activation of phospholipase c by g-protein βγ subunits in ddt1mf-2 cells
TRANSCRIPT

ejp European Journal of Pharmacology molecular pharmacology
Molecular Pharmacology Section 288 (1995) 393-398 ELSEVIER
Short communication
Activation of phospholipase C by G-protein/3y subunits in DDT1MF-2 cells
John M. Dickenson a,., Montserrat Camps b Peter Gierschik c, Stephen J. Hill a
a Department of Physiology and Pharmacology, Medical School, Queen's Medical Centre, Nottingham NG7 2UH, UK b Signal Transduction Section, Glaxo Institute for Molecular Biology, Geneva, Switzerland
c Department of Pharmacology and Toxicology, University of Ulm, Albert-Einstein-Allee 11, NZ6-429, 89069 Ulm, Germany
Received 19 October 1994; revised 5 December 1994; accepted 9 December 1994
Abstract
Adenosine A 1 receptors directly stimulate inositol phospholipid hydrolysis and Ca 2+ mobilization through a pertussis toxin sensitive mechanism in DDTIMF-2 cells. In the present study we have investigated whether G protein fly subunits (G/3y) are capable of stimulating phospholipase C in DDTIMF-2 cell membrane preparations using lipid vesicles containing [3H]phospha- tidylinositol 4,5-bisphosphate. DDTIMF-2 cell membrane and soluble fractions were found to contain phospholipase C activity which was stimulated by increases in free Ca 2+ ion concentration. G/3y purified from bovine retinal transducin produced significant increases in phospholipase C activity in DDT1MF-2 cell membranes. G/3y-dependent activation of phospholipase C, while virtually absent in the presence of low Ca 2+ ion concentrations, increased markedly with increasing free Ca 2+ ion concentration. These data suggest that membrane bound phospholipase C in DDT1MF-2 cells is sensitive to Ca 2+, and may be stimulated conditionally by G[3y subunits, i.e. G[3y subunits activate the enzyme only in the presence of Ca 2+. G/3y subunits also stimulated soluble phospholipase C in DDTIMF-2 cells. These findings support the hypothesis that G i /~y subunits are involved in adenosine A~ receptor stimulated phospholipase C/Ca 2+ signalling in DDT1MF-2 cells.
Keywords: DDTIMF-2 cell; Smooth muscle cell; G/3y subunit; Phospholipase C
1. Introduction
DDT1MF-2 cells express adenosine A~ receptors which are negatively coupled to adenylate cyclase through a pertussis toxin sensitive G i protein (Ramkumar et al., 1990). Our previous studies have shown that activating the adenosine A 1 receptor also stimulates the hydrolysis of inositol phospholipids and mobilization of intracellular Ca 2÷ through a pertussis toxin sensitive mechanism (White et al., 1992; Dicken- son and Hill, 1993). Recent findings suggest that the activation of phospholipase C through receptors cou- pled to pertussis toxin sensitive G i proteins may be mediated by G i protein /3y subunits (Camps et al., 1992; Clapham and Neer, 1993). In DDT1MF-2 cells o/i and fly subunits released from the G i coupled adeno- sine A 1 receptor may independently inhibit adenylate
* Corresponding author. Tel.: 0602 709468.
0922-4106/95/$09.50 © 1995 Elsevier Science B.V. All rights reserved SSDI 0922-4106(94)00201-0
cyclase and stimulate phospholipase C respectively (Gerwins and Fredholm, 1992a,b; Dickenson and Hill, 1994).
In the present study we have investigated whether G protein /3y subunits are capable of stimulating phos- pholipase C in DDTIMF-2 ceils.
2. Materials and methods
2.1. Cell culture
The hamster vas deferens smooth muscle cell line (DDT~MF-2) was obtained from the European Collec- tion of Animal Cell Cultures (Porton Down, Salisbury, UK). DDT~MF-2 cells were cultured at 37°C in a humidified a i r / C O 2 (90:10) atmosphere in 75 cm 2 flasks (Costar). The growth medium was Dulbecco's modified Eagle's medium supplemented with 2 mM L-glutamine and 10% (v/v) foetal calf serum. Cells

394 J.M. Dickenson et al. / European Journal of Pharmacology - Molecular Pharmacology Section 288 (1995) 393-398
were passaged twice a week (1/8 split ratio) by vigor- ous shaking of the flask and placed into 75 cm 2 flasks and fed with fresh growth medium every 48 h. Ceils for membrane preparation were grown in 175 cm 2 flasks.
2. 2. Membrane preparation
Cells from sixteen confluent 175 cm 2 flasks (suffi- cient to produce approximately 600 /~g of membrane protein) were detached mechanically (using a cell scraper) into a Ca2+/Mg 2+ free phosphate buffered saline solution (80 mM NazHPO4, 20 mM NaH2PO 4 100 mM NaC1, pH 7.5). After centrifugation (150 g for 5 min) cells were resuspended in 10 ml of ice-cold hypotonic Tris buffer (20 mM Tris-HCl, 2 mM EGTA, pH 7.4) which contained the following protease in- hibitors 2 tzg/ml soybean trypsin inhibitor, 3 mM benzamidine, 1 p~M pepstatin, 1 /zM leupeptin, 100 /zM phenylmethylsulphonyl fluoride and 100 nM tosyl lysine chloromethyl ketone. Cells were then ho- mogenised using a glass teflon homogeniser (approxi- mately 20 strokes) and centrifuged at 500 ×g for 10 min to remove unbroken cells and nuclei. The super- natant was removed and centrifuged at 36000 × g for 30 min. The resulting pellet was resuspended in hypo- tonic buffer and centrifuged at 36,000 g for 30 min, whereas the supernatant was used as the soluble (cyto- solic) fraction without further centrifugation and stored at - 80°C. Following centrifugation, the membrane pel- let was resuspended in hypotonic buffer to a final protein concentration of 4-6 mg/ml and stored at - 8 0 ° C until required.
2.3. Phospholipase C assay
Phospholipase C activity in membrane and soluble fractions from DDT~MF-2 cells was assayed using ex- ogenous radiolabelled substrate. Mixed phospholipid vesicles containing phosphatidylethanolamine (0.7 /.d/assay tube from 10 mg/ml stock solution in CHC13), PtdIns(4,5)P 2 (1.1 pA/assay tube from 1 mg/ml stock solution in CHC13) and [3H]PtdIns(4,5)P2 (201.7 GBq/mmol; 0.37 kBq/assay tube) were prepared by drying the lipids under a stream of N 2 and resuspend- ing them in an appropriate volume (20/zl/assay tube) of reaction mixture buffer [87.5 mM Tris-maleate, pH 7.0, 17.5 mM LiCI, 17.5 mM 2,3-bisphosphoglycerate, 5.25 mM EGTA, 8.75 mM MgCl 2 and 0.087% (mass/vol) sodium deoxycholate] followed by sonica- tion for 30 min. Assays were performed for 30 min at 25°C in a total volume of 35/zl which consisted of 5 ~1 membrane or soluble protein (usually 10 p.g of protein in 20 mM Tris-HCl buffer and protease inhibitors); 20 pA lipid substrate mixture; 5 p.1 /33' subunits [in 10 mM Tris-HC1, pH 7.5, 6 mM MgCl 2, 1 mM dithiothreitol, 20% (by vol) glycerol, 100 ~M phenylmethylsulphonyl
fluoride] and 5 /zl of CaC12 to give the indicated free Ca 2+ concentrations (calculated using the computer programme EqCal). The reaction was stopped by adding 175 /~1 of CHCIJCH3OH / concentrated HC1 (500:500:3, by vol), vortexing and adding 50/zl of 1 M HCI containing 5 mM EGTA. Phase separation was accelerated by centrifugation for 1 min in an Eppen- doff microcentrifuge. A 100 /~1 aliquot of the upper aqueous phase (containing tritiated products) was re- moved and radioactivity quantified by counting in the gel phase (scintillator 299, Packard).
2.4. Western blot analysis
Protein samples were separated by SDS/PAGE [4- 15% (w/v) gradient acrylamide gel] in 20 cm × 20 cm slab gels run overnight at 5 mA. Proteins were trans- ferred to nitrocellulose membranes using a Bio-Rad Trans-Blot system (5 h at 190 mA in 25 mM Tris, 192 mM glycine buffer without methanol). After transfer, the nitrocellulose membrane was stained with Ponceau S to visualise samples and molecular weight markers. The membrane was then washed with phosphate buffered saline (PBS) (80 mM NazHPO4, 20 mM NaHzPO 4, 100 mM NaC1, pH 7.5) and blocked for 2 h at room temperature with 5% (mass/vol) skimmed milk powder in PBS. Blots were then incubated overnight at 4°C with primary antibodies in 5% (mass/vol) skimmed milk powder dissolved in PBS containing 0.5% (by vol) Tween 20. The primary anti- body was removed and the blot extensively washed with PBS containing 0.5% (by vol) Tween 20. Blots were then incubated for 1 h at room temperature with the second antibody (swine anti-rabbit IgG coupled to horseradish peroxidase) at 1:1250 dilution in 5% (mass/vol) skimmed milk powder dissolved in PBS containing 0.5% (by vol) Tween 20. After removing the second antibody the blot was then extensively washed as above and developed using the Enhanced Chemilu- minescence detection system (Amersham).
2.5. Purification of /3y subunits from bouine retinal transducin
/32' subunits from bovine rod outer segments were purified as described previously (Camps et al., 1992). Briefly, heterotrimeric transducin was eluted from rod outer segment membranes with hypotonic buffer con- taining 100 /zM GTP and separated into a and /3y subunits by chromatography on Blue Sepharose CL 6B (Pharmacia-LKB).
2.6. Miscellaneous
Concentration-response curves were fitted to a lo- gistic equation using the non-linear regression pro-
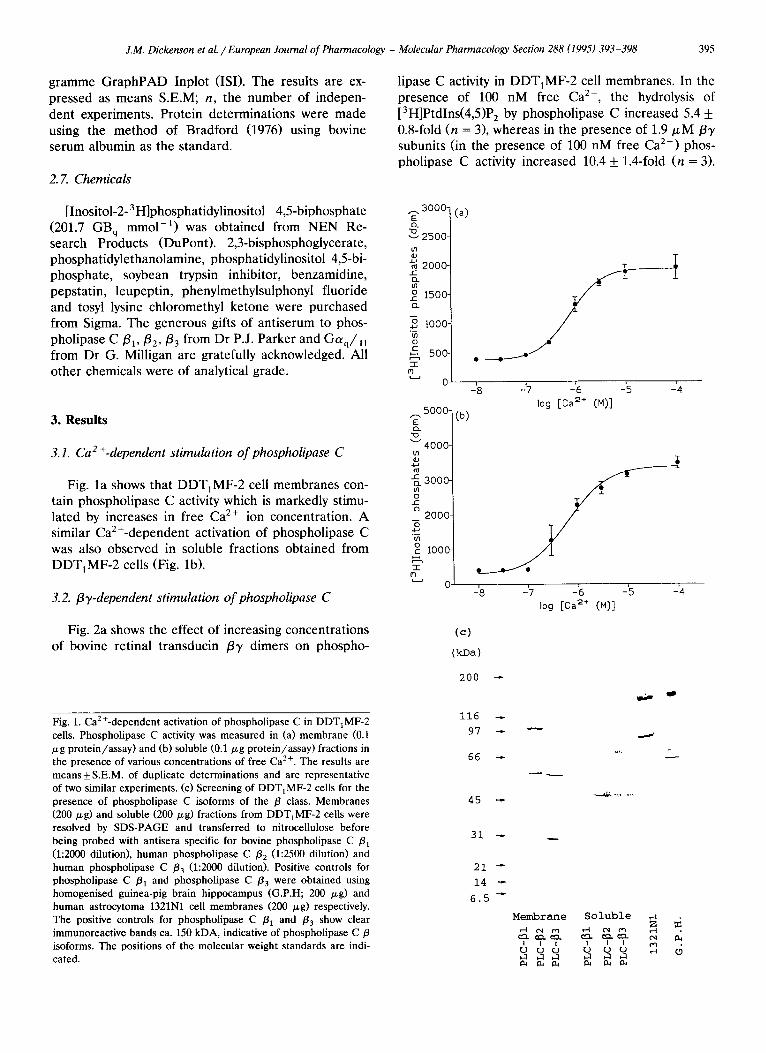
J.M. Dickenson et al. / European Journal of Pharmacology - Molecular Pharmacology Section 288 (1995) 393-398 395
gramme GraphPAD Inplot (ISI). The results are ex- pressed as means S.E.M; n, the number of indepen- dent experiments. Protein determinations were made using the method of Bradford (1976) using bovine serum albumin as the standard.
2. 7. Chemicals
lipase C activity in DDTtMF-2 cell membranes. In the presence of 100 nM free Ca 2+, the hydrolysis of [3H]Ptdlns(4,5)P2 by phospholipase C increased 5.4 + 0.8-fold (n = 3), whereas in the presence of 1.9/~M f y subunits (in the presence of 100 nM free Ca e+) phos- pholipase C activity increased 10.4 + 1.4-fold (n = 3).
[Inositol-2- 3 H]phosphatidylinositol 4,5-biphosphate (201.7 GBq mmol -x) was obtained from NEN Re- search Products (DuPont). 2,3-bisphosphoglycerate, phosphatidylethanolamine, phosphatidylinositol 4,5-bi- phosphate, soybean trypsin inhibitor, benzamidine, pepstatin, leupeptin, phenylmethylsulphonyl fluoride and tosyl lysine chloromethyl ketone were purchased from Sigma. The generous gifts of antiserum to phos- pholipase C f l , f2, f3 from Dr P.J. Parker and Gotq/ll from Dr G. Milligan are gratefully acknowledged. All other chemicals were of analytical grade.
3. Results
3.1. Ca 2 +-dependent stimulation of phospholipase C
Fig. la shows that DDT1MF-2 cell membranes con- tain phospholipase C activity which is markedly stimu- lated by increases in free Ca 2+ ion concentration. A similar Ca2+-dependent activation of phospholipase C was also observed in soluble fractions obtained from DDTIMF-2 cells (Fig. lb).
3.2. fly-dependent stimulation of phospholipase C
Fig. 2a shows the effect of increasing concentrations of bovine retinal transducin f y dimers on phospho-
r . 3OOO- E
~'~'2500-
o
2000-
1500-
i000-
o C 500-
0
5000- E
4000'
~_3000- o
2000- 3 m
1000-
I
~'~ 0
(a)
-'8
(b)
-7 -'6 -'5 -'4 log [ca 2+ (M)]
-18 --I7 -- '6 -- '5 -- '4 log [Ca 2+ (M)]
(e)
(kDa)
Fig. 1. Ca2+-dependent activation of phospholipase C in DDT1MF-2 cells. Phospholipase C activity was measured in (a) membrane (0.1 /xg protein/assay) and (b) soluble (0.1/zg protein/assay) fractions in the presence of various concentrations of free Ca 2+. The results are means_+S.E.M, of duplicate determinations and are representative of two similar experiments. (c) Screening of DDT1MF-2 cells for the presence of phospholipase C isoforms of the fl class. Membranes (200/zg) and soluble (200/zg) fractions from DDT1MF-2 cells were resolved by SDS-PAGE and transferred to nitrocellulose before being probed with antisera specific for bovine phospholipase C /31 (1:2000 dilution), human phospholipase C /32 (1:2500 dilution) and human phospholipase C /33 (1 :2000 dilution). Positive controls for phospholipase C /31 and phospholipase C /33 were obtained using homogenised guinea-pig brain hippocampus (G.P.H; 200 p,g) and human astrocytoma 1321N1 cell membranes (200 /zg) respectively. The positive controls for phospholipase C /31 and /33 show clear immunoreactive bands ca. 150 kDA, indicative of phospholipase C/3 isoforms. The positions of the molecular weight standards are indi- cated.
200
116
97
66
45
31
21
14
6.5
Membrane Soluble z =
I I t I I I
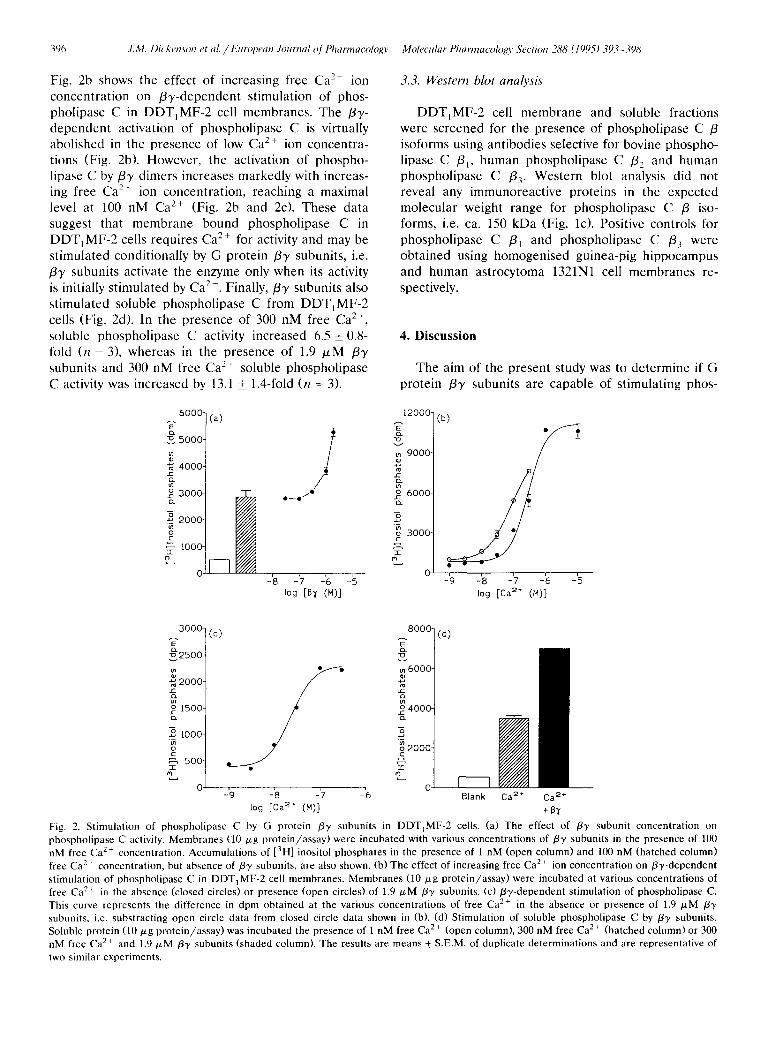
396 ,I.M. Dicken.son et al. / European Journal Of Pharmacology Molecular Pharmacolo~o, Section 288 (1995) 393-398
Fig. 2b shows the effect of increasing free Ca 2+ ion concentration on /3y-dependent stimulation of phos- pholipase C in DDT1MF-2 cell membranes. The /33'- dependent activation of phospholipase C is virtually abolished in the presence of low Ca 2+ ion concentra- tions (Fig. 2b). However, the activation of phospho- lipase C by/33' dimers increases markedly with increas- ing free Ca 2+ ion concentration, reaching a maximal level at 100 nM Ca 2+ (Fig. 2b and 2c). These data suggest that membrane bound phospholipase C in DDT, MF-2 cells requires Ca 2+ for activity and may be stimulated conditionally by G protein /33' subunits, i.e. /33' subunits activate the enzyme only when its activity is initially stimulated by Ca 2+. Finally,/3Y subunits also stimulated soluble phospholipase C from DDT,MF-2 cells (Fig. 2d). In the presence of 300 nM free Ca 2+, soluble phospholipase C activity increased 6.5 _+ 0.8- fold (n = 3), whereas in the presence of 1.9 # M /33' subunits and 300 nM free Ca 2+ soluble phospholipase C activity was increased by 13.1 _+ 1.4-fold (n = 3).
3.3. Western blot analysis
DDT,MF-2 cell membrane and soluble fractions were screened for the presence of phospholipase C /3 isoforms using antibodies selective for bovine phospho- lipase C /3,, human phospholipase C /32 and human phospholipase C /33- Western blot analysis did not reveal any immunoreactive proteins in the expected molecular weight range for phospholipase C /3 iso- forms, i.e. ca. 150 kDa (Fig. lc). Positive controls for phospholipase C /31 and phospholipase C /33 were obtained using homogenised guinea-pig hippocampus and human astrocytoma 1321N1 cell membranes re- spectively.
4. Discussion
The aim of the present study was to determine if G protein /33, subunits are capable of stimulating phos-
6000-
~ 5 0 0 0 -
4000-
3000-
.~ 2000-
~ , tO00-
0
(a)
F--I
/ 0 _ _ 0 . , / @
-'8 -'7 -'6 -'5 log [13y (M)]
12000-
m 9000"
C m ~ 6000-
m o 3000- C
I m
o
(b)
-'9 -'8 -'7 -'6 -'5 log [Ca =+ (M)]
3000-
~ 2 5 0 0 -
-~ 2ooo- C in
o 1500" .Z2
• ~ IO00-
w--, 500- I
M
(c) .~.8°°° 7 (d)
-'9 -'8 -'7 -'6 Blank C a 2 ÷ C a 2 ÷ - -
log rca 2÷ (M)] +By
Fig. 2. Stimulation of phospholipase C by G protein /37 subunits in DDTIMF-2 cells. (a) The effect of /33' subunit concentration on phospholipase C activity. Membranes (10 p,g p ro t e in / a s say )were incubated with various concentrations of/33' subunits in the presence of 100 nM free Ca 2+ concentration. Accumulat ions of [3H] inositol phosphates in the presence of 1 nM (open column) and 100 nM (hatched column) free Ca 2+ concentration, but absence of/33' subunits, are also shown. (b) The effect of increasing free Ca 2+ ion concentration on /3y-dependent stimulation of phospholipase C in DDT1MF-2 cell membranes . Membranes (10 p,g prote in /assay) were incubated at various concentrations of free Ca ~+ in the absence (closed circles) or presence (open circles) of 1.9 # M /3-), subunits. (c) /3",/-dependent stimulation of phospholipase C. This curve represents the difference in dpm obtained at the various concentrations of free Ca 2+ in the absence or presence of 1.9 /xM /37 subunits, i.e. substracting open circle data from closed circle data shown in (b). (d) Stimulation of soluble phospholipase C by /33, subunits. Soluble protein (10 p,g pro te in /assay) was incubated the presence of 1 nM free Ca 2+ (open column), 300 nM free Ca 2+ (hatched column) or 300 nM free Ca 2+ and 1.9 tzM /3y subunits (shaded column). The results are means + S.E.M. of duplicate determinations and are representative of two similar experiments.

J.M. Dickenson et al. / European Journal of Pharmacology - Molecular Pharmacology Section 288 (1995) 393-398 397
pholipase C in DDTIMF-2 cells. The data presented show that membrane and soluble fractions prepared from DDTIMF-2 cells possess phospholipase C activity which can be stimulated by G protein /33' subunits purified from bovine retinal transducin. The /33' sub- units used in this study are composed of /31 and 3'1 subunits.
It is now widely accepted that the /3 isoforms of phospholipase C (phospholipase C /31 -3 ) are regulated by both G a subunits belonging to the Gq class of G proteins (Gaq, G a l l , Gala and Goq6) and by G/33' subunits (Camps et al., 1992; Clapham and Neer, 1993). Furthermore, the G a subunits activate phospholipase C/3 isoforms in the order of PLC/31 > PLC/33 >> PLC /32, whereas /33' subunits activate in the order of PLC /33 > PLC /32 > PLC/31 (Park et al., 1993).
In contrast to the marked activation of phospho- lipase C by/33' subunits, direct activators of G protein a subunits G T P y S or A1F 4 did not stimulate mem- brane bound phospholipase C from DDT1MF-2 cells (data not shown). However, Western blot analysis did reveal the presence of G protein Otq/ll subunits in DDT1MF-2 cell membranes (data not shown). These data suggest that DDTIMF-2 cells may contain a phos- pholipase C with low sensitivity to stimulation by O/q/11 subunits, e.g. phospholipase C 132. This isoform of phospholipase C is not activated by aq in the presence of either AlF 4- or G T P y S (Park et al., 1992) but is markedly stimulated by 137 subunits (Park et al., 1993). Alternatively, the extremely low affinity of Gc~q and G a n for GTP3'S may explain the lack of G T P y S effect (Helper et al., 1993).
We screened DDTIMF-2 cell membrane and solu- ble fractions for the presence of phospholipase C /3 isoforms using antibodies selective for bovine phospho- lipase C /31, human phospholipase C /32 and human phospholipase C /33- These antibodies were raised against synthetic oligopeptides based upon C-terminal sequences specific for each of the phospholipase C /3 isoforms (Carozzi et al., 1992). Western blot analysis did not reveal any immunoreactive proteins in the expected molecular weight range for phospholipase C /3 isoforms, i.e. ca. 150 kDa (Fig. lc). However, several immunoreact ive proteins, with smaller molecular weights, were detected which may correspond to prote- olytic degradation products of phospholipase C. It is possible that the failure to detect phospholipase C /3 isoforms could be because antisera raised against bovine (phospholipase C /31) and human (phospho- lipase C /32 and /33) sequences do not react with phospholipase C 13 isoforms present in DDTIMF-2 cells, which originate from hamster vas deferens smooth muscle. The screening of numerous tissues and cell lines has revealed that phospholipase C /31 is found predominantly in the brain, whereas phospholipase C /33 is widely distributed (Smrcka and Sternweis, 1993).
In contrast, phospholipase C /32 has a much more limited distribution and to date has only been detected in HL60 ceils (Park et al., 1992; Smrcka and Sternweis, 1993; Jhon et al., 1993), and bovine and human neu- trophils (Camps and Gierschik, unpublished observa- tions). The identity of the phospholipase C sensitive to /33' subunits in DDTIMF-2 cells remains to be estab- lished.
The results presented in this communication sup- port our hypothesis that G i protein derived /33' sub- units are involved in the pertussis toxin sensitive activa- tion of the phospholipase C / C a 2÷ signalling pathway by the adenosine A~ receptor in DDTIMF-2 cells (White et al., 1992; Dickenson and Hill, 1993,1994). In this cell line ty i and /33, subunits released following adenosine A~ receptor activation may independently inhibit adenylate cyclase and stimulate phospholipase C respectively (Gerwins and Fredholm, 1992a,b; Dick- enson and Hill, 1994).
Acknowledgements
J.M.D. and S.J.H. thank the Wellcome Trust for financial support. M.C. and P.G. were supported by a grant from the Deutsche Forschungsgemeinschaft.
References
Bradford, M.M., 1976, A rapid and sensitive method for the quanti- tation of microgram quantities of protein utilizing the principle of protein-dye binding, Anal. Biochem. 72, 248.
Camps, M., C. Hou, D. Sidiropoulos, J.B. Stock, K.H. Jakobs and P. Gierschik, 1992, Stimulation of phospholipase C by guanine- nucleotide-binding protein /33' subunits, Eur. J. Biochem. 206, 821.
Carozzi, A.J., R.W. Kriz, C. Webster and P.J. Parker, 1992, Identifi- cation, purification and characterization of a novel phosphatidyl- inositol-specific phospholipase C, a third member of the /3 sub- family, Eur. J. Biochem. 210, 521.
Clapham, D.E. and E.J. Neer, 1993, New roles for G protein /3y-di- mers in transmembrane signalling, Nature 365, 403.
Dickenson, J.M. and S.J. Hill, 1993, Adenosine Avreceptor stimu- lated increases in intracellular calcium in the smooth muscle cell line DDTxMF-2, Br. J. Pharmacol. 108, 85.
Dickenson, J.M. and S.J. Hill, 1994, Interactions between adenosine A 1- and histamine Hi-receptors, Int. J. Biochem. 26, 959.
Gerwins, P. and B.B. Fredholm, 1992a, ATP and its metabolite adenosine act synergistically to mobilise intracellular calcium via the formation of inositol 1,4,5-trisphosphate in a smooth muscle cell line, J. Biol. Chem. 267, 16081.
Gerwins, P. and B.B. Fredholm, 1992b, Stimulation of adenosine Al-receptors and bradykinin receptors, which act via different G-proteins, synergistically raises inositol 1,4,5-trisphosphate and intracellular calcium in DDTIMF-2 smooth muscle cells, Proc. Natl. Acad. Sci. USA 89, 7330.
Helper, J.R., T. Kozasa, A.V. Smrcka, M.V. Simon, S.G. Rhee, P.C. Sternweis and A.G. Gilman, 1993, Purification from Sf9 cells and characterization of recombinant Gq,~ and Gll,~ , J. Biol. Chem. 268, 14367.

398 Z M. Dickenson et al. /European Journal o f Pharmacology - Molecular Pharmacology Section 288 (1995) 393-398
Jhon, D-Y., H.-H. Lee, D. Park, C.-W. Lee, K.-H. Lee, O.J. Yoo and S.G. Rhee, 1993, Cloning, sequencing, and purification, and Gq-dependent activation of phospholipase C /33, J. Biol. Chem. 268, 6654.
Park, D., D.-Y. Jhon, R. Kriz, J. Knop and S.G. Rhee, 1992, Cloning, sequencing, expression, and Gq-independent activation of phog- pholipase C /32, J- Biol. Chem. 267, 16048.
Park, D., D.-Y. Jhon, C.-W. Lee, K.-H. Lee and S.G. Rhee, 1993, Activation of phospholipase C isozymes by G protein /3,/ sub- units, J. Biol. Chem. 268, 4573.
Ramkumar, V., W.W. Barrington, K.A. Jacobson and G.L Stiles. 1990, Demonstration of both A l and A 2 adenosine receptors in DDT1MF-2 smooth muscle cells, Mol. Pharmacol. 37, 149.
Smrcka, A.V. and P.C. Sternweis, 1993, Regulation of purified subtypes of phosphatidylinositol-specific phospholipase C/3 by G protein a and /3,/ subunits, J. Biol. Chem. 268, 9667.
White, T.E., J.M. Dickenson, S.P.H. Alexander and S.J. Hill, 1992, Adenosine A~ receptor stimulation of inositol phospholipid hy- drolysis and Ca 2+ mobilization in DDTIMF-2 cells, Br. J. Phar- macol. 106, 215.