channel in the mouse thick ascending limb at low internal ph
TRANSCRIPT

Journal of Physiology (1995), 490.3, pp.759-765
Inhibition of a small-conductance cAMP-dependent C1-channel in the mouse thick ascending limb at low internal pH
Romain Guinamard, Marc Paulais and Jacques Teulon *
INSERM Unite 323, Faculte de Medecine Necker Enfants-Malades, 156 rue de Vaugirard,75730 Paris Cedex 15, France
1. A small-conductance Cl- channel that is stimulated by ATP and protein kinase A has beenidentified in the basolateral membranes of cortical thick ascending limbs (CTALs) of themouse nephron. The present study uses the cell-attached and inside-out variants of thepatch-clamp technique to investigate the pH sensitivity of this channel.
2. The open-state probability (P.) was dependent upon the internal pH in inside-out patches.Expressed as a percentage of the P. value at pH 7-2, P. increased to about 180% at pH 7 6,and decreased to 25% at pH 6 8. P0 was close to zero at pH 6-4. The internal pH had no
effect on the channel unit conductance.
3. The effect of pH on the CTAL Cl- channel was assessed in intact cells using NH4Cl toacidify the intracellular compartment. Experiments with the pH-sensitive fluorescent dye2',7'-(carboxyethyl)-5'(6')-carboxy fluorescein penta-acetoxymethyl ester (BCECF) indicatedthat 1 mmol 1-' NH4Cl acidified the cytoplasm by 0 15 pH units and 5 mmol F' NH4Cl by0 34 pH units. These concentrations of NH4Cl reduced the activity of the CTAL Cl-channel by 24 and 82% in cell-attached patches, showing that moderate changes in internalpH substantially altered the activity of this channel. NH4+ had no direct effect on channelactivity.
4. Inhibition at low pH is a newly discovered property of small-conductance Cl- channels inepithelia, which might help discriminate between types of Cl- channel.
The cortical thick ascending limb (CTAL) of the loop ofHenle is a part of the distal nephron in which NaCl isabsorbed via apical Na+-K+-Cl- cotransport, basolateralNa+-K+-ATPase and Cl diffusion across the basolateralmembrane (see Reeves & Andreoli, 1992). We recentlydescribed (Guinamard, Chrafbi & Teulon, 1995) a small-conductance Cl- channel with properties that areconsiderably different from those of previously identifiedCl channels in the same tubule (for reviews see Paulais &Teulon, 1990; Schwiebert, Lopes & Guggino, 1994). Thischannel probably dominates Cl- conductance in the basalstate; in accordance with the properties of the macroscopicCl- conductance of the basolateral membrane (see Reeves &Andreoli, 1992), it is stimulated by the cAMP transductionpathway and inhibited by 5-nitro-2-(3-phenylpropyl-amino)benzoic acid (NPPB). The CTAL Cl- channel also hasseveral properties of cystic fibrosis transmembraneconductance regulator (CFTR), including unit conductance(7-9 pS), inhibition by diphenylamine-2-carboxylic acid(DPC) at high concentration (> 1 mmol l-') andglibenclamide, activation by ATP and activation by thecatalytic subunit of protein kinase A (for review see Welsh
et al. 1992). However, little is known, as yet, of itsmolecular nature. It is possible that the CTAL channelfunctionally resembles CFTR while being a distinct entity.
This study shows that the CTAL Cl- channel is verysensitive to internal pH (pH,). This property may beimportant physiologically for NaCl reabsorption across theCTAL epithelium.
METHODSTubule isolationMale mice (15-20 g) were killed by cervical dislocation. Corticalfragments of CTAL were isolated from the kidneys by treatmentwith collagenase (Type CLS II; Worthington, Freehold, NJ, USA)and microdissection, as previously described (Paulais & Teulon,1990; Guinamard et al. 1995).
SolutionsIn the cell-attached configuration, the bath or standard Ringersolution contained (mmol F-): 140 NaCl, 4-8 KCI, 1P2 MgCl2, 1CaCl2, 10 glucose, 10 Hepes adjusted to pH 7-4 with NaOH. Thesame solution was used in the pipette, except for some experimentswhere NaCl was replaced by equimolar N-methyl-D-glucamine
* To whom correspondence should be addressed.
4549 759

R. Guinamard, M. Paulais and J Teulon
chloride or sodium gluconate. Low-NaCl (14 mmol 1-' withnominal [CaCl2] of 10-9 mol F') bath solutions were used for theexperiments with excised inside-out patches (mmol F'): 14 NaCl,1P2 MgCl,2 10 glucose, 200 sucrose, 2 EGTA. Solutions werebuffered with Hepes (pH 6'8-8 0) or 2-(N-morpholino)-ethanesulphonic acid (Mes; pH 6 0-6'6) at a concentration of10 mmol l-l. NPPB was a gift from Dr R. Greger (Freiburg,Germany). Unless otherwise stated, chemicals were obtained fromSigma.
Experiments were conducted at room temperature (20-25 °C) or at37 °C (when specified) on tubules preincubated for at least 15 minwith the adenylyl cyclase activator forskolin (10 ,umol 1-').Experiments at 37°C used a microchamber (Open PerfusionMicro-incubator) coupled to a cooler/heater with a temperaturecontroller (TC102; Medical Systems Corporation, Greenvale, NY,USA).
Current recordingsSingle-channel currents (i) were recorded with a patch-clampamplifier (LM-EPC 7; List Electronic, Darmstadt, Germany) frompatches of basolateral membranes using the cell-attached andinside-out variants of the patch-clamp technique (Hamill, Marty,Neher, Sakmann & Sigworth, 1981). The membrane potential (Vm),expressed as the potential at the inner side of the membranerelative to the pipette medium, is the sum of the spontaneousmembrane potential (Em) and of the clamp potential(VC = -pipette potential). Vm reduces to Vc in the excised, inside-out configuration. Anion currents from the outer to the inner sideof the membrane patch are given a positive sign and are shown inan upwards direction. The bath reference electrode was a0 5 mol 1-' KCl (in 4% agar) bridge connected to an Ag-AgCl half-cell.
Signals were filtered at 500 Hz with an 8-pole Bessel filter(Frequency Devices, Haverhill, MA, USA) and digitized at 1 kHzusing a Labmaster analog-to-digital interface and Fetchexsoftware (Axon Instruments, Foster City, CA, USA). Single-channel current recordings were then analysed with softwarekindly provided by Thierry Van Den Abbeele (Facultg deMedecine Lariboisiere, Paris, France). The mean current (I)passing through n channels in the patch, estimated from currentamplitude histograms, was used to calculate channel activity (nPO)or open-state probability (PO) using the equation I= nPoi.Throughout this paper PO at a given pH is expressed as apercentage of the control PO at pH 7'2.Measurement of intracellular pHIsolated CTALs were loaded with the pH-sensitive fluorescentprobe 2',7'-(carboxyethyl)-5'(6')-carboxy fluorescein penta-acetoxymethyl ester (BCECF AM; Calbiochem, San Diego, CA,USA) in standard medium containing 4 ,umol 1-' BCECF AM and0'1% bovine serum albumin (BSA) for 1 h at room temperature.The loaded tubules were then rinsed in BCECF AM- and BSA-freestandard Ringer solution and allowed to rest for at least 20 min.
BCECF fluorescence was monitored in ratio mode using amicrofluorometer (FilterscanTm RF-F3010; Photon TechnologyInternational Inc, South Brunswick, NJ, USA) equipped with442 + 10 and 492 + 10 nm interference filters (Optometrics Corp,Ayer, MA, USA) and connected to a Diaphot microscope (Nikon,Tokyo, Japan). One CTAL tubule was illuminated through aNikon x 40 Fluor objective lens (numerical aperture, 0'85) coupledto a dichroic mirror (Nikon DM400) and the light emitted at531 + 10 nm was detected by a photomultiplier tube (Oriel Corp,Stratford, CT, USA) connected to the side port of the microscope.
490/440 ratios were converted to pH, by the method of Thomas,Buchsbaum, Zimniak & Racker (1979). At the end of eachmeasurement, an in situ calibration was done using a high-K+solution containing 10 mol F' nigericin. The high-K+ solutioncontained (mmol F-'): 150 KCl, 1'2 MgCl2, 1 CaC12, 10 D-glucose,10 Hepes and was adjusted to pH 6-6-7'7 with tris-(hydroxymethyl) amino ethane (Trizma base). Autofluorescence,measured from unloaded tubules, accounted for about 2% of theBCECF signal and was therefore negligible.
Experimental values are given as means + S.E.M., n denotes thenumber of results. Student's t test was used to analyse unpairedresults, P < 0'05 was considered significant.
RESULTSEffects of pH on the small-conductance C1- channelin inside-out patchesFigure 1 shows the effects of internal pH on the activity ofthe small-conductance CF- channel recorded in excised,inside-out patches at room temperature (20-25 C). Thebath solution consisted of a low-NaCl (14 mmol 1-' NaCl,10-9 mol 1-' CaCl2) Ringer solution and the voltage was setto 50 mV, close to the equilibrium potential for cations.Under this condition, only Cl- channel openings aredetected and reasonably large currents (about 1 pA) aregenerated across the small-conductance Cl- channel. Thecurrent recording shown in Fig. 1A was obtained from oneinside-out patch containing thirteen small-conductance Cl-channels and two channels with a higher conductance.Openings of these latter channels are seen as large briefdeflections in Fig. 1B, where excerpts of the recordingshown in Fig. 1A can be seen on an expanded time scale.These openings probably correspond to a Cl- channel thatis not modulated by internal pH (Paulais & Teulon, 1990).Inspection of the recordings in Fig. 1A and B indicates thatthe small-conductance Cl- channel was very sensitive tointernal pH: channel activity, expressed as a percentage ofPO at pH 7'2, was close to zero at pH 6 4, increased to 32%at pH 6 8 and 194% at pH 7'6. Most channels were open atpH 8'0 (P., 256 %). The results obtained from four to sevenpatches are summarized in Fig. 2. The channel was verysensitive to pH at physiological values: P. values at pH 6'8and 7'6 were 25 + 5 and 185 + 14%, respectively. Sincethe activity of the small-conductance Cl- channel declinedspontaneously in excised patches, similar experiments weredone with 0 5 mmol F' ATP in the bath to stabilize channelactivity (Guinamard et at. 1995). The curves obtained withand without ATP were essentially similar, except at pH 8'0(Fig. 2). The larger PO increase observed at pH 8'0 withATP (305 + 70 %, n = 7) compared with no ATP(197 + 20%, n = 6) (P< 0 05) in the bath is puzzling, butwas not investigated further.
We checked that the effects of internal pH documented at20-25 °C were also observed at a physiological temperature(37 °C) in an independent series of experiments. At thistemperature, channel activity disappeared very rapidlyupon excision, even in the presence of ATP, precluding any
J. Physiol.490.3760

pH-sensitive small-conductance Cl- channel
B 2 pA
2 s
a b c
Figure 1. Effects of internal pH on the activity of the small-conductance Cl- channel frommouse CTAL in inside-out patchesA, current recording from one inside-out patch containing 13 small-conductance Cl- channels. Openingsof a Cl- channel with a larger conductance are seen as large brief deflections (asterisks, see also Fig. 1B),which are probably due to another Cl- channel (Paulais & Teulon, 1990). The internal pH was changedfrom 6-4 to 8-0 in 0 4 pH unit steps as shown below the trace. The pipette contained standard Ringersolution (140 mmol Fl NaCl), while the bath contained a low-NaCl solution (14 mmol F1 NaCl) devoid ofATP. The dashed line indicates the closed current level. Membrane potential, 50 mV. B, excerpts ofcurrent recording from the same inside-out patch are shown on an expanded time scale. Letters indicatethe position of the excerpt within the continuous recording shown in A; asterisks indicate openings of thelarger channel.
400 -
Figure 2. Mean effects of internal pH (pHJ) on the small- 300conductance CO- channelThe mean open probabilities (P.), expressed as a percentage of P0 2
Cat pH 7-2, are plotted against the pH1. Since the small- 0
conductance Cl- channel is subject to channel run-down in excised 200 -
patches, experimental series were done in the absence (0) andpresence ofATP (0 5 mmol F1, *) to stabilize channel activity. a? ru l
Measurements were obtained from 4-7 patches except at pH 7-6without ATP (n = 3). 100-
pHj
J Physiol.490.3 761
R. Guinamard, M. Paulais and J Teulon
i (pA)
-80 -40 0 40 80Vm (mV)
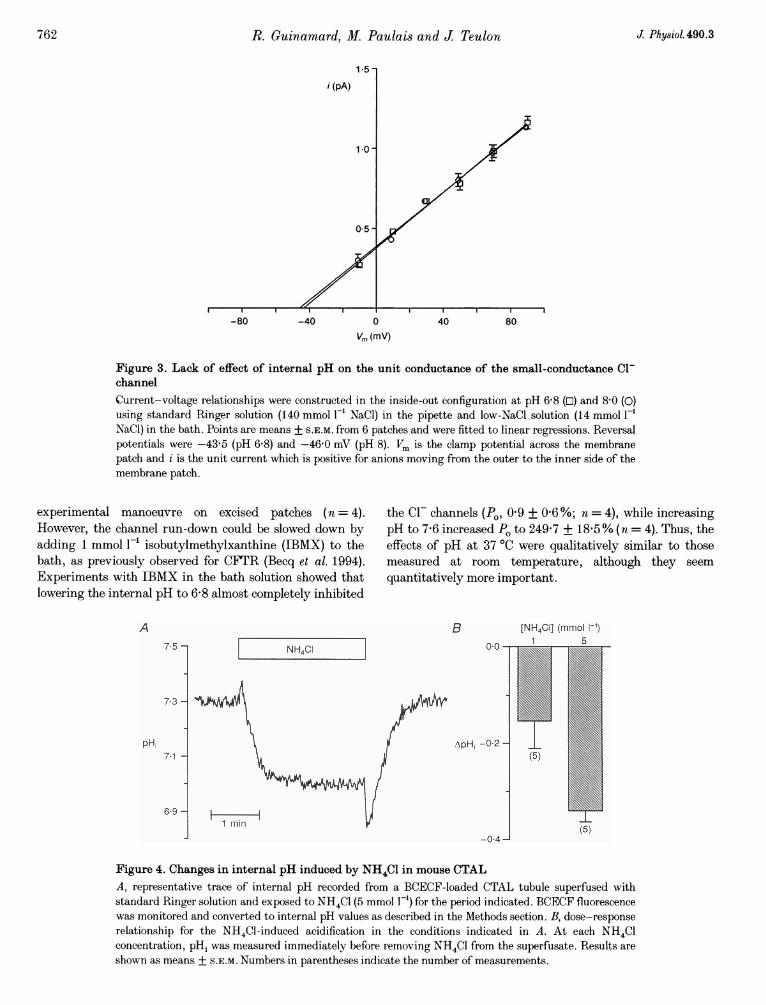
Figure 3. Lack of effect of internal pH on the unit conductance of the small-conductance Cl-channelCurrent-voltage relationships were constructed in the inside-out configuration at pH 6-8 (El) and 8.0 (O)using standard Ringer solution (140 mmol F' NaCl) in the pipette and low-NaCl solution (14 mmol F-'NaCl) in the bath. Points are means + S.E.M. from 6 patches and were fitted to linear regressions. Reversalpotentials were -43-5 (pH 6 8) and -46-0 mV (pH 8). Vm is the clamp potential across the membranepatch and i is the unit current which is positive for anions moving from the outer to the inner side of themembrane patch.
experimental manoeuvre on excised patches (n = 4).However, the channel run-down could be slowed down byadding 1 mmol F' isobutylmethylxanthine (IBMX) to thebath, as previously observed for CFTR (Becq et al. 1994).Experiments with IBMX in the bath solution showed thatlowering the internal pH to 6-8 almost completely inhibited
A7-5 -
73 -
pH,7-1 -
69 -
the Cl- channels (P., 0 9 + 0-6 %; n = 4), while increasingpH to 7-6 increased P0 to 249-7 + 18-5% (n = 4). Thus, theeffects of pH at 37 °C were qualitatively similar to thosemeasured at room temperature, although they seemquantitatively more important.
B0o0-
ApH, -02 -
1 min
-0-4-
[NH4CI] (mmol l-1)1 5
(5)
(5)
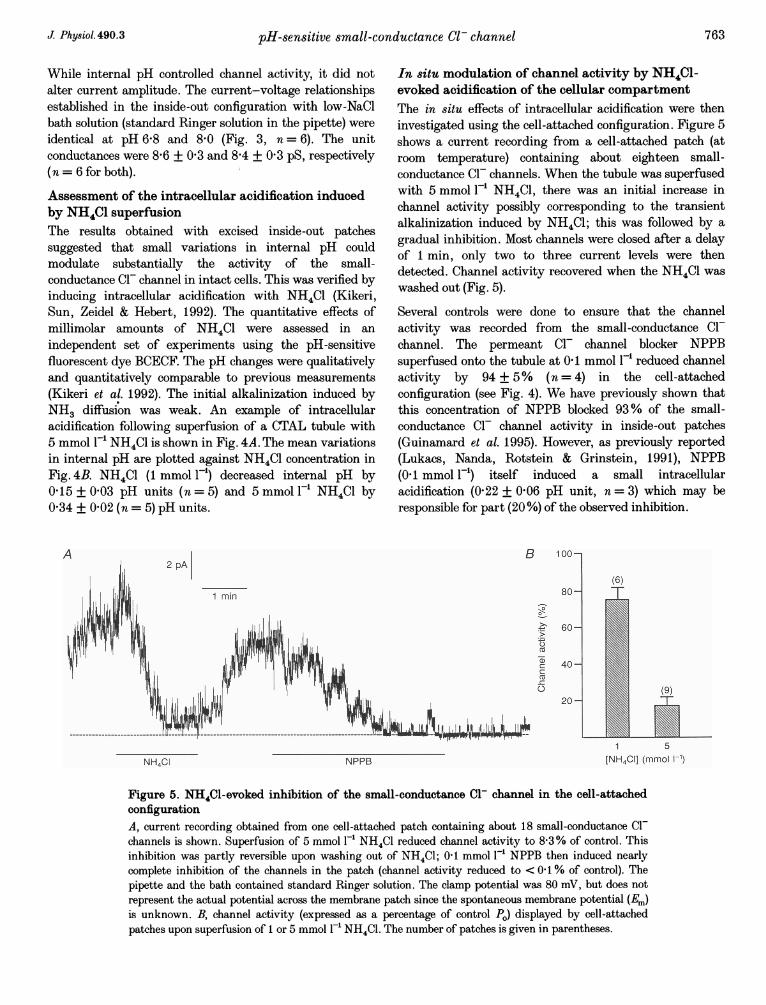
Figure 4. Changes in internal pH induced by NH4Cl in mouse CTALA, representative trace of internal pH recorded from a BCECF-loaded CTAL tubule superfused withstandard Ringer solution and exposed to NH4Cl (5 mmol F-') for the period indicated. BCECF fluorescencewas monitored and converted to internal pH values as described in the Methods section. B, dose-responserelationship for the NH4Cl-induced acidification in the conditions indicated in A. At each NH4C1concentration, pHi was measured immediately before removing NH4C1 from the superfusate. Results areshown as means + S.E.M. Numbers in parentheses indicate the number of measurements.
762 J Physiol. 490.3
pH-sensitive small-conductance Cl- channel
While internal pH controlled channel activity, it did notalter current amplitude. The current-voltage relationshipsestablished in the inside-out configuration with low-NaClbath solution (standard Ringer solution in the pipette) wereidentical at pH 6-8 and 8-0 (Fig. 3, n = 6). The unitconductances were 8-6 + 03 and 8-4 + 03 pS, respectively(n = 6 for both).
Assessment of the intracellular acidification inducedby NH4Cl superfusionThe results obtained with excised inside-out patchessuggested that small variations in internal pH couldmodulate substantially the activity of the small-conductance Cl- channel in intact cells. This was verified byinducing intracellular acidification with NH4Cl (Kikeri,Sun, Zeidel & Hebert, 1992). The quantitative effects ofmillimolar amounts of NH4Cl were assessed in anindependent set of experiments using the pH-sensitivefluorescent dye BCECF. The pH changes were qualitativelyand quantitatively comparable to previous measurements(Kikeri et al. 1992). The initial alkalinization induced byNH3 diffusion was weak. An example of intracellularacidification following superfusion of a CTAL tubule with5 mmol 1-' NH4Cl is shown in Fig. 4A. The mean variationsin internal pH are plotted against NH4Cl concentration inFig. 4B. NH4Cl (1 mmol F-l) decreased internal pH by0.15 + 0-03 pH units (n =5) and 5 mmol -' NH4Cl by0 34 + 0-02 (n = 5) pH units.
A
In situ modulation of channel activity by NH4Cl-evoked acidification of the cellular compartmentThe in situ effects of intracellular acidification were theninvestigated using the cell-attached configuration. Figure 5shows a current recording from a cell-attached patch (atroom temperature) containing about eighteen small-conductance Cl- channels. When the tubule was superfusedwith 5 mmol 1-' NH4Cl, there was an initial increase inchannel activity possibly corresponding to the transientalkalinization induced by NH4Cl; this was followed by agradual inhibition. Most channels were closed after a delayof 1 min, only two to three current levels were thendetected. Channel activity recovered when the NH4Cl waswashed out (Fig. 5).
Several controls were done to ensure that the channelactivity was recorded from the small-conductance Cl-channel. The permeant Cl- channel blocker NPPBsuperfused onto the tubule at 0 1 mmol 1-' reduced channelactivity by 94 + 5% (n =4) in the cell-attachedconfiguration (see Fig. 4). We have previously shown thatthis concentration of NPPB blocked 93% of the small-conductance Cl- channel activity in inside-out patches(Guinamard et al. 1995). However, as previously reported(Lukacs, Nanda, Rotstein & Grinstein, 1991), NPPB(0 1 mmol F-) itself induced a small intracellularacidification (0-22 + 0-06 pH unit, n = 3) which may beresponsible for part (20%) of the observed inhibition.
B 100-2 pA
1 min
.2:U
C'3
C5
c'c)
80-
60-
40-
20-
(6)
(9)
1 5
[NH4CI] (mmol 1-1)NH4Cm NPPB
Figure 5. NH4Cl-evoked inhibition of the small-conductance Cl- channel in the cell-attachedconfigurationA, current recording obtained from one cell-attached patch containing about 18 small-conductance Cl-channels is shown. Superfusion of 5 mmol F' NH4Cl reduced channel activity to 8-3% of control. Thisinhibition was partly reversible upon washing out of NH4Cl; 0.1 mmol F' NPPB then induced nearlycomplete inhibition of the channels in the patch (channel activity reduced to < 01% of control). Thepipette and the bath contained standard Ringer solution. The clamp potential was 80 mV, but does notrepresent the actual potential across the membrane patch since the spontaneous membrane potential (Em)is unknown. B, channel activity (expressed as a percentage of control P.) displayed by cell-attachedpatches upon superfusion of 1 or 5 mmol F' NH4Cl. The number of patches is given in parentheses.
763J. Physiol.490.3

R. Guinamard, M. Paulais and J Teulon
Other experiments were performed to determinequalitatively the ionic selectivity of the channel beingexamined in the cell-attached configuration. When pipetteNaCl was replaced by 140 mmol F' N-methyl-D-glucamine(NMDG) chloride, the current-voltage relationship had aunit conductance (g) of 7-8 + 0-2 pS (n = 5) and a reversalpotential (Er) of -0-8 + 1P9 mV (n = 5). These values arealmost identical to those obtained with 140 mmol 1-' NaClin the pipette (g, 7-2 pS; Er, -0 5 mV; Guinamard et al.1995), showing that NMDG did not alter the channelconductive properties. When 70 mmol 1-' NaCl wasreplaced by sodium gluconate in the pipette solution, Ershifted to 18-9 + 0-8 mV (n = 5), a value close to thatobtained under the same conditions in the previous study(Guinamard et al. 1995). These results suggest that the pH-sensitive channel recorded in the cell-attached mode is thepreviously described small-conductance Cl- channel.
The results for the NH4Cl-induced inhibition of thechannel, obtained at room temperature, are summarized inFig. 5B: 1 mmol 1-' NH4Cl, gave a mean intracellularacidification of 0.15 pH unit (see Fig. 4) and reducedchannel activity (nPi) to 76 + 7 % of control (n = 6) while5 mmol 1-' NH4Cl reduced channel activity to 18 + 5%(n = 9) and acidified the intracellular compartment by0'34 pH unit (see Fig. 4). Another set of experiments at37 °C gave comparable results, perfusion of 5 mmol 1-'NH4Cl onto the tubule surface reduced Cl- channel activityto 13-6 + 4 2% of control (n = 3).
The effect of NH4Cl may be attributed to intracellularacidification, since NH4Cl (5 mmol F-) did not directlyinhibit the small-conductance chloride channel in inside-outpatches (n = 4, data not shown).
DISCUSSIONThe small-conductance Cl- channel in the basolateralmembrane of the mouse thick ascending limb is inhibited atlow pH, unlike the similarly located 45 pS Cl- channel(Paulais & Teulon, 1990). This is of physiological interest,since Wingo (1986) reported that acute in vitro acidosisdecreases Cl- reabsorption across the rabbit CTAL by60-70% and suggested that this inhibition was mediatedby changes in cytosolic pH. The author concluded thatacidosis probably decreases the K+ conductance in theluminal membrane, thus reducing K+ recycling across theluminal membrane and Na+-K+-Cl- cotransport rate (seeReeves & Andreoli, 1992). It was also suggested that theacidosis-evoked inhibition had another, independentmechanism. Since then, patch-clamp studies have identifiedone K+ channel in the luminal membrane of the rat CTAL(Bleich, Schlatter & Greger, 1990; Bleich, Kottgen, Schlatter& Greger, 1995) which is inhibited by a decrease in internalpH. Our own results indicate that the second target ofacidosis for inhibiting NaCl absorption might be the small-conductance Cl- channel in the basolateral membrane. Theinternal pH would be then able to control both the apical
and basolateral steps of Cl- cotransport across the CTALepithelium. The influence of internal pH as a physiologicalmodulator of NaCl transport is supported by the fact thatthe thick ascending limb is a site for ammonia absorption(for review see Knepper, Packer & Good, 1989).
As stated in the introduction, the small-conductance CTALchannel resembles CFTR in a number of ways. However,although CFTR mRNA was found in several parts of thehuman and rat renal tubules, including CTAL (Morales,Morita, Stanton, Lopes. & Guggino, 1994; Schwiebert et at.1994), antibodies against CFTR showed immunoreactivityonly on the apical membranes of renal tubules (Crawford etal. 1991), while the CTAL Cl- channel is embedded in thebasolateral membrane. Furthermore, the CTAL channel hasseveral properties that are unlike those of humanrecombinant CFTR, particularly the blockade by internal4,4'-diisothiocyanatostilbene-2,2'-disulphonic acid (DIDS)and some aspects of the ion selectivity profile (Guinamardet at. 1995). Thus, the similarities with CFTR might bepurely functional and the CTAL channel is probably adistinct entity. This is not too unusual, Cl- channelsresembling CFTR have been found in cells not expressingCFTR (Gabriel, Price, Boucher & Stutts, 1992). The pHsensitivity of recombinant CFTR has not, to ourknowledge, been investigated directly using the patch-clamp method. Nevertheless, there is indirect evidence thatCFTR channels are not inhibited at low pH, since CFTR ispresent and functional in endosomes where the pH is low(Barasch, Kiss, Prince, Saiman, Gruenert & Al-Awqati,1991; Lukacs, Chang, Kartner, Rotstein, Riordan &Grinstein, 1992). Therefore, pH sensitivity is another (butnot conclusive) indication that the small-conductance Cl-channel discussed here is distinct from the CFTR.
Cl- channels having properties similar to those of the CTALchannel have been found in the basolateral membranes of therabbit proximal tubule (Segal, Geibel & Boulpaep, 1993) andin apical membranes of cultured cells from distal convolutedtubule (Poncet, Tauc, Bidet & Poujeol, 1994) and corticalcollecting tubule (Ling, Kokko & Eaton, 1994). There is noevidence that these channels are also sensitive to internalpH. Several molecular biology studies report that channelsrelated to the Cl- channel of Torpedo electric organ (C0Cchannel family) are present in human and rat kidneys (forreview see Pusch & Jentsch, 1994), including in the thickascending limb (Kieferle, Fong, Bens, Vandewalle & Jentsch,1994). While C0C channels are often activated at low pH(Pusch & Jentsch, 1994; Malinowska, Kupert, Bahinski,Sherry & Cuppoletti, 1995), a kidney-specific ClC-Klchannel, expressed in Xenopus oocytes, is inhibited at lowpH (Uchida, Sasaki, Uchida, Horita, Nihei & Marumo,1995). As a whole, the properties of the CTAL Cl- channeldo not seem incompatible with those of the C0C channels:these channels often have small unit conductances (forreview,Pc&es,s & ,1994) and at least one 010 channelis activated by protein kinase A (Malinowska et al. 1995).
J Physiol.490.3764

In conclusion, the precise nature of the small-conductanceC1- channel in the basolateral membrane of the thickascending limb remains unresolved and requires furtherstudy. However, it is probable that sensitivity of the C1-channel to pH, besides its importance for CTAL function,will be useful for distinguishing between different types ofsmall-conductance C1- channels.
BARASCH, J., Kiss, B., PRINCE, A., SAIMAN, L., GRUENERT, D & AL-AWQATI, Q. (1991). Defective acidification of intracellular organellesin cystic fibrosis. Nature 352, 70-72.
BECQ, F., JENSEN, T. J., CHANG, X.-B., SAVOIA, A., ROMMENS, J. M.,TsuI, L.-C., BUCHWALD, M., RIORDAN, J. R. & HANRAHAN, J. W.(1994). Phosphatase inhibitors activate normal and defective CFTRchloride channels. Proceedings of the National Academy of Sciencesof the USA 91, 9160-9164.
BLEICH, M., KOTTGEN, M., SCHLATTER, E. & GREGER, R. (1995).Effect of NH4/NH3 on cytosolic pH and the K+ channels of freshlyisolated cells from the thick ascending limb of Henle's loop. PfluigersArchiv 429, 345-354.
BLEICH, M., SCHLATTER, E. & GREGER, R. (1990). The luminal K+channel of the thick ascending limb of Henle's loop. Pfliigers Archiv415, 449-460.
CRAWFORD, I., MALONEY, P., ZEITHIN, P., GUGGINO, W. B., HYDE,S.C., TURLEY, H., GATTER, K. C., HARRIS, A. & HIGGINS, C. F.(1991). Immunocytochemical localization of the cystic fibrosis geneproduct CFTR. Proceedings of the National Academy of Sciences ofthe USA 88, 9262-9266.
GABRIEL, S. I., PRICE, E. M., BOUCHER, R. C. & STUTTS, M. J. (1992).Small chloride channels are endogenous to nonepithelial cells.American Journal of Physiology 263, C708-713.
GUINAMARD, R., CHRAIBI, A. & TEULON, J. (1995). A small-conductance C1- channel in the mouse thick ascending limb that isactivated by ATP and protein kinase A. Journal of Physiology485,97-112.
HAMILL, 0., MARTY, A., NEHER, E., SAKMANN, B. & SIGWORTH, F.(1981). Improved patch-clamp technique for high resolutionrecording from cells and cell-free membrane patches. PfliigersArchiv 391, 85-100.
KIEFERLE, S., FONG, P., BENS, M., VANDEWALLE, A. & JENTSCH, T. J.(1994). Two highly homologous members of the C1C chloride channelfamily in both rat and human kidney. Proceedings of the NationalAcademy of Sciences of the USA 91, 6943-6947.
KIKERI, D., SUN, A., ZEIDEL, M. L. & HEBERT, S. C. (1992). CellularNH4+/K+ transport pathways in mouse medullary thick limb ofHenle. Journal of General Physiology 99, 435-461.
KNEPPER, M. A., PACKER, R. & GOOD, D. W. (1989). Ammoniumtransport in the kidney. Physiological Reviews 69, 179-249.
LING, B. N., KOKKO, K. E. & EATON, D. C. (1994). Prostaglandin E2activates clusters of apical C1- channels in principal cells via a cyclicadenosine monophosphate-dependent pathway. Journal of ClinicalInvestigation 93, 829-837.
LUKACS, G. L., CHANG, X. B., KARTNER, N., ROTSTEIN, O. D.,RIORDAN, J. R. & GRINSTEIN, S. (1992). The cystic fibrosistransmembrane regulator is present and functional in endosomes.Journal of Biological Chemistry 267, 14568-14572.
LUKACS, G. L., NANDA, A., ROTSTEIN, O. R. & GRINSTEIN, S. (1991).The chloride channel blocker 5-nitro-2-(3-phenylpropyl-amino)
765
MALINOWSKA, D. H., KUPERT, E. Y., BAHINSKI, A., SHERRY, M. &CUPPOLETTI, J. (1995). Cloning, functional expression andcharacterization of a PKA-activated gastric C1- channel. AmericanJournal of Physiology 268, C191-200.
MORALES, M. M., MORITA, T., STANTON, B., LOPES, A. G. & GUGGINO,W. B. (1993). Localization of the cystic fibrosis transmembraneconductance regulator (CFTR) along the rat nephron using thepolymerase chain reaction. Journal of the American Society ofNephrology 3, 875a.
PAULAIS, M. & TEULON, J. (1990). A cAMP-activated chloride channelin the basolateral membrane of the thick ascending limb of themouse kidney. Journal of Membrane Biology 113, 253-260.
PONCET, V., TAUC, M., BIDET, M. & POUJEOL, P. (1994). Chloridechannels in the apical membrane of primary cultures of the rabbitdistal bright convoluted tubule. American Journal of Physiology266, F543-553.
PUSCH, M. & JENTSCH, T. J. (1994). Molecular physiology of voltage-gated chloride channels. Physiological Reviews 74, 813-827.
REEVES, W. B. & ANDREOLI, T. E. (1992). Sodium chloride transportin the loop of Henle. In The Kidney: Physiology andPathophysiology, ed. SELDIN, D. W. & GIEBISCH, G., pp. 1975-2001.Raven Press, New York.
SCHWIEBERT, E. M., LOPES, A. G. & GUGGINO, W. B. (1994). Chloridechannels along the nephron. In Chloride Channels, Current Topics inMembranes, vol. 42, ed. GUGGINO, W. B., pp. 265-315. AcademicPress, New York.
SEGAL, A. S., GEIBEL, J. & BOULPAEP, E. L. (1993). A chloride channelresembling CFTR on the basolateral membrane of rabbit proximaltubule. Journal of the American Society of Nephrology 4, 879a.
THOMAS, J. A., BUCHSBAUM, R. N., ZIMNIAK, A. & RACKER, E. (1979).Intracellular pH measurement in Ehrlich ascites tumor cellsutilizing spectroscopic generated in situ. Biochemistry 18,2210-2218.
UCHIDA, S., SASAKI, S., UCHIDA, K., HORITA, S., NIHEI, H. &MARUMO, F. (1995). Localization and functional characterization ofrat kidney-specific chloride channel, C1C-K1. Journal of ClinicalInvestigation 95, 104-113.
WELSH, M. J., ANDERSON, M. P., RICH, D. P., BERGER, H. A.,DENNING, G. M., OSTEDGAARD, L. S., SHEPPARD, D. N., CHENG,S.H., GREGORY, R. J. & SMITH, A. E. (1992). Cystic fibrosistransmembrane conductance regulator: a chloride channel withnovel regulation. Neuron 8, 821-829.
WINGO, C. S. (1986). Effect of acidosis on chloride transport in thecortical thick ascending limb of Henle perfused in vitro. Journal ofClinical Investigation 78, 1324-1330.
AcknowledgementsWe thank Dr R. Greger (University of Freiburg, Germany) forkindly providing NPPB, Dr 0. Parkes for editing the manuscript,M. Blonde for technical assistance and M. Poitou for secretarialhelp. This work was supported in part by the AssociationFranqaise de Lutte contre la Mucoviscidose. R.G. is the recipient ofa Ministere de la Recherche et de l'Espace (MRE) researchstudentship; M.P. is an INSERM researcher and J.T. is fromCentre National de la Recherche Scientifique (CNRS).
Received 28 April 1995; accepted 8 August 1995.
benzoic acid (NPPB) uncouples mitochondria and increases theproton permeability of the plasma membrane in phagocytic cells.FEBS Letters 288, 17-20.
J Physiol. 490.3 pH-sensitive small-conductance C- channel