zur mikrobiologisch-chemischen gewinnung von pyocyanin in...
TRANSCRIPT

This work has been digitalized and published in 2013 by Verlag Zeitschrift für Naturforschung in cooperation with the Max Planck Society for the Advancement of Science under a Creative Commons Attribution4.0 International License.
Dieses Werk wurde im Jahr 2013 vom Verlag Zeitschrift für Naturforschungin Zusammenarbeit mit der Max-Planck-Gesellschaft zur Förderung derWissenschaften e.V. digitalisiert und unter folgender Lizenz veröffentlicht:Creative Commons Namensnennung 4.0 Lizenz.
durch die Polarisierungsenergie, die eine Nerven-eigenschaft ist. Daher ist, in Übereinstimmung mit den experimentellen Befunden, das Ausmaß der auslösenden Störung, wenn sie den Schwellen-wert überschritten hat, ohne Einfluß auf die Stärke des Reizes.
Die refraktäre Phase, während der nach einem Reizablauf ein zweiter Reiz nicht gesetzt werden kann, ist gegeben durch die Zeit, die zur er-neuten Phosphorylierung der Amidkette benötigt wird. Die Ermüdung des Nerven durch viele hin-tereinander folgende Reizabläufe ist verursacht durch den Verbrauch der ATP, d.h. durch Sen-kung des Phosphatenergiespiegels. Da das Volu-men der leitenden Substanz des Nerven im Ver-gleich zum Muskel sehr gering ist, ist der Energie-bedarf bei der Nervenreizleitung entsprechend klein und der experimentelle Nachweis der Phos-phorylierungsvorgänge erschwert.
Der Mechanismus der Reizfortleitung als fort-schreitende Depolarisation der Amidketten und damit Polarisation der Ionenladungen steht in völliger Analogie zu der Auswirkung des Reizes auf Muskel oder elektrisches Organ, die eben-
falls eine Depolarisation darstellt, verursacht durch Abspaltung der Phosphorsäure. Das Auf-treten eines Aktionsstromes bei der Muskelbewe-gung weist darauf hin, daß die Polarisierung der Amidkette auch im Muskel trotz der Umfal-tung noch eine elektrische Auswirkung hat. Die Analogie macht es wahrscheinlich, daß die Pola-risation im Nerv in gleicher Weise die Folge einer Phosphorylierung ist, und daß in allen drei Organen der gleiche Übertragungsmechanismus vorliegt. Die Rolle des Acetylcholiris ist vornehm-lich in diesem Übertragungsmechanismus ^ zu suchen. Die Einschaltung der negativen Ionen-überschußladung in den Mechanismus der Ab-spaltung der Phosphorsäure aus der folgenden Amidkette im Nerv — und vom Nerv zum Muskel oder elektrischen Organ — stellt sich in den ex-perimentellen Befunden als eine völlige Ent-sprechung chemischer und elektrischer Reizung dar26'27.
Wir danken P. L y u e n und H. F r i e d r i c h -F r e k s a fü r wichtige Anregungen und Hinweise.
26 S. W. K u f f 1 e r , J . Neurophysiol . 9, 367 [1946]. 27 J . C. E c c l e s , Annu. Rev. Physiol. 1948, 93.
Zur mikrobiologisch-chemischen Gewinnung von Pyocyanin in der Submerskultur*
V o n A N N E M A R I E G R O S S E R u n d W I L H E L M FRIEDRICH
(Z. Naturforschg. 3b, 425—430 [1948]; eingegangen am 27. September 1948)
Es wird eine einfache Methode zur Gewinnung von Pyocyanin mit Hilfe von Pseudo-monas pyocyanea (Bacterium pyocyaneum) beschrieben. Unte r Anwendung moderner Gär technik (Submerskul tur mit intensivem Rühren und k r ä f t i g e r Be lü f tung ) wurden in geeigneten Nährmedien bereits nach 16 Stdn. bei 32° Lösungen erhal ten, die bis 17 mg-% Pyocyanin enthielten. Zu dessen Bes t immung wurde eine photometr ische Methode und zur p räpara t iven Gewinnung eine Adsorpt ionsmethode (un te r Verwen-dung von Carboraff in als Adsorpt ionsmit tel und Chloroform-Methanol als Elut ionsmit te l ) ausgearbei te t . Als Nebenprodukt des Pvocyanins wurde in k le iner Menge ein gelber Farbstoff C1 4H802N, isoliert.
Das Pyocyanin, der blaue Farbstoff des Bact. pyocyaneum, wurde bereits 1859 durch F o r -
d e s kristallisiert gewonnen. Die Konstitution
* 3. Mitt. in der Veröffent l ichungsreihe „Über die Bi ldung antibiot ischer Stoffe durch Mikroorganismen' ' von K. B e r n h a u e r und Mitarbb. Ans dem ehe-mal igen In s t i t u t f ü r Biochemie und Nahrungsmit te l -chemie der Deutschen Techn. Hochschule in P r a g . — Vgl. dazu W. F r i e d r i c h , Diss. P rag u. Graz 1947. — Die h i e r veröffent l ichten Versuche waren berei ts An- y fang 1945 abgeschlossen.
wurde von W r e d e und S t r a c k 1 ermittelt. Die Synthese gelang durch Methylierung von a-Oxy-phenazin mittels Dimethylsulfats2.
Von In teresse ist die antagonis t ische W i r k u n g der B. pyocyaneum-Kulturen gegenüber zahlreichen Mikro-
1 W r e d e u. S t r a c k , Hoppe-Seyler 's Z. phy-siol. Chem. 140. 1 [1924]; 142, 103 [1925]; 177, 177 [1928]; 181, 58 [1929].
2 W r e d e u. S t r a c k , Ber . dtsch. chem. Ges. 62, 2053 [1929]. — Hinsichtl ich einer neuen Arbei ts-vorschr i f t vgl. Org. Svntheses 26, 89 [1946].

Organismen. F ü r diese W i r k u n g ist nach ä l teren An-gaben 3 e inersei ts die in Alkohol, Äther , Benzol, Ace-ton und Chloroform lösliche Pyocyanase und anderer-seits das Chloroform- und wasserlösliche blaue Pigment verantwor t l ich . Die Pyocyanase besitzt Lipoidnatur und besteht aus höheren ungesä t t ig ten F e t t s ä u r e n 4 . Sie wurde wegen ihrer bactericiden und bacteriolyt i-schen Eigenschaf ten berei ts um die J ah rhunde r twende mit Hilfe von B. pyocyaneum technisch e rzeugt und in den Handel gebracht, doch hat sie die in sie gesetzten E r w a r t u n g e n nicht e r fü l l t 5 . Während die Pyocyanase vor allem in ä l teren Ku l tu ren au f t r i t t , ist die bacteri-cide W i r k u n g junger Pyocyaneusku l tu ren vorwiegend auf deren Pigmentgehal t z u r ü c k z u f ü h r e n 6 . Das Pyo-cyanin w i rk t cytolytisch sowie bactericid, u. a. auch auf Diphther iebazi l len; es war daher zur Spray-behandlung von DLphtheriebaci l lenträgern empfohlen worden 7 . Auch in neuerer Zeit beschäf t ig te man sich mit der bacter ios ta t i sch-bacter ic iden W i r k u n g des Pyocyanins und Hemipyopyan ins 7 a . In den letzten J a h r e n wurde eine Anzahl wei te re r ant ibacter ie l ler Substanzen aus Pyocyaneus-Ivul turen isoliert . Hier-her gehören zwei nicht näher identif izierte bacterio-stat ische Subs tanzen 7 b , f e rne r das a-Oxy-phenazin 7 c
und die gegenüber Mycobacterium tuberculosis wirk-same Pyo l ip in säu re 7 d . Außerdem wurden kürzl ich auch noch Chlororapliin und Oxychlororaphin (Phena-zin-a-carbonamid) als S tof fwechselprodukte des Bact. pyocyaneum a u f g e f u n d e n 7 6 . Schließlich konnten aus den Zellen einer unpigment ie r ten Var i an te des Bact. pyocyaneum zahlreiche ant ibacter ie l le Stoffe in kri-s ta l l i s ier ter Form isoliert werden, die als Pyo I, I I , I I I und IV sowie als l b und l c bezeichnet wurden. Der Körper Pyo I ist schwach antibiotisch wirksam und hat die Zusammensetzung CnHi0N2Oa7 f . In allen
3 Vgl. dazu die zusammenfass . Dar s t e l lung über die Gruppe der Fluorescenten von L e m b k e , Vor-ra tspf lege Lebensmit te l forschg. 5, 265 [1942-].
4 H. 0 . H e t t c h e , Z. Immuni tä t s forschg . exp. Therap . 83, 499, 506 [1934]. — Wei te re L i t e ra tu r -angaben hierzu vgl. bei W a l l e n f e l s , Die Chemie 58, 1 [1945].
5 Vgl. dazu z. B. R. L i e s k e , Morphologie und Biologie der Strahlenpilze. Born t r äge r , Leipzig 1921, S. 143.
0 0 . H e t t c h e u. W. V o g e l , Arch. Hyg. Bak-teriol. 117, 234 [1937].
7 Vgl. dazu H. O. H e 11 c h e , Arch. Hyg . Bakteriol . 107, 337 [1932].
7 a N. P r a d i n e s B r a z i 1, Arch. u r u g u a y . med. cirug. y especial. (Montevideo) 25, 627 [1944]: vgl. Chem. Abstr . 39, 4111 [1945].
7 b P. F r e d e r i c q , C. R. Seances Soc. Biol. Fil ia-les Associees 140, 683, 686 [1946].
7c J. Z w e i g , Veter in . J . 102, 55 [1946]; Exp. Stat. Ree. 95, 242 [1946]; vgl . Chem. Abstr . 42, 5950 [1948].
7d II. D a v i d e , Ark . Kein., Mineral. Geol., Ser. B 19, Nr. 7 [1945]; S. B e r g s t r ö m , E T l i e o r e l l u. H. D a v i d e , Arch. Biochemistry 10, 165 [1946] (vgl. Chem. Abstr . 40, 5794 [1946]); Ark . Kern., Mineral. Geol., Ser. A 23, Nr. 13 [1946] (vgl. Chem. Abstr . 41, 1278 [1947]).
diesen Fäl len handelt es sich um gelb gefärb te oder farblose Substanzen.
Im Stoffwechsel der B. pyocyaneum-Kulturen soll das Pyocyanin eine Funkt ion als Ho-Überträger be-sitzen, da es sich leicht zur Leukobase reduzieren läßt, die durch Luftsauers toff wieder in die Fa rb -base ü b e r f ü h r b a r ist8 . Bei Sauers toffabschluß soll eine P igmentvors tu fe gebildet werden, da bei Lu f t zu -t r i t t rasche F ä r b u n g erfolgt 8 . Im physiologischen pH-Bereicli (6—9) entspricht das Redox-Potential des Pyocyanins e twa dem des Methylenblaus9 . Es bewi rk t eine Atmungss te igerung lebender Bakter ien wie auch von deren Acetonpräparaten. In anaeroben Versuchen mit lebenden Bakterien können in Gegenwar t von Pyocyanin Lacta t und zahlreiche andere Substanzen als H2-Donatoren verwer te t werden 1 0 .
Mit der mikrobiologisch-chemischen Gewinnung des Pyocyanins beschäftigte sich eine ganze An-zahl von Autoren, wobei die verschiedensten Nähr-böden empfohlen wurden. Während W r e d e und S t r a c k ^ 2 Ragitbouillon benutzten, schlugen M a r x und W o i t h e 1 1 KBr-Bouillon vor . K r a m e r 1 2 verwendete Glycerin-Peptonw^asser. S t h e e m a n 8 erzielte unter Verwendung eines Peptonmediums bei manchen Bakterienstämmen durch Zusatz von Nitraten intensive Farbstoffbil-dung. Auch E 1 e m a und S a n d e r s 1 3 berichteten über kräftige Pvocyaninproduktion, und F r i e d -l e i n 1 4 empfahl die Verwendung von Na-Lactat als C-Quelle. In jüngerer Zeit beschäftigte sich C i c c o n i 1 5 mit den die Pyocyaninbildung fördern-den bzw. hemmenden Faktoren. Sie fand u. a., daß sich in Reiskleie, Malzextrakt, Leberextrakten und Hefewasser ein Hemmstoff befindet, der die Pyocyaninbildung durch B. pyocyaneum verhin-dert. ohne das Bakterienwachstum zu beeinträch-tigen. Ferner liegen aus der neuesten Zeit Unter-
7e L. B i r k o f e r , Ber. dtsch. chem. Ges. 80, 212 119471.
7 f E. E. II a y s , J . C. W e 11 s , Ph. A. K a t z m a n , C. K. C a i n , F. A . J a k o b s , S. A. T h a y e r , E. A. D o i s y , W. L. G a b y , E. C. R o b e r t s , R. D. M u i r , C. J . C a r r o 11, L. R. J o n e s u. N. J . W a d e , J . biol. Chemistry 159, 725 [1945].
8 A. A. S t h e e m a n , Biochem. Z. 191, 320 [19271. H B . E l e m a , Recueil Trav . chim. Pays-Bas 50, 807
[1931]. 10 O. E h r i s m a n n , Z. Hyg. Infekt . -Krankli . 116.
209 [1935]. 11 M a r x u. W o i t h e , Zbl. Bakteriol. , Pa ras i t en-
kunde Infekt ionskrankh. , Abt, I, 28, 1, 33, 65, 97 [1900]. 12 H. K r a m e r , Z. Immunitä tsforschg. exp. The rap .
84, 505 [1935]. 13 B. E l e m a u. A. C. S a n d e r s , Recueil T r a v .
chim. Pays-Bas 50, 796 [1931]. 14 F. F r i e d 1 e i n , Biochem. Z. 194, 273 [1928]. 15 M. C i c c o n i , Zbl. Bakteriol. , Pa ra s i t enkunde
Infekt ionskrankh. , Abt. I. Orig. 149, 102 [1942/43].

Nährsubstrat End-juH
Pyocyanin (Anfangs-pH 7,6) End-juH Gebalt (Anfangs-pH 7,6)
™g-% I Glycerin-Pepton-Wasser12 7,7 1,4
I I Fleischwasser mit l°/0 8,6 4,4 Pepton u. 0,5 °/0 KBr1 1 4,4
I I I Fleischwasser mit l°/0 8,4 3,7 Pepton u. 0,5 °/0 NaCl
3,7
IV 5°/0 Ca-Gluconat, 7,4 0,55 0 ,8% (NH4)„S04, 0 , 1 % KH 2 P0 4 ,
0,55 0 ,8% (NH4)„S04, 0 , 1 % KH 2 P0 4 , 0,025% MgSOj • 7 H , 0
V 5 % techn. Glucose, 5,1 0 Salze wie bei IV
0
VI 5 % Ca-Gluconat, 7,5 2,7 0 ,5% Hefeextrakt
7,5
„Vitambact" VI I 5 % Ca-Gluconat, 7,6 Spuren
0,1% Vitambact Spuren
VI I I 5 % techn. Glucose, 4,1 0 .
0 ,5% Vitambact 0
suchungen über die Wirkung von Aminosäuren1 5 a
und Mineralstoffen 15b auf die Pyocyaninbildung vor.
Bei allen bisherigen Untersuchungen über die Pyocyaningewinnung mittels Ps. pyocyanea wurde in der Oberflächenkultur (Ruhekultur) gearbei-tet. Wegen der bekannten Mängel derselben stell-ten wir uns die Aufgabe, die Herstellung des Pyocyanins unter Anwendung moderner Gär-technik, also unter submersen Bedingungen zu prüfen (Schüttel-, Durchlüftungs- und Rührkul-tur). Außerdem erschien die Ausarbeitung einer einfachen Methode zur Isolierung des Pyocyanins wünschenswert, da bei der bisher üblichen Extrak-tion mit Chloroform relativ große Volumina ver-arbeitet werden müssen. Eine Voraussetzung für die erstgenannte Aufgabe war die Ausarbeitung einer geeigneten Methode zur analytischen Ver-folgung des Pyocyaningehaltes des Kulturmediums.
Zur Bestimmung des Pyocyaningehaltes der Kulturflüssigkeiten versuchten wir zunächst, photometrische Messungen der blauen Kultur-filtrate durchzuführen. Da sich aber die schlei-migen Bakterienzellen weder durch Filtrieren noch durch Zentrifugieren schnell beseitigen ließen, mußten diese Versuche aufgegeben werden. Das Photometrieren der blauen Chloroformaus-züge ergab auch kein brauchbares Ergebnis, da in das Chloroform ein gelber Farbstoff mit über-ging, der die photometrischen Messungen un-geeignet machte. Es war deshalb notwendig, die Chloroformextrakte mit salzsaurem Wasser aus-zuschütteln und die so erhaltenen roten Lösungen des salzsauren Pyocyanins zu photometrieren. Arbeitsvorschrift vgl. unten.
Bei der Prüfung von 18 B. pyocyaneum-Stäm-men erwiesen sich nur 4 als Pyocyanin-Bildner, während die anderen trotz guten Wachstums nur wenig oder kein Pyocyanin erzeugten. Zur Fort-züchtung der Kulturen bewälirje sich neben Bouillon-Pepton-Agar ein Ca-Gluconat-Agar mit anorganischen Nährsalzen.
Versuche zur Auswahl eines günstigen Nähr-bodens wurden in der Schüttelkultur vorgenom-men, da diese zur Durchführung großer Versuchs-reihen am einfachsten zu handhaben ist. Als Vor-
M- 0 . B u r t o n , B. A. E a g 1 e s u. J . J . R. C a m p b e l l , Canad. J . Res., Sect. C 25 121 r19471 • vgl. Chem. Abstr. 42, 944 [1948].
1 5 b M. 0 . B u r t o n , J . J . R. C a m p b e l l u. B. 4 E a g l e s , Canad. J . Res., Sect, C 26, 15 [19481 • vgl Chem. Abstr . 42, 5503 [1948].
l a b . 1. Versuchsre ihe zur P r ü f u n g der E ignung ver-schiedener Nährsubs t ra te in der Schüt te lkul tur (ie 100 cm3 Nahrmedium, Versuchsdauer 72 Stdn.. Temp. 32°).
kultur fü r diese Versuche, wie auch für alle wei-teren, diente eine 24 Stdn. alte Schüttelkultur. In Tab. 1 sind die Ergebnisse einer Versuchsreihe zur Prüfung der Eignung verschiedener Nähr-substrate wiedergegeben. Die Pyocyaninbestim-tnung erfolgte dabei na^h 3-tägigem Schütteln bei 32°. ^ Aus der Versuchsre ihe ergibt sich, daß in dem von
K r a m e r 12 empfohlenen Glycer in-Peptonwasser (I) nur ger inge Pyocyaninbi ldung erfolgte. Wohl aber bewähr te sich die KBr-Bouil lon ( I I ) von M a r x und W o i t h e " ; a l lerdings war dieselbe der von uns zu Vergleichszwecken herangezogenen NaCl-Bouillon ( I I I ) nur wenig über legen. Im übr igen erwies sich auch ein Ca-Gluconat-Hefeextrakt-Nährboden (VI) als recht geeignet, wogegen Glucosemedien ungeeignet waren. , da dabei Gluconsäure entsteht , die — wenn nicht f ü r Neut ra l i sa t ion gesorgt wird — ein s ta rkes Absinken des pH-Wertes verursacht, wodurch die Organismen in ihrem Wachs tum s tark gehemmt werden.
Weitere Versuche wurden in der Durchlüftungs-kultur durchgeführt, wobei Hefebelüfter (Schott & Gen., Jena) verwendet werden. Als Beispiel für den Versuchsverlauf unter diesen Bedingun-gen sei ein Durchschnittsversuch mit der Nähr-lösung VI (vgl. Tab. 1) angeführt (1/-Hefebelüf-ter mit 300 cm3 Nährlösung):
Versuchsdauer Stdn. 24 32 42 48 Pyocyanin-Gehalt mg-% 8,8 11,15 12,15 11,6

Nährsubstrat Pyocyanin-Gehalt
in mg-% nach Stdn.
16 ' / , 21 V2 27 Y8
IX 3°/0 Ca-Gluconat, 0,5 °/0 Vitambact
11,1 — 13,0
X
X I
3°/o Ca-Gluconat, 0,2 O/o NH,N0 3 , 0 ,1% KH2PO4 , 0,025% MgS0 4 • 7 H 2 0
wie X, aber mit 0 ,25% Vitambact
4,9
17,0
8,3
20,0
9,2
20,7
X I I wie XI, außerdem mit 0,2 O/0 K N 0 3 .
16,9 17,0 16,6
Tab. 2. Versuchsre ihe mit geeigneten Nährsubs t ra ten in der R ü h r k u l t u r un ter Be lü f tung (je 300 cm3 Nähr-
medium. Temp. 32° ) .
Wie ersichtlich, wird bei dieser Versuchsanordnung die Pyocyanin-Bildung gegenüber den Schüt te lver-suchen bei Verwendung der gleichen Nähr lösung auf das 4—5-fache gesteigert , un ter gleichzeit iger Ver-minderung der Versuchsdauer auf e twa die Hä l f t e . Als recht störend erwies sich dabei aber das s t a rke Schäumen des Kul turmediums, wodurch häuf ig ein Über laufen desselben verursacht wurde.
Erst bei Anwendung der Rührkultur-Technik verliefen die Versuche einwandfrei. Wir benütz-ten dabei für Vergleichsversuclie 1/ - Rührgefäße mit je 300 cm3 Lösung und KPG-Rührwerk mit 720 U/min. Dabei wurde ein kräftiger Luftstrom auf die Oberfläche der rasch bewegten Flüssig-keit geblasen, da beim Einleiten der Luft in die Flüssigkeit gleichfalls starkes Schäumen eintrat. Unter diesen Bedingungen setzte die Pyocyanin-Bildung bereits 5 Stdn. nach der Beimpfung ein und erreichte nach 16—22 Stdn. ihr Maximum. Vergleichsversuche unter diesen Bedingungen zeigten, daß bei Nährboden VI die Ca-Gluconat-menge ohne Beeinträchtigung des Resultates auf 3% herabgesetzt werden kann; dagegen ver-ursachte eine weitere Verminderung auf 2 und 1% bereits ein deutliches Absinken der Ausbeute. Ca-Lactat wrar weniger geeignet als Ca-Gluconat, und Ca-Acetat ergab gar keine Pyocyanin-Bil-dung. Die von S t h e e m a n 8 vorgeschlagene Nähr-salzlösung mit Pepton-Witte versagte bei unseren B. pyocyaneum-Stämmen völlig (kein Bakterien-wachstum). In Tab. 2 werden einige Nährlösungen angeführt, die sich gut bewährt haben.
Wie ersichtlich, wurde die beste Ausbeute an Pyocyanin bei gleichzeit iger Anwesenheit von Nähr-salzen und Hefeex t rak t in einem Ca-Gluconat-Nälir-boden erzielt (XI ) . Ein Zusatz von KNO s , das sich
nach S t h e e m a n 8 bei manchen B a k u rienstämmen bewähr te , zu einem solchen Nährboden 1 ra hte keinen Vortei l ( X I I ) . Die K u r v e d r Abb. 1 l l l v . e i n -gehender den V e r s u c h t e . l a u f unter Benin ; i' v Nähr lösung XI. Es ist dabei ein Versuch r . i t reichen Probeentnahmen wiedergegeben.
Isolierung des Pyocyanins. Die von W r e d e und S t r a c k 1 - 2 beschriebene und auch von den nachfolgenden Autoren im Prinzip übernom-mene Methode der Isolierung des Pyocyanium-Pigmentes hat den Nachteil, daß dabei größere Mengen Flüssigkeit mit Chloroform ausgeschüt-telt werden müssen. Außer dem Chloroformauf-wand birgt dieses Verfahren oft kaum überwind-
r 5
Abb. 1. Versuchäver lauf bei Verwendung der Nähr-lösung XI (vgl. Tab. 2) .
bare Schwierigkeiten in sich, die auf der Ent-sleliung sehr beständiger, schwer trennbarer Emulsionen beruhen. Größere Mengen Kultur-flüssigkeit können mittels dieser Methode nicht verarbeitet werden. Um die Nachteile dieses Ver-fahrens zu umgehen, versuchten wir Pyocyanin durch Adsorption zu gewinnen. Wir fanden, daß Pyocyanin sehr leicht an Carboraffin adsorbiert wird. Es genügt ein Zusatz von 0,15% Carbo-raffin, um eine Kulturflüssigkeit von 10 mg-% Pvocyanin-Gehalt zu entfärben. Als Elutions-mittel bewährte sich ein Gemisch von 90 % Chloroform und 10 % Methanol (weder reines Chloroform, noch reines Methanol hatte eluie-rende Wirkung). Arbeitsvorschrift vgl. unten.
Bere i t s bei den Versuchen zur analytischen Be-s t immung des Pyocyanin-Gehal tes der Kul turf lüss ig-kei ten zeigte sich, daß neben dem Pyocyanin noch ein gelber Farbstoff gebildet wird. In der L i t e r a tu r finden sich zahlreiche Angaben über das Auf t re ten gelber bis gelbroter Fa rbs to f fe in B. pyocyaneum-Kulturen. Außer den bere i t s oben ange füh r t en Befunden über antibiotisch wi rksame gelbe Substanzen liegt noch
eine Anzah l ä l t e r e r Angaben übe r gelbe Stof fwechse l -p r o d u k t e des B. pyocyaneum vor . H i e r h e r g e h ö r t die P y o x a n t h o s e von G e s s a r d (1882) und B o 1 a n d (1899) sowie eine von E l e m a und S a n d e r s 1 3 be-obachte te gelbe Substanz, die als a -Oxy-phenaz in an-gesehen wurde , f e r n e r das P y o r u b i n von M e a d e r 1 6
und schl ießl ich eine bei 239° schmelzende gelbe Sub-s tanz, die in a l ten Pyocyaneum-Kulturen a u f t r i t t 1 7 . Bei dem von u n s i so l ie r ten gelben Fa rbs to f f Ci4H802N2
vom Schmp. 237,5—238° (Zers . ) könn te es sich even-tue l l um den be re i t s von S e i x a s P a l m a 1 7 beschr ie -benen K ö r p e r hande ln oder um ein D e h y d r i e r u n g s p r o -d u k t der oben e r w ä h n t e n 7 ' V e r b i n d u n g C I 4 H 1 0 O 2 N 2 .
B e s c h r e i b u n g d e r V e r s u c h e Die Stammkulturen von Ps- pyocyanea (Bact. pyocya-
neum) w u r d e n auf Bou i l lon -Pep ton-Agar f o r t g e f ü h r t . Z u r Auswahl geeigneter Bakterienstämme d iente ein
Ca-Glucona t -Agar , bes tehend aus 3 % Ca-Gluconat , 1,8% A g a r , 0 ,2% NH4NO3, 0 ,1% K H 2 P 0 4 und 0,02% M g S 0 4 - 7 H 2 0 vom p ^ l f i . D u r c h g u t e Bakter iens tämme wurde dieser Nährboden inne rha lb 48 Stdn. bei 3 2 ° in tens iv b l au ge fä rb t ,
Als Impfkultur b e w ä h r t e sich eine bei 3 2 ° f r i s ch angewachsene ( e twa 24-stdge.) S c h ü t t e l k u l t u r auf 3% Ca-Cluconat mit 0,5% Hefeextrakt („Vi tambact") . Z u r B e i m p f u n g d ienten 1—2% des Ansa tzes . (Das Be-impfen der Versuche mit e iner A g a r - V o r k u l t u r er-wies sich a ls u n g ü n s t i g . )
Schüttelversuche. Als G e f ä ß e w u r d e n 300 c m 3 - E r l e n -meyerko lben mit je 100 cm3 N ä h r l ö s u n g benü t z t . Die Schü t te lmasch ine w a r in e iner G ä r k a m m e r von 3 2 ° a u f g e s t e l l t und machte 80—100 Hin- und ebensovie le H e r g ä n g e p r o min; Hubwe i t e 5—6 cm.
Durchlüftungsversuche. E s w u r d e n 1 f - He febe lü f -t e r 1 8 mi t je 300 cm3 Näh r lö sung benü tz t . Die In t en -s i t ä t der B e l ü f t u n g wi rd durch das s t a r k e Schäumen der N ä h r f l ü s s i g k e i t begrenz t .
Rührversuche. Zumeist wurden 1 i - R ü h r g e f ä ß e (Re-akt ionsbecher mit p langesch l i f fenem, 4-fach tubu l i e r -tem Decke l ) mi t k le inem K P G - R ü h r w e r k 1 8 b e n ü t z t (720 U/min ) . S t a r k e B e l ü f t u n g durch E inb lasen s t e r i -ler L u f t (durch W a t t e f i l t e r filtriert) auf die Ober-f läche der ro t i e renden F lü s s igke i t , Die d i r e k t e Be-l ü f t u n g der N ä h r l ö s u n g e rwies sich a ls u n v o r t e i l h a f t , da dabei s t a r k e s Schäumen s t a t t f and . F ü r g r ö ß e r e Ansä t ze w u r d e n b i rnen fö rmige R e a k t i o n s b e h ä l t e r 1 8
benü tz t (von 10, 20 und 30 l I nha l t ) , die in sinn-gemäße r We i se ad ju s t i e r t und a n g e w e n d e t w u r d e n 1 9 .
Analytische Bestimmung des Pyocyanins. 10 cm3
K u l t u r f l ü s s i g k e i t werden , ohne zu filtrieren, mit 10 cm3
Chloro fo rm geschüt te l t . Dazu b e n ü t z t man zweck-m ä ß i g ein langes , n icht zu wei tes Rohr mit u n t e n an- N
gebrach tem Hahn . Das Schüt te ln m u ß durch vors ich-t iges Umschwenken e r fo lgen , da bei h e f t i g e m D u r c h -mischen eine sehr bes tänd ige Ch lo ro fo rm - W a s s e r -
16 M e a d e r , Zbl. Gewerbehyg . U n f a l l v e r h ü t . 12. 135 [1926].
17 S e i x a s P a l m a , Zbl. Bakte r io l . , P a r a s i t e n -k u n d e I n f e k t i o n s k r a n k h . , Abt. I, Or ig . 43, 417 [19071.
18 Von Schott & Gen., Jena .
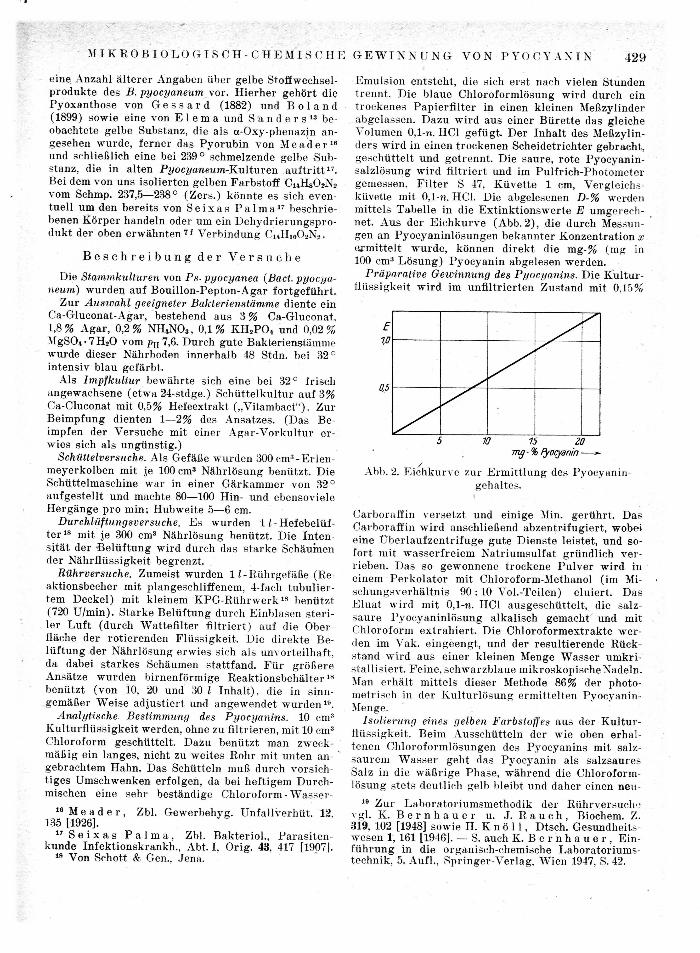
E m u l s i o n en t s t eh t , die sich e r s t nach v ie len S tunden t r e n n t . Die b l aue C h l o r o f o r m l ö s u n g w i r d durch ein t r o c k e n e s P a p i e r f i l t e r in e inen k le inen Meßzyl inder abge lassen . D a z u w i r d aus e iner B ü r e t t e das gleiche Volumen 0,1-n. HCl g e f ü g t . De r I n h a l t des Meßzylin-de r s w i r d in e inen t rockenen Sche ide t r ich te r gebrach t , ge schü t t e l t u n d ge t r enn t , Die saure , ro te Pyocyan in -sa l z lö sung w i r d filtriert und im P u l f r i c h - P h o t o m e t e r gemessen . F i l t e r S 47, K ü v e t t e 1 cm, Verg le ichs-küvette mit 0,1-n. HCl. Die abgelesenen D-% werden mi t t e l s Tabe l l e in die E x t i n k t i o n s w e r t e E umgerech -net . A u s der E i c h k u r v e (Abb. 2) , die durch Messun-gen an P y o c y a n i n l ö s u n g e n b e k a n n t e r Konzen t r a t i on x e r m i t t e l t w u r d e , können d i r e k t die mg-% (mg in 100 cm3 L ö s u n g ) P y o c y a n i n abge lesen werden .
Präparative Gewinnung des Pyocyanins. Die K u l t u r -f lü s s igke i t w i r d im u n f i l t r i e r t e n Z u s t a n d mit 0,15%
Abb. 2. E i c h k u r v e zur E r m i t t l u n g des Pvocyanin-geh al tes.
Carboraff ' in v e r s e t z t und e in ige Min. g e r ü h r t . Das C a r b o r a f f i n w i r d anschl ießend a b z e n t r i f u g i e r t , wobei e ine Ü b e r l a u f z e n t r i f u g e g u t e D iens t e le is te t , und so-f o r t mit w a s s e r f r e i e m N a t r i u m s u l f a t g ründ l i ch ver -r ieben . Das so gewonnene t rockene P u l v e r w i r d in e inem P e r k o l a t o r mi t Chloroform-Methanol (im Mi-s c h u n g s v e r h ä l t n i s 90 :10 Vol . -Tei len) e lu ie r t , Das E l u a t wi rd mi t 0,1-n. HCl ausgeschü t t e l t , die salz-s a u r e P y o c y a n i n l ö s u n g a lka l i sch gemacht und mit C h l o r o f o r m e x t r a h i e r t . Die C h l o r o f o r m e x t r a k t e wer-den im Vak . e ingeeng t , und der r e s u l t i e r e n d e Rück-s tand w i r d a u s e ine r k le inen Menge W a s s e r u m k r i -s t a l l i s i e r t . Fe ine , s c h w a r z b l a u e mikroskop i sche Nadeln. Man e r h ä l t m i t t e l s d ieser Methode 86% der photo-met r i sch in der K u l t u r l ö s u n g e rmi t t e l t en Pvocyanin-Menge.
Isolierung eines gelben Farbstoffes a u s der K u l t u r -f l ü s s igke i t . Beim A u s s c h ü t t e l n der wie oben erha l -t enen C h l o r o f o r m l ö s u n g e n des Pyocyan in s mi t salz-s a u r e m W a s s e r geh t das P y o c y a n i n a ls s a l z sau re s Salz in die w ä ß r i g e Phase , w ä h r e n d die Chloroform-lösung s t e t s deut l ich gelb ble ibt und daher einen neu-
19 Z u r L a b o r a t o r i u m s m e t h o d i k der R ü h r v e r s u c h e vgl . K. B e r n h a u e r u. J . R a u c h , Biochem. Z. 319, 102 [1948] sowie H. K n ö 11, Dtsch . Gesundhei t s -wesen 1, 161 [1946]. — S. auch K. B e r n h a u e r , Ein-f ü h r u n g in die organisch-chemische L a b o r a t o r i u m s -technik , 5. Auf l . , S p r i n g e r - V e r l a g , Wien 1947, S. 42.

t ra len oder sauren Bestandtei l enthal ten muß. Der bei der Vera rbe i tung eines 24-i-Ansatzes in der be-schriebenen Weise erhaltene, vom Pyocyanin bef re i te , gelb gefä rb te Chloroformauszug wurde mehrmals mit Wasser gewaschen, über entwässer tem Na t r iumsu l fa t getrocknet und eingeengt . Es resu l t i e r te eine gelbe, tei lweise kr is ta l l i s ie r te Masse. Nach mehrmal igem Umkris ta l l i s ie ren aus Äthanol wurden gelbe, e twa 6 mm lange Nadeln erhal ten. Ausb. 228 mg. Schmp. 237.5—238° (teilw. Zers ) . Die Substanz ist schwer
löslich in Wasser , kal tem Äthanol und Äther , gu t lös-lich in heißem Äthanol . In s ta rken Mineralsäuren (z. B. Schwefe lsäure 1: 1) löst sie sich unter Erha l -t ung der gelben Farbe . In Alkalien ist sie löslich, aber un te r nur schwacher Gelbfärbung. Sie läßt sich mit tels Dimethy lsu l fa t s nicht methylieren.
C14H«02N2. Ber. C 71,18, H3,41, N 11,86. Gef. C 70,91, H 3,68. N 11,71.
Mol.-Gew. ber. 236.2. gef. 226 (nach Rast) .
Über Chalkone III* Die bakteriostatische Wirkung von 4'-substituierten 2-Oxy-5-brom-chalkonen
V o n E R N S T S C H R A U F S T Ä T T E R * * u n d S I G I S M U N D D E U T S C H
Aus dem Labora to r ium Dr. R. P f l e g e r , Er langen (Z. Naturforschg. 3 b. 430—132 [1948]; eingegangen am 3. September 1948)
Von dem 2-Oxy-5-brom-chalkon wurden mehre re Der iva te mit verschiedenen Sub-st i tuenten in 4 '-Stellung herges te l l t . Die Verb indungen zeigten fas t alle eine sehr gute bakter iosta t ische Wi rkung . Auch die als Ausgangsmate r ia l i en dienenden Acetophenone wurden ausgetes te t und ließen in ihrem Hemmvermögen gewisse Beziehungen zu den entsprechenden Chalkonen erkennen.
Aus unserer letzten Mitteilung ist zu ersehen, .daß dem leicht aus Acetophenon und Brom-
salicylaldehyd zugänglichen 2-Oxy-5-brom-chal-kon eine sehr gute bakteriostatische Wirkung zu-kommt. Es wurden deshalb einige Derivate dieser Verbindung hergestellt, und zwar interessierte uns zunächst, welchen Einfluß die E in führung verschiedener Gruppen in 4'-Stellung auf das antibakterielle Verhalten des 2-Oxy-5-brom-chal-kons ausüben würde. Als Austestungsmethodc diente die gleiche wie in der vorhergehenden Mit-teilung. und als Testorganismus wurde wiederum Staph. aureus benützt. Die Ergebnisse sind, um eine leichte Übersicht zu ermöglichen, in Tabellen zusammengestellt.
D i s k u s s i o n d e r E r g e b n i s s e
Wie aus Tab. 1 ersichtlich ist, übertraf ein großer Teil der neuen Verbindungen die ur-sprüngliche Stammsubstanz, das 2-Oxy-5-brom-chalkon. Sehr wirksam waren besonders die 4'-Halogenderivate, von denen wiederum die Brom-verbindung die Spitzenstellung einnahm. Es ist interessant, daß die Einführung von Brom in das 2-Oxy-5-brom-chalkon in die 4'-Stellung, also in
* II . Mitt.: Z. Natur forschg . 3b, 163 [1948]. ** Er langen, Schuhst raße 19.
den noch unbesetzten Benzolkern, eine stärkere Wirkungssteigerung hervorruft als die Substitu-tion mit Brom in der 3-Stellung. In diesem Zu-sammenhang wräre zu erwähnen, daß eine ähn-liche Erscheinung, nämlich die Wirkungserhö-hung durch Halogensubstitution in 4'-Stellung, bereits von E. K l a r m a n n und J. v. W o w e r n 1
in der Diphenylmethanreihe beobachtet wTorden ist. So erwies sich das 4'-Chlor- bzw. 4'-Brom-2.4-dioxy-diphenylmethan dem 2.4-Dioxy-diphenyl-methan um etwa das 8—4-fache überlegen. Auch E. K1 a r m a n n , L. G a t e s und V. S h t e r n o v 2
konnten durch Einführung von Chlor oder Brom in 4'-Stellung bei 4-Oxy-3-chlor-diphenylmethan die Wi rkung verdoppeln bzw. verdreifachen.
Die Substitution mit einer Hydroxylgruppe in 4'-Stellung führte zu einem geringen Abfall des Hemmeffektes. Durch Verätherung dieser Hydroxylgruppe wurden aber wieder die gleichen antibakteriellen Eigenschaften wie bei der un-substituierten Verbindung erreicht. Einen ver-stärkenden Einfluß auf das Hemmvermögen zeig-ten die Thioverbindungen, wobei sich die Sulf-
1 E. K l a r m a n n u. -J. v. W o w e r n , J . Amer. chem. Soc. 51, 605 [1929].
2 E. K 1 a r m a n n , L. G a t e s u. V. S l i t e r n o v , J . Amer. chem. Soc. 54. 3315 [1932],