bacillus thuringiensis informe de toxicología
DESCRIPTION
Bacillus Thuringiensis informe de toxicologíaTRANSCRIPT

UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOSUniversidad del Perú, Decana de América
FACULTAD DE QUÍMICA E INGENIERÍA QUÍMICAE.A.P. Ingeniería Agroindustrial
2° SEMINARIO - Bacillus thuringiensis
CURSO: TOXICOLOGÍA DE LOS ALIMENTOS
PROFESOR: OSCAR SANTISTEBAN ROJAS
TURNO: VIERNES 5-7 PM
ALUMNO: HUARCAYA YNDIGOYEN MARCO
CÓDIGO: 12070067
FECHA DE ENTREGA: 16/09/2015

ÍNDICE
Pág.
INTRODUCCIÓN 3
1 CARACTERÍSTICAS GENERALES 4
2 SITUACIÓN TAXONÓMICA Y CLASIFICACIÓN 5
3 TOXINAS 5
4 MODO DE ACCIÓN 7
4.1. UNIÓN A RECEPTOR DE LARVAS DE LEDIDÓPTERO 8
4.2. FORMACIÓN DEL PORO 8
4.3. INSERCIÓN EN MEMBRANA 9
5 IMPORTANCIA DE LOS RECEPTORES 11
5.1. RECEPTOR CADERINA EN LIDÓPTERO 11
5.2. RECEPTORES ANCLADOS A GLICOSILFOSFATIL-INOSITOL (GPI) 11
6. VENTAJAS Y LIMITACIONES DEL USO DE BACILLUS THURINGIENSIS 12
7. OTROS FACTORES DE VIRULENCIA PRODUCIDOS POR B. THURINGIENSIS 13
8. PLANTAS TRANSGÉNICAS Bt 14
9 BIBLIOGRAFÍA 15
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

INTRODUCCIÓNBacillus thuringiensis (Bt) fue descubierta a principios del siglo XX en Japón por el profesor Shigetane Ishiwata a partir de larvas muertas del gusano de la seda (Ishiwata 1902), pero fue descrita por Berliner en 1911 cuando lo aisló a partir de larvas de Anagasta kuehniella (Zeller) (Lepidoptera: Pyralidae) en la provincia alemana de Thuringia.
Bacillus thuringiensis es una bacteria del suelo, anaerobia facultativa, móvil y esporogénica, perteneciente al grupo de bacterias Gram positivas conformado por Bacillus cereus y Bacillus anthracis (Lecadet, 1970). Bacillus thuringiensis se caracteriza porque luego de la fase de crecimiento exponencial, las células producen tanto una espora subapical como uno o varios cuerpos parasporales: inclusiones compuestas de una o más proteínas cristalinas (ICPs) que tienen actividad insecticida específica, incluso a nivel de especie. Las toxinas de Bt comenzaron a utilizarse comercialmente en Francia en 1938, y para 1958 su uso se había extendido a los Estados Unidos. A partir de los años 80 Bt se convierte en un plaguicida de interés mundial. Los productos comercializados de B. thuringiensis consisten principalmente en preparados de esporas y cristales, activados o no, que se rocían sobre los cultivos como si se tratara de un insecticida convencional. A finales de la década de los 70 se identifica en la ribera de un río en Israel una subespecie tóxica para varias especies de dípteros. La nueva subespecie, que se denominó Bt israelensis (Bti), comenzó a usarse con éxito para el control de poblaciones de especies de moscas y mosquitos vectores, presentando unos valores de LC50 de alrededor de 12 ng/mL, dando origen a productos comerciales de Bti utilizados estrictamente para el control de dípteros.
Hasta ahora las investigaciones han demostrado que los productos de Bt no suponen un riesgo para artrópodos no diana, aves, peces ni mamíferos, observándose, en todos los casos, valores de toxicidad de muy baja a inexistente y siempre con dosis mayores a las de uso . De hecho, los únicos efectos dañinos observados son indirectos por disminución de la disponibilidad de alimento para los depredadores de los organismos diana de Bt. Aunado a esto, también se ha determinado que los productos de Bt tampoco tienen efectos negativos sobre el medio ambiente. En el caso específico de Bti, un estudio llevado a cabo en un río de Pensilvania (EEUU) durante 8 años demostró que la aplicación de Bti no tiene efecto en la composición y densidad de las poblaciones de peces y macroinvertebrados no diana, ni siquiera en ciertas especies de quironómidos que previamente se habían reportado como sensibles a Bt.
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

BACILLUS THURINGIENSIS
1. CARACTERÍSTICAS GENERALES :
Bacillus thuringiensis es un bacilo gram positivo, de flagelación perítrica, que mide de 3 a 5 µm de largo por 1 a 1,2 µm de ancho y que posee la característica de desarrollar esporas de resistencia elipsoidales que no provocan el hinchamiento del perfil bacilar (Figura 1) . Es un microorganismo anaerobio facultativo, quimioorganótrofo y con actividad de catalasa. Los distintos aislamientos de B. thuringiensis presentan en general características bioquímicas comunes. Poseen la capacidad de fermentar glucosa, fructosa, trealosa, maltosa y ribosa, y de hidrolizar gelatina, almidón, glucógeno, esculina y N-acetil-glucosamina. Sin embargo, la característica principal de B. thuringiensis es que durante el proceso de esporulación produce una inclusión parasporal formada por uno o más cuerpos cristalinos de naturaleza proteica que son tóxicos para distintos invertebrados, especialmente larvas de insectos (Figura 1). Estas proteínas se llaman Cry (del inglés, Crystal) y constituyen la base del insecticida biológico más difundido a nivel mundial.
1. Figura 1-Imagen de microscopía electrónica de transmisión de una cepa de B.thuringiensis en estado de esporangio. C: cristal parasporal; E: espora. Barra, 0,5 um.
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

2. SITUACIÓN TAXONÓMICA Y CLASIFICACIÓN :
B. thuringiensis pertenece a la familia Bacillaceae y se ubica dentro del grupo 1 del género Bacillus; forma parte del grupo de Bacillus cereus, el que incluye a B. anthracis, B. cereus, B. mycoides, así como también a los más recientemente descritos B. pseudomycoides y B. weihenstephanensis. Se encuentra estrechamente relacionado con los dos primeros, de los que no logra distinguirse por completo debido a que no existen suficientes diferencias en sus características morfológicas y bioquímicas, por lo que la designación de B. thuringiensis como especie ha estado sometida a discusión. A pesar de esto, el Bergey´s Manual of Determinative Bacteriology reconoce la individualidad de estas especies basándose sobre todo en dos características diferenciales: la presencia de la inclusión o cristal parasporal y sus propiedades insecticidas. B. thuringiensis se clasifica en 84 serovares mediante serología del antígeno flagelar H. Se trató de establecer relaciones entre estos serovares y su actividad insecticida o la presencia de algún tipo de cristal en particular pero no se tuvo éxito, por lo que su significado biológico no está claro. Por ejemplo, el serovar morrisoni agrupa cepas que son tóxicas para lepidópteros, dípteros y coleópteros; no todas las cepas de B. thuringiensis svar aizawai son tóxicas para Spodoptera littoralis (Lepidoptera). Existen, además, cepas de B. thuringiensis que no pueden clasificarse utilizando esta metodología, ya que no pueden formar flagelos. Todas estas cepas se agruparon en un serovar, actualmente desaparecido de la literatura.
3. TOXINAS :
Existen dos tipos de d-endotoxinas: las proteínas Cry y las proteínas Cyt. A la fecha se han clonado y secuenciado más de 200 diferentes genes cry y 16 diferentes genes cyt. Esto es sin duda un arsenal muy valioso para el control de diferentes insectos plaga y de insectos transmisores de enfermedades. La nomenclatura de las d-endotoxinas está basada exclusivamente en la similitud de la secuencia primaria. La definición de proteínas Cry es cualquier proteína paraesporal de Bt que muestre un efecto tóxico hacia algún organismo, verificable por medio de bioensayos o cualquier proteína que muestre similitud con las proteínas Cry. Actualmente se han encontrado toxinas Cry en otras especies de bacterias como Clostridium bifermentans (clasificadas como Cry16A y Cry17A) con actividad hacia mosquitos. Las proteínas Cyt denotan a las proteínas paraesporales de Bt que muestren actividad hemolítica o tengan similitud a la secuencia de las toxinas Cyt.
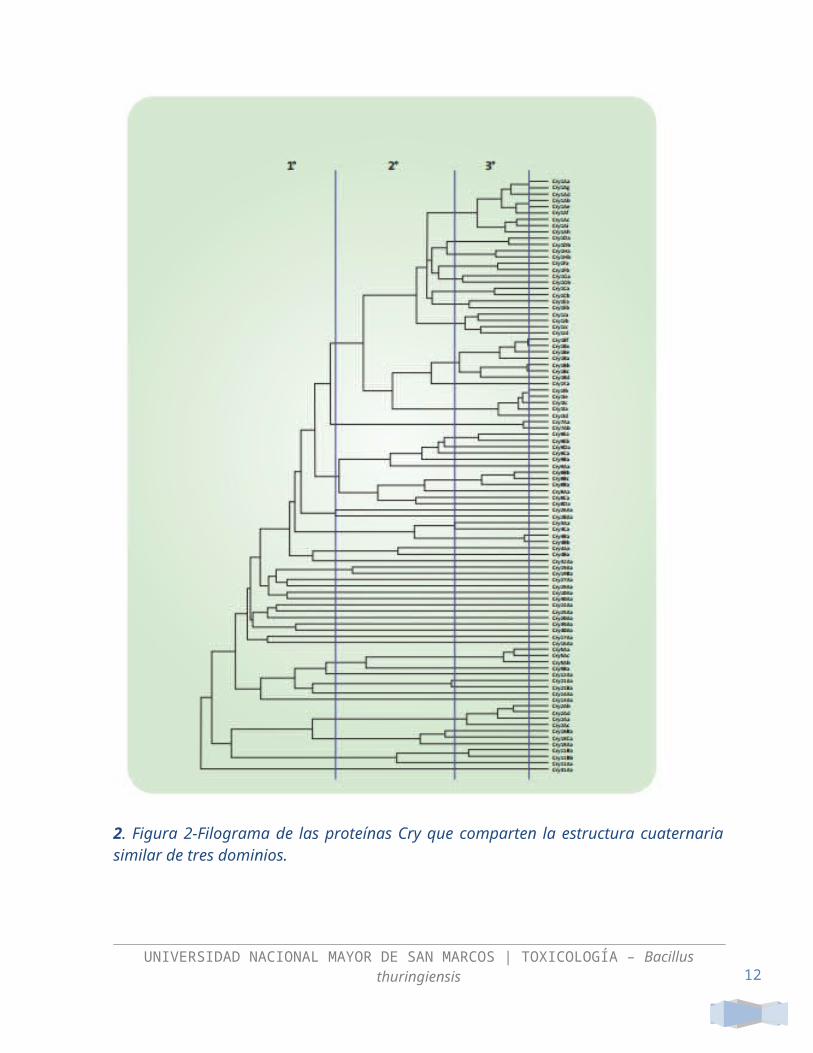
Las líneas verticales de la figura 2 representan los límites en identidad que marcan las diferentes categorías en la nomenclatura. El número arábigo se designa con la primera fila que corresponde hasta 45% de identidad (por ejemplo: Cry1, Cry2, etc.). La segunda hilera cataloga a las proteínas con una letra mayúscula y corresponde a identidades de 45 a 78% (Cry1A, Cry1B, etc.). La tercera fila asigna una letra minúscula y corresponde a identidades de 78 a
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis
95% (Cry1Aa, Cry1Ab, Cry1Ac, etc.). La última fila incluye un número arábigo al final de la nomenclatura indicando más de 95% de identidad (Cry1Aa1, Cry1Aa2, etc.). El grupo mayoritario de toxinas Cry se les conoce como la familia de tres-dominios, ya que están constituidas por tres dominios estructurales (figura 3).
2. Figura 2-Filograma de las proteínas Cry que comparten la estructura cuaternaria similar de tres dominios.
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

3.Figura 3-Estructura tridimensional de la proteína insecticida
4. MODO DE ACCIÓN
Las proteínas cristalinas se consideran de un gran interés teórico y práctico por la comunidad científica, por lo que es necesario conocer el efecto tóxico de las mismas, con el fin de poder realizar un correcto control de plagas y prevenir la aparición de insectos resistentes. El modo de acción de estas proteínas insecticidas ha sido principalmente caracterizado en lepidópteros. La principal acción de estas toxinas es lisar células epiteliales del intestino medio del insecto diana a partir de la formación de poros en la membrana apical de las microvellosidades de estas células. Por tanto, las proteínas Cry deben pasar de estar en forma de protoxina en el cristal de inclusión (que se disuelve por el pH alcalino del intestino medio), a toxina activada para unirse a receptores específicos de la misma y posteriormente insertarse en la membrana. Esto sucede una vez que la protoxina ha sido procesada por proteasas del intestino medio. De este modo se forman canales iónicos en las microvellosidades, la posterior formación del poro y finalmente se produce un desequilibrio osmótico y lisis celular. Durante la activación proteolítica, las protoxinas de alto peso molecular, entre 130- 140 kDa son ampliamente procesadas en su extremo carboxi-terminal, obteniéndose una proteína activa final de un peso molecular de entre 60-70 kDa. En cuanto a las proteínas de bajo peso molecular, de unos 70 kDa, el tamaño de la proteína cristalina activada es de un tamaño muy similar al de la protoxina.
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

4.1. UNIÓN A RECEPTOR EN LARVAS DE LEPIDÓPTERO.
Una vez que las toxinas están en su forma activada, éstas atraviesan la membrana peritrófica, que es una membrana que protege el epitelio pero permite el paso de moléculas de un tamaño menor o igual a 60 kDa hacia la membrana de las células epiteliales. El estudio de la unión de estas toxinas con las células del epitelio se ha realizado a partir del uso de técnicas histoquímicas, que utilizan anticuerpos frente a ellas o bien se las marca con biotina. A continuación, en preparaciones del intestino medio de larvas de insectos, incubadas in vitro con una proteína Cry, muestran la unión específica de éstas al borde en cepillo de las células epiteliales. Técnicas bioquímicas de ensayos de unión ligando-receptor, han permitido una mayor caracterización de la unión a la membrana celular. Cada una de las proteínas cristalinas se une a sitios de unión específicos en moléculas de membrana y, por este motivo, a estas moléculas se las denominó receptores.
Por ejemplo, para las toxinas Cry1A, se han encontrado diferentes receptores en distintas especies de lepidópteros, como son: proteínas de tipo cadherina (CADR), aminopeptidasa- N (APN), fosfatasa alcalina (ALP) y un glicoconjugado de 270 kDa. En esta especie se han encontrado dos de los cuatro tipos de receptores nombrados anteriormente: CADR y APN. Las caderinas son receptores transmembrana, con un dominio citoplásmico y un ectodominio extracelular con varias repeticiones de cadherina. El ectodominio tiene sitios de unión a calcio, secuencias de interacción a integrina y secuencias de unión a caderina. Por otro lado, se comprobó el grado de afinidad de la toxina Cry1A a ambos receptores y los resultados obtenidos fueron que, para la unión a caderina, la constante de disociación estaba en el rango de 1 nM, mientras que para el receptor APN era de 100 nM (Jenkins y Dean 2000).
4.2. FORMACIÓN DEL PORO.
En este paso de toxicidad, el modelo aceptado ha sido el de lisis coloideosmótica propuesto por Knowles y Ellar en 1987. Este modelo propone que las proteínas Cry producen pequeños poros en la membrana apical de las células columnares del intestino. Al formarse los poros en la membrana, se ve alterado el gradiente Na+ y K + hacia el exterior, permitiendo su libre movimiento. Además, las células, al ser de naturaleza hipertónica respecto a la luz intestinal, los poros formados permiten la entrada de agua al interior con el fin de producir un equilibrio osmótico con el exterior. Esto provoca un hinchamiento de la célula que desencadena la lisis celular (Worfersberger 1992). En el caso de la toxina Cry1Ab, la unión de ésta con su receptor en M. sexta, provoca un corte en el extremo N-terminal de la proteína, concretamente la α-hélice 1 del dominio I, lo que facilita la formación de la estructura oligomérica llamada pre-poro, importante para la inserción en membrana y para la toxicidad (Gómez et al. 2002b; Rausell et al. 2004a). Se han realizado numerosos trabajos para confirmar que la estructura oligomérica es importante en la formación del poro, como el uso de anticuerpos tipo scFv73, que imitan al receptor cadherina, o tratamientos con jugo de intestino medio de M. sexta
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis
(Gómez et al. 2002b, 2003). Además, las estructuras oligoméricas de Cry1Ab y Cry1Ac aumentan de 100 a 200 veces su afinidad de unión al receptor secundario APN, mostrando una constante de disociación de entre 0,75 - 1nM (Gómez et al. 2003; Pardo- López et al. 2006). Se ha demostrado la formación de estructuras oligoméricas en las toxinas: Cry1Aa, Cry1Ab, Cry1Ca, Cry1Da, Cry1Ea, Cry1Fa y Cry3 (Gómez et al. 2002b; Rausell et al. 2004a, 2004b; Muñoz-Garay et al. 2006). En todos estos casos, las toxinas que contienen estas estructuras muestran una alta correlación con la actividad formadora de poros, frente a estructuras monoméricas, que presentan niveles marginales de actividad de formación de poro. Todos estos trabajos apoyan la hipótesis de que la formación de un oligómero es un paso necesario para la actuación de estas proteínas insecticidas.
4.3. INSERCIÓN EN MEMBRANA.
El modelo de inserción en membrana no está del todo claro. Li et al. en 1991 propuso que las α-hélices 4 y 5 penetran en la membrana lipídica de las células epiteliales, mientras que el resto de las α-hélices permanecen en la superficie. A este modelo se le denominó “Modelo Paraguas”. Por otro lado, Hodgman y Ellar en 1990, formulan otro modelo, el “Modelo Navaja”, donde las α-hélices 5 y 6 sobresalen del dominio I y son las que penetran la membrana. Finalmente, Dean et al. en 1996, propuso que el dominio I se insertaba completamente. Se ha observado que el dominio I de estas proteínas Cry, es similar a otro de otras proteínas formadoras de poro, colicina A, toxina diftérica, o exotoxina de Pseudomonas, de hecho, los dos primeros modelos se basan en estas toxinas formadoras de poro. Todas estas toxinas bacterianas tienen en común el tipo de poro que forman, poro tipo α, que forma 3 capas de α-hélices; las α-hélices formadoras de un poro capilar hidrofóbico de forma helicoidal estarían protegidas en el centro y el poro se alinearía atravesando la membrana.
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

4.Figura 4-Modo de acción de toxinas Cry de Bacillus thuringiensis.
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

5. IMPORTANCIA DE LOS RECEPTORES EN EL MODO DE ACCIÓN DE LAS TOXINAS CRY :
Una de las mayores ventajas del uso de las toxinas Cry para el control de plagas es su alta especificidad en su acción insecticida que se limita a nivel de orden como categoría taxonómica. Esta cualidad está directamente relacionada con los receptores específicos que requiere la toxina para actuar, siendo solo algunos insectos quienes los expresan, lo cual también hace imposible que pueda afectar al ser humano si las consume, pues no expresa los receptores necesarios para que se lleve a cabo su toxicidad. El estudio bioquímico de las proteínas Cry se ha enfocado principalmente en la identificación de los receptores específicos que interaccionan con ellas una vez que fueron ingeridas por un insecto blanco. Se han identificado dos tipos principales de receptores involucrados en el modo de acción insecticida de las proteínas Cry en lepidópteros: el receptor transmembranal de caderina y los receptores acoplados a membrana por medio de un grupo glicosilfosfatidil-inositol (GPI) como la Aminopeptidasa N (APN) y la Alcalino Fosfatasa (ALP).
5.1. RECEPTOR CADERINA EN LEPIDOPTERO
La superfamilia de proteínas Caderina es diversa en sus funciones como en su expresión en diferentes organismos. Éstas, tienen funciones relacionadas en la adhesión celular, en la organización citoesquelética, respuesta inmune y en procesos como la morfogénesis (Angst et al., 2001). Las caderinas son glicoproteínas que se caracterizan estructuralmente por su composición en “microdominios” de aproximadamente 110 aminoácidos con un enlace a calcio, también llamados repeticiones de caderina (RC). En insectos esta proteína modular presenta tres dominios: un ectodominio extracelular formado de 11-12 RC, uno transmembranal y por último el intracelular (Vadlamudi et al., 1995). A diferencia de la caderina presente en vertebrados donde se encuentra localizada en el área baso-lateral de las membranas participando en el ensamblaje intercelular (Angst et al., 2001), en insectos está situada en las microvellosidades de las células epiteliales del intestino. Esto sucede en lepidópteros como M. sexta, H. virescens y B. mori (Aimanova et al., 2006; Chen et al., 2005). Aunque su nivel de expresión aumenta progresivamente con el desarrollo larval, su función intrínseca en el insecto no está identificada, sin embargo se propone que tiene un papel regulador en la estabilidad del tejido (Midboe et al., 2003) pero esto no explica su ausencia en el individuo adulto.
5.2. RECEPTORES ANCLADOS A GLICOSILFOSFATIDIL-INOSITOL (GPI)
La familia de enzimas aminopeptidasas N se encargan de cortar polipéptidos en zonas de aminoácidos neutros en su extremo terminal N. Sus funciones varían al encontrarse en una gran variedad de especies. En el caso específico del epitelio intestinal de lepidópteros, sus funciones
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

se enfocan a la digestión de proteínas derivadas de la dieta del insecto, papel que comparte con algunas endopeptidasas y carboxypeptidasas. La aminopeptidasa N (APN) fue de los primeros que se asociaron con las toxinas Cry (Knight et al., 1994). En lepidópteros, este receptor se encuentra anclado a la membrana a través de un grupo glicosilfosfatidil-inositol (GPI), a diferencia de cómo se encuentra en vertebrados, donde parte de su estructura en su extremo terminal N es hidrófóbica y funciona como pie de anclaje a la membrana celular (Piggot y Ellar 2007). La glicosilación es importante en la interacción de APN-Cry. Esto fue observado en la interacción de Cry1Ac con el receptor APN de M. sexta, donde el dominio III reconocía zonas específicas de unión de carbohidratos en aminopeptidasas N (Burton et al., 1999; revisado por Piggot y Ellar 2007)
6. VENTAJAS Y LIMITACIONES DEL USO DE BACILLUS THURINGIENSIS
Varios factores han hecho posible su éxito en la agricultura y en el control de mosquitos transmisores de enfermedades. El más importante es su alta especificidad hacia el insecto blanco y su inocuidad para mamíferos, otros vertebrados, plantas e inclusive otros insectos benéficos. Las toxinas de Bt se han utilizado como bioinsecticidas en agricultura durante los últimos 40 años, principalmente en cultivos de hortalizas y cereales. Como se mencionó antes, también Bti se ha ocupado con éxito durante 30 años para el control de mosquitos. Sólo existe un ejemplo de generación de resistencia a Bt en campo. Esto se debe a que los tiempos de permanencia de las proteínas Cry en el ambiente son muy cortos, por lo que la presión de selección es muy baja. También existen evidencias de mutaciones que afectan la expresión de los receptores tipo caderina, que resultan en insectos resistentes a las toxinas Cry1A, y tienen un costo en el desempeño de los insectos en la naturaleza, evitando que las mutaciones que afectan a esta proteína se fijen en la población de insectos. Paradójicamente, las ventajas de Bt se convierten en importantes desventajas para su uso comercial. El estrecho rango de huésped ocasiona que no se cuente con toxinas para cada plaga que afecta la actividad humana. También, la reducida permanencia en el ambiente hace necesario un profundo conocimiento de la biología y comportamiento de la plaga que se quiere controlar, ya que una toxina puede ser activa para los estadios larvarios, pero disminuir o incluso no ser tóxica para los adultos. Por lo tanto, los tiempos y formas de aplicación deben seleccionarse cuidadosamente.
Otra limitante ha sido la utilización de Bt para el control de insectos barrenadores y chupadores, ya que su aplicación se ha dado tradicionalmente como producto asperjado, y el hábito alimenticio de estos insectos impiden la ingestión de la toxina Cry. Este problema se ha resuelto con la creación de plantas transgénicas que producen sistémicamente la toxina Cry haciéndola accesible a insectos barrenadores. Por último, existe el riesgo de desarrollo de resistencias por el incremento en el uso de Bt como aspersiones de cristales y sobre todo en plantas transgénicas que expresen constitutivamente una o varias toxinas Cry. El objetivo es que la planta, una vez transformada con el gen de la toxina, exprese suficiente cantidad de ésta
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

como para aniquilar a las plagas susceptibles que las consumen. Desde 1987 aparecieron los primeros reportes sobre plantas de tabaco y tomate que presentaban suficiente expresión de la toxina de Bt como para conferir niveles altos de resistencia. En la actualidad, universidades, centros de investigación y compañías privadas llevan a cabo proyectos sobre el desarrollo de plantas transgénicas con capacidad insecticida, una gran variedad de importancia económica. La adopción de plantas transgénicas en la agricultura está ocurriendo a velocidad vertiginosa. Sólo en Estados Unidos, el 50% de la superficie sembrada con soya consiste de plantas transgénicas resistentes a herbicidas. En México, por ejemplo, el 60% del algodón que se cultiva es Bt. En la actualidad existen ocho cultivos importantes con cultivares transgénicos registrados: soya, maíz, algodón, canola, papa, tomate, tabaco y remolacha, y muchas otras plantas están próximas a registrarse.
7. OTROS FACTORES DE VIRULENCIA PRODUCIDOS POR B. THURINGIENSIS
Además de las proteínas Cry, B. thuringiensis posee otros factores de virulencia que le permitirían sobrevivir y multiplicarse dentro del huésped, evadir su sistema inmune y producir septicemia. Algunos de estos factores de virulencia, como fosfolipasas, enterotoxinas, β-exotoxinas, y quitinasas, no tienen especificidad por el orden de insectos que afectan y se encuentran ampliamente distribuidos dentro de los distintos serovares de B. thuringiensis y otros miembros del grupo B. cereus. Por otro lado, algunos B. thuringiensis poseen otros factores de virulencia que sí tienen especificidad por el orden de insectos que afectan. Tal es el caso de las proteínas Cyt que se encuentran formando parte principalmente de los cristales de cepas mosquitocidas de B. thuringiensis (ej. svar. israelensis, kyushuensis, darmstadiensis, medellin, etc.), aunque también se pueden llegar a encontrar rara vez en alguna cepa tóxica para lepidópteros (ej. svar. morrisoni HD-12). Estas proteínas son hemolíticas y citolíticas in vitro y son especialmente tóxicas para larvas de dípteros in vivo. Otro caso de factores de virulencia que tienen especificidad por el orden de insectos que afecta lo constituye el grupo de proteínas llamado Vip (vegetative insecticidal proteins). Las Vip son proteínas insecticidas no relacionadas con las Cry que no forman inclusiones cristalinas y que se secretan al medio durante el crecimiento vegetativo de la bacteriA. Incluyen la toxina binaria Vip1 y Vip2 con especificidad para coleópteros y la toxina Vip3 con especificidad para lepidópteros. Existen en la actualidad 40 genes vip3 distintos clasificados en 2 clases (A y B), que codifican para proteínas de aproximadamente 88 kDa. Si bien las Vip3 compartirían algunas etapas en el modo de acción con las Cry, utilizarían distintos receptores. Otras proteínas llamadas Sip (secreted insecticidal proteins) fueron recientemente descritas, pero muy poco se conoce de ellas. Sólo se conoce un miembro de este grupo: la proteína Sip1A, que también es producida y secretada por B. thuringiensis durante su etapa de crecimiento vegetativo y que posee propiedades insecticidas para coleópteros.
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

8. PLANTAS TRANSGÉNICAS Bt
La ingeniería genética desarrolló muchas especies de plantas que expresan genes cry de B. thuringiensis y las convirtió así en “plantas insecticidas”. Comúnmente se hace referencia a este tipo de plantas como “plantas o cultivos Bt” (por ejemplo, maíz Bt, algodón Bt, etc.). El primer informe de una planta transgénica con un gen cry de B. thuringiensis data de 1987. Se desarrollaron plantas de tabaco (Nicotiana tabacum) que producían cantidades suficientes de proteína Cry para controlar larvas de primer estadio de Manduca sexta. Desde entonces, al menos diez tipos de genes cry distintos se introdujeron en 26 especies vegetales: cry1Aa, cry1Ab, cry1Ac, cry1Ba, cry1Ca, cry1H, cry2Aa, cry3A, cry6A y cry9C. El empleo de este tipo de plantas posee la ventaja de reducir la necesidad de aplicar insecticidas y de proveer una protección duradera a lo largo de la temporada de cultivo. Otra característica muy importante que poseen es que los únicos insectos expuestos a la toxina son aquellos que se encuentran alimentándose de los cultivos y no otros. No obstante, la mayor ventaja de estas plantas Bt es que brindan a los agricultores una alternativa al uso de pesticidas químicos tradicionales y constituyen una herramienta para el control de plagas de importancia económica que son difíciles de controlar por los primeros. Todas estas ventajas se verían reflejadas finalmente en beneficios enormes para la producción de alimentos y en una calidad medioambiental mejor en todo el mundo.
Por otro lado, las plantas Bt poseen la gran desventaja que radica en la posible generación de resistencia en ciertas poblaciones naturales de insectos que se alimentan de ellas. Este fenómeno determinaría la inutilización de determinadas proteínas Cry para su control, ya sea mediante su empleo en plantas transgénicas o con los bioinsecticidas que los contengan (47).
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis

9. Referencias biliográficas.
DIEGO H. SAUKA, GRACIELA B. BENINTENDE. (2008). Bacillus thuringiensis: generalidades. Un acercamiento a su empleo en el biocontrol de insectos lepidópteros que son plagas agrícolas.
LUCÍA IZQUIERDO RUBIO. (2011). Actividad de las toxinas de Bacillus thuringiensis para el control de Tuta absoluta.
JOSÉ ALBERTO VALADÉZ LIRA. (2011). Modulación de la Inmunidad innata del Lepidóptero plaga Trichoplusia ni (Hübner) expuesto a Bacillus thuringiensis.
GERARDO DEL TORO DE LEÓN. (2010). Caracterización del espectro de acción de la toxina Cry1AbMod, activa contra insectos resistentes, y su comparación con la toxina convencional Cry1Ab de Bacillus thuringiensis.
Mancebo A, González Navarro B, Riera L, Lugo S, González Torres Y, Arteaga ME y Fuentes D. (2003) .Evaluación de la toxicidad/patogenicidad de una formulación de Bacillus thuringiensis var israelensis (Bactivec).
Mario Soberón y Alejandra Bravo. (2007). Las toxinas Cry de Bacillus thuringiensis: modo de acción y consecuencias de su aplicación.
Gustavo Ochoa & Jazzmin Arrivillaga. (2009). Bacillus thuringiensis: Avances y perspectivas en el control biológico de Aedes aegypti
UNIVERSIDAD NACIONAL MAYOR DE SAN MARCOS | TOXICOLOGÍA – Bacillus thuringiensis