analysis of bph operon from the polychlorinated biphenyl
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1992 by The American Society for Biochemistry and Molecular Biology, Inc
Vol. 267, No. 7, Issue of March 5, pp. 4M4-4853, 1992 Printed in U.S.A.
Analysis of bph Operon from the Polychlorinated Biphenyl-degrading Strain of Pseudomonas pseudoalcaligenes KF707*
(Received for publication, May 31,1991)
Kazunari Tairazf, Jun Hiroseli, Shinsaku Hayashidall, and Kensuke FurukawaSll From the $Fermentation Research Institute, Agency of Industrial Science & Technology, Ministry of International Trade and Industry, Tsukuba Science City 305 and the VDepartment of Agricultural Chemistry, Faculty of Agriculture, Kyushu University, Fukuoka812, Japan
The entire nucleotide sequences (6.8 kilobase pairs) of the bphABC genes and thier products involved in the initial dioxygenation and ring-meta-cleavage of biphenyls and polychlorinated biphenyls were deter- mined. The first bphA gene starts about a 100 base pairs downstream from the transcriptional initiation site. The bphA region, which encodes a cluster of en- zymes including biphenyl dioxygenase catalyzing the initial catabolic step, consists of five open reading frames (ORFs). Five proteins corresponding to these ORFs in the molecular masses were detected by in vitro protein synthesis, of which four ORFs are very similar to the recently reported todClC2BA genes coding for the corresponding enzymes catalyzing the initial diox- ygenation reactions of toluene (Zylstra, G. J., and Gib- son, D. T. (1989) J. Biol. Chem. 264, 14940-14946). The third open reading frame (ORF3) of the bphA region, missing its counterpart in the toluene dioxy- genase gene cluster, was site-specifically deleted, and the resulting enzymatically active mutant reveals that this ORF3 is not mandatory for the catabolism of bi- phenyls. Thus the biphenyl degradation pathway and the responsible enzymes/genes are very similar to those of toluene degradation despite their discrete substrate specificity.
Since identification of polychlorinated biphenyls (PCBs)’ in the environment in 1966, a number of PCB-degrading microorganisms have been isolated (1-10). These PCB de- graders utilize biphenyl as the sole source of carbon and energy. They cometabolize PCBs to chlorobenzoic acids by the major oxidative route as illustrated in Fig. 1 (4). We have cloned at least part of the catabolic genes from three classes of Pseudomonas strains: bphABCXD genes from Pseudomonas pseudoalcaligenes KF707, bphABCD genes from Pseudomonas putida KF715, and bphC gene from Pseudomonaspaucimobilis Q1 (11-15). In their first catabolic mechanistic stage, a mo-
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequencefs) reported in thispaper has been submitted to the GenBankTM/EMBL Data Bank with accession numbeds) M83673.
3 Correspondence can be addressed either to K.T. (MITI) or K.F. (Kyushu University). Correspondence to K.T. before October, 1992 should be addressed to: Counselor for Human Frontier Science Pro- gram, 1-3-1 Kasumigaseki, Chiyoda-ku, Tokyo 100, Japan. Tel.: 03- 3501-1367; Fax: 03-3501-5953.
The abbreviations used are: PCB, polychlorinated biphenyl; HPDA, 2-hydroxy-6-oxo-6-phenylhexa-2,4-dienoic acid; kb, kilobase pair(s); ORF, open reading frame; bp, base pair(s).
lecular oxygen is introduced at the 2,3 position of the non- chlorinated or less chlorinated ring to produce a dihydrodiol (compound I1 in Fig. 1) by the action of a group of enzymes including biphenyl dioxygenase (products of a gene cluster in bphA region; see below). The dihydrodiol is then dehydrogen- ated to a 2,3-dihydroxybiphenyl (111) by a dihydrodiol dehy- drogenase (product of bphB). The 2,3-dihydroxybiphenyl is cleaved at the 1,2 position by the 2,3-dihydroxybiphenyl diox- ygenase (product of bphC) to yield IV, which is eventually hydrolyzed to the corresponding chlorobenzoic acid, V, by 2- hydroxy-6-oxo-6-phenylhexa-2,4-dienoic acid (HPDA) hydro- lase (product of bphD). The function of bphX region which exists in P. pseudoalcaligenes KF707 but not in P. putida KF715 is not yet elucidated. The 2,3dihydroxybiphenyl diox- ygenases (bphC gene products) from strains KF707 and Q1 are enzymatically very similar (13, 14), but the bphC DNAs of KF707 and Q1 did not hybridize with each other, and the two enzymes showed no immunological cross-reactivity. Per- tinent to this observation, the nucleotide sequencing of the two bphC genes revealed that the overall sequence homology of the 2,3-dihydroxybiphenyl dioxygenases at the amino acid level was as low as 38% between the two functionally similar enzymes (13, 15).
This homology between the two bphC genes of KF707 and Q1 turned out to be even lower than that between the 2,3- dihydroxybiphenyl dioxygenase (KF707) and the recently re- ported 3-methylcatechol2,3-dioxygenase of toluene degrader P. putida F1, with the latter value being 55% (16). This result is surprising, when we consider their substrate specificities, since both KF707 and Q1 dioxygenases recognize similar substrates of mainly biphenyl analogues, whereas 3-methyl- catechol 2,3-dioxygenase from P. putida F1 recognizes much smaller substrates of catechol derivatives (16, 17). Here we report the first nucleotide sequence for the enzymes involved in the initial oxidation of biphenyl/PCB. We find that the trend of high similarity between the toluene degrader and the PCB degrader extends to the bphA region. The complete nucleotide sequence determination of the 6.8-kb far left X h I fragment of P. pseudoalcaligenes KF707 (Fig. 1) reveals that the gene organization as well as the size and homology of the corresponding enzymes between the PCB degrader (P. pseu- doalcaligenes KF707) and the toluene degrader (P. putida F1) are highly conserved, indicating the common ancestry despite their discrete substrate specificities.
MATERIALS AND METHODS
Sequence Determination-Both pHSG396 vector (2.24 kb; Takara Shuzo, Kyoto, Japan) and pMFB2 containing bphABC region (14) were digested by XhoI, mixed after phenol/chloroform treatment, ligated, and transformed into JM109. White colonies possessing the 6.8-kb XhoI fragment (formerly reported to be a 7.2-kb fragment) containing bphABC of pMFB2 (14) were identified by spraying with
4844

Analysis of bph Operon 4845
2,3-dihydroxy biphenyl. The resulting subclone pKTF10, where the orientation of the bphABC was reverse of lac2 gene, was obtained. The transformants harboring pKTFlO turned yellow upon being sprayed with 2,3-dihydroxy biphenyl. Then the XbaI-Hind111 frag- ment of pKTFlO containing the entire bphABC gene was further subcloned into the XbaI/HindIII sites of pUC118 and pUC119 to generate pKTF18 and pKTF20, respectively, of forward and reverse orientations with respect to lac2 gene. Various deletion mutants were constructed by treating KpnIIXbaI double-digested pKTF20, where the orientation of the bphABC was reverse of lac2 gene, with (i) exonuclease 111, (ii) mung bean nuclease, (iii) Klenow fragment with dNTP, and finally (iv) with ligase. The ligated DNA was transformed into competent Escherichia coli MV1184 cells and appropriately sized deletion mutants were selected by EcoRI/XhoI double digestion. Then single-stranded DNAs were isolated (18) for appropriately sized dele- tion mutants. Sequencing was performed on 370A DNA sequencer (Applied Biosystems). The deletion mutants of pKTF20 enabled us to sequence from the direction of bphA to bphC. The opposite strand was similarly sequenced either by taking advantage of already known restriction enzyme sites and deleting proper DNA regions between the restriction sites or synthesizing dye primers (Oliglow, Applied Biosystems). Thus, both the forward and reverse strands were se- quenced in their entirety to yield the nucleotide sequence of the 6780- base pair XhoI fragment shown in Fig. 2. Since there were many regions where full stops and/or band compressions prevented correct assignment of signals, the 6.8-kb XhoI fragment containing the bphABC of pMFB2 was further digested with PstI, yielding five smaller fragments and each fragments were individually subcloned into PstI site of pUC119. This fragmentation into smaller units, however, did not solve the compression problems. All sequencing was performed on single-stranded DNAs, initially utilizing Klenow frag- ment. With Klenow, however, since there were many regions where full stops and/or band compressions prevented correct assignment of signals, further sequencing was performed utilizing either Taq DNA polymerase or Sequenase with dITP. Especially with Sequenase, dye primers were designed to anneal -50 nucleotides upstream of the stacked regions. Although Sequenase lacks in handiness, the compres- sions were best avoided by the Sequenase/dITP.
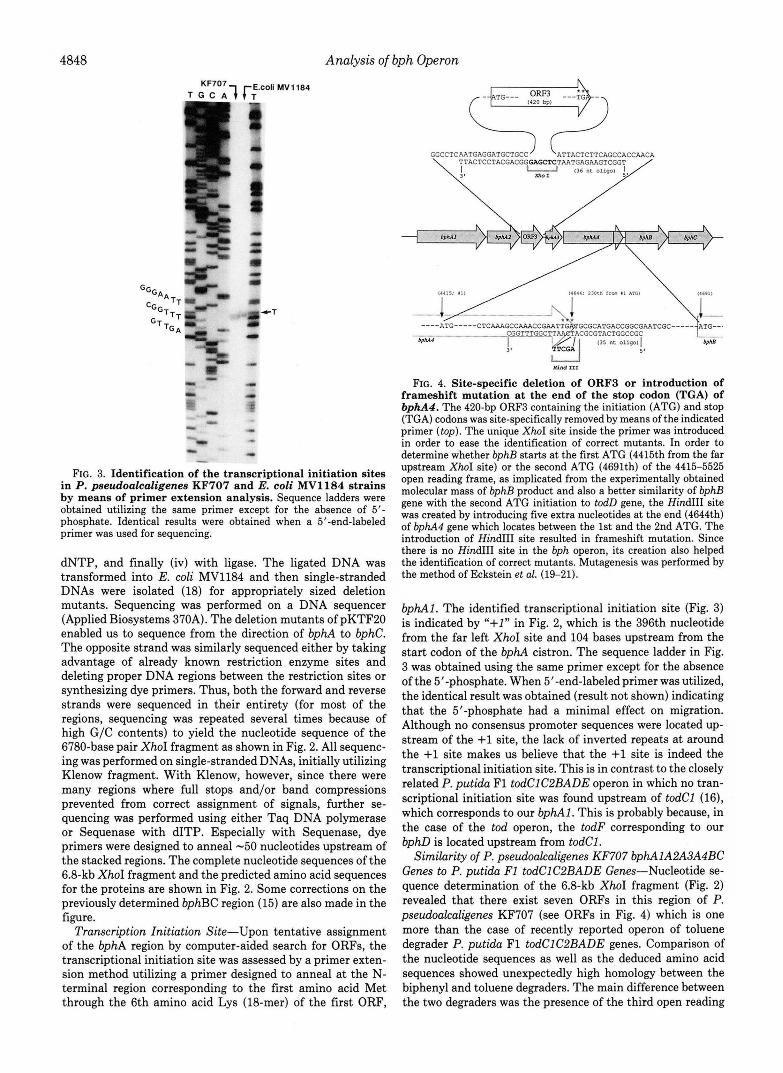
Primer Extension to Determine Transcription Initiation Site- Upon tentative assignment of the bphA region, the transcriptional initiation site was assessed by a primer extension method utilizing a primer designed to anneal at the N-terminal region corresponding to the first amino acid Met through the 6th amino acid Lys (M-mer, synthesized by Applied Biosystems 380B) of bphAI, and the result is shown in Fig. 3. The primer was labeled by T4 polynucleotide kinase.
mRNAs were prepared from either P. pseudoalcaligenes KF707 or E. coli MV1184 grown to the Klett value of 70-90. The samples were reverse-transcribed and loaded next to a sequencing sample. The sequence ladder was obtained using the same primer except for the absence of the 5”phosphate. When 5’-end-labeled primer was uti- lized, the identical result was obtained indicating that the 5”phos- phate had a minimal effect on migration.
Mutagenesis-A mutant in which ORF3 was site-specifically de- leted was constructed by the method of Eckstein et al. (19-21). For the construction of the ORF3 deletion mutant, 36-mer oligonucleotide was synthesized whose fifteen 5’-nucleotides were designed to bind just downstream of the stop codon (TGA) of ORF3 and similarly fifteen 3”nucleotides were designed to bind just upstream of the initiation codon (ATG) of ORF3 (see Fig. 4). In order to facilitate the identification of correct mutants, XhoI site was introduced, as shown in Fig. 4, between the 5’- and 3’-annealing regions. This primer was gradually annealed to single-stranded DNA of wild-type pKTF18 by dumping the tube containing the mixture into a boiling water. The whole boiling water bath was then moved to a cold-room and kept a t 4 “C. The initial polymerization/ligation was carried out with Klenow fragment and T4 DNA ligase at 16 “C utilizing dCTP(aS) (dCTP possessing thiophosphate linkage at the a-position), as described by Eckstein et al. (19-21, Amersham). After treatment with (i) NciI, (ii) exonuclease 111, and (iii) Klenow fragment with regular dNTP and T4 DNA ligase, competent E. coli MV1184 cells were transformed by the ligated mixture. When the single-stranded DNA containing C ( a S ) (cytidine with thiophosphate linkage) were used without the final polymerization/ligation step to transform the competent E. coli MV1184 cells, the transformation frequency was about 1/100 of the normally completed double stranded cases. One correct mutant was found among 36 arbitrarily chosen colonies. Sequencing of the mutant confirmed the site-specific deletion of the entire 420-bp ORF3 starting from the initiation codon ATG at 2598 and ending after the stop codon TGA at 3017. Another mutation (frameshift mutation) was site-specifically introduced just downstream of the stop codon TGA of bphA4, in a similar manner as shown in Fig. 4 (bottom). Similarly, various mutations were introduced into bphC region.
In Vitro Synthesis of Polypeptides-Zn vitro protein synthesis was carried out using prokaryotic DNA-directed translation kit (Amer- sham, Japan). We incubatedpUC118, pKTF18, andpKTFl8delORF3 with the reaction mixture containing E. coli extract (strain MRE6OO RNase I-), nucleotides, tRNA, energy-generating system, and inor- ganic salts. The polypeptides were labeled with ~-[4,5-~H]leucine. The samples were subjected to 15% sodium dodecyl sulfate-polyacryl- amide gel electrophoresis. After the electrophoresis, the gel was
A A
0 8 ? & n’ f ’ k o Y O O H f ’ bphC
T COOH
0 2 OH NAD+ NADH + H’ OH 0 2
I I1 I11 IV V
1 kb
(11.3 kb) FIG. 1. Catabolic pathway for degradation of biphenyl and chlorinated biphenyls and organization
of bph operon in P. pseudoalcaligenes KF707. The bphA region consists of a t least four genes and their products convert biphenyl (I) to dihydrodiol (11). By the action of dihydrodiol dehydrogenase (product of bphB), I1 is converted to 2,3-dihydroxybiphenyl (111). I11 is then cleaved at 1, 2-position by the action of 2,3-dihydroxy- biphenyl dioxygenase (product of bphC), yielding HPDA (IV) which is eventually hydrolyzed to benzoic acid (V) by HPDA hydrolase (product of bphD). bphX region is an extra DNA segment of unknown function.

4846
FIG. 2. Nucleotide sequence of the P. pseudoalcaligenee KF707 bph- AIA2A3A4BC genes and the corre- sponding amino acid sequences. The coding regions of bphA1, bphA2, ORF3, bphA3, bphA4, bphB, and bphC are at
3421, 3418-4644, 4691-5524, and 5546- 6442, respectively. Cysteine-histidine pairs separated by 17 amino residues which are characteristic of the [2Fe-2S] Rieske-type iron-sulfur proteins are in- dicated by underlines for bphA1 and bphA3. Several corrections were made on the previously reported bphEC regions (15).
500-1876, 1919-2560, 2598-3017, 3092-
1330 1340 1350 1360 1310 1380 1390 1400 1410 1420 1430 1440 G G G C T G G T T C G T C G A C W L G C G ~ ~ ~ A ~ C G G T W L
FAD binging rite?
G l y T r p P h e V a l A s p G l u P r o G l y " e U U a V a l M e t G l y P ~ ~ L Y s V a l T h ~ ~ n T y ~ T r p ~ ~ l ~ G ~ Y P r ~ ~ p ~ ~ a G l u G l ~ g l e u G l y H i s T h ~ t P ~ ~
G G T T C G A C G U L T G T T C G G C U U L ~ T G f f i C G T C I T C C C G 1450 1460 1470 1480 1490 1500 1510 1520 1530 1540 1550 1560
valAr~rgMetPheGlyGlnHiPHetSerValPh~P~~Th~y~s~~Phele~~~l~Il~nThrIleAe~h~TrpHi3ProArgGlyProAsnGluIleGluValTrpRlaPhe

Analysis of bph Operon 4847
FIG. 2. continued
C P A A G C A T C G A C W A G T ~ ~ ~ ~ G ~ T R C A ~ G ~ C G G ~ ~ C ~ T T G G C ~ G ~ ~ ~ G C 5530 5540 5550 5560 5570 5580 5590 5600 5610 5620 5630 5640
*** M e t s e r l l e l u g S e r L e u G l y T y M e t G l y P h e l l l a V a l S e r A S p V l ~
stained, fixed, and impregnated with 2,5-diphenyloxazole in dimethyl XhoI site of pHSG396, yielding pKTFlO where the orients- sulfoxide prior to drying and fluorographic exposure as described (49, tion of the b p ~ ~ c was the of gene. Then the 50). XbuI-Hind111 fragment of pKTFlO containing the entire
bphABC gene was further subcloned to the XbuIIHindIII sites of pUC118 and pUC119 to generate pKTF18 and pKTF20,
Sequence Determinution-The detailed procedures are de- respectively, of forward and reverse orientations with respect scribed under “Materials and Methods.” In short, the 6.8-kb to h c Z gene. Various deletion mutants were constructed by XhoI fragment (formerly reported to be a 7.2-kb fragment) treating KpnI/XbuI double-digested pKTF2O with (i) exonu- containing bphABC was subcloned from pMFB2 (14) to the clease 111, (ii) mung bean nuclease, (iii) Klenow fragment with
RESULTS
4848 Analysis of bph Operon
T G C A ~ F T KF707 E.coli MV1184
FIG. 3. Identification of the transcriptional initiation sites in P. pseudoalcaligenes KF707 and E. coli MV1184 strains by means of primer extension analysis. Sequence ladders were obtained utilizing the same primer except for the absence of 5’- phosphate. Identical results were obtained when a 5”end-labeled primer was used for sequencing.
dNTP, and finally (iv) with ligase. The ligated DNA was transformed into E. coli MV1184 and then single-stranded DNAs were isolated (18) for appropriately sized deletion mutants. Sequencing was performed on a DNA sequencer (Applied Biosystems 370A). The deletion mutants of pKTF2O enabled us to sequence from the direction of bphA to bphC. The opposite strand was similarly sequenced either by taking advantage of already known restriction enzyme sites and deleting proper DNA regions between the restriction sites or synthesizing dye primers. Thus, both the forward and reverse strands were sequenced in their entirety (for most of the regions, sequencing was repeated several times because of high G/C contents) to yield the nucleotide sequence of the 6780-base pair XhoI fragment as shown in Fig. 2. All sequenc- ing was performed on single-stranded DNAs, initially utilizing Klenow fragment. With Klenow, however, since there were many regions where full stops and/or band compressions prevented from correct assignment of signals, further se- quencing was performed using either Taq DNA polymerase or Sequenase with dITP. Especially with Sequenase, dye primers were designed to anneal -50 nucleotides upstream of the stacked regions. The complete nucleotide sequences of the 6.8-kb XhoI fragment and the predicted amino acid sequences for the proteins are shown in Fig. 2. Some corrections on the previously determined bphBC region (15) are also made in the figure.
Transcription Initiation Site-Upon tentative assignment of the bphA region by computer-aided search for ORFs, the transcriptional initiation site was assessed by a primer exten- sion method utilizing a primer designed to anneal at the N- terminal region corresponding to the first amino acid Met through the 6th amino acid Lys (18-mer) of the first ORF,
GGCCTCAATGAGGATGCTGCC’ ‘ATTACTCTTCAGCCACCAACA \ TTACTCCTACGACGGGAGCTCTAATGAGAAGTCGGT / GGCCTCAATGAGGATGCTGCC’ ‘ATTACTCTTCAGCCACCAACA
TTACTCCTACGACGGGAGCTCTAATGAGAAGTCGGT
h h h
U
FIG. 4. Site-specific deletion of ORF3 or introduction of frameshift mutation at the end of the stop codon (TGA) of bphA4. The 420-bp ORF3 containing the initiation (ATG) and stop (TGA) codons was site-specifically removed by means of the indicated primer (top). The unique XhoI site inside the primer was introduced in order to ease the identification of correct mutants. In order to determine whether bphB starts at the first ATG (4415th from the far upstream XhoI site) or the second ATG (4691th) of the 4415-5525 open reading frame, as implicated from the experimentally obtained molecular mass of bphB product and also a better similarity of bphB gene with the second ATG initiation to todD gene, the Hind111 site was created by introducing five extra nucleotides at the end (4644th) of bphA4 gene which locates between the 1st and the 2nd ATG. The introduction of Hind111 site resulted in frameshift mutation. Since there is no Hind111 site in the bph operon, its creation also helped the identification of correct mutants. Mutagenesis was performed by the method of Eckstein et al. (19-21).
Bind I11
bphA1. The identified transcriptional initiation site (Fig. 3) is indicated by “+l” in Fig. 2, which is the 396th nucleotide from the far left XhoI site and 104 bases upstream from the start codon of the bphA cistron. The sequence ladder in Fig. 3 was obtained using the same primer except for the absence of the 5“phosphate. When 5’-end-labeled primer was utilized, the identical result was obtained (result not shown) indicating that the 5”phosphate had a minimal effect on migration. Although no consensus promoter sequences were located up- stream of the +1 site, the lack of inverted repeats at around the +1 site makes us believe that the +1 site is indeed the transcriptional initiation site. This is in contrast to the closely related P. putida F1 todClC2BADE operon in which no tran- scriptional initiation site was found upstream of todCl (16), which corresponds to our bphA1. This is probably because, in the case of the tod operon, the todF corresponding to our bphD is located upstream from todC1.
Similarity of P. pseudoalcaligenes KF707 bphAlA2A3A4BC Genes to P. putida Fl todClC2BADE Genes-Nucleotide se- quence determination of the 6.8-kb XhoI fragment (Fig. 2) revealed that there exist seven ORFs in this region of P. pseudoalcaligenes KF707 (see ORFs in Fig. 4) which is one more than the case of recently reported operon of toluene degrader P. putida F1 todClC2BADE genes. Comparison of the nucleotide sequences as well as the deduced amino acid sequences showed unexpectedly high homology between the biphenyl and toluene degraders. The main difference between the two degraders was the presence of the third open reading

Analysis of bph Operon 4849
(e) bpk8 vs. 1 0 d D Homology Scare: 893 [59,9%f274aa] (a) bphAl vs. 1odCl Homology Scm:1752 [64,6$/455aa]
b p M 1 HPDFPGNVG-YVYAEEAARGMYHHWMRMMSEPSWATLKP 458 " :.I::: "*:I*."**:* " " : " " " : , * : " : : " " : tOdCl bPVYPGRISNNVYSEEAARGLVAHWLRMMTSPDWDALKATR 450
(C) b p W vs.rodB Homology Score: 339 159.8%B7aa]
12FeZSl binding site b p M 3 VE~SL#MGKFC~RTGKVKSPPPCEALKIFPlRlEDNDVLVDFEAGVLAP 109 ***:""~"******.**": I."" **."*"* **": I ( * t o m VEQTLWFGKFCVRTGKVKALPACKP~KVFPIKVEGDEVHYDLDNGELK 107
(d) bphA4 VI. 10dA Homology Scow: 1122 [52.5%/406aa]
bpha4 MIDTIAIIGAGLAGSTAARALRAQGYEGRIHLLGDESHQAYDRTTLSKTVLAGEQPEPPA 6 0 " : * * * * : * : : * .'.:."*":*;**"* .'".*:I ***::11.:**..: ** to& MATHVAIIGNGVGGFTTAQALRAEGFEGRISLIGDEPHLPYDRPSLSKAVLDGSLERPPI 60
bphA4 ILDSAWYASAHVDVQL?RRVSCt?LAN?Q!QFES?APtAY?RLLL~~?A??!R~?IR?GD : : . * : : * : : * ; 120
t0a LA~ADWYGEARIDMLTGPE;TALDVQTRT~S~~DGTTLSADAIVIATGSRARTMALPG~Q 120
bphA4 tAQIH! t?DLA?S?At :QALQPGQStV! I?~~t !?~~ I?~??!~ tSVH~1! t~??~~t t~ 180
tOa LPGVVTLRTVGDVQVLROSWTSATRLLIVGGGLIGCEVATTARKLGLSVTILEAGDELLV 180
bphA4 RVLGHRTGAWCRAELERMGVRVERNAQAARFEGQGQVRAVICA~?~RVP?~VV~VSI??~ 240
toda RVLGRRIGAWLRGLLTELGVQVELGTGVVGFSGEGQLEQVMASDGRSFVADSALICVGAE 240 ****:*:*** I I): :"":"* , , : : .:.:..: : * :
bphA4 PADELARAAGIACARGVLVDATGATSCPEVFAAGDVAAWPLRQGGQRSLETYLNSQMEAE 300
to& PADQLARQAGLACDRGVIVDHCGATLAKGVFAVGDVASWPLRAGGRRSLETYMNAQRQAA 300
bphA4 lAASAHLSQPVPAPQVPTSWTElAGHRIQMlGDAEGPGEIVYRGDAQSGQPlVLLRLL~G 360 to- AVAAAILGKNVSAPQLPVSWTEIAGHRMQMAGDIEGPGDFVSRGMPGSGAA-LLFRLQER 359
M .'*" * ' * * * " ~ " * * * * " " * " ~ " * .. ""I"::" *I I.,: .I.**
b P M 4 CVE?AT?IN~T:E~SVfL!i:GT~VSVSAEQ~QEVG5NLRDLLKAKPN 408
to& RIQAVVAVDAPRDFALATRLVEARAAIEPARLADLSNSM~~FVR~NEGDLT 410
*..:""":"":""~"""-*. I". *"":1".":"""*:"*:*.*I*I:*:*:~":
pseudoalcaligena KF707 bphAIA2A3APBC genes and P. pu- FIG. 5. Comparison of amino acid sequences coded by P.
tida F1 todCZC2BAIIE genes. For 2,3-dihydroxybiphenyl dioxy- genase coded by bphC gene, comparison is also made with the previ- ously reported P. paucimobilis Q1 bphC (13) and Pseudomonas sp. KKSlO2 bphC (51). Identical and similar amino acids are indicated by asterisks and colons, respectively. Cysteine-histidine pairs sepa- rated by 17 amino residues which are characteristic of the [2Fe-2S] Rieske-type iron-sulfur proteins are indicated by outlined letters for bphAl and bphA3.
Score: 948 [55.2%lL86aSl (0 QlbphC vs. KFlWbphC Homology S c m : 546 [38.7%R69aa]: KFlO7bphC "8. kdE Homology
QI MVAVTELGVLGLTVTNLDAWRSVAAEVAGMEIVDEGEGDR~YL~M~QWHHRIVLMASDS~ 60 bphC : : : :.(I.:*::*::::*MI*: M : : : : : : : : KF707 ~ ~ ! R S ~ C M ~ C A ~ S ~ ~ A A ~ ? S ~ L ! Q K ~ ? L ~ E ~ O T T D N G D L C ~ ? ! ~ ~ ? ~ ~ ? I A I Q ~ ? E V ~ 58
to& I** H S I Q R L G V L G F E V A D V R S ~ R T ~ A ~ T R ~ ~ M ~ E ? S A S E T E ? T F ~ ~ I ~ S ! ? ~ ~ L S ~ S R ~ P A ~ (I...*"* . ** 58
I*: : : : : :
KKSlO2 MSIERLGVLGFAVKDVPAWDHFLTKSVGLMAAGSAGDAALY-RADQRAWRIAVQPGELD 58
01 DLAYLGWRVADPVEFDAMVAKLTAAGISLTVASEAEARERRVLGLAKLADPGGNPTEIFY 120
KF707 ? L A ~ ~ ? ? Y E A D ~ ~ A ~ K ~ K ~ A ? I A ~ T T ? D A S ~ A R R ? ~ ~ T ~ ~ ! T F A ~ ~ ~ ? L P L ~ ! ~ ~ 118 bphC+r*: * : "**'.....*..**.-""I..*::::: "I " " I" ::I.* ". I":"
to& ?YLF?~FEI?~EQGLQEVKES~QAH?~TVKVEGGE~IAK:G~L~L; ISCT?~C?NRVE!~~ 118
KKS~O~DLAYAGLEVDDAAALERMADKLRQAGVAFTRGDEALM~QRKVMGLLCLQDPFGLPLEIYY 118
Ql GPQVDTHKPFHPGRPMYGKFVTGSEGIGHCILRQD?VPA~A~~~-GL~~LRG~VEVHLQL 179
KF707 I j t \ S ~ V r ~ K ~ e L P G A A ~ ~ ? ~ C L ~ ~ E ? ~ ~ ? ~ F ~ R C ~ P ? S ~ K ~ ~ ~ ~ ~ ? D V t ~ ~ ? ~ s ~ ~ ! ? M K M 177
todE ??lEL~ER~~ASPTG~s?IeQ??~??L?~Y~LSIA?VDA?L?~~?KAt?~QtA?V!?WTl 177
KKS~O~GAAEIFHEPFLPSAPVSG-FVTGDQGIGHFVRCVPDTAKAMAFVTEVLGFVLSD~IDIQM 177
bphC I : .#a "* : : *:"*::*:I. :
01 PNGMVAQPYFMHCNERQHSVAFGLGPMEKRINnLHFEYTDLDDLGLAHDlVRARKIDVAL 239
GPDVTVPVYFLHCNERHHTLAI~AFPLPKRIWMFMLEVASLDDVGFAFDRVDADGL-~TS 236 ":,::*::**":**:I**::*: I '.~:"**.";:;:..**:.:*: I.: " '
I "I I" """*** I I " ** """"".." ""1" I "
...* "I""."'.".: I : **..I.*,. . .**. .*.* " " . '
todb GHGLSVTLYFLVCNGRHHSFAFA~~PGSKRLHWFML~ANGMDDVGLAYDKFDAERA-VVM 236
~K~~O~GPETSVPAHFLHCNGRHHTIALAAFPIPKR~HWFMLQANTIDDVGVAFDRLDAAGR-~TS 236
Q;-., QLGKHANDQALTFYCANPS-GWLWEFGUGARKAPSQQEVYTRDIFGHGNEAAGYGMDIPLG 299 "I." . I . ...I .."._I 1.11.""
Q1 QLGKHANDQALTFYCANPS-GWLWEFGUGARKAPSQQEVYTRDIFGHGNEAAGYGMDIPLG 299 bphC *m:W.**: : : a " : : * W - m h:**:*N
W707 TLGRHTNDHMVSFYASTPS-GVEVEVGWSARTVDRSWVVVRHDSPSMWGMKSVRDKAAARNKA 298 .1*1.11*.1:***::**.-. : * h * w * : * * : w : * * **":a: I:****
"11" to& SLGRHTNDHMlSFVGATPS-GFAVEYG~QAREVTRH~S~V~VD~l5 lWG~KFQAPA 291
KKS~O~LLGRHTNDQTLSFVADTPSPMIEVEFGUGPRTVDSSWTVARHSRTAMWGHKSVRGQR 293 "**""** ""I ""*- ." I"" . " &yo; T~~RHTNDHMVSF~ASTPS-GVEVEVGWSARTVDRSWVVVRHDSPSMWGMKSVRDKAAARNKA 298
to& SLGRHTNDHMlSFVGATPS-GFAVEYG~QAREVTRH~S~V~VD~l5 lWG~KFQAPA 291
KKS~O~LLGRHTNDQTLSFVADTPSPMIEVEFGUGPRTVDSSWTVARHSRTAMWGHKSVRGQR 293
.1*1.11*.1:***::**.-. : * h * w * : * * : W : * * **":a: I:****
"11" "**""** ""I ""*- ." I"" . " FIG. 5. continued
frame (ORF3) in bph operon whose counterpart was absent in todCl CZBADE genes.
In order to find out whether this ORF3 is necessary for the specific degradation of biphenyls, a mutant in which ORF3 was site-specifically deleted was constructed. For the con- struction of the mutant, 36-mer oligonucleotide was synthe- sized whose fifteen 5'-nucleotides were designed to bind just downstream of the stop codon (TGA) of ORF3, and similarly fifteen 3'-nucleotides were designed to bind just upstream of the initiation codon (ATG) of ORF3, as shown in Fig. 4. In order to facilitate the identification of correct mutants, the unique XhoI site was introduced between the 5' - and 3'- annealing regions. This primer was annealed to single- stranded DNA of wild-type pKTF18, and polymerization was carried out utilizing dCTP(aS), as described by Eckstein et al. (19-21). One correct mutant was found among 36 arbitrar- ily chosen colonies. Sequencing of the mutant confirmed the site-specific deletion of the entire 420-bp ORF3 starting from the initiation codon ATG at 2598 and ending after the stop codon TCA at 3017. Most importantly, when the colony harboring the A(ORF3) mutation, designated pKTF18del- ORF3, was incubated with biphenyls, the mutant colony turned yellow, due to the production of compound IV in Fig. 1, indicating that the ORF3 is not mandatory for the degra- dation of biphenyls. It is noteworthy that disruption of any functional genes, e.g. by site-specific transposon mutagenesis, abolishes the corresponding enzyme's activity and accumu-
4850 Analysis of bph Operon TABLE I
Comparison of PCB degrader P. pseudoalcaligenes KF707 genes and their products to the corresponding genes and products of toluene degrader of P. putida Fl
P. pseudoalcaligenes KFIOI P. putida FIE Homologyb Enzymes Genes Nucleotides Amino acids
(G + C) residues
Terminal dioxygenase (ISP
Terminal dioxygenase (ISP
Ferredoxin Ferredoxin reductase Dehydrogenase 2,3-Dioxygenase
large subunit)
small subunit)
% bphA 1 62.2 458
bphA2 58.2 213
bphA3 56.3 109 bphA4 69.0 408 bphB 61.1 277 bphC 60.4 298
Genes Nucleotides Amino acids (G + C) residues
% todCl 60.4 450
todC2 58.7 187
todB 57.0 107 to& 63.7 410 todD 60.4 275 todE 58.3 291
Nucleotide Amino acid
% 68 65
64 60
62 60 59 53 62 60 61 55
2,3-Dioxygenase (Ql)' bphC (Ql) 62.2 298 54 39 These data are deduced from the published nucleotide sequence of Zylstra and Gibson (16). With respect to P. pseudoalcaligenes KF707. ' 2,3-Dioxygenase (Ql) is the product of bphC gene which originates from P. paucimobilis Q1 strain (13).
a-
b- -A 1 -A 4
c- "-6 - ,C - -A2
- 0 R F 3 -A 3 d-
1 2 3
FIG. 6. bphABC genes products synthesized in vitro. The fluorography was carried out to enhance the autoradiographic image of 3H-labeled products. Lane I , pUC118 as a control template; lane 2, pKTF1S; lane 3, pKTFlBdelORF3. Molecular mass markers: a, bovine serum albumin (66 kDa); b, ovalbumin (45 kDa); c, carbonic anhy- drase (29 kDa); d , cytochrome c (12.3 kDa).
lates degradation intermediates without producing yellow color (12, 22, 23).
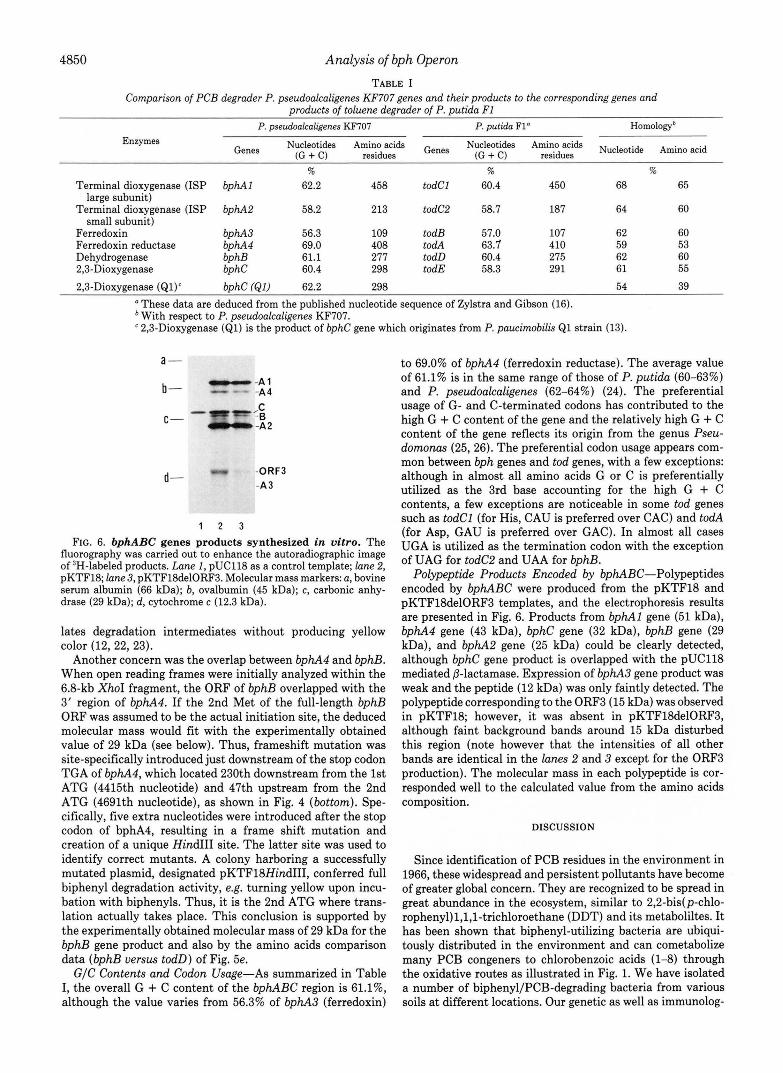
Another concern was the overlap between bphA4 and bphB. When open reading frames were initially analyzed within the 6.8-kb XhoI fragment, the ORF of bphB overlapped with the 3' region of bphA4. If the 2nd Met of the full-length bphB ORF was assumed to be the actual initiation site, the deduced molecular mass would fit with the experimentally obtained value of 29 kDa (see below). Thus, frameshift mutation was site-specifically introduced just downstream of the stop codon TGA of bphA4, which located 230th downstream from the 1st ATG (4415th nucleotide) and 47th upstream from the 2nd ATG (4691th nucleotide), as shown in Fig. 4 (bottom). Spe- cifically, five extra nucleotides were introduced after the stop codon of bphA4, resulting in a frame shift mutation and creation of a unique Hind111 site. The latter site was used to identify correct mutants. A colony harboring a successfully mutated plasmid, designated pKTF18HindII1, conferred full biphenyl degradation activity, e.g. turning yellow upon incu- bation with biphenyls. Thus, it is the 2nd ATG where trans- lation actually takes place. This conclusion is supported by the experimentally obtained molecular mass of 29 kDa for the bphB gene product and also by the amino acids comparison data (bphB versus todD) of Fig. 5e. G/C Contents and Codon Usage-As summarized in Table
I, the overall G + C content of the bphABC region is 61.1%, although the value varies from 56.3% of bphA3 (ferredoxin)
to 69.0% of bphA4 (ferredoxin reductase). The average value of 61.1% is in the same range of those of P. putida (60-63%) and P. pseudoalcaligenes (62-64%) (24). The preferential usage of G- and C-terminated codons has contributed to the high G + C content of the gene and the relatively high G + C content of the gene reflects its origin from the genus Pseu- domonas (25,26). The preferential codon usage appears com- mon between bph genes and tod genes, with a few exceptions: although in almost all amino acids G or C is preferentially utilized as the 3rd base accounting for the high G + C contents, a few exceptions are noticeable in some tod genes such as todCl (for His, CAU is preferred over CAC) and todA (for Asp, GAU is preferred over GAC). In almost all cases UGA is utilized as the termination codon with the exception of UAG for todC2 and UAA for bphB.
Polypeptide Products Encoded by bphABC-Polypeptides encoded by bphABC were produced from the pKTF18 and pKTFlSdelORF3 templates, and the electrophoresis results are presented in Fig. 6. Products from bphAl gene (51 kDa), bphA4 gene (43 kDa), bphC gene (32 kDa), bphB gene (29 kDa), and bphA2 gene (25 kDa) could be clearly detected, although bphC gene product is overlapped with the pUC118 mediated 0-lactamase. Expression of bphA3 gene product was weak and the peptide (12 kDa) was only faintly detected. The polypeptide corresponding to the ORF3 (15 kDa) was observed in pKTF18; however, it was absent in pKTFlSdelORF3, although faint background bands around 15 kDa disturbed this region (note however that the intensities of all other bands are identical in the lanes 2 and 3 except for the ORF3 production). The molecular mass in each polypeptide is cor- responded well to the calculated value from the amino acids composition.
DISCUSSION
Since identification of PCB residues in the environment in 1966, these widespread and persistent pollutants have become of greater global concern. They are recognized to be spread in great abundance in the ecosystem, similar to 2,2-bis(p-chlo- rophenyl)l,l,l-trichloroethane (DDT) and its metaboliltes. It has been shown that biphenyl-utilizing bacteria are ubiqui- tously distributed in the environment and can cometabolize many PCB congeners to chlorobenzoic acids (1-8) through the oxidative routes as illustrated in Fig. 1. We have isolated a number of biphenyl/PCB-degrading bacteria from various soils at different locations. Our genetic as well as immunolog-

Analysis of bph Operon 485 1
FIG. 7. Summary of P. peeudoal- caligenes KF707 bphAZA2A3A4BC operon. The products of bphAl, A2, A3, A4, I?, and bphC are, respectively, ter- minal dioxygenase (large subunit), ter- minal dioxygenase (small subunit), fer- redoxin, ferredoxin reductase, dehydro- genase, and 2,3-dioxygenase. The reduced form of ion-sulfur protein (ter- minal oxygenase), designated as ISP(Ied)BpH (the products of bphAl and AZ), catalyzes the initial addition of both atoms of molecular oxygen to the bi- phenyl ring. The resulting oxidized form of ISP(0x)BpH accepts electrons from fer- redOXin(red)BpH (the product of bphA3) which is a one-electron carrier, and in turn ferredoxin(ox)BpH accepts electrons from a flavoprotein, ferredoxin reduc- tase(red)BpH (the product of bphA4). The original source of electrons is NADH which initially transfers hydride equiva- lency to the flavoprotein, ferredoxin re- ductase(0x)BpH.
ical analyses have allowed us to classify PCB strains into four groups (11): in the first group the bph operons are very similar, if not identical to, that of P. pseudoalcaligenes KF707; the second group containing P. putida KF715 possesses highly homologous bph operon, but their restriction map profiles are different from that of the KF707 operon; the third group shows weak homology with KF707 bph operon; the fourth group, which includes the only one American isolate of P. paucimobilis Q1, shows neither genetic nor immunological homology with the KF707 operon (11-13). The gene organi- zation of the most extensively studied KF707 which belongs to the 1st group is bphABCXD, as shown in Fig. 1, where the function of bphX region is not yet elucidated. This bphX region is missing from the KF715 of the second group. Never- theless, the overall homology of the bphC genes in KF707 (first group) and KF715 (second group) at the level of DNA is as high as 92.4%, and even at the corresponding amino acid level it is 91.4%; however, a significant difference in nucleotide sequence is observed from 30 bp downstream of the bphC gene, reflecting the presence of the extra -3-kb DNA segment (bphX) in KF707 (12). From the sequencingof bphABC region of the KF707 and site-specific deletion of its ORF3, it is now apparent that bphA of the KF707 and most probably the corresponding regions of other groups consist of four genes,
1990)
(Zylslra. G. I. andGibson, D. T.. J.BiaL Chrm. 264,14940-14946. 1989)
bphAlA2A3A4 (27).2 It is particularly interesting to find out that the similarity
of the genetic organization and nucleotide sequences of indi- vidual genes between a biphenyl-catabolic bph operon and a toluene-catabolic tod operon is unexpectedly high. Even though structurally biphenyl is quite distinct from toluene, the enzymes catalyzing the initial incorporation of molecular oxygen in these two aromatic substrates, which have evolved in two different Pseudomonas species, appear to have a com- mon ancestry. P. putida F1 degrades toluene through cis- toluene dihydrodiol to 3-methylcatechol. The chemistry and biochemistry of the dihydrodiol pathway has been studied extensively and elegantly by Gibson and his associates (16, 28-39) in case of toluene dioxygenase. As Fig. 5 shows, the amino acid sequences involved in biphenyl catabolism and the analogous enzymes for toluene catabolism (16) can easily be aligned with each other. In contrast, the degree of align- ment between the enzymes for biphenyl degradation and the counterparts for benzene oxidation (17) is much weaker in- dicating that, since the genes for toluene catabolism seem to be identical to that for benzene catabolism, the sequences for the former genes appear more reliable. Based on the works of
All the sequencing and mutagenesis results presented in this paper are communicated at the 1990 Annual Meeting of the Japanese Society of Bioscience, Biotechnology, and Agrochemistry (27).

4852 Analysis of bph Operon
Gibson's group (16), we can now assign those genes in the bphA region: as shown in Fig. 7, the products of bphA1, A2, A3, A4, B, and bphC could be, respectively, by adapting Gibson's nomenclature: terminal dioxygenase (large subunit), terminal dioxygenase (small subunit), ferredoxin, ferredoxin reductase, dehydrogenase, and 2,3-dioxygenase. Reduced form of ion-sulfur protein (terminal dioxygenase), designated as ISP(red)spH (the products of bphAl and A2), catalyzes the initial addition of both atoms of molecular oxygen to the biphenyl ring. The resulting oxidized form of ISP(OX)B~H accepts electrons from ferredoxin(red)BpH (the product of bphA3) which is a one-electron carrier, and in turn ferre- doxin(ox)BpH accepts electrons from a flavoprotein, ferredoxin reductase(red)BpH (the product of bphA4). The original source of electrons is NADH which initially transfers hydride equiv- alency to the flavoprotein, ferredoxin reductase(ox)BpH. Thus, the multienzyme system of biphenyl/PCB oxygenase consists of a flavoprotein and two iron-sulfur proteins. All the expected polypeptides arising from the bphA1, A2, A3, A4, B, and bphC genes were detected by means of in vitro protein synthesis and the gel-measured molecular weights of each proteins corresponded well to the calculated values from the deduced amino acids composition. Since the terminal dioxygenase (ISPBpH) consists of large and small subunits (products of bphA1 and A2), their expression levels were similarly high.
Many of the multicomponent dioxygenases involved in aromatic degradation contain a Rieske-type iron-sulfur center either associated with the oxygenase itself or as part of a small electron transport protein. In contrast to the plant-type ferredoxins, which have four symmetrically placed cysteine sulfurs coordinating to the [2Fe-2S] core of the center, recent studies unequivocally established that two of the ligands to the Rieske [2Fe-2S] center are histidine nitrogens which coordinate to the ferrous ion site of the spin-coupled [Fez+ (S = 2), Fe3+ (S = 5/2)] pair of the reduced cluster (40, 41). The Rieske proteins contain a highly conserved sequence including two cysteine-histidine pairs separated by 16 or 17 amino acids (42-44). These conserved cysteine-histidine pairs are also present in the bphAl product (large iron-sulfur protein; Cys'"-His'02 and C y ~ ' ~ ~ - H i s ' ~ ~ ; see Figs. 2 and 5) and also in the bphA3 product (small iron-sulfur protein, ferredoxin;
and C y ~ ~ ~ - H i s ~ ~ ) . In both cases the cysteine-his- tidine pairs are separated by 17 amino residues. These cys- teine-histidine pairs separated by 17 amino acids are also found in the corresponding genes (todC1 and todB) of tod operon (see the discussion below), and the EPR spectra of the reduced ferredoxin(red)ToL (product of todB) revealed that its g,, value is more characteristic of Rieske proteins than the majority of [2Fe-2S] ferredoxins (29). In these enzyme sys- tems, the [2Fe-2S] center of ISP large subunit (product of bphA1) probably mediates electron transfer from the ferre- doxin [2Fe-2S] center (product of bphA3) to a mononuclear Fez+ at the oxygenation site, a role analogous to that in mitochondria. Although bphAl also contains the sequence - Gly268-X-Gly270-X-X-Gly273-, which is characteristic of Pa@ structures responsible for the binding of FAD or NAD (451, the score of the fingerprint region is 5. Moreover, the corre- sponding G1y268 is missing from the todC1, indicating that this sequence is just coincidental.
To our surprise the entire gene organization of the bphABC region as well as the homology of their products (- 60% as summarized in Table I), including their molecular sizes, are highly conserved among bph and tod operons. Zylstra and Gibson (16) also realized the significant homology between the amino acid sequences of 3-methylcatechol2,3-dioxygenase from P. putida F1 and 2,3-dihydroxybiphenyl1,2-dioxygenase
from P. pseudoalcaligenes KF707. There is no doubt that these two operons have arisen from a common evolutionary ances- tor. There are, however, minor differences between these two operons. Upstream of both bphAl and todC1, there appear to be putative open reading frames; 1-405 region for bph (Fig, 2) and 1-520 region for tod (16). The homology between these putative genes at the level of nucleotide is 51% which scores less than the other identified corresponding genes (Table I). Moreover, the translated amino acids of these open reading frames do not show any significant homology. Zylstra and Gibson (16) tentatively assigned the upstream open reading frame of tod operon as todF based on the polar effect on gene expression caused by Tn5 mutagenesis (28). The homology between todF and bphD of KF715 (12), both coding for hydrolase, are 52 and 35% at the level of nucleotide and amino acid, respectively, which are significantly lower than the other pairs in Table I. Moreover, in the case of KF707 and KF715, the bphD gene locates downstream from the bphC gene. Thus, the gene organizations of bph and tod operons outside of those depicted at the bottom of Fig. 7 (bphABC region) are less similar. In support of this conclusion, a transcriptional initi- ation site was identified just upstream of bphA1 (Fig. 3), although sequences homologous to known Pseudomonas pro- moters (46) and translation terminators typical of those found in enteric bacteria (47, 48) were not detected. The presence of the transcriptional initiation site upstream of bphA1 con- trasts to the absence of such initiation site upstream of the corresponding todC1 gene, reflecting the existence of todF upstream of todC1 in case of the tod operon.
Since bph and tod proteins appear to share a common ancestry, the identification of their nucleotide sequences at various dioxygenase levels and their comparison should pro- vide important information on substrate specificity. It is of particular interest that, notwithstanding the apparent enzy- matic similarities of 2,3-dihydroxybiphenyl dioxygenase (product of bphC) of P. pseudoalcaligenes KF707 and P. paucimobilis Q1 such that both enzymes (i) exhibit a native molecular mass of 260 kDa, (ii) contain ferrous form of ion, (iii) are composed of eight identical subunits, (iv) are inhibited by the ferric form of ion, and (v) cannot catalyze the oxidative cleavage of the substrate's positional isomer, 3,4-dihydroxy- biphenyl (13), the homology at the level of amino acids between bphC(KF707) and bphC(Q1) is much lower (39%, see Fig. Sf) than the corresponding value of 55% between bphC(KF707) and todE. Catalytically indispensable amino residues must be highly conserved in these sequen- ces, and the difference in substrate specificity between bphC(KF707) and todE must be hidden in the nonconserved region (note that the 2,3-dihydroxybiphenyl dioxygenase coded by bphC(KF707) does not metabolize benzene deriva- tives which are smaller in sizes than biphenyl derivatives). In fact, a site-specific mutagenesis of the strictly conserved Hiszo9 (KF707 bphC numbering) to alanine completely abolished the 2,3-dihydroxybiphenyl dioxygenase activity, implying that this site is a candidate for the Fe(I1) binding site. On the other hand, mutagenesis of the other nonconserved histidines such as Hiszw and Hisza4 to alanine is inert to the 2,3- dihydroxybiphenyl dioxygenase a~t iv i ty .~
It was possible to isolate many chromosomally encoded bph genes possessing different degrees of homology. Thus, there must be a mechanism governing bph gene-shuffling among various species which probably operates on tod genes as well, because they all possess high similarity and all are encoded on chromosomes. To our surprise, despite the discrete sub- strate specificities between the PCB degraders and benzene/
K. Taira and K. Furukawa, unpublished data.

Analysis of bph Operon 4853
toluene degraders, bph and tod operons are very similar not 23. Furukawa, K., Hayashida, S., and Taira, K. (1991) Gene (Amst. ) only in gene organizations but also the sizes and sequences of 98,21-28 the deduced amino acids, especially in the the regions coding 24. Normore, W. M. (1976) in Handbook of Biochemistry and Molec-
for the initial oxidation steps (bphABC). It is thus of signifi- ulur Biology (Fasman, G. D., ed) 3rd Ed., pp. 65-235, The Chemical Rubber Co., Cleveland, OH
cant value to check which comPonent(s) is (are) mutually 25. Nakai, C., Kagamiyama, H., Nozaki, M., Nakazawa, T., Inoue, replaceable among bph and tod gene products. At least todD S., Ebina, Y., and Nakazawa, A. (1983) J. Biol. Chem. 258, product (dehydrogenase) can utilize biphenyl dihydrodiol (I1 2923-2928 in Fig. 1) as a substrate (32). It would be more attractive to 26. Brown, N. L., Ford, S. J., Pridmore, R. D., and Fritzinger, D. c.
ilarity of the bph and tod genes, and manipulate substrate Annual Meeting of the Japanese Society of Bioscience, Biotech- specificities. rwlogy, and Agrochemistry, March, Fukuokn, Japan, Abstract,
construct various chimeric because Of the high sim- 27. Taira, K., Makuura, A., Kokean, y., and Furukawa, K. (1990) (1983) Biochemistry 22,4089-4095
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18. 19.
20.
21.
22.
REFERENCES
Ahmed, M., and Focht, D. D. (1973) Can. J . Microbiology l9,47-
Furukawa, K., Matsumura, F., and Tonomura, K. (1978) Agric.
Yagi, O., and Sudo, R. (1980) J. Water Pollut. Control Fed. 52,
Furukawa, K. (1982) in Biodegradation and Detoxification of Environmental Pollutants (Chakrabarty, A. M., ed) pp. 33-57, CRC Press, Boca Raton, FL
Furukawa, K., and Chakrabarty, A. M. (1982) Appl. Enuiron. Microbiol. 44,619-626
Furukawa, K., Simon, J. R., and Chakrabarty, A. M. (1983) J. Bacteriol. 154,1356-1362
Bedard, D. L., Unterman, R., Bopp, L. H., Brennan, M. J., Haberl, M. L., and Johnson, C. (1986) Appl. Environ. Microbiol. 51,
Bedard, D. L., Wagner, R. E., Brennan, M. J., Haberl, M. L., and Brown, J. F., Jr. (1987) Appl. Environ. Microbiol. 53, 1094- 1102
Ruisinger, S., Klages, U., and Lingens, F. (1976) Arch. Microbiol. 110,253-256
Bedard, D. L., Brennan, M. J., and Unterman, R. (1984) Proceed- ings of the 1983 PCB Seminar (Addis, G., and Komai, R., eda) Electrical Power Research Institute, Palo Alto, CA
Furukawa, K., Hayase, N., Taira, K., and Tomizuka, N. (1989) J. Bacteriol. 171,5467-5472
Hayase, N., Taira, K., and Furukawa, K. (1990) J. Bacteriol. 172,
Taira, K., Hayase, N., Arimura, N., Yamashita, S., Miyazaki, T.,
Furukawa, K., and Miyazaki, T. (1986) J. Bacteriol. 166, 392-
Furukawa, K., Arimura, N., and Miyazaki, T. (1987) J. Bacteriol.
Zylstra, G. J., and Gibson, D. T. (1989) J. Biol. Chem. 264,
Irie, S., Doi, S., Yorifuji, T., Takagi, M., and Yano, K. (1987) J.
Vieira, J., and Messing J. (1987) Methods Enzymol. 153, 3-11 Taylor, J. W., Schmidt, W., Cosstick, R., Okruszek, A., and
Taylor, J. W., Ott, J., and Eckstein, F. (1985) Nucleic Acids Res.
Nakamaye, K. L., and Eckstein, F. (1986) Nucleic Acids Res. 14,
Furukawa, K., and Suzuki, H. (1989) Agric. Biol. Chem. 42,543-
52
Biol. Chem. 42,543-548
1035-1043
761-768
1160-1164
and Furukawa, K. (1988) Biochemistry 27,3990-3996
398
169,427-429
14940-14946
Bacteriol. 169,5174-5179
Eckstein, F. (1985) Nucleic Acids Res. 13,8749-8764
13,8765-8785
9679-9698
548
p. 496 28. Zylstra, G. J., McCombie, W. R., Gibson, D. T., and Finette, B.
A, (1988) Appl. Enuiron. Microbiol. 54,1498-1503 29. Subramanian, V., Liu, T.-N., Yeh, W.-K., Serdar, C. M., Wackett,
L. P., and Gibson, D. T. (1985) J. Biol. Chem. 260,2355-2363 30. Subramanian, V., Liu, T.-N., Yeh, W.-K., Narro, M., and Gibson,
D. T. (1981) J. Biol. Chem. 256,2723-2730 31. Subramanian, V., Liu, T.-N., Yeh, W.-K., and Gibson, D. T.
(1979) Biochem. Biophys. Res. Commun. 91,1131-1139 32. Rogers, J. E., and Gibson, D. T. (1977) J. Bacteriol. 130, 1117-
1124 33. Yeh, W.-K., Gibson, D. T., and Liu, T.-N. (1977) Biochem.
Biophys. Res. Commun. 78,401-410 34. Ziffer, H., Jerina, D. M., Gibson, D. T., and Kobal, V. M. (1973)
J. Am. Chem. SOC. 95,4048-4049 35. Kobal, V. M., Gibson, D. T., Davis, R. E., and Garza, A. (1973)
J. Am. Chem. SOC. 95,4420-4421 36. Gibson, D. T., Hensley, M., Yoshioka, H., and Mabry, T. J. (1970)
Biochemistry 9,1626-1630 37. Gibson, D. T., Cardini, G. E., Maseles, F. C., and Kallio, R. E.
(1970) Biochemistry 9, 1631-1635 38. Haigler, B. E., and Gibson, D. T. (1990) J. Bacteriol. 172,457-
464 39. Haigler, B. E., and Gibson, D. T. (1990) J. Bacteriol. 172, 465-
468 40. Gurbiel, R. J., Batie, C. J., Sivaraja, M., True, A. E., Fee, J. A.,
Hoffman, B. M., and Ballou, D. P. (1989) Biochemistry 28,
41. Tsang, H.-T., Batie, C. J., Ballou, D. P., and Penner-Hahn, J. E. (1989) Biochemistry 28, 7233-7240
42. Fee, J. A., Findling, K. L., Yoshida, T., Hille, R., Tarr, G. E., Hearshen, D. O., Dunham, W. R., Day, E. P., Kent, T. A., and Munck, E. (1984) J. Biol. Chem. 259, 124-133
43. Nordlund, I, Powlowski, J., and Shingler, V. (1990) J. Bacteriol.
44. Powlowski, J., and Shingler, V. (1990) J. Bacteriol. 172, 6834-
45. Wierenga, R. K., Terpstra, P., and Hol, W. G. J. (1986) J. Mol.
46. Deretic, V., Gill, J. F., and Chakrabarty, A. M. (1987) J. Biotech-
47. Adhya, S., and Gottesman, M. (1978) Annu. Rev. Biochem. 47,
48. Platt, T. (1986) Annu. Rev. Biochem. 55,339-372 49. Bonner, W. M., and Laskey, R. A. (1974) Eur. J . Biochem. 46,
50. Laskey, R. A., and Mills, A. D. (1975) Eur. J . Biochem. 56,335-
51. Kimbara, K., Hashimoto, T., Fukuda, M., Koana, T., Takagi, M.,
4861-4871
172,6826-6833
6840
Biol. 187,101-107
ml. 5,469-477
967-996
83-88
341
Oishi, M., and Yano, K. (1989) J. Bacteriol. 171, 2740-2747