cardiotonic pill reduces myocardial ischemia-reperfusion injury...
TRANSCRIPT

1521-009X/44/7/878–887$25.00 http://dx.doi.org/10.1124/dmd.116.069914DRUG METABOLISM AND DISPOSITION Drug Metab Dispos 44:878–887, July 2016Copyright ª 2016 by The American Society for Pharmacology and Experimental Therapeutics
Cardiotonic Pill Reduces Myocardial Ischemia-Reperfusion Injury viaIncreasing EET Concentrations in Rats s
Meijuan Xu,1 Haiping Hao,1 Lifeng Jiang, Yidan Wei, Fang Zhou, Jianguo Sun, Jingwei Zhang,Hui Ji, Guangji Wang, Wenzheng Ju, and Ping Li
State Key Laboratory of Natural Medicines (M.X., H.H., L.J., Y.W., H.J., P.L.) and Key Laboratory of Drug Metabolism andPharmacokinetics (H.H., F.Z., J.S., J.Z., G.W.), China Pharmaceutical University, Nanjing, China; and Department of Clinical
Pharmacology, Affiliated Hospital of Nanjing University of Chinese Medicine, Nanjing, China (M.X., W.J.)
Received February 4, 2016; accepted May 4, 2016
ABSTRACT
Accumulating data suggest that epoxyeicosatrienoic acids (EETs)and 20-hydroxyeicosatetraenoic acid, both cytochrome P450 (P450)enzymemetabolites of arachidonic acid (AA), play important roles incardiovascular diseases. For many years, the cardiotonic pill (CP),an herbal preparation derived from Salviae Miltiorrhizae Radix etRhizoma, Notoginseng Radix et Rhizoma, and Borneolum Syntheti-cum, has been widely used in China for the treatment of coronaryartery disease. However, its pharmacological mechanism has notbeen well elucidated. The purpose of this study was to investigatethe chronic effects of the CP on myocardial ischemia-reperfusioninjury (MIRI) and AA P450 enzyme metabolism in rats (in vivo) andH9c2 cells (in vitro). The results showed that CP dose dependently(10, 20, and 40 mg/kg/d; 7 days) mitigated MIRI in rats. The plasmaconcentrations of EETs inCP-treated ischemia-reperfusion (I/R) rats
(40 mg/kg/d; 7 days) were significantly higher (P < 0.05) than those incontrols. Cardiac Cyp1b1, Cyp2b1, Cyp2e1, Cyp2j3, and Cyp4f6were significantly induced (P < 0.05); CYP2J and CYP2C11 proteinswere upregulated (P < 0.05); and AA-epoxygenases activity wassignificantly increased (P < 0.05) after CP (40 mg/kg/d; 7 days) ad-ministration in rats. In H9c2 cells, the CP also increased (P < 0.05) theEET concentrations and showed protection in hypoxia-reoxygenation(H/R) cells.However, anantagonist ofEETs, 14,15-epoxyeicosa-5(Z)-enoic acid, displayed a dose-dependent depression of the CP’sprotective effects in H/R cells. In conclusion, upregulation of cardiacepoxygenases after multiple doses of the CP—leading to elevatedconcentrations of cardioprotective EETs after myocardial I/R—maybe the underlying mechanism, at least in part, for the CP’s cardiopro-tective effect in rats.
Introduction
Cardiovascular disease (CVD), including coronary artery disease andacute myocardial infarction, is a human health crisis that remains one ofthe leading causes of death worldwide (Elbekai and El-Kadi, 2006).Increasing evidence has shed light on the roles of the cytochrome P450(P450) enzymes in the onset, progression, and prognosis of CVD. Overthe past three decades, P450 enzymes have been found expressed in thecardiovascular system (Enayetallah et al., 2004; Delozier et al., 2007).Endogenous P450 enzymemetabolites, such as epoxyeicosatrienoic acids(EETs), aldosterone, and sex hormones, are shown to be implicated
in the maintenance of cardiovascular health (Elbekai and El-Kadi,2006).Arachidonic acid (AA) is a polyunsaturated fatty acid located in body
cell membranes in an inactive state, and can be released by phospho-lipase A2 in response to stimuli such as ischemia (Sato et al., 2011).Recent investigations indicate that the P450 enzyme metabolites of AA,EETs, and 20-hydroxyeicosatetraenoic acid (20-HETE), are involved inmany cardiovascular pathophysiological mechanisms, and play impor-tant roles in myocardial ischemia-reperfusion injury (MIRI) (Elbekaiand El-Kadi, 2006; Sato et al., 2011; Chen and Wang, 2013). EETs,mainly generated by members of the CYP2 family, including CYP2C8,CYP2C9, and CYP2J2, are the epoxidation metabolites of AA. Theiractions are terminated by conversion to the less biologically activedihydroxyeicosatrienoic acids (DHETs) by soluble epoxide hydrolase(sEH). 20-HETE is the v-hydroxylation metabolite of AA, and isprimarily produced by members of the CYP4 family, including CYP4Aand CYP4F isoforms (Sato et al., 2011). EETs act as vasodilatorysubstances in several vascular beds via activation of Ca2+-activated Kchannels (Campbell et al., 1996), endothelial nitric oxide synthase(Wang et al., 2003), and the TRPV4 channel (Vriens et al., 2005). In theheart, EETs regulate L-type Ca2+, ATP-sensitive K+, and Na+ channels
This work was sponsored by the National Natural Science Foundation of China[Grants 81130068 and 81202984]; the National Science and Technology MajorProject “Creation of Major New Drugs” [Grant 2012ZX09303009-002]; the PriorityAcademic Program Development of Jiangsu Higher Education Institutions [PAPD];the 12th Six Talent Peaks Project in Jiangsu Province [Grant WSN-054]; andthe Leading Talents of Scientific Research in TCM of Jiangsu Province [GrantLJ200906].
1M.X. and H.H. contributed equally to this work.dx.doi.org/10.1124/dmd.116.069914.s This article has supplemental material available at dmd.aspetjournals.org.
ABBREVIATIONS: AA, arachidonic acid; CDP, compound Danshen prescription; CK-MB, creatine kinase MB; CP, cardiotonic pill; CVD,cardiovascular disease; DHET, dihydroxyeicosatrienoic acid; EET, epoxyeicosatrienoic acid; 14,15-EEZE, 14,15-epoxyeicosa-5(Z)-enoic acid; HE,ethidium; 20-HETE, 20-hydroxyeicosatetraenoic acid; H/R, hypoxia-reoxygenation; I/R, ischemia-reperfusion; LC-MS/MS, liquid chromatography–tandem mass spectrometry; LDH, lactate dehydrogenase; MIRI, myocardial ischemia-reperfusion injury; m/z, mass-to-charge ratio; P450,cytochrome P450; PBS, phosphate-buffered saline; PCR, polymerase chain reaction; ROS, reactive oxygen species; sEH, soluble epoxidehydrolase; SPE, solid-phase extraction; TTC, 2,3,5-triphenyltetrazolium chloride.
878
http://dmd.aspetjournals.org/content/suppl/2016/05/05/dmd.116.069914.DC1Supplemental material to this article can be found at:
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from

(Konkel and Schunck, 2011) and inhibit apoptosis by a caspase-dependent pathway (Wang et al., 2012). Notably, EETs inhibit theinflammatory response by repressing NF-kB activation and inhibitingTNF-a–induced VCAM-1 expression (Node et al., 1999). Moreover,EETs are potent endogenous angiogenic factors (Xu et al., 2011).However, 20-HETE has been demonstrated to have vasoconstrictive(Miyata and Roman, 2005), proinflammatory (Ishizuka et al., 2008) andproapoptosis (Nilakantan et al., 2008) functions. Furthermore, althoughCYP2J2 and CYP2C both generate cardioprotective EETs from AA,studies have demonstrated that CYP2C also produces detrimentalreactive oxygen species (ROS) (Fleming et al., 2001). In animal models,effective protection against MIRI has been shown by increasing EETconcentrations through direct administration of EETs, upregulatingthe expression of AA epoxygenases using transgenic technology, orinhibiting the activity of sEH (Seubert et al., 2004; Motoki et al., 2008;Batchu et al., 2011; Oni-Orisan et al., 2014); decreasing 20-HETEgeneration through CYP4A/4F inhibition (Nithipatikom et al., 2004);and depressing ROS formation via CYP2C inhibition (Granville et al.,2004). AA-metabolizing P450 enzymes are thus considered to betherapeutic targets for MIRI and other CVDs.For many years, the compound Danshen prescription (CDP), an
herbal preparation derived from SalviaeMiltiorrhizae Radix et Rhizoma,Notoginseng Radix et Rhizoma, and Borneolum Syntheticum has beenwidely used in China to improve cardiac function and coronarycirculation in the therapy of stable angina pectoris (Luo et al., 2013).Numerous CDP preparations, such as tablets, capsules, injectables,granules, oral liquid, and dripping pills, are commercially available inChina. Among them, the cardiotonic pill (CP), also known as thedantonic pill or the compound Danshen dripping pill, is the best known.The CP is now available in countries such as Singapore, Korea, India, theUnited Arab Emirates, Russia, Cuba, and South Africa as a prescriptionor over-the-counter drug (Lu et al., 2008). It is in phase III clinicaldevelopment in the United States (Lei et al., 2014). Although the CDP isrecognized as clinically effective for coronary artery disease and iswidely used, its pharmacological mechanism has not yet been fullydocumented. Moreover, to our knowledge, neither chronic CP’s pro-tective effects against MIRI, nor the association between its pharmaco-logical effects and AA P450 enzyme metabolism has been reported.This study used the CP as the typical CDP preparation, investigating
its effects on cardiac function, serum creatine kinase MB (CK-MB) andlactate dehydrogenase (LDH) concentrations, myocardial infarction andsuperoxide generation in ischemia-reperfusion (I/R) rats, and hypoxia-reoxygenation (H/R) injury in H9c2 cells. This study also examined theCP’s effects on the concentrations of AA P450 enzyme metabolites, andevaluated its influence on the cardiac expressions and activities of AA-metabolizing P450 enzymes in rats.
Materials and Methods
Chemicals and Reagents
14,15-EET; 11,12-EET; 8,9-EET; 5,6-EET; 14,15-DHET; 11,12-DHET; 8,9-DHET; 5,6-DHET; 14,15-epoxyeicosa-5(Z)-enoic acid (14,15-EEZE); 14,15-DHET-d11; 14,15-EET-d11; and 20-HETE were obtained from CaymanChemicals (Ann Arbor, MI). AA, NADPH, MgCl2, 2,3,5-triphenyltetrazoliumchloride (TTC), and dihydroethidium were obtained from Sigma-Aldrich Co. (St.Louis, MO). The CK-MB isoenzyme and LDH assay kits were purchased fromNanjing Jiancheng Bioengineering Institute (Nanjing, China). The TRIzol reagentand Prime Script RTMaster Mix Perfect Real Time Kit were bought from Takara(Tokyo). The primary antibody of CYP2C11 (ab3571) was purchased fromAbcam (Cambridge, MA), and antibodies of CYP2J (sc-67276), sEH (sc-24797),and glyceraldehyde-3-phosphate dehydrogenase (sc-365062) were obtained fromSanta Cruz Biotechnology, Inc. (Paso Robles, CA). The Pierce ECL WesternBlotting Kit was bought from Thermo Fisher Scientific (Grand Island, NY). The CP
used in this research (27 mg each pill) was obtained from Tasly PharmaceuticalGroup Co., Ltd. (Tianjin, China). The content levels of various Danshen componentswere determined in a previous study (Liu et al., 2014) using three different CPbatches. The average levels were as follows: 30.4 mg/g of Danshensu; 10.2 mg/g ofprotocatechuic aldehyde; 8.1 mg/g of salvianolic acid A; 4.5 mg/g of salvianolic acidB; 8.2 mg/g of salvianolic acid D; 1.6 mg/g of lithospermic acid; 4.1 mg/g ofrosmarinci acid; 0.74 mg/g of tanshinone I; 0.63 mg/g tanshinone IIA; 0.39 mg/g ofcryptotanshinone; 0.63 mg/g of dihydrotanshinone I; 22.0 mg/g of ginseno-sides Rg1; 19.1 mg/g of ginsenosides Rb1; and 10.6 mg/g of ginsenosides Rh1(see Supplemental Table 1). High-performance liquid chromatography–gradeformic acid, ammonium acetate, and acetonitrile were obtained from Merck(Darmstadt, Germany). Water was obtained from a Milli-Q water system(Millipore, Bedford, MA). All other chemicals used were the highest gradecommercially available.
Animals
Adult male Sprague-Dawley rats (2406 20 g body weight) were supplied bythe Nanjing Qinglongshan Experimental Animal Co. Ltd. (Nanjing, China). Allexperimental procedures and protocols were in compliance with the NationalInstitutes of Health Guide for the Care and Use of Laboratory Animals (http://grants.nih.gov/grants/olaw/Guide-for-the-Care-and-Use-of-Laboratory-Animals.pdf)and approved by the Animal Ethics Committee of China Pharmaceutical Univer-sity. The animals were maintained in a room with a temperature of 25�C6 2�C, a12-hour day-night cycle, and 50% 6 10% relative humidity. Water and food(laboratory rodent chow pellets, Sikerui Biological Technology Co. Ltd.,Nanjing,China) were provided ad libitum.
Groups and Administration of the CP in Rats
As shown in Table 1, the present in vivo study was comprised of threesubstudies, using separate rat samples. Rats were divided into groups at random(randomized block experiment design) and gavage administered normal saline orCP solution. The CP solution was freshly prepared before use each day bycrushing the pills with a mortar and then dissolving them in normal saline with thehelp of ultrasound. No organic solvent was added. The animal doses were derivedaccording to the daily dose of the CP applied in the phase II clinical trials (low-dosage group: 250 mg per person daily; high-dosage group: 375 mg per persondaily), which was calculated by the body surface area difference between humansand rats, with minor adjustments to conform to pharmacological research con-ventions. All assessors in the study were blinded to the group assignment of thespecimens they were assessing.
Establishment of the MIRI Model
After 7 days of administration of normal saline or CP, the rats wereanesthetized with intraperitoneal injections of chloral hydrate (300 mg/kg).Following endotracheal intubation, the heart was exposed via a left thoracotomyin the fourth intercostal space. For induction of the myocardial ischemia model,the left anterior descending coronary artery, together with a silica rubber cannula,was ligated with a 6-0 silk suture. After 45 minutes of occlusion, the coronaryartery was reperfused by cutting the suture. The length of reperfusion was 15minutes for superoxide generation measurements, 30 minutes and 3 hours forAA P450 enzyme metabolites determinations, and 24 hours for cardiac injury
TABLE 1
Groups, numbers, and administration of the CP in rats
GroupNumber of Rats
CP Treatment (7 Days)Study 1a Study 2b Study 3c
Total 45 21 18 —
Sham/control 9 7 9 Normal salineModel 9 7 — Normal salineCP (low dose) 9 — — 10 mg/kg/dayCP (middle dose) 9 — — 20 mg/kg/dayCP (high dose) 9 7 9 40 mg/kg/day
aPharmacological study.bAA P450 enzyme metabolite determination.cAA-metabolizing P450 enzyme expression and activity.
CP Reduces MIRI via Increasing EET levels in Rats 879
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from

measurements. Alterations of color in the myocardium and electrocardiographicchanges were used to confirm successful surgery. Rats in the sham groupexperienced the same surgical progress without the ligation.
Assessment of Myocardial Injury
After 45 minutes of myocardial ischemia and 24 hours of reperfusion, all ratswere narcotized by urethane (1 g/kg). Hemodynamics and cardiac function weremeasured using a biologic mechanism experiment system (BL420, Taimeng Co.Ltd., Chengdu, China) as described previously (Wei et al., 2014). A pressure-volume catheter, filled with heparin saline (500 U/ml), was inserted into the rightcarotid artery and then advanced into the left ventricle for monitoring heart rate,left-ventricular end-diastolic pressure, left-ventricular systolic pressure, andmaximum/minimum rates of developed left ventricular pressure (6 dp/dtmax).Subsequently, all rats were sacrificed to harvest the hearts immediately for infarctsize determination.
The size of the infarcted myocardium was determined through TTC staining.The hearts were kept at220�C for 10 minutes, and then cut into five 1- to 2-mm-thick slices and incubated for 10 minutes in 1% (w/v) TTC in buffer (pH 7.4) at37�C. Next, the slices were fixed in 10% formalin and pictures were taken. Thesizes of infarcted area (white coloration) and at-risk areas (the entire scannedsection) were determined by the Image-Pro Plus image analysis software (version4.1, Media Cybernetics, LP, Silver Spring, MD). The infarct size was calculatedas a percentage of the risk area for assessing the degree of myocardial infarction.
Myocardial injury was assessed bymeasuring the amount of CK-MB and LDHin serum collected after 24 hours of reperfusion. CK-MB and LDH are expressedin the heart muscle and released during tissue damage, thus they are used asindicators of cardiac injury (Liu et al., 2014). The activities of CK-MB and LDHin serum were assessed by commercially available kits, and measured spectro-photometrically at 340 and 450 nm, respectively. All procedures adhered to themanufacturer’s instructions.
Superoxide Generation Measurement in Heart Tissues
Superoxide generation in the I/R heart tissue was determined as previouslyreported (Khan et al., 2007) with dihydroethidium, which can be oxidized tofluorescent ethidium (HE) by superoxide and then intercalated into DNA. Sincesuperoxide generation in hearts subjected to I/R was reported to occur during thefirst 15 minutes of reperfusion, HE fluorescence was determined at this period(after 15 minutes of reperfusion) (Khan et al., 2007). Fluorescence intensity wasquantified using ImageJ software (version 1.48, National Institutes of Health).
Real-time Polymerase Chain Reaction (PCR) Analysis of P450 Enzymes inthe Rat Heart
Total RNA from the heart tissues was isolated using a TRIzol reagentaccording to the manufacturer’s instructions. Next, first-strand cDNA synthesiswas performed by using the Prime Script RT Master Mix Perfect Real Time Kit(Takara) according to the manufacturer’s instructions. Quantitative analysis ofspecific mRNA expression was performed with real-time PCR by subjecting theresulting cDNA to PCR amplification using 96-well optical reaction plates in theABI Prism 7500 System (Applied Biosystems, Foster City, CA). The primersemployed in the current study (Table 2) were chosen from previously publishedstudies (Zordoky et al., 2011; Alsaad et al., 2012). No-template controls wereincorporated into the same plate to test for contamination of any assay reagents. Anoptical adhesive cover was used to seal the plate; thereafter, thermocycling conditionswere initiated at 95�C for 10 minutes, followed by 40 PCR cycles of denaturationat 95�C for 15 seconds, annealing at 60�C for 15 seconds, and extension at 72�C for30 seconds. Dissociation curves were performed at the end of each cycle to confirmthe specificity of the primers and purity of the final PCR product.
Microsomal Protein Preparation
Cardiac microsomal protein was prepared from heart tissues as describedpreviously (Aboutabl et al., 2009). In brief, heart tissues were washed with ice-cold phosphate-buffered saline (PBS), cut into pieces, and homogenized using anIKA T10 basic ULTRA-TURRAX homogenizer (IKA, Königswinter, Germany)in additional cold PBS solution. The homogenates from three separate hearts inthe same group were mixed, and the microsomal and cytosolic proteins wereseparated by differential ultracentrifugation from the homogenized tissues.
Thereafter, the final microsomal pellet was reconstituted in cold PBS containing20% glycerol and stored at280�C. The bicinchonininc acid method using bovineserum albumin as a standard (Beyotime, Nanjing, China) was used to determinethe concentration of heart microsomal and cytosolic proteins.
Western Blotting Analysis
Next, 50 mg of cardiac microsomal (for P450 enzymes) or cytosolic (for sEH)protein from each treatment group was separated by 10% SDS-PAGE, and thenelectrophoretically transferred to nitrocellulose membrane. Protein blots werethen blocked overnight at 4�C in blocking solution containing 0.15 M sodiumchloride, 3 mM potassium chloride, 25 mM Tris base (Tris-buffered saline), 5%skim milk, 2% bovine serum albumin, and 0.5% Tween 20. After blocking, theblots were incubated with the primary antibodies of CYP2C11 (1:1000), CYP2J,sEH, and glyceraldehyde-3-phosphate dehydrogenase (1:200) for 2 hours.Incubation with a peroxidase-conjugated secondary antibody was performed for2 hours at room temperature. The bands were visualized using the enhancedchemiluminescence method according to the manufacturer’s instructions. Theintensity of the protein bands was quantified, relative to the signals obtained forglyceraldehyde-3-phosphate dehydrogenase, using the ImageJ software (version1.48, National Institutes of Health).
Activity Determination of AA-Metabolizing P450 Enzymes
Rat heart microsomes (1 mg/ml) were incubated at 37�C in a shaking waterbath (50 rpm). The incubation buffer consisted of 0.1 M potassium phosphatebuffer containing 0.15 M KCl and 1 mM EDTA (pH 7.4). After pre-equilibrationfor 5 minutes, the reaction was initiated by the addition of 1 mM NADPH. AAwas added to the incubation mixture with a final concentration of 50 mM andincubated at 37�C for 30 minutes. The reaction was terminated by the addition of200 ml of ice-cold acetonitrile followed by the internal standards, 14,15-EET-d11(40 ng/ml) and 14,15-DHET-d11 (40 ng/ml). After a vortex for 1 minute, theincubation mixtures were centrifuged at 12,000 rpm for 5 minutes at 4�C, and analiquot of 10 ml of the supernatant was injected for liquid chromatography–tandem mass spectrometry (LC-MS/MS) analysis.
sEH activity was determined using its natural substrate 14,15-EET. Thecytosolic protein was diluted to 0.4 mg/ml with potassium phosphate buffer(0.1 M, pH 7.4), supplemented with bovine serum albumin (2.5 mg/ml) (Alsaadet al., 2012). The reaction was initiated by adding 14,15-EET (final concentration2 mg/ml) to the preheated cytosolic solution (37�C for 5 minutes), after which thereaction was conducted at 37�C for another 5 minutes. After incubation, thereactionwas terminated by adding 1ml of ice-cold ethyl acetate and 10ml of 14,15-DHET-d11 (10 ng/ml). After shaking for 3 minutes, the tubes were centrifuged andthe organic phase was transferred to a new tube. The organic phase was evaporatedto dryness using a CentriVap Centrifugal Concentrator (Labconco, Kansas City,MO). The residue was dissolved in 120ml of dehydrated alcohol and vortex mixedfor 1 minute. The tubes were then centrifuged at 12,000g for 10 minutes, and 5 mlof the resulting supernatant was injected into the LC-MS/MS system.
Cell Culture and H/R Treatments
H9c2 cells (American Type Culture Collection, Manassas, VA) weremaintained in Dulbecco’s modified Eagle’s medium, without phenol red,supplemented with 0.45% glucose, 0.11% sodium pyruvate, 0.15% sodiumbicarbonate, 20 mM L-glutamine, 10% fetal bovine serum, 100 IU/ml penicillin,and 10 mg/ml streptomycin. Cells were grown in 75-cm2 culture plates at 37�C ina 5% CO2 humidified incubator (Thermo Fisher Scientific). To mimic hypoxicinjury in vitro, the cells were incubated for 9 hours in a hypoxic solutioncontaining 0.9 mM NaH2PO4, 6.0 mM NaHCO3, 1.0 mM CaCl2, 1.2 mMMgSO4, 40 mM natrium lacticum, 20 mMHEPES, 98.5 mMNaCl, and 10.0 mMKCl (pH adjusted to 6.8). The hypoxic condition was produced by placing theplates of cultured cardiomyocytes in a hypoxic incubator (SANYO, Osaka,Japan); oxygen was adjusted to 1.0% and CO2 was adjusted to 5.0%. Afterhypoxia, the cells were reincubated in a normal Dulbecco’s modified Eagle’smedium at 37�C in a 5% CO2 humidified incubator for 3 hours.
Treatment of H9c2 Cells with the CP and 14,15-EEZE
To investigate the cardioprotective effect of the CP, cardiac-derived H9c2 cellswere pretreated with various concentrations (0.025–0.02 mg/ml) of the CP
880 Xu et al.
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from

(dissolved in the cell culture with the help of ultrasonic and vortex mixing) for 12hours. FollowingH/R, the cell viability was determined. To investigate the actionsof 14,15-EEZE on the cardioprotective effect of the CP in H9c2 cell H/R injury,various concentrations of 14,15-EEZE (0.01, 0.1, and 1 mM) were added to thehypoxia solution and incubated with H9c2 cells in a hypoxic condition. 14,15-EEZEwas dissolved in ethanol, in which the final concentration was less than 1%(v/v) in the mixture, and the same amount of ethanol was added to the controlgroup. After H/R, the cell viability was determined by a Cell Counting Kit-8 assay(CCK-8, Dojindo, Shanghai, China).
Sample Pretreatment for AA P450 Enzyme Metabolite Determination
Plasma Sample. Next, 300 ml of rat plasma was added with 315 ml methanol,30 ml internal standard (14,15-DHET-d11, 5 ng/ml), and 30 ml butylatedhydroxytoluene (10 mg/ml). After vortex mixing for 1 minute, the mixture wasthen centrifuged at 12,000 rpm at 4�C for 5 minutes. The supernatant was drawnoff and treated with a solid-phase extraction (SPE) method described previously(Zhang et al., 2007;Martin-Venegas et al., 2011)withminormodification. Briefly,the supernatant was applied to a 3-ml Oasis HLB SPE column (3 cc/60 mg, Waters,Milford, MA) that had been preconditioned with 3 ml methanol and 3 ml water. Thecolumns were washed with 3 ml of water and allowed to run dry. Then, AA P450enzyme metabolites were eluted from the column with 5 ml of ethyl acetate. Becausethere was still a small amount of water left on the SPE column, the elution liquid wasdivided into two layers. The ethyl acetate layer was carefully removed from the waterlayer and dried using a CentriVap Centrifugal Concentrator (Labconco). The samplewas then redissolved in 125ml ethanol. After being vortexed for 1 minute, the contentwas centrifuged at 12,000 rpm at 4�C for 10 minutes, and an aliquot of 5 ml of thesupernatant was injected for LC-MS/MS analysis.
Cell Sample. After control or CP treatment, the cells were washed with ice-cold PBS, collected in 1 ml PBS, and disrupted with an ultrasonic crusher (HaishuKesheng Ultrasonic Equipment, Ningbo, China). After being centrifuged at12,000 rpm for 5 minutes at 4�C, the supernatant was loaded onto the SPEcolumn, with the same extraction procedure as was used for the plasma sample.
Measurement of AA P450 Enzyme Metabolites Using LC-MS/MS
The P450 enzyme metabolites of AA, EETs, DHETs, and 20-HETE weresimultaneously quantitated using the LC-MS/MS system (API 4000 LC-MS/MS;Applied Biosystems Sciex, Ontario, Canada) equipped with an electrosprayionization interface. Chromatography was performed using a C18 column(2.6 mm, 100 � 2.1 mm; Kinetex, Phenomenex, Torrance, CA) and an Agilent(Agilent Technologies, Foster City, CA) 1200 high-performance liquid chroma-tography system. The mobile phase, which consisted of a 0.1% formic acidaqueous solution (A) and acetonitrile (B), was delivered with a gradient elution ata flow rate of 0.6 ml/min: 0 minutes, 50% B; 3–5 minutes, 62% B; 6–10 minutes,85% B; and 11–18 minutes, 50% B. The column temperature was maintained at40�C. Ion spray voltage was set at 4.5 kV for negative ionization, and the heatergas temperature was 650�C. Nitrogen was used as the nebulizing gas (50 psi),auxiliary gas (70 psi), and curtain gas (15 psi). The Multiple Reaction Monitoring
experiments were conducted by monitoring the precursor ion to product iontransitions for 20-HETE [mass-to-charge ratio (m/z) 319.2–289.3]; 14,15-DHET(m/z 337.2–207.0); 11,12-DHET (m/z 337.2–208.1); 8,9-DHET (m/z 337.2.2–127.1); 5,6-DHET (m/z 337.2–145.1); 14,15-EET (m/z 319.2–219.2); 11,12-EET(m/z 319.2–208.1); 8,9-EET (m/z 319.2–155.1); 14,15-DHET-d11 (m/z 348.3–207.2); and 14,15-EET-d11 (m/z 330.3–219.2). The lower limits of quantificationof AA P450 enzyme metabolites are shown in Supplemental Table 2.
Statistical Analysis
The results are presented as the mean6 S.E. Data were subjected to statisticalanalysis using Graphpad Prism 5.0 (GraphPad Software, La Jolla, CA). TheStudent’s t test was used for data comparison between two groups. One-wayanalysis of variance with the Dunnett’s post hoc test was carried out forcomparison of more than two groups. In all cases, a value of P , 0.05 wasconsidered significant.
Results
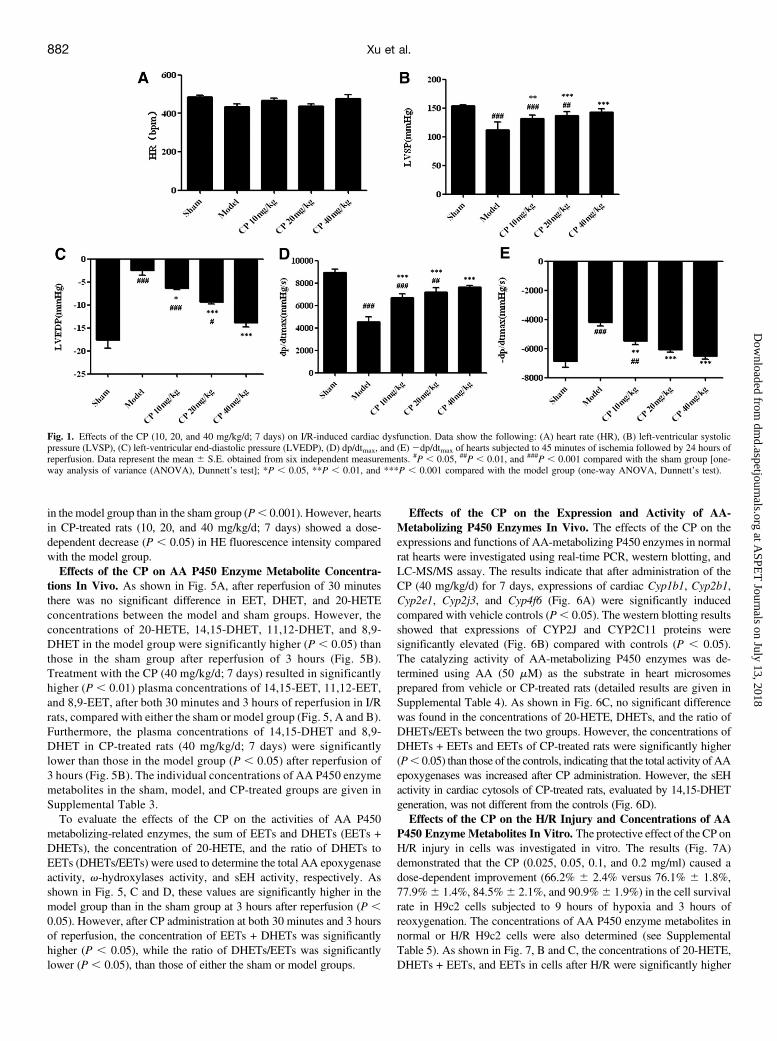
Effect of the CP on MIRI in Rats. The effects of the CP on the heartrate, hemodynamics, and cardiac function were determined after heartswere subjected to 45 minutes of ischemia followed by 24 hours ofreperfusion. There was no significant difference (P. 0.05) in the heartrate among all groups (Fig. 1A). Hearts in the model group showed asignificant decrease in left-ventricular systolic pressure (P, 0.001) anddp/dtmax (P , 0.001), and a significant increase in left-ventricular end-diastolic pressure (P , 0.001) and 2dp/dtmax (P , 0.001) whencompared with those in the sham group (Fig. 1). However, hearts in CP-treated rats (10, 20, and 40 mg/kg/d; 7 days) showed a dose-dependentrecovery (P, 0.05) of left-ventricular systolic pressure, left-ventricularend-diastolic pressure, dp/dtmax, and 2dp/dtmax when compared withthose in the model group (Fig. 1).The serum levels of LDH and CK-MB were regarded as significant
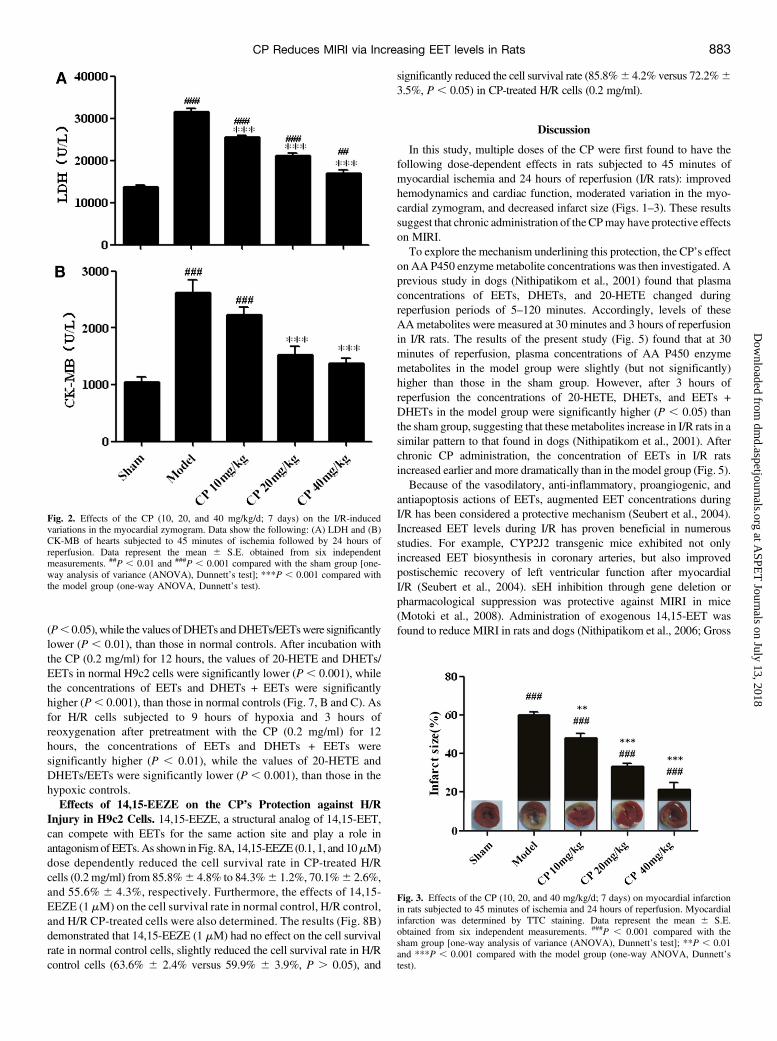
biomarkers of MIRI. As presented in Fig. 2, levels in the model groupwere significantly higher (P , 0.001) than those in the sham group.However, treatment with the CP (10, 20, and 40 mg/kg/d; 7 days)significantly decreased the levels of LDH and CK-MB compared withthe model group.As shown in Fig. 3, TTC staining of hearts in the sham group showed
no infarct and ischemic area. Hearts in the model group showed aninfarction of 59.7%6 4.1% in the risk area. However, infarct size in CP-treated rats (10, 20, and 40mg/kg/d; 7 days) was significantly reduced to48.0% 6 5.4% (P , 0.01), 33.1% 6 4.2% (P , 0.001), and 21.2% 68.0% (P , 0.001), respectively.Effects of the CP on Myocardial Superoxide Generation. As
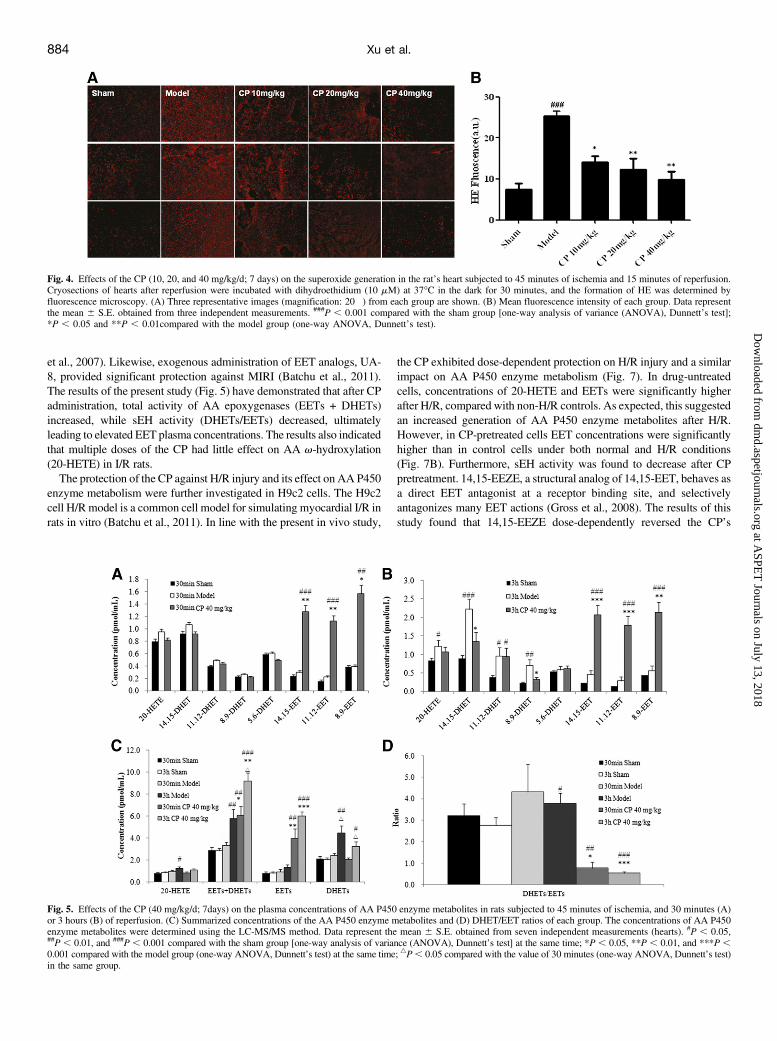
shown in Fig. 4, the HE fluorescence intensity was significantly higher
TABLE 2
Primer sequences used for real-time PCR reactions
Gene Forward Primer Reverse Primer
Cyp1a1 CCCGAGTTCCGGCCT TGCCCCCGAGAATGACyp1b1 CACGACGATGCCGAGTTC AGGACGAAGTTGCTGAAGTTGCyp2b1 AACCCTTGATGACCGCAGT TGTGGTACTCCAATAGGGACAAGATCCyp2c11 CACCAGCTATCAGTGGATTTGG GTCTGCCCTTTGCACAGGAACyp2e1 GCGTGTGTGTGTTGGAGAA AGAGACTTCAGGTTATGCTGCACyp2j3 CATTGAGCTCACAAGTGGCTTT CAATTCCTAGGCTGTGATGTCGCyp4a1 TTGAGCTACTGCCAGATCCCAC CCCATTTTTGGACTTCAGCACACyp4a3 CTCGCCATAGCCATGCTTATC CCTTCAGCTCATTCATGGCCCyp4f1 CCCCCAAGGCTTTTTGATG GAGCGCAACGGCAGCTCyp4f4 AAGGTCTGAAGCAGGTAACTAAGC ACGTCAGGGTGGCACAGAGTCyp4f5 AGGATGCCGTGGCTAACTG GGCTCCAAGCAGCAGAAGACyp4f6 TCACTTGACCTTGATGAAGAACAAC AAGAGAGGTGGATATCACGGAAGEphx1 CACATCCAAGCCACCAAGCC CAGGCCTCCATCCTCCAGEphx2 TCATCAAGTGGCTGAAGAC CTGCTGGCTGTGGATAAGb-Actin CCAGATCATGTTTGAGACCTTCAA GTGGTACGACCAGAGGCATACA
CP Reduces MIRI via Increasing EET levels in Rats 881
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from
in the model group than in the sham group (P, 0.001). However, heartsin CP-treated rats (10, 20, and 40 mg/kg/d; 7 days) showed a dose-dependent decrease (P , 0.05) in HE fluorescence intensity comparedwith the model group.Effects of the CP on AA P450 Enzyme Metabolite Concentra-
tions In Vivo. As shown in Fig. 5A, after reperfusion of 30 minutesthere was no significant difference in EET, DHET, and 20-HETEconcentrations between the model and sham groups. However, theconcentrations of 20-HETE, 14,15-DHET, 11,12-DHET, and 8,9-DHET in the model group were significantly higher (P , 0.05) thanthose in the sham group after reperfusion of 3 hours (Fig. 5B).Treatment with the CP (40 mg/kg/d; 7 days) resulted in significantlyhigher (P , 0.01) plasma concentrations of 14,15-EET, 11,12-EET,and 8,9-EET, after both 30 minutes and 3 hours of reperfusion in I/Rrats, compared with either the sham or model group (Fig. 5, A and B).Furthermore, the plasma concentrations of 14,15-DHET and 8,9-DHET in CP-treated rats (40 mg/kg/d; 7 days) were significantlylower than those in the model group (P , 0.05) after reperfusion of3 hours (Fig. 5B). The individual concentrations of AA P450 enzymemetabolites in the sham, model, and CP-treated groups are given inSupplemental Table 3.To evaluate the effects of the CP on the activities of AA P450
metabolizing-related enzymes, the sum of EETs and DHETs (EETs +DHETs), the concentration of 20-HETE, and the ratio of DHETs toEETs (DHETs/EETs) were used to determine the total AA epoxygenaseactivity, v-hydroxylases activity, and sEH activity, respectively. Asshown in Fig. 5, C and D, these values are significantly higher in themodel group than in the sham group at 3 hours after reperfusion (P ,0.05). However, after CP administration at both 30 minutes and 3 hoursof reperfusion, the concentration of EETs + DHETs was significantlyhigher (P , 0.05), while the ratio of DHETs/EETs was significantlylower (P , 0.05), than those of either the sham or model groups.
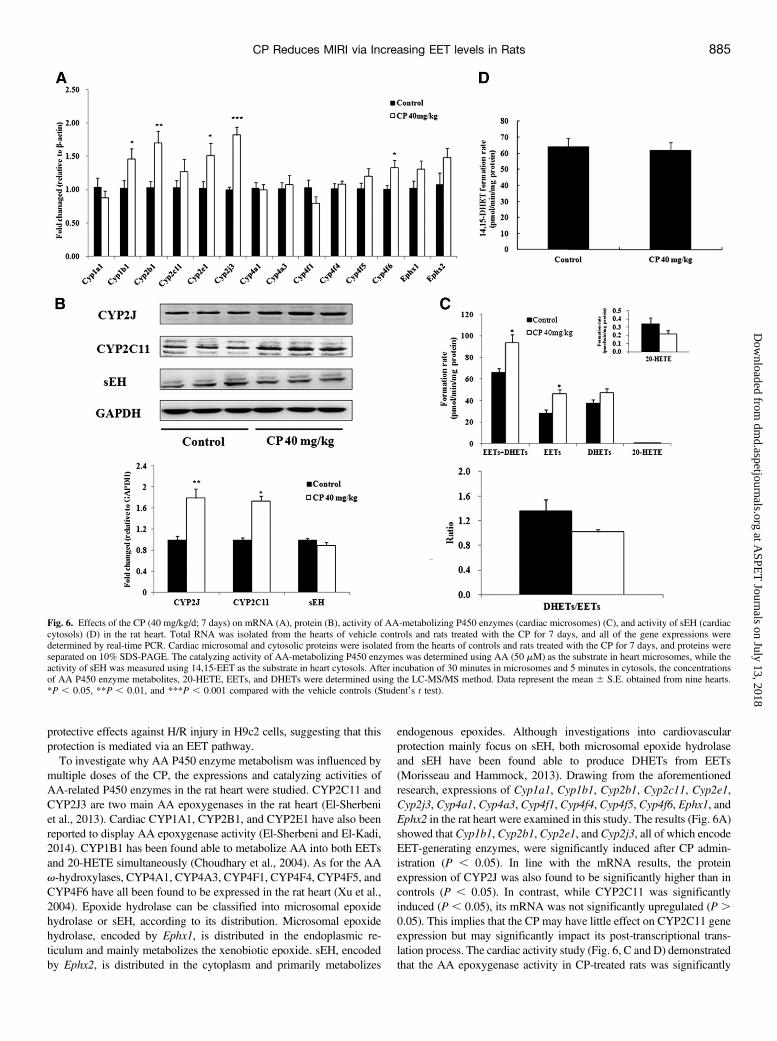
Effects of the CP on the Expression and Activity of AA-Metabolizing P450 Enzymes In Vivo. The effects of the CP on theexpressions and functions of AA-metabolizing P450 enzymes in normalrat hearts were investigated using real-time PCR, western blotting, andLC-MS/MS assay. The results indicate that after administration of theCP (40 mg/kg/d) for 7 days, expressions of cardiac Cyp1b1, Cyp2b1,Cyp2e1, Cyp2j3, and Cyp4f6 (Fig. 6A) were significantly inducedcompared with vehicle controls (P, 0.05). The western blotting resultsshowed that expressions of CYP2J and CYP2C11 proteins weresignificantly elevated (Fig. 6B) compared with controls (P , 0.05).The catalyzing activity of AA-metabolizing P450 enzymes was de-termined using AA (50 mM) as the substrate in heart microsomesprepared from vehicle or CP-treated rats (detailed results are given inSupplemental Table 4). As shown in Fig. 6C, no significant differencewas found in the concentrations of 20-HETE, DHETs, and the ratio ofDHETs/EETs between the two groups. However, the concentrations ofDHETs + EETs and EETs of CP-treated rats were significantly higher(P, 0.05) than those of the controls, indicating that the total activity of AAepoxygenases was increased after CP administration. However, the sEHactivity in cardiac cytosols of CP-treated rats, evaluated by 14,15-DHETgeneration, was not different from the controls (Fig. 6D).Effects of the CP on the H/R Injury and Concentrations of AA
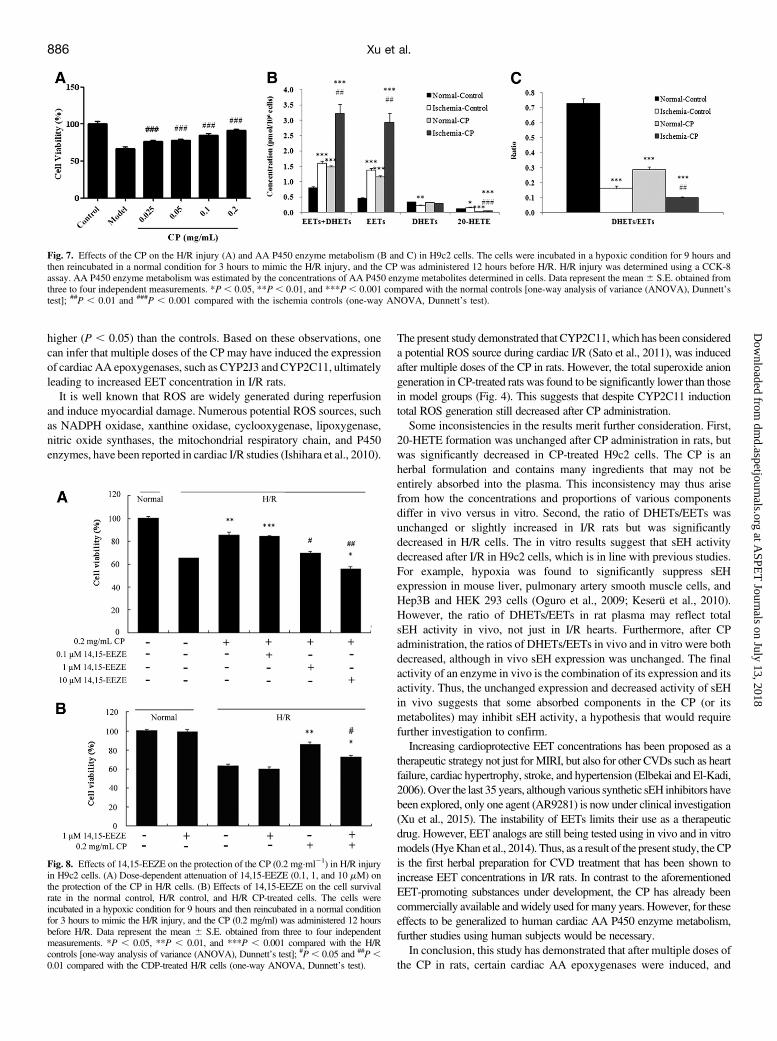
P450 Enzyme Metabolites In Vitro. The protective effect of the CP onH/R injury in cells was investigated in vitro. The results (Fig. 7A)demonstrated that the CP (0.025, 0.05, 0.1, and 0.2 mg/ml) caused adose-dependent improvement (66.2% 6 2.4% versus 76.1% 6 1.8%,77.9%6 1.4%, 84.5%6 2.1%, and 90.9%6 1.9%) in the cell survivalrate in H9c2 cells subjected to 9 hours of hypoxia and 3 hours ofreoxygenation. The concentrations of AA P450 enzyme metabolites innormal or H/R H9c2 cells were also determined (see SupplementalTable 5). As shown in Fig. 7, B and C, the concentrations of 20-HETE,DHETs + EETs, and EETs in cells after H/R were significantly higher
Fig. 1. Effects of the CP (10, 20, and 40 mg/kg/d; 7 days) on I/R-induced cardiac dysfunction. Data show the following: (A) heart rate (HR), (B) left-ventricular systolicpressure (LVSP), (C) left-ventricular end-diastolic pressure (LVEDP), (D) dp/dtmax, and (E)2dp/dtmax of hearts subjected to 45 minutes of ischemia followed by 24 hours ofreperfusion. Data represent the mean 6 S.E. obtained from six independent measurements. #P , 0.05, ##P , 0.01, and ###P , 0.001 compared with the sham group [one-way analysis of variance (ANOVA), Dunnett’s test]; *P , 0.05, **P , 0.01, and ***P , 0.001 compared with the model group (one-way ANOVA, Dunnett’s test).
882 Xu et al.
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from
(P, 0.05),while the values ofDHETs andDHETs/EETswere significantlylower (P , 0.01), than those in normal controls. After incubation withthe CP (0.2 mg/ml) for 12 hours, the values of 20-HETE and DHETs/EETs in normal H9c2 cells were significantly lower (P, 0.001), whilethe concentrations of EETs and DHETs + EETs were significantlyhigher (P, 0.001), than those in normal controls (Fig. 7, B and C). Asfor H/R cells subjected to 9 hours of hypoxia and 3 hours ofreoxygenation after pretreatment with the CP (0.2 mg/ml) for 12hours, the concentrations of EETs and DHETs + EETs weresignificantly higher (P , 0.01), while the values of 20-HETE andDHETs/EETs were significantly lower (P , 0.001), than those in thehypoxic controls.Effects of 14,15-EEZE on the CP’s Protection against H/R
Injury in H9c2 Cells. 14,15-EEZE, a structural analog of 14,15-EET,can compete with EETs for the same action site and play a role inantagonismofEETs.As shown in Fig. 8A, 14,15-EEZE (0.1, 1, and 10mM)dose dependently reduced the cell survival rate in CP-treated H/Rcells (0.2 mg/ml) from 85.8%6 4.8% to 84.3%6 1.2%, 70.1%6 2.6%,and 55.6% 6 4.3%, respectively. Furthermore, the effects of 14,15-EEZE (1 mM) on the cell survival rate in normal control, H/R control,and H/R CP-treated cells were also determined. The results (Fig. 8B)demonstrated that 14,15-EEZE (1 mM) had no effect on the cell survivalrate in normal control cells, slightly reduced the cell survival rate in H/Rcontrol cells (63.6% 6 2.4% versus 59.9% 6 3.9%, P . 0.05), and
significantly reduced the cell survival rate (85.8%6 4.2% versus 72.2%63.5%, P , 0.05) in CP-treated H/R cells (0.2 mg/ml).
Discussion
In this study, multiple doses of the CP were first found to have thefollowing dose-dependent effects in rats subjected to 45 minutes ofmyocardial ischemia and 24 hours of reperfusion (I/R rats): improvedhemodynamics and cardiac function, moderated variation in the myo-cardial zymogram, and decreased infarct size (Figs. 1–3). These resultssuggest that chronic administration of the CPmay have protective effectson MIRI.To explore the mechanism underlining this protection, the CP’s effect
on AA P450 enzyme metabolite concentrations was then investigated. Aprevious study in dogs (Nithipatikom et al., 2001) found that plasmaconcentrations of EETs, DHETs, and 20-HETE changed duringreperfusion periods of 5–120 minutes. Accordingly, levels of theseAAmetabolites were measured at 30 minutes and 3 hours of reperfusionin I/R rats. The results of the present study (Fig. 5) found that at 30minutes of reperfusion, plasma concentrations of AA P450 enzymemetabolites in the model group were slightly (but not significantly)higher than those in the sham group. However, after 3 hours ofreperfusion the concentrations of 20-HETE, DHETs, and EETs +DHETs in the model group were significantly higher (P , 0.05) thanthe sham group, suggesting that thesemetabolites increase in I/R rats in asimilar pattern to that found in dogs (Nithipatikom et al., 2001). Afterchronic CP administration, the concentration of EETs in I/R ratsincreased earlier and more dramatically than in the model group (Fig. 5).Because of the vasodilatory, anti-inflammatory, proangiogenic, and
antiapoptosis actions of EETs, augmented EET concentrations duringI/R has been considered a protective mechanism (Seubert et al., 2004).Increased EET levels during I/R has proven beneficial in numerousstudies. For example, CYP2J2 transgenic mice exhibited not onlyincreased EET biosynthesis in coronary arteries, but also improvedpostischemic recovery of left ventricular function after myocardialI/R (Seubert et al., 2004). sEH inhibition through gene deletion orpharmacological suppression was protective against MIRI in mice(Motoki et al., 2008). Administration of exogenous 14,15-EET wasfound to reduce MIRI in rats and dogs (Nithipatikom et al., 2006; Gross
Fig. 2. Effects of the CP (10, 20, and 40 mg/kg/d; 7 days) on the I/R-inducedvariations in the myocardial zymogram. Data show the following: (A) LDH and (B)CK-MB of hearts subjected to 45 minutes of ischemia followed by 24 hours ofreperfusion. Data represent the mean 6 S.E. obtained from six independentmeasurements. ##P , 0.01 and ###P , 0.001 compared with the sham group [one-way analysis of variance (ANOVA), Dunnett’s test]; ***P , 0.001 compared withthe model group (one-way ANOVA, Dunnett’s test).
Fig. 3. Effects of the CP (10, 20, and 40 mg/kg/d; 7 days) on myocardial infarctionin rats subjected to 45 minutes of ischemia and 24 hours of reperfusion. Myocardialinfarction was determined by TTC staining. Data represent the mean 6 S.E.obtained from six independent measurements. ###P , 0.001 compared with thesham group [one-way analysis of variance (ANOVA), Dunnett’s test]; **P , 0.01and ***P , 0.001 compared with the model group (one-way ANOVA, Dunnett’stest).
CP Reduces MIRI via Increasing EET levels in Rats 883
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from
et al., 2007). Likewise, exogenous administration of EET analogs, UA-8, provided significant protection against MIRI (Batchu et al., 2011).The results of the present study (Fig. 5) have demonstrated that after CPadministration, total activity of AA epoxygenases (EETs + DHETs)increased, while sEH activity (DHETs/EETs) decreased, ultimatelyleading to elevated EET plasma concentrations. The results also indicatedthat multiple doses of the CP had little effect on AA v-hydroxylation(20-HETE) in I/R rats.The protection of the CP against H/R injury and its effect on AA P450
enzyme metabolism were further investigated in H9c2 cells. The H9c2cell H/Rmodel is a common cell model for simulating myocardial I/R inrats in vitro (Batchu et al., 2011). In line with the present in vivo study,
the CP exhibited dose-dependent protection on H/R injury and a similarimpact on AA P450 enzyme metabolism (Fig. 7). In drug-untreatedcells, concentrations of 20-HETE and EETs were significantly higherafter H/R, compared with non-H/R controls. As expected, this suggestedan increased generation of AA P450 enzyme metabolites after H/R.However, in CP-pretreated cells EET concentrations were significantlyhigher than in control cells under both normal and H/R conditions(Fig. 7B). Furthermore, sEH activity was found to decrease after CPpretreatment. 14,15-EEZE, a structural analog of 14,15-EET, behaves asa direct EET antagonist at a receptor binding site, and selectivelyantagonizes many EET actions (Gross et al., 2008). The results of thisstudy found that 14,15-EEZE dose-dependently reversed the CP’s
Fig. 4. Effects of the CP (10, 20, and 40 mg/kg/d; 7 days) on the superoxide generation in the rat’s heart subjected to 45 minutes of ischemia and 15 minutes of reperfusion.Cryosections of hearts after reperfusion were incubated with dihydroethidium (10 mM) at 37�C in the dark for 30 minutes, and the formation of HE was determined byfluorescence microscopy. (A) Three representative images (magnification: 20�) from each group are shown. (B) Mean fluorescence intensity of each group. Data representthe mean 6 S.E. obtained from three independent measurements. ###P , 0.001 compared with the sham group [one-way analysis of variance (ANOVA), Dunnett’s test];*P , 0.05 and **P , 0.01compared with the model group (one-way ANOVA, Dunnett’s test).
Fig. 5. Effects of the CP (40 mg/kg/d; 7days) on the plasma concentrations of AA P450 enzyme metabolites in rats subjected to 45 minutes of ischemia, and 30 minutes (A)or 3 hours (B) of reperfusion. (C) Summarized concentrations of the AA P450 enzyme metabolites and (D) DHET/EET ratios of each group. The concentrations of AA P450enzyme metabolites were determined using the LC-MS/MS method. Data represent the mean 6 S.E. obtained from seven independent measurements (hearts). #P , 0.05,##P , 0.01, and ###P , 0.001 compared with the sham group [one-way analysis of variance (ANOVA), Dunnett’s test] at the same time; *P , 0.05, **P , 0.01, and ***P ,0.001 compared with the model group (one-way ANOVA, Dunnett’s test) at the same time; nP, 0.05 compared with the value of 30 minutes (one-way ANOVA, Dunnett’s test)in the same group.
884 Xu et al.
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from
protective effects against H/R injury in H9c2 cells, suggesting that thisprotection is mediated via an EET pathway.To investigate why AA P450 enzyme metabolism was influenced by
multiple doses of the CP, the expressions and catalyzing activities ofAA-related P450 enzymes in the rat heart were studied. CYP2C11 andCYP2J3 are two main AA epoxygenases in the rat heart (El-Sherbeniet al., 2013). Cardiac CYP1A1, CYP2B1, and CYP2E1 have also beenreported to display AA epoxygenase activity (El-Sherbeni and El-Kadi,2014). CYP1B1 has been found able to metabolize AA into both EETsand 20-HETE simultaneously (Choudhary et al., 2004). As for the AAv-hydroxylases, CYP4A1, CYP4A3, CYP4F1, CYP4F4, CYP4F5, andCYP4F6 have all been found to be expressed in the rat heart (Xu et al.,2004). Epoxide hydrolase can be classified into microsomal epoxidehydrolase or sEH, according to its distribution. Microsomal epoxidehydrolase, encoded by Ephx1, is distributed in the endoplasmic re-ticulum and mainly metabolizes the xenobiotic epoxide. sEH, encodedby Ephx2, is distributed in the cytoplasm and primarily metabolizes
endogenous epoxides. Although investigations into cardiovascularprotection mainly focus on sEH, both microsomal epoxide hydrolaseand sEH have been found able to produce DHETs from EETs(Morisseau and Hammock, 2013). Drawing from the aforementionedresearch, expressions of Cyp1a1, Cyp1b1, Cyp2b1, Cyp2c11, Cyp2e1,Cyp2j3, Cyp4a1, Cyp4a3, Cyp4f1, Cyp4f4, Cyp4f5, Cyp4f6, Ephx1, andEphx2 in the rat heart were examined in this study. The results (Fig. 6A)showed that Cyp1b1, Cyp2b1, Cyp2e1, and Cyp2j3, all of which encodeEET-generating enzymes, were significantly induced after CP admin-istration (P , 0.05). In line with the mRNA results, the proteinexpression of CYP2J was also found to be significantly higher than incontrols (P , 0.05). In contrast, while CYP2C11 was significantlyinduced (P , 0.05), its mRNA was not significantly upregulated (P .0.05). This implies that the CP may have little effect on CYP2C11 geneexpression but may significantly impact its post-transcriptional trans-lation process. The cardiac activity study (Fig. 6, C and D) demonstratedthat the AA epoxygenase activity in CP-treated rats was significantly
Fig. 6. Effects of the CP (40 mg/kg/d; 7 days) on mRNA (A), protein (B), activity of AA-metabolizing P450 enzymes (cardiac microsomes) (C), and activity of sEH (cardiaccytosols) (D) in the rat heart. Total RNA was isolated from the hearts of vehicle controls and rats treated with the CP for 7 days, and all of the gene expressions weredetermined by real-time PCR. Cardiac microsomal and cytosolic proteins were isolated from the hearts of controls and rats treated with the CP for 7 days, and proteins wereseparated on 10% SDS-PAGE. The catalyzing activity of AA-metabolizing P450 enzymes was determined using AA (50 mM) as the substrate in heart microsomes, while theactivity of sEH was measured using 14,15-EET as the substrate in heart cytosols. After incubation of 30 minutes in microsomes and 5 minutes in cytosols, the concentrationsof AA P450 enzyme metabolites, 20-HETE, EETs, and DHETs were determined using the LC-MS/MS method. Data represent the mean 6 S.E. obtained from nine hearts.*P , 0.05, **P , 0.01, and ***P , 0.001 compared with the vehicle controls (Student’s t test).
CP Reduces MIRI via Increasing EET levels in Rats 885
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from
higher (P , 0.05) than the controls. Based on these observations, onecan infer that multiple doses of the CP may have induced the expressionof cardiac AA epoxygenases, such as CYP2J3 andCYP2C11, ultimatelyleading to increased EET concentration in I/R rats.It is well known that ROS are widely generated during reperfusion
and induce myocardial damage. Numerous potential ROS sources, suchas NADPH oxidase, xanthine oxidase, cyclooxygenase, lipoxygenase,nitric oxide synthases, the mitochondrial respiratory chain, and P450enzymes, have been reported in cardiac I/R studies (Ishihara et al., 2010).
The present study demonstrated that CYP2C11, which has been considereda potential ROS source during cardiac I/R (Sato et al., 2011), was inducedafter multiple doses of the CP in rats. However, the total superoxide aniongeneration in CP-treated rats was found to be significantly lower than thosein model groups (Fig. 4). This suggests that despite CYP2C11 inductiontotal ROS generation still decreased after CP administration.Some inconsistencies in the results merit further consideration. First,
20-HETE formation was unchanged after CP administration in rats, butwas significantly decreased in CP-treated H9c2 cells. The CP is anherbal formulation and contains many ingredients that may not beentirely absorbed into the plasma. This inconsistency may thus arisefrom how the concentrations and proportions of various componentsdiffer in vivo versus in vitro. Second, the ratio of DHETs/EETs wasunchanged or slightly increased in I/R rats but was significantlydecreased in H/R cells. The in vitro results suggest that sEH activitydecreased after I/R in H9c2 cells, which is in line with previous studies.For example, hypoxia was found to significantly suppress sEHexpression in mouse liver, pulmonary artery smooth muscle cells, andHep3B and HEK 293 cells (Oguro et al., 2009; Keserü et al., 2010).However, the ratio of DHETs/EETs in rat plasma may reflect totalsEH activity in vivo, not just in I/R hearts. Furthermore, after CPadministration, the ratios of DHETs/EETs in vivo and in vitro were bothdecreased, although in vivo sEH expression was unchanged. The finalactivity of an enzyme in vivo is the combination of its expression and itsactivity. Thus, the unchanged expression and decreased activity of sEHin vivo suggests that some absorbed components in the CP (or itsmetabolites) may inhibit sEH activity, a hypothesis that would requirefurther investigation to confirm.Increasing cardioprotective EET concentrations has been proposed as a
therapeutic strategy not just for MIRI, but also for other CVDs such as heartfailure, cardiac hypertrophy, stroke, and hypertension (Elbekai and El-Kadi,2006).Over the last 35 years, although various synthetic sEH inhibitors havebeen explored, only one agent (AR9281) is now under clinical investigation(Xu et al., 2015). The instability of EETs limits their use as a therapeuticdrug. However, EET analogs are still being tested using in vivo and in vitromodels (HyeKhan et al., 2014). Thus, as a result of the present study, the CPis the first herbal preparation for CVD treatment that has been shown toincrease EET concentrations in I/R rats. In contrast to the aforementionedEET-promoting substances under development, the CP has already beencommercially available andwidely used for many years. However, for theseeffects to be generalized to human cardiac AA P450 enzyme metabolism,further studies using human subjects would be necessary.In conclusion, this study has demonstrated that after multiple doses of
the CP in rats, certain cardiac AA epoxygenases were induced, and
Fig. 8. Effects of 14,15-EEZE on the protection of the CP (0.2 mg×ml21) in H/R injuryin H9c2 cells. (A) Dose-dependent attenuation of 14,15-EEZE (0.1, 1, and 10 mM) onthe protection of the CP in H/R cells. (B) Effects of 14,15-EEZE on the cell survivalrate in the normal control, H/R control, and H/R CP-treated cells. The cells wereincubated in a hypoxic condition for 9 hours and then reincubated in a normal conditionfor 3 hours to mimic the H/R injury, and the CP (0.2 mg/ml) was administered 12 hoursbefore H/R. Data represent the mean 6 S.E. obtained from three to four independentmeasurements. *P , 0.05, **P , 0.01, and ***P , 0.001 compared with the H/Rcontrols [one-way analysis of variance (ANOVA), Dunnett’s test]; #P, 0.05 and ##P ,0.01 compared with the CDP-treated H/R cells (one-way ANOVA, Dunnett’s test).
Fig. 7. Effects of the CP on the H/R injury (A) and AA P450 enzyme metabolism (B and C) in H9c2 cells. The cells were incubated in a hypoxic condition for 9 hours andthen reincubated in a normal condition for 3 hours to mimic the H/R injury, and the CP was administered 12 hours before H/R. H/R injury was determined using a CCK-8assay. AA P450 enzyme metabolism was estimated by the concentrations of AA P450 enzyme metabolites determined in cells. Data represent the mean 6 S.E. obtained fromthree to four independent measurements. *P , 0.05, **P , 0.01, and ***P, 0.001 compared with the normal controls [one-way analysis of variance (ANOVA), Dunnett’stest]; ##P , 0.01 and ###P , 0.001 compared with the ischemia controls (one-way ANOVA, Dunnett’s test).
886 Xu et al.
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from

cardioprotective EET concentrations were increased following I/R. Theresults suggest that the mechanism underlying the CP’s protection inMIRI may be, at least in part, upregulation of EET-generating P450enzymes.
Acknowledgments
The authors are grateful for the helpful technical assistance from Li Liu,Jingyong Zhou, Jian Wu, Fei Ke, and Banghong Gu during the study.
Authorship ContributionsParticipated in research design: Li, Hao, Ju, Wang.Conducted experiments: Xu, Jiang, Wei, Zhang.Contributed new reagents or analytic tools: Sun, Ji.Performed data analysis: Xu, Zhou.Wrote or contributed to the writing of the manuscript: Xu, Hao.
References
Aboutabl ME, Zordoky BN, and El-Kadi AO (2009) 3-Methylcholanthrene and benzo(a)pyrenemodulate cardiac cytochrome P450 gene expression and arachidonic acid metabolism in maleSprague Dawley rats. Br J Pharmacol 158:1808–1819.
Alsaad AM, Zordoky BN, El-Sherbeni AA, and El-Kadi AO (2012) Chronic doxorubicin car-diotoxicity modulates cardiac cytochrome P450-mediated arachidonic acid metabolism in rats.Drug Metab Dispos 40:2126–2135.
Batchu SN, Lee SB, Qadhi RS, Chaudhary KR, El-Sikhry H, Kodela R, Falck JR, and Seubert JM(2011) Cardioprotective effect of a dual acting epoxyeicosatrienoic acid analogue towards is-chaemia reperfusion injury. Br J Pharmacol 162:897–907.
Campbell WB, Gebremedhin D, Pratt PF, and Harder DR (1996) Identification of epoxyeicosatrienoicacids as endothelium-derived hyperpolarizing factors. Circ Res 78:415–423.
Chen C and Wang DW (2013) CYP epoxygenase derived EETs: from cardiovascular protection tohuman cancer therapy. Curr Top Med Chem 13:1454–1469.
Choudhary D, Jansson I, Stoilov I, Sarfarazi M, and Schenkman JB (2004) Metabolism of retinoidsand arachidonic acid by human and mouse cytochrome P450 1B1. Drug Metab Dispos 32:840–847.
Delozier TC, Kissling GE, Coulter SJ, Dai D, Foley JF, Bradbury JA, Murphy E, Steenbergen C,Zeldin DC, and Goldstein JA (2007) Detection of human CYP2C8, CYP2C9, and CYP2J2 incardiovascular tissues. Drug Metab Dispos 35:682–688.
Elbekai RH and El-Kadi AO (2006) Cytochrome P450 enzymes: central players in cardiovascularhealth and disease. Pharmacol Ther 112:564–587.
El-Sherbeni AA, Aboutabl ME, Zordoky BN, Anwar-Mohamed A, and El-Kadi AO (2013) De-termination of the dominant arachidonic acid cytochrome P450 monooxygenases in rat heart,lung, kidney, and liver: protein expression and metabolite kinetics. AAPS J 15:112–122.
El-Sherbeni AA and El-Kadi AO (2014) Characterization of arachidonic acid metabolism by ratcytochrome P450 enzymes: the involvement of CYP1As. Drug Metab Dispos 42:1498–1507.
Enayetallah AE, French RA, Thibodeau MS, and Grant DF (2004) Distribution of soluble epoxidehydrolase and of cytochrome P450 2C8, 2C9, and 2J2 in human tissues. J Histochem Cytochem52:447–454.
Fleming I, Michaelis UR, Bredenkötter D, Fisslthaler B, Dehghani F, Brandes RP, and Busse R(2001) Endothelium-derived hyperpolarizing factor synthase (cytochrome P450 2C9) is afunctionally significant source of reactive oxygen species in coronary arteries. Circ Res 88:44–51.
Granville DJ, Tashakkor B, Takeuchi C, Gustafsson AB, Huang C, Sayen MR, Wentworth P, Jr,Yeager M, and Gottlieb RA (2004) Reduction of ischemia and reperfusion-induced myocardialdamage by cytochrome P450 inhibitors. Proc Natl Acad Sci USA 101:1321–1326.
Gross GJ, Gauthier KM, Moore J, Falck JR, Hammock BD, Campbell WB, and Nithipatikom K(2008) Effects of the selective EET antagonist, 14,15-EEZE, on cardioprotection produced byexogenous or endogenous EETs in the canine heart. Am J Physiol Heart Circ Physiol 294:H2838–H2844.
Gross GJ, Hsu A, Falck JR, and Nithipatikom K (2007) Mechanisms by which epoxyeicosatrienoicacids (EETs) elicit cardioprotection in rat hearts. J Mol Cell Cardiol 42:687–691.
Hye Khan MA, Pavlov TS, Christain SV, Necká�r J, Staruschenko A, Gauthier KM, Capdevila JH,Falck JR, Campbell WB, and Imig JD (2014) Epoxyeicosatrienoic acid analogue lowers bloodpressure through vasodilation and sodium channel inhibition. Clin Sci (Lond) 127:463–474.
Ishihara Y, Sekine M, Hamaguchi A, Kobayashi Y, Harayama T, Nakazawa M, and Shimamoto N(2010) Effects of sulfaphenazole derivatives on cardiac ischemia-reperfusion injury: associationof cytochrome P450 activity and infarct size. J Pharmacol Sci 113:335–342.
Ishizuka T, Cheng J, Singh H, Vitto MD, Manthati VL, Falck JR, and Laniado-Schwartzman M(2008) 20-Hydroxyeicosatetraenoic acid stimulates nuclear factor-kB activation and the pro-duction of inflammatory cytokines in human endothelial cells. J Pharmacol Exp Ther 324:103–110.
Keserü B, Barbosa-Sicard E, Schermuly RT, Tanaka H, Hammock BD, Weissmann N, FisslthalerB, and Fleming I (2010) Hypoxia-induced pulmonary hypertension: comparison of solubleepoxide hydrolase deletion vs. inhibition. Cardiovasc Res 85:232–240.
Khan M, Mohan IK, Kutala VK, Kumbala D, and Kuppusamy P (2007) Cardioprotection bysulfaphenazole, a cytochrome P450 inhibitor: mitigation of ischemia-reperfusion injury byscavenging of reactive oxygen species. J Pharmacol Exp Ther 323:813–821.
Konkel A and Schunck WH (2011) Role of cytochrome P450 enzymes in the bioactivation ofpolyunsaturated fatty acids. Biochim Biophys Acta 1814:210–222.
Lei X, Chen J, Liu CX, Lin J, Lou J, and Shang HC (2014) Status and thoughts of Chinese patentmedicines seeking approval in the US market. Chin J Integr Med 20:403–408.
Liu P, Yang H, Long F, Hao HP, Xu X, Liu Y, Shi XW, Zhang DD, Zheng HC, and Wen QY, et al.(2014) Bioactive equivalence of combinatorial components identified in screening of an herbalmedicine. Pharm Res 31:1788–1800.
Lu T, Yang J, Gao X, Chen P, Du F, Sun Y, Wang F, Xu F, Shang H, and Huang Y, et al. (2008)Plasma and urinary tanshinol from Salvia miltiorrhiza (Danshen) can be used as pharmacokineticmarkers for cardiotonic pills, a cardiovascular herbal medicine. Drug Metab Dispos 36:1578–1586.
Luo J, Xu H, and Chen K (2013) Systematic review of compound Danshen dropping pill: a chinesepatent medicine for acute myocardial infarction. Evid Based Complement Alternat Med 2013:808076.
Martín-Venegas R, Casillas R, Jáuregui O, and Moreno JJ (2011) Rapid simultaneous analysis ofcyclooxygenase, lipoxygenase and cytochrome P-450 metabolites of arachidonic and linoleicacids using high performance liquid chromatography/mass spectrometry in tandem mode. JPharm Biomed Anal 56:976–982.
Miyata N and Roman RJ (2005) Role of 20-hydroxyeicosatetraenoic acid (20-HETE) in vascularsystem. J Smooth Muscle Res 41:175–193.
Morisseau C and Hammock BD (2013) Impact of soluble epoxide hydrolase and epoxyeicosanoidson human health. Annu Rev Pharmacol Toxicol 53:37–58.
Motoki A, Merkel MJ, Packwood WH, Cao Z, Liu L, Iliff J, Alkayed NJ, and Van Winkle DM(2008) Soluble epoxide hydrolase inhibition and gene deletion are protective against myocardialischemia-reperfusion injury in vivo. Am J Physiol Heart Circ Physiol 295:H2128–H2134.
Nilakantan V, Maenpaa C, Jia G, Roman RJ, and Park F (2008) 20-HETE-mediated cytotoxicityand apoptosis in ischemic kidney epithelial cells. Am J Physiol Renal Physiol 294:F562–F570.
Nithipatikom K, DiCamelli RF, Kohler S, Gumina RJ, Falck JR, Campbell WB, and Gross GJ(2001) Determination of cytochrome P450 metabolites of arachidonic acid in coronary venousplasma during ischemia and reperfusion in dogs. Anal Biochem 292:115–124.
Nithipatikom K, Gross ER, Endsley MP, Moore JM, Isbell MA, Falck JR, Campbell WB,and Gross GJ (2004) Inhibition of cytochrome P450v-hydroxylase: a novel endogenous car-dioprotective pathway. Circ Res 95:e65–e71.
Nithipatikom K, Moore JM, Isbell MA, Falck JR, and Gross GJ (2006) Epoxyeicosatrienoic acidsin cardioprotection: ischemic versus reperfusion injury. Am J Physiol Heart Circ Physiol 291:H537–H542.
Node K, Huo Y, Ruan X, Yang B, Spiecker M, Ley K, Zeldin DC, and Liao JK (1999) Anti-inflammatory properties of cytochrome P450 epoxygenase-derived eicosanoids. Science 285:1276–1279.
Oguro A, Sakamoto K, Suzuki S, and Imaoka S (2009) Contribution of hydrolase and phosphatasedomains in soluble epoxide hydrolase to vascular endothelial growth factor expression and cellgrowth. Biol Pharm Bull 32:1962–1967.
Oni-Orisan A, Alsaleh N, Lee CR, and Seubert JM (2014) Epoxyeicosatrienoic acids and car-dioprotection: the road to translation. J Mol Cell Cardiol 74:199–208.
Sato M, Yokoyama U, Fujita T, Okumura S, and Ishikawa Y (2011) The roles of cytochrome P450in ischemic heart disease. Curr Drug Metab 12:526–532.
Seubert J, Yang B, Bradbury JA, Graves J, Degraff LM, Gabel S, Gooch R, Foley J, Newman J,and Mao L, et al. (2004) Enhanced postischemic functional recovery in CYP2J2 transgenichearts involves mitochondrial ATP-sensitive K+ channels and p42/p44 MAPK pathway. CircRes 95:506–514.
Vriens J, Owsianik G, Fisslthaler B, Suzuki M, Janssens A, Voets T, Morisseau C, Hammock BD,Fleming I, and Busse R, et al. (2005) Modulation of the Ca2 permeable cation channel TRPV4by cytochrome P450 epoxygenases in vascular endothelium. Circ Res 97:908–915.
Wang H, Lin L, Jiang J, Wang Y, Lu ZY, Bradbury JA, Lih FB, Wang DW, and Zeldin DC (2003)Up-regulation of endothelial nitric-oxide synthase by endothelium-derived hyperpolarizing factorinvolves mitogen-activated protein kinase and protein kinase C signaling pathways. J PharmacolExp Ther 307:753–764.
Wang HX, Zhang DM, Zeng XJ, Mu J, Yang H, Lu LQ, and Zhang LK (2012) Upregulation ofcytochrome P450 2J3/11,12-epoxyeicosatrienoic acid inhibits apoptosis in neonatal rat car-diomyocytes by a caspase-dependent pathway. Cytokine 60:360–368.
Wei B, Li WW, Ji J, Hu QH, and Ji H (2014) The cardioprotective effect of sodium tanshinone IIAsulfonate and the optimizing of therapeutic time window in myocardial ischemia/reperfusioninjury in rats. Atherosclerosis 235:318–327.
Xu F, Falck JR, Ortiz de Montellano PR, and Kroetz DL (2004) Catalytic activity and isoform-specific inhibition of rat cytochrome P450 4F enzymes. J Pharmacol Exp Ther 308:887–895.
Xu M, Hao H, Jiang L, Long F, Wei Y, Ji H, Sun B, Peng Y, Wang G, and Ju W, et al. (2015) Invitro inhibitory effects of ethanol extract of Danshen (Salvia miltiorrhiza) and its components onthe catalytic activity of soluble epoxide hydrolase. Phytomedicine 22:444–451.
Xu X, Zhang XA, and Wang DW (2011) The roles of CYP450 epoxygenases and metabolites,epoxyeicosatrienoic acids, in cardiovascular and malignant diseases. Adv Drug Deliv Rev 63:597–609.
Zhang JH, Pearson T, Matharoo-Ball B, Ortori CA, Warren AY, Khan R, and Barrett DA (2007)Quantitative profiling of epoxyeicosatrienoic, hydroxyeicosatetraenoic, and dihydroxyeicosate-traenoic acids in human intrauterine tissues using liquid chromatography/electrospray ionizationtandem mass spectrometry. Anal Biochem 365:40–51.
Zordoky BN, Anwar-Mohamed A, Aboutabl ME, and El-Kadi AO (2011) Acute doxorubicintoxicity differentially alters cytochrome P450 expression and arachidonic acid metabolism in ratkidney and liver. Drug Metab Dispos 39:1440–1450.
Address correspondence to: Dr. Ping Li, China Pharmaceutical University, No. 24Tongjia Lane, Nanjing 210009, China. E-mail: [email protected] or Dr. WenzhengJu, Affiliated Hospital of Nanjing University of Traditional Chinese Medicine, No. 155Hanzhong Road, Nanjing 210029, China. E-mail: [email protected]
CP Reduces MIRI via Increasing EET levels in Rats 887
at ASPE
T Journals on July 13, 2018
dmd.aspetjournals.org
Dow
nloaded from