veterinary immunology and immunopathology · s.m. steelman et al. / veterinary immunology and...
TRANSCRIPT

R
C
SAa
7b
Uc
ARRA
KNCA
1
toTli1
0h
Veterinary Immunology and Immunopathology 153 (2013) 217– 226
Contents lists available at SciVerse ScienceDirect
Veterinary Immunology and Immunopathology
j ourna l h omepa ge: www.elsev ier .com/ locate /vet imm
esearch paper
ellular and humoral immunity in chronic equine laminitis
amantha M. Steelmana,∗, Daisy Johnsona, Bettina Wagnerb,shley M. Stokesc, Bhanu P. Chowdharya
Veterinary Integrative Biosciences, College of Veterinary Medicine & Biomedical Sciences, Texas A&M University, College Station, TX7843-4458, United StatesDepartment of Population Medicine and Diagnostics Sciences, College of Veterinary Medicine, Cornell University, Ithaca, NY 14853,nited StatesDepartment of Human Nutrition, Food, and Animal Sciences, University of Hawaii, Honolulu, HI 96848, United States
a r t i c l e i n f o
rticle history:eceived 25 November 2012eceived in revised form 1 March 2013ccepted 4 March 2013
eywords:eutrophilytokinentibody
a b s t r a c t
Chronic equine laminitis causes persistent pain and lameness in affected animals andoften necessitates euthanasia when pain management strategies become ineffective. Pub-lished studies as well as anecdotal reports suggest that this chronic inflammatory diseaseis associated with systemic alterations in immune responsiveness, perhaps involving anautoimmune component. We investigated this broad hypothesis by measuring a variety ofimmune indicators in healthy control horses (CON) and horses with chronic laminitis (LMN).We found that white blood cells from LMN horses produced more IFN� than did cells fromCON horses when stimulated in vitro with polyinosinic-polycytidylic acid [poly(I:C)], pos-sibly due to an elevated number of circulating monocytes. No differences between groupswere observed in plasma concentrations of IgG, IgA, IgM, IgE, or rheumatoid factor. Laminartissue from LMN horses expressed elevated levels of keratinocyte damage-related genesas well as inflammatory cytokines and chemokines, which corresponded with a modestamount of neutrophil infiltration as shown by histological staining of fixed tissue and accu-
mulation of neutrophil elastase protein. Taken together, our results do not support thehypothesis of an autoimmune component in chronic laminitis, although the strong induc-tion of neutrophil chemokines and the presence of tissue neutrophils suggests that this celltype is likely involved in perpetuating the inflammation and tissue damage associated withthis disease.© 2013 Elsevier B.V. All rights reserved.
. Introduction
Laminitis is a debilitating disease of the equine foothat often occurs secondary to gastrointestinal diseaser metabolic syndrome (Slater et al., 1995; USDA, 2000).he acute phase of the disease is characterized by severe
ameness, disintegration of the laminar tissue connect-ng the hoof wall to the underlying third phalanx (Pollitt,996), and in some cases, dorsopalmar rotation of the third∗ Corresponding author. Tel.: +1 979 862 1436; fax: +1 979 847 8981.E-mail address: [email protected] (S.M. Steelman).
165-2427/$ – see front matter © 2013 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.vetimm.2013.03.001
phalanx. This condition is usually not reversible and horsesthat survive an episode of acute laminitis often experiencepersistent lameness for years. This stage of the disease istermed chronic laminitis or “founder” (Hood, 1999). Acutelaminitis transitions to the chronic form of the diseasein an estimated 75% of cases (Moore, 2010), leaving themajority of afflicted horses with some degree of continuedlameness. Whereas much effort has been expended toidentify the causes of acute laminitis, the chronic form of
the disease remains relatively uncharacterized.Existing publications regarding chronic laminitis pri-marily focus on management and shoeing strategies toreduce pain and improve quality of life (Hunt and Wharton,

218 S.M. Steelman et al. / Veterinary Immunology and Immunopathology 153 (2013) 217– 226
Table 1Real time PCR primers used in the current study.
Gene Forward Primer Reverse Primer
IFN� TACCTATTACTGCCAGGCCGCG ATCCAGGAAAAGAGGCCCACCAIL-6 ACTCCTTCTTCACAAGCACCGTC AGTGGGGTGGGGAAAGCAGTAGIL-1 CCAGAGGCGGCCGGGACATAAC GGGAAGGCAGCTGGGCATTGATTTNF� CTGGCCCAGACACTCAGATCATCT CATTTGCACGCCCACTCAGCACTB CCAGCACGATGAAGATCAAG GTGGACAATGAGGCCAGAATCD3 AATGCAGCCACTGTGTTGGGC TCTGAAGCTCTTGACTGGCGAACCD14 TGGCGACATGGTCCTCTCGT CGCGCACCATGGTCTATAGGTCCD19 AGACTCCTTCTCCAACGGTAA CCACCTCTTCATCCTCATTCTCCL2 TCCAGTCACCTGCTGCTATAC ATTCTTGGCTTTTGGAGTAGGCCL4 AGAGAACCAGGGAAAATCAA GTCCAGATTAGGAACCAAGGCCL7 TCAATAAGAAGATCCCCATCC TCTTGTCCAGGTAGTTCGTGAIL-8 ACAAGTCCCCAGGGACAGCA ACAACCGCAGCTTCACACAGADEFB4 ACTTGCCTTCCTCATTGTCTT AGCAGTTTCTCCGCTTTCTATPI3 CTTCTTGATCCTGGTGGTGTT ACTGAATCTTGCCCTTTGACTSOD2 ACTTTGGTTCCTTCGACAAAT CAGGGGAATAAGACCTGTTGTTLR4 GGCCAGTGATTTTCCAGTATT AGGGTGGTCAGGTTAGTCATCS100A8 ACGGATCTGGAGAATGCTATC TGATGTCCAACTCTTTGAACCS100A9 CGACCTAGAGACCATCATCAA TGCTTGTCCTCATTAGTGTCC
blood were then treated with 1 �g/ml lipopolysaccharide
MMP13 CTTGAGCTGGACTCGTTGTT
2010; Morrison, 2010; Orsini et al., 2010); only a handfulhave sought to identify the pathological processes under-lying this disease (Carter et al., 2011; Faleiros et al., 2004;Johnson et al., 2004; Kuwano et al., 2005). In particular,one study suggested that chronic laminitis might have anautoimmune component (Wagner et al., 2003b), a hypoth-esis which has yet to be tested. Autoimmune diseases aregenerally characterized by circulating autoantibodies orthe presence of autoreactive T cells (Rose and Bona, 1993) inaddition to disease-specific criteria such as elevated whiteblood cell (WBC) cytokine levels and increased serum con-centrations of acute phase proteins or immunoglobulins(Aho et al., 1997; Badolato and Oppenheim, 1996; Daviset al., 2010). We recently found evidence of dysregulationof IgM, IgA, and the acute phase proteins fibrinogen, fetuinsA and B, alpha-2 macroglobulin, and alpha-1 antitrypsin inthe plasma of horses with chronic laminitis (Steelman andChowdhary, 2012), thus providing support for the explo-ration of a possible autoimmune component of this disease.
In the present study, we investigated parameters ofhumoral and cellular immunity at both the local and sys-temic levels in horses with chronic laminitis in order tocharacterize the immune status of foundered horses. Ourresults suggest a role for neutrophils in perpetuating thechronic inflammation and tissue damage seen in this dis-ease.
2. Methods
2.1. Animals and tissue collection
All animal protocols were approved by the Texas A&MUniversity College of Veterinary Medicine Clinical ResearchReview Committee and/or the Institutional Animal Careand Use Committee, as appropriate, and were performed
with the consent of the owner or the owner’s agent.Details of the horses used for each experiment are includedin Supplementary Table 1. Horses with chronic lamini-tis were diagnosed by a licensed veterinarian based onCAGCAGGATTAAGGGGATAGT
clinical presentation, case history, and radiographic evi-dence of dorsopalmar rotation of the distal phalanx. Adetailed description of all laminitis cases, including Obelgrade of lameness (Obel, 1948), is provided in Supplemen-tary Table 2.
Blood samples (n = 35) were collected from horses andponies admitted to the Texas A&M University Large Ani-mal Clinic (College Station, TX), the Hoof Diagnostic andRehabilitation Clinic (Bryan, TX), or that were owned byTexas A&M University. Blood samples were collected fromthe jugular vein into evacuated blood tubes containing hep-arin or EDTA. Samples were either centrifuged at 1200 x gfor plasma collection, processed immediately for WBC RNAisolation, or used as described below for in vitro culture ofWBCs. Complete blood counts were performed by the TexasA&M University Clinical Pathology service on a subset ofsamples.
Tissue samples were collected at necropsy from the dor-sal aspect of the hoof of either forelimb of horses admittedto the Texas A&M University Large Animal Clinic, the HoofDiagnostic and Rehabilitation Clinic, or the Louisiana StateUniversity Veterinary Teaching Hospital (Baton Rouge, LA).Horses in the control group were euthanized for reasonsunrelated to laminitis or other lameness. Control sampleswere not collected from unaffected limbs of unilaterallylame horses, as it would be impossible to rule out potentialeffects of increased weight bearing on the unaffected limb.
2.2. Blood cell culture
Heparinized blood samples from control (CON, n = 10)and chronic laminitis (LMN, n = 10) horses were thor-oughly mixed by inversion and the total number of WBCswas counted for each sample. Sterile aliquots of whole
(LPS; from E. coli 0111:B4, Sigma-Aldrich, St. Louis, MO),50 �g/ml polyinosinic:polycytidylic acid (poly (I:C); Sigma-Aldrich), or vehicle control (PBS) for 6 h at 37 ◦C. Uponcompletion of the culture, RNA was isolated immediately.

S.M. Steelman et al. / Veterinary Immunology and Immunopathology 153 (2013) 217– 226 219
Table 2Summary of complete blood counts performed on healthy control horses (n = 5) and horses with chronic laminitis (n = 12). Lymphos: total lymphocytes,Monos: monocytes, Eosin: eosinphils, Total WBC: white blood cell count, RBC morph: red blood cell morphology.
Case # Group Neutrophils Lymphos Monos Eosin Total WBC RBC morph
21 CON 5934 2494 86 86 8.6 rouleaux22 CON 4392 2376 – 432 7.223 CON 5610 2635 85 170 8.524 CON 6175 3040 95 95 9.526 CON 4160 1920 128 128 6.428 LMN 4160 2015 325 – 6.5 rouleaux29 LMN 4745 2409 73 73 7.3 rouleaux30 LMN 5251 3204 267 89 8.9 rouleaux31 LMN 4176 2520 216 216 7.2 rouleaux33 LMN 8284 2180 327 109 11 rouleaux34 LMN 3968 1426 248 434 6.2 rouleaux35 LMN 8510 2530 345 115 11.536 LMN 5002 2788 – 410 8.237 LMN 3596 2170 186 186 6.2
2
MuCScsliw(wT4miawcs
2
cptMw4SW(ptt(au
38 LMN 4686 1775
39 LMN 5369 3367
40 LMN 5355 2720
.3. Gene expression
RNA was isolated from WBCs using the RNeasy Bloodini Kit (Qiagen, Valencia, CA) and from laminar tissue
sing Qiazol Lysis Buffer (Qiagen) followed by the RNeasyleanup Kit (Qiagen). RNA was reverse transcribed (Super-cript III First Strand Synthesis Kit or Superscript VILODNA Synthesis Kit, Invitrogen, Carlsbad, CA) and the sub-equent cDNA was used to determine gene expressionevels using real time PCR. Twenty-five nanograms of lam-nar tissue cDNA or 2.5 ng of WBC RNA per PCR reaction
as added to LightCycler 480 SYBR Green I master mixRoche Applied Science, Indianapolis, IN) and 0.6 �M for-ard and reverse primers. Primer sequences are listed in
able 1. PCR reactions were performed on a LightCycler80 (Roche) and the data was analyzed using the Pfafflethod (Pfaffl, 2001). Negative controls included samples
n which the reverse transcriptase was omitted as wells PCR reactions containing no template. Melting curvesere examined for each reaction to ensure that the fluores-
ence detected came from a single product of the expectedize.
.4. ELISAs
Capture ELISAs were performed to determine plasmaoncentrations of IgA and IgM using validated antibodyairs (Bethyl Labs, Montgomery, TX) and standard pro-ocols. Briefly, high protein-binding 96 well plates (Nunc
axiSorp, Thermo Scientific, Rockford, IL) were coatedith 100 �g of unlabelled capture antibody overnight at◦C. Plates were blocked with SuperBlock buffer (Thermocientific, Waltham, MA) for 30 min at room temperature.hole plasma samples were diluted 1:4000 in assay buffer
10% SuperBlock in PBS) and 100 �l of each diluted sam-le was pipetted in duplicate onto the plates, which werehen incubated at 37 ◦C for 90 min. HRP-conjugated detec-
ion antibodies were diluted 1:50,000 (IgM) or 1:150,000IgA) in assay buffer and incubated on the plates for 2 ht room temperature. SigmaFast OPD (Sigma-Aldrich) wassed as a substrate for HRP. Sample concentrations were284 355 7.1 rouleaux273 91 9.1 rouleaux255 170 8.5
calculated using dilutions of a reference serum of knownconcentration. Plasma concentrations of IgG were deter-mined using a similar protocol adapted for use with a directELISA. Plates were coated overnight at 4 ◦C with plasmasamples diluted 1:80,000 in PBS and the HRP-conjugateddetection antibody was used at a dilution of 1:50,000; theremainder of the protocol was unchanged. IgE was mea-sured as described previously (Wagner et al., 2003a).
Rheumatoid factor (RF) assays were also performed byELISA using heat-aggregated equine IgG (10 min at 70 ◦C)diluted 1:100 in PBS as the capture antibody. Detectionantibodies were anti-IgA or anti-IgM (Bethyl Laboratories)diluted 1:5000 in assay buffer. Plasma samples were seri-ally diluted from 1:31.25 to 1:8000 (IgM-RF) or 1:125 to1:2000 (IgA-RF) in order to determine titer.
2.5. Western blots
Western blotting was performed according to publishedprotocols (Steelman and Chowdhary, 2012). Antibodiesused are as follows: polyclonal rabbit anti-human neu-trophil elastase (ab68672, Abcam, Cambridge, UK) andgoat anti-rabbit secondary antibody (ab97080, Abcam);monoclonal rabbit anti-human beta actin (clone 13E5, CellSignaling Technology, Danvers, MA) and goat anti-rabbitsecondary antibody (#7074, Cell Signaling Technology).Resulting bands were quantified using ImageJ software(Rasband, 1997–2011).
2.6. Histology
Laminar tissue was fixed in neutral buffered formalinfor at least 48 h before being embedded, sectioned to athickness of 10 �m, and stained with H&E. Observation ofneutrophils was performed by manual inspection of 10–2040x fields of view per animal.
2.7. Statistics
Comparisons between CON and LMN groups were madeby Student’s t test or two-way ANOVA, as appropriate.

logy and
220 S.M. Steelman et al. / Veterinary ImmunoAs previous studies (Wang, 2010) suggested that inflam-matory gene expression would be upregulated in laminartissue from horses with laminitis, a one-tailed t test wasused to evaluate these data. A one-tailed t-test was alsoused to assess differences in IgA and IgM concentra-tions between groups, as previous proteomics experimentsin our lab suggested that these immunoglobulins wereelevated in horses with chronic laminitis (SMS, unpub-lished data; Steelman and Chowdhary, 2012). Calculationswere performed using Prism (GraphPad Software Inc., LaJolla CA) or a variation of the S programming languagecalled R, which is commonly used for statistical analysis(http://www.r-project.org). A P value < 0.05 was consid-ered sufficient to reject the null hypothesis.
3. Results
3.1. Cellular immunity
Complete blood counts (CBCs) were performed on 5healthy control horses and 12 horses with chronic lamini-tis. Absolute counts of all cell types fell within normalranges, although the number of monocytes was higher inLMN horses than in controls (99 ± 10 CON vs. 254 ± 38 LMN,P < 0.01). Rouleaux were observed in 8 LMN horses (75%)and 1 CON horse (20%). Total WBC number did not differbetween groups.
WBC expression of the inflammatory cytokinesinterleukin-1� (IL-1), interleukin-6 (IL-6), tumor necrosisfactor alpha (TNF�), and interferon gamma (IFN�) wasassessed in horses with chronic laminitis (n = 9) and inhealthy controls (n = 9). As the 18 horses used for thisexperiment had not had CBCs performed, the total numberof WBCs was counted manually; no differences wereseen between groups. In RNA isolated from freshly col-lected blood samples, we found no significant differencesbetween the two groups in any of the cytokines tested
(Fig. 1).To assess the response of WBCs to exposure topathogen-associated molecular patterns (PAMPs), wholeblood samples from control (n = 9) and foundered (n = 9)
Fig. 1. Basal cytokine expression in equine white blood cells. RNA wasisolated from whole blood of healthy controls (CON, n = 9) and horses withchronic laminitis (LMN, n = 9). Pro-inflammatory cytokine expression wasdetermined by real time PCR. *P < 0.05.
Immunopathology 153 (2013) 217– 226
horses were exposed to LPS, poly(I:C), or a vehicle con-trol for 6 h. LPS and poly(I:C), a double-stranded RNAmolecule, were chosen as they are often associated withpathogenic bacteria and viruses, respectively. LPS andpoly(I:C) induced expression of IL-1, TNF�, IL-6, and IFN�(P < 0.01) in both groups; induction of IFN� was higher inthe LMN group than in controls (P < 0.05; Fig. 2). Post hoctesting revealed a significant elevation of IFN� in LMN sam-ples treated with poly(I:C) and a trend toward elevation insamples treated with LPS. No differences were observed inIL-1, TNF�, or IL-6 between CON and LMN groups.
3.2. Humoral immunity
Total plasma concentrations of IgM, IgA, IgE, and IgGin healthy horses and those with chronic laminitis weremeasured by ELISA. No differences were found between theLMN group (n = 8) and controls (n = 14), with the exceptionof IgA, which was significantly elevated in the LMN group(P = 0.025, Fig. 3).
As chronic laminitis shares several similarities withautoimmune diseases such as rheumatoid arthritis, welooked for the presence of IgM and IgA rheumatoid fac-tors in the plasma of LMN horses, young controls (YC),and mature, age-matched controls (MC). The YC group uni-formly tested negative for both types of rheumatoid factor;we thus used the YC values to establish the minimum valuefor a positive titer, indicated by the dashed horizontal linesin Fig. 4. Two MC horses and two LMN horses exceeded thislimit for IgA rheumatoid factor, although the titers for theMC horses were 500 and 1000, whereas the titers for theLMN horses were 62 and 250 (Fig. 4). Similar results wereseen with IgM rheumatoid factor, with 3 and 2 horses test-ing positive in the MC and LMN groups, respectively (Fig. 4).No significant differences were seen between the MC andLMN groups for either rheumatoid factor.
3.3. Laminar inflammation
RNA expression of innate inflammatory cytokines,antimicrobial and damage-related genes, chemokines, andWBC markers was assessed in laminar tissue of horses withchronic laminitis (n = 6) as well as in control horses (n = 5).Chemokine and WBC marker genes were chosen to screenfor the presence of infiltrating inflammatory cells in thelaminar tissue. Other genes of interest were chosen basedon microarray data describing transcriptional changes inlaminar tissue in three different experimental models ofacute laminitis (Budak et al., 2009; Noschka et al., 2008;Wang, 2010). The genes investigated herein thus representa small group of consistently upregulated transcripts thatwe believe to be integral to the pathology of acute laminitisand possibly involved in chronic laminitis.
We found significant upregulation of pro-inflammatorycytokine genes, antimicrobial/damage-related genes, andchemokine genes in laminar tissue from foundered horses(Fig. 5). IL-1 and IL-6 were both significantly increased
(P < 0.05) in chronic laminitis, although TNF� and IFN�were not (Fig. 5). The antimicrobial peptides beta-defensin4 (DEFB4) and peptidase inhibitor 3 (PI3) were elevated 22-and 132-fold, respectively (P < 0.05), although no change
S.M. Steelman et al. / Veterinary Immunology and Immunopathology 153 (2013) 217– 226 221
Fig. 2. Cytokine expression in PAMP-stimulated equine white blood cells. Whole blood of healthy controls (CON, n = 9) and horses with chronic laminitis(LMN, n = 9) was cultured for 6 h with vehicle (VEH), LPS (1 �g/ml), or poly(I:C) (50 ng/ml). RNA was isolated from whole blood and pro-inflammatorycytokine expression was determined by real time PCR. *P < 0.05.
Fig. 3. Plasma immunoglobulin concentrations. Immunoglobulin concentrations were measured by ELISA in control horses (CON, n = 14) and horses withchronic laminitis (LMN, n = 8). *P < 0.05.

222 S.M. Steelman et al. / Veterinary Immunology and Immunopathology 153 (2013) 217– 226
Fig. 4. IgA and IgM rheumatoid factor titration curves. Titers were measured by ELISA in mature (> 5 y.o., n = 8) control horses [CON(M)] and horses withchronic laminitis (LMN, n = 8).
Fig. 5. Laminar expression of inflammatory cytokines, damage-related genes, chemokines, and cell surface markers. RNA was isolated from laminar tissueof control horses (CON, n = 5) and horses with chronic laminitis (LMN, n = 6). Gene expression was determined by real time PCR. *P < 0.05.
S.M. Steelman et al. / Veterinary Immunology and
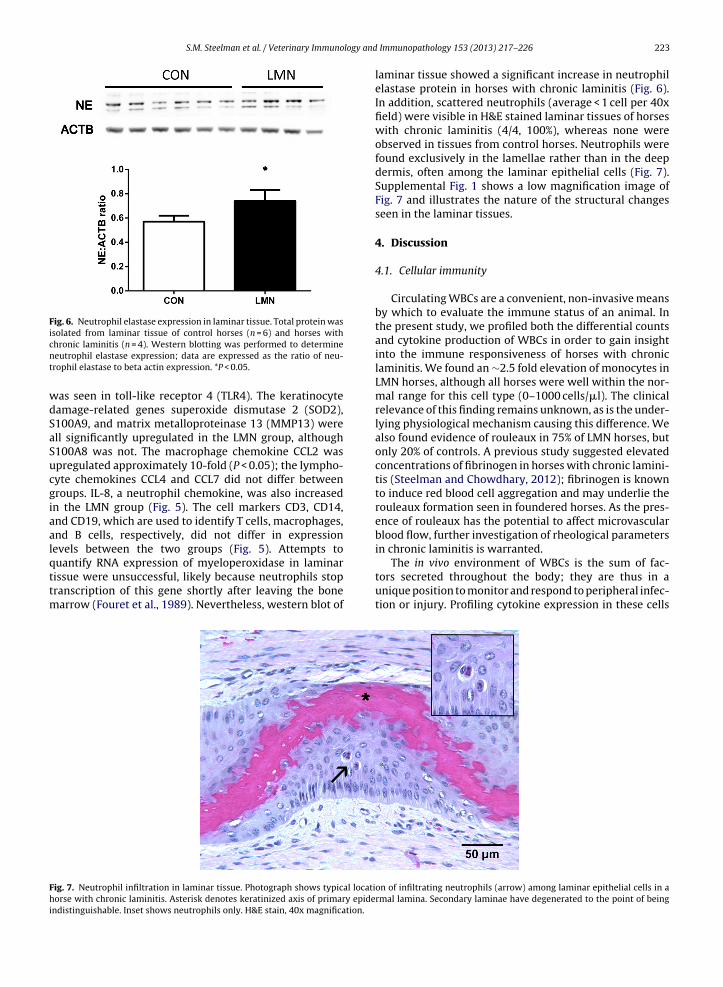
Fig. 6. Neutrophil elastase expression in laminar tissue. Total protein wasisolated from laminar tissue of control horses (n = 6) and horses withcnt
wdSaSucgiaalqttm
Fhi
hronic laminitis (n = 4). Western blotting was performed to determineeutrophil elastase expression; data are expressed as the ratio of neu-rophil elastase to beta actin expression. *P < 0.05.
as seen in toll-like receptor 4 (TLR4). The keratinocyteamage-related genes superoxide dismutase 2 (SOD2),100A9, and matrix metalloproteinase 13 (MMP13) werell significantly upregulated in the LMN group, although100A8 was not. The macrophage chemokine CCL2 waspregulated approximately 10-fold (P < 0.05); the lympho-yte chemokines CCL4 and CCL7 did not differ betweenroups. IL-8, a neutrophil chemokine, was also increasedn the LMN group (Fig. 5). The cell markers CD3, CD14,nd CD19, which are used to identify T cells, macrophages,nd B cells, respectively, did not differ in expressionevels between the two groups (Fig. 5). Attempts to
uantify RNA expression of myeloperoxidase in laminarissue were unsuccessful, likely because neutrophils stopranscription of this gene shortly after leaving the bonearrow (Fouret et al., 1989). Nevertheless, western blot of
ig. 7. Neutrophil infiltration in laminar tissue. Photograph shows typical locatiorse with chronic laminitis. Asterisk denotes keratinized axis of primary epide
ndistinguishable. Inset shows neutrophils only. H&E stain, 40x magnification.
Immunopathology 153 (2013) 217– 226 223
laminar tissue showed a significant increase in neutrophilelastase protein in horses with chronic laminitis (Fig. 6).In addition, scattered neutrophils (average < 1 cell per 40xfield) were visible in H&E stained laminar tissues of horseswith chronic laminitis (4/4, 100%), whereas none wereobserved in tissues from control horses. Neutrophils werefound exclusively in the lamellae rather than in the deepdermis, often among the laminar epithelial cells (Fig. 7).Supplemental Fig. 1 shows a low magnification image ofFig. 7 and illustrates the nature of the structural changesseen in the laminar tissues.
4. Discussion
4.1. Cellular immunity
Circulating WBCs are a convenient, non-invasive meansby which to evaluate the immune status of an animal. Inthe present study, we profiled both the differential countsand cytokine production of WBCs in order to gain insightinto the immune responsiveness of horses with chroniclaminitis. We found an ∼2.5 fold elevation of monocytes inLMN horses, although all horses were well within the nor-mal range for this cell type (0–1000 cells/�l). The clinicalrelevance of this finding remains unknown, as is the under-lying physiological mechanism causing this difference. Wealso found evidence of rouleaux in 75% of LMN horses, butonly 20% of controls. A previous study suggested elevatedconcentrations of fibrinogen in horses with chronic lamini-tis (Steelman and Chowdhary, 2012); fibrinogen is knownto induce red blood cell aggregation and may underlie therouleaux formation seen in foundered horses. As the pres-ence of rouleaux has the potential to affect microvascularblood flow, further investigation of rheological parametersin chronic laminitis is warranted.
The in vivo environment of WBCs is the sum of fac-tors secreted throughout the body; they are thus in aunique position to monitor and respond to peripheral infec-tion or injury. Profiling cytokine expression in these cells
on of infiltrating neutrophils (arrow) among laminar epithelial cells in armal lamina. Secondary laminae have degenerated to the point of being

logy and
224 S.M. Steelman et al. / Veterinary Immunothus provides a static assessment of the animal’s overallhealth and well-being. In this study, however, we foundno change in basal production of IL-1, IL-6, TNF�, or IFN�.In vitro antigen-stimulated cytokine production, on theother hand, has been shown to be an excellent predictor ofsome disease states (Davis et al., 2010). In particular, geneexpression of WBCs cultured in their in vivo environment(i.e., whole blood) is thought to be more representative ofthe true physiology of the animal than WBCs cultured inartificial media (Chen et al., 2010; Hodge et al., 2000). Wefound that, while WBCs from both groups responded toexposure to LPS or poly(I:C), foundered horses expressedsignificantly more IFN� than did controls. It is possible thatthis is due to the elevated number of circulating monocytesin LMN horses, although unpublished data from our labora-tory suggests that highly purified neutrophils isolated fromLMN horses produce elevated amounts of IFN� (S.M. Steel-man, unpublished observations). Although neutrophils arenot generally known for expression of interferons, someevidence suggests that IFN� can be induced by stimulationwith IL-12 (Ethuin et al., 2004). Importantly, the differ-ential response of LMN horses to perceived pathogens,although conducted in vitro with mixed WBCs rather thantissue-resident antigen presenting cells, might underly theadverse reactions to vaccination in some LMN horses. Thishypothesis was not specifically tested in the present study,but is currently under investigation in our laboratory.
4.2. Humoral immunity
Elevated immunoglobin levels are a hallmark of a num-ber of chronic inflammatory and autoimmune diseases,including hepatitis, asthma, and rheumatoid arthritis (Ahoet al., 1997; Mieli-Vergani and Vergani, 2011; Platts-Mills,2001). Previous work in our laboratory suggested dif-ferences in plasma immunoglobulin concentrations in alimited number of animals (Steelman and Chowdhary,2012), so we sought to confirm and expand upon these datain the present study. Herein, we found that IgA was signif-icantly elevated in horses with chronic laminitis. Whereasthe functions of secretory IgA, primarily found at mucosalsurfaces, are well described (Pabst, 2012), the immuno-logical role of serum IgA is not completely understood.Serum IgA, which exists predominantly in the monomericform, is evelated in several chronic inflammatory diseasessuch as ankylosing spondylitis and alcoholic liver diseaseand has been shown to have anti-inflammatory proper-ties (reviewed in Monteiro, 2010). Specifically, IgA inhibitsthe respiratory burst in human neutrophils and mono-cytes (Wolf et al., 1994b), decreases monocyte productionof TNF� and IL-6 (Wolf et al., 1994a), and impairs neu-trophil phagocytosis (Wilton, 1978) and chemotaxis (VanEpps and Williams, 1976). Notably, our previous workuncovered a similar increase in the anti-inflammatoryprotein apolipoprotein A-IV (APOA-IV) in the plasma ofhorses with chronic laminitis (Steelman and Chowdhary,2012). The elevation of plasma IgA and APOA-IV might
represent a compensatory effort of the immune system todownregulate inflammation in chronic laminitis. Furtherinvestigation of the anti-inflammatory response in chroniclaminitis is warranted.Immunopathology 153 (2013) 217– 226
It is also interesting to note that some older horsesexhibited positive rheumatoid factor titers, whereas theyounger ones did not; the reason for the occurrence ofrheumatoid factor in otherwise healthy horses is unknown.A previous study found no increase in C1q-binding immunecomplexes in the serum of horses with chronic inflamma-tory diseases, including osteoarthritis and osteochondrosisdissecans (Osborne et al., 1995), although Stanek et al.(1984) found synovial fluid immune complexes in horseswith laminitis [as reported by Osborne et al. (1995)]. Ourdata, however, do not support the hypothesis of immunecomplexes as contributors to the pathology of chroniclaminitis.
4.3. Laminar inflammation
Although it is generally accepted that some level ofinflammation is ongoing in the laminar tissue of founderedhorses, there is a lack of data describing the nature of thisinflammation. For this reason, we measured the expressionlevels of several inflammation-related genes as well as a“core” group of genes that we and others have found to beconsistently upregulated in several experimental modelsof acute laminitis. We believe that these core genes areintegral to the pathology of laminitis and are possiblecandidates for targeted intervention. Although not con-sistently upregulated in our previous studies in acutelaminitis (Wang, 2010), the innate cytokines IL-1, IL-6,IFN�, and TNF� are potentially important given their majorregulatory roles in initiating and sustaining the immuneresponse. Similar to results seen in acute, carbohydrate-induced laminitis (Leise et al., 2011), we found that IL-1and IL-6, but not TNF�, were overexpressed in the laminartissue of foundered horses. The source of these cytokines isunknown, but could be laminar keratinocytes or infiltrat-ing phagocytes. No increase was found in laminar tissueIFN�, despite the elevation seen in circulating WBCs. How-ever, we also found no indication of increased numbers ofantigen presenting cells (macrophages, dendritic cells), Tcells, or B cells during chronic laminitis, as evidenced bythe expression levels of CD14, CD3, and CD19, respectively.RNA expression of cell markers is often used to assesstissue infiltration of inflammatory cells (Michalak et al.,2000; Ponnuswamy et al., 2012), although the possibilityremains that RNA level might not necessarily correlatewith cell number. In the present study, however, the lackof change in cell marker expression was supported byan absence of inflammatory cells visible in histologicalsections, with the exception of neutrophils. This is inter-esting in light of the fact that the monocyte/macrophagechemokine CCL2 was overexpressed in foundered horses,although the lymphocyte chemokines CCL4 and CCL7did not differ between groups. IL-8, the prototypicalneutrophil chemokine, was highly upregulated and, in thisstudy, was accompanied by the presence of neutrophilswithin the laminar tissue, as evidenced by both histologyand neutrophil elastase protein levels. With the exception
of a single horse, neutrophil infiltration appeared to beminimal, although similarly low numbers of laminarneutrophils have been found in both the developmentaland Obel grade I stages of experimental laminitis (Black
logy and
epctgspwlaiit
gttdob2tdicaemttdcduinpdSa(aSioti
5
mwtowmicac
S.M. Steelman et al. / Veterinary Immuno
t al., 2006). Given the ability of neutrophils to releaseroteinases, reactive oxygen species, and inflammatoryytokines, it is likely that these cells contribute to laminarissue damage during chronic laminitis. Indeed, someroups have hypothesized that neutrophils drive bothystemic and laminar inflammation during acute laminitis,articularly in the black walnut extract induction model,here they are likely the source of IL-1 (Belknap, 2010; de
a Rebiere de Pouyade and Serteyn, 2011). As neutrophilsre generally the first cell type to respond to injury ornfection, it is tempting to speculate that their presencen chronic laminitis is the result of repeated trauma to theissue from everyday weight-bearing activity.
The last class of genes examined, the damage-relatedenes, is an essential component of the epithelial responseo infection or trauma, particularly the antimicrobial pep-ides (AMPs). The major keratinocyte AMPs include theefensins (such as DEFB4) and PI3 (also known as SKALPr elafin), which defend against microbes by disruptingacterial cell membrane integrity (Wiesner and Vilcinskas,010). Interestingly, PI3 is also a serine protease inhibitorhat protects against excessive tissue damage by neutrophilegranulation. Both molecules are produced in response to
nflammation resulting from infection or trauma and arehemotactic for several types of immune cells (Wiesnernd Vilcinskas, 2010; Wilkinson et al., 2009). The over-xpression of these peptides could be a link between theechanical damage to the laminar tissue resulting from
he instability of the decompensated foundered foot andhe chronic inflammation characteristic of this disease. Theramatic upregulation of MMP13, a collagenase enzyme,ould also promote mechanical instability of the foot viaegradation of laminar collagen. We also found a modestpregulation of the antioxidant enzyme SOD2, suggest-
ng the occurrence of oxidative injury, perhaps from theeutrophil respiratory burst. Finally, the calcium-bindingrotein S100A9, but not its binding partner S100A8, wasramatically overexpressed in laminar tissue. S100A8 and100A9 are generally co-regulated in laminar keratinocytess part of the pro-inflammatory calprotectin complexFaleiros et al., 2009), although there is some evidence thatctivated neutrophils express S100A9 in the absence of100A8 (Kumar et al., 2001). Although immunohistochem-stry would be necessary to definitively localize expressionf S100A9 to neutrophils, it is possible that the upregula-ion of this gene in chronic laminitis is due to neutrophilnfiltration.
. Conclusion
Despite previous speculation that chronic laminitisight have an autoimmune component (Hood et al., 1990),e found no evidence to support this hypothesis, either in
he form of circulating immune complexes or expressionf the T cell marker CD3 within laminar tissue. Instead,e suggest a working hypothesis in which laminar inflam-ation results from the keratinocytes themselves and
nfiltrating neutrophils, likely in response to continual sub-linical injury sustained during everyday weight-bearingctivity. The current study provides evidence that spe-ific targeting of keratinocyte mediators of inflammation
Immunopathology 153 (2013) 217– 226 225
or blockade of neutrophil infiltration might provide a moreeffective approach to limiting pain and inflammation infoundered horses.
Acknowledgements
The authors wish to thank Dr. David Hood of the HoofDiagnostic and Rehabilitation Clinic and Dr. Carolyn Arnoldof the Texas A&M Veterinary Medical Teaching Hospitalfor access to samples and Dr. Jianrong Li for use of lab-oratory equipment. This research was supported in partby the United States Department of Agriculture (award #2011-67012-30685 to SMS and 2010-65205-20446 to BPC).
Appendix A. Supplementary data
Supplementary data associated with this article can befound, in the online version, at http://dx.doi.org/10.1016/j.vetimm.2013.03.001.
References
Aho, K., Heliovaara, M., Knekt, P., Reunanen, A., Aromaa, A., Leino, A., Kurki,P., Heikkila, R., Palosuo, T., 1997. Serum immunoglobulins and the riskof rheumatoid arthritis. Ann. Rheum. Dis. 56, 351–356.
Badolato, R., Oppenheim, J.J., 1996. Role of cytokines, acute-phase pro-teins, and chemokines in the progression of rheumatoid arthritis.Semin. Arthritis Rheum. 26, 526–538.
Belknap, J.K., 2010. Black walnut extract: an inflammatory model. TheVeterinary clinics of North America. Equine Pract. 26, 95–101.
Black, S.J., Lunn, D.P., Yin, C., Hwang, M., Lenz, S.D., Belknap, J.K., 2006.Leukocyte emigration in the early stages of laminitis. Vet. Immunol.Immunopathol. 109, 161–166.
Budak, M.T., Orsini, J.A., Pollitt, C.C., Rubinstein, N.A., 2009. Gene expres-sion in the lamellar dermis-epidermis during the developmentalphase of carbohydrate overload-induced laminitis in the horse. Vet.Immunol. Immunopathol. 131, 86–96.
Carter, R.A., Engiles, J.B., Megee, S.O., Senoo, M., Galantino-Homer, H.L.,2011. Decreased expression of p63, a regulator of epidermal stem cells,in the chronic laminitic equine hoof. Equine Vet. J. 43, 543–551.
Chen, J., Bruns, A.H., Donnelly, H.K., Wunderink, R.G., 2010. Comparativein vitro stimulation with lipopolysaccharide to study TNFalpha geneexpression in fresh whole blood, fresh and frozen peripheral bloodmononuclear cells. J. Immunol. Methods 357, 33–37.
Davis 3rd, J.M., Knutson, K.L., Strausbauch, M.A., Crowson, C.S., Therneau,T.M., Wettstein, P.J., Matteson, E.L., Gabriel, S.E., 2010. Analysis of com-plex biomarkers for human immune-mediated disorders based oncytokine responsiveness of peripheral blood cells. J. Immunol. 184,7297–7304.
de la Rebiere de Pouyade, G., Serteyn, D., 2011. The role of activated neu-trophils in the early stage of equine laminitis. Vet. J. 189, 27–33.
Ethuin, F., Gerard, B., Benna, J.E., Boutten, A., Gougereot-Pocidalo, M.A.,Jacob, L., Chollet-Martin, S., 2004. Human neutrophils produce inter-feron gamma upon stimulation by interleukin-12. Lab. Invest. 84,1363–1371.
Faleiros, R.R., Nuovo, G.J., Belknap, J.K., 2009. Calprotectin in myeloid andepithelial cells of laminae from horses with black walnut extract-induced laminitis. J. Vet. Intern. Med. 23, 174–181.
Faleiros, R.R., Stokes, A.M., Eades, S.C., Kim, D.Y., Paulsen, D.B., Moore, R.M.,2004. Assessment of apoptosis in epidermal lamellar cells in clinicallynormal horses and those with laminitis. Am. J. Vet. Res. 65, 578–585.
Fouret, P., du Bois, R.M., Bernaudin, J.F., Takahashi, H., Ferrans, V.J., Crystal,R.G., 1989. Expression of the neutrophil elastase gene during humanbone marrow cell differentiation. J. Exp. Med. 169, 833–845.
Hodge, G., Hodge, S., Han, P., 2000. Increased levels of apoptosis of leuko-cyte subsets in cultured PBMCs compared to whole blood as shownby Annexin V binding: relevance to cytokine production. Cytokine 12,
1763–1768.Hood, D.M., 1999. The pathophysiology of developmental and acutelaminitis. Vet. Clin. North Am. Equine Pract. 15, 321–343.
Hood, D.M., Amoss, M.S., Grosenbaugh, D.A., 1990. Equine laminitis: apotential model of Raynaud’s phenomenon. Angiology 41, 270–277.

logy and
226 S.M. Steelman et al. / Veterinary ImmunoHunt, R.J., Wharton, R.E., 2010. Clinical presentation, diagnosis, and prog-nosis of chronic laminitis in North America. Vet. Clin. North Am.Equine Pract. 26, 141–153.
Johnson, P.J., Ganjam, V.K., Slight, S.H., Kreeger, J.M., Messer, N.T., 2004.Tissue-specific dysregulation of cortisol metabolism in equine lamini-tis. Equine Vet. J. 36, 41–45.
Kumar, R.K., Yang, Z., Bilson, S., Thliveris, S., Cooke, B.E., Geczy, C.L., 2001.Dimeric S100A8 in human neutrophils is diminished after phagocy-tosis. J. Leukoc. Biol. 70, 59–64.
Kuwano, A., Ueno, T., Katayama, Y., Nishiyama, T., Arai, K., 2005. Uni-lateral basement membrane zone alteration of the regeneratedlaminar region in equine chronic laminitis. J. Vet. Med. Sci. 67,685–691.
Leise, B.S., Faleiros, R.R., Watts, M., Johnson, P.J., Black, S.J., Belknap, J.K.,2011. Laminar inflammatory gene expression in the carbohydrateoverload model of equine laminitis. Equine Vet. J. 43, 54–61.
Michalak, T.I., Hodgson, P.D., Churchill, N.D., 2000. Posttranscriptionalinhibition of class I major histocompatibility complex presentationon hepatocytes and lymphoid cells in chronic woodchuck hepatitisvirus infection. J. Virol. 74, 4483–4494.
Mieli-Vergani, G., Vergani, D., 2011. Autoimmune hepatitis. Nat. Rev. Gas-troenterol. Hepatol. 8, 320–329.
Monteiro, R.C., 2010. Role of IgA and IgA fc receptors in inflammation. J.Clin. Immunol. 30, 1–9.
Moore, R.M., 2010. Vision 20/20 – conquer laminitis by 2020. J. Equine Vet.Sci. 30, 74–76.
Morrison, S., 2010. Chronic laminitis: foot management. Vet. Clin. NorthAm. Equine Pract. 26, 425–446.
Noschka, E., Vandenplas, M.L., Hurley, D.J., Moore, J.N., 2008. Temporalaspects of laminar gene expression during the developmental stagesof equine laminitis. Vet. Immunol. Immunopathol.
Obel, N.J., 1948. Studies on the histopathology of acute laminitis. Disser-tation. Swedish University of Agricultural Sciences.
Orsini, J.A., Wrigley, J., Riley, P., 2010. Home care for horses with chroniclaminitis. Vet. Clin. North Am. Equine Pract. 26, 215–223.
Osborne, A.C., Carter, S.D., May, S.A., Bennett, D., 1995. Anti-collagen anti-bodies and immune complexes in equine joint diseases. Vet. Immunol.Immunopathol. 45, 19–30.
Pabst, O., 2012. New concepts in the generation and functions of IgA. Nat.Rev. Immunol. 12, 821–832.
Pfaffl, M.W., 2001. A new mathematical model for relative quantificationin real-time RT-PCR. Nucleic Acids Res. 29, e45.
Platts-Mills, T.A., 2001. The role of immunoglobulin E in allergy and
asthma. Am. J. Respir. Crit. Care Med. 164, S1–S5.Pollitt, C.C., 1996. Basement membrane pathology: a feature of acuteequine laminitis. Equine Vet. J. 28, 38–46.
Ponnuswamy, P., Schrottle, A., Ostermeier, E., Gruner, S., Huang, P.L., Ertl,G., Hoffmann, U., Nieswandt, B., Kuhlencordt, P.J., 2012. eNOS protects
Immunopathology 153 (2013) 217– 226
from atherosclerosis despite relevant superoxide production by theenzyme in apoE mice. PLoS ONE 7, e30193.
Rose, N.R., Bona, C., 1993. Defining criteria for autoimmune dis-eases (Witebsky’s postulates revisited). Immunol. Today 14,426–430.
Slater, M.R., Hood, D.M., Carter, G.K., 1995. Descriptive epidemiologicalstudy of equine laminitis. Equine Vet. J. 27, 364–367.
Stanek, C., Mayr, B., Graninger, W., Hofmann, R., 1984. Immune complexesin the peripheral blood of healthy horses and horses with laminitis.Berl. Munch. Tierarztl. Wochenschr. 97, 25–28.
Steelman, S.M., Chowdhary, B.P., 2012. Plasma proteomics shows an ele-vation of the anti-inflammatory protein APOA-IV in chronic equinelaminitis. BMC Vet. Res. 8, 179.
USDA, 2000. Lameness and laminitis in U.S. horses. United States Depart-ment of Agriculture Report #N318.0400.
Van Epps, D.E., Williams Jr., R.C., 1976. Suppression of leukocytechemotaxis by human IgA myeloma components. J. Exp. Med. 144,1227–1242.
Wagner, B., Radbruch, A., Rohwer, J., Leibold, W., 2003a. Monoclonalanti-equine IgE antibodies with specificity for different epitopeson the immunoglobulin heavy chain of native IgE. Vet. Immunol.Immunopathol. 92, 45–60.
Wagner, I.P., Rees, C.A., Dunstan, R.W., Credille, K.M., Hood, D.M., 2003b.Evaluation of systemic immunologic hyperreactivity after intrader-mal testing in horses with chronic laminitis. Am. J. Vet. Res. 64,279–283.
Wang, J., 2010. Genome-wide transcriptome analysis of laminar tissueduring the early stages of experimentally induced equine laminitis.Dissertation. Texas A&M University.
Wiesner, J., Vilcinskas, A., 2010. Antimicrobial peptides: the ancient armof the human immune system. Virulence. 1, 440–64.
Wilkinson, T.S., Dhaliwal, K., Hamilton, T.W., Lipka, A.F., Farrell, L.,Davidson, D.J., Duffin, R., Morris, A.C., Haslett, C., Govan, J.R., Greg-ory, C.D., Sallenave, J.M., Simpson, A.J., 2009. Trappin-2 promotesearly clearance of Pseudomonas aeruginosa through CD14-dependentmacrophage activation and neutrophil recruitment. Am. J. Pathol. 174,1338–1346.
Wilton, J.M., 1978. Suppression by IgA of IgG-mediated phagocytosisby human polymorphonuclear leucocytes. Clin. Exp. Immunol. 34,423–428.
Wolf, H.M., Fischer, M.B., Puhringer, H., Samstag, A., Vogel, E., Eibl, M.M.,1994a. Human serum IgA downregulates the release of inflamma-tory cytokines (tumor necrosis factor-alpha, interleukin-6) in human
monocytes. Blood 83, 1278–1288.Wolf, H.M., Vogel, E., Fischer, M.B., Rengs, H., Schwarz, H.P., Eibl, M.M.,1994b. Inhibition of receptor-dependent and receptor-independentgeneration of the respiratory burst in human neutrophils and mono-cytes by human serum IgA. Pediatr. Res. 36, 235–243.