vital statistics of triatominae (hemiptera - ecopaedia
TRANSCRIPT

J. Med. Ent. Vol. 9, no. 4: 351-370 1 August 1972
VITAL STATISTICS OF TRIATOMINAE (HEMIPTERA: REDUVIIDAE) UNDER LABORATORY CONDITIONS
������������� ������������
By Jorge E. Rabinovich�
Abstract: A statistical evaluation of the population dynamics of T. infestans is based on cohort experiments conducted under controlled laboratory conditions. Egg hatching, nymphal developmental time and mortality, adult male and female longevity and mortality, and female fecundity were computed, and results compared to information in the existing literature; in most cases there is good agreement, and large deviations can be attributed to differences in experimental conditions, particularly temperature, feeding and density. In addition to previously studied characteristics of the life cycle, the following population parameters were evaluated, and their numerical values interpreted both in terms of population dynamics and control approaches: life expectancy, intrinsic rate of natural increase, generation time, reproductive value, finite rate of increase, and stable age distribution. The impor-tance of age distributions and dispersal in designing control strategies is pointed out. Finally the population characteristics and adaptation of T. infestans to its man-modified environment lead to the conclusion that this species is a "K-strategist", existing at or near saturation levels in a stable environment.
This is the first of a series of papers dealing with the demographic features (life cycle, reproduction, and mortality) and population parameters of several reduviid species that are vectors of Chagas' Disease in the Neotropical region. The goals of this series are twofold: (a) to give statistical information about populations kept under controlled laboratory conditions, and (b) to compare closely related species in terms of their biologicar characteristics. The former will fill a gap in the existing literature, providing statistical life tables and variations in population responses that have rarely, if ever, been published for reduviids. The latter will provide knowledge about relative population growth potential for different species, information that is particularly important when 2 species coexist in a given habitat. Also, this type of biodemographic analysis, when coupled with field studies of the population dynamics, provides critical information for the design of optimal control strategies.
Triatoma infestans ranges over a considerable part of South America, south of 12º S Lat., al- though it is restricted to semiarid regions (Usinger et al. 1966). It is found in the coastal area of
1The first part of the experimental work of this investigation was carried out while the author was a researcher at the Instituto de Zoología Tropical, Universidad Central de Venezuela.
2Instituto Venezolano de Investigaciones Cientificas, Depart-ment of Ecology, Apartado 1827, Caracas, Venezuela.
southern Peru, in the eastern and southern parts of Bolivia, in Brazil from Bahia southward, in the coastal area of central and northern Chile, over all of Paraguay, in the central and northern regions of Argentina, and in northwestern Uruguay (Romaña 1963). Neiva (1913) refers to a personal communication by Dr Sisto Alberto Padilla indicating that T. infestans was found in El Salvador, but this has not been confirmed by later investigators. Thus species is the most important vector of Chagas' Disease within its geographical range. Perlowagora-Szumlewicz (1953) states that 96.3 to 98.6% of all triatomine bugs found in the State of Sao Paulo, Brazil, are T. infestans; Borda (1972) attributes 100% of Chagas' Disease trasmission within houses and 99% of peridomiciliary transmission to this species in eastern and southeastern Bolivia. Abalos & Wygodzinsky (1951) and Usinger et al. (1966) believe that the present status of T. infestans is that of an exclusively domestic and peridomestic species, for no reliable reference indicates its presence far from permanent human dwellings. Since Chagas' (1909) discovery of Schizotrypanum cruzi in the intestinal contents of Panstrongylus megistus, a huge literature has been published on the biology, ecology, and epidemiology of many species of triatomine vectors of this disease. The papers about T. infestans are particularly numerous, starting with the 1st biological account by Neiva (1913). This was followed by many field and laboratory studies as those, among others, of Maggio & Rosenbusch (1915), Dias (1938, 19555, b), Dias & Zeledón (1955), Pessoa & Barros (1939), Nicolle & Lwoff (1942), Abalos & Wygodzinsky (1951), Ryckman (1951, 1952), Pellegrino (1952), Correa (1954), Correa & Schiavi (1954), Perlowagora-Szumlewicz (1953, 1954, 1969), Wiesinger (1955), Hack (1955), Pessoa (1959), Joërg (1962), Wood (1964), Pinto Dias (1965), Franca Rodriguez & Ceruzzi-Romeo (1965), Neghme et al. (1967), Danilov (1968), and Borda (1972). With a few exceptions (Pessoa & Barros 1939, Joërg 1962, Perlowagora-Szumlewicz 1969) the papers mentioned refer to studies with T. infestans
352 J. Med. Ent. Vol. 9, no. 4
that were either made under laboratory conditions subject to local climatic changes or with a number of individuals too small for any statistical treatment of the results. This paper presents the results of an experiment designed to obtain not only the numerical value of demographic statistics under controlled constant environmental conditions, but also their biological variability.
MATERIALS AND METHODS
The population of Triatoma infestans used in this study was made available by J. C. Gómez-Nuñez, of the Venezuelan Ministerio de Sanidad y Asistencia Social; it originated in Chile and had been reared in the laboratory for about 2 years.
The experiment was carried out in a climatic room with constant conditions of temperature (26 +- PC) and humidity (60 ± 10% RH). Photoperiod did not need to be controlled, for at the latitude of Caracas there is a very small change between seasons. The experimental design involved cohort populations initiated at the egg stage and replicated 5 times. Each replicate was started with 100 recently laid (0-48-hr-old) eggs; these were kept in 150 cc glass containers until all viable eggs hatched. After hatching the 1st instar nymphs were transferred to 3.785-liter jars, covered with nylon mesh, and with vertically placed strips of paper inside that served both as resting places and for climbing to the top at feeding time.
Each cohort was fed weekly using hens placed on a special wooden box with holes at the bottom, through which the tops of the cohort jars could be inserted. The T. infestans climbed to the top and fed through the nylon mesh. Food was offered
for 1 hr, and during the following hour the jars were horizontally exposed to a high speed fan. Use of the fan avoided the accumulation of excessive moisture that is very common after the insects become engorged while in glass containers.
After being exposed to the fan, each jar was opened in order to check the number of dead individuals (identified by their nymphal instar, and by sex if adults), and to count the number of eggs laid. In some instances the weekly schedule of feeding and counting could not be maintained, and in those cases the recorded information was subjected to linear interpolation to keep the week as the time unit for life tables and for calculation of population statistics.
The weekly death schedule of a cohort initiated at the egg stage provided the necessary information to construct a complete "horizontal" life table (Deevey 1947). To carry out the calculations the method of Dublin et al. (1949) was followed; definitions of the components of a life table, with their formulae, are given in the Appendix.
The same mortality schedule, coupled with female age-specific fecundity, was used to calculate such statistics as the intrinsic rate of natural increase, the net reproductive rate, the instantaneous birth and death rate (Birch 1948), and the age specific reproductive value (Fisher 1930). In addition, the stable age distribution was calculated. All definitions and formulae are given in the Appendix. Some calculations were carried out with the 360/50 IBM System of the School of Sciences, Universidad Central de Venezuela, and others with the 1130 IBM computer of the Instituto Venezolano de Investigaciones Científicas.

1972 Rabinovich: Vital statistics of Triatominae 353
RESULTS
Life cycle In TABLE 1 a statistical summary of life-cycle
statistics is given. Average time from oviposition to hatching was about 3 weeks. Adult longevity was calculated simply from the frequency table of the number of weeks lived by each individual; it is expressed for the total population and for each sex separately. On the average, after entering the adult stage, males lived 1.59 times longer than females. Maximum longevity was also greater for males than for females, by a factor of 1.8; the most extreme case was that of a male (in cohort N ° 3) that lived for 72.5 weeks as an imago and a total of 630 days (~1.9 years) from oviposition. A similar relationship between males and females was obtained by analyzing the life expectancy after they had molted to adults; males had a life expectancy 1.61 times greater than females.
As each member of the cohort was not followed individually, a frequency table of the time in instar was used to provide an estimate of the duration of each nymphal instar. The tables were based on time of and number of individuals (a) entering a particular instar, (b) dying in that instar, and (c) molting to the following instar. The results, also given in TABLE 1, show that the 1 st and 2nd nymphal instar took about the same time (~1 month) to complete their development (although the develop-mental time was much more variable in the 1st nymphal instar than in the 2nd); that the 3rd nymphal instar was about 1/2 that long, and that the 4th nymphal instar was completed, on the average, in only 11 days. However, this diminish-
ing trend in developmental time was not maintained in the last nymphal instar; the 5th instar was longest (about 49 days) and at the same time, the least variable. By adding the average developmental time of all nymphal instars, a total preadult period of 141 days (~20.1 weeks) is obtained. Mortality
Mortality information is given in 2 different ways; as an age specific schedule of mortality, and as mortality by developmental stage.
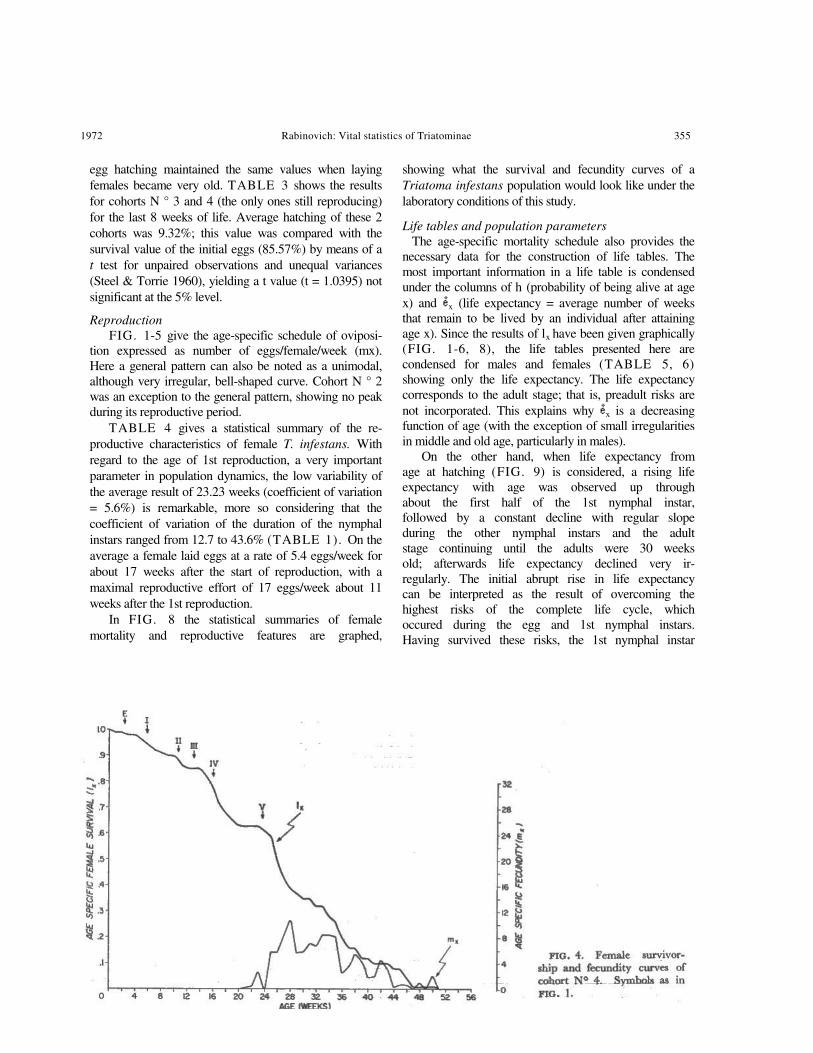
FIG. 1-5 show, for each cohort, female age specific mortality expressed as lx, i.e., the probability of being alive at age x. There are some differences between cohorts. Cohort N° 2 showed the largest deviation from the general pattern: in the 1st nymphal instar survival dropped abruptly to about 40%, and remained static for the rest of the nymphal instars. Cohorts N° 1, 3, 4, and 5 showed the general pattern: a relatively steep slope up to the 1st nymphal instar, a very gentle decrease during the 2nd, 3rd, and 4th nymphal instars, and another abrupt drop during the 5th nymphal instar. A remarkably constant feature for almost all cohorts was that after becoming imagoes, survival remained stable for a few weeks, as if with the last molt a severe mortality risk had been passed. After those few weeks of constant survival values, adult females showed a fairly linear age-specific schedule of mortality, with the exception of cohort N° 3, which had no mortality from week 30 through week 40.
The male age-specific mortality is shown for all cohorts in FIG. 6, where it can be observed that, with the exception of cohort N° 3 which had a

consistently higher survival probability, all cohorts had an almost identical pattern of mortality.
TABLE 2 shows the mortality pattern in terms of developmental stages. Mortality at the egg stage was fairly low (about 15% of all eggs fail to hatch), but more than 1/2 of all nymphs died before reaching the adult stage.
Thus total preadult mortality can be broken down by nymphal instars in 2 different ways, as relative or absolute mortality. In both cases they followed the same pattern, with the heaviest mortality in the 1st nymphal instar, followed by the 5th instar, nymphs in instars 2, 3, and 4 having a much lighter toll and the 3rd nymphal instar having the lowest mortality.
The sex ratio was not 1: 1 when entering the adult stage; females were favored, on the average, by a proportion of 1.27 to 1.00; this suggests that males have slightly higher mortality risks during development than females.
It was of interest to determine if nymphal mortality was in some way related to developmental time. FIG. 7 shows no linear relationship between absolute nymphal mortality and developmental time (r = 0.230, not statistically significant). There seems to be a slight non-linear relationship in FIG. 7 ; however, due to the completely out-oftrend position of point 12, the non-linear fit was not considered worthy of further analysis.
It was also of interest to determine if percentage
354 J. Med. Ent. Vol. 9, no. 4
1972 Rabinovich: Vital statistics of Triatominae 355
egg hatching maintained the same values when laying females became very old. TABLE 3 shows the results for cohorts N ° 3 and 4 (the only ones still reproducing) for the last 8 weeks of life. Average hatching of these 2 cohorts was 9.32%; this value was compared with the survival value of the initial eggs (85.57%) by means of a t test for unpaired observations and unequal variances (Steel & Torrie 1960), yielding a t value (t = 1.0395) not significant at the 5% level.
Reproduction FIG. 1-5 give the age-specific schedule of oviposi-
tion expressed as number of eggs/female/week (mx). Here a general pattern can also be noted as a unimodal, although very irregular, bell-shaped curve. Cohort N ° 2 was an exception to the general pattern, showing no peak during its reproductive period.
TABLE 4 gives a statistical summary of the re-productive characteristics of female T. infestans. With regard to the age of 1st reproduction, a very important parameter in population dynamics, the low variability of the average result of 23.23 weeks (coefficient of variation = 5.6%) is remarkable, more so considering that the coefficient of variation of the duration of the nymphal instars ranged from 12.7 to 43.6% (TABLE 1). On the average a female laid eggs at a rate of 5.4 eggs/week for about 17 weeks after the start of reproduction, with a maximal reproductive effort of 17 eggs/week about 11 weeks after the 1st reproduction.
In FIG. 8 the statistical summaries of female mortality and reproductive features are graphed,
showing what the survival and fecundity curves of a Triatoma infestans population would look like under the laboratory conditions of this study.
Life tables and population parameters The age-specific mortality schedule also provides the
necessary data for the construction of life tables. The most important information in a life table is condensed under the columns of h (probability of being alive at age x) and x (life expectancy = average number of weeks that remain to be lived by an individual after attaining age x). Since the results of lx have been given graphically (FIG. 1-6, 8), the life tables presented here are condensed for males and females (TABLE 5, 6) showing only the life expectancy. The life expectancy corresponds to the adult stage; that is, preadult risks are not incorporated. This explains why x is a decreasing function of age (with the exception of small irregularities in middle and old age, particularly in males).
On the other hand, when life expectancy from age at hatching (FIG. 9) is considered, a rising life expectancy with age was observed up through about the first half of the 1st nymphal instar, followed by a constant decline with regular slope during the other nymphal instars and the adult stage continuing until the adults were 30 weeks old; afterwards life expectancy declined very ir- regularly. The initial abrupt rise in life expectancy can be interpreted as the result of overcoming the highest risks of the complete life cycle, which occured during the egg and 1st nymphal instars. Having survived these risks, the 1st nymphal instar

attained the highest life expectancy. This interpretation can be checked against TABLE 1 and 2 which show that the highest nymphal mortality occurred in the 1st nymphal instar (41.6%), and that this mortality took place during a short period of 31.2 days (only about 30% of the whole nymphal developmental period).
TABLE 7 summarizes the statistics of population characteristics. The average generation time (=mean length of a generation; see Appendix) is 30.9 weeks (~216 days). The average net reproduction rate (R0) is 25.04. The former means that successive generations will be spaced 31 weeks apart, and the latter, ratio of total female births
in 2 successive generations (Lotka 1945), indicates that a T. infestans population (under the laboratory conditions used here) will increase 25-fold during each generation.
The average intrinsic rate of natural increase, also called the Malthusian parameter, was 0.101 per individual per week, and it represents the difference between the instantaneous birth and the instantaneous death rates. However, in many circumstances it is more useful to know the finite rate of increase (λ), that is, the number of times a T. infestans population will multiply per week, and here the average value of λ was 1.107.
Any statement about rates of population increase,
356 J. Med. Ent. Vol. 9, no. 4

1972 Rabinovich: Vital statistics of Triatominae 357
either considering r or λ, is incomplete without reference to the age distribution of the population (Birch 1948). When the stable age distribution is calculated (TABLE 8), it is seen that, with no changes in the mortality and fecundity schedules and an unlimited environment, a T. infestans population will gradually assume a stable age distribution composed of 38.66% eggs, 58.56% nymphs, and only 2.79% adults.
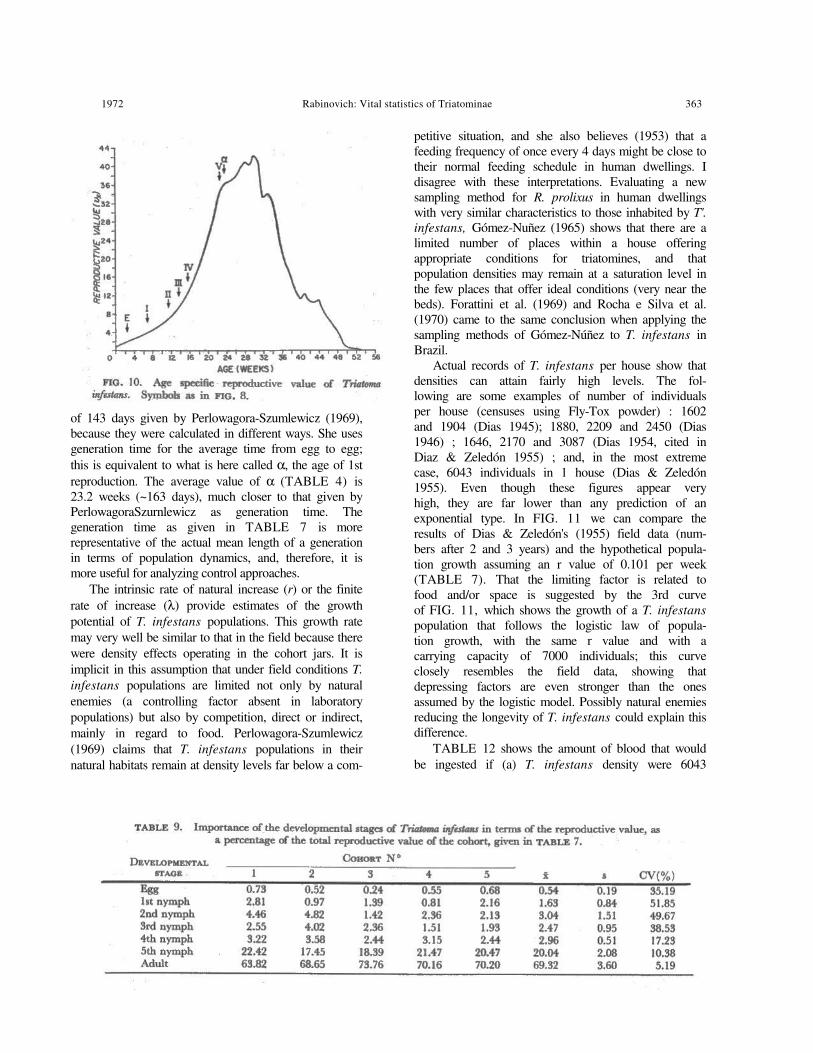
The reproductive value of the population was calculated (TABLE 7). This characteristic is more valuable when it is analyzed by age classes. FIG. 10 shows that the reproductive value changes with age and follows a typical bell-shaped curve with its peak occurring a few weeks after the start of the reproductive period. TABLE 9 shows, for each cohort, the reproductive values grouped by developmental stages; these figures indicate how much each developmental stage or instar is worth in terms of offspring it will contribute to the next generation (Wilson & Bossert 1971). With the exception of the 2nd nymphal instar, which has a reproductive value larger than the preceding instar but smaller than the succeeding one, there is a trend of increasing reproductive value as the individuals advance in their development.
TABLE 9 gives only a relative measure of the im-portance of each developmental stage in terms of their reproductive value; the actual value of each stage depends on the population's age composition. TABLE 10 was prepared to show the same information as TABLE 9, but weighting it under the asumption that the population has achieved a stable age distribution; this provides a very different picture. Although adults have a higher reproductive value,
they are not as important as the 5th instar nymphs because of the small number in a population with stabilized age distribution. First and 2nd nymphs follow in importance for the same reason. Finally eggs are more important than the 3rd and 4th nymphal instar individuals, not because of their intrinsic reproductive value, but because of the high proportion of eggs found in a population with stable age distribution (See TABLE 8).
DISCUSSION
It is difficult to compare the results of this study with others that have been published, because most of the previous work with T. infestans was carried out under a variety of laboratory conditions. Temperature and humidity (usually uncontrolled), size of jars, feeding frequency and duration, density of insects in each jar, species of host offered for feeding, and other factors rarely coincide in different investigations. Of the various papers based on laboratory experimentation similar to that of the present work, Perlowagora-Szumlewicz' (1969) is the most appropriate for comparison because (a) temperature, although uncontrolled, was recorded and many results are grouped for given temperature intervals; (b) jars were only slightly larger than the ones used in this experiment; (c) feeding was carried out at the same frequency and in a similar fashion; and (d) large numbers of insects were analyzed and the degree of variability for most results is indicated.
In evaluating the results, it is important that the following aspects of this cohort study be taken into account:
(1) Histories of individual insects are not known.

Because of the amount of labor that individual analysis would have involved, each cohort was followed as a group; thus possible interactions between individuals of a given cohort are not known. For example, Ryckman (1951) notes that in Triatoma phyllosoma pallidipennis, older nymphs crowd up to the host to the exclusion of younger members of the colony. Perlowagora-Szumlewicz (1953) shows the importance of the time of the 1st meal, and in general it is known (Goodchild 1955, Danilov 1968, Perlowagora-Szumlewicz 1969) that the volume of the blood meal in one or successive feedings drastically affects the process of molting and of production and viability of eggs. This interference, if it also occurred with T. infestans, may have greatly influenced the results of this cohort study. Also, the considerable amount of warm blood ingested by older nymphs stimulates
a probably thermotrophic response in unengorged younger nymphs; this elicits a probing reaction that leads to a type of cannibalism called "klepto-hemodeipnonism" by Ryckman (1951). This reaction has been reported in T. infestans, and it is not known to what degree, if at all, it might have affected these experiments. Another disad- vantage of treating the cohort as a group is that the reproductive performance cannot be evaluated individually; some information is lost, such as individual periodicity in oviposition.
(2) Density varies with time. As each cohort becomes older and individuals die, the density (number of individuals per jar) decreases. Perlowagora-Szumlewicz (1969) demonstrates an apparent density effect upon longevity and fecundity.
(3) Calendar age does not equal physiological age. As a result of interactions referred to above as well as normal biological variability, individuals of T. infestans do not molt synchronously even if they develop from eggs laid simultaneously. Consequently as the insects develop into more advanced stages there is a larger amount of overlap between stages, and individuals with the same calendar age may belong to, say, the 3rd, 4th, or 5th nymphal instar. It is not known if these differences affected the results of this study. For example, the prob-ability of dying may differ between a 1st instar nymph 25 days old and a 1st instar nymph 40 days old.
With these 3 reservations in mind, however, most of the results are comparable to those found in the literature. A discussion of some particular aspects of the results of this cohort study follows.
Duration of egg stage. The average incubation period, 19.8 ± 2.7 days is in line with 20-24 days at 25ºC and 11-13 days at 33ºC given by Hack (1955), and 16-33 days (but most commonly 20-25 days) by Neiva (1913). Dias (1955b) claims that an observed incubation of 22.5 days was a result of the climatic conditions of his Rio de Janeiro
358 J. Med. Ent. Vol. 9, no. 4

1972 Rabinovich: Vital statistics of Triatominae 359
laboratory. Borda (1972) reports a fairly long egg developmental time of 35 days, probably due to the relatively low ambient temperatures of Cochabamba, Bolivia. One of the largest figures for egg development found in the literature is 46 days (Hack 1955). The results of ParlowagoraSzumlewicz (1969) are the most consistent with those presented here; she reports an average hatching time of 20.2 ± 2.4 days (obtained from analyzing a total of J78,689 eggs, under the climatic conditions of a laboratory in Rio de Janeiro) and of 18.9 ± 1.6 days when grouping all those eggs subjected to temperatures of 25-27ºC. (These figures, given as mean ± 1 standard deviation, were calculated by me from Perlowagora-Szumlewicz' data)
Egg-hatching percentage. The average hatch for all cohorts, 85.6 ± 8.4%, is very close to the 88% given by Perlowagora-Szumlewicz (1969). Hack (1955) observed a similar percentage of 82% at 25ºC, but a much lower value at 33ºC; however, Perlowagora-Szumlewicz (1969) does not detect any effect of temperature upon hatching between 18 and 28°C. Borda's (1972) observation of 85% is also consistent with the results of this cohort experiment. The information provided here in- dicates that survival during embryonic development
is not affected by the age of the female parent. Nymphal developmental time. The total duration
of the nymphal stage is 141 days, much longer than the 102.4 ± 10.9 days of Perlowagora-Szum-lewicz (1969) (mean and 1 standard deviation, calculated by me from Table 3 in Perlowagora-Szumlewicz, based on 1328 males and females). There is an even greater difference with the value of 76.5 days given earlier by Perlowagora-Szum- lewicz (J953). The figure is almost identical with the 143 days given by Dias (1955b) ; however, this very close similarity may well be coincidental, because the distribution of developmental time by nymphal instars is completely different. (See TABLE 11, where developmental time of each nymphal instar, expressed as a percentage of the total time required from hatching to imago, is compared for all sources known.) The most conspicuous difference in relative duration of nymphal instars is the inversion that occurs in the 5th nymph; this is the briefest instar in 1 case (Dias 1955b) and the longest in others (Perlowagora-Szumlewicz 1953, present study). However, Dias' experiment was carried out with only 2 individuals, kept separately and with assured weekly feeding unlike the present study.

The published developmental time from hatching to imago of 220-240 days (Neiva 1913), about 300 days (Pessoa & Barros 1939), and about 1.5 years (from egg to adult) (Maggio & Rosembush 1915) are not comparable with the values presented here because of large differences in environmental conditions and feeding schedules. For example, it is not known how frequency of feeding affects nymphal developmental time in T. infestans; lightly-fed 4th-instar nymphs of Rhodnius prolixus molt to the 5th instar more rapidly than completely engorged bugs (Bennet-Clark 1966).
There is no reference in the literature to diapause in T. infestans, with the exception of Joërg (1962) who suggests that some specimens (reared at 32ºC and 52-78% RH) that remained as 5th instar nymphs might be in diapause.
Nymphal mortality. This cohort study yielded a total nymphal mortality of 54.6 ± 7.2%, a figure close to the 60% given by Borda (1972) but much
larger than the 25% and 5.3% of Perlowagora-Szumlewicz (1953), 39% of Perlowagora-Szum- lewicz (1969), and 24.5% of Joërg (1962). The differences again can be attributed to differences in feeding schedules and temperature conditions. The importance of feeding in nymphal mortality is shown by the contrasting results in 2 of Perlowagora-Szumlewicz' (1953, 1969) experiments, when bugs were fed every 4 days, mortality was 25 and 5.3%, but it increased to 39% with 7-day feeding intervals. The importance of temperature is shown by the results from experiments with controlled temperature carried out by Joërg (1962); a statistically significant negative linear correlation (r = -0.837, P<0.01) between temperature and percentage nymphal mortality was found [correla-tion and regression analyses were performed by me using data given in Fig. 1 of Joërg's (1962) paper]. The regression equation that shows the relationship between temperature and nymphal mortality (Y =
360 J. Med. Ent. Vol. 9, no. 4

1972 Rabinovich: Vital statistics of Triatominae 361
100.09 - 2.91 X) predicts a 24.33% mortality at 26°C; this value is much smaller than the 54.6% obtained with the cohort populations and indicates the importance of factor interaction (feeding and temperature in this case).
When gross nymphal mortality is broken down into specific nymphal instar mortality, there is a heavy toll (19.7%) in the 1st instar (TABLE 2) not found by Perlowagora-Szumlewicz (1953, 1969). Since the number of insects per jar was still relatively high during this instar, the result might be at- tributed to some kind of density effect. Mortality in the 5th nymphal instar (16.3%) is also high in
comparison with 7.5% (for group 1) and 5.3% (for group 2) given by Perlowagora-Szumlewicz (1953) and 6% given by Borda (1972).
Adult longevity. Males had greater average lon-gevity (182.4 days) than females (114.8 days). These values are much smaller than 319.5 days (for females) obtained by Perlowagora-Szumlewicz (1969) and 390 days by Borda (1972). Again the difference may be a function of either temperature or density effects. The former is clearly shown in Hack's (1955) experiments where adult longevity of 146 days at 25 °C drops to 66 days at 33ºC. Assuming that males and females are

present in equal number, the average for adult longevity in this cohort study was 148 days, almost the same as Hack's (1955) results at 25ºC (a temperature very close to that of the cohort jars). The effect of density is demonstrated by Perlowagora-Szumlewicz (1969); female longevity of 319.5 days with bugs held individually dropped to 114.4 days when they were maintained at a density of 10 females per jar. In the cohort experiments the density decreased continuously, but the average number of individuals/jar/week was 13.1; this density is fairly close to that in the experiment of Perlowagora-Szumlewicz (1969) that resulted in a longevity of 114.4 days, almost identical to the 114.8 days obtained for females in the cohort jars.
Fecundity. Females in cohort jars began repro-duction an average 21.7 days after becoming adults; this value is intermediate between the 15.2 days given by Perlowagora-Szumlewicz (1969) and about 30 days given by Neiva (1913). Variation in the number of eggs/female/week, although very irregular, apparently followed a pattern of increasing and then decreasing with the age of the female. Maximum reproductive effort occurred at an average age of 33.8 weeks, and about 10.6 weeks after the initiation of reproduction. PerlowagoraSzumlewicz (1969, see her Table 9) does not recognize any significant change in reproduction as a function of female age, even though her data show variation of nearly 100% in the number of eggs/female/week with age.
The average weekly reproduction in cohort jars (5.4 eggs/female/week) was lower than the 9.8
Eggs/female/week at 25ºC given by Hack (1955) and 20.2 ± 4.6 eggs/female/week by PerlowagoraSzumlewicz (1969) (mean ± 1 standard deviation, calculated by me from her Table 9).
Total number of eggs per female was 149.1 91.1; Hack's (1955) figures are 209 and 99 eggs at 25 and 33ºC respectively; assuming a linear relationship between temperature and fecundity, Hack's results indicate a total of 195 eggs/female at 26°C. The results of Perlowagora-Szumlewicz (1969) demonstrate the influence of density, production of 649.4 eggs per individually maintained female dropped to 54.2 eggs when 20-39 females were maintained together. Perlowagora-Szumlewicz (1969) shows that several other factors are important in the reproductive performance of T. infestans: female age at time of pairing, duration of exposure to males, and feeding frequency. These factors were not evaluated in the cohort jars, but it can be assumed that they also caused differences in fecundity between the values given here and those obtained when insects were reared individually and with controlled mating and feeding schedules. Some other factors also should be investigated in T. infestans; for example, it has been demonstrated in R. prolixus that egg retention is followed by a smaller meal in subsequent feedings (Davey 1967) and that sperm are able to fertilize ova up to 8 weeks after copulation (Trejos et al. 1966).
Population parameters and control strategies. Generation time (=mean length of a generation) was 30.9 weeks (~216 days). This value should not be compared with the much shorter generation time
362 J. Med. Ent. Vol. 9, no. 4
1972 Rabinovich: Vital statistics of Triatominae 363
petitive situation, and she also believes (1953) that a feeding frequency of once every 4 days might be close to their normal feeding schedule in human dwellings. I disagree with these interpretations. Evaluating a new sampling method for R. prolixus in human dwellings with very similar characteristics to those inhabited by T'. infestans, Gómez-Nuñez (1965) shows that there are a limited number of places within a house offering appropriate conditions for triatomines, and that population densities may remain at a saturation level in the few places that offer ideal conditions (very near the beds). Forattini et al. (1969) and Rocha e Silva et al. (1970) came to the same conclusion when applying the sampling methods of Gómez-Núñez to T. infestans in Brazil.
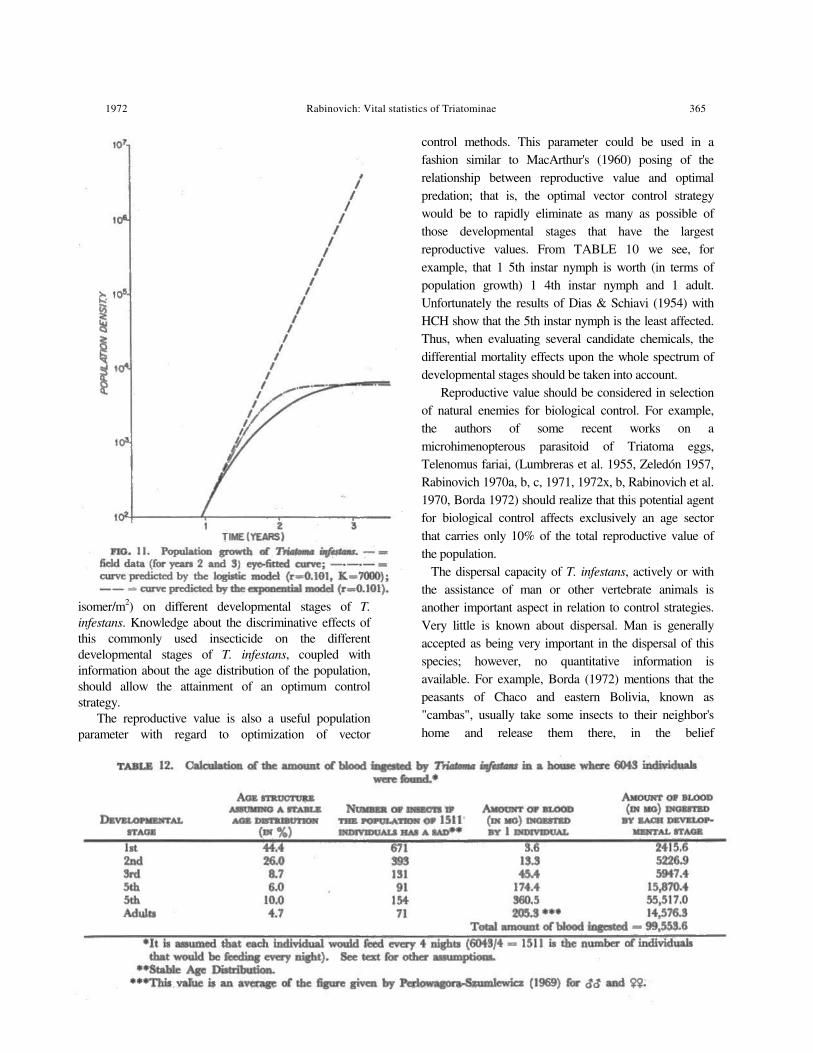
Actual records of T. infestans per house show that densities can attain fairly high levels. The fol- lowing are some examples of number of individuals per house (censuses using Fly-Tox powder) : 1602 and 1904 (Dias 1945); 1880, 2209 and 2450 (Dias 1946) ; 1646, 2170 and 3087 (Dias 1954, cited in Diaz & Zeledón 1955) ; and, in the most extreme case, 6043 individuals in 1 house (Dias & Zeledón 1955). Even though these figures appear very high, they are far lower than any prediction of an exponential type. In FIG. 11 we can compare the results of Dias & Zeledón's (1955) field data (num- bers after 2 and 3 years) and the hypothetical popula- tion growth assuming an r value of 0.101 per week (TABLE 7). That the limiting factor is related to food and/or space is suggested by the 3rd curve of FIG. 11, which shows the growth of a T. infestans population that follows the logistic law of popula- tion growth, with the same r value and with a carrying capacity of 7000 individuals; this curve closely resembles the field data, showing that depressing factors are even stronger than the ones assumed by the logistic model. Possibly natural enemies reducing the longevity of T. infestans could explain this difference.
TABLE 12 shows the amount of blood that would be ingested if (a) T. infestans density were 6043
of 143 days given by Perlowagora-Szumlewicz (1969), because they were calculated in different ways. She uses generation time for the average time from egg to egg; this is equivalent to what is here called α, the age of 1st reproduction. The average value of α (TABLE 4) is 23.2 weeks (~163 days), much closer to that given by PerlowagoraSzurnlewicz as generation time. The generation time as given in TABLE 7 is more representative of the actual mean length of a generation in terms of population dynamics, and, therefore, it is more useful for analyzing control approaches.
The intrinsic rate of natural increase (r) or the finite rate of increase (λ) provide estimates of the growth potential of T. infestans populations. This growth rate may very well be similar to that in the field because there were density effects operating in the cohort jars. It is implicit in this assumption that under field conditions T. infestans populations are limited not only by natural enemies (a controlling factor absent in laboratory populations) but also by competition, direct or indirect, mainly in regard to food. Perlowagora-Szumlewicz (1969) claims that T. infestans populations in their natural habitats remain at density levels far below a com-

individuals, as reported by Dias & Zeledón (1955), (b) every individual fed once every 4 days, as assumed by Perlowagora-Szumlewicz (1953), (c) feeding rates by stages are as given by PerlowagoraSzumlewicz (1969), (d) stable age distribution is as given in TABLE 8 (adjusted to exclude the nonfeeding egg stage), and (e) human occupants provide the only source of food (which is not strictly true). It can be seen that about 100 cc of blood would be ingested each night, or an average blood amount of about 17 cc/night/person in the house described by Dias & Zeledón (1955) that had 6 occupants. It is very doubtful that a family could accept this amount of bleeding, and even less other effects of having 252 bugs biting each person every night. Thus, although there is no direct evidence, it is almost certain that competition, or at least interference, is a factor limiting population growth of T. infestans in human dwellings. Of course, the example used is an extremely high infestation, but the fact that in most houses T. infestans populations usually remain at a much lower level reinforces the hypothesis of density-dependence (involving intraspecific competition) and/or natural enemies as factors in population regulation.
The analysis of the stable age distribution shows some parallels with the little available field in- formation. One of the most extensive censuses it that reported by Dias (1955a), refering to a period of 7 years and giving percentages of adults and nymphs month by month. In general, the results show a relative abundance of adults much
larger than expected for a stable age distribution (between 23 and 36%, as against 4.7%; see TABLE 12 for percentage of adults in a stable age distribution adjusted to ignore the egg stage). However, although a natural population rarely attains a stable age distribution, in this particular case 2 methodological factors of the field data contribute to enlargement of differences: (a) the field information is not the result of a designed sampling program (some houses may have been sampled only once, other several times; data from houses in different stages of "colonization" have been mixed), and (b) the younger and smaller nymphs are more difficult to see and collect, thus inflating the proportion of larger nymphs and adults taken.
The results of Dias & Zeledón (1955) show a better agreement with a stable age distribution. The 1st census 2 years after the house was built and previous to any collecting or insecticide treatment, yields the following figures (for field and stable age distribution, in percent, respectively) 1 st to 4th nymphs = 74.8 and 85.1, 5th nymphs = 15.3 and 10.2, and adults = 9.9 and 4.7. Con-sidering the above-mentioned factor of underestimating small nymphs in field collections, the agreement can be considered quite satisfactory.
The age structure of the population has usually not been given sufficient emphasis in analyzing control strategies and control evaluation. In this respect the work of Correa & Schiavi (1954) is very important, because it provides results of the effect of HCH (hexachlorocyclohexane) (0.5 γ-
364 J. Med. Ent. Vol. 9, no. 4
isomer/m2) on different developmental stages of T. infestans. Knowledge about the discriminative effects of this commonly used insecticide on the different developmental stages of T. infestans, coupled with information about the age distribution of the population, should allow the attainment of an optimum control strategy.
The reproductive value is also a useful population parameter with regard to optimization of vector
control methods. This parameter could be used in a fashion similar to MacArthur's (1960) posing of the relationship between reproductive value and optimal predation; that is, the optimal vector control strategy would be to rapidly eliminate as many as possible of those developmental stages that have the largest reproductive values. From TABLE 10 we see, for example, that 1 5th instar nymph is worth (in terms of population growth) 1 4th instar nymph and 1 adult. Unfortunately the results of Dias & Schiavi (1954) with HCH show that the 5th instar nymph is the least affected. Thus, when evaluating several candidate chemicals, the differential mortality effects upon the whole spectrum of developmental stages should be taken into account.
Reproductive value should be considered in selection of natural enemies for biological control. For example, the authors of some recent works on a microhimenopterous parasitoid of Triatoma eggs, Telenomus fariai, (Lumbreras et al. 1955, Zeledón 1957, Rabinovich 1970a, b, c, 1971, 1972x, b, Rabinovich et al. 1970, Borda 1972) should realize that this potential agent for biological control affects exclusively an age sector that carries only 10% of the total reproductive value of the population.
The dispersal capacity of T. infestans, actively or with the assistance of man or other vertebrate animals is another important aspect in relation to control strategies. Very little is known about dispersal. Man is generally accepted as being very important in the dispersal of this species; however, no quantitative information is available. For example, Borda (1972) mentions that the peasants of Chaco and eastern Bolivia, known as "cambas", usually take some insects to their neighbor's home and release them there, in the belief
1972 Rabinovich: Vital statistics of Triatominae 365

366 J. Med. Ent. Vol. 9, no. 4
that the rest of the population of their own house will follow. The dispersal features of T. infestans may play a very important role in the population growth process; Rabinovich (1972a), with computer simulation techniques, has shown that a random immigration of only 1 or 2 adults per day very rapidly damps population fluctuations. Investigations on dispersal of T. infestans using radioactive tracers, as carried out with R. prolixus and T. maculata by Gómez Núñez (1969, 1970) would provide critical information that we now lack.
The colonizing ability of T. infestans can be considered in relation to dispersion and other population parameters. The following 4 features merit discussion.
(a) Population growth rate is very low. An intrinsic rate of natural increase of 0.101 per head per week is very low. TABLE 13 was prepared to give a relative idea of this figure's meaning. The intrinsic rates of natural increase of 24 species are compared (list not exhaustive but believed to cover most of the information available in the literature). To facilitate comparisons the r value was transformed to weeks when expressed in other times unit. It can be seen that T. infestans ranks 22nd out of a total of 24 species; in other words it is an insect with an extremely low population growth rate.
(b) Average longevity is remarkably high. Lon-gevity values of 115 days for females and 182 days for males can be considered extremely high for an insect species.
(c) Dispersal capacity is apparently very low. As mentioned before, there is no quantitative information about the dispersal capacity of T. infestans. However, if the assumption is made that this capacity is similar to the one found in R. prolixus and T. maculata (Gómez-Núñez 1969), it is very poor. Dispersal of R. prolixus over distances greater than 15 m is probably accomplished only with vertebrate assistance.
(d) Starvation can be resisted for long periods. T. infestans can withstand starvation for periods between 14 and 158 days, depending on the developmental stage and frequency of previous feeding (Perlowagora-Szumlewicz 1969). Borda (1972) states that with only 1 feeding T. infestans can live up to 140, 220, 230, 240, 150 and 210 days, for 1 st, 2nd, 3rd, 4th, 5th instar nymphs and adults, respectively.
These 4 characteristics indicate that T. infestans should be considered a "K-strategist", and not an "r-strategist", in the sense of Wilson & Bossert (J971). An "r-strategist" is a species well adapted to short-lived, unpredictable habitats through (a) a high dispersal power, allowing the quick discovery of adequate newly formed habitats, and (b) a high population growth rate, allowing it to compete successfully against other species in exploiting the habitat and/or to use up an available resource before it deteriorates. In contrast, a "K-strategist" is a species that lives in a more stable habitat. Its populations are, consequently, at or near their

1972 Rabinovich: Vital statistics of Triatominae 367
saturation level K and do not derive much advantage in having a high r. The "K-strategist" is characterized by a high degree of specialization (to avoid interference from competitors) and the ability to maintain a dense population at equilibrium. It is quite clear that the characteristics of T. infestans fit the definition of a "K-strategist", and this is in agreement with the occurrence of T. infestans almost exclusively in or near human dwellings that usually provide a very stable and undisturbed habitat.
The lack of power of dispersal and the low population growth rate can be explained on theoretical grounds. MacArthur & Wilson (1967) give an expression to calculate the extinction time of a population affected by a death density-dependent growth process, after the arrival of a propagule (a mated female) ; this extinction time is a function of b, d, and K (the instantaneous birth- and deathrates and the carying capacity, respectively). From data on T. infestans, the model predicts that the population resulting from 1 mated female, when K is only J0 individuals, would survive for about 100 years; for larger values of K the figures for the time necessary for extinction to occur become astronomical, completely out of scale with MacArthur & Wilson's (1967) graphs.
Thus with a long-lived environment, stable populations, and long extinction times, a high dispersal power is superflous. A low r value is also reasonably to be expected, for as Levins (1970) shows, selection will act most strongly in favor of r if the population is subject to wide fluctuations, and most strongly in favor of K if the population is usually near the carrying capacity. Kimura (1958) also shows that under natural selection the intrinsic rate of natural increase may be reduced, even under conditions of random mating.
TABLE 13, showing a very low r value for T. infestans, can be used to evaluate the degree of fitness of this species to its environment. Evans & Smith (1952) claim that the distance the maximal r deviates from zero reflects the degree to which the natural environment departs from conditions conducive to greatest growth. Furthermore, Levins (1970) shows that fitness is reduced by variance of r for a fixed mean r; from TABLE 7 we see that the variance of r is very low (0.0004). Thus the low r value of T. infestans with its low variance, in the stable environment provided by human dwellings, indicates that this species, as a result of its K strategy, has adapted exceedingly well to its habitat.
In spite of the stability of the environment of T. infestans populations, there is an indication of
some degree of population fluctuation from year to year (Dias 1955a). Even disregarding a correlation between population numbers and climate (which probably exists and plays an important role) the existence of time lags is a factor that can produce population fluctuations. It has been accepted (Birch 1953b) that the interval between the laying of eggs and the emergence of adults is a measure of the time lag which occurs in the influence of density on rate of population increase. If these cohort studies are repeated under a variety of combinations of temperature and humidity conditions, within the "zone" of temperature and moisture tolerance of T. infestans, its distribution can be determined (so far as these 2 factors are concerned) as that area beyond which the finite rate of increase (J) is less than 1 (Birch 1953a).
Dispersal seems to be a critical factor in the population growth process of Triatominae. The following questions need investigation with T. infestans: (a) which are the most active stages in the dispersal process? (b) how does dispersal relate to the reproductive value? (c) is there active movement of individuals between different places within a house? and (d) does each section of a wall act as a more or less isolated habitat?
Model building and simulation techniques play a major role in pest control (MacDonald 1953, Watt 1963, 1964, 1968, Rabinovich 1970e, 1972a, Conway 1970). The information thus far accumulated about demography, insecticidal effects, field data, and the biology of the egg parasite Telenomus fariai of T. infestans, coupled with modern computers and systems analysis techniques, gives us the capacity to build complex and realistic models for this vector, and from them to develop optimal least-cost strategies of control in a far more efficient manner than before.
Acknowledgments:Dr J. C. Gómez-Nuñez, of the Venezuelan ministerio de Sanidad y Asistencia Social, very kindly provided the specimens that were used to start the cohorts, and was most helpful ín editing the manuscript, incorporating many useful suggestions. Dr R. E. Ryckman, of Loma Linda University, California, pointed out some important references that were used in this paper.
LITERATURE CITED Abalos, J. W. & P. Wygodzinsky. 1951. Las Tratominae
Argentinas ((Reduaidae, Hentiptera). Publication N° 601, Instituto de Medicina Regional, Univcrsidad National do Tucuman. 178 p.
Bennet-Clark, H. C. 1966. Abdominal stretch and inhibi-tion of moulting in Rhodnius prolixus (Hemiptera). /. Insect. Physiol. 12: 1019-28.
Birch, L. C. 1948. The intrinsic rate of natural increase of an insect population. f. Anim. Ecol. 17: 15-26.

368 J. Med. Ent. Vol. 9, no. 4 1953x. Experimental background to the study of the
distribution and abundance of insects. 1. The influence of temperature, moisture and food on the innate capacity of increase of three grain beetles. Ecology 34: 698-711.
1953b. Experimental background to the study of the dis-tribution and abundance of insects. 11. The relation between the innate capacity for increase in numbers and the abundance in three grain beetles in experi-mental populations. Ecology 34: 712-26.
Borda, M. R 1972. Algunos nuevos aspectos sobrc biologfa y ccologfa de Triatoma infestans Klug, 1834, y su cnemigo natural Teknomus fariai Lima, 1927. Breves notas referentcs a Tripanosoma cruzi Chagas, 1909. Primer Congruo Latinoamericano de Entomologia. Cuzco, Peril. 12-18 de Abril de 1971. (In press).
Chagas, C. 1909. Nova tripanozorniaze humana. Estudos sobre la morfologia e o ciclo evolutivo do Schizotryparwm cruzi n. gen. n. sp., ajente etiologico de nova entidade morbida do homen. Mem. Inst. Osw. Cruz 1: 159-218.
Conway, G. R. 1970. Computer simulation as an aid to developing strategies for Anopheline control. Mist. Publ. Ent. Soc. Amer. 7: 181-93.
Correa, R R. 1954. Alguns dados sobrc a criariao de Triatomineos em laboraaooryo (Hemiptera: Reduviidae). Folia (7inica et Biol. 22: 51-56.
Correa, R R & A. Schiavi. 1954. ResistMcia sos insccti-cidas, do Triatoma infestans em suas divcrsas fases evolutivas. Folia Clinica et Biol. 22: 57-64.
Danilov, V. N. 1968. The effect of blood meal size taken in the nymphal stage on moulting into imago in tri-atomid bugs, Rhodnius prolixus and Triatoma infestans. Medskaya Parasitol. 46: 218-23 (Trop. Dis. Bull. 65: 982).
Davey, K. G. 1967. Some consequences of copulation in Rhodnius prolixus. ,J. Insect. Physiol. 13: 629-36.
Deevey, E. S. 1947. Life Tables for natural populations of animals. Quart. Rev. Biol. 22: 283-314.
DeWitt, R. M. 1954. The intrinsic rate of natural increase in a pond snail (Phyla gyrina Say). Amer. Nat. 88: 353-59.
Dias, E. 1938. Criaoo de Triatomineos no laboratbrio. Afem. Inst. Osw. Cruz. 33: 407-12.
1945. Um ensaio de profelaxia de molistia de Chagas. Impresa National Rio de Janeiro. 116 p.
1946. Profilaxia da Doenca de Chagas. Resumo dal prinpais atividades to Centro de Estudos Instituto Oswaldo Cruz em Bambui, Minas Gerais. Brasil-Midico 60: 161-63.
1955x. Variacioes mensais da incidencia dal formal evolutivas do Triatoma infestans e do Panstrongylus megistus no Municipio de Bambui, Estado de Minas Gerais. Mem. Inst. Osw. Cruz 53: 457-72.
1955b. Notas sobrc o tempo de evolutao de a1gumas especies de Triatomineos em laborat6rio. Rev. Brash. Biol. 15: 157-58.
Dias, E. & R. Zeledón. 1955. Infestat`ito domiciliaria em grau extremo per Triatoma infestans. Mem. Inst. Osw. 53: 4736.
Dublin, L.I ., A. J. Lotka & M. Spiegeltuan. 1949. Length of life. (Rev. Edit.). Ronald Press, New York.
Evans, R. C. & F. E. Smith. 1952. The intrinsic rate of natural increase for the human louse, Pediculus humanus L. Amer. Nat. 86: 299-310.
Filipponi, A. & M. G. Petrelli. 1967. Autoecologfa e capacity moltiplicata di Macrocheles muscaedomesticae (Scopoli) (Atari: Mesostigmata). Rev. di Parasitol. 28: 129-56.
Fisher, R A. 1930. The Genetical Theory of Natural Selection. Claredon Press, Oxford. 268 p.
Forattini, O. P., E. Juarez & R. de R. Correa. 1969. Mcdida da infestaciio domiciliar por Triatoma infestans. Rev. Saude publ., S. Paulo 3: I1-16.
Franca Rodriguez, M. E. & O. Ceruzzi-Romeo. 1965. Observaciones sobre el comportamiento de Triatoma infestans en el laboraaooryo. An. Fac. Med. Univ. Montevideo. 50: 431-34.
French, N. R & H. W. Kaaz. 1968. The intrinsic rate of natural increase of irradiated Peromyscus in the laboratory. Ecology 49: 1172-79.
Gómez-Núñez, J. C. 1965. Desarrollo de un nuevo metodo para evaluar la infestaci6n intradomiciliaria por Rhodnius prolixus. Acta Cientif. Venezolana 16: 26-31.
1969.Resting places, dispersal and survival of Co., tagged adult Rhodnius prolixus. J. Med. Ent. 6: 83-86.
1970. Ionizing radiation and vectors of Chagas' Disease. The Manitoba Ent. 4: 70-75.
Goodchild, A. J. P. 1955. Some observations on growth and egg production of the blood-sucking Reduviids, Rhodnius prolixus and Triatoma infestans. Proc. Roy. Ent. Soc. (London), Series A, 30: 137-44.
Hack, W. H. 1955. Estudios sobre biologia do Triatoma infestans (Klug, 1834) (Hemiptera, Reduviidae). An. Inst. Med. Regional 4: 125-47.
Joërg, M. E. 1962. Influencia de temperaturas fijas en perfodos anuales sobre metamorfosis y fertilidad de Triatoma infestans. Bol. Chil. Parasitol. 17: 17-19.
Kimura, M. 1958. On the change of population fitness by natural selection. Heredity 12: 145--67.
Leslie, P. H. 1945. On the use of matrices in certain popula-tion mathematics. Biometrika 33: 183-212.
Leslie, P. H. & T. Park. 1949. The intrisic rate of natural increase of Tribolium castaneum Herbst. Ecology 30: 469-77.
Leslie, P. H. & R M. Ranson. 1940. The mortality, fertility and rate of natural increase of the vole (Microtus agrestis) as observed in the laboratory. ,J. Anirn. Fsol. 9: 27-52.
Levins, R. 1970. Fitness and optimization. In: Mathematical Topics in Population Genetics (K. Kojima, Ed.). SpringerVcrlag. 400 p.
Lotka, A. J. 1945. Population analysis as a chapter ín the Mathematical Therory of Evolution. In: Essays on Growth and Form (LeGros Clark, W. E. & P. B. Medawar, Ed.). Oxford. p. 355-85.
Lumbreras, H., J. Arrarte & B. Guevara. 1955. Control biol6gico de los vectores de la enfermedad de Chagas. Rev. Med. Peruana 26: 63-73.
MacArthur, R. H. 1960. On the relation between repro-ductive value and optimal predation. Proc. Nat. Acad. Sci. 46: 143-45.
MacArthur, R. H. & E. O. Wilson. 1967. The theory of Island Biogeography. Princeton University Press. 203 p. Macdonald, G. 1953. The analysis of malaria epidemics. Trop. Dis. Bull. 50: 871-89.
Maggio, C. & F. Rosenbuch. 1915. Studien uber die Chagaskrankheit in Argentinien and die Trypanosomen der "Vinchucas" (Wanzen, Triatoma infestans Klug). Centralbl. f. bakt. 77: 40-46.
Marshall, J. S. 1962. The effects of continuous gamma radiation on the intrinsic rate of natural increase of Daphnia pulex. Ecology 43: 59807.
Messenger, P. S. 1964. Use of Life Tables in a bioclimatic study of an experimental aphid-braconid wasp host-parasite system. Ecology 45: 119-31.
Neghme, A., E. Alfaro, H. Reyes & H. Schenone. 1967.

1972 Rabinovich: Vital statistics of Triatominae 369
hletodo para la crianza de laboratorio de Triatoma infestans (Klug, 1834) (Hemiptera, Reduviidac). Bol. Chil. Parasitol. 22: 107-12.
Neiva, A. 1913. Informações sobre la biologia da Vinchuca, Triatoma infestans Klug. Afem. Inst. Osw. Cruz 5: 29--31.
Nicolle, P. &. M. Lwoff. 1942. Recherches sur la nutrition des reduvides hematophages. I. Developpement des stades larvaires de Triatoma infestans Klug dans les conditions habituelles d'elevage. Bull. Soc. Path. Ex. 35: 219-32.
Ohba, S. 1967. Chromosomal polymorphism and capacity for increase under near optimal conditions. Heredity 22: 169-85.
Pellegrino, J. 1952. Observações sôbre a resistencia do Triatoma infestans ao jejum. Rev. Brasil. Biol. 12: 31720.
Perlowagora-Szunulewicz, A. 1953. Ciclo evolutivo do Triatoma infestans em condigaes de laborat6rio. Rev. Bras. Malario. D. Trop. 5: 35-47.
1954. A eficicia do expurgo domiciliArio tom Hexaclorocicloexana no contr6le do vetor da doenga de Chagas. (A importincia de a1guns caracteristicos biologicos dos triatomineos no planejamento do ciclo de aplicação do insecticida). Rev. Bras. Afalariol. D. Trop. 6: 63-100.
1969. Estudos sôbre la biologia do Triatoma infestans, o principal vetor da doenga de Chagas no Brasil. (Im-portância de algumas de suas caracteristicas bio-lógicas no plenejamento de esquemas de combate a esse vetor). Rev. Bras. Malarial. D. Trop. 21: 117-59.
Pessoa, S. B. 1959. Biologia dos triatomincos. Rev. Goiana Med. 5: 3-I1.
Pessoa, S. B. & N. V. Barros. 1939. Criação do Triatoma infestans na temperatura de estufa. Fôlha Med. 20: 285-87.
Philipp, J. S. & T. F. Watson. 1971. Influence of Tem-perature on population growth of the Pink Bollworm, Pectonophora gossypiella (Lepidoptera: Gelechiidae). Ann. Ent. Sac. Amer. 64: 334-40.
Pinto Dias, J. C. 1965. Observações s6bre o comportamento de Triatomineos brasileiros frente ao jejum em laboratório. Rev. Bras. Malariol. D. Trop. 17: 55-63.
Rabinovich, J. E. 1968. Contribuci6n al estudio de la dinamica de poblaciones. I. Anilisis poblacional de Nasonia vitripennis Walk (Hymenoptera: Pteromalidae). Acta Biol. Fen. 6: 68-81.
1970a. Population dynamics of Telenomus fariai (Hy-menoptera: Scelionidae), a parasite of Chagas' Disease vectors. 11. Effect of host-egg age. J. Med. Ent. 7: 477-81.
1970b. Population dynamics of Telenomus fariai (Hy-menoptera: Scelionidae), a parasite of Chagas' Disease vectors. IV. Effect of female size on progeny. J. Med. Ent. 7: 561-65.
1970c. Population dynamics of Telenomus fariai (Hymenoptera: Scelionidae), a parasite of Chagas' Disease Vectors. IV. Ability to discriminate between parasitized and nonparasitized hosts. Acta Cientif. Venezolana 21: 154-56.
1970d. Vital statistics of Synthesiomyia nudiseta (Diptera: Muscidae). Ann. Ent. Soc. Amer. 63: 749-52.
1970e. El analisis de Sistemas en Ecologia. In: La Ciencia en Venezuela 1970. Direction de Cultura. Universidad de Carabobo, Valencia, Venezuela. p. 193-223.
1971. Population dynamics of Telenomus fariai (Hymenoptera: Scelionidae), a parasite of Chagas' Disease vectors. II. Preferences for and progeny from different age classes of host eggs. Ann. Ent. Soc. Amer. 64: 29-32.
1972a. Simulacidn en una computadora digital de la regulation poblacional de triatominos vectores de la enfermedad de Chagas por parte del parisito Telenomus fariai (Hymenoptera: Scelionidae), y de las estrategias para el control integrado. Anales el ler Congreso Latinoamericano de Entomologia. Cuzco, Peru, 12-18 Abril, 1971. (In press).
1972b. Comparaci6n experimental entre la longevidad y cl tiempo de desarrollo de dos poblacinones alopitridas de Telenomus fariai (Hymenoptera: Scelionidae) a dos temperaturas constantes. Anales del ler Congreso Latinoamercano de Fntomologia. Cuzco, Perd, 12-18 Abril, 1971. (In press).
Rabinovich, J. E., G. Escalante & A. Rodriguez. 1970. Dindmica poblacional de Telenomus fariai (Hyrrtenoptera: Scelionidae). I. Eflecto de la carencia de huespedcs (Triatoma phyllosoma) (Hemiptera: Reduviidae) sobre la produccidn de progenie. Acta Biol. Ven. 7: 5-21.
Rocha e Silva, E. O., J. Maluf & R. de R. Correa. 1970. Doencga de Chagas-Actividades de vigilancia ento-mologica dcsenvolvida numa area do Estado de Sao Paulo, Brasil. Rev. Saude publ., S. Paulo. 4: 129-45.
Romaña, C. 1963. Enfermedad de Chagas. Lopez libreros Editores, S. R. L. Buenos :fires, Argentina. 242
Root, R. B. 1960. An estimate of the intrinsic rate of natural increase in the Planarian, Dugesia tigrina. Ecology 41: 369-72.
Ryckman, R. E. 1951. Recent observations of cannibalism in Triatoma (Hemiptera: Reduviidae). J. Parasitol. 37: 433-34.
1952. Laboratory culture of Triatominae with observa-tions on behaviour and a new feeding device. J. Par-asitol. 38: 210-14.
Smith, L. B. 1965. The intrinsic rate of natural increase of Cryptolestes ferrugineus (Stephens) (Colcoptera, Cucujidae). J. Stored Prod. Res. 1: 35-49.
Steel, R. G. D. & J. H. Torrie. 1960. Principles and procedures of statistics, with special reference to the biological sciences. McGraw-Hill Book Co., Inc. New York. 481 p.
Sterling, W. L. & P. L. Adkisson. 1970. Seasonal rates of increase for a population of the Boll Weevil, An-thonomus grandis, in the High and Rolling Planes of Texas. Ann. Ent. Soc. Amer. 63: 1696-1700.
Trejos, A. R., Rosabal & A. Y. de Hernández. 1966. Fecundacion de una hembra de Rhodnius prolixus Stal, 1852, por dos machos. Rev. Biol. Trop. 14: 105-10.
Usinger, R. L., P. Wygodzinsky & R. E. Ryckman. 1966. The Biosystematics of triatominae. Ann. Rev. Ent. 11: 309-30.
Watt, K. E. F. 1963. Dynamic programming, "Look-ahead programing", and the strategy of insect pest control. Can. Ent. 95: 525-36.
1964. The use of mathematics and computers to determine optimal strategy and tactics for a given pest control problem. Can. Ent. 96: 202-20.
1968. Ecology and Resource Management. A quantitative approach. MacGraw-llill Co., Inc. New York. 450 p.
Wiesinger, D. 1955. Die Bcdentung der Umweltfaktorem fur den Saugakt van Triatoma infestans. Doctoral Thesis, University of Basilea. 114 p.
Wilson, E. O. & W. H. Bossert. 1971. A primer af' popula-lion Biology. Sinauer Associates, Inc. Publishers. Stanford. 192 p.
Wood, S. 1964. The laboratory Culture of Triatoma (Hemip-tera, Reduviidae). Bull. Wld Hlth Organ. 31: 579-81.
Zeledón, R. 1957. Sobre la biologia de Teknomus fariai Lima, 1927. (Hymenoptera: Scelionidae), parisito endbfago de huevos de algunos Triatominae. Rev. Biol. Trop. 5: 1-17.

368 J. Med. Ent. Vol. 9, no. 4
Appendix Definition of terms and formulae used in the calculation of population parameters of Triatoma infestans.
SYMBOL NAME AND/OR DEFINITION CALCULATION b Instantaneous birth-rate. (r�)/(e-r -1)
c(j) Percentage distribution of deve-lopmental stage j in a population with stable age distribu tion.
�=
δ
γx
xc )(
C(X) Percentage distribution of age class x in a population with stable age distribution. 100 � Lx er(x+1)
d Instantaneous death-rate. B - r Expectation of life or average
future lifetime. Tx /lx j Identification for a given de-
velopmental stage; j=γ, and j= δ are the average initial and final ages, respectively, of a given developmental stage. Observed values
Lx Number of weeks lived by the cohort between ages x and x+1. (lx + lx+1)/2
lx Probability of an individual being alive at the end of age x. Nx/N0
mx Age specific fecundity (n° eggs/�/week). Observed value
m´x Age specific fecundity corrected for sex ratio. mxp
Nx Number of animals alive at the end of age x. Observed value
p Sex ratio (�: �). Observed value r Intrinsic rate of natural increase. The value of r that
satisfies the equation:
1´ =�=
−ω
αx
rxxx eml
R0 Net reproduction rate. �
=
ω
αxxxml ´
T Generation time loge (R0)/r Tx Total number of weeks lived by
the cohort from x until all have died.
�=
x
iiL
φ
V Reproductive value of cohort �
=
φ
0xxv
vj Developmental stage specific reproductive value �
=
δ
γxxv
*
vj Developmental stage specific reproductive value weighted after the stable age distribution.
Vjc(j)
vx Age specific reproductive value (worth of an individual age x in terms of the number of off-spring it is destined to contribute in the next generation).
�=
−φ
xiii
ri
x
rx
mlele
x Age (here in units of weeks) ; x= 0, is the initiation of the cohorts. Observed value
� Age of first reproduction. Observed value � Finite birth-rate.
���
����
��
=
+φ
0
)1(
1
x
xrxel
∅ Maximun life span (last age group of the cohort). Observed value
λ Finite rate of increase. Antiloge (r) ω Age of last reproduction. Observed value