characterization of chlorobenzoate degraders isolated from polychlorinated biphenyl-contaminated...
TRANSCRIPT

Journal of Applied Microbiology 1999, 87, 381–386
Characterization of chlorobenzoate degraders isolated frompolychlorinated biphenyl-contaminated soil and sediment in theCzech Republic
L. Pavlu 1, J. Vosa hlova 1, H. Klierova 2, M. Prouza 2, K. Demnerova 2 and V. Brenner 1
1Institute of Microbiology, Academy of Sciences of the Czech Republic and 2Institute of Chemical Technology,Prague, Czech Republic
7021/01/99: received 8 January 1999, revised and accepted 30 April 1999
L. PAVLU, J. VOSAHLOVA, H. KLIEROVA, M. PROUZA, K. DEMNEROVA AND V. BRENNER. 1999.
Two polychlorinated biphenyl-contaminated sites in the Czech Republic, a soilat Zamberk and a sediment sludge at Milevsko, were screened for the presence ofchlorobenzoate degraders. Sixteen different chlorobenzoate degraders were isolated from thesoil compared with only three strains isolated from the sediment. From these strains,only four soil degraders and one strain isolated from the sediment, respectively, were shownto possess a complete chlorobenzoate (CB) pathway. Bacteria isolated from the soil haveexpressed more flexibility for CB degradation, namely in the case of ortho-chlorinatedbenzoates. They all possessed large plasmids, the restriction patterns of which werecompared. Plasmids in Pseudomonas sp. A7, A8, A18 and A19, respectively, werecured and found to encode at least part of the metabolic pathway involved in the growthon ortho-chlorinated benzoates.
INTRODUCTION
Chlorobenzoates are environmentally important as com-pounds produced during and affecting the microbial degra-dation of polychlorinated biphenyls (PCBs). While the lattercompounds are widespread soil pollutants in all westernnations, the problems related to their presence are particularlysevere in the former Soviet block countries such as the CzechRepublic.
In the past, a number of strategies to isolate PCB degraderswith enhanced ability to degrade more chlorinated congenershave been evaluated (Brenner et al. 1994). Chlorobenzoate-degrading bacteria were shown frequently to be absent insoils while PCB-co-metabolizing strains were routinely iso-lated from contaminated soil by the enrichment on biphenyl(Hickey et al. 1993). Hernandez et al. (1991) isolated Pseudo-monas putida P111 which was shown to have the greatest rangeof chlorobenzoate utilization yet reported. Further studies(Brenner et al. 1993) revealed the presence of the gene forortho-chlorobenzoate-1,2-dioxygenase (OCBD) on trans-missible plasmid pPB111 which was shown to be responsible
Correspondence to: V. Brenner, Institute of Microbiology, Academy ofSciences of the Czech Republic, Vıdenska 1083, 142 20 Prague, Czech Republic.
© 1999 The Society for Applied Microbiology
for the extraordinarily broad degradation activities of thisstrain. The same type of dioxygenase was found on andcloned from the plasmid pBAH1 in the strain Ps. cepacia2CBS (Haak et al. 1995). The frequency and diversity ofchlorobenzoate degraders on PCB-contaminated sites, andtheir impact on PCB degradation, have not yet been fullyinvestigated, although molecular diagnostics in such soilshave been performed for classical biphenyl degraders carryinggenes of the bph operon (Layton et al. 1994).
A number of authors (Furukawa et al. 1978; Thiem et al.1994; Hernandez et al. 1995) have shown that soils which werenot exposed to chloro-organics do not harbour chloroaromaticdegraders. In particular, Brunsbach and Reineke (1993)showed that soils require inoculation with chlorobenzenedegraders before the degradation will take place. Other stud-ies showed that the inoculation of soil with chlorobenzoatedegraders increased the mineralization of PCBs in soil morethan the addition of chlorobiphenyl-co-metabolizing strains(Hickey et al. 1993; Focht et al. 1996). To our knowledge,there has not yet been a comprehensive study reflecting thediversity of the chlorobenzoate-degrading population onPCB-contaminated sites. Therefore, in this study weintended to compare the chlorobenzoate microbial populationisolated from both PCB-contaminated soil and sediment.

382 L. PAVLU ET AL.
MATERIALS AND METHODS
Site characterization
The soil sample (Zamberk) contained 300mg PCB mixtureDelor 103 per kg dry soil (condensators from ZEZ SILKO,Zamberk 1984–92). The sediment sludge (Milevsko) con-tained 200mg PCB mixture Delor 103 per kg dry matter(coating plant).
The PCB mixture Delor 103 is a moderately chlorinatedcommercial product with a prevalence of tri- and tetra-chlori-nated congeners (Chemko Strazske, Slovakia). It roughly cor-responds to the mixture Aroclor 1242 (USA).
Media and reagents
The mineral salts (MS) (Hickey and Focht 1990) and com-plete Luria-Bertani (Sambrook et al. 1989) media were used.All chemicals used were of analytical grade and availablecommercially. The enzymes used were provided by Amersham(Buckinghamshire, UK).
Characterization of bacteria
Bacteria were characterized microscopically, morphologicallyand by the commercial kits NEFERM (Lachema, CzechRepublic) and Biolog (Biolog Inc., USA).
Enrichment procedures
Soil or sludge (1g) was added to 50ml MS medium (seebelow) containing 500mg l−1 2-, 3- or 4-chlorobenzoate (CB)or 2,5-dichlorobenzoate (DCB) as the sole source of carbonand energy. The cultivation was carried out at 28 °C on anorbital shaker. After 1 week 0·5ml of the suspension wastransferred into a fresh MS medium under the sameconditions. The transfer was repeated four times.
Enzyme assays
Cells were grown aerobically in 100ml MS medium con-taining 500mg l−1 substrate at 28 °C on an orbital shaker,harvested by centrifugation and washed twice with phosphatebuffer (0·1mol l−1, pH 7·5). The cells were resuspended inthe same buffer and disrupted by sonication (6× 15 s). Celldebris was removed by centrifugation (22 000 g, 20min, 4°C). Catechol dioxygenase activities were assayed spec-trophotometrically by measuring the rate of cis,cis-muconicacid (260 nm) and 2-hydroxymuconic semialdehyde(375 nm), respectively (Miguez et al. 1990). The reaction mix-ture was composed of 20 ml crude extract, 970 ml phosphatebuffer and 10 ml catechol (0·5mmol l−1). Protein con-centration was determined by the method of Bradford withbovine serum albumin as a standard.
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 87, 381–386
DNA manipulation
Plasmid DNA isolation and DNA digestions by restrictionendonucleases were performed as described by Sambrooket al. (1989) and total DNA was isolated by the methoddescribed by Adams et al. (1992). A non-radioactive DNAlabelling and detection kit (Genius; Boehringer Mannheim,Indianapolis, IN, USA) was used according to the manu-facturer’s instructions. The ability to utilize CBs was elim-inated by repeated subculture of Pseudomonas sp. A7, A8,A18 and A19 five times on complete medium.
Analytical procedures
The decrease in 3-CB, 4-CB and 2,5-DCB concentrations incultivation media was monitored by high-performance liquidchromatography (HPLC). During the degradation of 3-CBthe accumulation of 3-chlorocatechol (3-ClC) was monitored.Samples were centrifuged to remove cells (11 000 g, 3min)and measured using an HP 1090L with diode-array detector(Hewlett-Packard, Amstelveen, The Netherlands). Allmeasurements were carried out under the following con-ditions: stationary phase, LiChrospher RP-18, 5 mm; column,125× 4mm i.d. (Merck, Darmstadt, Germany); guardcolumn, 40× 4mm i.d., 30 mm; mobile phase, wat-er:acetonitrile in ratio 62:38 (v/v); flow rate, 1·0mlmin−1;temperature, 40 °C; detection, u.v. at 230 nm. The retentiontimes of the following compounds were: 3-CB, 4·99min; 4-CB, 5·39min; 2,5-DCB, 3·10min and 3-ClC, 4·22min. Thelimit of detection (counted as 3 × baseline noise) was: 3-CB,4·5 mg l−1; 4-CB, 2·6 mg l−1; 2,5-DCB, 3·0 mg l−1 and 3-ClC,4·9 mg l−1 (injection volume 10 ml).
Chloride release was measured electrochemically by anion-selective electrode (model 96–17B; Orion, Boston, MA,USA). A 5-ml sample of cultures was collected and dilutedwith 50ml distilled water and 0·5ml low-level ISA (1mol l−1
NaNO3) was added. The concentration of released chlorideswas determined from the calibration curve (standard solutionsof 1× 10–5–4·8× 10–4 mol l−1 NaCl).
RESULTS
The method of continual enrichment on selected CBs wasused to collect six mixed bacterial cultures (Table 1). TheCBs used for the first enrichment included 2-, 3- and 4-CB and 2,5-DCB. These substrates were supplied into themineral salts media as the sole sources of carbon and energy.Four mixed cultures were isolated from the soil at Zamberkon each of the supplied substrates, while only two wereobtained from the sediment originating from Milevsko. Thesetwo mixed cultures were growing on 3-CB and 4-CB, respec-tively, but we have failed to isolate from the sediment anybacterial mixtures growing on ortho-chlorinated benzoates as
CHARACTERIZATION OF CHLOROBENZOATE DEGRADERS 383
Table 1 Cultures collected by the method of continualenrichment on selected chlorobenzoates—–––––––––––––––––––––––––––––––––––––––––––––––––––––
Mixture Purified strains Substrate Sample—–––––––––––––––––––––––––––––––––––––––––––––––––––––SM1 A7, A8 2-CB Zamberk
A13, A14, A15 2,4-DCBA16, A17 2,3-DCBA18, A19 2,5-DCB
SM2 A10, A11 3-CB ZamberkSM3 A9, A12 4-CB ZamberkSM4 A4 3,5-DCB ZamberkSM6 B3 3-CB MilevskoSM7 B5, B6 4-CB Milevsko—–––––––––––––––––––––––––––––––––––––––––––––––––––––
CB, Chlorobenzoate; DCB, dichlorobenzoate.
the sole source of carbon and energy. These results cor-respond with the data obtained from cell counts when6·1× 105 cells g−1 were found in the soil sample and8·5× 104 cells g−1 were obtained from the sediment (per1 g dry matter). The bacteria from the mixed cultures werepurified on a complete Luria agar and pure cultures obtainedwere distinguished and characterized on the basis of mor-phological differences. All strains isolated were mobile Gram-negative rods with smooth or mucoid colonies, white andyellow colour and an elevated profile. Further identification ofthese strains was performed on NEFERM diagnostic titrationplates and Biolog GN titration plates. The strains designatedA7 and A8 were closest to Ps. fluorescens, strain A18 toPs. pseudoalcaligenes and strains A19 and B3 to Ps. stutzeri.However, due to the limitations of the taxonomic methodsused, we have decided to use the temporary designationPseudomonas sp. in all cases.
After the purification, the ability of individual strains tocompletely utilize substrates initially supplied to the mixedcultures was tested in a set of batch cultures containing MSmedia and 500mg l−1 of selected CBs. In this experiment wecould collect only five pure utilizers, four of them of soilorigin and only one pure culture from the sediment. Sur-prisingly, the soil pure cultures were able to utilize only ortho-chlorinated benzoates, while the sediment isolate could growon all monochlorinated benzoates. The utilization of CBs bypure cultures is summarized in Table 2. The degradationspectrum of these strains is not as broad as that of versatileCB degraders such as Ps. putida P111 (Brenner et al. 1993) orPs. aeruginosa JB2 (Hickey and Focht 1990) and all bacteriaisolated from the soil had quite similar CB utilization patterns.We suggest that the presence of transmissible specializedortho-chlorinated dioxygenase could be responsible for thisphenomenon. We have isolated large plasmids from all strainsdegrading ortho-chlorinated benzoates and compared their
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 87, 381–386
Table 2 Utilization of chlorobenzoates by the isolated strains—–––––––––––––––––––––––––––––––––––––––––––––––––––––
Tested strain—––––––––––––––––––––––––––––––––––––––––
Substrate B3 A7 A8 A18 A19—–––––––––––––––––––––––––––––––––––––––––––––––––––––2-CB + + + + +3-CB + − − − −4-CB + − − − −3,5-DCB − − − − −2,3-DCB − − − − −2,4-DCB − − − − −2,5-DCB − + + + +2,6-DCB − − − − −2,3,5-TCB − − − 2 2—–––––––––––––––––––––––––––––––––––––––––––––––––––––
CB, Chlorobenzoate; DCB, dichlorobenzoate; TCB, trichloro-benzoate.
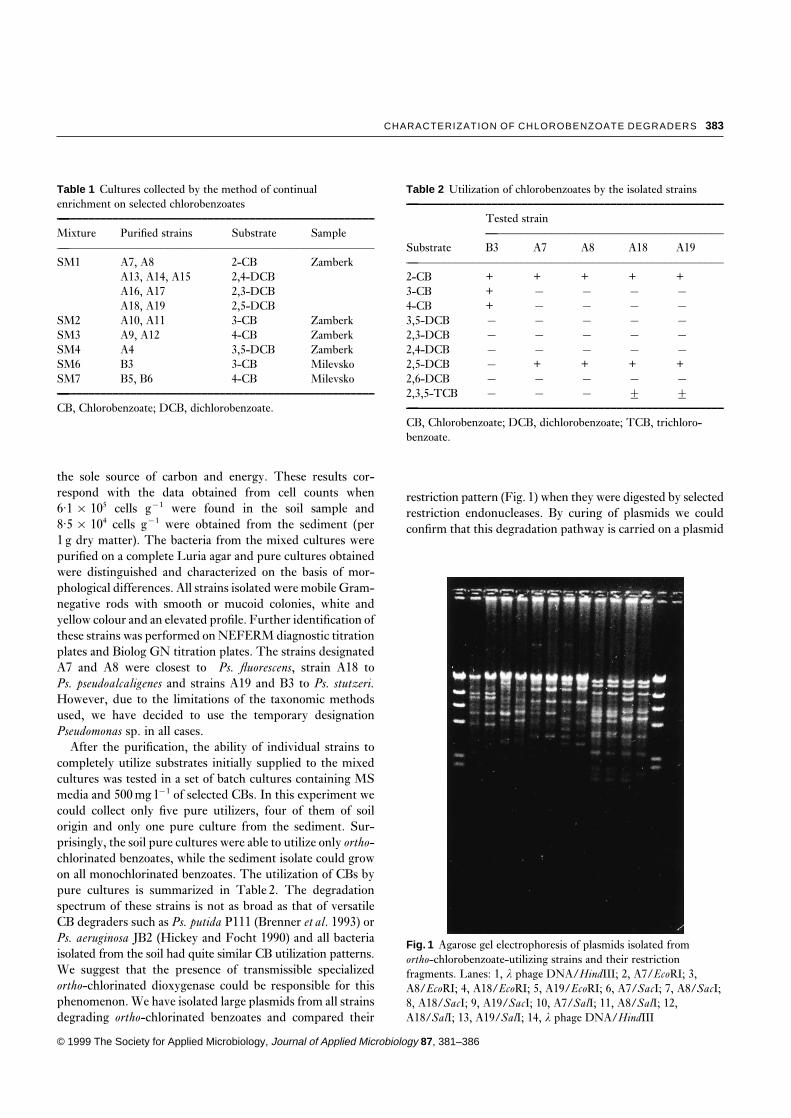
restriction pattern (Fig. 1) when they were digested by selectedrestriction endonucleases. By curing of plasmids we couldconfirm that this degradation pathway is carried on a plasmid
Fig. 1 Agarose gel electrophoresis of plasmids isolated fromortho-chlorobenzoate-utilizing strains and their restrictionfragments. Lanes: 1, l phage DNA/HindIII; 2, A7/EcoRI; 3,A8/EcoRI; 4, A18/EcoRI; 5, A19/EcoRI; 6, A7/SacI; 7, A8/SacI;8, A18/SacI; 9, A19/SacI; 10, A7/SalI; 11, A8/SalI; 12,A18/SalI; 13, A19/SalI; 14, l phage DNA/HindIII
384 L. PAVLU ET AL.
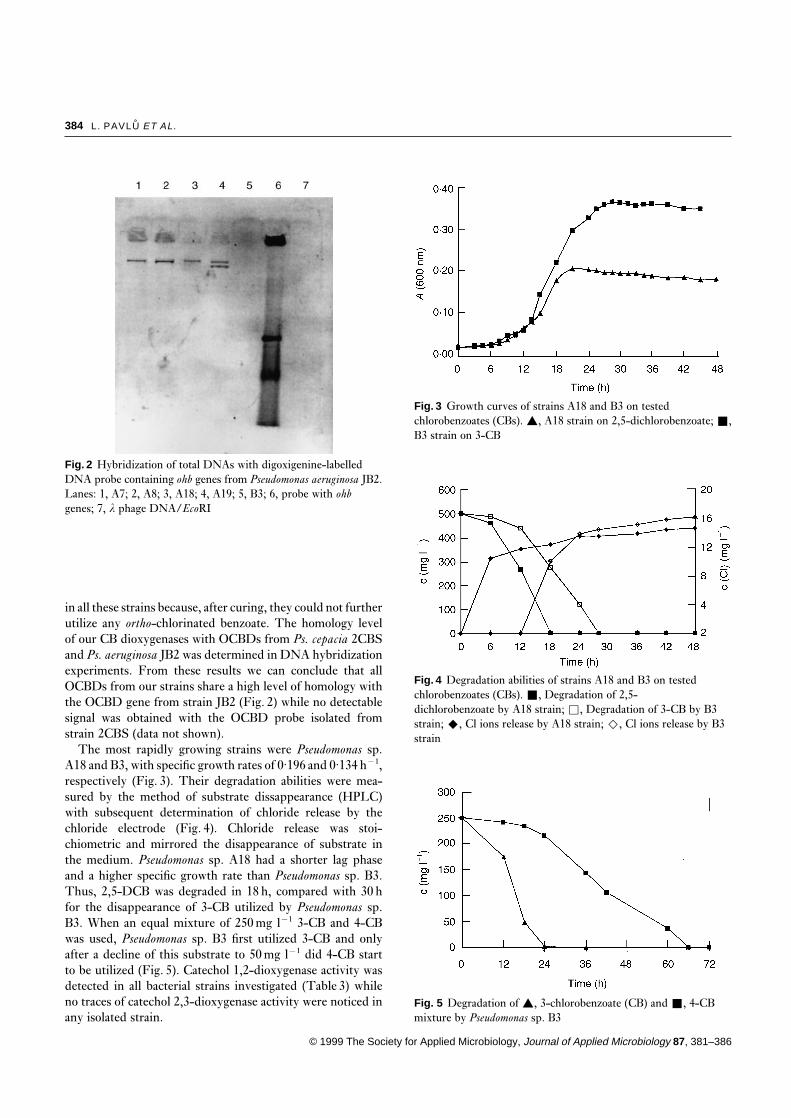
Fig. 2 Hybridization of total DNAs with digoxigenine-labelledDNA probe containing ohb genes from Pseudomonas aeruginosa JB2.Lanes: 1, A7; 2, A8; 3, A18; 4, A19; 5, B3; 6, probe with ohbgenes; 7, l phage DNA/EcoRI
in all these strains because, after curing, they could not furtherutilize any ortho-chlorinated benzoate. The homology levelof our CB dioxygenases with OCBDs from Ps. cepacia 2CBSand Ps. aeruginosa JB2 was determined in DNA hybridizationexperiments. From these results we can conclude that allOCBDs from our strains share a high level of homology withthe OCBD gene from strain JB2 (Fig. 2) while no detectablesignal was obtained with the OCBD probe isolated fromstrain 2CBS (data not shown).
The most rapidly growing strains were Pseudomonas sp.A18 and B3, with specific growth rates of 0·196 and 0·134 h−1,respectively (Fig. 3). Their degradation abilities were mea-sured by the method of substrate dissappearance (HPLC)with subsequent determination of chloride release by thechloride electrode (Fig. 4). Chloride release was stoi-chiometric and mirrored the disappearance of substrate inthe medium. Pseudomonas sp. A18 had a shorter lag phaseand a higher specific growth rate than Pseudomonas sp. B3.Thus, 2,5-DCB was degraded in 18 h, compared with 30 hfor the disappearance of 3-CB utilized by Pseudomonas sp.B3. When an equal mixture of 250mg l−1 3-CB and 4-CBwas used, Pseudomonas sp. B3 first utilized 3-CB and onlyafter a decline of this substrate to 50mg l−1 did 4-CB startto be utilized (Fig. 5). Catechol 1,2-dioxygenase activity wasdetected in all bacterial strains investigated (Table 3) whileno traces of catechol 2,3-dioxygenase activity were noticed inany isolated strain.
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 87, 381–386
Fig. 3 Growth curves of strains A18 and B3 on testedchlorobenzoates (CBs). R, A18 strain on 2,5-dichlorobenzoate; Ž,B3 strain on 3-CB
Fig. 4 Degradation abilities of strains A18 and B3 on testedchlorobenzoates (CBs). Ž, Degradation of 2,5-dichlorobenzoate by A18 strain; �, Degradation of 3-CB by B3strain; E, Cl ions release by A18 strain; e, Cl ions release by B3strain
Fig. 5 Degradation of R, 3-chlorobenzoate (CB) and Ž, 4-CBmixture by Pseudomonas sp. B3

CHARACTERIZATION OF CHLOROBENZOATE DEGRADERS 385
Table 3 Specific activities of catechol-1,2-dioxygenase—–––––––––––––––––––––––––––––––––––––––––––––––––––––
Strains Substrates Specific activity (mU mg−1 protein)—–––––––––––––––––––––––––––––––––––––––––––––––––––––B3 3-CB 647
4-CB 1153A7 2-CB 52
2,5-DCB 63A8 2-CB 71
2,5-DCB 55A18 2-CB 126
2,5-DCB 96A19 2-CB 234
2,6-DCB 81—–––––––––––––––––––––––––––––––––––––––––––––––––––––
CB, Chlorobenzoate; DCB, dichlorobenzoate.
DISCUSSION
The data regarding the utilization of specific CBs suggestthat the CB pathways utilized on two investigated sites are ofdifferent origin. This is supported by the fact that even mixedcultures isolated from the sediment at Milevsko cannot utilizeany ortho-dichlorinated benzoate while all pure culturesfound in the soil at Zamberk are utilizing 2,5-DCB. On theother hand, pure soil cultures could not utilize other thanortho-chlorinated benzoates which supports the hypothesisthat, in that particular microbial community, a specializedOCBD which cannot attack meta- and para-chlorinated ben-zoates was spread on a transmissible plasmid encoding at leastpart of the metabolic pathway for the degradation of ortho-chlorinated benzoates. This hypothesis is also supported bythe comparison of plasmid restriction patterns in all strainsdegrading ortho-chlorinated benzoates which were found tobe identical in Pseudomonas sp. A7 and A8 and only slightlymodified in Pseudomonas sp. A18 and A19, respectively. Noplasmid was found in Pseudomonas sp. B3. Thus, the presenceof a less specific benzoate dioxygenase, probably coded onthe chromosome, was suggested in sediment isolates whilethe more specialized OCBD is missing in that particularenvironment. The benzoate dioxygenase is apparently presenteven in the soil mixed cultures, but no single strain isolatedfrom the soil was found to possess a complete pathway forthe degradation of meta- or para-chlorinated benzoates. Ofall four soil isolates degrading ortho-CBs Pseudomonas sp.A19 has the highest versatility in the number of degradedchlorobenzoic isomers (Table 2). As the origin of genes uti-lized for the degradation of ortho-CBs in these strains appearsto be the same (Figs 1 and 2) we speculate that broadeningof the benzoate-1,2-dioxygenase specificity is due to eithermutation events or to plasmid recombination in individualstrains. The nature and character of these events will be thesubject of our further studies.
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 87, 381–386
The significance of the CB metabolic pathway for thedegradation of PCBs has already been indicated (Fava et al.1993; Hickey et al. 1993; Focht et al. 1996). In our recentexperiment focusing on the degradation of commercial PCBmixture Delor 103 by co-metabolizing biphenyl degraders wehave noticed the accumulation of high amounts of ortho-chlorinated benzoates, namely 2,5-DCB, in a reactionmixture. This finding is relevant to the already known inhi-bition effects of metabolic products from the lower part ofthe PCB metabolic pathway on the enzymes from the upperregion, namely on 3-phenylcatechol 2,3-dioxygenase (Adamset al. 1992). Therefore, this study should be followed bymore experiments focusing on the degradation activity of CBdegraders on PCB-contaminated sites to better understandtheir bioremediation potentials.
REFERENCES
Adams, R.H., Huang, C.-M., Higson, F.K., Brenner, V. and Focht,D.D. (1992) Construction of a 3-chlorobiphenyl-utilizing recom-binant from an intergeneric matting. Applied EnvironmentalMicrobiology 58, 647–654.
Brenner, V., Arensdorf, J.J. and Focht, D.D. (1994) Genetic con-struction of PCB degraders. Biodegradation 5, 359–377.
Brenner, V., Hernandez, B.S. and Focht, D.D. (1993) Variationin chlorobenzoate catabolism by Pseudomonas putida P111 as aconsequence of genetic alterations. Applied Environmental Micro-biology 59, 2790–2794.
Brunsbach, F.R. and Reineke, W. (1993) Degradation of chlo-robenzoates in soil slurry by special organisms. Applied Micro-biological Biotechnology 39, 117–122.
Fava, F., Di Gioia, D., Marchetti, L., Quattroni, G. and Maraffa,V. (1993) Aerobic mineralization of chlorobenzoates by a naturalpolychlorinated biphenyl-degrading mixed bacterial culture.Applied Microbiological Biotechnology 40, 541–548.
Focht, D.D., Sealers, D.B. and Koh, S.-Ch. (1996) Genetic exch-ange in soil between introduced chlorobenzoate degraders andindigenous biphenyl degraders. Applied Environmental Micro-biology 62, 3910–3913.
Furukawa, K., Matsumura, F. and Tonomura, K. (1978) Alcaligenesand Acinetobacter strains capable of degrading polychlorinatedbiphenyls. Agriculture Biological Chemistry 42, 543–548.
Haak, B., Fetzner, S. and Lingens, F. (1995) Cloning, nucleotidesequence, and expression of the plasmid encoded genes for thetwo-component 2-halobenzoate 1,2-dioxygenase from Pseudo-monas cepacia 2CBS. Journal of Bacteriology 177, 667–675.
Hernandez, B.S., Arensdorf, J.J. and Focht, D.D. (1995) Cataboliccharacteristics of biphenyl-utilizing isolates which cometabolizePCBs. Biodegradation 6, 75–82.
Hernandez, B.S., Higson, K., Kondrat, R. and Focht, D.D. (1991)Metabolism of and inhibition by chlorobenzoates in Pseudomonasputida P111. Applied Environmental Microbiology 57, 3361–3366.
Hickey, W.J. and Focht, D.D. (1990) Degradation of mono-, di-,and trihalogenated benzoic acids by Pseudomonas aeruginosa JB2.Applied Environmental Microbiology 56, 3842–3850.
Hickey, W.J., Searles, D.B. and Focht, D.D. (1993) Enhanced

386 L. PAVLU ET AL.
mineralization of polychlorinated biphenyls in soil inoculatedwith chlorobenzoate-degrading bacteria. Applied EnvironmentalMicrobiology 59, 1194–1200.
Layton, A.C., Lajoie, C.A., Easter, J.P., Jernigan, R., Sanseverino,J. and Sayler, G.S. (1994) Molecular diagnostics and chemicalanalysis for assessing biodegradation of polychlorinated biphenylsin contaminated soils. Journal of Industrial Microbiology 13, 392–401.
Miguez, C.B., Greer, C.W. and Ingram, J.M. (1990) Degradation
© 1999 The Society for Applied Microbiology, Journal of Applied Microbiology 87, 381–386
of mono- and dichlorobenzoic acid isomers by two natural isolatesof Alcaligenes denitrificans. Archives of Microbiology 154, 139–143.
Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Appendix. InMolecular Cloning, 2nd edn. Cold Spring Harbor LaboratoryPress.
Thiem, S.M., . Krumme, M.L., Smith, R.L. and Tiedje, J.M.(1994) Use of molecular techniques to evaluate the survival of amicroorganism injected into an aquifer. Applied EnvironmentalMicrobiology 60, 1059–1067.