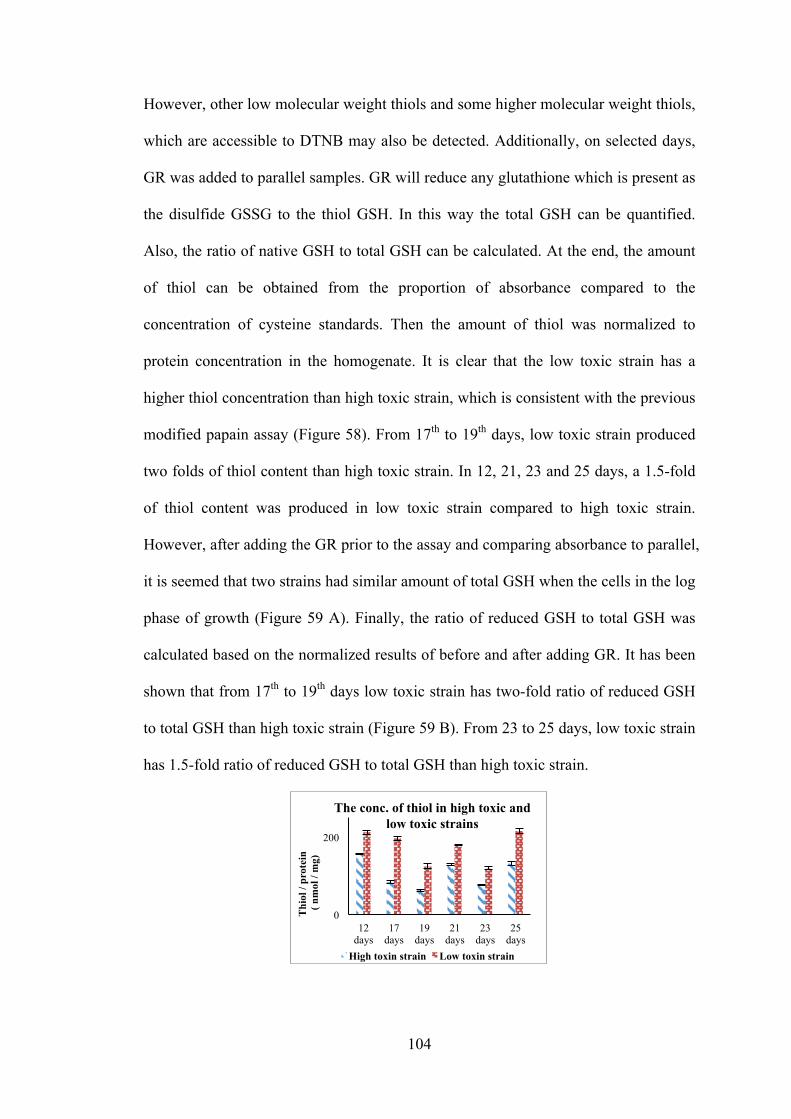
104
However, other low molecular weight thiols and some higher molecular weight thiols,
which are accessible to DTNB may also be detected. Additionally, on selected days,
GR was added to parallel samples. GR will reduce any glutathione which is present as
the disulfide GSSG to the thiol GSH. In this way the total GSH can be quantified.
Also, the ratio of native GSH to total GSH can be calculated. At the end, the amount
of thiol can be obtained from the proportion of absorbance compared to the
concentration of cysteine standards. Then the amount of thiol was normalized to
protein concentration in the homogenate. It is clear that the low toxic strain has a
higher thiol concentration than high toxic strain, which is consistent with the previous
modified papain assay (Figure 58). From 17th to 19th days, low toxic strain produced
two folds of thiol content than high toxic strain. In 12, 21, 23 and 25 days, a 1.5-fold
of thiol content was produced in low toxic strain compared to high toxic strain.
However, after adding the GR prior to the assay and comparing absorbance to parallel,
it is seemed that two strains had similar amount of total GSH when the cells in the log
phase of growth (Figure 59 A). Finally, the ratio of reduced GSH to total GSH was
calculated based on the normalized results of before and after adding GR. It has been
shown that from 17th to 19th days low toxic strain has two-fold ratio of reduced GSH
to total GSH than high toxic strain (Figure 59 B). From 23 to 25 days, low toxic strain
has 1.5-fold ratio of reduced GSH to total GSH than high toxic strain.
0
200
12 days
17 days
19 days
21 days
23 days
25 days
Thi
ol /
prot
ein
( nm
ol /
mg)
The conc. of thiol in high toxic and low toxic strains
High toxin strain Low toxin strain
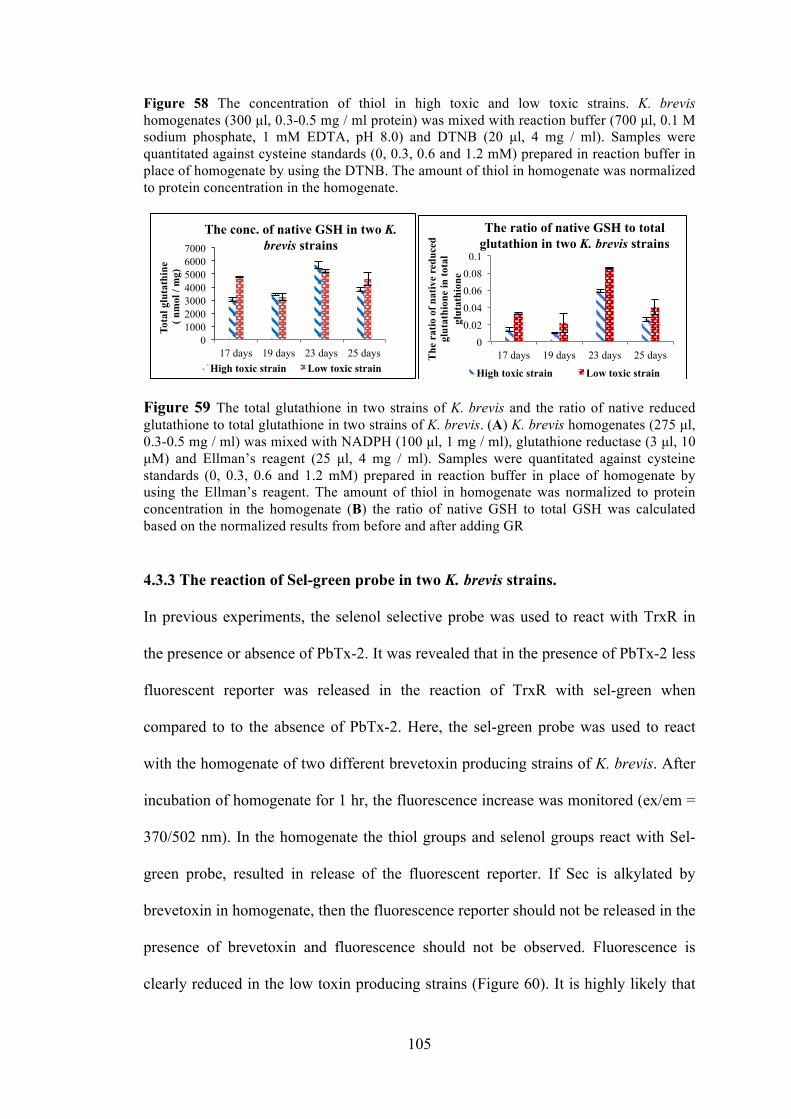
105
Figure 58 The concentration of thiol in high toxic and low toxic strains. K. brevis homogenates (300 µl, 0.3-0.5 mg / ml protein) was mixed with reaction buffer (700 µl, 0.1 M sodium phosphate, 1 mM EDTA, pH 8.0) and DTNB (20 µl, 4 mg / ml). Samples were quantitated against cysteine standards (0, 0.3, 0.6 and 1.2 mM) prepared in reaction buffer in place of homogenate by using the DTNB. The amount of thiol in homogenate was normalized to protein concentration in the homogenate.
Figure 59 The total glutathione in two strains of K. brevis and the ratio of native reduced glutathione to total glutathione in two strains of K. brevis. (A) K. brevis homogenates (275 µl, 0.3-0.5 mg / ml) was mixed with NADPH (100 µl, 1 mg / ml), glutathione reductase (3 µl, 10 µM) and Ellman’s reagent (25 µl, 4 mg / ml). Samples were quantitated against cysteine standards (0, 0.3, 0.6 and 1.2 mM) prepared in reaction buffer in place of homogenate by using the Ellman’s reagent. The amount of thiol in homogenate was normalized to protein concentration in the homogenate (B) the ratio of native GSH to total GSH was calculated based on the normalized results from before and after adding GR
4.3.3 The reaction of Sel-green probe in two K. brevis strains.
In previous experiments, the selenol selective probe was used to react with TrxR in
the presence or absence of PbTx-2. It was revealed that in the presence of PbTx-2 less
fluorescent reporter was released in the reaction of TrxR with sel-green when
compared to to the absence of PbTx-2. Here, the sel-green probe was used to react
with the homogenate of two different brevetoxin producing strains of K. brevis. After
incubation of homogenate for 1 hr, the fluorescence increase was monitored (ex/em =
370/502 nm). In the homogenate the thiol groups and selenol groups react with Sel-
green probe, resulted in release of the fluorescent reporter. If Sec is alkylated by
brevetoxin in homogenate, then the fluorescence reporter should not be released in the
presence of brevetoxin and fluorescence should not be observed. Fluorescence is
clearly reduced in the low toxin producing strains (Figure 60). It is highly likely that
0 1000 2000 3000 4000 5000 6000 7000
17 days 19 days 23 days 25 days
Tota
l glu
tath
ine
( nm
ol /
mg)
The conc. of native GSH in two K. brevis strains
High toxic strain Low toxic strain
0 0.02 0.04 0.06 0.08 0.1
17 days 19 days 23 days 25 days The
rat
io o
f nat
ive
redu
ced
glut
athi
one
in to
tal
glut
athi
one
The ratio of native GSH to total glutathion in two K. brevis strains
High toxic strain Low toxic strain
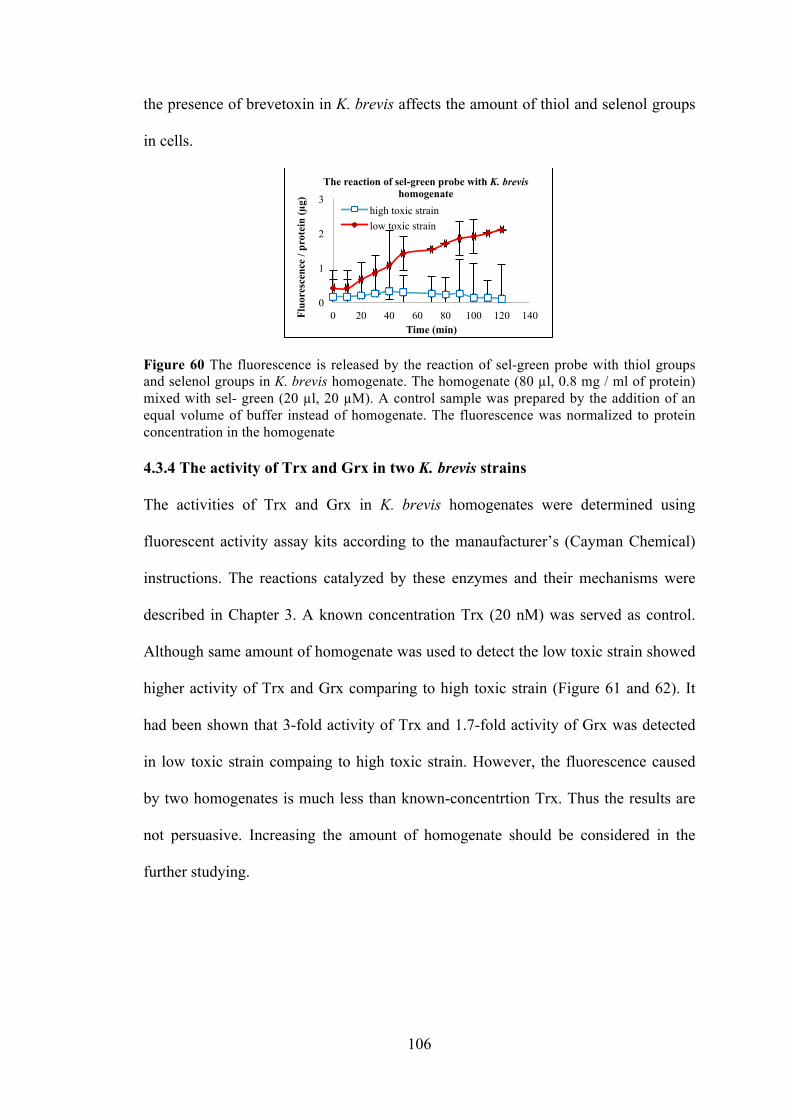
106
the presence of brevetoxin in K. brevis affects the amount of thiol and selenol groups
in cells.
Figure 60 The fluorescence is released by the reaction of sel-green probe with thiol groups and selenol groups in K. brevis homogenate. The homogenate (80 µl, 0.8 mg / ml of protein) mixed with sel- green (20 µl, 20 µM). A control sample was prepared by the addition of an equal volume of buffer instead of homogenate. The fluorescence was normalized to protein concentration in the homogenate 4.3.4 The activity of Trx and Grx in two K. brevis strains
The activities of Trx and Grx in K. brevis homogenates were determined using
fluorescent activity assay kits according to the manaufacturer’s (Cayman Chemical)
instructions. The reactions catalyzed by these enzymes and their mechanisms were
described in Chapter 3. A known concentration Trx (20 nM) was served as control.
Although same amount of homogenate was used to detect the low toxic strain showed
higher activity of Trx and Grx comparing to high toxic strain (Figure 61 and 62). It
had been shown that 3-fold activity of Trx and 1.7-fold activity of Grx was detected
in low toxic strain compaing to high toxic strain. However, the fluorescence caused
by two homogenates is much less than known-concentrtion Trx. Thus the results are
not persuasive. Increasing the amount of homogenate should be considered in the
further studying.
0
1
2
3
0 20 40 60 80 100 120 140 Fluo
resc
ence
/ pr
otei
n (µ
g)
Time (min)
The reaction of sel-green probe with K. brevis homogenate
high toxic strain low toxic strain
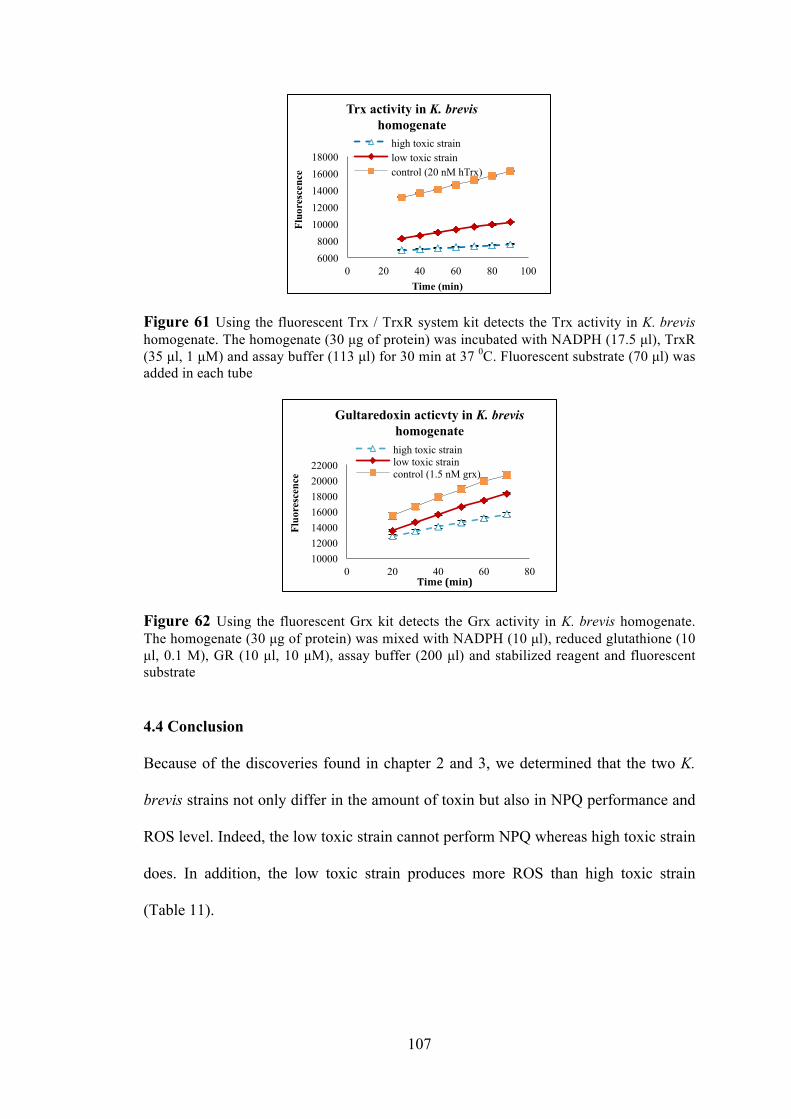
107
Figure 61 Using the fluorescent Trx / TrxR system kit detects the Trx activity in K. brevis homogenate. The homogenate (30 µg of protein) was incubated with NADPH (17.5 µl), TrxR (35 µl, 1 µM) and assay buffer (113 µl) for 30 min at 37 0C. Fluorescent substrate (70 µl) was added in each tube
Figure 62 Using the fluorescent Grx kit detects the Grx activity in K. brevis homogenate. The homogenate (30 µg of protein) was mixed with NADPH (10 µl), reduced glutathione (10 µl, 0.1 M), GR (10 µl, 10 µM), assay buffer (200 µl) and stabilized reagent and fluorescent substrate
4.4 Conclusion
Because of the discoveries found in chapter 2 and 3, we determined that the two K.
brevis strains not only differ in the amount of toxin but also in NPQ performance and
ROS level. Indeed, the low toxic strain cannot perform NPQ whereas high toxic strain
does. In addition, the low toxic strain produces more ROS than high toxic strain
(Table 11).
6000 8000
10000 12000 14000 16000 18000
0 20 40 60 80 100
Fluo
resc
ence
Time (min)
Trx activity in K. brevis homogenate
high toxic strain low toxic strain control (20 nM hTrx)
10000 12000 14000 16000 18000 20000 22000
0 20 40 60 80
Fluo
resc
ence
Time (min)
Gultaredoxin acticvty in K. brevis homogenate high toxic strain low toxic strain control (1.5 nM grx)

108
Table 11 Comparison of NPQ and ROS in two K. brevis strains
NPQ Relative rate of ROS High toxic strain able 21.9 Low toxic strain disable 12.16
The modified papain assay revealed that the low toxic strain of K. brevis has twice
thiol content when it compared to the high toxic strain. The free thiol was monitored
via the Ellman’s assay over the lifetime of the cultures, which revealed that low toxic
strain has twice amount of reduced thiol than high toxic strains from 17 to 19 growing
days. This is also consistent with the previous modified papain assay. Additionally, on
selected days during the log phase of the living cells’ growth cycle, GR that could
reduce any oxidized GSSG to the GSH, was added to parallel samples to quantify
total glutathione and asses the ratio of reduced GSH to total GSH. This showed that
the high toxic strain and low toxic strain had similar total GSH when the cells in the
log phase of growth. However, the low toxic strain has a maximum 2.4-fold ratio of
reduced GSH to total GSH than high toxic strain. In addition to thiols, antioxidant
capacity and ascorbic acid content are not the same in the high toxic and low toxic
strains. At 19th growing days, the high toxic strain has 1.16-fold antioxidant capacity
than low toxic strain. However, 4.5-fold less of ascorbic acid was present in high
toxic strain than low toxic strain, which matched the ratio of native GSH to total GSH
(Table 12). Because in cells GSH can reduce dehydroascorbic acid to ascorbic acid.
Table 12 Comparison of various biochemical parameters in two K. brevis strains at 19th days
Thiol content
Ratio of native GSH to total GSH
Ascorbic acid Antioxidant capacity
High toxic strain 1 1 1 1.16-fold
Low toxic strain 1.97-fold 2.2-fold 4.5-fold 1

109
Taking the results from various assays under consideration, we do find a difference of
redox state, antioxidant capacity, GSH and ascorbic acid content between high and
low toxin-producing strains. It has been determined that TrxR is related to the
production of GSH and ascorbic acid, such as the reduction of dehydroascorbic acid
to ascorbic acid by TrxR. In addition, the discoveries found in chapter 3 reveal that
brevetoxin impacts the activity of Trx/TrxR system, which is the major reductive
system in the cell. Experimental evidence indicates that the effect is caused by the
alkylation of TrxR by brevetoxin. Certain antioxidants such as thiols and ascorbic
acid are negatively correlated with brevetoxin content in K. brevis. These discoveries
provide evidence to support the hypothesis that brevetoixn impacts the reductive
system in cells through reaction with TrxR or other thiol containing proteins. Also,
the low toxin-producing strain has higher level of ROS and ascorbic acid than high
toxin-producing, which is able to support the difference at NPQ performance and
ROS level in two strains. Although it seems that total antioxidant capacity is in
conflict with our expectations, antioxidant capacity includes many enzymatic or non-
enzymatic systems, such as the TrxR/Trx and GSH systems, carotenoids, vitamin E
and so on. It is likely that brevetoxin deficiency or the reaction of brevetoxin with
proteins is the factor that causes the different performance or capacity in K. brevis
cells. But it still need further study to explore.
Furthermore, it is likely that those discoveries perhaps create a new horizon to
understand the role of brevetoxin in K. brevis or the impacts of damage caused by
brevetoxin.

110
4.5 Materials and Methods
4.5.1 Culture methods
Method: K. brevis culture (Wilson strains) were obtained from Mote Marine
Laboratory (Sarasota, Florida) and maintained in L1-Si medium, with the exception
that the NH 15 vitamin supplement (Gates and Wilson 1960) replaced the L-1
supplement, in a growth chamber at ~20ºC under 35 µmol.photons.m-2.s-1. Growth
was monitored by counting a 1:10 dilution of culture in Z pak reagent using a
Beckman Z-series Coulter Counter with aperture size between 10~30 µm according to
the manufacturer’s instructions.
4.5.2 Preparation of K. brevis homogenate
Method K. brevis culture (200–500 ml) was concentrated by centrifugation (5 min at
466 X g) and the supernatant was discarded. The cells were resuspended in
phosphate-buffered saline (PBS) and vortexed for 1 min. This suspension was
centrifuged (10 min at 14,000 X g) to remove cellular debris and the supernatant was
transferred to a clean tube. An aliquot (50 µl) was reserved for a protein assay.
Samples were quantitated against BSA standards (0, 1, 0.5, 0.25, 0.125, and 0.0625
mg /ml) prepared in PBS using the Coomassie Protein Assay Reagent (BioRad)
according to the manufacturer’s instructions. Final protein concentrations ranged from
0.2–1 mg / ml
4.5.3 ABTS assay
The materials for ABTS assay ABTS (2,2-azinobis (3-ethenelbenzothiazoline-6-
sulfonic acid)), (NH4)2S2O8, Trolox (6-hydroxy-2,5,7,8-tetramethylchroman-2-
carboxylic acid), ascorbic acid oxidase. High and low toxic K. brevis strain

111
homogenate was prepared through the pervious method. UV/Visible measurements
were performed in 96 or 384 well microplates using a Synergy® 2 (Biotek Instrument,
Inc.)
The method for ABTS assay Total antioxidant capacity of K. brevis homogenate
was determined by 2,2-azinobis (3-ethenelbenzothiazoline-6-sulfonic acid) radical
cation (ABTS.+) decolorization and expressed as trolox equivalents. ABTS.+ was
produced by oxidation of aqueous ABTS (1.3 mL, 7 mM) with ammonium persulfate
(21.2 µl, 2.45 mM). The mixture was allowed to stand at room temperature in the
dark for 12-16 h before use. The ABTS.+ solution was diluted with PBS to an
absorbance of 0.7. Either 50 ul K. brevis homogenate or trolox standards (0 µM, 25
µM, 50 µM, 100µM, 200 µM) were added to the ABTS.+ solution (100 µl). The
absorbance was read at 734 nM after incubation for 5 min. Ascorbic acid
concentration (as trolox equivalents) of K. brevis homogenate was estimated by
incubating with of ascorbic acid oxidase (2 µl, 0.01 U/ µL) for 30 min prior to the
assay and comparing absorbance to parallel samples.
4.5.4 Modified papain assay
Materials for modified papain assay: Papain, methyl methanethiosulfonate, N-
benzoyl-L-arginine-p-nitroanilide and cystamine were purchased from Thermo
Scientific Co.. High and low toxic K. brevis strain homogenate was prepared through
the pervious method. UV/Visible measurements were performed in 96 or 384 well
microplates using a Synergy® 2 (Biotek Instrument, Inc.)

112
Preparation of papain-S-SCH3 (~1.2 mg / ml)1.5 ml of papain stock solution (26.4
mg / ml) was added to a solution of cysteine (0.6 mM, 28.5 ml of 20 mM sodium
phosphate – 1 mM EDTA buffer). The solution was kept at room temperature for 30
min. The methyl methanethiosulfonate (117 µmol) was added in mixture. The
reaction mixture was kept in ice for 4 hr. The mixture was dialyzed (Mr cutoff 6000-
8000) against 5 mM sodium acetate buffer, containing 50 mM NaCl, pH 4.7 at 4oC 12
hr.
Method for modified papain assay: The culture was treated under dark adapt or sun
light for 1 hr before making homogenate. Papain-S-SCH3 solution (0.5 ml, 0.6 mg /
ml), 0.7 ml substrate and 15 µl cystamine were added in one test tube which contained
200 µl supernatant. A tube containing papain-S-SCH3, without homogenate or
substrate, serves as control or background. The mixture was carried out in triplicate in
plate. Each well was added 300 µl mixture. Record the absorbance at 410 nm
(Synergy™ 2, Bioteck Instrument, Inc.) every 60 min up to 15 hr.
4.5.5 Ellman’s assay
Materials for Ellman’s assay: DTNB and cysteine was purchased from Thermo
Scientific Co.. High and low toxic K. brevis strain homogenate was prepared through
the pervious method. Glutathione reductase was from Grx kit purchased from
Cayman Chemical. UV/Visible measurements were performed in 96 or 384 well
microplates using a Synergy® 2 (Biotek Instrument, Inc.)
Method for Ellman’s assay: K. brevis homogenates (300 µl, 0.3-0.5 mg / ml protein)
was mixed with reaction buffer (700 µl, 0.1 M sodium phosphate, 1 mM EDTA, pH

113
8.0) and Ellman’s reagent (20 µl, 4 mg / ml). After incubation for 15 min at room
temperature absorbance was read at 410 nm (Synergy™ 2, Bioteck Instrument, Inc).
Homogenate prepared similarly, but without Ellman’s reagent was used to determine
background absorbance which was subtracted from each sample. Ellman’s reagent in
reaction buffer served as the control. Samples were quantitated against cysteine
standards (0, 0.3, 0.6 and 1.2 mM) prepared in reaction buffer in place of homogenate
by using the Ellman’s reagent according to the manufacturer’s instructions (Thermo
Fisher). Samples and cysteine standards were analyzed in triplicate. The amount of
thiol in homogenate was normalized to protein concentration in the homogenate.
Ellman’s assay of two strains with GR K. brevis homogenates (275 µl, 0.3-0.5 mg /
ml) was mixed with NADPH (100 µl, 1 mg / ml), glutathione reductase (3 µl, 10 µM)
and Ellman’s reagent (25 µl, 4 mg / ml). Immediately the absorbance was read at 410
nm at room temperature up to 30 min. Homogenate prepared similarly, but without
Ellman’s reagent was used to determine background absorbance which was subtracted
from each sample. Ellman’s reagent in reaction buffer served as the control. Samples
were quantitated against cysteine standards (0, 0.3, 0.6 and 1.2 mM) prepared in
reaction buffer in place of homogenate by using the Ellman’s reagent. Samples and
cysteine standards were analyzed in triplicate.
4.5.6 Sel- green reacts with homogenate of tow K. brevis strains
Materials: Sel-Green was synthesized by Freddy Rodriguez. High and low toxic K.
brevis strain homogenate was prepared through the pervious method. UV/Visible
measurements were performed in 96 or 384 well microplates using a Synergy® 2
(Biotek Instrument, Inc.)

114
Method: The selenol selective probe, Sel-green, was synthesized according to
published methods (Zhang, Ge et al. 2015). The homogenate was made resulting in
0.8 mg / ml of protein both in high toxic and low toxic homogenate. Sel- green (20 µl,
20 µM) was mixed with high toxic strain or low toxic strain homogenate (80 µl).
Samples were prepared in triplicate. A control sample was prepared by the addition of
an equal volume of buffer instead of homogenate. A blank sample was prepared
without Sel-green. Fluorescence at 502 (λex = 370 nm) was monitored for 120 mins.
4.5.7 Thioredoxin activity in homogenate of two strains K. brevis
Materials: Enzyme assay kit was purchased from Cayman Chemical. Assay kit was
used according to the manufacturer’s instructions with noted exceptions. High and
low toxic K. brevis strain homogenate was prepared through the pervious method.
Fluorescence measurements were performed in 96 or 384 well microplates using an
Infinite® M1000 PRO (Tecan Group Ltd.) microplate readers.
Method: The homogenate was made resulting in 1.0 mg / ml and 1.5 mg / ml of
protein high toxic and low toxic homogenate. Assay buffer (113 µl, 0.2 mg / ml
bovine serum albumin in 50 mM Tris-HCl and 1 mM EDTA, pH 7.5) was mixed with
17.5 µl NADPH in one clean tube. Thioredoxin reductase (35 µl, 1 µM) was added
into the mixture. The homogenate was incubated with the mixture for 30 min at 37 0C.
70 µl fluorescent substrate was added in each tube. The mixture was carried out in
triplicate in plate. Each well is added 100 µl mixture. Record its fluorescence at the
emission / excitation at 545 nm / 520 nm every 10 min for 3 hr.

115
4.5.8 Grx activity in homogenate of two strains K. brevis
Materials: Fluorescent Grx kit was purchase from Cayman Chemical. High and low
toxic K. brevis strain homogenate was prepared as mentioned above. Fluorescence
measurements were performed in 96 or 384 well microplates using an Infinite®
M1000 PRO (Tecan Group Ltd.) microplate readers.
Method: The homogenate was made resulting in 1.0 mg / ml and 1.5 mg / ml of
protein high toxic and low toxic homogenate. Assay buffer (200 µl) was mixed with
reduced glutathione (10 µl, 0.1 M) in one clean tube. NADPH (10 µl) and stabilized
reagent were added with mixture. Glutathione reductase (10 µl, 10 µM) was applied
into the mixture. The distilled water was added it up to 1 ml. The homogenate and
fluorescent substrate were added with mixture in each tube. Immediately the mixture
was carried out in triplicate in plate. Each well was added 100 µl mixture. Record its
fluorescence at the emission / excitation at 545 nm / 520 nm every 10 min for 3 hr.

116
Chapter 5 Summary and future work
5.1 Summary
This dissertation describes efforts to understand the principle role that brevetoxin
plays in K. brevis. The dissertation includes four components: i) localization of
brevetoxin in K. brevis cells ii) identification of the target proteins of brevetoxin iii)
characterization of the interaction between a target protein and brevetoxin iv)
comparison of the various metabolic and biochemical parameters in two strains of K.
brevis having different toxin content.
Brevetoxin was localized to the chloroplast of live K. brevis cells through the
fluorescently labeled brevetoxin probe. Using a brevetoxin photoaffinity probe, two
native receptors of brevetoxin were discovered from K. brevis. Based on the BLAST
analysis, LHC II and Trx were identified as binding partners for brevetoxin in K.
brevis. An interaction between brevetoxin and a transmembrane target protein, LHC
II, was discovered, which is related to the NPQ mechanism to protect the cells from
damage by ROS. Also, PbTx-2 appeared to inhibit the Trx/TrxR system. However,
PbTx-2 promoted the reduction of DTNB by TrxR. Although there is no direct
evidence to prove PbTx-2 reacts with mammalian TrxR by alkylation of the α, β-
unsaturated aldehyde of brevetoxin with selenocysteine in TrxR, several pieces of
indirect evidence supported this conclusion. Because of this deactivation, mammalian
TrxR could not reduce the Trx, and thus, the insulin disulfide bonds could not be
reduced by Trx either. However, the reaction center on N-terminal of mammalian
TrxR still can reduce some small molecule such as DTNB.
Thus, according to the identification of the two native receptors and interaction
between brevetoxin with mammalian Trx/TrxR system, follows that the redox state

117
and antioxidant capacity of K. brevis may be related to toxin content. It had been
revealed that high toxic strain could perform NPQ whereas low toxic strain could not
perform NPQ. The low toxic strain had higher levels of ROS than the high toxic strain.
The modified papain assay, Ellman’s assay, and ABTS assay were used to compare
the free thiol groups and antioxidants in the high and low toxic strains. In general, the
high toxic strain had a higher antioxidant capacity than the low toxic strain. However,
the low toxic strain consistently exhibited higher levels of free thiol groups and
ascorbic acid. Both of these would be influenced by the TrxR/Trx activity in the cells.
The low toxic strain has slightly higher activity of Trx system and Grx compared to
high toxic strain. Therefore, brevetoxin content seems to be correlated with redox
homeostasis of the cells.
It is likely that brevetoxins plays an important role in K. brevis. It is highly possible
that brevetoxin affects the reductive system in cells, leading to the impact on various
metabolic and biochemical parameters in two strains of K. brevis. In order to better
understand these phenomenons, additional experiments are still needed to be done to
prove our hypothesis.
5.2 Future work
5.2.1 Identify all biological receptors for brevetoxin in K. brevis and localize the
binding site on native receptors
Our receptor binding experiments at pH 7 have consistently shown one additional
high MW bands between 57 and 75 kD (Figure 63). A third band at 37 kD has been
observed sporadically. These two bands have been excised from gels and can be sent
immediately to the mass spectrometry facility at the University of Massachusetts at

118
Amherst to be identified.
Figure 63 Spinach homogenate was incubated with photoaffinity probe in PBS (pH=7) for 1 hour. Lane 1: Molecular Weight markers Lane 2: The first elution. Lane3: The second elution. Lane4: Total protein. Because the increase effect of TrxR by PbTx-3 was observed as well in DTNB assay.
Also, PbTx-3 and PbTx-2 both have an aldehyde at A-ring. We suspect PbTx-3 may
bind or react with Trx or TrxR enzyme as well. Perhaps, the A-ring is another active
site for brevetoxin B. Thus, a brevetoxin (PbTx-2 or PbTx-3) photoaffinity probe
(Figure 64) which includes a photoactive diazirine to interact with its receptor can be
synthesized to incubate with K. brevis homogenate. This time, the ligand will be
introduced A-ring position in PbTx-2 or PbTx-3. The photolysis, purification and
identification are same as previous experiment. We may or may not discover new
target proteins.
Figure 64 A-ring derivative from PbTx-2 or PbTx-3 A-ring diol If in order to identify the specific binding amino acid in target protein, the same
photoaffinity probe (K-ring derivative or A- ring derivative) is incubated with K.
O
CF3
N
N
3
O
O
MeO
HOH
H
B

119
brevis homogenate. After photolysis, the probe may react to form a covalent cross-
link to a variety of amino acid residues within the target protein. This time, the protein
sample will be digested by trypsin to its peptide constituents before purification. The
same procedure will be used to isolate the full protein by passing the digest through a
strepavadin column. The target fragment, linked with biotin will be eluted from the
streptavidin column by heat treatment. SDS-PAGE will be performed to analyze the
captured fragments. Finally, send the bands to the mass spectrometry facility at the
University of Massachusetts at Amherst.
5.2.2 Analysis of the essential proteins, carotenoids and lipids for NPQ in low vs
high toxin K. brevis
The low toxic strain does not perform NPQ. It is still unclear if the inability to
perform NPQ is due to the absence of brevetoxin, or if the lack of brevetoxin is a
result of the NPQ deficiency. It is necessary to determine if the low toxic strain is
deficient in one or more of the essential components of NPQ: zeaxanthin and
violaxanthin.
Quantitative polymerase chain reaction (qPCR) can be used to amplify and quantify
the VDE and ZE mRNA in low and high toxic strains. If we can determine the RNA
of VDE and ZE in two strains, we will do further study at translational level. Thus,
western blot analysis can be used in further studies to compare protein extracts of the
low and high toxic strains of K. brevis using three different antibodies: LHC II, VDE
and ZE. These three antibodies can be purchased from Swedish company Agrisera.
Hopefully it is possible to determine if one or more of these essential components is
absent or reduced in the low-toxin culture.

120
5.2.3 Determination the role of brevetoxin in two K. brevis strains
Although the difference of NPQ, ROS, antioxidant capacity and biochemical
parameters in high toxin and low toxin producing strains of K. brevis were determined,
the reasons caused this phenomenon were still unknown. Does brevetoxin induce
those difference? In order to understand it, my idea is adding PbTx-2 with low toxic
strain for 25 days. The cells are used unused medium to wash at least twice. The
homogenate is made as previous method. DTNB assay, ROS and ABTS assay can be
applied again to compare the difference between high toxic strain and low toxic strain
which is treated with PbTx-2. The difference becomes smaller or totally disappear,
which means brevetoxin is one of the reasons that cause the distinction between high
and low toxic strains.

121
Reference
Abraham, A., S. M. Plakas, L. J. Flewelling, K. R. El Said, E. L. Jester, H. R. Granade, K. D. White and R. W. Dickey (2008). "Biomarkers of neurotoxic shellfish poisoning." Toxicon 52(2): 237-245.
Abraham, A., S. M. Plakas, Z. Wang, E. L. Jester, K. R. El Said, H. R. Granade, M. S. Henry, P. C. Blum, R. H. Pierce and R. W. Dickey (2006). "Characterization of polar brevetoxin derivatives isolated from Karenia brevis cultures and natural blooms." Toxicon 48(1): 104-115.
Abraham, W. M. and D. G. Baden (2006). "Aerosolized Florida red tide toxins and human health effects." Oceanography 19(2): 107-109.
Abraham, W. M., A. J. Bourdelais, A. Ahmed, I. Serebriakov and D. G. Baden (2005). "Effects of inhaled brevetoxins in allergic airways: toxin-allergen interactions and pharmacologic intervention." Environmental Health Perspectives 113(6): 632-637.
Abraham, W. M., A. J. Bourdelais, J. R. Sabater, A. Ahmed, T. A. Lee, I. Serebriakov and D. G. Baden (2005). "Airway responses to aerosolized brevetoxins in an animal model of asthma." American Journal of Respiratory and Critical Care Medicine Home 171(1): 26-34.
Aebi, H. (1984). "Catalase in vitro." Methods in Enzymology 105: 121-126.
Anderson, C. R., S. K. Moore, M. C. Tomlinson, J. Silke and C. K. Cusack (2015). "Living with harmful algal blooms in a changing world: strategies for modeling and mitigating their effects in coastal marine ecosystems." Coastal and Marine Hazards, Risks, and Disasters. Boston, Elsevier: 495-561.
Anestål, K., S. Prast-Nielsen, N. Cenas and E. S. J. Arnér (2008). "Cell death by SecTRAPs: thioredoxin reductase as a prooxidant killer of cells." Plos One 3(4): e1846.
Araie, H., I. Suzuki and Y. Shiraiwa (2008). "Identification and characterization of a selenoprotein, thioredoxin reductase, in a unicellular marine haptophyte alga, Emiliania huxleyi." Journal of Biological Chemistry 283(51): 35329-35336.
Armstrong, R. N. (1997). "Structure, catalytic mechanism, and evolution of the glutathione transferases." Chemical Research in Toxicology 10(1): 2-18.
Arnér, E. S. (2009). "Focus on mammalian thioredoxin reductases—important selenoproteins with versatile functions." Biochimica et Biophysica Acta 1790(6): 495-526.
Arnér, E. S. and A. Holmgren (2000). "Physiological functions of thioredoxin and thioredoxin reductase." European Journal of Biochemistry 267(20): 6102-6109.

122
Arnér, E. S., L. Zhong and A. Holmgren (1999). "Preparation and assay of mammalian thioredoxin and thioredoxin reductase." Methods in Enzymology 300: 226-239.
Arsalane, W., B. Rousseau and J. C. Duval (1994). "Influence of the pool size of the xanthophyll cycle on the effects of light stress in a diatom: competition between photoprotection and photoinhibition." Photochemical and Photobiological Sciences 60(3): 237-243.
Arscott, L. D., S. Gromer, R. H. Schirmer, K. Becker and C. H. Williams (1997). "The mechanism of thioredoxin reductase from human placenta is similar to the mechanisms of lipoamide dehydrogenase and glutathione reductase and is distinct from the mechanism of thioredoxin reductase from Escherichia coli." Proceedings of the National Academy of Sciences 94(8): 3621-3626.
Aslund, F., B. Ehn, A. Miranda-Vizuete, C. Pueyo and A. Holmgren (1994). "Two additional glutaredoxins exist in Escherichia coli: glutaredoxin 3 is a hydrogen donor for ribonucleotide reductase in a thioredoxin/glutaredoxin 1 double mutant." Proceedings of the National Academy of Sciences 91(21): 9813-9817.
Azzone, G. F. (1972). "Biochemistry and biophysics of mitochondrial membranes." Yale Journal of Biology and Medicine 47(1): 68.
Backer, L., L. Fleming, A. Rowan and D. Baden (2003). "Epidemiology and public health of human illnesses associated with harmful marine phytoplankton." Handbook of Developmental Neurotoxicology: 725-750.
Backer, L. C., B. Kirkpatrick, L. E. Fleming, Y. S. Cheng, R. Pierce, J. A. Bean, R. Clark, D. Johnson, A. Wanner, R. Tamer, Y. Zhou and D. G. Baden (2005). "Occupational exposure to aerosolized brevetoxins during Florida Red Tide events: effects on a healthy worker population." Environmental Health Perspectives 113(5): 644-649.
Baden, D. G. (1989). "Brevetoxins: unique polyether dinoflagellate toxins." The FASEB Journal 3(7): 1807-1817.
Baden, D. G., A. J. Bourdelais, H. Jacocks, S. Michelliza and J. Naar (2005). "Natural and derivative brevetoxins: historical background, multiplicity, and effects." Environmental Health Perspectives 113(5): 621-625.
Baden, D. G., T. J. Mende, W. Lichter and L. Wellham (1981). "Crystallization and toxicology of T34: A major toxin from Florida's red tide organism (Ptychodiscus brevis)." Toxicon 19(4): 455-462.
Baker, A., C. M. Payne, M. M. Briehl and G. Powis (1997). "Thioredoxin, a gene found overexpressed in human cancer, inhibits apoptosis in vitro and in vivo." Cancer Research 57(22): 5162-5167.

123
Bauer, H., S. Gromer, A. Urbani, M. Schnölzer, R. H. Schirmer and H. M. Müller (2003). "Thioredoxin reductase from the malaria mosquito Anopheles gambiae." European Journal of Biochemistry 270(21): 4272-4281.
Becker, K., S. Gromer, R. H. Schirmer and S. Müller (2000). "Thioredoxin reductase as a pathophysiological factor and drug target." European Journal of Biochemistry 267(20): 6118-6125.
Benson, J., F. Hahn, B. Tibbetts, L. Bowen, T. March, R. Langley, T. Murray, A. Bourdelais, J. Naar and J. Zaias (2002). "Florida red tide: inhalation toxicity of Karenia brevis extract in rats." Harmful Algae 2004(10)501-504
Benson, J. M., B. B. Stagner, G. K. Martin, M. Friedman, S. E. Durr, A. Gomez, J. McDonald, L. E. Fleming, L. C. Backer and D. G. Baden (2005). "Cochlear function in mice following inhalation of brevetoxin-3." Journal of Comparative Physiology A 191(7): 619-626.
Benson, J. M., D. L. Tischler and D. G. Baden (1999). "Uptake, tissue distribution, and excretion of brevetoxin 3 administered to rats by intratracheal instillation." Journal of Toxicology and Environmental Health Part A 57(5): 345-355.
Berta, P., S. Phaneuf, J. Derancourt, J. Casanova, M. Durand-Clement, C. le Peuch, J. Haiech and J.-C. Cavadore (1988). "The effects of maitotoxin on phosphoinositides and calcium metabolism in a primary culture of aortic smooth muscle cells." Toxicon 26(2): 133-141.
Betteridge, D. J. (2000). "What is oxidative stress?" Metabolism 49(2): 3-8.
Bodeanu, N., S. Moncheva, G. Ruta and L. Popa (1998). "Long-term evolution of the algal blooms in Romanian and Bulgarian Black Sea waters." Cercetari Marine/Recherches Marines 31: 37-55.
Bogdan, C. (2001). "Nitric oxide and the regulation of gene expression." Trends in Cell Biology 11(2): 66-75.
Bormann, B. J., W. J. Knowles and V. T. Marchesi (1989). "Synthetic peptides mimic the assembly of transmembrane glycoproteins." The Journal of Biological Chemistry 264(7): 4033-4037.
Bossart, G. D., D. G. Baden, R. Y. Ewing, B. Roberts and S. D. Wright (1998). "Brevetoxicosis in manatees (Trichechus manatus latirostris) from the 1996 epizootic: gross, histologic, and immunohistochemical features." Toxicologic Pathology 26(2): 276-282.
Bourdelais, A. J., S. Campbell, H. Jacocks, J. Naar, J. L. Wright, J. Carsi and D. G. Baden (2004). "Brevenal is a natural inhibitor of brevetoxin action in sodium channel receptor binding assays." Cellular and Molecular Neurobiology 24(4): 553-563.
Bradshaw, T. D., C. S. Matthews, J. Cookson, E.-H. Chew, M. Shah, K. Bailey, A. Monks, E. Harris, A. D. Westwell, G. Wells, C. A. Laughton and M. F. G. Stevens

124
(2005). "Elucidation of thioredoxin as a molecular target for antitumor quinols." Cancer Research 65(9): 3911-3919.
Bricker, S. B., B. Longstaff, W. Dennison, A. Jones, K. Boicourt, C. Wicks and J. Woerner (2008). "Effects of nutrient enrichment in the nation's estuaries: A decade of change." Harmful Algae 8(1): 21-32.
Bulaj, G., T. Kortemme and D. P. Goldenberg (1998). "Ionization-reactivity relationships for cysteine thiols in polypeptides." Biochemistry 37(25): 8965-8972.
Burkholder, J. M., D. A. Dickey, C. A. Kinder, R. E. Reed, M. A. Mallin, M. R. McIver, L. B. Cahoon, G. Melia, C. Brownie, J. Smith, N. Deamer, J. Springer, H. B. Glasgow and D. Toms (2006). "Comprehensive trend analysis of nutrients and related variables in a large eutrophic estuary: A decadal study of anthropogenic and climatic influences." Limnology and Oceanography 51(1part2): 463-487.
Butterfield, D. A., T. Koppal, B. Howard, R. Subramaniam, N. Hall, K. Hensley, S. Yatin, K. Allen, M. Aksenov and M. Aksenova (1998). "Structural and functional changes in proteins Iinduced by free radical-mediated oxidative stress and protective action of the antioxidants N‐tert‐butyl‐α‐phenylnitrone and vitamin E." Annals of the New York Academy of Sciences 854(1): 448-462.
Cadenas, E. and K. J. Davies (2000). "Mitochondrial free radical generation, oxidative stress, and aging." Free Radical Biology and Medicine 29(3): 222-230.
Carvalho, A. P., P. A. Fernandes and M. J. Ramos (2006). "Similarities and differences in the thioredoxin superfamily." Progress in Biophysics and Molecular Biology 91(3): 229-248.
Cassell, R. T., W. Chen, S. Thomas, L. Liu and K. S. Rein (2015). "Brevetoxin, the dinoflagellate neurotoxin, localizes to thylakoid membranes and interacts with the light‐harvesting complex II (LHCII) of photosystem II." ChemBioChem 16(7): 1060-1067.
Catterall, W. A. (1977). "Activation of the action potential Na+ ionophore by neurotoxins. An allosteric model." The Journal of Biological Chemistry 252(23): 8669-8676.
Catterall, W. A. (1980). "Neurotoxins that act on voltage-sensitive sodium channels in excitable membranes." Annual Review of Pharmacology and Toxicology 20(1): 15-43.
Catterall, W. A. (1986). "Molecular properties of voltage-sensitive sodium channels." Annual Review Biochemistry 55: 953-985.
Catterall, W. A., C. Morrow, J. Daly and G. Brown (1981). "Binding of batrachotoxinin A 20-alpha-benzoate to a receptor site associated with sodium channels in synaptic nerve ending particles." The Journal of Biological Chemistry 256(17): 8922-8927.

125
Cenas, N., H. Nivinskas, Z. Anusevicius, J. Sarlauskas, F. Lederer and E. S. J. Arnér (2004). "Interactions of quinones with thioredoxin reductase: a challenge of the antioxidant role of the mammalian selenoprotein." The Journal of Biological Chemistry 279(4): 2583-2592.
Chae, H. Z., S. W. Kang and S. G. Rhee (1999). "Isoforms of mammalian peroxiredoxin that reduce peroxides in presence of thioredoxin." Methods in Enzymology 300: 219-226.
Chae, H. Z., H. J. Kim, S. W. Kang and S. G. Rhee (1999). "Characterization of three isoforms of mammalian peroxiredoxin that reduce peroxides in the presence of thioredoxin." Diabetes Research and Clinical Practice 45(2): 101-112.
Checchetto, V., E. Teardo, L. Carraretto, E. Formentin, E. Bergantino, G. M. Giacometti and I. Szabo (2013). "Regulation of photosynthesis by ion channels in cyanobacteria and higher plants." Biophysical Chemistry 182: 51-57.
Cheng, Q., T. Sandalova, Y. Lindqvist and E. S. Arnér (2009). "Crystal structure and catalysis of the selenoprotein thioredoxin reductase 1." Journal of Biological Chemistry 284(6): 3998-4008.
Cheng, Y. S., J. D. McDonald, D. Kracko, C. M. Irvin, Y. Zhou, R. H. Pierce, M. S. Henry, A. Bourdelaisa, J. Naar and D. G. Baden (2005). "Concentration and particle size of airborne toxic algae (brevetoxin) derived from ocean red tide events." Environmental Science and Technology 39(10): 3443-3449.
Cheng, Y. S., T. A. Villareal, Y. Zhou, J. Gao, R. H. Pierce, D. Wetzel, J. Naar and D. G. Baden (2005). "Characterization of red tide aerosol on the Texas coast." Harmful Algae 4(1): 87-94.
Cheng, Y. S., Y. Zhou, C. M. Irvin, R. H. Pierce, J. Naar, L. C. Backer, L. E. Fleming, B. Kirkpatrick and D. G. Baden (2005). "Characterization of marine aerosol for assessment of human exposure to brevetoxins." Environmental Health Perspectives 113(5): 638-643.
Collet, J.-F. and J. Messens (2010). "Structure, function, and mechanism of thioredoxin proteins." Antioxidants and Redox Signaling 13(8): 1205-1216.
Collins, M., R. Knutti, J. Arblaster, J. Dufresne, T. Fichefet, P. Friedlingstein, X. Gao, W. Gutowski, T. Johns and G. Krinner (2013). "Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change." Climate Change.
Darcie, E. R., E. P. Alan and L. Campbell (2014). "De novo assembly and characterization of the transcriptome of the toxic dinoflagellate Karenia brevis." BioMedCentral Genomics15(1) 888
Dechraoui, M. Y., J. Naar, S. Pauillac and A.-M. Legrand (1999). "Ciguatoxins and brevetoxins, neurotoxic polyether compounds active on sodium channels." Toxicon 37(1): 125-143.

126
Dechraoui, M.-Y. B. and J. S. Ramsdell (2003). "Type B brevetoxins show tissue selectivity for voltage-gated sodium channels: comparison of brain, skeletal muscle and cardiac sodium channels." Toxicon 41(7): 919-927.
Demers, S., S. Roy and R. Gagnon (1991)." Rapid light-induced changes in cell fluorescence and in xanthophyll-cycle pigments of Alexandrium excavatum (dinophyceae) and Thalassiosira pseudonana (bacillariophyceae): a photo-protection mechanism." Marine Ecology Progress Series 76: 185-193.
Derick, H., E. Williams and E. Cadenas (2001). "Mitochondrial respiratory chain-dependent generation of superoxide anion and its release into the intermembrane space." Biochemical Journal 353(2): 411-416.
Dickinson, B. C. and C. J. Chang (2008). "A targetable fluorescent probe for imaging hydrogen peroxide in the mitochondria of living cells." Journal of the American Chemical Society 130(30): 9638-9639.
Dormán, G. and G. D. Prestwich (2000). "Using photolabile ligands in drug discovery and development." Trends in Biotechnology 18(2): 64-77.
Dyson, K. and D. D. Huppert (2010). "Regional economic impacts of razor clam beach closures due to harmful algal blooms (HABs) on the Pacific coast of Washington." Harmful Algae 9(3): 264-271.
Eckenroth, B. E., B. M. Lacey, A. P. Lothrop, K. M. Harris and R. J. Hondal (2007). "Investigation of the C-Terminal redox center of high-Mr thioredoxin reductase by protein engineering and semisynthesis." Biochemistry 46(33): 9472-9483.
Errera, R. M., A. Bourdelais, M. Drennan, E. Dodd, D. Henrichs and L. Campbell (2010). "Variation in brevetoxin and brevenal content among clonal cultures of Karenia brevis may influence bloom toxicity." Toxicon 55(2): 195-203.
Errera, R. M. and L. Campbell (2011). "Osmotic stress triggers toxin production by the dinoflagellate Karenia brevis." Proceedings of the National Academy of Sciences, USA 108(26): 10597-10601.
Evans, G. and L. Jones (2001). "Economic impact of the 2000 red tide on Galveston County, Texas: A case study." Department of Agricultural Economics, Texas A&M University.
Fang, J. and A. Holmgren (2006). "Inhibition of thioredoxin and thioredoxin reductase by 4-hydroxy-2-nonenal in vitro and in vivo." Journal of the American Chemical Society 128(6): 1879-1885.
Fang, J., J. Lu and A. Holmgren (2005). "Thioredoxin reductase is irreversibly modified by curcumin: a novel molecular mechanism for its anticancer activity " Journal of Biological Chemistry 280(26): 25284-25290.
Finkel, T. (1998). "Oxygen radicals and signaling." Current Opinion in Cell Biology 10(2): 248-253.

127
Finkel, T. and N. J. Holbrook (2000). "Oxidants, oxidative stress and the biology of ageing." Nature 408(6809): 239-247.
Fischer, A. H., K. A. Jacobson, J. Rose and R. Zeller (2008). "Hematoxylin and eosin staining of tissue and cell sections." Cold Spring Harbor Protocols 2008(5): pdb. prot4986.
Fleming, L. E., L. C. Backer and D. G. Baden (2005). "Overview of aerosolized Florida red tide toxins: exposures and effects." Environmental Health Perspectives 113(5): 618-620.
Fleming, L. E., E. Jerez, W. B. B. Stephan, A. Cassedy, J. A. Bean, A. Reich, B. Kirkpatrick, L. Backer, K. Nierenberg and S. Watkins (2007). "Evaluation of harmful algal bloom outreach activities." Marine Drugs 5(4): 208-219.
Fleming, L. E., B. Kirkpatrick, L. C. Backer, J. A. Bean, A. Wanner, D. Dalpra, R. Tamer, J. Zaias, Y. S. Cheng and R. Pierce (2005). "Initial evaluation of the effects of aerosolized Florida red tide toxins (brevetoxins) in persons with asthma." Environmental Health Perspectives 113(5): 650-657.
Fleming, L. E., B. Kirkpatrick, L. C. Backer, J. A. Bean, A. Wanner, A. Reich, J. Zaias, Y. S. Cheng, R. Pierce and J. Naar (2007). "Aerosolized red-tide toxins (brevetoxins) and asthma." Chest Journal 131(1): 187-194.
Fleming, L. E., B. Kirkpatrick, L. C. Backer, C. J. Walsh, K. Nierenberg, J. Clark, A. Reich, J. Hollenbeck, J. Benson and Y. S. Cheng (2011). "Review of Florida red tide and human health effects." Harmful Algae 10(2): 224-233.
Fleming, S. A. (1995). "Chemical reagents in photoaffinity labeling." Tetrahedron 51(46): 12479-12520.
Flewelling, L. J. (2008). "PhD dissertation: vectors of brevetoxins to marine mammals."
Flewelling, L. J., J. P. Naar, J. P. Abbott, D. G. Baden, N. B. Barros, G. D. Bossart, M.-Y. D. Bottein, D. G. Hammond, E. M. Haubold and C. A. Heil (2005). "Brevetoxicosis: red tides and marine mammal mortalities." Nature 435(7043): 755-756.
Frei, B. (1999). "On the role of vitamin C and other antioxidants in atherogenesis and vascular dysfunction." Experimental Biology and Medicine 222(3): 196-204.
Fridovich, I. (1999). "Fundamental aspects of reactive oxygen species, or what's the matter with oxygen?" Annals of the New York Academy of Sciences 893(1): 13-18.
Friedman, M. A., L. E. Fleming, M. Fernandez, P. Bienfang, K. Schrank, R. Dickey, M.-Y. Bottein, L. Backer, R. Ayyar and R. Weisman (2008). "Ciguatera fish poisoning: treatment, prevention and management." Marine Drugs 6(3): 456-479.

128
Fritz-Wolf, K., S. Kehr, M. Stumpf, S. Rahlfs and K. Becker (2011). "Crystal structure of the human thioredoxin reductase-thioredoxin complex." Nature Communications 2: 383.
Fuciman, M., M. M. Enriquez, T. Polívka, L. Dall’Osto, R. Bassi and H. A. Frank (2012). "Role of xanthophylls in light harvesting in green plants: a spectroscopic investigation of mutant LHCII and Lhcb pigment–protein complexes." The Journal of Physical Chemistry B 116(12): 3834-3849.
Galter, D., S. Mihm and W. Droge (1994). "Distinct effects of glutathione disulphide on the nuclear transcription factors kappa B and the activator protein‐1." European Journal of Biochemistry 221(2): 639-648.
Gates, J. A. and W. B. Wilson (1960). "The toxicity of Gonyaulax monilata Howell to Mugil cephalus." Limnology and Oceanography 5(2): 171-174.
Griffin, B. A., S. R. Adams and R. Y. Tsien (1998). "Specific covalent labeling of recombinant protein molecules inside live cells." Science 281(5374): 269-272.
Grimes, D. J. (2009). "Oceans and human health: risks and remedies from the sea." Environmental Health Perspectives 117(3): A124-A124.
Gromer, S., R. H. Schirmer and K. Becker (1999). "News and views on thioredoxin reductases." Redox Report 4(5): 221-228.
Grzebyk, D., C. Béchemin, C. J. Ward, C. Vérité, G. A. Codd and S. Y. Maestrini (2003). "Effects of salinity and two coastal waters on the growth and toxin content of the dinoflagellate Alexandrium minutum." Journal of Plankton Research 25(10): 1185-1199.
Gusovsky, F. and J. W. Daly (1990). "Maitotoxin: a unique pharmacological tool for research on calcium-dependent mechanisms." Biochemical Pharmacology 39(11): 1633-1639.
Hall, M., A. Mata-Cabana, H. E. Åkerlund, F. J. Florencio, W. P. Schröder, M. Lindahl and T. Kieselbach (2010). "Thioredoxin targets of the plant chloroplast lumen and their implications for plastid function." Proteomics 10(5): 987-1001.
Hallegraeff, G. M. (1993). "A review of harmful algal blooms and their apparent global increase." Phycologia 32(2): 79-99.
Hallegraeff, G. M. (2010). "Ocean climate change, phytoplankton community responses, and harmful algal blooms: a formidable predictive challenge1." Journal of Phycology 46(2): 220-235.
Halliwell, B. (1993). "The role of oxygen radicals in human disease, with particular reference to the vascular system." Pathophysiology of Haemostasis and Thrombosis 23(Suppl. 1): 118-126.

129
Halliwell, B. (1996). "Antioxidants in human health and disease." Annual Review of Nutrition 16(1): 33-50.
Halliwell, B., A. Zentella, E. O. Gomez and D. Kershenobich (1997). "Antioxidants and human disease: a general introduction." Nutrition Reviews 55(1): S44.
Hansen, R. E., H. Østergaard and J. R. Winther (2005). "Increasing the reactivity of an artificial dithiol-disulfide pair through modification of the electrostatic milieu." Biochemistry 44(15): 5899-5906.
Harrison, J. A., N. Caraco and S. P. Seitzinger (2005). "Global patterns and sources of dissolved organic matter export to the coastal zone: Results from a spatially explicit, global model." Global Biogeochemical Cycles 19(4): GB4S04
Hashimoto, M., Y. Hatanaka, J. Yang, J. Dhesi and G. D. Holman (2001). "Synthesis of biotinylated bis (D-glucose) derivatives for glucose transporter photoaffinity labelling." Carbohydrate Research 331(2): 119-127.
Heinemann, S. H., H. Terlau, W. Stühmer, K. Imoto and S. Numa (1992). "Calcium channel characteristics conferred on the sodium channel by single mutations." Nature 356(6368): 441-443.
Heisler, J., P. M. Glibert, J. M. Burkholder, D. M. Anderson, W. Cochlan, W. C. Dennison, Q. Dortch, C. J. Gobler, C. A. Heil and E. Humphries (2008). "Eutrophication and harmful algal blooms: a scientific consensus." Harmful Algae 8(1): 3-13.
Heuck, A. P. and R. A. Wolosiuk (1997). "Fluoresceinthiocarbamyl-insulin: a potential analytical tool for the assay of disulfide bond reduction." Journal of Biochemical and Biophysical Methods 34(3): 213-225.
Hille, B. (2001). Book: Ion channels of excitable membranes
Hirota, K., M. Matsui, S. Iwata, A. Nishiyama, K. Mori and J. Yodoi (1997). "AP-1 transcriptional activity is regulated by a direct association between thioredoxin and Ref-1." Proceedings of the National Academy of Sciences 94(8): 3633-3638.
Hirota, K., M. Murata, Y. Sachi, H. Nakamura, J. Takeuchi, K. Mori and J. Yodoi (1999). "Distinct roles of thioredoxin in the cytoplasm and in the nucleus a two-step mechanism of redox regulation of transcription factor NF-κB." Journal of Biological Chemistry 274(39): 27891-27897.
Hitchcock, G. L., J. W. Fourqurean, J. L. Drake, R. N. Mead and C. A. Heil (2012). "Brevetoxin persistence in sediments and seagrass epiphytes of east Florida coastal waters." Harmful Algae 13: 89-94.
Hitchcock, G. L., G. Kirkpatrick, P. Minnett and V. Palubok (2010). "Net community production and dark community respiration in a Karenia brevis (Davis) bloom in West Florida coastal waters, USA." Harmful Algae 9(4): 351-358.

130
Hoagland, P., D. Anderson, Y. Kaoru and A. White (2002). "The economic effects of harmful algal blooms in the United States: estimates, assessment issues, and information needs." Estuaries 25(4): 819-837.
Hoagland, P., D. Jin, A. Beet, B. Kirkpatrick, A. Reich, S. Ullmann, L. E. Fleming and G. Kirkpatrick (2014). "The human health effects of Florida red tide (FRT) blooms: An expanded analysis." Environment International 68: 144-153.
Hoagland, P., D. Jin, L. Y. Polansky, B. Kirkpatrick, G. Kirkpatrick, L. E. Fleming, A. Reich, S. M. Watkins, S. G. Ullmann and L. C. Backer (2009). "The costs of respiratory illnesses arising from Florida Gulf Coast Karenia brevis blooms." Environmental Health Perspectives 117(8): 1239.
Hogg, N. and B. Kalyanaraman (1998). "Nitric oxide and low-density lipoprotein oxidation." Free Radical Research 28(6): 593-600.
Holmgren, A. (1985). "Thioredoxin." Annual Review of Biochemistry 54(1): 237-271.
Holmgren, A. (1989). "Thioredoxin and glutaredoxin systems." The Journal of Biological Chemistry 264(24): 13963-13966.
Holmgren, A. (1995). "Thioredoxin structure and mechanism: conformational changes on oxidation of the active-site sulfhydryls to a disulfide." Structure 3(3): 239-243.
Holmgren, A., B. Söderberg, H. Eklund and C. Brändén (1975). "Three-dimensional structure of Escherichia coli thioredoxin-S2 to 2.8 A resolution." Proceedings of the National Academy of Sciences 72(6): 2305-2309.
Huang, J. M., C. H. Wu and D. G. Baden (1984). "Depolarizing action of a red-tide dinoflagellate brevetoxin on axonal membranes." Journal of Pharmacology and Experimental Therapeutics 229(2): 615-621.
Ip, C., M. Birringer, E. Block, M. Kotrebai, J. F. Tyson, P. C. Uden and D. J. Lisk (2000). "Chemical speciation influences comparative activity of selenium-enriched garlic and yeast in mammary cancer prevention." Journal of Agricultural and Food Chemistry 48(6): 2062-2070.
Jeglitsch, G., K. Rein, D. G. Baden and D. J. Adams (1998). "Brevetoxin-3 (PbTx-3) and its derivatives modulate single tetrodotoxin-sensitive sodium channels in rat sensory neurons." Journal of Pharmacology and Experimental Therapeutics 284(2): 516-525.
Jeng, M. F., M. T. Reymond, L. L. Tennant, A. Holmgren and H. J. Dyson (1998). "NMR characterization of a single-cysteine mutant of Escherichia coli thioredoxin and a covalent thioredoxin-peptide complex." European Journal of Biochemistry 257(2): 299-308.

131
Jensen, K. S., R. E. Hansen and J. R. Winther (2009). "Kinetic and thermodynamic aspects of cellular thiol-disulfide redox regulation." Antioxidants and Redox Signaling 11(5): 1047-1058.
Johnson, L. V., M. L. Walsh and L. B. Chen (1980). "Localization of mitochondria in living cells with rhodamine 123." Proceedings of the National Academy of Sciences 77(2): 990-994.
Kamata, H. and H. Hirata (1999). "Redox regulation of cellular signalling." Cellular Signalling 11(1): 1-14.
Kanzok, S. M., R. H. Schirmer, I. Türbachova, R. Iozef and K. Becker (2000). "The thioredoxin system of the malaria parasite Plasmodium falciparum glutathione reduction revisited." Journal of Biological Chemistry 275(51): 40180-40186.
Keller, A., A. I. Nesvizhskii, E. Kolker and R. Aebersold (2002). "Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search." Analytical Chemistry 74(20): 5383-5392.
Kirkpatrick, B., J. A. Bean, L. E. Fleming, G. Kirkpatrick, L. Grief, K. Nierenberg, A. Reich, S. Watkins and J. Naar (2010). "Gastrointestinal emergency room admissions and Florida red tide blooms." Harmful Algae 9(1): 82-86.
Kirkpatrick, B., L. E. Fleming, L. C. Backer, J. A. Bean, R. Tamer, G. Kirkpatrick, T. Kane, A. Wanner, D. Dalpra and A. Reich (2006). "Environmental exposures to Florida red tides: effects on emergency room respiratory diagnoses admissions." Harmful Algae 5(5): 526-533.
Kirkpatrick, B., L. E. Fleming, D. Squicciarini, L. C. Backer, R. Clark, W. Abraham, J. Benson, Y. S. Cheng, D. Johnson and R. Pierce (2004). "Literature review of Florida red tide: implications for human health effects." Harmful Algae 3(2): 99-115.
Koide, Y., Y. Urano, S. Kenmoku, H. Kojima and T. Nagano (2007). "Design and synthesis of fluorescent probes for selective detection of highly reactive oxygen species in mitochondria of living cells." Journal of the American Chemical Society 129(34): 10324-10325.
König, J., M. Baier, F. Horling, U. Kahmann, G. Harris, P. Schürmann and K.-J. Dietz (2002). "The plant-specific function of 2-Cys peroxiredoxin-mediated detoxification of peroxides in the redox-hierarchy of photosynthetic electron flux." Proceedings of the National Academy of Sciences 99(8): 5738-5743.
Konoki, K., M. Hashimoto, K. Honda, K. Tachibana, R. Tamate, F. Hasegawa, T. Oishi and M. Murata (2009). "Maitotoxin-photoactive probe binds to membrane proteins in blood cells." Heterocycles 79(1): 1007-1017.
Kontos, H. A., E. P. Wei, E. F. Ellis, L. W. Jenkins, J. T. Povlishock, G. T. Rowe and M. L. Hess (1985). "Appearance of superoxide anion radical in cerebral extracellular space during increased prostaglandin synthesis in cats." Circulation Research 57(1): 142-151.

132
Krause, G. and E. Weis (1991). "Chlorophyll fluorescence and photosynthesis: the basics." Annual Review of Plant Biology 42(1): 313-349.
Krauth-Siegel, R. and G. Coombs (1999). "Enzymes of parasite thiol metabolism as drug targets." Parasitology Today 15(10): 404-409.
Kubanek, J., T. W. Snell and C. Pirkle (2007). "Chemical defense of the red tide dinoflagellate Karenia brevis against rotifer grazing." Limnology and Oceanography 52(3): 1026-1035.
Kumar-Roiné, S., M. Matsui, S. Pauillac and D. Laurent (2011). "Ciguatera fish poisoning and other seafood intoxication syndromes: A revisit and a review of the existing treatments employed in ciguatera fish poisoning." The South Pacific Journal of Natural and Applied Sciences 28(1): 1-26.
Kumar, S., M. Bjornstedt and A. Holmgren (1992). "Selenite is a substrate for calf thymus thioredoxin reductase and thioredoxin and elicits a large non-stoichiometric oxidation of NADPH in the presence of oxygen." European Journal of Biochemistry 207(2): 435-439.
Landino, L. M., T. E. Skreslet and J. A. Alston (2004). "Cysteine oxidation of tau and microtubule-associated protein-2 by peroxynitrite modulation of microtubule assembly kinetics by the thioredoxin reductase system." Journal of Biological Chemistry 279(33): 35101-35105.
Laurent, T. C., E. C. Moore and P. Reichard (1964). "Enzymatic synthesis of deoxyribonucleotides IV. Isolation and characterization of thioredoxin, the hydrogen donor from Escherichia coli B." Journal of Biological Chemistry 239(10): 3436-3444.
Lee, M. S., G. Qin, K. Nakanishi and M. G. Zagorski (1989). "Biosynthetic studies of brevetoxins, potent neurotoxins produced by the dinoflagellate Gymnodinium breve." Journal of the American Chemical Society 111(16): 6234-6241.
Lee, S.-R., S. Bar-Noy, J. Kwon, R. L. Levine, T. C. Stadtman and S. G. Rhee (2000). "Mammalian thioredoxin reductase: oxidation of the C-terminal cysteine/selenocysteine active site forms a thioselenide, and replacement of selenium with sulfur markedly reduces catalytic activity." Proceedings of the National Academy of Sciences of the United States of America 97(6): 2521-2526.
Lehane, L. and R. J. Lewis (2000). "Ciguatera: recent advances but the risk remains." International Journal of Food Microbiology 61(2): 91-125.
Lennon, B. W. and C. H. Williams (1997). "Reductive half-reaction of thioredoxin reductase from Escherichia coli." Biochemistry 36(31): 9464-9477.
Levine, D. (1995). "Ciguatera: current concepts." The Journal of the American Osteopathic Association 95(3): 193-198.
Lillig, C. H. and A. Holmgren (2007). "Thioredoxin and related molecules-from biology to health and disease." Antioxidants and Redox Signaling 9(1): 25-47.

133
Lin, M. and O. Alfi (1976). "Detection of sister chromatid exchanges by 4′-6-diamidino-2-phenylindole fluorescence." Chromosoma 57(3): 219-225.
Lin, M., D. Comings and O. Alfi (1977). "Optical studies of the interaction of 4′-6-diamidino-2-phenylindole with DNA and metaphase chromosomes." Chromosoma 60(1): 15-25.
Lin, Y.-Y., M. Risk, S. M. Ray, D. Van Engen, J. Clardy, J. Golik, J. C. James and K. Nakanishi (1981). "Isolation and structure of brevetoxin B from the" red tide" dinoflagellate Ptychodiscus brevis (Gymnodinium breve)." Journal of the American Chemical Society 103(22): 6773-6775.
Lincoln, D. T., E. E. Ali, K. F. Tonissen and F. M. Clarke (2002). "The thioredoxin-thioredoxin reductase system: over-expression in human cancer." Anticancer Research 23(3B): 2425-2433.
Lothrop, A. P., E. L. Ruggles and R. J. Hondal (2009). "No selenium required: reactions catalyzed by mammalian thioredoxin reductase that are independent of a selenocysteine residue." Biochemistry 48(26): 6213-6223.
Lowther, W. T., N. Brot, H. Weissbach, J. F. Honek and B. W. Matthews (2000). "Thiol–disulfide exchange is involved in the catalytic mechanism of peptide methionine sulfoxide reductase." Proceedings of the National Academy of Sciences 97(12): 6463-6468.
Lu, J., E.-H. Chew and A. Holmgren (2007). "Targeting thioredoxin reductase is a basis for cancer therapy by arsenic trioxide." Proceedings of the National Academy of Sciences 104(30): 12288-12293.
Lu, J. and A. Holmgren (2014). "The thioredoxin antioxidant system." Free Radical Biology and Medicine 66: 75-87.
Luthman, M. and A. Holmgren (1982). "Rat liver thioredoxin and thioredoxin reductase: purification and characterization." Biochemistry 21(26): 6628-6633.
Magaña, H. A., C. Contreras and T. A. Villareal (2003). "A historical assessment of Karenia brevis in the western Gulf of Mexico." Harmful Algae 2(3): 163-171.
Marnett, L. J. (2000). "Oxyradicals and DNA damage." Carcinogenesis 21(3): 361-370.
Martin, D. F. and B. B. Martin (1976). "Red tide, red terror. Effects of red tide and related toxins." Journal of Chemical Education 53(10): 614.
Martin, H. and M. Dean (1991). "Identification of a thioredoxin—related protein associated with plasma membranes." Biochemical and Biophysical Research Communications 175(1): 123-128.
Martin, J. L. (1995). "Thioredoxin —a fold for all reasons." Structure 3(3): 245-250.

134
MatÉs, J. M., C. Pérez-Gómez and I. N. De Castro (1999). "Antioxidant enzymes and human diseases." Clinical Biochemistry 32(8): 595-603.
Matile, S., N. Berova and K. Nakanishi (1996). "Exciton coupled circular dichroic studies of self-assembled brevetoxin-porphyrin conjugates in lipid bilayers and polar solvents." Chemistry and Biology 3(5): 379-392.
Matsuo, G., K. Kawamura, N. Hori, H. Matsukura and T. Nakata (2004). "Total synthesis of brevetoxin-B." Journal of the American Chemical Society 126(44): 14374-14376.
Mattei, P., A. Virdis, L. Ghiadoni, S. Taddei and A. Salvetti (1996). "Endothelial function in hypertension." Journal of Nephrology 10(4): 192-197.
Messens, J. and S. Silver (2006). "Arsenate Reduction: thiol cascade chemistry with convergent evolution." Journal of Molecular Biology 362(1): 1-17.
Milian, A., K. Nierenberg, L. E. Fleming, J. A. Bean, A. Wanner, A. Reich, L. C. Backer, D. Jayroe and B. Kirkpatrick (2007). "Reported respiratory symptom intensity in asthmatics during exposure to aerosolized Florida red tide toxins." Journal of Asthma 44(7): 583-587.
Miranda-Vizuete, A., A. E. Damdimopoulos, J.-Å. Gustafsson and G. Spyrou (1997). "Cloning, expression, and characterization of a novel Escherichia coli thioredoxin." Journal of Biological Chemistry 272(49): 30841-30847.
Moisan, T. A., M. Olaizola and B. G. Mitchell (1998). "Xanthophyll cycling in Phaeocystis antarctica: changes in cellular fluorescence." Marine Ecology Progress Series 169: 113-121.
Montano, S. J., J. Lu, T. N. Gustafsson and A. Holmgren (2014). "Activity assays of mammalian thioredoxin and thioredoxin reductase: fluorescent disulfide substrates, mechanisms, and use with tissue samples." Analytical Biochemistry 449: 139-146.
Moore, E. C., P. Reichard and L. Thelander (1964). "Enzymatic synthesis of deoxyribonucleotides V. Purification and properties of thioredoxin reductase from Escherichia coli B." Journal of Biological Chemistry 239(10): 3445-3452.
Morgenstern, R. (2005). Microsomal glutathione transferase 1. Methods in Enzymology. S. Helmut and P. Lester, Academic Press. 401: 136-146.
Mugesh, G. and H. B. Singh (2000). "Synthetic organoselenium compounds as antioxidants: glutathione peroxidase activity." Chemical Society Reviews 29(5): 347-357.
Müller, P., X.-P. Li and K. K. Niyogi (2001). "Non-photochemical quenching. A response to excess light energy." Plant Physiology 125(4): 1558-1566.

135
Murata, M., M. Kumagai, J. S. Lee and T. Yasumoto (1987). "Isolation and structure of yessotoxin, a novel polyether compound implicated in diarrhetic shellfish poisoning." Tetrahedron Letters 28(47): 5869-5872.
Murata, M., A. M. Legrand, Y. Ishibashi, M. Fukui and T. Yasumoto (1990). "Structures and configurations of ciguatoxin from the moray eel Gymnothorax javanicus and its likely precursor from the dinoflagellate Gambierdiscus toxicus." Journal of the American Chemical Society 112(11): 4380-4386.
Murata, M., A. M. Legrand, Y. Ishibashi and T. Yasumoto (1989). "Structures of ciguatoxin and its congener." Journal of the American Chemical Society 111(24): 8929-8931.
Murrell, R. N. and J. E. Gibson (2009). "Brevetoxins 2, 3, 6, and 9 show variability in potency and cause significant induction of DNA damage and apoptosis in Jurkat E6-1 cells." Archives of Toxicology 83(11): 1009-1019.
Mustacich, D. and G. Powis (2000). "Thioredoxin reductase." Biochemical Journal 346(1): 1-8.
Nair, D. P., M. Podgorski, S. Chatani, T. Gong, W. Xi, C. R. Fenoli and C. N. Bowman (2013). "The Thiol-Michael Addition Click Reaction: A Powerful and Widely Used Tool in Materials Chemistry." Chemistry of Materials 26(1): 724-744.
Nakamura, H., M. Matsuda, K. Furuke, Y. Kitaoka, S. Iwata, K. Toda, T. Inamoto, Y. Yamaoka, K. Ozawa and J. Yodoi (1994). "Adult T cell leukemia-derived factor/human thioredoxin protects endothelial F-2 cell injury caused by activated neutrophils or hydrogen peroxide." Immunology Letters 42(1): 75-80.
Nakanishi, K. (1985). "The chemistry of brevetoxins: A review." Toxicon 23(3): 473-479.
Nesvizhskii, A. I., A. Keller, E. Kolker and R. Aebersold (2003). "A statistical model for identifying proteins by tandem mass spectrometry." Analytical Chemistry 75(17): 4646-4658.
Netto, L. E. S., M. A. de Oliveira, C. A. Tairum and J. F. da Silva Neto (2016). "Conferring specificity in redox pathways by enzymatic thiol/disulfide exchange reactions." Free Radical Research 50(2): 206-245.
Neyens, E. and J. Baeyens (2003). "A review of classic Fenton’s peroxidation as an advanced oxidation technique." Journal of Hazardous Materials 98(1): 33-50.
Nicolaou, K., Z. Yang, G.-q. Shi, J. L. Gunzner, K. A. Agrios and P. Gärtner (1998). "Total synthesis of brevetoxin A." Nature 392(6673): 264-269.
Nicolaou, K. C. (1996). "The total synthesis of brevetoxin B: a twelve-year odyssey in organic synthesis." Angewandte Chemie International Edition in English 35(6): 588-607.

136
Nicolaou, K. C., C. K. Hwang, M. E. Duggan, D. A. Nugiel, Y. Abe, K. B. Reddy, S. A. DeFrees, D. R. Reddy and R. A. Awartani (1995). "Total synthesis of brevetoxin B. 1. First gneration atrategies and new approaches to oxepane systems." Journal of the American Chemical Society 117(41): 10227-10238.
Nicolaou, K. C., F. P. J. T. Rutjes, E. A. Theodorakis, J. Tiebes, M. Sato and E. Untersteller (1995). "Total synthesis of brevetoxin B. 3. Final strategy and completion." Journal of the American Chemical Society 117(41): 10252-10263.
Nicolaou, K. C., E. A. Theodorakis, F. P. J. T. Rutjes, M. Sato, J. Tiebes, X. Y. Xiao, C. K. Hwang, M. E. Duggan and Z. Yang (1995). "Total synthesis of brevetoxin B. 2. Second generation strategies and construction of the dioxepane region [DEFG]." Journal of the American Chemical Society 117(41): 10239-10251.
Nikkanen, L. and E. Rintamäki (2014). "Thioredoxin-dependent regulatory networks in chloroplasts under fluctuating light conditions." Philosophical Transactions of the Royal Society B 369(1640): 20130224.
Nordberg, J. and E. S. J. Arnér (2001). "Reactive oxygen species, antioxidants, and the mammalian thioredoxin system1." Free Radical Biology and Medicine 31(11): 1287-1312.
O'Shea, T. J., G. B. Rathbun, R. K. Bonde, C. D. Buergelt and D. K. Odell (1991). "An epizootic of Florida manatees associated with a dinoflagellate bloom." Marine Mammal Science 7(2): 165-179.
Østergaard, H., A. Henriksen, F. G. Hansen and J. R. Winther (2001). "Shedding light on disulfide bond formation: engineering a redox switch in green fluorescent protein." The European Molecular Biology Organization journal 20(21): 5853-5862. Patel, R. P., D. Moellering, J. Murphy-Ullrich, H. Jo, J. S. Beckman and V. M. Darley-Usmar (2000). "Cell signaling by reactive nitrogen and oxygen species in atherosclerosis." Free Radical Biology and Medicine 28(12): 1780-1794.
Pearn, J. (2001). "Neurology of ciguatera." Journal of Neurology, Neurosurgery and Psychiatry 70(1): 4-8.
Pierce, R., M. Henry, P. Blum, S. Hamel, B. Kirkpatrick, Y. Cheng, Y. Zhou, C. Irvin, J. Naar and A. Weidner (2005). "Brevetoxin composition in water and marine aerosol along a Florida beach: Assessing potential human exposure to marine biotoxins." Harmful Algae 4(6): 965-972.
Pierce, R., M. Henry, P. Blum, J. Lyons, Y. Cheng, D. Yazzie and Y. Zhou (2003). "Brevetoxin concentrations in marine aerosol: human exposure levels during a Karenia brevis harmful algal bloom." Bulletin of Environmental Contamination and Toxicology 70(1): 161-165.
Poli, M., T. J. Mende and D. G. Baden (1986). "Brevetoxins, unique activators of voltage-sensitive sodium channels, bind to specific sites in rat brain synaptosomes." Molecular Pharmacology 30(2): 129-135.

137
Powis, G., D. Mustacich and A. Coon (2000). "The role of the redox protein thioredoxin in cell growth and cancer." Free Radical Biology and Medicine 29(3): 312-322.
Rahman, I. and I. Adcock (2006). "Oxidative stress and redox regulation of lung inflammation in COPD." European Respiratory Journal 28(1): 219-242.
Raj, A., P. Van Den Bogaard, S. A. Rifkin, A. Van Oudenaarden and S. Tyagi (2008). "Imaging individual mRNA molecules using multiple singly labeled probes." Nature Methods 5(10): 877-879.
Re, R., N. Pellegrini, A. Proteggente, A. Pannala, M. Yang and C. Rice-Evans (1999). "Antioxidant activity applying an improved ABTS radical cation decolorization assay." Free Radical Biology and Medicine 26(9–10): 1231-1237.
Rein, K. S., D. G. Baden and R. E. Gawley (1994). "Conformational analysis of the sodium channel modulator, brevetoxin A, comparison with brevetoxin B conformations, and a hypothesis about the common pharmacophore of the" site 5" toxins." The Journal of Organic Chemistry 59(8): 2101-2106.
Rein, K. S. and J. Borrone (1999). "Polyketides from dinoflagellates: origins, pharmacology and biosynthesis." Comparative Biochemistry and Physiology Part B: Biochemistry and Molecular Biology 124(2): 117-131.
Rein, K. S., B. Lynn, R. E. Gawley and D. G. Baden (1994). "Brevetoxin B: chemical modifications, synaptosome binding, toxicity, and an unexpected conformational effect." The Journal of Organic Chemistry 59(8): 2107-2113.
Rhee, S. G. (1999). "Redox signaling: hydrogen peroxide as intracellular messenger." Experimental and Molecular Medicine 31(2): 53-59.
Riener, C. K., G. Kada and H. J. Gruber (2002). "Quick measurement of protein sulfhydryls with Ellman's reagent and with 4, 4′-dithiodipyridine." Analytical and Bioanalytical Chemistry 373(4-5): 266-276.
Rogers, J. C., Y. Qu, T. N. Tanada, T. Scheuer and W. A. Catterall (1996). "Molecular determinants of high affinity binding of α-scorpion toxin and sea anemone toxin in the S3-S4 extracellular loop in domain IV of the Na+ channel α subunit." Journal of Biological Chemistry 271(27): 15950-15962.
Roos, G., N. Foloppe, K. Van Laer, L. Wyns, L. Nilsson, P. Geerlings and J. Messens (2009). "How thioredoxin dissociates its mixed disulfide." PLOS Computational Biology 5(8): e1000461.
Ross, C., R. Ritson-Williams, R. Pierce, J. B. Bullington, M. Henry and V. J. Paul (2010). "Effects of the Florida red tide dinoflagellate, Karenia brevis, on oxidative stress and metamorphosis of larvae of the coral Porites astreoides." Harmful Algae 9(2): 173-179.

138
Rubbo, H., R. Radi, D. Anselmi, M. Kirk, S. Barnes, J. Butler, J. P. Eiserich and B. A. Freeman (2000). "Nitric oxide reaction with lipid peroxyl radicals spares α-tocopherol during lipid peroxidation greater oxidant protection from pair nitric oxide/α-tocopherol than α- tocopherol/ascorbate." Journal of Biological Chemistry 275(15): 10812-10818.
Satake, M., A. Campbell, R. M. Van Wagoner, A. J. Bourdelais, H. Jacocks, D. G. Baden and J. L. Wright (2009). "Brevisin: an aberrant polycyclic ether structure from the dinoflagellate Karenia brevis and its implications for polyether assembly." The Journal of Organic Chemistry 74(3): 989-994.
Satake, M., L. MacKenzie and T. Yasumoto (1997). "Identification of Protoceratium reticulatum as the biogenetic origin of yessotoxin." Natural Toxins 5(4): 164-167.
Sayer, A., Q. Hu, A. J. Bourdelais, D. G. Baden and J. E. Gibson (2006). "The inhibition of CHO-K1-BH4 cell proliferation and induction of chromosomal aberrations by brevetoxins in vitro." Food and Chemical Toxicology 44(7): 1082-1091.
Schettini, G., K. Koike, A. Judd, M. Cronin, T. Yasumoto and R. MacLeod (1984). "Maitotoxin stimulates hormonal release and calcium flux in rat anterior pituitary cells in vitro." American Journal of Physiology-Endocrinology and Metabolism 247(4): E520-E525.
Schweizer, D. (1976). "DAPI fluorescence of plant chromosomes prestained with actinomycin D." Experimental Cell Research 102(2): 408-413.
Schweizer, D. (1976). "Reverse fluorescent chromosome banding with chromomycin and DAPI." Chromosoma 58(4): 307-324.
Seitzinger, S. P., J. A. Harrison, E. Dumont, A. H. W. Beusen and A. F. Bouwman (2005). "Sources and delivery of carbon, nitrogen, and phosphorus to the coastal zone: An overview of Global Nutrient Export from Watersheds (NEWS) models and their application." Global Biogeochemical Cycles 19(4): doi:10.1029/2005GB002606
Setterdahl, A. T., P. T. Chivers, M. Hirasawa, S. D. Lemaire, E. Keryer, M. Miginiac-Maslow, S.-K. Kim, J. Mason, J.-P. Jacquot and C. C. Longbine (2003). "Effect of pH on the oxidation-reduction properties of thioredoxins." Biochemistry 42(50): 14877-14884.
Seyedrezazadeh, E., A. Ostadrahimi, S. Mahboob, Y. Assadi, J. Ghaemmagami and M. Pourmogaddam (2008). "Effect of vitamin E and selenium supplementation on oxidative stress status in pulmonary tuberculosis patients." Respirology 13(2): 294-298.
Shaked, Z. e., R. P. Szajewski and G. M. Whitesides (1980). "Rates of thiol-disulfide interchange reactions involving proteins and kinetic measurements of thiol pKa values." Biochemistry 19(18): 4156-4166.

139
Shimizu, Y., H. Bando, H.-N. Chou, G. Van Duyne and J. C. Clardy (1986). "Absolute configuration of brevetoxins." Journal of the Chemical Society, Chemical Communications (22): 1656-1658.
Sikorski, Z. E. (2006). Book :Chemical and functional properties of food components
Simon, J. and D. Dauer (1972). "A quantitative evaluation of red-tide induced mass mortalities of benthic invertebrates in Tampa Bay, Florida." Environmental Letters 3(4): 229-234.
Singh, R., W. A. Blattler and A. R. Collinson (1993). "An amplified assay for thiols based on reactivation of papain." Analytical Biochemistry 213(1): 49-56.
Sinigalliano, C. D., J. Winshell, M. A. Guerrero, G. Scorzetti, J. W. Fell, R. W. Eaton, L. Brand and K. S. Rein (2009). "Viable cell sorting of dinoflagellates by multiparametric flow cytometry." Phycologia 48(4): 249-257.
Smayda, T. J. (2007). "Reflections on the ballast water dispersal—harmful algal bloom paradigm." Harmful Algae 6(4): 601-622.
Snider, G. W., C. M. Dustin, E. L. Ruggles and R. J. Hondal (2014). "A mechanistic investigation of the C-terminal redox motif of thioredoxin reductase from Plasmodium falciparum." Biochemistry 53(3): 601-609.
Stadtman, E. R. (1992). "Protein oxidation and aging." Science 257(5074): 1220-1224.
Stadtman, E. R. and B. S. Berlett (1997). "Reactive oxygen-mediated protein oxidation in aging and disease." Chemical Research in Toxicology 10(5): 485-494.
Steinberg, D. (1997). "Low density lipoprotein oxidation and its pathobiological significance." Journal of Biological Chemistry 272(34): 20963-20966.
Stephen, J. and J. G. Kettle (1997). "Synthesis of brevetoxin sub-units by sequential ring-closing metathesis and hydroboration." Tetrahedron Letters 38(1): 123-126.
Stewart, E. J., F. Åslund and J. Beckwith (1998). "Disulfide bond formation in the Escherichia coli cytoplasm: an in vivo role reversal for the thioredoxins." The EMBO journal 17(19): 5543-5550.
Stuart, A. M. and D. G. Baden (1988). "Florida red tide brevetoxins and binding in fish brain synaptosomes." Aquatic Toxicology 13(4): 271-279.
Stumpf, R. P., M. E. Culver, P. A. Tester, M. Tomlinson, G. J. Kirkpatrick, B. A. Pederson, E. Truby, V. Ransibrahmanakul and M. Soracco (2003). "Monitoring Karenia brevis blooms in the Gulf of Mexico using satellite ocean color imagery and other data." Harmful Algae 2(2): 147-160.
Sudarsanam, S., G. D. Virca, C. March and S. Srinivasan (1992). "An approach to computer-aided inhibitor design: Application to cathepsin L." Journal of Computer-Aided Molecular Design 6(3): 223-233.

140
Sugimoto, T., T. Fujii, Y. Idutu, S. Yamamura and M. Ueda (2004). "Detection of potential membrane receptor proteins concerning circadian rhythmic leaf movement of legumes using novel photoaffinity probe compounds." Tetrahedron Letters 45(2): 335-338.
Sun, P., C. Leeson, X. Zhi, F. Leng, R. H. Pierce, M. S. Henry and K. S. Rein (2016). "Characterization of an epoxide hydrolase from the Florida red tide dinoflagellate, Karenia brevis." Phytochemistry 122: 11-21.
Sunda, W. G., C. Burleson, D. R. Hardison, J. S. Morey, Z. Wang, J. Wolny, A. A. Corcoran, L. J. Flewelling and F. M. Van Dolah (2013). "Osmotic stress does not trigger brevetoxin production in the dinoflagellate Karenia brevis." Proceedings of the National Academy of Sciences of the United States of America 110(25): 10223-10228.
Sundaresan, M., Z.-X. Yu, V. J. Ferrans, K. Irani and T. Finkel (1995). "Requirement for generation of H2O2 for platelet-derived growth factor signal transduction." Science 270(5234): 296.
Suzuki, T., T. Matsuzaki, H. Hagiwara, T. Aoki and K. Takata (2007). "Recent advances in fluorescent labeling techniques for fluorescence microscopy." Acta Histochemica et Cytochemica 40(5): 131-137.
Taglialatela, M., S. Amoroso, T. Yasumoto, G. Di Renzo and L. Annunziato (1986). "Maitotoxin and BAY-K-8644: two putative calcium channel activators with different effects on endogenous dopamine release from tuberoinfundibular neurons." Brain Research 381(2): 356-358.
Takahashi, W., H. Kawamura, T. Omura and K. Furuya (2009). "Detecting red tides in the eastern Seto inland sea with satellite ocean color imagery." Journal of Oceanography 65(5): 647-656.
Taniguchi, Y., Y. Taniguchi-Ueda, K. Mori and J. Yodoi (1996). "A novel promoter sequence is involved in the oxidative stress-induced expression of the adult T-cell leukemia-derived factor (ADF)/human thioredoxin (Trx) gene." Nucleic ACids Research 24(14): 2746-2752.
Tejedor, F. J. and W. A. Catterall (1988). "Site of covalent attachment of alpha-scorpion toxin derivatives in domain I of the sodium channel alpha subunit." Proceedings of the National Academy of Sciences of the United States of America 85(22): 8742-8746.
Tester, P. A. and K. A. Steidinger (1997). "Gymnodinium breve red tide blooms: Initiation, transport, and consequences of surface circulation." Limnology and Oceanography 42(5part2): 1039-1051.
Thyng, K. M., R. D. Hetland, M. T. Ogle, X. Zhang, F. Chen and L. Campbell (2013). "Origins of Karenia brevis harmful algal blooms along the Texas coast." Limnology and Oceanography: Fluids and Environments 3(1): 269-278.

141
Toppo, S., S. Vanin, V. Bosello and S. C. Tosatto (2008). "Evolutionary and structural insights into the multifaceted glutathione peroxidase (Gpx) superfamily." Antioxidants and Redox Signaling 10(9): 1501-1514.
Trainer, V. L. and D. G. Baden (1991). "An enzyme immunoassay for the detection of Florida red tide brevetoxins." Toxicon 29(11): 1387-1394.
Trainer, V. L. and D. G. Baden (1999). "High affinity binding of red tide neurotoxins to marine mammal brain." Aquatic Toxicology 46(2): 139-148.
Trainer, V. L., D. G. Baden and W. A. Catterall (1994). "Identification of peptide components of the brevetoxin receptor site of rat brain sodium channels." Journal of Biological Chemistry 269(31): 19904-19909.
Trainer, V. L., W. J. Thomsen, W. A. Catterall and D. G. Baden (1991). "Photoaffinity labeling of the brevetoxin receptor on sodium channels in rat brain synaptosomes." Molecular Pharmacology 40(6): 988-994.
Twiner, M. J., M.-Y. Bottein Dechraoui, Z. Wang, C. M. Mikulski, M. S. Henry, R. H. Pierce and G. J. Doucette (2007). "Extraction and analysis of lipophilic brevetoxins from the red tide dinoflagellate Karenia brevis." Analytical Biochemistry 369(1): 128-135.
Ujihara, S., T. Oishi, K. Torikai, K. Konoki, N. Matsumori, M. Murata, Y. Oshima and S. Aimoto (2008). "Interaction of ladder-shaped polyethers with transmembrane α-helix of glycophorin A as evidenced by saturation transfer difference NMR and surface plasmon resonance." Bioorganic and Medicinal Chemistry Letters 18(23): 6115-6118.
Urig, S. and K. Becker (2006). "On the potential of thioredoxin reductase inhibitors for cancer therapy." Seminars in cancer biology 16(6):452-65
Valko, M., C. Rhodes, J. Moncol, M. Izakovic and M. Mazur (2006). "Free radicals, metals and antioxidants in oxidative stress-induced cancer." Chemico Biological Interactions 160(1): 1-40.
Van Bladeren, P. J. (2000). "Glutathione conjugation as a bioactivation reaction." Chemico-biological Interactions 129(1): 61-76.
Van Deventer, M., K. Atwood, A. Vargo Gabriel, J. Flewelling Leanne, H. Landsberg Jan, P. Naar Jerome and D. Stanek (2012). "Karenia brevis red tides and brevetoxin-contaminated fish: a high risk factor for Florida’s scavenging shorebirds? "Botanica Marina. 55(1): 31.
Vargo, G. A., C. A. Heil, D. N. Ault, M. B. Neely, S. Murasko, J. Havens, K. M. Lester, L. K. Dixon, R. Merkt and J. Walsh (2002). "Four Karenia brevis blooms: a comparative analysis." Harmful Algae: 14-16.

142
Vijayan, A. K. and S. A. Somayajula (2014). "Effect of accessory pigment composition on the absorption characteristics of dinoflagellate bloom in a coastal embayment." Oceanologia 56(1): 107-124.
Walsh, C., S. Leggett, K. Strohbehn, R. Pierce and J. Sleasman (2008). "Effects of in vitro brevetoxin exposure on apoptosis and cellular metabolism in a leukemic T cell line (Jurkat)." Marine Drugs 6(2): 291-307.
Walsh, C. J., M. Butawan, J. Yordy, R. Ball, L. Flewelling, M. de Wit and R. K. Bonde (2015). "Sublethal red tide toxin exposure in free-ranging manatees (Trichechus manatus) affects the immune system through reduced lymphocyte proliferation responses, inflammation, and oxidative stress." Aquatic Toxicology 161: 73-84.
Walsh, C. J., S. R. Leggett, B. J. Carter and C. Colle (2010). "Effects of brevetoxin exposure on the immune system of loggerhead sea turtles." Aquatic Toxicology 97(4): 293-303.
Walsh, C. J., S. R. Leggett, M. S. Henry, P. C. Blum, S. Osborn and R. H. Pierce (2009). "Cellular metabolism of brevetoxin (PbTx-2) by a monocyte cell line (U-937)." Toxicon 53(1): 135-145.
Walsh, C. J., J. E. Stuckey, H. Cox, B. Smith, C. Funke, J. Stott, C. Colle, J. Gaspard and C. A. Manire (2007). "Production of nitric oxide by peripheral blood mononuclear cells from the Florida manatee, Trichechus manatus latirostris." Veterinary Immunology and Immunopathology 118(3–4): 199-209.
Walsh, J., R. Weisberg, J. Lenes, F. Chen, D. Dieterle, L. Zheng, K. Carder, G. Vargo, J. Havens and E. Peebles (2009). "Isotopic evidence for dead fish maintenance of Florida red tides, with implications for coastal fisheries over both source regions of the West Florida shelf and within downstream waters of the South Atlantic Bight." Progress in Oceanography 80(1): 51-73.
Watkins, S. M., A. Reich, L. E. Fleming and R. Hammond (2008). "Neurotoxic shellfish poisoning." Marine Drugs 6(3): 431-455.
Weber, P. J. and A. G. Beck-Sickinger (1997). "Comparison of the photochemical behavior of four different photoactivatable probes." The Journal of Peptide Research 49(5): 375-383.
Williams, C. (1995). "Mechanism and structure of thioredoxin reductase from Escherichia coli." The Federation of American Societies for Experimental Biology journal 9(13): 1267-1276.
Williams, C. H. (2000). "Thioredoxin–thioredoxin reductase–a system that has come of age." European Journal of Biochemistry 267(20): 6101-6101.
Williams Jr, C. H. (1992). "Lipoamide dehydrogenase, glutathione reductase, thioredoxin reductase, and mercuric ion reductase—a family of flavoenzyme transhydrogenases." Chemistry and Biochemistry of Flavoenzymes 3: 121-211.

143
Williamson, D. and D. Fennell (1975). "The use of fluorescent DNA-binding agent for detecting and separating yeast mitochondrial DNA." Methods in Cell Biology 12: 335-351.
Woo, S. P. S., W. Liu, D. W. T. Au, D. M. Anderson and R. S. S. Wu (2006). "Antioxidant responses and lipid peroxidation in gills and erythrocytes of fish (Rhabdosarga sarba) upon exposure to Chattonella marina and hydrogen peroxide: implications on the cause of fish kills." Journal of Experimental Marine Biology and Ecology 336(2): 230-241.
Yla‐Herttuala, S. (1999). "Oxidized LDL and atherogenesisa." Annals of the New York Academy of Sciences 874(1): 134-137.
Yost, F. J. and I. Fridovich (1973). "An iron-containing superoxide dismutase from Escherichia coli." Journal of Biological Chemistry 248(14): 4905-4908.
Young, F. M., C. Thomson, J. S. Metcalf, J. M. Lucocq and G. A. Codd (2005). "Immunogold localisation of microcystins in cryosectioned cells of Microcystis." Journal of Structural Biology 151(2): 208-214.
Zhang, B., C. Ge, J. Yao, Y. Liu, H. Xie and J. Fang (2015). "Selective selenol fluorescent probes: design, synthesis, structural determinants, and biological applications." Journal of the American Chemical Society 137(2): 757-769.
Zhong, L., E. S. Arnér, J. Ljung, F. Åslund and A. Holmgren (1998). "Rat and calf thioredoxin reductase are homologous to glutathione reductase with a carboxyl-terminal elongation containing a conserved catalytically active penultimate selenocysteine residue." Journal of Biological Chemistry 273(15): 8581-8591.
Zhong, L. and A. Holmgren (2000). "Essential role of selenium in the catalytic activities of mammalian thioredoxin reductase revealed by characterization of recombinant enzymes with selenocysteine mutations." Journal of Biological Chemistry 275(24): 18121-18128.
Zhou, J. and L. Fritz (1994). "Okadaic acid antibody localizes to chloroplasts in the DSP-toxin-producing dinoflagellates Prorocentrum lima and Prorocentrum maculosum." Phycologia 33(6): 455-461.

144
VITA
WEI CHEN
EDUCATION
2007 - 2011 B.S., Chemistry
Wuhan institute of technology, Wuhan, China
2011 - 2016 Doctoral Candidate
Florida International University, Miami, Florida, USA
PUBLICATIONS and PRESENTATIONS
*Cassell, R. T., *W. Chen, S. Thomas, L. Liu and K. S. Rein “Brevetoxin, the dinoflagellate neurotoxin, localizes to thylakoid membranes and interacts with the light-harvesting complex II (LHCII) of photosystem II.” ChemBioChem Volume 16, Issue 7, pages 1060–1067, May 4, 2015 (* represents the equal contribution)
W. Chen, F. Rodriguez del Rey, T. Anupama, C. E. Zavala and K. S. Rein “Brevetoxin-2, inhibits the C-terminal redox center of mammalian thioredoxin reductase-1: Implications for a new mechanism of toxicity.” Chemical Research in Toxicology (in- progress)
W. Chen, F. Rodriguez del Rey, R. Colon, M. Durham and K. S. Rein “Brevetoxin-2, is an Inhibitor of thioredoxin reductase: lmplcations for endogenous function and toxicity” Harmful Algae (in- progress)
W. Chen and K. S. Rein “The inhibition of the mammalian thioredoxin / thioredoxin reductase system by brevetoxins” 2016 Florida Annual Meeting and Exposition of the Florida Section of the American Chemical Society, Tampa, FL 05/06-05/08 (Oral Presentation)
W. Chen, F. Rodriguez and K. S. Rein “Characterizing the redox status of high and low toxin producing K. brevis strains” 2016 Florida Annual Meeting and Exposition of the Florida Section of the American Chemical Society, Tampa, FL 05/06-05/08 (Poster Presentation) W. Chen and K. S. Rein “The interaction between the dinoflagellate neurotoxin and the thioredoxin system” 2015 8th Symposia of Harmful Algae in the U.S., Long beach, CA, 11/15-11/19. (Oral Presentation) W. Chen and K. S. Rein “The interaction between the dinoflagellate neurotoxin and the thioredoxin system” 2015 8th Symposia of Harmful Algae in the U.S., Long beach, CA, 11/15-11/19. (Poster Presentation) W. Chen and K. S. Rein “The interaction between the dinoflagellate neurotoxin and its native receptors” 2015 Florida Annual Meeting and Exposition of the Florida Section of the American Chemical Society, Tampa, FL, 05/08-05/10. (Oral Presentation)
W. Chen, Cassell, R. T. and K. S. Rein “Localiaztion of brevetoxins and photoaffinity labeling of target proteins” 2013 7th Symposia of Harmful Algae in the U.S. Sarasota, FL, 10/27-10/31 (Poster Presentation)

145
T. Anupama, W. Chen, S. Rolle, Y. Liu and K. S. Rein “The effect of trolox on brevetoxicosis in human lymphoblast cells” 2016 XII congress of the pan American section of the international society of toxinology, Miami beach, FL, 09/18-09/23 (Poster presentation)