acetoin catabolic system of klebsiella cg43: sequence,jb.asm.org/content/176/12/3527.full.pdf ·...
TRANSCRIPT

Vol. 176, No. 12JOURNAL OF BACrERIOLOGY, June 1994, p. 3527-35350021-9193/94/$04.00+0Copyright X 1994, American Society for Microbiology
Acetoin Catabolic System of Klebsiella pneumoniae CG43: Sequence,Expression, and Organization of the aco Operon
WEN-LING DENG,' HWAN-YOU CHANG,2 AND HWEI-LING PENGI*
Departments of Microbiology and Immunology,' and Molecular and Cellular Biology, 2Chang Gung College of Medicine and Technology, Kwei San, Tao Yuan, Taiwan
Received 6 January 1994/Accepted 25 March 1994
A cosmid clone which was capable of depleting acetoin in vivo was isolated from a library of Klebsiellapneumoniae CG43 cosmids. The smallest functional subclone contained a 3.9-kb DNA fragment of the cosmidclone. Sequencing of the DNA fragment revealed three open reading frames (ORFs A, B, and C) encodingpolypeptides of 34, 36, and 52 kDa, respectively. The presence of these proteins was demonstrated by expressionof the recombinant DNA clone in Escherichia coli. Considerable similarities between the deduced amino acidsequences of the ORFs and those of the following enzymes were found: acetoin dissimilation enzymes, pyruvatedehydrogenase complex, 2-oxoglutarate dehydrogenase complex, and branched-chain 2-oxo acid dehydroge-nase complex of various origins. Activities of these enzymes, including acetoin-dependent dichlorophenolin-dophenol oxidoreductase and dihydrolipoamide acetyltransferase, were detected in the extracts of E. coliharboring the genes encoding products of the three ORFs. Although not required for acetoin depletion in vivo,a possible fourth ORF (ORF D), located 39 nucleotides downstream of ORF C, was also identified. The deducedN-terminal sequence of the ORF D product was highly homologous to the dihydrolipoamide dehydrogenasesof several organisms. Primer extension analysis identified the transcriptional start of the operon as an Aresidue 72 nucleotides upstream of ORF A.
Acetoin (3-hydroxy-2-butanone) is a major fermentationproduct of many bacteria when they are grown on a mediumwith excess carbohydrates. This neutral product allows thebacteria to ferment large amounts of carbohydrates withoutself-inhibition. It also can be reutilized after the carbohydratesare exhausted and is therefore considered an energy-storingmetabolite.Many bacterial species are able to degrade acetoin, and
some of the catabolic pathways have been elucidated. Accord-ing to Juni and Heym (17), the dissimilation of acetoin byAcinetobacter spp. proceeds via a 2,3-butanediol cycle, in whichthe acetoin is an intermediate and is finally converted toacetate. Bacillus subtilis also degrades acetoin. The complete2,3-butanediol cycle, however, is not essential for the dissimi-lation of acetoin in this bacterium (21). Instead, acetoin iscatabolized by an oxidative cleavage (22) with acetaldehydeand acetyl coenzyme A as the end products, as in Alcaligeneseutrophus (10), Streptococcus faecalis (9), and Pelobactercarbinolicus (25).The cleavage of acetoin inA. eutrophus is catalyzed by enzymes
encoded by the acoXABC operon (32). acoA and acoB are thestructural genes for the a and ,B subunits of acetoin:dichlorophe-nolindophenol oxidoreductase (Ao:DCPIP OR). The aminoacid sequences deduced from acoA and acoB exhibit significanthomology to the Ela and E1B sequences of various 2-oxo aciddehydrogenase complexes. Amino acid sequence similaritiesbetween the fast-migrating protein encoded by acoC and thedihydrolipoamide acetyltransferase (DHLAT) of E. coli havealso been found. The function of the first gene productencoded by acoX has not been identified. Additional geneproducts, including the acetaldehyde dehydrogenase II whichis encoded by acoD (33) and the regulator AcoR (19), are also
* Corresponding author. Mailing address: Department of Microbi-ology and Immunology, Chang Gung College of Medicine and Tech-nology, Kwei San, Tao Yuan, Taiwan. Phone: 886-3-3283016, ext. 5134.Fax: 886-3-3283031.
required for acetoin catabolism in A. eutrophus. Most recently,the P. carbinolicus aco genes were also isolated (26). There areconsiderable similarities in the amino acid sequences corre-sponding to acoA, acoB, and acoC between the two acetoindehydrogenase enzyme systems. Molecular organizations ofthe structural genes for Ela, Eip, and E2 components are alsosimilar in the two bacteria. The P. carbinolicus acoL genecoding for dihydrolipoamide dehydrogenase (DHLDH), how-ever, was not found in the A. eutrophus aco operon.
In Klebsiella spp., acetoin is an intermediate of the 2,3-butanediol biosynthetic pathway (16, 46). Three enzymes areinvolved in the pathway, a-acetolactate decarboxylase, a-ace-tolactate synthase, and acetoin (diacetyl) reductase. The genesencoding these enzymes compose the budABC operon (2).Acetoin reductase (EC 1.1.1.5) catalyzes the reversible conver-sion of acetoin into 2,3-butanediol. However, under normalphysiological conditions, acetoin reductase only carries out theforward reaction, i.e., the synthesis of 2,3-butanediol. Although2,3-butanediol is thought to be the end product of the pathway,a significant increase in the amount of acetoin parallel with thedecrease in 2,3-butanediol concentration occurred after 14 hwith batch cultures (16). Thus, acetoin is eventually accumu-lated in the culture containing excess carbohydrates. No evi-dence that acetoin could be metabolized by enzymes otherthan acetoin reductase in Klebsiella spp. has been reported. Inthis paper, we report the isolation of an operon from Klebsiellapneumoniae CG43 which is able to provide E. coli cells with anacetoin dissimilation activity. The results demonstrate theexistence of an alternative route to the 2,3-butanediol pathwayfor acetoin metabolism in this organism.
MATERIALS AND METHODS
Bacterial strains, plasmids, and growth conditions.K pneu-moniae CG43, a K2 strain presenting a strong Voges-Proskauer reaction (29, 30), was the source for the genomicDNA preparation. K pneumoniae ATCC 13883, a type strain,
3527
on May 21, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

3528 DENG ET AL.
was obtained from the Culture Collection and Research Cen-ter, Hsin Chu, Taiwan; K pneumoniae M5al was obtainedfrom M. J. Merrick, Agriculture and Food Research CouncilUnit of Nitrogen Fixation, University of Sussex, Brighton,United Kingdom. E. coli DH1 [F- supE44 recAl endA41 gyrA96thi hsdR17 (rK- MK+) relAl] was the host for cosmid libraryconstruction. E. coli JM109 [F' traD36 lacjq A(lacZ)M15proA+B+ A(lac-proAB) thi gyrA96 end4l hsdl7 (rK-mK+)relAl supE44 recAl] was the recombinant DNA host. E. coliBL21 (DE3), a A DE3 lysogen with a T7 RNA polymerase genein an isopropyl-f-D-thiogalactopyranoside (IPTG)-inducibleform (47), was used for in vivo protein analysis. Cosmid vectorpHC79 was used for the construction of the K pneumoniaecosmid library. Plasmid pGEM-1 was the vector for expressionpurposes, and pUC18 was routinely used as the cloning vector.Both K pneumoniae and E. coli were grown at 37°C in Luriabroth (LB) unless otherwise indicated. M9 (39) supplementedwith 0.2% acetoin served as minimal medium.Enzymes and chemicals. Restriction endonucleases and
DNA-modifying enzymes, including avian myeloblastosis virusreverse transcriptase and T4 DNA ligase, were purchased fromPromega and were used under the conditions recommended bythe supplier. All chemicals were reagent grade and wereobtained either from Merck (Darmstadt, Germany) or SigmaChemical Co. (St. Louis, Mo.). Isotopes (including [a- 2P]dCTP and 35S-dATP) were purchased from Amersham Corp.(Buckinghamshire, England).Assay for acetoin. LB (pH 6.0) supplemented with 0.01%
(vol/vol) acetoin was the culture medium for screening clonescarrying acetoin dissimilation activity. The amount of acetoinleft in the broth grown overnight was determined by themethod of Krampitz (18), which is based on the specificreaction of acetoin with creatine and a-naphthol at an alkalinepH. The reaction produces a bright red product, which wasmeasured spectrophotometrically at 540 nm.
Construction of cosmid library. Genomic DNA of K pneu-moniae CG43 was prepared by the method of Caswell et al. (6).The DNA was partially digested with Sau3AI, and the frag-ments were size fractionated and ligated to the BamHI site ofcosmid vector pHC79. The ligation mixture was packaged withX-coat proteins by using an in vitro packaging kit (Promega).The resulting phage particles were transfected into E. coliDH1.Recombinant DNA techniques and DNA sequencing. Plas-
mid DNA preparation and DNA manipulation were carriedout essentially as described previously (39). DNA sequencedetermination was performed by the dideoxy chain termina-tion method (40) with the Sequenase kits (U.S. Biochemical,Cleveland, Ohio). Both universal M13 primer and syntheticoligonucleotides for primer-hopping strategy were used. Thesites of compressions were resolved by using dITP, and thesequence was confirmed in both strands. The nucleotide andamino acid sequences were analyzed with the DNASTARprogram (DNASTAR Inc., Madison, Wis.) on a Macintosh LCcomputer.
Expression of acoA, acoB, and acoC in E. coli. E. coli strainBL21 containing the recombinant plasmid was grown in LBsupplemented with 100 ,ug of ampicillin per ml at 37°C withvigorous shaking until the optical density at 600 nm (OD600)reached 0.4. IPTG was added to a final concentration of 1.0mM, and the incubation was continued at 37°C for 120 min.The cell pellet was suspended in 2x Laemmli sample bufferand boiled before being loaded on a sodium dodecyl sulfate(SDS)-polyacrylamide gel as described by Laemmli (20). Theprotein profiles were visualized with Coomassie brilliant blueR-250.
Ao:DCPIP OR activity assay. Activity of Ao:DCPIP OR wasdetermined by the method of Frund et al. (10) with slightmodification. Bacteria grown overnight were harvested bycentrifugation and suspended in 100 mM potassium phosphatebuffer (pH 7.0) for Ao:DCPIP OR activity assay. The assaymixture contained 100 mM potassium phosphate (pH 7.0), 0.08mM thiamine pyrophosphate, 0.5 mM magnesium chloride,0.10 mM DCPIP, 0.4 mM acetoin, 50 pl of toluene, and thebacterial suspension in a final volume of 1.0 ml. The reactionmixture was incubated at 37°C, and the amount of clarifiedsupernatant at OD578 was measured. The presence of toluenerendered the cells permeable and allowed the reaction tocontinue without detectable interference. We have also foundthat Ao:DCPIP OR activity obtained through this modifi-cation is higher than that obtained from the cell extract bysonication. One unit of DCPIP OR activity is defined as thereduction of 1 ,umol of DCPIP per min. Specific activity isrecorded in units per milligram of protein. The amount ofprotein is determined by assuming that 109 cells yieldapproximately 150 ,ug of protein as estimated according toreference 24.DHLAT activity assay. DL-Dihydrolipoamide, the substrate
for the DHLAT assay, was prepared by reduction of DL-lipoamide with potassium borohydride (35). DHLAT activitywas determined by measuring the formation of the coloredhydroxamate ferric complex according to the procedure ofReed and Willms (34) with slight modification. The assaymixture contained 100 mM Tris-HCl (pH 7.5), 10 mM acetylphosphate, 4 mM DL-dihydrolipoamide, 0.13 mM coenzyme A,10 U of phosphotransacetylase, 50 pl of toluene, and thebacterial suspension in a final volume of 1.0 ml. The reactionmixture was incubated at 30°C for 30 min, terminated by theaddition of 0.1 ml of HCl (1 N), and heated at 100°C for 5 min.Hydroxylamine and FeCl3 were then added to the supematant,and its OD540 was measured. One specific unit of DHLATactivity is defined as the formation of 1 ,umol of acetylphosphate per min per mg of protein.Primer extension assay. RNA was isolated (48) from K
pneumoniae grown in acetoin-containing minimal medium.The oligonucleotides were obtained from TIB MOLBIOL,Berlin, Germany. To ensure the accuracy of the results, twodifferent primers were used independently. Primers CH61(5'-GCCTGTITGCTGAGCATCGT-3') and CH62 (5'-TCTGATCTCCCGCATCTT-3') are the reverse complements ofpositions 327 to 346 and 366 to 383 of the nucleotide sequence,respectively. The assay mixture for primer extension contained10 pmol of either CH61 or CH62 synthetic oligonucleotide, 10,ug of cellular RNA, 0.2 mM (each) dATP, dTTP, and dGTP,10 ,ICi of [a-32P]dCTP (111 TBq/mmol; New England Nucle-ar), 5 U of RNasin, and 5 U of avian myeloblastosis virusreverse transcriptase. The reaction was performed at 42°C for15 min. Excess cold dCTP was added, and the reaction wascontinued for another 10 min. The primer extension productwas analyzed on a sequencing gel by using the sequence laddergenerated by the same primer.
Nucleotide sequence accession number. The nucleotide se-quence has been submitted to GenBank, and accession num-ber U00985 has been assigned.
RESULTS
Acetoin catabolism in K. pneumoniae. To determine if Kpneumoniae CG43 is able to utilize acetoin as its sole carbonsource, the bacteria were inoculated to an OD6o0 of 0.1 intoM9 minimal medium supplemented with varied concentrationsof acetoin and grown at 37°C with vigorous shaking. The
J. BACTERIOL.
on May 21, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

acoABCD OPERON OF K PNEUMONIAE 3529
growth of bacteria at OD600 was monitored. A doubling time ofapproximately 8 h was observed for K pneumoniae CG43 inthe presence of 0.2% acetoin. The growth of the bacteria wasaccompanied by a decrease in the acetoin concentration. E.coli, however, did not grow under the same conditions, and nosignificant amount of acetoin in the medium was lost duringincubation. Very little growth was observed forK pneumoniaeCG43 when 2,3-butanediol was used as the carbon source.Thus, the utilization of acetoin by K pneumoniae does notadopt 2,3-butanediol as an intermediate. The presence of analternative pathway in K pneumoniae for the dissimilation ofacetoin was therefore investigated.
Isolation of the genes responsible for acetoin catabolism.Since E. coli does not have the ability to utilize acetoin, it maybe used to screen for genes which code for the acetoinutilization activity. To screen for the presence of genes in-volved in acetoin catabolism, about 1,000K pneumoniae CG43cosmid clones in E. coli DH1 were transferred individually to96-well plates containing LB (pH 6.0) and 0.01% acetoin. Afterovernight incubation at 37°C, the amounts of acetoin remain-ing in the medium were determined. Two positive cosmidclones with acetoin-dissimilating capability, as demonstratedby their inability to produce the bright red color in thescreening reaction, were obtained. The two clones were foundto contain DNA fragments overlapping each other, and henceonly one of the clones, designated pHP654, was furthercharacterized. A functional subclone (designated pHP679; seeFig. 5A) containing a 3.9-kb DNA fragment was obtained byserial deletions of pHP654 and confirmed by acetoin depletionactivity assay.
Since the depletion of acetoin in the cell culture grownovernight could also be due to the conversion of acetoin into2,3-butanediol by acetoin reductase which was encoded by thebudC gene (2), acetoin conversion in vivo by acetoin reductasewas examined by using cosmid clone pCD64, which containsthe entire bud operon of K pneumoniae (29). E. coli DH1(pCD64), however, exhibited a much weaker acetoin-depletingactivity than E. coli DH1(pHP654). In addition, no cross-hybridization between the inserts of pCD64 and pHP654 wasobserved, which indicates that these two clones are unrelated.These results further supported the postulate that there is analternative to the 2,3-butanediol pathway for acetoin catabo-lism in K pneumoniae.
Nucleotide sequence of the DNA involved in acetoin catab-olism. The nucleotide sequence of the 3.9-kb insert of thesmallest functional clone, pHP679, was determined and isshown in Fig. 1. Analysis of the nucleotide sequence revealedthree major open reading frames (ORFs) (designated A, B,and C) which are capable of encoding polypeptides of 319, 339,and 493 amino acid residues with calculated molecular weightsof 34,021, 36,561, and 52,216, respectively. There are 11 basesbetween ORF A and ORF B and only 3 bases between ORF Band ORF C. A possible fourth ORF (designated ORF D),located 39 bp downstream of ORF C, was also observed. Thetranslation start codons of all four ORFs are preceded byShine-Dalgarno sequences.Amino acid sequence comparison. The deduced amino acid
sequences of the ORFs were used to search for homologousdata files in GenBank. Striking similarities to the followingenzymes were observed: A. eutrophus aco operon (32)-encod-ing enzymes, P. carbinolicus acetoin dehydrogenase (26), pyru-vate dehydrogenase (PDH) complex (13-15, 27, 43-45), 2-oxo-glutarate dehydrogenase (OGDH) complex (36, 42, 49), andbranched-chain 2-oxo acid dehydrogenase (BCODH) complex(3, 4, 41) of various origins (Fig. 2 through 4). The greatestamino acid homologies were found to be those between the
ORF A product and A. eutrophus Ao:DCPIP OR subunit cx(59.4%) (32), between the ORF B product and A. eutrophusAo:DCPIP OR subunit , (56.8%) (32), and between the ORFC product and E. coli PDH E2 (28.7%) (44).A putative thiamine pyrophosphate binding motif, GDG-
n7-8-E-n4-A-n5^-P-n6-NN (12), was identified in the deducedamino acid sequence of ORF A but not in that of ORF B (Fig.2). As with E. coli PDH E2 (1, 44), a lipoyl-binding site and theE3-binding sequence were also noted in the gene product ofORF C (Fig. 3A and B). The C-terminal region of the ORF Bproduct, which represents the El-binding sequence and theinner core domain, however, exhibited a greater homology(Fig. 3C) to E. coli OGDH E2 (42) and to OGDH E2 of otherorigins (4, 36, 49). In the partial amino acid sequence of theputative ORF D product, a conserved sequence representingthe flavine adenine dinucleotide (FAD)-binding domain I (7)in Pseudomonas lipoamide dehydrogenase, P. carbinolicusAcoL (26), and PDH E3 of E. coli (45) and other organisms (5,14, 28) was identified (Fig. 4).These homology studies suggest that the enzymes encoded
by ORFs A, B, C, and D work together in a way similar to thatof enzymes in the acetoin dehydrogenase systems of A. eutro-phus and P. carbinolicus and that of 2-oxo acid dehydrogenasemultienzyme complexes of various organisms. The gene clusterthat codes for the ORFs is therefore designated acoABCD.
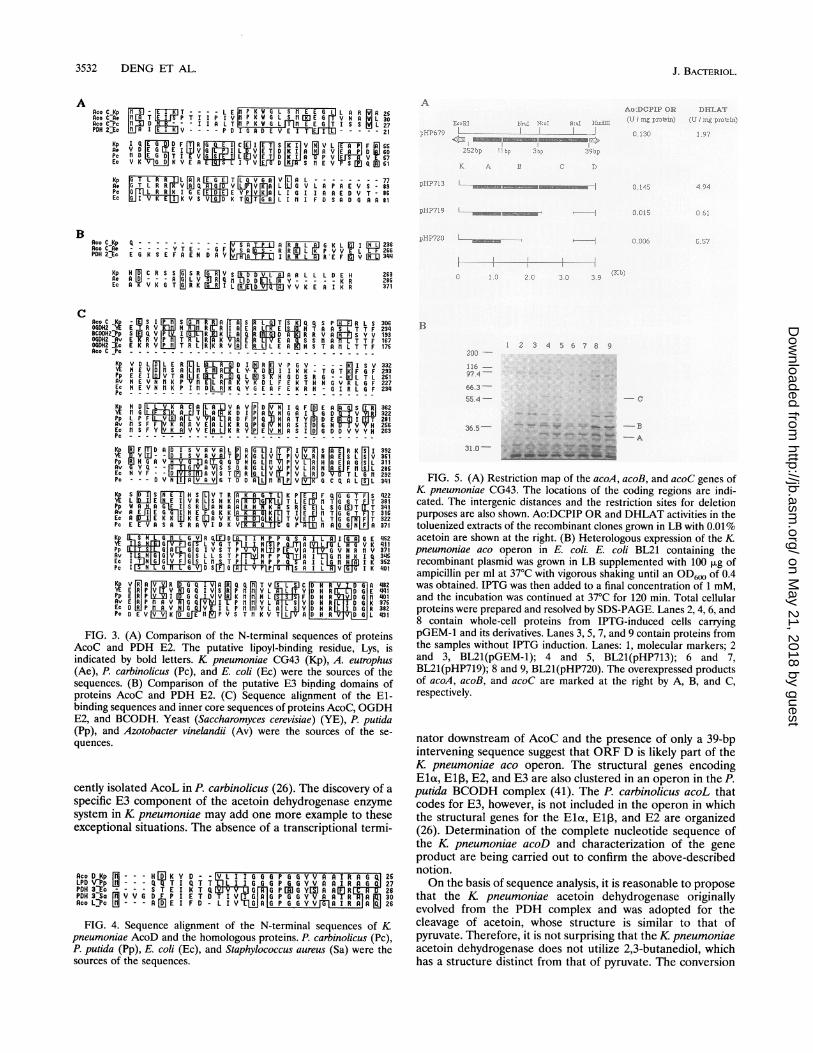
Expression of the K. pneumoniae aco genes in E. coli. In orderto identify the aco gene products encoded by the insert DNAof pHP679, plasmid pHP713 (Fig. 5A) was constructed byinserting the 3.9-kb EcoRI-HindIII fragment of pHP679 intothe expression vector, pGEM-1. Examination of the proteinssynthesized in vivo following induction with IPTG indicatedthat pHP713 was capable of synthesizing three proteins. Asshown in Fig. SB, the proteins could be easily identified evenin the absence of IPTG. The sizes of these three proteins,as determined by SDS-polyacrylamide gel electrophoresis(PAGE), are approximately 34, 36, and 52 kDa, which agreewell with those of the deduced proteins of the ORFs. To assigneach of the in vivo-synthesized proteins to the correspondinggene, several deletion subclones of pHP713 were constructed(Fig. SA). Plasmid pHP719 was derived from pHP713 with adeletion of an NcoI-StuI fragment containing part of the ORFC sequence. Plasmid pHP720 was generated by deleting anNruI-StuI fragment which contains parts of both the ORF Bsequence and the ORF C sequence of pHP713. The proteinprofiles of bacteria containing pHP719 and pHP720 werethen analyzed in parallel with those of the cells harboringthe parental plasmid, pHP713. As shown in Fig. 5B, twosmaller proteins (A and B) from pHP719 were identified,and the 34-kDa protein was the only protein from pHP720that was identified. Therefore, the 52-kDa protein wasdesignated AcoC, and the 36-kDa protein was called theacoB gene product. The sizes of the proteins are alsoconsistent with the molecular weights calculated from thededuced amino acid sequences of the corresponding geneproducts.
Enzymatic activities of the K. pneumoniae aco genes. Inaddition to K pneumoniae CG43, the K pneumoniae typestrain (ATCC 13883) and MSal were also grown in M9supplemented with 0.1% Casamino Acids and 0.2% acetoin.Comparable Ao:DCPIP OR activities in the extracts of theseK pneumoniae strains were detected, which further supportsthe presence of an acetoin utilization system inK pneumoniae.On the basis of sequence analysis, ORF A and ORF B most
likely encode an enzyme with Ao:DCPIP OR activity, whereasthe ORF C protein should possess DHLAT activity. To verifythis theory, the enzymatic activities in the extracts of K
VOL. 176, 1994
on May 21, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

3530 DENG ET AL.
acoK CGCAACCCTCICITCAICCCCAATIIrCCCACACrACTCAICCTCATT CICACCT58 TGTAAATACCAATICTTCATCCGTGGTGTICTACTCACTTTGTTTTTTTCAATTGTACGCCAGCCCCAGGGATGACATGG137 GGCAGCCAAACTTTATTGCTACACAAAATTTAGAAAGTTTGACCTCAAGCATGAAAACCCCCACCTCCCCCCCATGAA216 GGTGGCTTCGCACTCCCTTTAACCGCTCGTAGATTCATTGCATGGGTGTGGCCAGGGTTATCCGCCCTGTCTGTTGGCC
acoA Met Leu Ser Lys GIn Ala Leu Leu Gin Ala Tyr295 CCGCCTGGGTGACCTTTATTGACCAIAATTAAACG ATG CTC AGC AAA CAG GCG TTA TTG CAG GCT TAC
Arg Lys Met Arg Glu lie Arg Thr Phe Glu Glu Arg Leu His Gin Glu Asn Thr Ser Gly363 CGC AAG ATG CGG GAG ATC AGA ACA TTT GAA GAG CGG TTG CAT CAG GAA AAT ACC AGC GGT
Asp lie Pro Gly Phe lie His Leu Tyr Thr Gly Glu Glu Ala lie Ala Val Gly Val Cys423 GAT ATT CCG GGC TTC ATT CAC CTT TAC ACC GGT GAG GAG GCC ATC GCG GTG GGG GTC TGC
Glu Asn Leu Thr Ser Ala Asp Phe Ile Gly Ser Thr His Arg Gly His Gly His Cys Ile483 GAA AAT TTA ACG AGC GCA GAT TTT ATT TCA ACA CAC CGT GGA CAT GGC CAC TGT ATT
Ala Lys Gly Cys Asp lie His Gly Met Met Ala Glu lie Phe Gly Lys Asp Ser Gly Leu543 GCT AAA GGT TGC GAC ATT CAC GGC ATG ATG GCC GAA ATA TTC GGT AAG GAC AGC GGG TTA
Cys Arg Gly Lys Gly Gly Ser Met His lie Alo Asp Leu Ser Lys Gly Met Leu Gly Ala603 TGT CGC GGT AAG GGC GGG TCG ATG CAC ATT GCC GAT CTG TCG AAA GGA ATG CTG GGG GCG
Asn Ala lie Val Gly Gly Ala Pro Pro Leu Ala lie Gly Ala Ala Leu Thr Ala Lys Thr663 AAC GCT ATC GTT GGG GGA GCC CCT CCC CTG GCC ATC GGC GCC GCG CTA ACG GCA AAG ACG
Leu Lys Thr Gly Asn Vol Gly Val Ser Phe Thr Gly Asp Gly Gly Ser Asn Gln Gly Leu723 CTA AAA ACC GGC AAC C GGT GTC TCT TTT ACG GGC GAT GGT GGT TCT AAT CAG GGC CTG
Val Phe Glu Ala lie Asn Met Ala Val Val Leu Gin Leu Pro Ala Val Phe lie Phe Glu783 GTC TTT GAA GCC ATC AAT ATG GCC GTC GTG CTC CAG CTT CCA GCC GTC TTT ATT TTC GAG
Asn Asn Gly Tyr Gly Glu Gly Thr Gly His Asp Tyr Ala Val Gly Gly Arg Asp lie Ala843 AAT AAC GGT TAC GGC GAA GGA ACC GGC CAT GAC TAC GCC GTG GGT GGG CGT GAT ATC GCC
Arg Arg Ala Ala Gly Phe Gly Leu Pro Ala Val Thr Val Asp Gly Thr Asp Phe Phe Ala903 CGG CGC GCC GCT GGC TTC GGC CTG CCG GCA GTG ACC C GAT GGC ACC GAT TTC TTT GCC
Val Tyr Glu Ala Thr Ser Glu Ala Val Lys Arg Ala Arg Glu Gly Gly Gly Pro Ser Val963 GTT TAT GAG GCA ACC TCA GAG GCG GTC AAG CGT GCG CGA GAA GGC GGT GGC CCA AGC GTC
lie Glu Ala Lys Ala Phe Arg Trp His Gly His Phe Glu Gly Asp Pro Ala Leu Tyr Arg1023 ATT GAG GCC AAA GCC TTC CGC TGG CAT GGT CAT TTT GAG GGC GAT CCC GCG CTA TAT CGT
Ala Glu Gly Glu Val Gln Arg Leu Arg Clu Gln His Asp Pro Leu Lys lie Phe Thr Ala1083 GCG GAA GGT GAA GTG CAA CGC CTG CGT GAA CAA CAT GAT CCG CTG AAG ATT TTC ACC GCT
Lys Val Lys Gin His lie Thr Gin Glu Glu Leu Ala Ala lie Asp Glu Glu Val Glu Ala1143 AAG GTC AAG CAA CAT ATC ACC CAG GAA GM CTG GCG GCG ATT GAC GAG GAA GTA GAA GCC
Leu Val Asn Asp Ala Val Leu Lys Ala Arg Ala Ala Ala Tyr Pro Ala Pro Glu Asp Leu1203 CTG GTC AAC GAT GCC GTA TTG AAG GCC CGC GCC GCT GCC TAC CCG GCT CCG GAA GAC CTG
Leu Thr Asp Val Tyr Val Ser Tyr TER acoB Met Thr lie Lys Thr Tyr Arg1263 CTG ACA GAC GTG TAC C TCA TAC T115111 TGA TAA CC ATG ACA ATA AAA ACC TAT CGT
Glu Ala Val Lys Glu Ala Leu Ala Gln Glu Met Glu Arg Asp Glu Arg Vol Val Leu lie1322 GAA GCG GTC AAG GAA GCC CTG GCT CAG GAA ATG GAA CGC GAT GAA CGC GTG GTG CTC ATC
Ala Gly Glu Asp Leu Arg Gly Gly His Gly Gly Asn Ala Pro Glu Glu Alo Lys lie Glu1382 GGT GAS GAC TTG CGT GGC GGT CAT GGC GGA AAT GCG CCC GAA GAG GCT AAA ATA GAA GCC
Phe Gly Gly Val Leu Gly Val Thr Lys Gly Leu Trp Thr Gin Phe Gly Ser Asp Arg Val1442 TTT GGC GGT GTG CTG GGT GTC ACT AAA GGG CTC TGG ACG CAG TTC GGC TCC GAT CGG GTG
lie Asp Thr Pro lie Thr Giu Ser Ala lie lie Gly Met Ala Alo Gly Ala Ala Ala Thr1502 ATC GAC ACG CCC ATT ACC GAG TCG GCC ATC ATC GGT ATG GCG GCC GGC GCC GCA GCG ACG
Gly Leu Arg Pro Val Ala Glu Leu Met Phe Met Asp Phe Phe Gly Val Ser His Asp Ala1562 GGT TTG CGG CCG GTC GCG GAA TTG ATG TTC ATG GAT TTT TTT GGC GTG AGC CAC GAT GCA
Leu Tyr Asn Gln Ala Ala Lys Phe Arg Tyr Met Phe Gly Gly Lys Ala Arg Ala Pro Leu1622 GCG GCT AAG TTC CGC TAC ATG TTT GGT GGC AAA GCC CTG TAC AAC CAG AGA GCT CCG CTG
Val Met Arg Gly Met lie Gly Ala Gly Phe Ser Ala Ala Ala Gln His Ser Gln Ser Pro1682 GTG ATG CGA GGG ATG ATC GGC GCG GGG TTT TCT GCC GCA GCC CAG CAT TCA CAG TCA CCC
Tyr Asn lie Phe Ala Thr Thr Pro Gly Leu Lys Val Val Val Pro Ser Thr Pro Tyr Asp1742 TAT AAT ATC TTT GCC SIC ACG CCT GGA CTG AAG GTG GTG GTG CCC TCG ACG CCT TAT GAC
Vol Lys GIly Leu Leu Ile GIn Ser Ile Arg Asp Asp Asp Pro Val Val Phe Cys GCu His1802 GTC AAG GGT CTG TTA ATC CAG TCC ATT CGC GAC GAC GAC CCG GTG GTA TIC TGC GAG CAT
Lys Met Leu Tyr Asp Leu Lys Gly Glu Vol Pro Asp Giu le Tyr Thr Ile Pro Leu Gly1862 AAA ATG CTI TAC GAC CTC AAG GGC GAG GTA CCG GAC GAG ATC TAT ACC ATC CCG CTA GGT
Vol Ala Asn Tyr Thr Arg Glu Gly Glu Asp Val Thr Ile Ile Alo Leu Ser Ala Met Val1922 GTA GCC AAC TAC ACT CGC GAA GGG GAG GAC GTC ACC ATC ATT GCG TTG TCG GCA ATG GTA
His Lys Ala Asn Gin Val Ala Asp Lys Leu Ala Arg Glu Gly lie Ser Val Glu Val Val1982 CAT AAA GCA AAC CAG GTG GCG GAC AAA CTG GCC AGA GAG GGG ATC TCG GTC GAG GTG GTC
Asp Pro Arg Thr Ile Ser Pro Leu Asp Glu Glu Gly Ile Leu Glu Ser Val Ala Ser Thr2042 GAC CCI CIV SIC ATT TCG CCG CTG GAT GAG GAA GGT ATT CTG GAA TCG GTG GCG TCC ACG
Gly Arg Val Val Ile Val Asp Glu Ser Ala Ala Arg Phe Gly Phe Ala His Asp Val Ala
2102 GGG CGG GTC GTG ATT GTC GAC GAA TCC GCT GCA CGC TTC GGT TTT GCT CAT GAT GTC GCGAla Leu Ile Ala Ser Gin Ala Phe His Phe Leu Lys Ala Pro Val Leu Leu Val Thr Pro
2162 GCG CTG ATT GCG TCC CAG GCA TTC CAT TTC CTC AAA GCG CCC GTT CTG CTG GTG ACG CCGPro His Thr Pro Val Pro Phe Ser Pro Ala Leu Glu Lys Leu Trp Ile Pro Gly Val Glu
2222 CCA CAC ACG CCG GTC CCG TTC TCC CCT GCT CTC GAA AAA CTC TGG ATC CCT GGC GTT GAA
Arg Ile Glu Ala Ala Vol Arg Gln Val Leu Glu Asp TERacoCMet Ser Glu Ile Lys Thr
2282 CGT ATC GAA GCC GCC GTC CGT CAA GTG CTG 1CAGAT TAA CCT ATG AGC GAA ATC AAG ACGLeu Glu Met Pro Lys Trp Gly Leu Ser Met Glu Glu Gly Leu Leu Ala Arg Trp Ala Ile
2342 CTT GAA ATG CCA AAG TGG GGG CTT TCC ATG GAG GAA GGC TTG CTC GCT CGA TGG GCA ATC
GCn Glu Gly Asp Asp Phe Thr Arg Gly Gln Glu Ile Cys Glu Ile Glu Thr Ser Lys Ile2402 CAG GAG GGT GAC GAT TTC ACC AGA GGG CAG GAA ATA TGT GAG ATT GAA ACC AGT AAA ATC
Val Asn Val Leu Glu Ala Pro Phe Ala Gly Thr Leu Arg Arg Ile Leu Ala Arg Glu Gly2462 GTC AAT GTG CTG GAG GCC CCC TTT GCC GGT ACC TTA CGT CGG ATA CTC GCC CGC GAG GGT
Glu Thr Leu GCn Val Gly Ala Val Leu Ala Leu Ala Ala Asp Ala Ser Val Ser Asp Ala2522 GAG ACG CTT CAG GTA GGC GCC GTG CTG GCC CTG GCG GCT GAC GCG TCG GTC AGC GAT GCT
Glu Leu Asp Glu Phe Val Ala Arg Leu Ala Thr Ala Lys Pro Ala Ala Pro Gly Pro Glu2582 GAA CTG GAC GAA TTT GTT GCC CGC CTG GCG ACC GCG AM CCC GCA GCC CCA GGC CCG GAG
Ala Ala Ala Pro Asp Val Ala Ala GCn Ala Gly Ala Lys Pro Ala Ser Val Val Ser Pro2642 GCT GCC GCG CCG GAC GTA GCG GCA CAC CA GGC GCT AAG CCA GCT TCC GTT GTT TCG CCG
Pro Ser Asn Ser Pro Glu Pro Pro Vol Gly Gln Thr Val lie Pro Val Ser Leu Gin Gly2702 CCA TCC AAC AGC CCC GAG CCC CCT GTT GGG CAG ACC GTC ATC CCC GTC AGT CTG CAA GGT
Val Thr Asp Val Thr Gln Val Asn Ala Thr Pro His Ala Leu Arg Leu Ser Ala Arg Trp2762 GTG ACC GAT GTG ACT CAG GTT AAT GCC ACG CCC CAT GCG TTA CCV CTG TCT GCC CGC TGG
Gly Val Asp Leu Lys Lys Val Ala Ala Ala Gly Ala Gly lie Val Ser Leu Phe Leu lle2822 GGT GTC GAC CTG AAA AM GTC GCG GCA GCG GGC GCG GGG ATC GTA TCT CTG TTT CTG ATC
Trp Lys Ala Arg Ser Leu Pro Arg Gly Arg Leu Ala Ser Pro Thr Pro Pro Val Arg Arg2882 TGG AAA GCG CGA TCG TTG CCG CGC GGT CGC CTG GCC TCT CCG ACG CCC CCC GTT CGT CGC
Ser Lys Ala Pro Arg Ser His Ala Asp Asp Ser GIn Val Ser AIa Thr Pro Leu Ala Arg2942 AGC MA GCG CCC CGC TCG CAT GCC GAT GAC AGC CAG GTA TCG GCC ACC CCG CTG GCG CGC
Arg Leu Ala Gly Lys Leu Gly Ile Asn Lea His Asp Cys Arg Ser Ser Gly Ser Arg Gly3002 CGC CTG GCC GGC AAA CTG GGT ATC MC TTG CAT GAC TGC CGA AGC AGC GGT TCA CGC GGT
Arg Val Ser Arg Asp Asp Val Leu Ala Ala Ala Leu Leu Leu Asp Glu His Pro Gln Thr3062 CGG GTA AGT CGC GAT GAT GTG CTG GCC GCG GCG TTG TTA CTC GAC GAG CAC CCG CAG ACC
Ser Pro Val Gin Glu Ser Ala Pro Ala Pro Phe Glu Ser lie Pro Met Ser Gly Met Arg3122 AGT CCT GTG CAG GAG AGT GCC CCG GCC CCC TTT GAG AGC ATC CCG ATG TCC GGT ATG CGG
Arg Ala Ile Ala Ser Arg Leu Gln Thr Ser Lys Gln Gin Ser Pro His Phe Arg Leu Ser
3182 CGG GCT ATC GCT TCT CGC CTG CAA ACG TCC AAGCCAA CAG TCT CCC CAT TTC CGT TTG AGCVol Asp Leu Asp Leu CIu Arg Leu Leu AIa Leu Arg GCn Asp Ile Asn Arg GCu Val Pro
3242 GTC GAT CTC GAT CTG GAG CGA CTG TTA GCG TTA CGT CAG GAC ATT MC CGC GAA GTA CCC
Gly Val Lys Ile Ser Val Asn Asp Leu Leu Val Lys Ala Cys Ala Leu AIa Leu Val Ala
3302 GGC GTC AAG ATT TCG GTG AAT GAC CTG CTG GTG AAG GCC TGC GCC CTG GCG CTG GTG GCCVal Pro Asp Vol Asn Ile GCn Phe Asp Glu Ala Ala Gin Ser Ile Arg Arg Phe Thr Asp
3362 GTG CCT GAC GTC AAT ATC CAG TTT GAT GAA GCG GCA CM AGT ATC CII CGC TTT ACG GATAla Asp lie Ser Val Ala Val Ala Leu Pro Ala Gly Leu lie Thr Pro lie Val Arg Ser
3422 GCC GAC ATT TCG GTG GCC GTT GCG CTG CCT GCC GGG CTA ATT ACC CCT ATT GTT CGC TCG
Ala Glu Arg Lys Ser Ile Ser Asp Ile Ser Asn Glu Ile His Ser Leu Val Thr Arg Ala
3482 GCG GAA CGG AAA TCG ATT AGC GAC ATA TCG AAC GAA ATT CAC TCG CTG GTG ACC AGA GCC
Lys Ala Gly Thr Leu Lys Pro Glu Glu Phe Gln Gly Gly Thr Phe Ser Leu Ser Asn Leu3542 AAA GCG GGC ACG CTI AAG CCG GAA GAG TTC CAG GGC GGG ACC TTT AGC CTG TCA AAT CTG
Gly Met Leu GIly Val Arg Gin Phe Asp Ala Ile Ile Asn Pro Pro GIn Ser Ala Ile Leu3602 GGG ATG CTC GGC GTC CGG CAG TTT GAC CCC ATC ATC AAT CCA CCG CAA AGC GCA ATC CTC
Ala Ile Gly Alo Gly Glu Val Arg Ala Val Val Arg Asp Gly Gln Ile Val Ala Arg Gln3662 GCC ATT GGC GCC GGC GAG GTC CGG GCG GTA GTA CGC GAC GGG CAA ATC GTC GCG CIA CAG
Gln Met Thr Val Ser Leu Ser Cys Asp His Arg Val Ile Asp Gly Ala Ala Gly Ala Ala3722 CAA ATG ACG GTA TCG CTA TCC TGC CAT CAC CGG GTC ATC GAT GGC GCG GCA GGT GCG GCT
Phe Leu Arg Asn Ser Ser Asp TER3782 TTT CTC CGG AAC TCA AGC CAC TGA TTG AAA CCC CGA CCT TGA TGT TTA TCC ACIGAA CGA
acoD Met His Asp Lys Tyr Asp Val Leu le Ile Gly Gly Gly Pro Gly Gly Tyr Val Ala
3842 GCT ATG CAC GAT AAA TAC GAT GTG CTG ATC ATC GGC GGG GGT CCT GGG GGA TAC GTT GCCAla I le Arg Ala Gly Gin
3902 GCC ATC CGT GCC GGC CAG 3919
FIG. 1. Nucleotide sequence of the K pneumoniae CG43 aco operon. The amino acids encoded by the ORFs are shown above the nucleotidesequence. The putative Shine-Dalgarno sequences are underlined. The potential -10 and -35 sequences are indicated with dashed lines. Theposition and direction of ORF K (acoK) are also indicated.
pneumoniae CG43 and of several E. coli strains containingdifferent derivatives of pHP654 were analyzed. Both Ao:DCPIP OR and DHLAT activities could be demonstrated inthe extracts of E. coli DH1(pHP654) and K pneumoniae CG43cells grown on acetoin. The addition of acetoin to the culturemedium greatly enhanced Ao:DCPIP OR activity in E. coliDH1(pHP654). The Ao:DCPIP OR activity in the crudeextract of the cells induced with 0.01% acetoin for 60 minincreased approximately fourfold compared with that of unin-duced cells. The results indicated that acetoin could act as an
inducer. We also found that acetoin was the most appropriatesubstrate for the DCPIP OR activity. The enzymatic activitiescould also be detected by using diacetyl but neither 2,3-butanediol nor pyruvate as the substrate (Table 1). The resultssuggest that an acetyl group on a four-carbon compound iscrucial for the interaction with DCPIP OR.As shown in Fig. 5A, both Ao:DCPIP OR and DHLAT
activities were detected in the cell extracts of E. coliJM109(pHP679) and E. coli BL21(pHP713). Even withoutacetoin induction, the enzyme activities in these cell extractswere comparable to those of K pneumoniae CG43 and E. coliDH1(pHP654), suggesting that a regulatory gene in the origi-
nal cosmid clone for the aco operon was missing. On the basisof sequence analysis, the Ao:DCPIP OR activity can only bedetected in the extracts of recombinant E. coli clones thatcontain intact acoA and acoB sequences. On the other hand,DHLAT activity should be observed only in bacteria possessinga complete acoC gene. Thus, comparative studies of theenzymatic activities of E. coli BL21 harboring pHP713 contain-ing the three aco genes, pHP719 containing complete acoA andacoB sequences, and pHP720 containing only the acoA genewere carried out. In the crude extract of E. coli BL21(pHP720),which contained only the acoA gene product, neither Ao:DCPIP OR nor DHLAT activity was demonstrable. Theextract of E. coli BL21(pHP719), which contained both acoAand acoB gene products, exhibited Ao:DCPIP OR activity butno DHLAT activity. Only the strain carrying pHP713 showedboth Ao:DCPIP OR and DHLAT activities. These resultsindicated that proteins encoded by acoA and acoB indeed codefor the Ao:DCPIP OR activity and that the presence of theacoC gene product is necessary for DHLAT activity. Theseresults were further supported by the finding that purifiedAo:DCPIP OR from DH1(pHP654) was composed of twosubunits of 34 and 36 kDa (data not shown).
J. BACTERIOL.
on May 21, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

VOL.176,1994~~~~~~~~acoABCDOPERON OF K PNEUMONIAE 3531
~~~~BAco AKp -I QKOA F TQIYWR K nR EI RT 13 AcoAIKpfMlT IK T YR E 51V KFF A1ERItFWi?RTDR V2 D? 30R -ATe T ARA S D S AAL IPIKE T TV YR KRn RmOIR D 30 Rco BR-eAFR K L SIK LAI NIEID RR RD 30
Aco A-Pe n KTO-QLOKE D0 1LtK . RK R E IRE 22 AcoRBPc A RK OI n RLsYYLIGM 31
AeIFEL GV2F ROD GP F VH ATYORE ERA sojVRIIKRDeOVIERIRDOEA]R-KW1GJGIVRLRVYTLKRI 03Ae VEF AK 5T L pGFfV[H Y SRGEE J V G 62 PoYAG6AGTVTLDKERDS- wGGVLGylKR 07
KpE[ O mF IRTT5W W OWI[5I J~Kp s1RQ[lFf DTPGATF1~1AR5T~ A?8ReTI SAAAITINDRIDIRIOGAISTAROARCGAKOI rGJPV 7O WI RT4QJDT T SE A INGO A OA C R
~~~ R2 ~ ~ ~ ~ ~ ~RaVIAAJ31A~~~~~~~~~~~ICAKp~KeP N H NDFG M RO[AG L TR[ G 5GH TWITATT51fl 1 09 Aep AEI0I1I1T ?A~1 5iol1l 4 A AFRA 117ReRAf~~jEOYOKK OIOIOICIRIOKH ROGHOOCIAOALK RAYLE R]120F0 ROV FDI L N A KF Y 12
A OAAR]KEI GAOAF6GnCOA H1~ IFRA IRLISIVOG L 100IPennAE__AGRe IRFO A TI PIY JSRCA FJOR F JA ARO S A 1Pcj55jjJo~~~IVo~AoF IAOAAIOO[5JLR~~~JDI§JSL~~DK 1L2n11
A e T c A RQo T F L lAO
PC F AR[S N G T N NF D L P!JRI FJV 172
Kp F N NNGV fl SG 100H 1A V -- ARD RJR2A GRF 197
PceL IE N N OG YI EI AlD 202E~~S T OS P KY 0K V DON0
KP AfVTV D G T DF F AvVTW E[ K ATR 5E 227
PJ0 9 VT V AT D F F A E A A OR6 232
Kp VKE A F ROOGHF E RD PPAL v iAE E QV0207Re L0P ]EICIKIA IYG H F EOGDI A Q T VIV Al A D 2K 8
Pc D A A ROD C P L K F RODA A 0000A L V E RAA D OIE 292
Kp DE 1E D IiR[ PA Y P A PIE5T LT DY 310Ae IDR VA E VQE~o Q PRLE DL L TL D VI 328PC JK YL P D EAEKLJK AIDEI 322
KpReV332
PC 32K
Kp I P LKV VP TI LL Q I 177HLF I P C IP S T K RG 1...L AJ I 72
YIRW[EJREVPIEIOIAL OOL:IKSVO
V IinIEF[EDS
N~D P HD E 120K
KpID
VIVIOVii7E HKpc YNVDTREOGE D I S AQV. V [5D 23K
A eE R E IA 2322
Kp EIG ARe EO~ A~I DRTKR3376
FIG. 2. (A) Amino acid sequence comparison ofK pneumoniae (Kp) AcoA, A. eutrophus (Ae) AcoA, and P. carbinolicus (Pc) AcoA. The exact
matches are boxed, and the putative thiamine pyrophosphate binding consensus sequence is indicated by bold letters. (B) Amino acid sequence
comparison of K pneumoniae AcoB, A. eutrophus AcoB, and P. carbinolicus AcoB. The matched residues are boxed.
Primer extension analysis of the K. pneumoniae aco operon.
To determine the precise initiation site of transcription in the
acoABCD operon, primer extension was performed by usingthe RNA prepared from K pneumoniae CG43 under acetoin
induction. By using two different primers, the primer extension
products were mapped to the same adenyl residue 72 bp
upstream of acoA. The data obtained with one of the primersare shown in Fig. 6. The control region for the K pneumoniaeaco operon resembled the uF70 consensus elements in E. coli
(38). Upstream of the transcription initiation site, a potential-10 region, TAGATIT, was identified. A putative -35 site,
TTCGCA (Fig. 1), which is 17 bp away from the 10 site, was
also observed.
DISCUSSION
The sequencing data indicate that the gene organization of
the K pneumoniae CG43 acoABCD operon is similar to the
operons of the A. eutrophus and P. carbinolicus acetoin utili-
zation systems and to the 2-oxo acid dehydrogenase multien-
zyme complexes of several bacteria. It is conceivable that the
biochemical properties of these proteins are very similar. Thus,
acetoin is likely to be cleaved into tWo C2 compounds in the
acetoin utilization pathway ofK pneumoniae. The acetyl groupof one of the C2 compounds is transferred to a coenzyme A to
form acetyl coenzyme A by the catalysis of the acetoin-
dependent DCPIP OR, DHLAT, and DHLDH encoded by the
acoABCD operon. The other C2 compound, presumably an
acetaldehyde, is probably captured by an acetoin-dependent
acetaldehyde dehydrogenase.A significant homology was observed among the acoABCD
operon of K pneumoniae CG43, the acoXABC operon of A.
eutrophus (32), and the aco genes of P. carbinolicus (26). The
very high degree of similarity in the deduced amino acid
sequences corresponding to the two subunits of El (Fig. 2)
suggests that the El activity of the acetoin dehydrogenase
enzyme system is acetoin specific. The dependence of El on
acetoin is shown in Table 1. Blot may catalyze the initial step
of a nucleophilic attack on acetoin, as suggested by Opper-
mann and Steinbuchel (26). Elp is required either for correct
assembly of the El heterotetramers or for binding of El to the
E2 core (8, 13, 50). It also likely interacts directly with acetoin
in the reaction.
However, several noteworthy differences among the three
acetoin dehydrogenase enzyme systems were also demon-
strated. K pneumoniae AcoC is a protein composed of 493
amino acid residues which is 119 amino acids longer than the
corresponding fast-moving protein in A. eutrophus. The size of
K pneumoniae AcoC is, however, comparable to those of P.
carbinolicus AcoC and most E2 components of various 2-oxo
acid dehydrogenase multienzyme complexes identified so far,
whose molecular masses range from 46 to 52 kDa (4, 15, 36,
42). In contrast to the two lipoyl domains of the P. carbinolicus
AcoC, there is only one lipoyl domain in the E2s of K
pneumoniae and A. eutrophus. A single lipoyl domain, however,
is sufficient for the dehydrogenase complex activity (31). A
truncated ORF (ORF D) of 25 codons in length (Fig. 1) has
been identified in the nucleotide sequence of the insert of
pHP679. The deduced amino acid sequence has significant
homology (Fig. 4) with the N-terminal sequences of many
enzymes with DHLDH activity, including P. carbinolicus AcoL
(5, 14, 26, 28, 45). In contrast, analysis of the nucleotide
sequence of the A. eutrophus acoXA4BC operon did not identifya gene which encodes a DHLDH (32). An identical E3
component is usually shared by the various multienzyme
complexes of a given organism. Two exceptions found so far
are Pseudomonas putida, which has three different genes
encoding respective E3s with DHLDH4 activity, and the re-
A
VOL. 176, 1994
on May 21, 2018 by guest
http://jb.asm.org/
Dow
nloaded from
3532 DENG ET AL.
AAcoCKIp n P K W L S nE E G L R IA1?1Aco Nee T PTII [PIVPK WGLSn EE lET V N
L 3
AcRcCcn D I A L TI PR L T EG TI S S WL 27PDD2HE28 I KI Y - - - -PDIG D E WE E I1TE ---21
K p Im IS K I[V 1 SIW T1YLE Yl FW1 55
Ne V D E I VGI L Y VET D V PJD 1060
Pc n D T IEN E VI
QSTIT 1VAT DKIE m1PVV ES7ECV K V K YER AS I T V D1 Sn E Y S 61
S5
OpG TL R AIIL JIRGEIT[ GV1RA L-77Ae G T L RR AjQ yGLVV 1LI LG V L AP
PA Ev -
3
EcGil IGERR IEID IE Y V L IG I IP E D V T -88Ec gjI VKE KV S L£ KTETGR L I nI F D SARD G A A81
Aco CKp Q.-FTT-- -IYSAT-E I1 A' I I KIV -JIN230Aco CAe- TE- - FR L K
I V
PDE2Sc Ea EvKG T KE F A E N D yEIT a REFIr1 v1 341
Kp N RE NL L LD EH 263
a I AO 286
EC A0 K6 T lJR K OLDI L 0JJID YIVYK E A I R 371
r ..._ ..
i~~~~~~~~~~~~~~~~~~~~~~~~~~~~.';
AcoC P - IS I n R RSG QlAfLT1 QQSP[R JRLS 306
OODD 2 XE R N Han.R1RII R EZRLAERL E N T A ASLTTF 234BCODH 2-PpS Q V7 I Gr JLR KIVK AIQ R 51f751D R RVR A ET] IYV 193
O GD2 2AEv V R Pn T L IR K A E E A S AR
FL T T F 167OGDH2 EcE K RV P T RLRRK RV A E
L ER NS T An
LTTF171RCOC _PC
VD I L LIE R L DID I IRR vP G---IR1ISV 332
NE VD AaLI LE RL Y I I K - T GTTKIFGF 293PP EEI VT ALI E LR EL IHIS H
H AGD SR - K L TL 261N EVN K PV AEL K KD L F E K THN GVRLGF 227
Ec NE VNn K P I nD K Q V GEa FE K R H- GIRLGF 234
KP N DIL L AKLRClRILRVRV A_V PD ICqF1 ERIR S 362YE n G IL AF...R LDNIRD P A N G I D I 322Pp LPFP aL QILY A LRDF PQ N ATV D E 281AVn FF
YAVAREL KRQ P
IN S ID G N DIV
H 266EC S F Y R V LK R Y PE V N
S I D V V YH 263
KP FT D NIS V l WRG I YR SAAV ERA1
I 39 2YELYIII -- ID I S Y RVAR TIP GL VIT P V R NIElS L i 5Pp H A VYQ 6 NG n PV L R N
SIL 311
A G Y -- - RVIS1 S D R G V L IR N lalFn L 286EC N YF2--32S?RG V P VLR D TLGn 232PC - - -D V N TD D WIni v jEXG C QaLII]L 341KP Sl; SIN E Il H NSmV TRIRK R G T K P T FIS '122
L E E I RILS NK ANGI1nI TLIErG n TIGG TFIT 381
Pp w a ANG IE I IS RLAN NAA 153K S RIE L SGIG TT T341
v a IE IE N E R6jKKIRKKRA 61Kj T II El n TI T T 31TEAR EEK XK E AVK IGIKILl T VLTJ L TIGGrR1FIT 322Pc E E V S R I DKVl GilJG n aNLA 371
KILSNLG n-LG VR QMD1 I I P QSRA a IrG AlGE452IISNOOGI G LY 6 TP I IN a L HFr VK411
PPIT T IItGIPIfGI S TPP V N Rnv 371
V L L ST P N P P
SOIln H KIQ 34
Ec I NG GLSLns T PII N P P A1 G 11 16MIK362PC IrE n L GVD S L A I LV0I K401
VRN~~Q 4VO'82"EV [NR ID G Q IR1WRR1; T VIS L CID H R V I DGIRA8
ER PIYT:V V. IVISIRIPI YL Y D GIE 441PpEIR PIV V1 I aG Q VLIYIRI K n IIIIN LS IS IIF D N R D
6n 401
v EIRIPn AV G QYVJI LIP In Y L ISII H RILII DGIK 376EC D RPAn V GQ IL P nInI YL AIL S YIDHRIL GIR 382Pc D EV YrVY K PEPV S T K VT L A!. 'HR DL 431
FIG. 3. (A) Comparison of the N-terminal sequences of proteinsAcoC and PDH E2. The putative lipoyl-binding residue, Lys, isindicated by bold letters. K pneumoniae CG43 (Kp), A. eutrophus(Ae), P. carbinolicus (Pc), and E. coli (Ec) were the sources of thesequences. (B) Comparison of the putative E3 binding domains ofproteins AcoC and PDH E2. (C) Sequence alignment of the El-binding sequences and inner core sequences of proteins AcoC, OGDHE2, and BCODH. Yeast (Saccharomyces cerevisiae) (YE), P. putida(Pp), and Azotobacter vinelandii (Av) were the sources of the se-
quences.
cently isolated AcoL in P. carbinolicus (26). The discovery of a
specific E3 component of the acetoin dehydrogenase enzyme
system in K pneumoniae may add one more example to theseexceptional situations. The absence of a transcriptional termi-
Aco D Kp li--- H K Y D - -VL I I G GG P 6 G Y V A A I RAGQ 26
LPD Vp - - - Q T I Q TT L I I G G G P G G Y V a a I R a G l 27PDH 3OEc - - - - S T E I K T Gl P [I G Y A A RIC R D 26PDH 3 Sa IIV V G D .. P I E T D T IV P G G Y V a R A F1 30Aco L_Pc jl - - ODE I F D L IV A G P G Y v RAI AgJ 26
FIG. 4. Sequence alignment of the N-terminal sequences of Kpneumoniae AcoD and the homologous proteins. P. carbinolicus (Pc),P. putida (Pp), E. coli (Ec), and Staphylococcus aureus (Sa) were thesources of the sequences.
B
200-116 -97.4-
66.355.4 -
i 2 3 4 5 6 7 8 9
c
36.5 _ -B~~ --~~A
31.0 .a ,
FIG. 5. (A) Restriction map of the acoA, acoB, and acoC genes ofK pneumoniae CG43. The locations of the coding regions are indi-cated. The intergenic distances and the restriction sites for deletionpurposes are also shown. Ao:DCPIP OR and DHLAT activities in thetoluenized extracts of the recombinant clones grown in LB with 0.01%acetoin are shown at the right. (B) Heterologous expression of the Kpneumoniae aco operon in E. coli. E. coli BL21 containing therecombinant plasmid was grown in LB supplemented with 100pug ofampicillin per ml at 37°C with vigorous shaking until an OD600 of 0.4was obtained. IPTG was then added to a final concentration of 1 mM,and the incubation was continued at 37°C for 120 min. Total cellularproteins were prepared and resolved by SDS-PAGE. Lanes 2, 4, 6, and8 contain whole-cell proteins from IPTG-induced cells carryingpGEM-1 and its derivatives. Lanes 3, 5, 7, and 9 contain proteins fromthe samples without IPTG induction. Lanes: 1, molecular markers; 2and 3, BL21(pGEM-1); 4 and 5, BL21(pHP713); 6 and 7,BL21(pHP719); 8 and 9, BL21(pHP720). The overexpressed productsof acoA, acoB, and acoC are marked at the right by A, B, and C,respectively.
nator downstream of AcoC and the presence of only a 39-bpintervening sequence suggest that ORF D is likely part of theK pneumoniae aco operon. The structural genes encodingEla, E1,B, E2, and E3 are also clustered in an operon in the P.putida BCODH complex (41). The P. carbinolicus acoL thatcodes for E3, however, is not included in the operon in whichthe structural genes for the Elot, Elr, and E2 are organized(26). Determination of the complete nucleotide sequence ofthe K pneumoniae acoD and characterization of the geneproduct are being carried out to confirm the above-describednotion.On the basis of sequence analysis, it is reasonable to propose
that the K pneumoniae acetoin dehydrogenase originallyevolved from the PDH complex and was adopted for thecleavage of acetoin, whose structure is similar to that ofpyruvate. Therefore, it is not surprising that theK pneumoniaeacetoin dehydrogenase does not utilize 2,3-butanediol, whichhas a structure distinct from that of pyruvate. The conversion
A
p.-1PE.9
J. BACTERIOL.
B
Aw:)CIfPOiIn.;F -1 D)iL.AT
p-iPH:3 8MMM
on May 21, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

acoABCD OPERON OF K PNEUMONIAE 3533
TABLE 1. Substrate specificity of DCPIP OR'
Substrate" % Activity
Acetoin ............................................. 100.0Acetate............................................. 3.8Acetaldehyde............................................. 6.7Acetoacetate ............................................. 1.92,3-Butanediol ............................................. 3.8Diacetyl ............................................. 85.6Ethyl alcohol ............................................. 3.8Glycerol ............................................. 1.9Pyruvate ............................................. 3.8None............................................. 3.8
" The DCPIP OR (89 ,ug) of E. coli (pHP654), partially purified by chroma-tography on DEAE-agarose (Bio-Rad), was incubated at 37°C in 1 ml of 100 mMpotassium phosphate buffer (pH 7.0) in the presence of 0.10 mM DCPIP, 0.08mM thymine PPi, 0.5 mM MgCl2, and 0.4 mM substrate.
h The concentration of each substrate was 0.4 mM.
of 2,3-butanediol to acetoin in K pneumoniae CG43 is quitesluggish (data not shown), which may explain the poor growthof K pneumoniae CG43 on 2,3-butanediol. When K pneu-moniae CG43 was grown in M9 medium containing 0.2%acetoin, a doubling time of approximately 8 h was recorded.Although an acetoin depleting activity could be demonstratedin vivo for E. coli DH1(pHP654), the strain grew poorly withacetoin as the sole carbon source. This result is consistent withthe finding that E. coli cells carrying the entire aco operon ofA. eutrophus do not grow when acetoin is used as the solecarbon source (32). With A. eutrophus H16, at least anadditional locus encoding the acetoin-dependent acetaldehydedehydrogenase is required for the growth of the organism onacetoin (33). It is possible that the energy derived from thedissimilation of acetoin is produced sluggishly in the presence
G
T R C G TC_E.'.. ,.,'.'....
FIG.6.PireesnnliotenT, , ~~~~I
_n
-._ U v RThetnI
_-.vrtrsrtsanter6n
t. ~~~T
FIG.e 6. PrimenxeninaalssotheKaepierae Tneumoniaacdsoperon.
sequencing reaction products. The primer extension product and therelevant region of the sequence are shown together with the start site (*).
of only acoABCD and may not be sufficient to sustain thegrowth of bacteria. Since a similar acetoin catabolic pathwaymay be adopted from A. eutrophus, an acetoin-dependentacetaldehyde dehydrogenase is probably also required for thegrowth ofK pneumoniae CG43 on acetoin. Two closely linkedORFs with unknown biochemical functions which are requiredfor the utilization of acetoin in B. subtilis have recently beenreported (11). No homology between those two gene productsand the acoABCD products was observed. The likelihood ofthe presence of similar genes in K pneumoniae involved inacetoin catabolism is still to be determined.The aco operon ofK pneumoniae CG43 is highly active in E.
coli BL21, even in the absence of an inducer. We believe thatthe expression of the K pneumoniae aco operon contained inpHP713 is under the control of its own potent promoter. Incontrast to the or54-dependent transcription suggested for A.eutrophus acoXABC (32), the transcription start site of Kpneumoniae acoABCD is preceded by a U70-type promoter. It islikely that the K pneumoniae acoABCD operon resembles theP. putida BCODH operon, which does not require the rpoNsigma factor for expression (23).Although many of its features are homologous to those of
the A. eutrophus aco operon, acoX could not be found in theKpneumoniae acoABCD operon. On the other hand, we foundan ORF (acoK; Fig. 1) of at least 475 amino acids in lengthrunning in the opposite direction of the acoABCD operon.Thus, acoK is divergently transcribed from a promoter whichprobably overlaps the promoter of the acoABCD operon.Analysis of the partial AcoK sequence revealed a significanthomology with the regulatory protein MalT of the E. coli malregulon (37). Our preliminary result has shown that the acoKgene is essential for the acetoin-induced expression of theacoABCD operon and that deletion of the gene results in aconstitutive expression. It is likely that the AcoK protein playsa regulatory role in the acetoin-inducible expression of the Kpneumoniae aco operon.
ACKNOWLEDGMENTS
We thank A. Herp, T. Liu, J. Ou, and T. Wang for their criticalcomments on the manuscript.The study was supported in part by grants from Chang Gung College
of Medicine and Technology (CMRP 391 to H.-L.P. and CMRP 385 toH.-Y.C.) and from the National Science Council, Taiwan (NSC81-0412-B182-530 to H.-L.P. and NSC 82-0412-B182-093 to H.-Y.C.).
REFERENCES
1. Allen, A. G., and R. N. Perham. 1989. Reductive acetylation oftandemly repeated lipoyl domains in the pyruvate dehydrogenasemultienzyme complex of Escherichia coli is random order. J. Mol.Biol. 208:623-633.
2. Blomqvist, K., M. Nikkola, P. Lehtovaara, M. L. Suihko, U.Airaksinen, K. B. Straby, J. K. C. Knowles, and M. E. Penttila.1993. Characterization of the genes of the 2,3-butanediol operonsfrom Klebsiella tettigena and Enterobacter aerogenes. J. Bacteriol.175:1392-1404.
3. Burns, G., T. Brown, K. Hatter, J. M. Idriss, and J. R. Sokatch.1988. Similarity of the El subunits of branched-chain-oxoaciddehydrogenase from Pseudomonas putida to the correspondingsubunits of mammalian branched-chain-oxoacid and pyruvatedehydrogenase. Eur. J. Biochem. 176:311-317.
4. Burns, G., T. Brown, K. Hatter, and J. R. Sokatch. 1988. Com-parison of the amino acid sequences of the transacylase compo-nents of branched chain oxoacid dehydrogenase of Pseudomonasputida, and the pyruvate and 2-oxoglutarate dehydrogenase ofEscherichia coli. Eur. J. Biochem. 176:311-317.
VOL. 176, 1994
on May 21, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

3534 DENG ET AL.
5. Burns, G., T. Brown, K. Hatter, and J. R. Sokatch. 1989. Sequenceanalysis of the lpdV gene for lipoamide dehydrogenase ofbranched chain oxoacid dehydrogenase of Pseudomonas putida.Eur. J. Biochem. 179:61-69.
6. Caswell, R. C., P. Gacesa, K. E. Lutrell, and A. J. Weightman.1989. Molecular cloning and heterologous expression of a Kleb-siella pneumoniae gene encoding alginate lyase. Gene 75:127-134.
7. Cohen, G., M. Yanko, M. Mislovati, A. Argaman, R. Schreiber, Y.Av-Gay, and Y. Aharonowitz. 1993. Thioredoxin-thioredoxin re-ductase system of Streptomyces clavuligerus: sequence, expression,and organization of the genes. J. Bacteriol. 175:5159-5167.
8. Davie, J. R., R. M. Wynn, R. P. Cox, and D. T. Chuang. 1992.Expression and assembly of a functional El component (a232) ofmammalian branched-chain ox-ketoacid dehydrogenase complex inEscherichia coli. J. Biol. Chem. 267:16601-16606.
9. Dolin, M. I. 1955. Diacetyl oxidation by Streptococcus faecalis, alipoic acid-dependent reaction. J. Bacteriol. 69:51-58.
10. Frund, C., H. Priefert, A. Steinbuchel, and H. G. Schlegel. 1989.Biochemical and genetic analysis of acetoin catabolism in Alcali-genes eutrophus. J. Bacteriol. 171:6539-6548.
11. Grundy, F. J., D. A. Waters, T. Y. Takova, and T. M. Henkin. 1993.Identification of genes involved in utilization of acetate andacetoin in Bacillus subtilis. Mol. Microbiol. 10:259-271.
12. Hawkins, C. F., A. Borges, and R. N. Perham. 1989. A commonstructural motif in thiamine pyrophosphate-binding enzymes.FEBS Lett. 255:77-82.
13. Hawkins, C. F., A. Borges, and R. N. Perham. 1990. Cloning andsequence analysis of the genes encoding the cx and ,B subunits ofthe El component of the pyruvate dehydrogenase multienzymecomplex of Bacillus stearothermophilus. Eur. J. Biochem. 191:337-346.
14. Hemila, H. 1991. Lipoamide dehydrogenase of Staphylococcusaureus: nucleotide sequence and sequence analysis. Biochim.Biophys. Acta 1129:119-123.
15. Hemila, H., A. Palva, L. Paulin, S. Arvidson, and I. Palva. 1990.Secretory S complex of Bacillus subtilis: sequence analysis andidentity to pyruvate dehydrogenase. J. Bacteriol. 172:5052-5063.
16. Johansen, L., K. Bryn, and F. C. St0rmer. 1975. Physiological andbiochemical role of the butanediol pathway in Aerobacter (Entero-bacter) aerogenes. J. Bacteriol. 123:1124-1130.
17. Juni, E., and G. A. Heym. 1956. A cyclic pathway for the bacterialdissimilation of 2,3-butanediol, acetylmethylcarbinol and diacetyl.II. The synthesis of diacetylmethylcarbinol from diacetyl, a newdiphosphothiamin catalyzed reaction. J. Bacteriol. 72:746-753.
18. Krampitz, L. 0. 1955. Preparation and determination of acetoin,diacetyl, and acetolactate. Methods Enzymol. 13:277-283.
19. Kruger, N., and A. Steinbuchel. 1992. Identification of acoR, aregulatory gene for the expression of genes essential for acetoincatabolism in Alcaligenes eutrophus H16. J. Bacteriol. 174:4391-4400.
20. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
21. Lopez, J. M., B. Thoms, and P. Fortnagel. 1973. Mutants ofBacillus subtilis blocked in acetoin reductase. Eur. J. Biochem.40:479-483.
22. Lopez, J. M., B. Thoms, and H. Rehbein. 1975. Acetoin degrada-tion in Bacillus subtilis by direct oxidative cleavage. Eur. J.Biochem. 57:425-430.
23. Madhusudhan, K. T., G. Huang, G. Burns, and J. R. Sokatch.1990. Transcriptional analysis of the promoter region of thePseudomonas putida branched-chain keto acid dehydrogenaseoperon. J. Bacteriol. 172:5655-5663.
24. Miller, J. H. 1972. Experiments in molecular genetics, p. 352-355.Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y.
25. Oppermann, F. B., B. Schmidt, and A. Steinbuchel. 1991. Purifi-cation and characterization of acetoin:2,6-dichlorophenolindophe-nol oxidoreductase, dihydrolipoamide dehydrogenase, and dihy-drolipoamide acetyltransferase of the Pelobacter carbinolicusacetoin dehydrogenase enzyme system. J. Bacteriol. 173:757-767.
26. Oppermann, F. B., and A. Steinbuchel. 1994. Identification andmolecular characterization of the aco genes encoding the Pe-
lobacter carbinolicus acetoin dehydrogenase enzyme system. J.Bacteriol. 176:469-485.
27. Packman, L. C., G. Hale, and R. N. Perham. 1984. Repeatingfunctional domains in the pyruvate dehydrogenase multienzymecomplex of Escherichia coli. EMBO J. 3:1315-1319.
28. Palmer, J. A., K. T. Madhusudhan, K. Hatter, and J. R. Sokatch.1991. Cloning, sequence and transcriptional analysis of the struc-tural gene for LPD-3, the third lipoamide dehydrogenase ofPseudomonas putida. Eur. J. Biochem. 202:231-240.
29. Peng, H.-L., P.-Y. Wang, C.-M. Wu, D.-C. Hwang, and H.-Y.Chang. 1992. Cloning, sequencing and heterologous expression ofa Klebsiella pneumoniae gene encoding an FAD-independentacetolactate synthase. Gene 117:125-130.
30. Peng, H.-L., P.-Y. Wang, J.-L. Wu, C.-T. Chiu, and H.-Y. Chang.1991. Molecular epidemiology of Klebsiella pneumoniae. Chin. J.Microbiol. Immunol. (Taipei) 24:264-271.
31. Perham, R. N. 1991. Domains, motifs, and linkers in 2-oxo aciddehydrogenase multienzyme complexes: a paradigm in the designof a multifunctional protein. Biochemistry 30:8501-8511.
32. Priefert, H., S. Hein, N. Kruger, K. Zeh, B. Schmidt, and A.Steinbuchel. 1991. Identification and molecular characterizationof the Alcaligenes eutrophus H16 aco operon genes involved inacetoin catabolism. J. Bacteriol. 173:4056-4071.
33. Priefert, H., N. Kruger, D. Jendrossek, B. Schmidt, and A.Steinbuchel. 1992. Identification and molecular characterizationof the gene coding for acetaldehyde dehydrogenase II (acoD) ofAlcaligenes eutrophus. J. Bacteriol. 174:899-907.
34. Reed, L., and C. R. Willms. 1966. Purification and resolution of thepyruvate dehydrogenase complex (Escherichia coli). Methods En-zymol. 9:247-265.
35. Reed, L. J., M. Koike, M. E. Lewitch, and F. R. Leach. 1958.Studies on the nature and reactions of protein-bound lipoic acid.J. Biol. Chem. 232:143-158.
36. Repetto, B., and A. Tzagoloff. 1990. Structure and regulation ofKGD2, the structural gene for yeast dihydrolipoyl transsuccinylase.Mol. Cell. Biol. 10:4221-4232.
37. Richet, E., and 0. Raibaud. 1989. MalT, the regulatory protein ofthe Escherichia coli maltose system, is an ATP-dependent tran-scriptional activator. EMBO J. 8:981-987.
38. Rosenberg, M., and D. Court. 1979. Regulatory sequences in-volved in the promotion and termination of RNA transcription.Annu. Rev. Genet. 13:319-353.
39. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecularcloning: a laboratory manual, 2nd ed. Cold Spring Harbor Labo-ratory, Cold Spring Harbor, N.Y.
40. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequencingwith chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
41. Skyes, P. J., G. Burns, J. Menard, K. Hatter, and J. R. Sokatch.1987. Molecular cloning of genes encoding branched-chain ketoacid dehydrogenase of Pseudomonas putida. J. Bacteriol. 169:1619-1625.
42. Spencer, M. E., M. G. Darlison, P. E. Stephens, I. K. Duckenfield,and J. R. Guest. 1984. Nucleotide sequence of the sucB geneencoding the dihydrolipoamide succinyltransferase of Escherichiacoli and homology with the corresponding acetyltransferase. Eur.J. Biochem. 141:361-373.
43. Stephens, P. E., M. G. Darlison, H. M. Lewis, and J. R. Guest.1983. The pyruvate dehydrogenase complex of Eschenchia coliK12: nucleotide sequence encoding the pyruvate dehydrogenasecomponent. Eur. J. Biochem. 133:155-162.
44. Stephens, P. E., M. G. Darlison, H. M. Lewis, and J. R. Guest.1983. The pyruvate dehydrogenase complex of Escherichia coliK12: nucleotide sequence encoding the dihydrolipoamide acetyl-transferase component. Eur. J. Biochem. 133:481-489.
45. Stephens, P. E., H. M. Lewis, M. G. Darlison, and J. R. Guest.1983. Nucleotide sequence of the lipoamide dehydrogenase geneof Escherichia coli K12. Eur. J. Biochem. 135:519-527.
46. St0rmer, F. C. 1975. 2,3-Butanediol biosynthetic system in Aero-bacter aerogenes. Methods Enzymol. 41:518-533.
47. Studier, F. W., and B. A. Moffatt. 1986. Use of bacteriophage T7RNA polymerase to detect selective high-level expression ofcloned genes. J. Mol. Biol. 189:113-130.
J. BAC-FERIOL.
on May 21, 2018 by guest
http://jb.asm.org/
Dow
nloaded from

acoABCD OPERON OF K PNEUMONIE 3535
48. Summers, W. C. 1970. A simple method for extraction of RNAfrom Escherichia coli utilizing diethylpyrocarbonate. Anal. Bio-chem. 33:459-463.
49. Westphal, A. H., and A. de Kolk 1990. The 2-oxoglutarate dehy-drogenase complex from Azobacter vinelandii. Molecular cloningand sequence analysis of the gene encoding the succinyltransferase
component. Eur. J. Biochem. 187:235-239.50. Wynn, R. M., J. L. Chuang, J. R. Davie, C. W. Fisher, M. A. Hale,
R. P. Cox, and D. T. Chuang. 1992. Cloning and expression inEscherichia coli of mature E1 subunit of bovine mitochondrialbranched-chain a-ketoacid dehydrogenase complex. Mapping ofthe E1l-binding region on E2. J. Biol. Chem. 267:1881-1887.
VOL. 176, 1994
on May 21, 2018 by guest
http://jb.asm.org/
Dow
nloaded from