biologÍa celular y molecular capítulo 16....
TRANSCRIPT

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Cáncer
Capítulo 16
16.1 Propiedades básicas de una célula cancerosa 16.2 Causas del cáncer 16.3 Genética del cáncer 16.4 Nuevas estrategias para combatir el cáncer VÍAS EXPERIMENTALES: El descubrimiento de los oncogenes
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
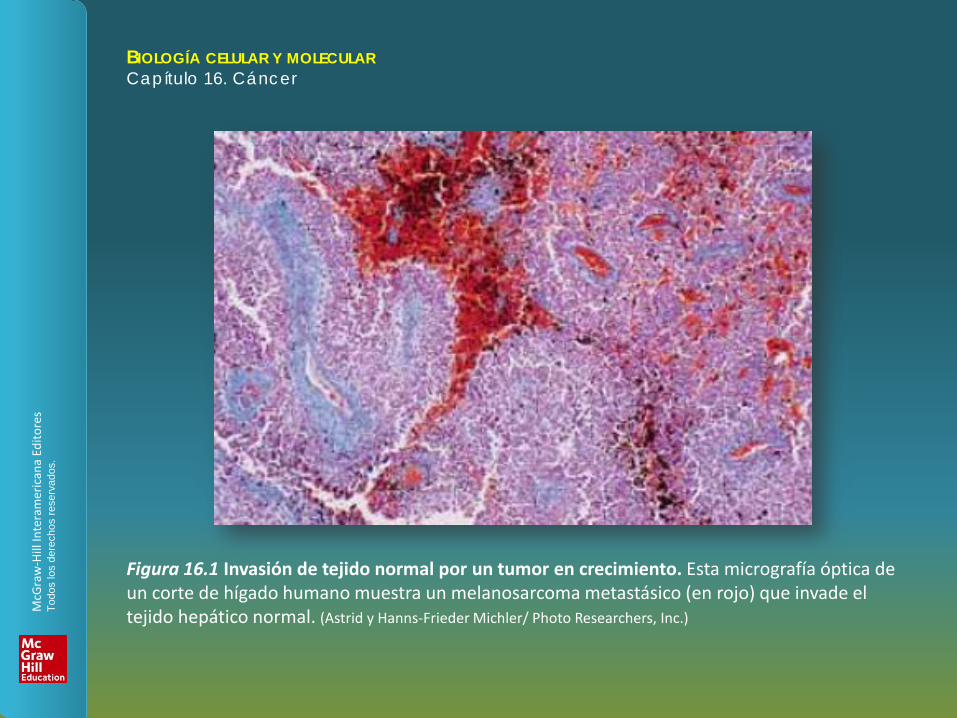
Figura 16.1 Invasión de tejido normal por un tumor en crecimiento. Esta micrografía óptica de un corte de hígado humano muestra un melanosarcoma metastásico (en rojo) que invade el tejido hepático normal. (Astrid y Hanns-Frieder Michler/ Photo Researchers, Inc.)
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
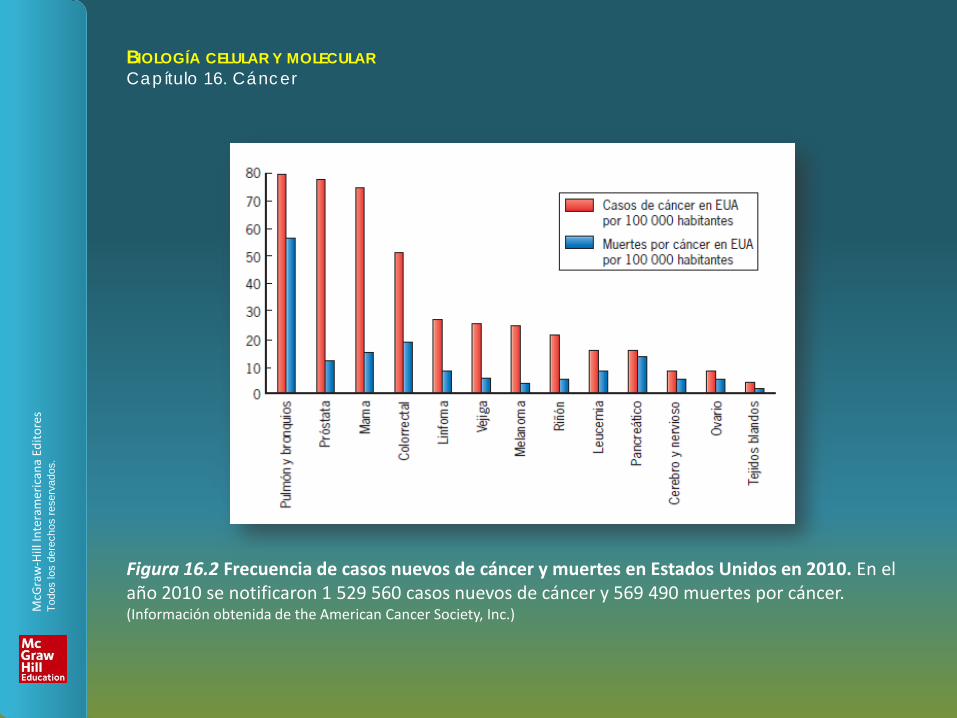
Figura 16.2 Frecuencia de casos nuevos de cáncer y muertes en Estados Unidos en 2010. En el año 2010 se notificaron 1 529 560 casos nuevos de cáncer y 569 490 muertes por cáncer. (Información obtenida de the American Cancer Society, Inc.)
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
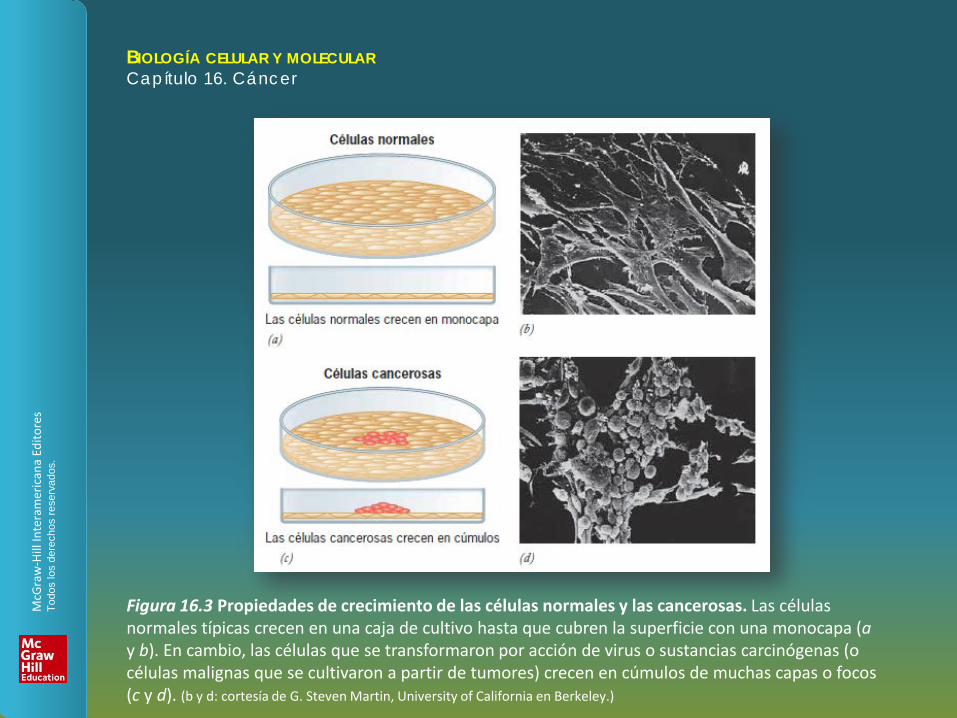
Figura 16.3 Propiedades de crecimiento de las células normales y las cancerosas. Las células normales típicas crecen en una caja de cultivo hasta que cubren la superficie con una monocapa (a y b). En cambio, las células que se transformaron por acción de virus o sustancias carcinógenas (o células malignas que se cultivaron a partir de tumores) crecen en cúmulos de muchas capas o focos (c y d). (b y d: cortesía de G. Steven Martin, University of California en Berkeley.)
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
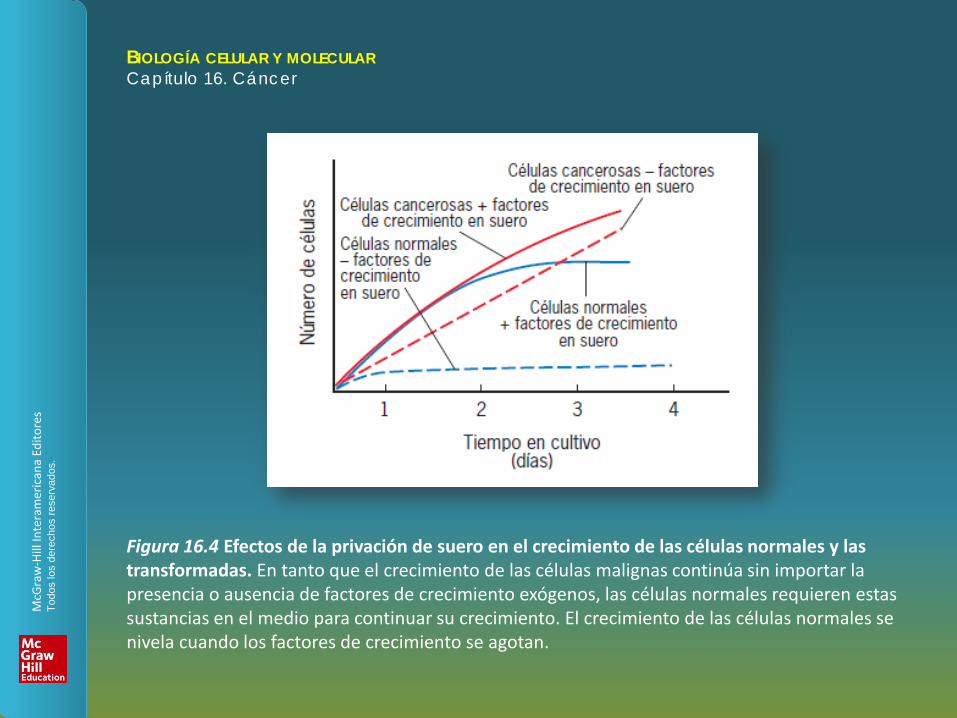
Figura 16.4 Efectos de la privación de suero en el crecimiento de las células normales y las transformadas. En tanto que el crecimiento de las células malignas continúa sin importar la presencia o ausencia de factores de crecimiento exógenos, las células normales requieren estas sustancias en el medio para continuar su crecimiento. El crecimiento de las células normales se nivela cuando los factores de crecimiento se agotan.

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.5 Cariotipo de una célula de una línea de cáncer mamario que muestra un complemento cromosómico muy anormal. Una célula diploide normal tendría 22 pares de autosomas y dos cromosomas sexuales. Los dos miembros de un par serían idénticos y cada cromosoma tendría un solo color continuo (como en el cariotipo de una célula normal en la figura 12.22b que utiliza la misma técnica de visualización espectral.) Los cromosomas de esta célula están muy alterados, como lo demuestra la presencia de cromosomas adicionales y faltantes (p. ej., aneuploidia), así como cromosomas con más de un color. Estos cromosomas multicolores reflejan la gran cantidad de translocaciones que ocurrieron en las generaciones celulares previas. Una célula con puntos de revisión normales en el ciclo celular y vías apoptóticas normales nunca alcanzaría un complemento cromosómico similar al observado aquí. (Cortesía de J. Davidson y Paul A. W. Edwards.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.6 Incidencia cambiante de cáncer en personas de ascendencia japonesa después de la inmigración a Hawai. La incidencia de cáncer estomacal declina, mientras que la de cáncer de mama y colon aumenta. Sin embargo, de los tres tipos de cáncer, sólo el de colon ha alcanzado tasas equivalentes a las de los hawaianos caucásicos en la segunda generación. (Tomada de L. N. Kolonel et al., reimpresa con autorización de Nature Revs. Cancer 4:3, 2004; copyright 2004. Nature Reviews Cancer por Nature Publishing Group. Reimpresa con autorización de Nature Publishing Groups en el formato de reutilización en un libro/libro de texto vía copyright Clearance Center.)
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
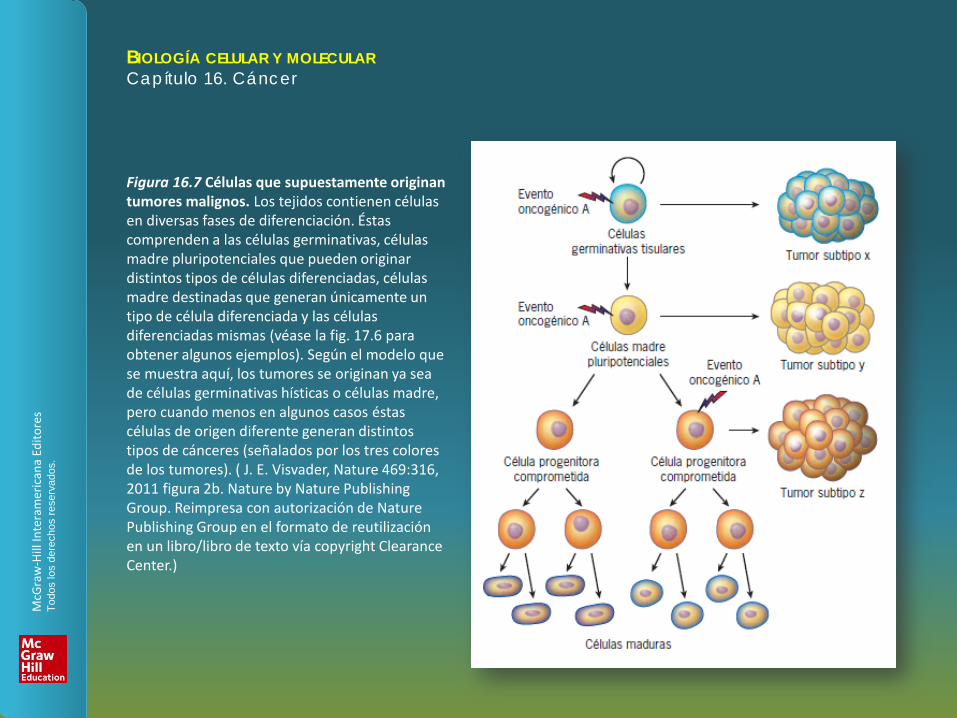
Figura 16.7 Células que supuestamente originan tumores malignos. Los tejidos contienen células en diversas fases de diferenciación. Éstas comprenden a las células germinativas, células madre pluripotenciales que pueden originar distintos tipos de células diferenciadas, células madre destinadas que generan únicamente un tipo de célula diferenciada y las células diferenciadas mismas (véase la fig. 17.6 para obtener algunos ejemplos). Según el modelo que se muestra aquí, los tumores se originan ya sea de células germinativas hísticas o células madre, pero cuando menos en algunos casos éstas células de origen diferente generan distintos tipos de cánceres (señalados por los tres colores de los tumores). ( J. E. Visvader, Nature 469:316, 2011 figura 2b. Nature by Nature Publishing Group. Reimpresa con autorización de Nature Publishing Group en el formato de reutilización en un libro/libro de texto vía copyright Clearance Center.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.8 Detección de células anormales (premalignas) en un extendido de Papanicolaou. (a) Células epiteliales escamosas normales del cuello uterino. Las células tienen una morfología uniforme con núcleo central y pequeño. (b) Células anormales de una tumoración in situ, un cáncer preinvasivo del cuello uterino. Las células tienen formas heterogéneas y núcleos grandes. (a: Dr. E. Walker/ Photo Researchers, Inc.; b: Spl/Photo Researchers, Inc.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.9 Efectos contrastantes de las mutaciones en los genes supresores tumorales (a) y los oncogenes (b). Mientras que una mutación en una de las dos copias (alelos) de un oncogén puede ser suficiente para hacer que la célula pierda el control del crecimiento, para inducir el mismo efecto deben eliminarse ambas copias de un gen supresor tumoral. Como se explica en breve, los oncogenes surgen de protooncogenes como efecto de mutaciones con ganancia de función, esto es, mutaciones que hacen que el producto del gen tenga nuevas funciones que conducen a la transformación maligna. Por el contrario, los genes supresores tumorales sufren mutaciones con pérdida de función o desactivación epigenética que los vuelven incapaces de limitar el crecimiento celular.

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.10 Activación de un protooncogén a un oncogén. La activación puede realizarse de varias maneras, como se indica en esta figura. En la vía a, una mutación del gen altera la estructura y función de la proteína codificada. En la vía b, la amplificación del gen produce una expresión excesiva de éste. En la vía c, un reordenamiento del DNA pone un nuevo segmento de DNA cerca o junto al gen, lo que altera su expresión o la estructura de la proteína codificada.

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.11 Mutaciones en el gen RB que pueden conducir al retinoblastoma. (a) En casos esporádicos (no familiares) de la enfermedad, un individuo comienza su vida con dos copias normales del gen RB en el cigoto y el retinoblastoma se desarrolla sólo en los raros sujetos en los que una célula retiniana determinada acumula mutaciones independientes en ambos alelos del gen. (b) En los casos familiares (hereditarios) de la enfermedad, una persona comienza su vida con un alelo anormal del gen RB, casi siempre una deleción. Por lo tanto, todas las células de la retina tienen por lo menos uno de sus genes RB sin función. Si el otro alelo RB en una célula retiniana se desactiva, las más de las veces como resultado de una mutación puntual, esa célula da origen a un tumor retiniano.
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
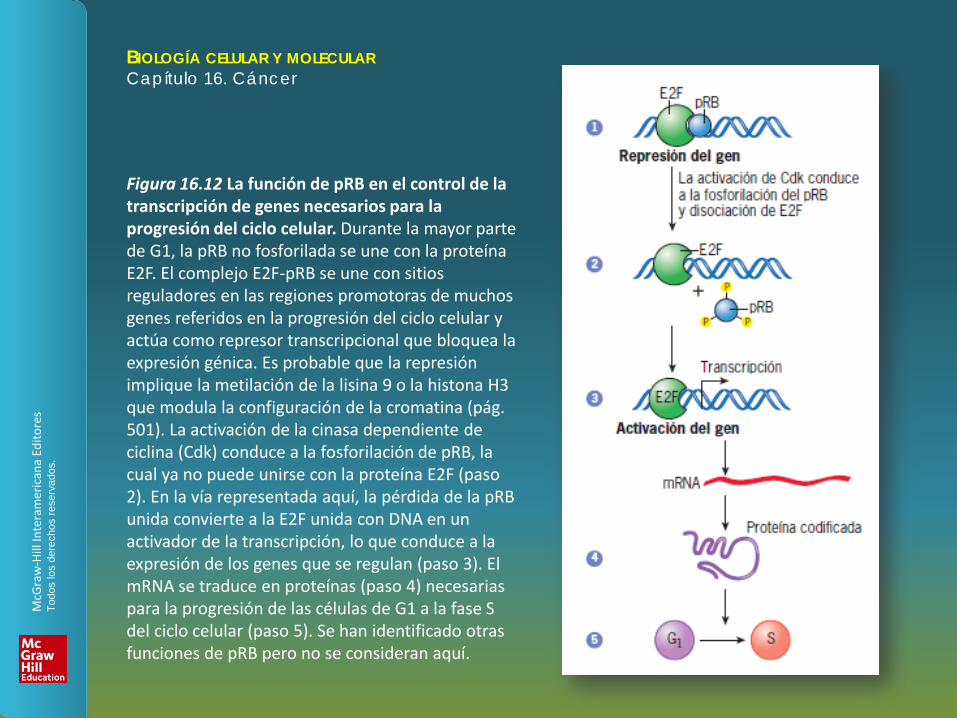
Figura 16.12 La función de pRB en el control de la transcripción de genes necesarios para la progresión del ciclo celular. Durante la mayor parte de G1, la pRB no fosforilada se une con la proteína E2F. El complejo E2F-pRB se une con sitios reguladores en las regiones promotoras de muchos genes referidos en la progresión del ciclo celular y actúa como represor transcripcional que bloquea la expresión génica. Es probable que la represión implique la metilación de la lisina 9 o la histona H3 que modula la configuración de la cromatina (pág. 501). La activación de la cinasa dependiente de ciclina (Cdk) conduce a la fosforilación de pRB, la cual ya no puede unirse con la proteína E2F (paso 2). En la vía representada aquí, la pérdida de la pRB unida convierte a la E2F unida con DNA en un activador de la transcripción, lo que conduce a la expresión de los genes que se regulan (paso 3). El mRNA se traduce en proteínas (paso 4) necesarias para la progresión de las células de G1 a la fase S del ciclo celular (paso 5). Se han identificado otras funciones de pRB pero no se consideran aquí.

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.13 Función del gen supresor tumoral TP53 en el cáncer humano. (a) La frecuencia con que ambos alelos del gen TP53 mutan en los diversos tipos de cánceres. La información se refiere a la variedad más frecuente de cada uno de estos 10 tipos de cánceres. (b) La función de p53 es especialmente sensible a las mutaciones en su dominio de unión a DNA; p53 funciona como tetrámero y cada subunidad consta de varios dominios con diferentes funciones. En esta imagen se muestra un listón del dominio de unión a DNA. Los seis residuos de aminoácidos que mutan con más frecuencia en las moléculas p53 que se han debilitado en los cánceres humanos se señalan con una sola letra (fig. 2.26). Estos residuos aparecen en o cerca de la interfase proteína-DNA y repercuten directamente en la unión de la proteína con el DNA o bien alteran su conformación. (a: Cosmic v54 Release Forbes et al., 2011. mar 39, D945,’11 base de datos. Cell 148:412, derechos reservados 2012 Elsevier Inc. Cell por Cell Press. Reimpresa con autorización de Cell Press en el formato de reutilización en libros/libros de texto vía copyright Clearence Center; b: tomado de Y. Cho, S. Gorina, P. D. Jeffrey, N. P. Pavletich, Science 265:352, 1994. Reimpresa con autorización de AAAS.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.14 Un modelo de la función de p53. (a) La división celular normal no requiere la participación de p53. (b) Empero, si el DNA de una célula se daña como resultado de la exposición a mutágenos, el nivel de p53 se eleva y actúa para detener la progresión de la célula por G1 o dirigir la célula hacia la apoptosis. (c) Si se desactivan ambas copias de TP53, la célula pierde la capacidad para detener el ciclo celular o derivar la célula hacia la apoptosis después del daño del DNA. Como resultado, la célula muere por falla de la mitosis o continúa su proliferación con anomalías genéticas, lo cual puede conducirla a la formación de una neoplasia maligna. (D. P. Lane, reimpresa con autorización de Nature 358:15, 1992. Derechos reservados 1992. Nature by Nature Publishing Group. Reimpreso con autorización de Nature Publishing Group en el formato de reutilización en un libro/libro de texto vía copyright Clearance Center.)
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
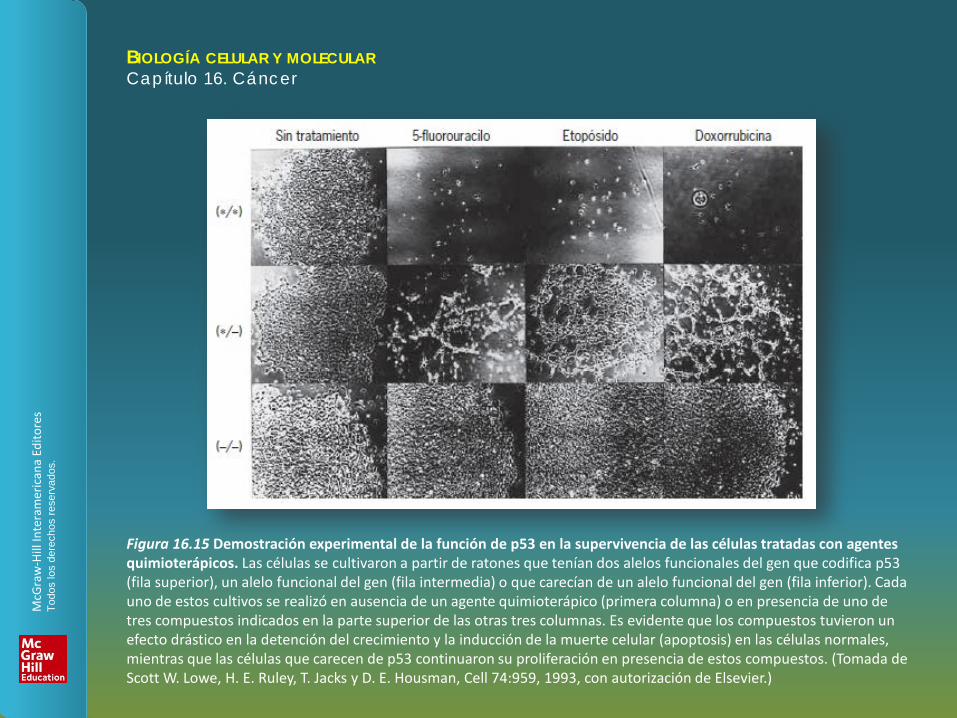
Figura 16.15 Demostración experimental de la función de p53 en la supervivencia de las células tratadas con agentes quimioterápicos. Las células se cultivaron a partir de ratones que tenían dos alelos funcionales del gen que codifica p53 (fila superior), un alelo funcional del gen (fila intermedia) o que carecían de un alelo funcional del gen (fila inferior). Cada uno de estos cultivos se realizó en ausencia de un agente quimioterápico (primera columna) o en presencia de uno de tres compuestos indicados en la parte superior de las otras tres columnas. Es evidente que los compuestos tuvieron un efecto drástico en la detención del crecimiento y la inducción de la muerte celular (apoptosis) en las células normales, mientras que las células que carecen de p53 continuaron su proliferación en presencia de estos compuestos. (Tomada de Scott W. Lowe, H. E. Ruley, T. Jacks y D. E. Housman, Cell 74:959, 1993, con autorización de Elsevier.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.16 El daño en el DNA inicia la actividad de varias proteínas codificadas por genes supresores tumorales y protooncogenes. En esta figura se advierte que el daño al DNA causa roturas en la doble cadena del DNA (paso 1) que se reparan por un complejo multiproteínico que incluye BRCA1 y BRCA2 (paso 2a). Las mutaciones en los genes que codifican estas proteínas pueden bloquear el proceso de reparación (paso 2b). Si no se repara el daño en el DNA, se activa un punto de comprobación que conduce al incremento del nivel de actividad de p53 (paso 3a). La proteína p53 normal se inhibe por la interacción con la proteína MDM2 (paso 3b). El p53 es un factor de transcripción que activa la expresión de: (1) el gen p21 (paso 4a), cuyo producto (p21) detiene el ciclo celular o (2) el gen BAX (paso 4b), cuyo producto (Bax) conduce a la apoptosis. La activación de p53 también puede fomentar el envejecimiento celular, pero no se conoce la vía. (Reimpresa con autorización de J. Brugarolas y T. Jacks. Nature Med 3:721, 1997, copyright 1997, Nature Medicine by Nature Publishing Group. Reimpresa con autorización de Nature Publishing Group en el formato de reutilización en un libro/libro de texto vía Copyright Clearance Center.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.17 Esquema que resume los tipos de proteínas codificadas por los protooncogenes. Éstas incluyen factores de crecimiento (1), receptores para factores de crecimiento (2), proteína cinasas y las proteínas que las activan (3), proteínas que regulan el ciclo celular (4), factores de transcripción (5), proteínas que modifican la cromatina (6), enzimas metabólicas (7), y proteínas que inhiben la apoptosis (8). No se muestran las proteínas que participan en la mitosis, invasión de los tejidos y metástasis.

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.18 Generalidades de varias vías de señalización implicadas en la oncogénesis que se explican en esta sección. Los supresores tumorales y la supresión tumoral se presentan en rojo, mientras que los oncogenes y la estimulación tumoral están en azul. Las flechas indican activación, las líneas perpendiculares inhibición. Entre las proteínas mostradas en esta figura, están factores de transcripción (p53, MYC y E2F), un coactivador o correpresor de la transcripción (pRB), una cinasa de lípidos (PI3K) y fosfatasa de lípidos (PTEN), una tirosina cinasa citoplásmica (RAF) y su activador (RAS), una GTP-asa que activa la proteína RAS (NF1), una cinasa que fomenta la supervivencia celular (PKB/AKT), una proteína que percibe las roturas en el DNA (BRCA), subunidades de una cinasa dependiente de ciclina (CICLINA D1 y CDK4), un inhibidor de Cdk (p21), una proteína antiapoptótica (BCL-2), una ligasa de ubicuitina (MDM2), una enzima que prolonga el DNA (telomerasa) y una proteína que se une con factores de crecimiento (p. ej., EGFR). Las flechas y líneas no necesariamente señalan la activación o inhibición directas. Por ejemplo, PTEN inhibe a PKB eliminando un fosfato de PIP3 y EGFR activa a RAS a través de GRB2 y SOS. La línea punteada indica una acción directa por activación de la expresión del gen MYC.

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.19 El paisaje genómico de los cánceres colorrectales. Estos mapas bidimensionales muestran los genes mutados con mayor frecuencia en los tumores colorrectales. Cada proyección rojiza representa un gen diferente. Los cinco genes que están mutados en un alto porcentaje de tumores están representados por proyecciones más altas conocidas como “montañas”, y reciben nombres específicos. Los 50 o más genes mutados con frecuencia mucho menor constituyen las “colinas” más pequeñas del paisaje genómico. Para mostrar el grado en que los tumores colorrectales de distintos pacientes comparten genes mutados comunes, en esta ilustración se presentan los paisajes de mutación de dos tumores individuales (identificados como Mx38 y Mx32). Los genes que presentaron mutación somática en los dos tumores individuales están indicados con círculos blancos. Resulta evidente que hay muy pocas mutaciones compartidas entre los tumores de estas dos personas. En el ejemplo mostrado, sólo los genes APC y TP53 están mutados en ambos casos de enfermedad. (Nota: las posiciones de los genes en este paisaje bidimensional están ordenadas con loci de un extremo del cromosoma 1 en la parte inferior izquierda del paisaje, proceden de cada uno de los autosomas en orden ascendente, hasta que al final llega a los loci del cromosoma X en el borde derecho del paisaje.) (Tomada de Laura D. Wood et al., por cortesía de Bert Vogelstein, Science 318:1113, 2007; copyright 2007, reimpresa con autorización de AAAS.)
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
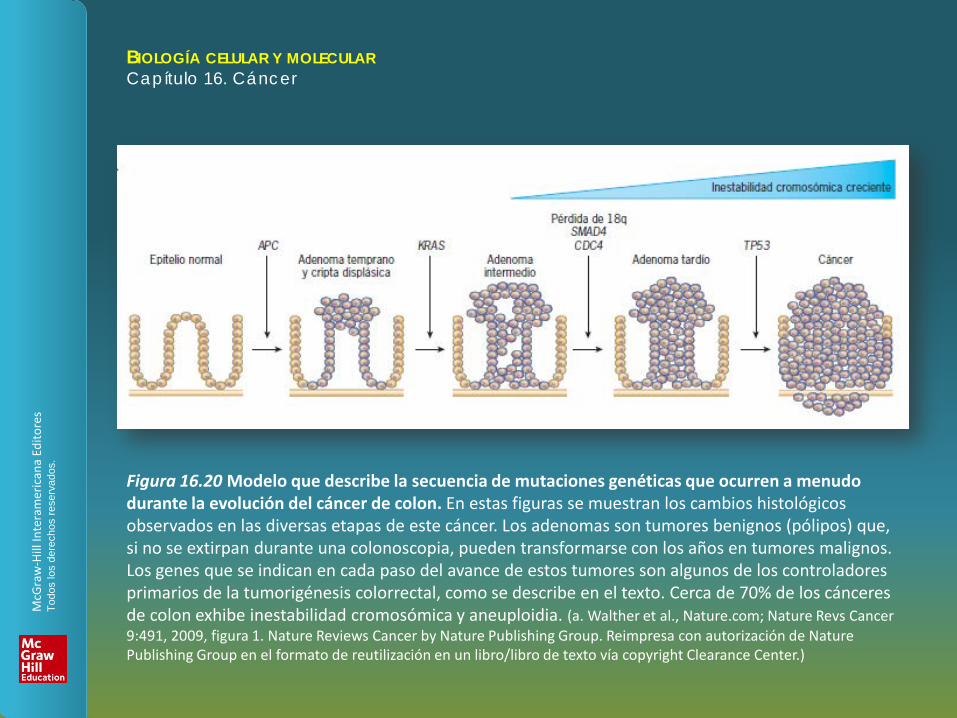
Figura 16.20 Modelo que describe la secuencia de mutaciones genéticas que ocurren a menudo durante la evolución del cáncer de colon. En estas figuras se muestran los cambios histológicos observados en las diversas etapas de este cáncer. Los adenomas son tumores benignos (pólipos) que, si no se extirpan durante una colonoscopia, pueden transformarse con los años en tumores malignos. Los genes que se indican en cada paso del avance de estos tumores son algunos de los controladores primarios de la tumorigénesis colorrectal, como se describe en el texto. Cerca de 70% de los cánceres de colon exhibe inestabilidad cromosómica y aneuploidia. (a. Walther et al., Nature.com; Nature Revs Cancer 9:491, 2009, figura 1. Nature Reviews Cancer by Nature Publishing Group. Reimpresa con autorización de Nature Publishing Group en el formato de reutilización en un libro/libro de texto vía copyright Clearance Center.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.21 Perfil de expresión génica que distingue dos tipos de leucemia. Cada fila muestra el nivel de expresión de un solo gen cuyo nombre aparece a la derecha de la fila. Se indican los niveles de expresión de 50 genes distintos. La clave de color se muestra al pie de la figura e indica que el nivel más bajo de expresión es el azul oscuro y el más alto el rojo intenso. Cada columna representa los datos de una muestra (paciente) diferente. Las columnas de la izquierda muestran los perfiles de expresión de personas con leucemia linfoblástica aguda (ALL), mientras que las columnas de la derecha señalan los perfiles de expresión de individuos con leucemia mieloide aguda (AML) (indicadas por las llaves en la parte superior). Resulta evidente que los genes del cuadro superior se expresan con un nivel mucho mayor en sujetos con ALL, mientras que los genes del cuadro inferior se expresan en un nivel mucho más alto en pacientes con AML. (Los genes incluidos en la figura se eligieron por estas diferencias en la expresión entre ambas enfermedades.) (Tomada de T. R. Golub et al., Science 286:534, 1999; copyright 1999, Science por Moses King. Reimpresa con autorización de the American Association for the Advancement of Science en el formato de reutilización en un libro/libro de texto vía copyright Clearance Center.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.22 Uso de los datos de una micromatriz de DNA para elegir el tratamiento. Cada gráfica muestra el índice de supervivencia respecto del tiempo de pacientes con cáncer mamario que tenían buen o mal pronóstico, de acuerdo con el nivel de expresión de 70 genes seleccionados. Las pacientes de a no mostraron evidencia visible de que el cáncer se diseminara a los ganglios linfáticos cercanos al momento de la intervención quirúrgica. Como se indica en la gráfica: (1) no todas estas personas sobrevivieron y (2) la probabilidad de supervivencia puede predecirse en gran medida por los perfiles de expresión génica de sus tumores. Esto permite a los médicos tratar a las pacientes con mal pronóstico en forma más agresiva en comparación con las mujeres con buen pronóstico. Las pacientes en b revelaron evidencia visible de diseminación de células cancerosas a los ganglios linfáticos cercanos. Como se indica en la gráfica, la probabilidad de sobrevivir en este grupo también puede predecirse con los datos de la expresión génica. En condiciones normales, todos los sujetos de este grupo recibirían tratamiento muy agresivo, que tal vez no sea necesario para los que tienen buen pronóstico. (Tomada de M. J. Van de Vijver et al., New Engl. J. Med. 347:2004, 2005, copyright 2002, the New England Journal Medicine by Massachusetts Medical Society, reimpresa con autorización de Massachusetts Medical Society, en el formato de reutilización de un libro/libro de texto (derechos básicos) vía copyright Clearance Center.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Figura 16.23 Efectos terapéuticos del Zelboraf (PLX4032) en pacientes con melanoma metastásico que presenta una oncoproteína BRAFV600E mutante. Estas PET de un paciente antes de (a) y dos semanas después de haber empezado el tratamiento (b) demuestran los efectos benéficos dramáticos de esta sustancia. Las regiones oscuras de la tomografía muestran la ubicación de las lesiones cancerosas con actividad metabólica. En muchos pacientes el tumor crece de nuevo por el surgimiento de clones farmacorresistentes. (Tomada de Gideon Bollag, et al., Nature 467:599, 2010; © 2010, reimpresa con autorización de Macmillan Publishers Ltd.)
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
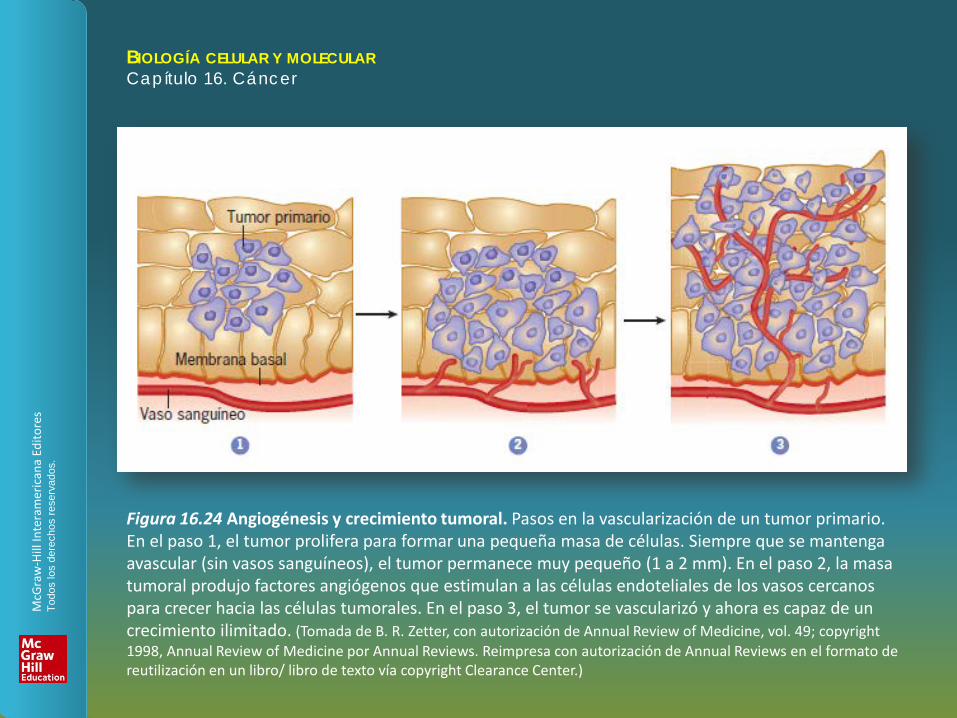
Figura 16.24 Angiogénesis y crecimiento tumoral. Pasos en la vascularización de un tumor primario. En el paso 1, el tumor prolifera para formar una pequeña masa de células. Siempre que se mantenga avascular (sin vasos sanguíneos), el tumor permanece muy pequeño (1 a 2 mm). En el paso 2, la masa tumoral produjo factores angiógenos que estimulan a las células endoteliales de los vasos cercanos para crecer hacia las células tumorales. En el paso 3, el tumor se vascularizó y ahora es capaz de un crecimiento ilimitado. (Tomada de B. R. Zetter, con autorización de Annual Review of Medicine, vol. 49; copyright 1998, Annual Review of Medicine por Annual Reviews. Reimpresa con autorización de Annual Reviews en el formato de reutilización en un libro/ libro de texto vía copyright Clearance Center.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Vías Experimentales Figura 1 Micrografía electrónica de un virus de la leucemia del ratón Friend que se desprende por gemación de la superficie de una célula leucémica cultivada. (Por cortesía de E. de Harven.)

McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
Vías Experimentales Figura 2 Incorporación de radiactividad de [3H]TTP en un precipitado insoluble en ácido por la DNA polimerasa del virus de la leucemia murina de Rauscher en presencia y ausencia de ribonucleasa. (Nota: el precursor TTP marcado se convierte en TMP cuando se incorpora en el DNA.) Curva 1, sin ribonucleasa agregada; curva 2, preincubada sin ribonucleasa agregada durante 20 min antes de añadir [3H]TTP; curva 3, ribonucleasa adicionada a la mezcla de reacción; curva 4, preincubada con ribonucleasa antes de añadir [3H]TTP. (Tomada de D. Baltimore Nature 226:1210, 1970, copyright 1970. Nature por Nature Publishing Group. Reimpreso con autorización de Nature Publishing Group en el formato de reutilización en un libro/libro de texto vía copyright Clearance Center.)
McG
raw
-Hill
Inte
ram
eric
ana
Edito
res
Todo
s lo
s de
rech
os re
serv
ados
. BIOLOGÍA CELULAR Y MOLECULAR Capítulo 16. Cáncer
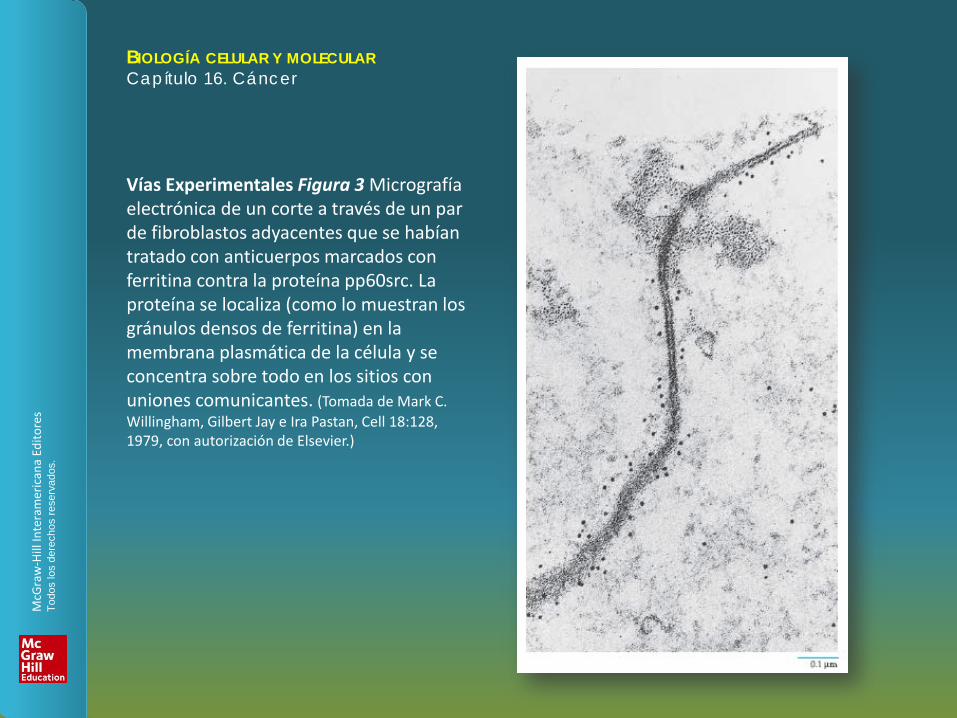
Vías Experimentales Figura 3 Micrografía electrónica de un corte a través de un par de fibroblastos adyacentes que se habían tratado con anticuerpos marcados con ferritina contra la proteína pp60src. La proteína se localiza (como lo muestran los gránulos densos de ferritina) en la membrana plasmática de la célula y se concentra sobre todo en los sitios con uniones comunicantes. (Tomada de Mark C. Willingham, Gilbert Jay e Ira Pastan, Cell 18:128, 1979, con autorización de Elsevier.)