3. orthodromic i.p.s.p.s in normal conditions appear to be complex
TRANSCRIPT

J. Physiol. (1982), 328, pp. 105-123 105With 11 text-figures
Printed in Great Britain
FEED-FORWARD DENDRITIC INHIBITION IN RAT HIPPOCAMPALPYRAMIDAL CELLS STUDIED IN VITRO
BY B. E. ALGER* AND R. A. NICOLLFrom the Departments of Pharmacology and Physiology, University of California,
San Francisco, CA 94143, U.S.A.
(Received 6 July 1981)
SUMMARY
1. Intracellular recordings from CAI pyramidal cells in the rat hippocampal slicepreparation have been used to study the neuronal pathways involved in hippocampalsynaptic inhibition.
2. When direct comparisons are made in a single pyramidal cell, orthodromicstimulation delivered to stratum (s.) radiatum in normal recording conditions is foundto be more effective than antidromic stimulation in producing inhibitory post-synapticpotentials (i.p.s.p.s).
3. Orthodromic i.p.s.p.s in normal conditions appear to be complex, multiphasicevents, whereas antidromic i.p.s.p.s are relatively simple. The orthodromic i.p.s.p.involves both a GABA-mediated dendritic component and a non-GABA-mediatedcomponent neither of which is activated by antidromic stimulation.
4. Barbiturates induce a late depolarizing phase of the orthodromic response, a'depolarizing i.p.s.p.', which is mediated by GABA. The depolarizing i.p.s.p. is notproduced by antidromic stimulation.
5. Injections of tetrodotoxin and bicuculline methiodide localized to either somaticor apical dendritic regions reveal that the depolarizing i.p.s.p. is produced by GABAreleased from neuronal elements in the dendritic field which acts on pyramidal celldendrites.
6. The depolarizing i.p.s.p. is strongly temperature-dependent and increases inamplitude and duration progressively as slices are cooled from 37 to 22 °C.
7. Depolarizing i.p.s.p.s can be produced by orthodromic stimulation in s. oriensas well as in s. radiatum. In each case the depolarizing i.p.s.p.s appear localized tothe dendrites in the field stimulated.
8. We conclude that the depolarizing i.p.s.p. evident in the presence of barbituratesis caused by the same synaptic release ofGABA which in normal conditions produceshyperpolarizing dendritic i.p.s.p.s.
9. Numerous comparisons between orthodromic and antidromic stimulation indi-cate that dendritic i.p.s.p.s are activated by feed-forward pathways.
* Present address: Department of Physiology, School of Medicine, University of Maryland,Baltimore, MD 21201, U.S.A.

B. E. ALGER AND R. A. NICOLL
INTRODUCTION
The work of Kandel, Spencer & Brinley (1961) and of Andersen, Eccles & L0yning(1964a, 1964b) showed that the prominent inhibitory post-synaptic potential (i.p.s.p.)in hippocampal pyramidal cells can largely be accounted for by the activation of arecurrent inhibitory system which terminates on the pyramidal cell soma. While thishypothesis has generally withstood the test of time, evidence suggests that it is notsufficient to explain every aspect of hippocampal inhibition. Purpura and hiscoworkers (Purpura & Pappas, 1968; Purpura, Prelevic & Santini, 1968; Schwartz,Pappas & Purpura, 1968) noted that, in neonatal animals, there must be some othersystem besides the recurrent somatic one, since it is possible to record large i.p.s.p.sin these animals prior to development of the axosomatic terminal plexus whichdominates cells in the adult animal (see also Schwartzkroin & Altschuler, 1977). Inadult animals it is possible to record i.p.s.p.s intradendritically (Wong & Prince, 1979;Wong, Prince & Basbaum, 1979). Hippocampal i.p.s.p.s are thought to be mediated byy-aminobutyric acid (GABA) (Curtis, Felix & McLennan, 1970; see Storm-Mathisen,1977, for review) and a variety of biochemical, immuno-histochemical and anatomicalevidence would be consistent with a hypothesis of GABA-mediated dendriticinhibition in the hippocampus (see Discussion). The possibility of a feed-forwardinhibitory system has been suggested by many investigators (Andersen, Gross, L0mo& Sveen, 1969; Lynch, Jensen, McGaugh, Davila & Oliver, 1981; Lee, Dunwiddie &Hoffer, 1980; Dingledine & Gjerstad, 1980).
In the experiments presented in this paper we have used pharmacological andphysiological techniques to examine the neuronal circuitry involved in the productionof i.p.s.p.s in the hippocampal slice. During this investigation we observed thatsuperfusion with pentobarbitone, an anaesthetic drug known to enhance hippocampali.p.s.p.s (Nicoll, Eccles, Oshima & Rubia, 1975) results in a biphasic (hyperpolarizing-depolarizing) i.p.s.p. in response to orthodromic synaptic stimulation (cf. Thalmann,Peck & Ayala, 1981). Because of the very unusual nature of this response we haveexamined it in some detail and will present evidence that the depolarizing aspect ofthe pentobarbitone-induced response is a manifestation of feed-forward dendriticinhibition. The companion paper describes a series of ionophoretic experiments withGABA and considers the nature of the depolarizing synaptic response (Alger & Nicoll,1982). A preliminary report of this work has appeared (Alger & Nicoll, 1979).
METHODS
The rat hippocampal slice preparation was used in these experiments. The modified experimentalchamber which we now use (Nicoll & Alger, 1981 b) incorporates a Peltier heating-coolingthermo-electric unit (Cambion Inc.) for temperature control. Following standard procedures fordissecting the hippocampus and cutting slices, one slice is submerged in a small, constantly perfusedwell the temperature of which is controlled by the thermo-electric unit. Other slices are maintaineduntil needed in a separate incubation chamber at room temperature at the interface of thephysiological medium and 95% 02-5%C02. Our standard bathing medium consists of(mM): NaCl,116-4; KCl, 5-4; MgSO4, 1-3; CaCl2, 2-5; NaH2PO4, 0O92; NaHCO3, 26-2; glucose, 11. In mediacontaining low concentrations of chloride, sodium isethionate replaced NaCl in equimolar amounts.In low potassium media NaCl replaced KC1. Experiments on pentobarbitone-induced responses weredone in the presence ofeither 100 or 125 /sM-sodiumpentobarbitone (Sigma). Phenobarbitone (Sigma)was bath-applied at concentrations of 100 ,eM-1 0 mM and diazepam (Hoffman-LaRoche) at
106

HIPPOCAMPAL DENDRITIC INHIBITION
0-1-10 /M. Switching between media is controlled by a latching solenoid valve (General Valve Inc).In assessing the effects of bath-applied drugs we have used a given cell as its own control, firstrecording responses in a control condition, and subsequently applying and reversing the effects ofan experimental treatment. Except where noted, all traces in a figure were obtained from the samecell. Temperature was monitored by a small thermistor within 1 mm of the slice and was usuallyheld at 30-32 TC, although in some experiments it was increased or decreased for short (< 20 min)periods of time.
Conventional intracellular recording techniques were used. Intracellular recording electrodes(80-120 MCI) pulled from 'omegadot' glass tubing were filled with 2M-potassium methylsulphate,unless otherwise stated. A bridge circuit was used for altering the membrane potential by passingcurrent through the balanced recording electrode. Field potentials were recorded with either thehigh impedance intracellular micropipette outside a cell, or with a low impedance (10MQ) glasspipette filled with 3M-NaCl. Field potential population spike amplitudes were determined as theamplitude of the negative portion of the triphasic field potential measured from the initial positivityto the depth of the negative trough.We used a hydraulic stepping microdrive (Kopf) equipped with a counter which permitted precise
determination of the depth of an impalement within a slice. This measurement was essential tocheck field potential responses recorded in the same location. Data were recorded on-line with a
pen recorder (Gould) or photographed from the face of an oscilloscope.The experiments in this and the following paper were performed on over 150 CAl pyramidal cells.
All cells had resting membrane potentials greater than -50 mV (range -50 to -73 mV). Actionpotentials ranged from 65 to 110 mV and input impedances from 25-80miQ. Membrane potentialswere stable from 30 min to 6 h.Throughout these experiments we have made extensive comparisons between 'orthodromic' and
'antidromic' activation of the CAI pyramidal cells. Orthodromic stimulation was delivered tostratum (s.) radiatum in most cases. Usually the orthodromic stimulating electrode was on thesubicular side of the recording electrode. Antidromic stimulation was always delivered to the outerportion of the alveus, also on the subicular side of the recording electrode. To produce a 'pure'antidromic input we occasionally placed lesions, made with a razor blade shard, across the widthof the s. oriens region, sparing only the outer two-thirds of the alveus (cf. Dingledine & Langmoen,1980). Stimulating the alveus then evoked antidromic responses with no sign of contamination bystimulation of the orthodromic fibres in s. oriens.A central technique in these experiments was selective ionophoretic application of drugs to
different regions of a pyramidal cell. The drugs used for ionophoresis were prepared as follows:GABA (Sigma),
1 M
in distilled water, pH adjusted to 40 with HCI; bicuculline methiodide, 10mmin 0-2M-NaCI; tetrodotoxin (Sigma), 100FM in 0'2M-NaCl. The methiodide derivative was preparedfrom bicuculline (Sigma) according to the method of Pong & Graham (1973). A WPI 160ionophoresis programmer was used for drug ionophoresis. Somatic injections occurred in s.pyramidale with the ionophoretic pipette positioned within 50 Fm of the recording micro-electrode.In some cases double-barrelled pipettes were used for the somatic injections of bicucullinemethiodide or tetrodotoxin. One barrel was filled with GABA and the hyperpolarizing responses
(cf. Alger & Nicoll, 1982) were used to guide the electrode to the soma of the impaled cell so thatthe effects of the other drug were as localized as possible. Applications of drugs to dendrites occurreda minimum of 150 Fm from the edge of s. pyramidale in s. radiatum. For optimal placement ofbicuculline methiodide-and tetrodotoxin-containing pipettes, field potentials were recorded fromthe ionophoretic pipette and it was positioned in the region which gave the largest potential inresponse to orthodromic stimulation. In testing for the selectivity of a drug effect on antidromicor orthodromic responses we alternated antidromic and orthodromic stimulation. Only when a drugeffect on the response stimulated by one input was bracketed by unaltered responses to the otherinput was the application considered to be localized.
RESULTS
Antidromic and orthodromic activation of i.p.8.p.8 in standard mediaA clue to the existence of an inhibitory pathway besides the recurrent somatic one
can be obtained by directly comparing the ability of orthodromic and antidromicstimuli to produce iLp.s.p.s in a pyramidal cell. Making a valid comparison requires
107
B. E. ALGER AND R. A. NICOLL
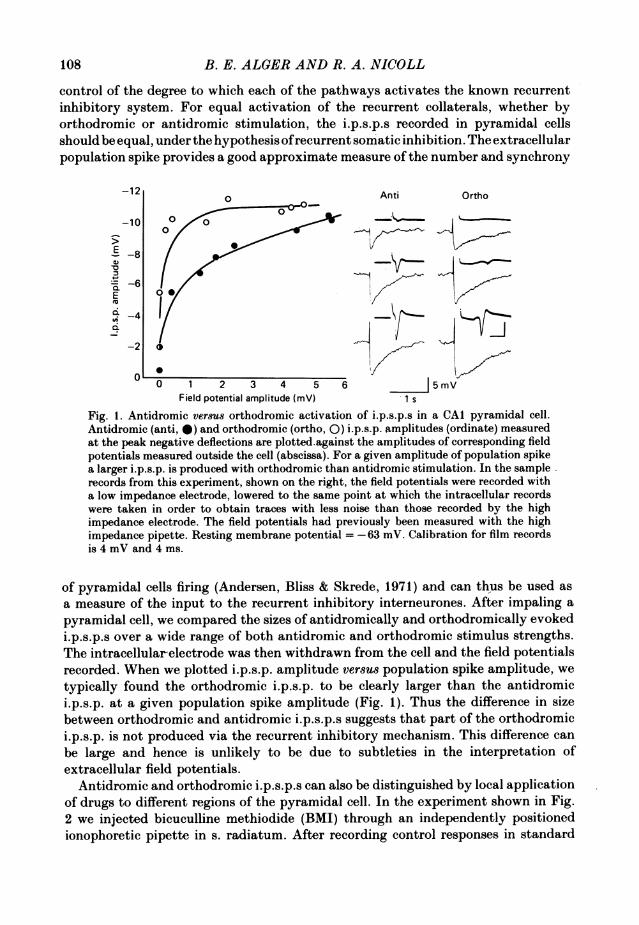
control of the degree to which each of the pathways activates the known recurrentinhibitory system. For equal activation of the recurrent collaterals, whether byorthodromic or antidromic stimulation, the i.p.s.p.s recorded in pyramidal cellsshould be equal, under the hypothesis ofrecurrent somatic inhibition. The extracellularpopulation spike provides a good approximate measure of the number and synchrony
-12 O Anti Ortho
E-80 O B _0.-o0
0
E _8 ° J _
:-4 0/
C 1 2 3 4 5 6 j5mVField potential amplitude (mV) 1 s
Fig. 1. Antidromic versus orthodromic activation of i.p.s.p.s in a CAI pyramidal cell.Antidromic (anti, 0) and orthodromic (ortho, 0) i.p.s.p. amplitudes (ordinate) measuredat the peak negative deflections are plotted.against the amplitudes of corresponding fieldpotentials measured outside the cell (abscissa). For a given amplitude of population spikea larger i.p.s.p. is produced with orthodromic than antidromic stimulation. In the samplerecords from this experiment, shown on the right, the field potentials were recorded witha low impedance electrode, lowered to the same point at which the intracellular recordswere taken in order to obtain traces with less noise than those recorded by the highimpedance electrode. The field potentials had previously been measured with the highimpedance pipette. Resting membrane potential =-63 mV. Calibration for film recordsis 4 mV and 4 ms.
of pyramidal cells firing (Andersen, Bliss & Skrede, 1971) and can thus be used asa measure of the input to the recurrent inhibitory interneurones. After impaling apyramidal cell, we compared the sizes of antidromically and orthodromically evokedi.p.s.p.s over a wide range of both antidromic and orthodromic stimulus strengths.The intracellular-electrode was then withdrawn from the cell and the field potentialsrecorded. When we plotted i.p.s.p. amplitude versus population spike amplitude, wetypically found the orthodromic i.p.s.p. to be clearly larger than the antidromici.p.s.p. at a given population spike amplitude (Fig. 1). Thus the difference in sizebetween orthodromic and antidromic i.p.s.p.s suggests that part of the orthodromici.p.s.p. is not produced via the recurrent inhibitory mechanism. This difference canbe large and hence is unlikely to be due to subtleties in the interpretation ofextracellular field potentials.
Antidromic and orthodromic i.p.s.p.s can also be distinguished by local applicationof drugs to different regions of the pyramidal cell. In the experiment shown in Fig.2 we injected bicuculline methiodide (BMI) through an independently positionedionophoretic pipette in s. radiatum. After recording control responses in standard
108

HIPPOCAMPAL DENDRITIC INHIBITION
medium (left column) BMI was injected into the apical dendritic region. BMI quicklyreduced the early phase of the orthodromic i.p.s.p. without affecting the antidromici.p.s.p. Subsequently the antidromic i.p.s.p. was blocked, presumably due to diffusionof the BMI to s. pyramidale; however, the late hyperpolarizing potential (l.h.p.) ofthe orthodromic response was not blocked. We have recently studied the l.h.p. (Nicoll
Control BMI-dend Recovery
Ortho .._
400 msAnti
Fig. 2. Independent antagonism of orthodromic and antidromic i.p.s.p.s. A bicucullinemethiodide (BMI)-containing pipette was positioned in s. radiatum and the records in themiddle columns were obtained 30 s after starting to eject BMI (100 nA). BMI blocks theearly negative peak of the orthodromic response and the same stimulus now producesa secondary action potential (compare oscilloscope trace insets). The antidromici.p.s.p. is not affected at this point, however. The records labelled 'recovery' were obtained7 min after turning off the ionophoretic current. The control and experimental recordshave been overlapped and traced in the far right-hand column to facilitate comparison.Resting membrane potential = -53 mV. Calibration for all pen traces is 5 mV, 400 ms.Calibration for oscilloscope insets is 40 mV, 20 ms.
& Alger, 1981 a) and found that, unlike the early phase, it is not sensitive to changesin either intra- or extracellular chloride or to high doses of bath-applied GABAantagonists.
Figs. 1 and 2 illustrate that there are two components of the orthodromichyperpolarization which are not activated by the recurrent pathway. One componentis a non-GABA-mediated potential. The other component however is blocked byGABA antagonists and appears to be produced in the apical dendritic region.
Effects of pentobarbitone, phenobarbitone and diazepamThe late hyperpolarizing potential clearly overlaps and obscures the GABA-
mediated orthodromic i.p.s.p. Thus to study the i.p.s.p. in more detail we appliedpentobarbitone, an anaesthetic agent which enhances the action of GABA-mediatedi.p.s.p.s (Nicoll et al. 1975). Surprisingly, we found that with orthodromic activationof pyramidal cells in the presence of 100,uM-pentobarbitone a biphasic, hyper-polarizing-depolarizing i.p.s.p. replaced the hyperpolarizing responses (Fig. 3). Giventhe observation that dendritic application of GABA also depolarizes pyramidal celldendrites (Alger & Nicoll, 1982), it appeared likely that this pentobarbitone-induceddepolarizing potential might provide a method for studying GABA-mediated dendritici.p.s.p.s. Therefore we have made a detailed characterization of the inhibitorypotentials in the presence of pentobarbitone.The depolarization induced in the presence of pentobarbitone is associated with
a large conductance increase and is usually inhibitory, blocking both direct and
109
B. E. ALGER AND R. A. NICOLL
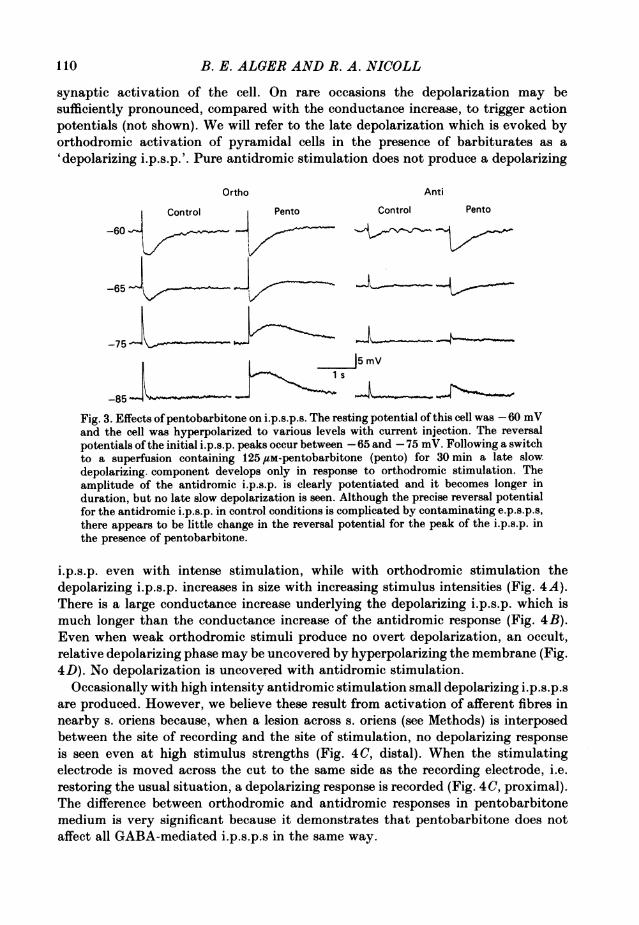
synaptic activation of the cell. On rare occasions the depolarization may besufficiently pronounced, compared with the conductance increase, to trigger actionpotentials (not shown). We will refer to the late depolarization which is evoked byorthodromic activation of pyramidal cells in the presence of barbiturates as a'depolarizing i.p.s.p.'. Pure antidromic stimulation does not produce a depolarizing
Ortho Anti
Control Pento Control Pento
-60 Ct Pento
-65 I
-75
J 5 mV
-85
Fig. 3. Effects ofpentobarbitone on i.p.s.p.s. The resting potential of this cell was -60 mVand the cell was hyperpolarized to various levels with current injection. The reversalpotentials ofthe initial i.p.s.p. peaks occur between -65 and -75 mV. Following a switchto a superfusion containing 125 /tM-pentobarbitone (pento) for 30 min a late slowdepolarizing. component develops only in response to orthodromic stimulation. Theamplitude of the antidromic i.p.s.p. is clearly potentiated and it becomes longer induration, but no late slow depolarization is seen. Although the precise reversal potentialfor the antidromic i.p.s.p. in control conditions is complicated by contaminating e.p.s.p.s,there appears to be little change in the reversal potential for the peak of the i.p.s.p. inthe presence of pentobarbitone.
i.p.s.p. even with intense stimulation, while with orthodromic stimulation thedepolarizing i.p.s.p. increases in size with increasing stimulus intensities (Fig. 4A).There is a large conductance increase underlying the depolarizing i.p.s.p. which ismuch longer than the conductance increase of the antidromic response (Fig. 4B).Even when weak orthodromic stimuli produce no overt depolarization, an occult,relative depolarizing phase may be uncovered by hyperpolarizing the membrane (Fig.4D). No depolarization is uncovered with antidromic stimulation.
Occasionally with high intensity antidromic stimulation small depolarizing i.p.s.p.sare produced. However, we believe these result from activation of afferent fibres innearby s. oriens because, when a lesion across s. oriens (see Methods) is interposedbetween the site of recording and the site of stimulation, no depolarizing responseis seen even at high stimulus strengths (Fig. 4C, distal). When the stimulatingelectrode is moved across the cut to the same side as the recording electrode, i.e.restoring the usual situation, a depolarizing response is recorded (Fig. 4C, proximal).The difference between orthodromic and antidromic responses in pentobarbitonemedium is very significant because it demonstrates that pentobarbitone does notaffect all GABA-mediated i.p.s.p.s in the same way.
110

HIPPOCAMPAL DENDRITIC INHIBITION
We have also tested phenobarbitone and diazepam, two other drugs which enhanceGABA-mediated inhibition, on eight cells. Phenobarbitone produces depolarizingi.p.s.p.s in response to orthodromic stimulation, although it requires five to ten timeshigher concentrations than those of pentobarbitone to produce a similar response.A 1 0 mm concentration of phenobarbitone is approximately as effective as 100 /LM-
A Anti Ortho B ASub J___
C; Distal Proximal
j mV2 s
4x D Ortho Anti I10mV
Fig. 4. Comparison of antidromic and orthodromic responses in pentobarbitone. In A aseries of stimuli of increasing strengths from subthreshold (sub) to multiples of thresholdwas delivered to the alveus (anti) or to s. radiatum (ortho) in pentobarbitone (125 /M)A lesion had been placed in s. oriens (see Methods). With weak stimuli both i.p.s.p.sappeared similar; however with increasing stimulus strengths a late depolarizing i.p.s.p.develops in response to ortho but not anti stimulation. Resting membrane potential= -62 mV. Responses in B are from the same cell as in A at 4 x threshold. Current pulses,40 ms in duration at 4 Hz, were passed across the membrane to monitor input impedance.A larger and much longer conductance increase accompanies the ortho than the antiresponse. C, from another cell, demonstrates that a high intensity 'pure' antidromicstimulus, delivered to alvear fibres following a microlesion which cut orthodromic fibresin s. oriens, is not associated with a depolarizing i.p.s.p. (C, distal). When the stimulatingelectrode is moved across the cut to the same side as the recording micro-electrode thesame stimulus (proximal) produces a depolarizing i.p.s.p. following the hyperpolarizingi.p.s.p. This indicates that the orthodromic fibres in s. oriens are responsible for thedepolarizing i.p.s.p. (compare Fig. 10). Resting membrane potential = -58 mV. In D itcan be seen that, even when there is no overt depolarizing i.p.s.p. at the resting potential(left trace), a relative depolarization can be uncovered by hyperpolarizing the membrane.No depolarization is uncovered with antidromic stimulation. Resting membrane potential= -70 mV. Voltage calibration for A, B and C is 5 mV, time calibration for C is the sameas for B, 4 s.
pentobarbitone. At concentrations of 0'1-1.0 /M, diazepam produces no overtdepolarizations, although it usually increases the size of submaximal hyperpolarizingi.p.s.p.s (not shown). Occasionally slight relative depolarizations are seen when themembrane is hyperpolarized to the reversal potential of the hyperpolarizing i.p.s.p.At levels higher than 10/M, diazepam has a general depressant effect on neuronalactivity (cf. Lee, Dunwiddie & Hoffer, 1979).
Properties of the depolarizing i.p.s.p.The induction of the depolarizing i.p.s.p. by barbiturates suggests that the i.p.s.p.
is a GABA-mediated event. This is supported by the finding that it is entirely blocked
ill
B. E. ALGER AND R. A. NICOLL
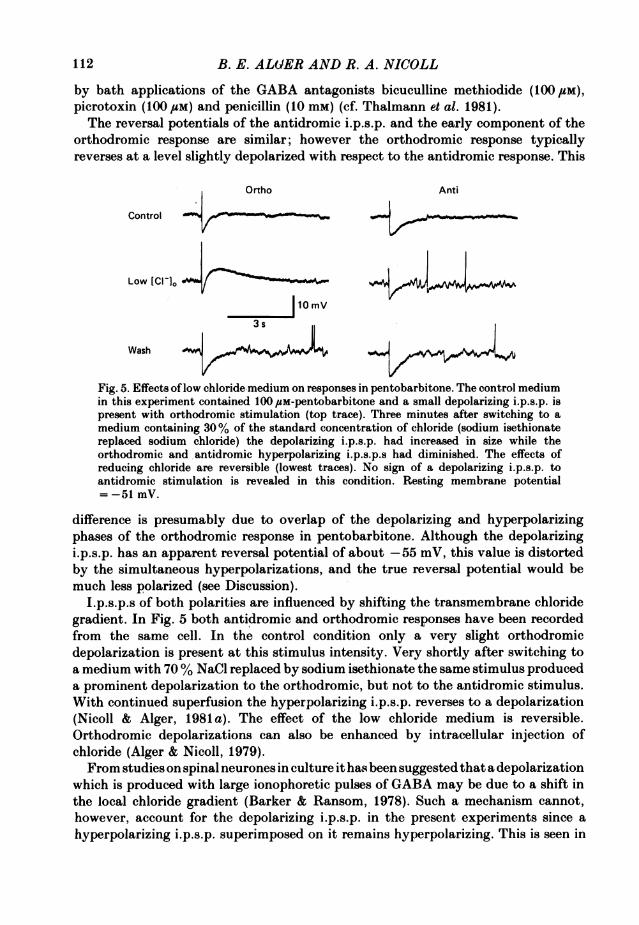
by bath applications of the GABA antagonists bicuculline methiodide (100 UM),picrotoxin (100 #sM) and penicillin (10 mM) (cf. Thalmann et al. 1981).The reversal potentials of the antidromic i.p.s.p. and the early component of the
orthodromic response are similar; however the orthodromic response typicallyreverses at a level slightly depolarized with respect to the antidromic response. This
Ortho Anti
Control
Low [Cl-l0
10mV
3 s
Wash J
Fig. 5. Effects oflow chloride medium on responses in pentobarbitone. The control mediumin this experiment contained 100 /SM-pentobarbitone and a small depolarizing i.p.s.p. ispresent with orthodromic stimulation (top trace). Three minutes after switching to amedium containing 30% of the standard concentration of chloride (sodium isethionatereplaced sodium chloride) the depolarizing i.p.s.p. had increased in size while theorthodromic and antidromic hyperpolarizing i.p.s.p.s had diminished. The effects ofreducing chloride are reversible (lowest traces). No sign of a depolarizing i.p.s.p. toantidromic stimulation is revealed in this condition. Resting membrane potential= -51 mV.
difference is presumably due to overlap of the depolarizing and hyperpolarizingphases of the orthodromic response in pentobarbitone. Although the depolarizingi.p.s.p. has an apparent reversal potential of about -55 mV, this value is distortedby the simultaneous hyperpolarizations, and the true reversal potential would bemuch less polarized (see Discussion).
I.p.s.p.s of both polarities are influenced by shifting the transmembrane chloridegradient. In Fig. 5 both antidromic and orthodromic responses have been recordedfrom the same cell. In the control condition only a very slight orthodromicdepolarization is present at this stimulus intensity. Very shortly after switching toa medium with 70% NaCl replaced by sodium isethionate the same stimulus produceda prominent depolarization to the orthodromic, but not to the antidromic stimulus.With continued superfusion the hyperpolarizing i.p.s.p. reverses to a depolarization(Nicoll & Alger, 1981 a). The effect of the low chloride medium is reversible.Orthodromic depolarizations can also be enhanced by intracellular injection ofchloride (Alger & Nicoll, 1979).From studies on spinal neurones in culture it has been suggested that a depolarization
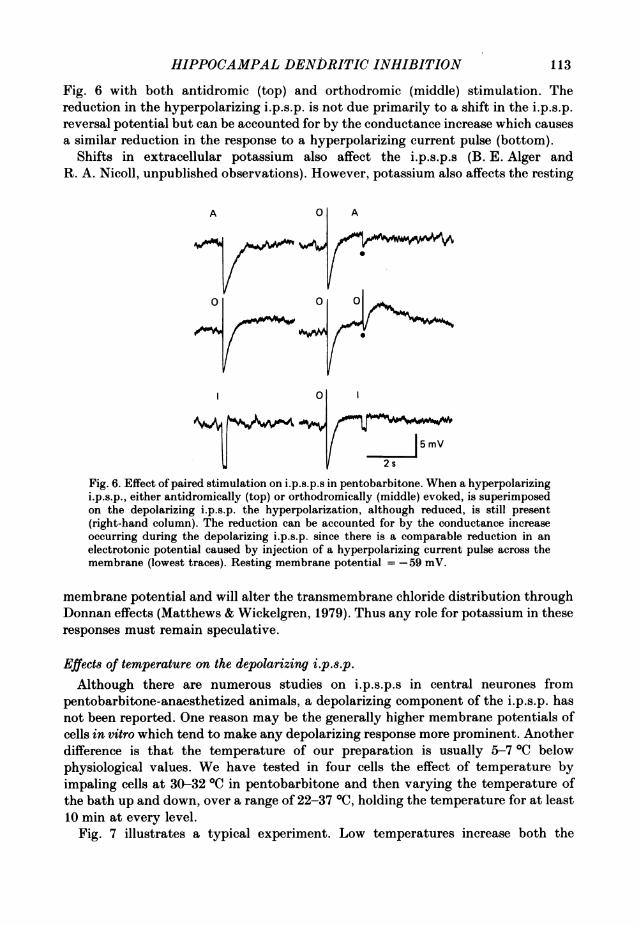
which is produced with large ionophoretic pulses of GABA may be due to a shift inthe local chloride gradient (Barker & Ransom, 1978). Such a mechanism cannot,however, account for the depolarizing i.p.s.p. in the present experiments since ahyperpolarizing i.p.s.p. superimposed on it remains hyperpolarizing. This is seen in
112
HIPPOCAMPAL DENDRITIC INHIBITION
Fig. 6 with both antidromic (top) and orthodromic (middle) stimulation. Thereduction in the hyperpolarizing i.p.s.p. is not due primarily to a shift in the i.p.s.p.reversal potential but can be accounted for by the conductance increase which causesa similar reduction in the response to a hyperpolarizing current pulse (bottom).
Shifts in extracellular potassium also affect the i.p.s.p.s (B. E. Alger andR. A. Nicoll, unpublished observations). However, potassium also affects the resting
A 0 A
0~~~~
| ~~~5mV
Fig. 6. Effect ofpaired stimulation on i.p.s.p.s in pentobarbitone. When a hyperpolarizingi.p.s.p., either antidromically (top) or orthodromically (middle) evoked, is superimposedon the depolarizing i.p.s.p. the hyperpolarization, although reduced, is still present(right-hand column). The reduction can be accounted for by the conductance increaseoccurring during the depolarizing i.p.s.p. since there is a comparable reduction in anelectrotonic potential caused by injection of a hyperpolarizing current pulse across themembrane (lowest traces). Resting membrane potential =-59 mV.
membrane potential and will alter the transmembrane chloride distribution throughDonnan effects (Matthews & Wickelgren, 1979). Thus any role for potassium in theseresponses must remain speculative.
Effect of temperature on the depolarizing i.p.s.p.Although there are numerous studies on i.p.s.p.s in central neurones from
pentobarbitone-anaesthetized animals, a depolarizing component of the i.p.s.p. hasnot been reported. One reason may be the generally higher membrane potentials ofcells in vitro which tend to make any depolarizing response more prominent. Anotherdifference is that the temperature of our preparation is usually 5-7 0C belowphysiological values. We have tested in four cells the effect of temperature byimpaling cells at 30-32 0C in pentobarbitone and then varying the temperature ofthe bath up and down, over a range of 22-37 0C, holding the temperature for at least10 min at every level.
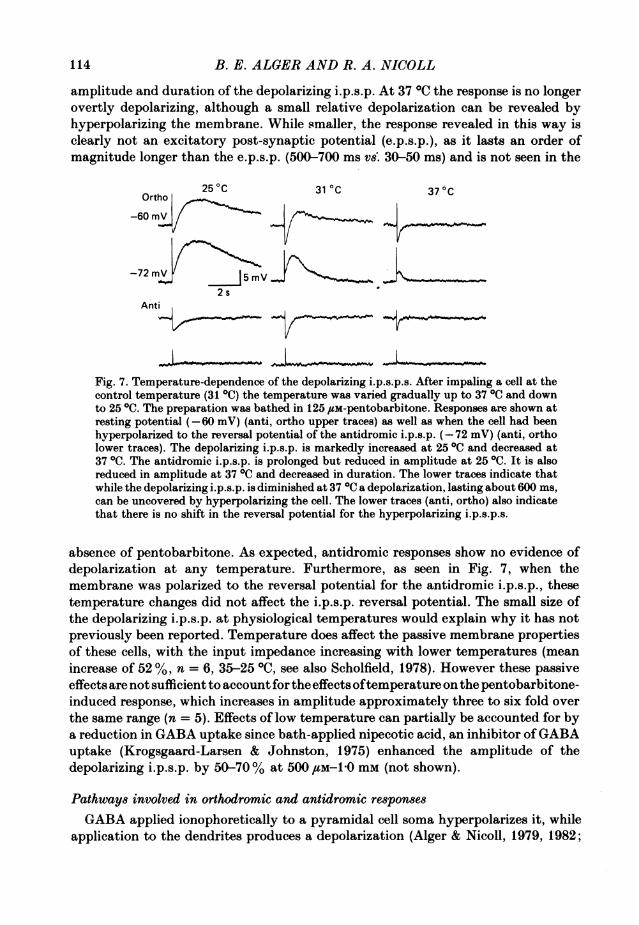
Fig. 7 illustrates a typical experiment. Low temperatures increase both the
113
B. E. ALGER AND R. A. NICOLL
amplitude and duration of the depolarizing i.p.s.p. At 37 IC the response is no longerovertly depolarizing, although a small relative depolarization can be revealed byhyperpolarizing the membrane. While smaller, the response revealed in this way isclearly not an excitatory post-synaptic potential (e.p.s.p.), as it lasts an order ofmagnitude longer than the e.p.s.p. (500-700 ms vs. 30-50 ms) and is not seen in the
Ortho 250C 31 0C 370C-60m~~rnV
-72mVjVmV2 s
Anti
Fig. 7. Temperature-dependence of the depolarizing i.p.s.p.s. After impaling a cell at thecontrol temperature (31 0C) the temperature was varied gradually up to 370 and downto 250C. The preparation was bathed in 125 /sM-pentobarbitone. Responses are shown atresting potential (-60 mV) (anti, ortho upper traces) as well as when the cell had beenhyperpolarized to the reversal potential of the antidromic i.p.s.p. (-72 mV) (anti, ortholower traces). The depolarizing i.p.s.p. is markedly increased at 250C and decreased at37 0C. The antidromic i.p.s.p. is prolonged but reduced in amplitude at 250C. It is alsoreduced in amplitude at 37 0C and decreased in duration. The lower traces indicate thatwhile the depolarizing i.p.s.p. is diminished at 37 0C a depolarization, lasting about 600 ms,can be uncovered by hyperpolarizing the cell. The lower traces (anti, ortho) also indicatethat there is no shift in the reversal potential for the hyperpolarizing i.p.s.p.s.
absence of pentobarbitone. As expected, antidromic responses show no evidence ofdepolarization at any temperature. Furthermore, as seen in Fig. 7, when themembrane was polarized to the reversal potential for the antidromic i.p.s.p., thesetemperature changes did not affect the i.p.s.p. reversal potential. The small size ofthe depolarizing i.p.s.p. at physiological temperatures would explain why it has notpreviously been reported. Temperature does affect the passive membrane propertiesof these cells, with the input impedance increasing with lower temperatures (meanincrease of 52 %, n = 6, 35-25 °C, see also Scholfield, 1978). However these passiveeffects are not sufficient to account for the effects oftemperature on the pentobarbitone-induced response, which increases in amplitude approximately three to six fold overthe same range (n = 5). Effects of low temperature can partially be accounted for bya reduction in GABA uptake since bath-applied nipecotic acid, an inhibitor ofGABAuptake (Krogsgaard-Larsen & Johnston, 1975) enhanced the amplitude of thedepolarizing i.p.s.p. by 50-70% at 500 /LM-1O mm (not shown).
Pathways involved in orthodromic and antidromic responsesGABA applied ionophoretically to a pyramidal cell soma hyperpolarizes it, while
application to the dendrites produces a depolarization (Alger & Nicoll, 1979, 1982;
114

HIPPOCAMPAL DENDRITIC INHIBITION
Andersen, Dingledine, Gjerstad, Langmoen & Mosfeldt Laursen, 1980; Thalmann etal. 1981) both in standard bathing media and in the presence of pentobarbitone. Wehypothesize that the depolarizing i.p.s.p. results from synaptically released GABAacting on dendritic receptors and have tested this hypothesis by using localizedinjections of BMI (twelve cells) and tetrodotoxin (eight cells) to either the somaticor dendritic regions. In Fig. 8A antidromic and orthodromic responses were evokedin the same cell before (left) and during (centre) an injection of BMI in s. radiatum.
A Control BMI-Dend Recovery
Ortho
j5mV3 s
Anti
B Control BM I -Soma Recovery
Ortho
Anti
Fig. 8. Localized injection of bicuculline methiodide. In A a BMI-containing pipette wasplaced in s. radiatum and the records in the middle columns were obtained 8 min afterswitching on the ionophoretic current (20 nA). At this time the depolarizing i.p.s.p. wasselectively blocked, leaving the fast orthodromic i.p.s.p. and the antidromic i.p.s.p. Therecovery was obtained 20 min after switching off the ionophoretic current. Restingmembrane potential = -60 mV. In B, performed on a different cell, the BMI-containingpipette was positioned in s. pyramidale. Injection in this region (300 nA for 4 min) resultedin a marked reduction of the antidromic i.p.s.p., a lesser reduction of the orthodromichyperpolarizing i.p.s.p. and no change in the depolarizing i.p.s.p. The recovery wasobtained 7 min after switching off the ionophoretic current. Resting membrane potential= -63 mV.
BMI eliminates the depolarizing i.p.s.p. before either of the hyperpolarizing i.p.s.p.sis affected. In Fig. 8B, an experiment done on another cell, a BMI injection localizedto the somatic region has the opposite effect, i.e. hyperpolarizing i.p.s.p.s areconsiderably reduced before the depolarizing i.p.s.p. is affected. These experimentsindicate that the orthodromic depolarizing response and the antidromic i.p.s.p. areproduced on separate portions of the pyramidal cell and specifically suggest that thedepolarizing i.p.s.p. is due to the activation of dendritic GABA receptors. GABAmight either diffuse to the dendrites from somatic release sites, or it could be releaseddirectly from presynaptic terminals onto dendritic receptors.To distinguish between these alternatives we used the same paradigm as shown
115

B. E. ALGER AND R. A. NICOLL
in Fig. 8, except that we injected TTX instead of BMI. Responses prior to TTXinjection are shown in the left column. In Fig. 9A the TTX injection was into s.radiatum. Results similar to those obtained with BMI were obtained with TTX.Localized TTX injections in s. radiatum block the depolarizing i.p.s.p. before theantidromic response, or the directly evoked somatic action potential (Alger & Nicoll,
A Control TTX-Dend Recovery
Ortho
J5mV
Anti
B Control TTX-Soma Recovery
Ortho
Anti
Fig. 9. Localized injections of tetrodotoxin. In A a TTX-containing pipette was positionedin s. radiatum. An injection ofTTX (400 nA for 3 min) selectively blocks the depolarizingi.p.s.p. The recovery was obtained 12 min after switching off the ionophoretic current. TheTTX-containing pipette was then positioned in s. pyramidale (B) and injection of TTXat this site (400 nA for 80 s) produces a relatively greater reduction in the hyperpolarizingresponses than in the depolarizing i.p.s.p. The recovery was obtained 10 min afterswitching off the ionophoretic current. Resting membrane potential = -54 mV.
1979), is affected. The converse is true when the TTX injection is into the s.pyramidale region (Fig. 9B). This experiment rules out the possibility that thedendritically acting GABA diffuses to the dendrites from somatic release points, sinceTTX does not blockGABA receptorsper se. The effects ofTTX must be due to blockingneuronal circuitry in s. radiatum which is involved in producing depolarizing i.p.s.p.s.
In the experiments illustrated in Figs. 8 and 9 the fast hyperpolarizing phase of the orthodromici.p.s.p. is not apparently depressed when the depolarizing i.p.s.p. is first blocked. This can beexplained in part by the differential sensitivity of depolarizations and hyperpolarizations toantagonists (Alger & Nicoll, 1982) and in part by the overlap of depolarizing and hyperpolarizingphases. Reduction of the depolarization will at first unmask part of the orthodromic hyperpolar-ization, partially counteracting a concomitant depression of the hyperpolarization. Finally, theorthodromic i.p.S.p. may have a somatic component due either to stimulation of the recurrent orfeed-forward pathways (see Discussion).
Apical vs. basal dendritic inputsAs mentioned earlier (see Fig. 4C), it is also possible to evoke a depolarizing i.p.s.p.
by stimulating fibres in s. oriens. This raises the questions ofwhether the depolarizing
116

HIPPOCAMPAL DENDRITIC INHIBITION
i.p.s.p. is produced on the apical dendrites only and, if not, whether there is only oneset of interneurones responsible for producing depolarizing i.p.s.p.s on differentdendritic trees. Application of GABA to the basal dendrites of CAI pyramidal cellsdepolarizes the cells (B. E. Alger & R. A. Nicoll, unpublished observations). Fig. 10illustrates evidence indicating that apical and basal dendrites receive separate GABA
Control BMI-s. rad. Recovery
S. rad.
J5mV
S. oriens
Fig. 10. Depolarizing i.p.s.p.s produced on different parts ofthe dendritic tree. Depolarizingi.p.s.p.s were produced in the same pyramidal cell by stimulating afferent fibres in s.radiatum (s. rad., upper) and s. oriens (lower). An injection of BMI via an ionophoreticpipette into s. radiatum (40 nA for 3 min) selectively reduces the depolarizing i.p.s.p. tos. radiatum stimulation. The recovery was obtained 8 min after stopping the ionophoreticcurrent. This indicates the depolarizing i.p.s.p. can be produced at spatially separatedregions of the pyramidal cell dendritic tree. Resting membrane potential = -60 mV.
inputs. In the control records (left) stimulation of both s. oriens and s. radiatumproduce depolarizing i.p.s.p.s in the same cell. Local application ofBMI in s. radiatumgreatly decreases the s. radiatum response without affecting the s. oriens response.It is likely that two separate interneuronal networks are involved because the s.radiatum-evoked response can be entirely abolished by infusion of BMI into thatregion (see Fig. 8). If the s. radiatum stimulus had activated a single set ofinterneurones which produced depolarizing i.p.s.p.s on both apical and basal dendrites,then BMI in the apical dendritic region should have spared the basal dendriticresponse.
DISCUSSION
We have presented a variety of evidence, both physiological and pharmacological,indicating that in addition to the well known recurrent somatic inhibitory system,there is also feed-forward dendritic inhibition in the hippocampus. There are twoissues involved: evidence for dendritic inhibition and evidence for feed-forwardinhibition.
Previous evidence for feed-forward inhibition includes the observations that weakorthodromic stimuli, not evoking obvious population spikes can none the less evokei.p.s.p.s (Andersen et al. 1969; Dingledine & Gjerstad, 1980) and can have aninhibitory effect in paired-pulse tests (Lynch et al. 1981). Presumed inhibitoryinterneurones can also be directly, i.e. not recurrently, activated (Andersen et al. 1969;Lee et al. 1980). Evidence in favour of dendritic inhibition is of several kinds. (1)Electrophysiological: interpretations offield potentials have suggested the occurrenceofi.p.s.p.s on the dendrites (Fugita & Sakata, 1962; Leung, 1979) as have intracellularrecordings, indicating that the early part of the i.p.s.p. reverses before the later partwith either KCl (Kandel et al. 1961; Eccles, Nicoll, Oshima & Rubia, 1977; Alger &
117

B. E. ALGER AND R. A. NICOLL
Nicoll, 1979) or current injection (Eccles et al. 1977; Dingledine & Langmoen, 1980).I.p.s.p.s have been recorded intradendritically, although it was not clear whetherthese originated in the dendrites (Wong & Prince, 1979; Wong, Prince & Basbaum,1979). (2) Biochemical: there are large amounts of GABA, GABA-transaminase andglutamic acid decarboxylase (GAD) in the dendritic regions (Storm-Mathisen, 1977).Binding of the GABA receptor agonist muscimol has also been demonstrated in s.
radiatum and s. oriens (Chan-Palay, 1979). (3) Immuno-histochemical: immuno-reactive GAD has been detected in cell bodies, dendrites and axon terminals in s.
radiatum and s. oriens. Some GAD-containing terminals were found in contact withpresumed pyramidal cell dendritic shafts (Barber & Saito, 1976; Ribak, Vaughn &Saito, 1978). (4) Anatomical: synaptic terminals containing flattenable vesicles havebeen found making symmetrical synaptic contacts on presumed pyramidal celldendrites (Gottlieb & Cowan, 1972). Purpura and his colleagues demonstratednumerous synapses on dendrites of neonatal animals, and recorded i.p.s.p.s intra-cellularly prior to the development of axosomatic synapses (Purpura & Pappas, 1968;Purpura et al. 1968; Schwartz et al. 1968). These developmental experiments havebeen extended by Schwartzkroin & Altschuler (1977). The present study providesfurther support for the hypothesis of feed-forward dendritic inhibition.
Since antidromic stimulation will selectively activate a recurrent collateral system,we have made extensive comparisons between antidromic and orthodromic responses.
We found that the recurrent system is not capable of accounting entirely fororthodromically evoked i.p.s.p.s. In standard medium orthodromic stimulation ismore effective than antidromic stimulation in eliciting i.p.s.p.s when the size of theinput to the recurrent inhibitory pathway is taken into account. Furthermore, instandard medium, the orthodromic i.p.s.p. is a complex multiphasic event (cf. Fugita,1979), while the antidromic i.p.s.p. is simple. In previous work we have shown thata late hyperpolarizing potential evoked orthodromically (l.h.p.) is not blocked by bathapplication of several GABA antagonists or by changes in the transmembranechloride gradient which rapidly abolish the early hyperpolarizing i.p.s.p. (Nicoll &Alger, 1981 a). It appears that the l.h.p. is a potassium potential possibly activatedby calcium. Thel.h.p. may be initiated in the dendrites, as proposed by Fugita (1979)but we have no evidence on this point. In the present paper we have demonstratedthat the fast orthodromic hyperpolarization is also complex and involves a componentwhich is produced by GABA released in s. radiatum. We propose that the dendriticcomponent is due to the activation of a feed-forward inhibitory network.
While the large size ofthel.h.p. has prevented a detailed analysis of the feed-forwardinhibitory pathway in standard medium, barbiturates provide a way to studyfeed-forward inhibition. Pentobarbitone or phenobarbitone in the bathing mediumlead to the appearance of a late bicuculline-sensitive depolarizing component of theorthodromic i.p.s.p. following a fast hyperpolarization, while the antidromic i.p.s.p.is devoid of a depolarizing component. This selective action on orthodromic responsesindicates that the barbiturate effect is not a generalized one on GABA-mediatedi.p.s.p.s. Interestingly, diazepam, although it interacts with GABA responses andenhances inhibition in the hippocampus (Lee et al. 1980, Wolf & Haas, 1979; Tsuchiya& Fukushima, 1980), does not produce a depolarizing i.p.s.p. High concentrations ofdiazepam do produce a small relative depolarization when the membrane is polarizedto the reversal potential of the hyperpolarizing i.p.s.p.
118

HIPPOCAMPAL DENDRITIC INHIBITION
We have been unsuccessful so far in identifying the ionic mechanism of the depolarizing i.p.s.p.Measurements of conductance changes and reversal potential of the depolarizing i.p.s.p. arecomplicated by its overlap with the fast hyperpolarizing i.p.s.p. and the non-GABA-mediated l.h.p.While there is clearly a large increase in chloride conductance during the depolarizing i.p.s.p. wehave evidence that the chloride gradient is inward throughout the pyramidal cell (Alger & Nicoll,1982) and therefore some ion in addition to chloride is required to account for the depolarizingresponse.
Fig. 1 1. Diagrammatic respresentation of proposed inhibitory circuits ofCAI region. Cellsand synapses in black represent inhibitory elements, while those that are open representexcitatory elements. A basket cell (a) receives excitation from collaterals ofpyramidal cells(b) and it synapses on the pyramidal cell soma. A separate population of inhibitoryinterneurones (c) receives excitation from afferents. These interneurones make inhibitorysynapses on pyramidal cell dendrites.
Observations that ionophoretic GABA responses in the apical dendritic region arepredominantly depolarizing while somatic GABA responses are hyperpolarizing(Alger & Nicoll, 1982; Andersen et al. 1980; Thalmann et al. 1981) suggested that thedepolarizing i.p.s.p. might be a dendritic event. Local TTX injections in s. radiatumselectively block the s. radiatum-evoked depolarizing i.p.s.p., indicating it dependson neuronal activity in this region. Since the depolarizing i.p.s.p. can also be blockedby local dendritic application of GABA antagonists, we conclude that the GABAreceptors for the depolarizing i.p.s.p. exist on the dendrites. In addition we have foundthat orthodromic stimulation in s. oriens generates a depolarizing i.p.s.p., and thisappears to be spatially distinct from that produced on the apical dendrities.
Fig. 11 summarizes these findings. First there is the well established recurrentcollateral pathway which involves the activation of interneurones (cell a in Fig. 11)that appear to end exclusively on the pyramidal cell somata (cell b), because pureantidromic stimulation does not lead to activation of dendritic GABA receptors. As
119

B. E. ALGER AND R. A. NICOLL
suggested previously (Andersen et al. 1964 b), the basket cells are the most likelycandidates for the recurrent collateral interneurones. Orthodromic stimulation in s.radiatum leads to the activation of GABA receptors on the apical dendrites whileorthodromic stimulation in s. oriens leads to the activation of GABA receptors onthe basal dendrites. These inhibitory pathways must also be feed-forward, since,again, recurrent collateral stimulation fails to activate them. Although we have notidentified the feed-forward interneurones involved in the pathways, both Lorente deNo' (1934) and Ramon y Cajal (1968) describe interneurones in s. radiatum and s.oriens, at least some of which are probably GABA-ergic since numerous GAD-containing cells have been described both in s. oriens and s. radiatum (Ribak et al.1978). These pathways are diagrammed in Fig. 11 (see cells labelled c). Our resultsdo not exclude the possibility of either a direct synaptic excitation of basket cellsby afferent fibres or that feed-forward interneurones form somatic as well as dendriticsynapses.The strong temperature dependence of the depolarizing i.p.s.p., which is markedly
increased by low temperatures, presumably accounts for its not having been previouslyreported. The effects of temperature are probably presynaptic on interneuroneexcitability (Andersen, Gjerstad & Pasztor, 1972) and on the transmitter releaseprocess (Hubbard, Jones & Landau, 1971). Low temperatures will also affect theGABA uptake system (Iversen & Neal, 1968) and lead to a prolongation of GABAaction. This latter idea is supported by our finding that nipecotic acid, an inhibitorofGABA uptake (Krogsgaard-Larsen & Johnston, 1975), also enhances the amplitudeof the depolarizing i.p.s.p. We would stress that in the absence of barbiturates wehave no evidence for a depolarizing dendritic i.p.s.p., and indeed based on observationssuch as those in Fig. 2 we believe that the dendritic i.p.s.p.s are normally hyperpo-larizing. In the following paper we will discuss how pentobarbitone brings out adepolarizing i.p.s.p.The dendrites of hippocampal CAI pyramidal cells are capable of producing large
depolarizing potentials which can trigger repetitive ('burst') firing when stimulateddirectly (Wong & Prince, 1979). Somatic current injections do not trigger bursts.Under ordinary conditions synaptic excitation, which occurs primarily via dendriticsynapses (Andersen, Blackstad & L0mo, 1966), is also incapable of initiating bursts.However, when synaptic inhibition is blocked, bursts are produced following synapticexcitation (Yamamoto & Kawai, 1968; Wong & Prince, 1979; Schwartzkroin &Prince, 1980). This indicates that inhibition plays a crucial role in controlling thegeneration of dendritic bursting. Since bursts can be elicited at a time when thesomatic membrane is hyperpolarized by current injection (Alger & Nicoll, 1980) itappears unlikely that inhibition generated on the soma alone would be sufficient tocontrol dendritic excitability. We have found in the absence of barbiturates thatapplication of bicuculline methiodide to the dendritic region reduces the orthodromici.p.s.p. and results in multiple spike discharge to orthodromic stimulation at a timewhen the antidromic (somatic) i.p.s.p. is unaffected (Fig. 2). Similar bursting has beenreported in response to localized dendritic penicillin application (Silfvenius, Olofsson& Ridderheim, 1980). These findings suggest that following orthodromic activationof pyramidal cells, a local dendritic i.p.s.p. inhibits the dendritic membrane. Theproposed feed-forward pathway will also temporally couple excitation and inhibition
120

HIPPOCAMPAL DENDRITIC INHIBITION
on the pyramidal cells. Our findings in the rat hippocampus are therefore inagreement with previous observations (see Llinas, 1975, for review) and suggest thatlocal inhibitory control of dendritic excitability is a widespread phenomenon.
This work was supported by N.I.H. Postdoctoral Fellowships 9-F32 NS 05744 and NS 07067 toB. E. A. and Grant NS 15674, RCDA NS 00287 and the Klingenstein Fund to R. A. N.
REFERENCES
ALGER, B. E. & NICOLL, R. A. (1979). GABA-mediated biphasic inhibitory responses in hippo-campus. Nature, Lond. 281, 315-317.
ALGER, B. E. & NIcoLL, R. A. (1980). The epileptiform burst after-hyperpolarization: A calcium-dependent potassium potential in hippocampal pyramidal cells. Science, N. Y. 210, 1122-1124.
ALGER, B. E. & NICOLL, R. A. (1982). Pharmacological evidence for two kinds of GABA receptoron rat hippocampal pyramidal cells studied in vitro. J. Physiol. 328, 125-141.
ANDERSEN, P., BLACKSTAD, T. W. & L0Mo, T. (1966). Location and identification of excitatorysynapses on hippocampal pyramidal cells. Expl Brain Res. 1, 236-248.
ANDERSEN, P., BLISS, T. V. P. & SKREDE, K. K. (1971). Unit analysis of hippocampal populationspikes. Expl Brain Res. 13, 208-221.
ANDERSEN, P., DINGLEDINE, R., GJERSTAD, L., LANGMOEN, I. A. & MOSFELDT LAURSEN, A. (1980).Two different responses of hippocampal pyramidal cells to application of gamma-aminobutyricacid. J. Physiol. 305, 279-296.
ANDERSEN, P., ECCLES, J. C. & L0YNING, Y. (1964a). Location of postsynaptic inhibitory synapseson hippocampal pyramids. J. Neurophysiol. 27, 592-607.
ANDERSEN, P., ECCLES, J. C. & L0YNING, Y. (1964b). Pathway of postsynaptic inhibition in thehippocampus. J. Neurophysiol. 27, 608-619.
ANDERSEN, P., GJERSTAD, L. & PASZTOR, E. (1972). Effects of cooling on inhibitory processes inthe cuneate nucleus. Acta physiol. scand. 84, 448461.
ANDERSEN, P., GROSS, G. N., L0Mo, T. & SVEEN, 0. (1969). Participation of inhibitory andexcitatory interneurons in the control of hippocampal cortical output. In The Interneuron, ed.BRAZIER, M., pp. 415-465. Los Angeles: University of California Press.
BARBER, R. & SAITO, K. (1976). Light microscopic visualization of GAD and GABA-T inimmunocytochemical preparations of rodent CNS. In GABA in Nervous Sy8tem Function, ed.ROBERTS, E., CHASE, T. N. & TOWER, D. B., pp. 113-132. New York: Raven.
BARKER, J. L. & RANSOM, B. R. (1978). Amino acid pharmacology of mammalian central neuronesgrown in tissue culture. J. Physiol. 280, 331-354.
CHAN-PALAY, V. (1978). Quantitative visualization of y-aminobutyric acid receptors in hippocam-pus and area dentata demonstrated by [3H] muscimol autoradiography. Proc. natn. Acad. Sci.U.S.A. 75, 5 2516-2520.
CURTIS, D. R., FELIX, D. & MCLENNAN, H. (1970). GABA and hippocampal inhibition. Br. J.Pharmac. 40, 881-883.
DINGLEDINE, R. & GJERSTAD, L. (1980). Reduced inhibition during epileptiform activity in thein vitro hippocampal slice. J. Physiol. 305, 297-313.
DINGLEDINE, R. & LANGMOEN, I. A. (1980). Conductance changes and inhibitory actions ofhippocampal recurrent i.p.s.p.s. Brain Res. 185, 277-287.
ECCLES, J. C., NICOLL, R. A., OSHIMA, T. & RUBIA, F. J. (1977). The anionic permeability of theinhibitory postsynaptic membrane of hippocampal pyramidal cells. Proc. R. Soc. B 198, 345-361.
FUGITA, Y. (1979). Evidence for the existence of inhibitory postsynaptic potentials in dendrites andtheir functional significance in hippocampal pyramidal cells ofadult rabbits. Brain Res. 175, 59-69.
FUGITA, Y. & SAKATA, H. (1962). Electrophysiological properties of CA1 and CA2 apical dendritesof rabbit hippocampus. J. Neurophysiol. 25, 209-222.
GOTTLIEB, D. I. & COWAN, W. M. (1972). On the distribution of axonal terminals containingspheroidal and flattened synaptic vesicles in the hippocampus and dentate gyrus of the rat andcat. Z. Zellforsch. mikrosk. Anat. 129, 413-419.
HUBBARD, J. I., JONES, S. F. & LANDAU, E. M. (1971). The effect of temperature change upontransmitter release, facilitation and post-tetanic potentiation. J. Physiol. 216, 591-609.
121

B. E. ALGER AND R. A. NICOLL
IVERSEN, L. L. & NEAL, M. J. (1968). The uptake of [3H] GABA by slices of rat cerebral cortex.J. Neurochem. 15, 1141-1149.
KANDEL, E. R., SPENCER, W. A. & BRINLEY, F. J. (1961). Electrophysiology of hippocampalneurons. I. Sequential invasion and synaptic organization. J. Neurophysiol. 24, 225-242.
KROGSGAARD-LARSEN, P. & JOHNSTON, G. A. R. (1975). Inhibition of GABA uptake in rat brainslices by nipecotic acid, various isoxazoles and related compounds. J. Neurochem. 25, 787-802.
LEE, H. K., DUNWIDDIE, T. V. & HOFFER, B. J. (1979). Interaction of diazepam with synaptictransmission in the in vitro rat hippocampus. Naunyn-Schmiedebergs Arch. Pharmak. 309,131-136.
LEE, H. K., DUNWIDDIE, T. & HOFFER, B. (1980). Electrophysiological interactions of enkephalinwith neuronal circuitry in the rat hippocampus. II. Effects on interneuron excitability. BrainRes. 184, 331-342.
LEUNG, L. S. (1978). Hippocampal CA1 region - demonstration of antidromic dendritic spike anddendritic inhibition. Brain Res. 158, 219-222.
LLINAS, R. (1975). Electroresponsive properties of dendrites in central neurons. In Advances inNeurology, ed. KREUTZBERG, G. W., vol. 12, pp. 1-13. New York: Raven.
LORENTE DE No', R. (1934). Studies on the structure of the cerebral cortex. II. Continuation ofthe structure of the ammonic system. J. Psychol. Neurol., Lpz. 46, 225-242.
LYNCH, G. S., JENSEN, R. A., MCGAUGH, J. L., DAVILA, K. & OLIVER, M. W. (1981). Effects ofenkephalin, morphine and naloxone on the electrical activity of the in vitro hippocampal slicepreparation. Expl Neurol. 71, 527-540.
MATTHEWS, G. & WICKELGREN, W. 0. (1979). Glycine, GABA and synaptic inhibition of reticulo-spinal neurones of lamprey. J. Physiol. 293, 393-415.
NICOLL, R. A. & ALGER, B. E. (1981 a). Synaptic excitation may activate a calcium-dependentpotassium conductance in hippocampal pyramidal cells. Science, N. Y. 212, 957-959.
NICOLL, R. A. & ALGER, B. E. (1981 b). A simple chamber for recording from submerged brain slices.J. Neurosci. Meth. 4, 153-156.
NICOLL, R. A., ECCLES, J. C., OSHIMA, T. & RUBIA, F. (1975). Prolongation of hippocampalinhibitory postsynaptic potentials by barbiturates. Nature, Lond. 258, 625-627.
PONG, S. F. & GRAHAM, L. T. (1973). A simple preparation of bicuculline methiodide, a watersolubleGABA antagonist. Brain Res. 58, 266-267.
PURPURA, D. D. & PAPPAS, G. D. (1968). Structural characteristics of neurons in the felinehippocampus during postnatal ontogenesis. Expl Neurol. 22, 379-393.
PURPURA, D. P., PRELEVIC, S. & SANTINI, M. (1968). Postsynaptic potentials and spike variationsin feline hippocampus during postnatal ontogenesis. Expl Neurol. 22, 408-417.
RAMON Y CAJAL, S. (1968). The structure of Ammon's Horn, translated Kraft, L. M., Springfield:Charles C. Thomas.
RIBAK, C. E., VAUGHN, J. E. & SAITO, K. (1978). Immunocytochemical localization ofglutamic aciddecarboxylase in neuronal somata following colchicine inhibition of axonal transport. Brain Res.140, 315-332.
SCHOLFIELD, C. N. (1978). Electrical properties of neurones in the olfactory cortex slice in vitro.J. Physiol. 275, 535-546.
SCHWARTZ, I. R., PAPPAS, G. D. & PURPURA, D. D. (1968). Fine structure of neurons and synapsesin the feline hippocampus during postnatal ontogenesis. Expl Neurol. 22, 394-407.
SCHWARTZKROIN, P. A. & ALTSCHULER, R. J. (1977). Development of kitten hippocampal neurons.Brain Res. 134, 429-444.
SCHWARTZKROIN, P. A. & PRINCE, D. A. (1980). Changes in excitatory and inhibitory synapticpotentials leading to epileptogenic activity. Brain Re8. 183, 61-76.
SILFVENIUS, H., OLOFSSON, S. & RIDDERHEIM, P-A. (1980). Induced epileptiform activity evokedfrom dendrites of hippocampal neurones. Acta. physiol. scand. 108, 109-111.
STORM-MATHISEN, J. (1977). Localization of transmitter candidates in the brain: the hippocampalformation as a model. Progr. Neurobiol. 8, 119-181.
THALMANN, R. H., PECK, E. J. & AYALA, G. F. (1981). Biphasic response ofhippocampal pyramidalneurons to GABA. Neurosci. Lett. 21, 319-324.
TsUCHIYA, T. & FUKUSHIMA, H. (1978). Effects of benzodiazepines and pentobarbitone on theGABA-ergic recurrent inhibition of hippocampal neurons. Eur. J. Pharmac. 48, 421-424.
122

HIPPOCAMPAL DENDRITIC INHIBITION 123
WOLF, P. & HAAS, H. L. (1977). Effects of diazepines and barbiturates on hippocampal recurrentinhibition. Naunyn-Schmiedebergs Arch. Pharmak. 299, 211-218.
WONG, R. K. S. & PRINCE, D. A. (1979). Dendritic mechanisms underlying penicillin-inducedepileptiform activity. Science, N. Y. 204, 1228-1231.
WONG, R. K. S., PRINCE, D. A. & BASBAUM, A. I. (1979). Intradendritic recordings from hippo-campal neurons. Proc. natn. Acad. Sci. U.S.A. 76, 986-990.
YAMAMOTO, C. & KAWAI, N. (1968). Generation of the seizure discharge in thin sections from theguinea pig brain in choride free medium in vitro. Jap. J. Physiol. 18, 620-631.