universidade federal do amapá pró-reitoria de … with the increase of the concentration of grains...
TRANSCRIPT

1
Universidade Federal do Amapá
Pró-Reitoria de Pesquisa e Pós-Graduação
Programa de Pós-Graduação em Biodiversidade Tropical
Mestrado e Doutorado
UNIFAP / EMBRAPA-AP / IEPA / CI-Brasil
ANTONIO FERREIRA DE OLIVEIRA
Estudo da viabilidade da produção de biofilmes de kefir e suas interações com
extratos de açaí (Euterpe oleracea Martius) e de gérmen de soja (Glycine max
(L.) Merrill)
MACAPÁ, AP
2016

2
ANTONIO FERREIRA DE OLIVEIRA
Estudo da viabilidade da produção de biofilmes de kefir e suas interações com
extratos de açaí (Euterpe oleracea Martius) e de gérmen de soja (Glycine max
(L.) Merrill)
Tese apresentada ao Programa de Pós-
Graduação em Biodiversidade Tropical da
Universidade Federal do Amapá (UNIFAP),
como parte dos requisitos para obtenção do
título de Doutor em Biodiversidade
Tropical.
Linha de Pesquisa: Uso sustentável da
biodiversidade.
Orientador: Prof. Tit. José Carlos Tavares
Carvalho
MACAPÁ, AP
2016

3
Dados Internacionais de Catalogação na Publicação (CIP) Biblioteca Central da Universidade Federal do Amapá
502.82
O48e Oliveira, Antonio Ferreira.
Estudo da viabilidade da produção de biofilmes de kefir e suas
interações com extratos de açaí (Euterpe oleracea Martius) e de gérmen
de soja (Glycine max(L.) Merrill) / Antonio Ferreira Oliveira;
orientador, José Carlos Tavares Carvalho. – Macapá, 2016. 180 f.
Tese (doutorado) – Fundação Universidade Federal do Amapá,
Programa de Pós-Graduação em Biodiversidade Tropical.
1. Biofilmes. 2. Kefir. 3. Microscopia de força atômica (AFM) I.
Carvalho, José Carlos Tavares, orientador. II. Fundação Universidade
Federal do Amapá. III. Título.

4
ANTONIO FERREIRA DE OLIVEIRA
Estudo da viabilidade da produção e padronização de biofilmes de kefir e suas
interações com extratos de açaí (Euterpe oleracea Martius) e de gérmen de soja
(Glycine max (L.) Merrill)
_________________________________________
Prof. Tit. José Carlos Tavares Carvalho
(Orientador)
Universidade Federal do Amapá (UNIFAP)
____________________________________________
Dra. Elizabeth Portal Viana
Universidade Federal do Amapá (UNIFAP)
____________________________________________
Dr. Rodrigo Alves Soares Cruz
Universidade Federal do Amapá (UNIFAP)
____________________________________________
Dr. Caio Pinho Fernandes
Universidade Federal do Amapá (UNIFAP)
____________________________________________
Dr. Robert Maguiña Zamora
Universidade Federal do Amapá (UNIFAP)
Aprovada em 26 de Agosto de 2016, Macapá, AP, Brasil

5
Dedico este trabalho ao meu melhor Amigo
e companheiro dentro do meu coração, pois
sem as orações ouvidas e atendidas por Ele
nada disso teria sido possível

6
AGRADECIMENTOS
Agradeço ao meu Orientador pelo direcionamento, apoio e empurrões para que eu seguisse
em frente.Seu exemplo de pesquisador competente e dedicação e inestimável auxílio tornaram
possível a cosecução desta pesquisa.
À Universidade Federal do Amapá através do Programa de Pós-Graduação em
Biodiversidade Tropical (PPGBIO).
Ao CNPq pela bolsa de doutorado concedida nos dois últimos anos.
Agradeço à técnica do Laboratório de Pesquisa em Fármacos e amiga Adriana Maciel pelo
apoio e toda ajuda prestada para a construção da tese.
Agradeço aos professores que contribuíram significativamente: Dr Alexandro Cezar
Florentino, Dr Caio Pinho Fernandes, Dr Irlon Maciel Ferreira, Dr Jesus Rodrigues Amado, Dr
Robert Maquiña Zamora, Dr Roberto Messias Bezerra, Dr Rodrigo Alves Soares Cruz.
Aos companheiros do Laboratório de Pesquisa em Fármacos, em especial ao Jonatas
Lobato Hélison e Jéssica Vilhena e Beatriz de Sá pelo apoio cientifico e pelas palavras de
incentivo.
Ao pesquisador Mauricio Abdon (IEPA), o qual contribuiu com a análise estatística.
Aos meus irmãos que, mesmo distante, me incentivaram, e especialmente ao meu irmão
Carlos Fernandes e a minha irmã Regina que me apoiaram, advindo pessoalmente ao Amapá
várias vezes com o intuito de me auxiliar.
Enfim a todos que me ajudaram direta e indiretamente que, eventualmente, não tenham sido
citados, o meu muito obrigado.

7
“Há de a esperança transmudar-se em alegre fruto, a
fé em realidade,
a oração em louvor!”

8
RESUMO
Oliveira, Antonio Ferreira. Estudo da produção de biofilmes de kefir em diferentes substratos, e
suas interações com extratos de açaí (Euterpe oleracea Martius) e de gérmen de soja (Glycine
max (L.) Merrill). Macapá, 2016. Tese (Doutorado em Biodiversidade Tropical) – Programa de
Pós-graduação em Biodiversidade Tropical – Pró-Reitoria de Pesquisa e Pós-Graduação -
Universidade Federal do Amapá.
O kefir é um probiótico composto por microrganismos que vivem em simbiose principalmente
Lactobacillus e leveduras. O objetivo deste estudo foi padronizar a produção de biofilmes de
kefir em solução de açúcar mascavo, açúcar branco e aferir suas interações com extrato de açaí e
de gérmen de soja. Para tanto os grãos de kefir foram inoculados em meio de cultura com
diferentes concentrações açúcar mascavo (AM) e açúcar branco (AB). Após 20 dias, foram
aferidas as biomassas dos biofilmes formados. Foi também determinada a composição de macro e
microelementos dos biofilmes e sua estrutura foi analisada usando microscopia ótica,
microscopia de força atômica (AFM) e microscopia eletrônica de varredura (MEV). A
concentração necessária de substrato para formação de biofilme com o açúcar mascavo foi de 40
g/L e a concentração grãos de kefir foi de 20 g/L. Com relação ao açúcar branco a melhor
concentração de substrato foi de 20 g/L e a concentração de grãos de kefir foi de 20 g/L. Quanto
a composição de macro e microelementos dos biofilmes de kefir a concentração de Fe++ foi de
2,725 mg/L em AM e 1,570 mg/L em AB. O Ca++ foi maior nos biofilmes formados em AB
(2,150 mg/L) do que nos formados em AM (0,550 mg/L). A análise por AFM revelou diferença
de rugosidade à medida que se aumentou a concentração de grãos de kefir no meio e a
microscopia eletrônica de varredura (MEV) mostrou a estrutura do biofilme bem como a adesão
de bactérias às hifas. Para se obter biofilmes de kefir com extrato de açaí (EHEEo) diferentes
concentrações de EHEEo, de grãos de kefir e açúcar mascavo foram testadas. Após 20 dias foi
aferida a biomassa dos biofilmes e analisada a estrutura desses biofilmes através AFM e MEV.
Buscou-se confirmar a incorporação de antocianinas nestes biofilmes, a composição de macro e
microelementos e a rugosidade (RMS) com o uso de AFM. Procedeu-se a dosagem de
antocianinas tanto nos biofilmes como no meio. A melhor concentração de açúcar mascavo para a
formação de biofilme foi de 40 g/L com 100 mL de EHEEo (12,62+/-0,1626). de fungos. As
antocianinas se mostraram estáveis em biofilmes por pelo menos 26 meses (18 µg/100g).
Diferentes concentrações de grãos de kefir e de gérmen de soja foram cultivados em meio de
cultura com solução de açúcar mascavo (40g/L) por 20 dias quando os biofilmes foram retirados
e as isoflavonas foram extraídas dos biofilmes e quantificadas através de cromatografia líquida de
alta eficiência (CLAE). A melhor concentração de grãos de kefir foi 40 g/L com 20 g/L de
gérmen de soja e o doseamento de isoflavonas foi de 18 µg/L para gliciteína e para genisteína 22
µg/L. A análise por AFM revelou uniformidade da rugosidade à medida que se aumentou a
concentração de gérmen de soja no meio e a microscopia eletrônica de varredura (MEV) mostrou
a partículas adesivadas hifas e a estrutura do biofilme. A fusão das propriedades do kefir com
compostos bioativos abre um potencial de aplicação terapeutica considerável.
Palavras-chave: Biofilmes. Kefir. Lactobacillus. Leveduras. Microscopia de Força Atômica
(AFM). Açaí. Euterpe oleracea Mart. Antocianina. Gérmen de Soja. Isoflavonas.

9
ABSTRACT
Oliveira, Antonio Ferreira. Estudo da viabilidade da produção de biofilmes de kefir e suas
interações com extratos de açaí (Euterpe oleracea Martius) e de gérmen de soja (Glycine max
(L.) Merrill). Macapa, 2016. Thesis (Doctorate in Tropical Biodiversity) - Postgraduate in
Tropical Biodiversity Program - Dean of Research and Graduate Studies - Federal University of
Amapá.
Kefir is a probiotic composed by microorganisms that live in symbiosis mainly Lactobacillus and
yeast. This study has objective the padronization of biofilms of kefir in brown and white sugar
solution and his interactions with açai extract (Euterpe oleracea Mart.) and soybean extract
(Glycine max (L.) Merril). They were formed tree kinds of biofilms: kefir sugar (boh brown ans
white sugar), kefir with sugar and açai and kefir sugar and soybean. Kefir grains were inoculated
in the culture medium with different concentration of brown sugar (BS) and white sugar (WS).
After twenty days the biomass of the biofilms were weighted. It was also determined the
composition of macro and micro elements of biofilms and its structure was analyzed using optical
microscopy (OM), atomic force microscopy (AFM) and scanning electron microscopy (SEM).
The concentration of white sugar substrate was 20 g/L and the concentration of kefir grains was
20 g/L. The concentration of brown sugar to form biofilm was 40 g/L and the concentration of
kefir grains was 20 g/L. The composition of macro and microelements kefir were iron 2,725
mg/L in BS solution and 1570 mg/L in WS solution. The calcium was higher in biofilms WS
(2,150 mg/L) than those formed in BS (0.550 mg / L). Analysis by AFM revealed a difference of
roughness with the increase of the concentration of grains of kefir in the culture medium.
Scanning electron microscopy (SEM) showed the structure of the biofilm and bacterial adhesion
to fungal hyphae. For kefir biofilms with acai extract (EHEEo), different concentrations of
EHEEo, kefir grains and brown sugar were tested. After 20 days, it was measured biomass of
biofilms and the structure of the biofilms were analyzed by AFM and SEM. The incorporation of
anthocyanins in these biofilms was confirmed and macro and microelements composition were
determined . The surface roughness (RMS) was studied using AFM. The dosage of anthocyanins
in both the biofilm and in the medium was determinated. The concentration of brown sugar to
form biofilm was 40 g/L with 100 mL/L of EHEEo (12.62 +/- 0.1626 g). Anthocyanins in
biofilms were stable for at least 26 months (18 mg / 100 g). Different kefir grains concentrations
and soy germ were cultivated in culture medium with brown sugar solution (40g/L) for 20 days
when biofilms were removed and isoflavones were extracted from the biofilms and quantified by
high performance liquid chromatography (HPLC). The concentration of kefir grains was 40 g/L
and soy germ was 20 g/L and determination of isoflavones was 18 µg/L of genistein and glycitein
22 µg/L. The merger of kefir properties with bioactive compounds opens a significant therapeutic
application potential.
Key Words: Biofilms. Kefir. Lactobacillus. Yeast. Probiotic. Atomic Force Microscopy (AFM).
Açaí. Euterpe oleracea Mart.. Anthocyanins.

10
LISTA DE ILUSTRAÇÕES
Figura 1 - Estrutura do kefirano....................................................................................... 32
Figura 2 - Diagrama de funcionamento do microscópio de varredura de força............... 49
Figura 3 - Estrutura do Núcleo Flavílico........................................................................... 56
Figura 4 - Principais antocianinas encontradas nas frutas................................................. 57
Figura 5 - Principais antocianinas encontradas nos alimentos.......................................... 58
Figura 6 - Mapa conceitual apresentando as propriedades e os principais compostos
bioativos do extrato de açaí com a incorporaração de antocianina do açaí (Euterpe
oleracea Mart.) em biofilme de kefir.................................................................................
62
CAPÍTULO 1 - ESTUDOS DA VIABILIDADE DE OBTENÇÃO E
PADRONIZAÇÃO DE BIOFILMES DE KEFIR
Figura 1 - Biofilme de kefir em açúcar mascavo 40 g/L com concentração dos grãos de
kefir de 20 g/L....................................................................................................................
104
Figura 2 - Biofilme de kefir (20 g/L) em açúcar mascavo (40 g/L). Observa-se os biofilmes
formados no fundo do frasco se expandindo para a superfície.......................................................
104
Figura 3 - Meio de cultura de kefir (20 g/L) em açúcar mascavo (40 g/L). a) Material
aglutinado no fundo do béquer e b) Grãos de kefir adesivados por substância
aglutinante............................................................................................................................
104
Figura 4 - Foto de biofilme de kefir (20g/L) com açúcar mascavo (40 g/L) através de
microscopia óptica com resolução 400x mostrando a estrutura de formação de biofilmes e a
ausência de porosidade do biofilme................................................................................................
107
Figura 5 - Imagem topógráfica de biofilmes de kefir através de microscopia de força
atômica (AFM). Concentraçao de GK a) 20 g/L, b) 60 g/L e c) 80 g/L............................
107
Figura 6 - Imagem topográfica de biofilme de kefir com açúcar mascavo obtida através de
microscopia de varredura de força (AFM). Concentração AM 40 g/L e GK 20 g/L. Observa-se
uniformidade do tecido do biofilme e a rugosidade........................................................................
108
Figura 7 - Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (20 g/L) em
açúcar mascavo (40 g/L) visualizado de perfil onde se observa as camadas de
exopolissacarídeo que se sobrepõem formando o biofilme. Dados de magnificação são
mostrados na figura.........................................................................................................................
108

11
Figura 8 – Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (20 g/L) em
açúcar mascavo (40 g/L) onde se observa as bactérias adesivada às hifas de fungos.
Magnificação de a) 800x b)2000x e c) 3000x.............................................................................
1088
CAPÍTULO 2 - ESTUDO DE BIOFILME DE KEFIR ASSOCIADO AO EXTRATO
HIDROETANÓLICO DE Euterpe oleracea MART. (AÇAÍ). UM POSSÍVEL
PROBIÓTICO DA AMAZÔNIA
Figura 1 - Biofilmes de kefir com extrato hidroetanólico de Euterpe oleracea (BEHEEo)
formado com grão de kefir (GK) 20 g/L, açúcar mascavo, 40 g/L e extrato de açaí (100 mL).
Biofilme retirado do meio de cultura (1a) e biofilme liofilizado (1b)...........................
132
Figura 2 - Imagem gráfica através de microscopia de força atômica (AFM) de biofilme de
kefir a) Imagem gráfica da superfície do biofilme quando utilizado 20 g/L de grãos de kefir e
40 g/L açúcar mascavo. b) Imagem topográfica do biofilme quando utilizado 20 g/L de
grãos de kefir e 40 g/L açúcar mascavo. c) Imagem gráfica da superfície do biofilme
quando utilizado 60 g/L de grãos de kefir e 40 g/L açúcar mascavo. d) Imagem
topográfica do biofilme quando utilizado 60 g/L de grãos de kefir e 40 g/L açúcar
mascavo.Em todo este experimento ao meio de cultura foi adicionado 100 g
EHEEo................................................................................................................................
137
Figura 3 - Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (20 g/L) com
açúcar mascavo (40 g/L) e açaí (100 g/L) (BEHEEo) obtida com resolução 2500x onde se
observa a presença de emaranhado de hifas................................................................................
138
CAPÍTULO 3 - ESTUDO DO BIOFILME DE KEFIR ASSOCIADO AO GÉRMEN DE
SOJA (Glycine max L. Merril)
Figura 1 - Perfil cromatográfico de agliconas padrão (Sigma Aldrich) através de
cromatografia líquida de alta eficiência (CLAE) para gliciteína e genisteína com tempo
de retenção 9,2 minutos e 18,7 min. respectivamente........................................................
159
Figura 2 - Perfil cromatográfico obtido por CLAE composto padrão extrato de gérmen
de soja para gliciteína e genisteína com tempo de retenção 9,60 minutos e 18,50 min.
respectivamente..................................................................................................................
159
Figura 3 - Perfil cromatográfico do meio de cultura com kefir 20 g/L com açúcar
mascavo 40 g/L, mais extrato de gérmen de soja a 10 g/L obtido através de
cromatografia líquida de alta eficiência (CLAE) para gliciteína e genisteína com tempo

12
de retenção 9,8 minutos e 19,8 min. respectivamente........................................................ 161
Figura 4 - Foto de biofilmes de kefir (20 g/L) com açúcar mascavo (40 g/L) com gérmen de
soja a) 20 g/L, b) 40 g/L, c) 80 g/L e d) 100 g/L............................................................................
162
Figura 5 - Perfil cromatográfico obtido por CLAE de kefir mas extrato de gérmen de
soja cultivado por 5 dias para gliciteína e genisteína com tempo de retenção 9,60
minutos e 18,50 min. respectivamente...............................................................................
163
Figura 6 - Imagem gráfica obtida através de microscopia de força atômica (AFM) mostrando
a rugosidade (0,287 µm) de biofilme de kefir formado 2 g/L de gérmen de soja. Observa-se a
presença de Lactobacillus...............................................................................................................
Figura 7 - Imagem gráfica de biofilme de kefir com gérmen de soja (10g/L) obtida com
microscopia de força atômica (AFM). O aumento da concentração não alterou a rugosidade (0,
283 µm)...........................................................................................................................................
Figura 8 - Imagem gráfica de biofilme de kefir com gérmen de soja (80g/L) obtida com
microscopia de força atômica (AFM). A rugosidade RMS (0,167 µm) naõ foi alterada
significativamente com o aumento da concentração de gérmen de soja.........................................
Figura 9 -Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (20 g/L) com
açúcar mascavo (40 g/L) e gérmen de soja (20g/L) visualizada de perfil onde se observa as
camadas que formam os biofilmes se sobrepondo..........................................................................
Figura 10 - Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (20 g/L) com
açúcar mascavo (40 g/L) com gérmen de soja (20 g/L) onde se visualiza hifa com moléculas
adesivadas à mesma........................................................................................................................
164
164
165
165
166

13
LISTA DE TABELAS
CAPÍTULO 1 - ESTUDO DA OBTENÇÃO DE BIOFILMES DE KEFIR EM
DIFERENTES SUBSTRATOS
Tabela 1 - Formação de biofilmes em diferentes concentrações de grãos de kefir e
substratos (açúcar mascavo e açúcar branco).....................................................................
101
Tabela 2 - Formação de biofilmes em função da concentração de açúcar branco e
mascavo..............................................................................................................................
102
Tabela 3 - Concentração de macro e microelementos presentes em grãos de kefir e em
biofilmes e obtidos em açúcar mascavo e açúcar branco...................................................
105
Tabela 4 - Compostos voláteis majoritários encontrados no meio de cultura do kefir..... 106
CAPÍTULO 2 - ESTUDO DE BIOFILME DE KEFIR ASSOCIADO AO EXTRATO
HIDROETANÓLICO DE Euterpe oleracea MART. (AÇAÍ). UM POSSÍVEL
PROBIÓTICO DA AMAZÔNIA
Tabela 1 - Formação de BEHEEo associado ao EHEEo em diferentes concentrações de
GK in AM (40 g/L)............................................................................................................
131
Tabela 2 - Formação de BEHEEo associado ao EHEEo em diferentes concentrações de
AM com GK (20 g/L).........................................................................................................
133
Tabela 3 - Concentração de antocianinas no BEHEEo formado com grãos de kefir GK
(20 g/L) com AM (40 g/L) em diferentes concentrações de EHEEo.................................
134
Tabela 4 - Perfil de liberação e dissolução de antocianina do BEHEEo........................... 134
Tabela 5 - Quantificação da concentração de antocianinas in BEHEEo em diferentes
períodos..............................................................................................................................
135
Tabela 6 - Concentração de macro e microelementos do EHEEo e do
BEHEEo.............................................................................................................................
135
Tabela 7 - Compostos voláteis majoritários encontrado no kefir com EHEEo................. 136
Tabela 8 - Efeito da temperatura sobre a síntese de BEHEEo................................... 139

14
CAPÍTULO 3 - ESTUDO DO BIOFILME DE KEFIR ASSOCIADO AO GÉRMEN DE
SOJA (Glycine max L. Merril)
Tabela 1 - Formação do biofilme com GK (20 g/L) e AM (40 g/L) em diferentes
concentrações de gérmen de soja (GS)...............................................................................
160
Tabela 2 - Formação do biofilme com GS (40 g/L) e AM (40 g/L) em diferentes
concentrações de grãos de kefir (GK)................................................................................
161
Tabela 3 - Determinação de período de tempo necessário para formação de biofilme e
liberação de isoflavonas do biofilme de kefir com germen de soja...................................
162
Tabela 4 - Concentração de macro e microelementos do biofilme de kefir com gérmen
de soja.................................................................................................................................
163

15
ABREVIATURAS E SIGLAS
°C Graus Celsius
°GL Grau Gay-Lussac
µ Micrômetro
µg Microgramas
µL Microlitro
AB Açúcar branco
AFM Atomic Force Microscopy ou Microscopia de Força Atômica
AI Índice Aritmético
AM Açúcar mascavo
AOAC Association of Official Analytical Chemists
BEHEEo Biofilme de extrato hidroetanólico de Euterpe oleracea
CG Cromatografia gasosa
CG-MS Cromatografia gasosa acoplada a espectrometria de massa
CLAE Cromatografia Líquida de Alta Eficiência
D Dimensões
EHEEo Extrato hidroetanólico de Euterpe oleracea
eV Eletrónvolt
g Grama
GK Grãos de kefir
kg kilograma
KHz Kilohertz

16
L Litro
M Molar
m Metro
MEV Microscopia Eletrônica de Varredura
mg miligrama
min minuto
mL mililitro
mm milimetro
mmol milimol
nN nanoNewtons
p/v peso por volume
PA Puro para análise
pH Potencial hidrogeniônico
Rm Rugosidade média
RMS rugosidade
rpm Rotações por minuto

17
SUMÁRIO
1 INTRODUÇÃO GERAL ................................................................................................ 210
2 REVISÃO BIBLIOGRÁFICA ....................................................................................... 243
2.1. KEFIR ..................................................................................................................... 24
2.1.1. Efeito probiótico do kefir .................................................................................. 25
2.1.2 Compostos voláteis do meio de cultura ............................................................ 26
2.1.3 Processos fermentativvos de kefir e produção de etanol ............................... 28
2.1.4 A associação simbiótica do kefir ....................................................................... 28
2.1.5 O kefirano ........................................................................................................... 31
2.1.5.1 Kefirano: possíveis aplicações .......................................................................... 35
2.1.5.2 Utilização do kefirano como biopolímero ......................................................... 36
2.1.5.3 Fatores para a produção de kefirano .................................................................. 37
2.2 BIOFILMES MICROBIANOS ............................................................................... 38
2.2.1 Aplicações de biofilmes ...................................................................................... 43
2.2.2. Biofilmes e biorremediação ............................................................................. 43
2.2.2.1 Biorremediação de pesticidas ............................................................................ 42
2.2.3 Forças de adesão envolvidas em síntese de biofilmes ...................................... 44
2.2.4 Propriedades reológicas dos biopolímeros ....................................................... 47
2.2.5 Métodos físicos para estudo dos biofilmes ....................................................... 48
2.2.5.1 Utilização de microscopia de força atômica (AFM) ......................................... 48
2.2.5.2 Microscopia Eletrônica de Varredura (MEV) ................................................... 51
2.3 COMPOSTOS FENÓLICOS E SUA IMPORTÂNCIA ........................................ 521
2.4 FLAVONÓIDES ..................................................................................................... 55
2.5 ANTOCIANINAS ................................................................................................... 57
2.6. O GÊNERO EUTERPE .......................................................................................... 61
2.6.1 O açaizeiro (Euterpe oleracea Mart.) ............................................................... 62
2.6.1.1 Classificação de extrato de açaí (Euterpe oleracea Mart.) de acordo com o pH64
2.6.1.2 Açaí (Euterpe oleracea Mart.) e fotoproteção .................................................. 64
2.6.1.3 Componentes do extrato de açaí (Euterpe oleracea MART.) ........................... 64
2.7 ISOFLAVONAS PRESENTES EM GÉRMEN DE SOJA ...................................... 65
REFERÊNCIAS ................................................................................................................. 668
3 OBJETIVOS ...................................................................................................................... 91
3.1 GERAL .................................................................................................................... 91
3.2 ESPECÍFICOS ......................................................................................................... 91
CAPÍTULO 1: VIABILIDADE DE OBTENÇÃO E PADRONIZAÇÃO DE
BIOFILMES DE KEFIR .................................................................................................... 93
RESUMO ............................................................................................................................... 94
ABSTRACT...........................................................................................................................94
1 INTRODUÇÃO ................................................................................................................. 95
2 MATERIAL E MÉTODOS .............................................................................................. 98
2.1 Seleção da concentração de grãos de kefir ................................................................... 98
2.2 Determinação da concentração de substrato para a formação de biofilme ............ 987
2.3 Controle da temperatura para produção de biofilme de kefir ................................... 98
2.4 Doseamento de macro e microelementos nos grãos de kefir e biofilme por
espectrometria de absorção atômica ................................................................................ 998

18
2.5 Análise dos compostos voláteis do meio de cultura do kefir por cromatografia
gasosa acoplada a espectrometria de massa (CG-EM) ..................................................... 99
2.6 Análise estrutural de biofilmes de kefir por microscopia óptica e por AFM .......... 100
2.7 Análise estrutural de biofilmes de kefir por microscopia eletrônica de varredura
(SEM) ................................................................................................................................... 100
2.8 Análise estatística .......................................................................................................... 100
3. RESULTADOS ................................................................................................................ 101
3.1 Seleção da concentração de grãos de kefir ................................................................. 101
3.2 Determinação da concentração de substrato para a formação de biofilme ............ 103
3.3 Controle da temperatura para produção de biofilme de kefir ................................. 105
3.4 Doseamento de macro e microelementos nos grãos de kefir e biofilme por
espectrometria de absorção atômica ................................................................................. 106
3.5 Análise dos compostos voláteis do meio de cultura do kefir por cromatografia
gasosa acoplada a espectrometria de massa (CG-EM) ................................................... 106
3.6 Análise estrutural de biofilmes de kefir por microscopia óptica e por AFM .......... 107
3.7 Análise estrutural de biofilmes de kefir por microscopia eletrônica de varredura
(SEM) ................................................................................................................................... 109
4 DISCUSSÃO .................................................................................................................... 110
5 CONCLUSÃO .................................................................................................................. 116
REFERÊNCIAS ................................................................................................................. 117
CAPÍTULO 2: ESTUDO DO BIOFILME DE KEFIR ASSOCIADO AO EXTRATO
HIDROETANOLICO DE Euterpe Oleracea MART. (AÇAÍ). UM POSSÍVEL
PROBIÓTICO AMAZÔNICO ......................................................................................... 121
RESUMO ............................................................................................................................. 122
ABSTRACT ........................................................................................................................ 123
1 INTRODUÇÃO ............................................................................................................. 1243
2 MATERIAL E MÉTODOS .......................................................................................... 1276
2.1 OBTENÇÃO DE EHEEO. .......................................................................... ..........127
2.2 OBTENÇÃO DE BIOFILMES ASSOCIADOS AO EXTRATO
HIDROETANÓLICO DE EUTERPE OLERACEA MART. (BEHEEO). .......................... 127
2.3 DETERMINAÇÃO DA CONCENTRAÇÃO DE SUBSTRATO COM EHEEO
PARA FORMAR BIOFILMES. ................................................................................ 1287
2.4 DETERMINAÇÃO DA CONCENTRAÇÃO DE EHEHO OBJETIVANDO
FORMAÇÃO DE BIOFILME ..................................................................................... 128
2.5 EXTRAÇÃO E QUANTIFICAÇÃO DE ANTOCIANINAS. .............................. 128
2.6 PERFIL DA LIBERAÇÃO E DISSOLUÇÃO DE ANTOCIANINAS DE BEHEEO
..................................................................................................................................... 129
2.7 DOSAGEM DE MACRO E MICROELEMENTOS DE BEHEEO. ................. 13029
2.8 ANÁLISE ESTRUTURAL BEHEEO POR MICROSCOPIA DE FORÇA
ATÔMICA (AFM). ..................................................................................................... 130
2.9 ANÁLISE DE BIOFILMES DE KEFIR ATRAVÉS DE MICROSCÓPIO
ELETRÔNICA DE VARREDURA (MEV) ................................................................ 131
2.10 ANÁLISE ESTATÍSTICA. ................................................................................. 131
3 RESULTADOS ................................................................................................................ 132
3.1 OBTENÇÃO DE BEHEEO. .................................................................................. 132
3.2 DETERMINAÇÃO DA CONCENTRAÇÃO DE SUBSTRATO COM EHEEO
PARA FORMAR BIOFILMES. .................................................................................. 133

19
3.3 DETERMINAÇÃO DA CONCENTRAÇÃO DE EHEEO PARA A FORMAÇÃO
DE BIOFILME. ........................................................................................................... 134
3.4. PERFIL DA LIBERAÇÃO E DISSOLUÇÃO DE ANTOCIANINAS DE BEHEEO
..................................................................................................................................... 135
3.5 EXTRAÇÃO E DOSAGEM DE ANTOCIANINA DO BEHEEO ........................ 135
3.6. DOSAGEM DE MACRO E MICROELEMENTOS DE BEHEEO. .................... 136
3.7 COMPOSTOS VOLÁTEIS DO EHEEO ............................................................ 1365
3.8 ANÁLISE ESTRUTURAL BEHEEO POR MICROSCOPIA DE FORÇA
ATÔMICA. ................................................................................................................. 137
3.9 ANÁLISE ESTRUTURAL BEHEEO POR MICROSCOPIA ELETRÔNICA DE
VARREDURA (MEV) ................................................................................................ 139
3.10. DETERMINAÇÃO DA TEMPERATURA NA FORMAÇÃO DE BEHEEO. .. 139
4. DISCUSSÃO ................................................................................................................... 141
5 CONCLUSÃO .................................................................................................................. 145
REFERÊNCIAS ............................................................................................................... 1465
CAPÍTULO 3: ESTUDO DO BIOFILME DE KEFIR ASSOCIADOAO GÉRMEN DE
SOJA (GLYCINE MAX L. MERRIL) ................................................................................ 151
RESUMO ............................................................................................................................. 152
ABSTRACT ........................................................................................................................ 153
1 INTRODUÇÃO ............................................................................................................... 154
2 MATERIAL E MÉTODOS .......................................................................................... 1565
2.1 DETERMINAÇÃO DA CONCENTRAÇÃO DE GÉRMEN DE SOJA
NECESSÁRIA PARA A FORMAÇÃO DE BIOFILMES COM KEFIR E AM ....... 1565
2.2 DETERMINAÇÃO DA CONCENTRAÇÃO DE GRÃOS DE KEFIR
NECESSÁRIA PARA A FORMAÇÃO DE BIOFILMES COM GÉRMEN DE SOJA
................................................................................................................................... 1565
2.3 DETERMINAÇÃO DA CONCENTRAÇÃO DE ISOFLAVONAS NO MEIO DE
CULTURA E NOS BIOFILMES ................................................................................ 157
2.4 DETERMINAÇÃO DE PERÍODO INCORPORAÇÃO DE ISOFLAVONAS 1576
2.5 DOSAGEM DE MACRO E MICROELEMENTOS DE BIOFILMES DE KEFIR
COM GÉRMEN DE SOJA .......................................................................................... 158
2.6 ANÁLISE ESTRUTURAL DE BIOFILMES DE KEFIR COM GÉRMEN DE SOJA
ATRAVÉS DE MICROSCOPIA DE FORÇA ATÔMICA (AFM) ............................ 157
2.7 ANÁLISE DE BIOFILMES DE KEFIR COM GÉRMEN DE SOJA ATRAVÉS DE
MICROSCÓPIO ELETRÔNICA DE VARREDURA (MEV) .................................... 159
2.8 ANÁLISE ESTATÍSTICA. ................................................................................... 159
3 RESULTADOS ................................................................................................................ 160
3.1 DETERMINAÇÃO DA CONCENTRAÇÃO DE GÉRMEN DE SOJA
NECESSÁRIA PARA A FORMAÇÃO DE BIOFILMES COM KEFIR E GÉRMEN DE
SOJA ......................................................................................................................... 1609
3.2 DETERMINAÇÃO DA CONCENTRAÇÃO DE GRÃOS DE KEFIR
NECESSÁRIA PARA A FORMAÇÃO DE BIOFILMES COM GÉRMEN DE SOJA162
3.3 DETERMINAÇÃO DE PERÍODO NECESSÁRIO PARA INCORPORAÇÃO DE
ISOFLAVONAS ......................................................................................................... 163
3.4 DOSAGEM DE MACRO E MICROELEMENTOS DE BIOFILME COM GÉRMEN
DE SOJA ..................................................................................................................... 164

20
3.7. ANÁLISE ESTRUTURAL BIOFILMES DE KEFIR E GÉRMEN DE SOJA POR
MICROSCOPIA DE FORÇA ATÔMICA E MICROSCOPIA ELETRÔNICA DE
VARREDURA (MEV) .............................................................................................. 1654
4 DISCUSSÃO .................................................................................................................. 1687
5. CONCLUSÃO.................................................................................................................171
CONSIDERAÇÕES FINAIS.............................................................................................172
REFERÊNCIAS ........................................................................................................ .........175

21
1 INTRODUÇÃO GERAL
Através da História da Microbiologia os microrganismos têm sido descritos como seres
unicelulares. Analisando-se seu metabolismo a partir deste prisma têm sido descritos com base
em suas características em um meio de cultura nutricionalmente rico. Contudo na natureza, em
geral, não se encontram meios com disponibilidade de todos nutrientes, por isso estes seres
desenvolveram estratégias que lhes permitem comunicar-se entre si e agir como um ser
multicelular, o que se dá através da formação de biofilmes, os quais cumprem diversas funções
que lhes possibilitam a sobrevivência nos mais diversos habitats da Biosfera (FLEMMING;
WINGENDER, 2001a; SUTHERLAND, 2001; VAN HULLEBUSCH, 2003; VU et al., 2009).
Sabe-se que os microrganismos existem predominantemente em biofilmes e através
desta estratégia adquirem alta tolerância ao estresse tanto biológico como químico e físico
(GORBUSHINA; BROUGHTON, 2009). Em torno de 70 % da energia destes seres é despendida
neste processo (POLI et al., 2010), o que apenas evidencia o quanto isto é vital para os mesmos
(DAVEY; O'TOOLE, 2000).
Os biofilmes exercem várias funções desde a sustentação como a proteção contra
estresse ambiental. A formação de biofilme protege a bactéria contra bacteriófagos, amebas,
contra diversos biocidas e antibióticos (COSTERTON et al., 1999). Essa proteção é tanto física
pois retém e impede o contato com antibióticos como química criando uma barreira de proteção
com íons por exemplo (GRUMBEIN et al., 2014) que veta a entrada de compostos lesivos.
Neste estudo aborda-se o uso do complexo microbiano do kefir aferindo sua capacidade
de formar biofilmes ou de se agregarem a biofilmes.
O kefir constitui uma bebida obtida a partir da fermentação do leite ou de sucos de frutas
ou mesmo da água com açúcar. Obtido na temperatura ambiente é metabolizado por colônias de
lactobacilos, leveduras e fungos filamentosos que formam os “grãos de kefir”
Tradicionalmente costumamos pensar na célula como a unidade básica de toda a vida, os
grãos de kefir, no entanto se comportam como uma exceção na qual vários seres se fundem numa
relação mutualística formando um ser simbionte onde todos usufruem do mesmo substrato em
sucessão.
Quando se trata sobre biofilmes bacterianos são abordados quase exclusivamente os que
são formados por bactérias patogênicas, causando ferimentos crônicos, ou biofilmes que se

22
formam sobre instrumentos médicos (VU et al., 2009; VIDIGAL; SVIDZINSKI, 2009). Embora
haja uma abordagem sobre a utilidade dos mesmos em biorremediação (HONG et al., 2012), a
ênfase predomina sobre seus efeitos deletérios à saúde bem como sobre como impedir sua
formação e encontrar mecanismos que o limitem (GHOSH et al., 2015). Com efeito, em sua
forma séssil as bactérias se tornam até mil vezes mais resistentes a antibióticos. Por isso uma das
alternativas para o problema da resistência dos microrganismos a antibióticos tem sido a
compreensão e obstrução do mecanismo de formação de biofilmes (FURIGA et al., 2015;
GHOSH et al., 2015).
O uso de probíóticos como o kefir para induzir a formação de biofilmes tem perspectivas
para aplicações terapêuticas, pois estes biofilmes têm grande probabilidade de serem benéficos à
saúde. Probióticos repelem microrganismos patológicos ao competir por sitio e nutrientes
(VIDYALAXME et al., 2014) e impedem a formação de biofilmes por bactérias patogênicas e
anaeróbias (FERNÁNDEZ et al., 2015).
Uma das características dos biofilmes é sua capacidade de agregarem componentes do
meio que podem ser ácidos nucleicos, proteína e até mesmo sangue, portanto tendo em vista esta
capacidade ao se obter biofilmes de kefir conjecturou-se a possibilidade de esses biofilmes
incorporarem composto bioativos encontrado em plantas, como por exemplo as antocianinas do
açaí (Euterpe oleracea Martius) ou as isoflavonas do gérmen de soja (Glycine max (L) Merril).
O açaí (Euterpe oleracea Martius) tem despertado a atenção do mundo todo devido ser
uma planta da Amazônia que pode inclusive contribuir para a preservação da mesma
(MALCHER, 2011) devido ao interesse de sua exploração com sustentabilidade, exploração essa
que é decorrente do seu alto conteúdo de polifenóis dentre os quais se destacam as antocianinas
(PEIXOTO et al, (2016). Foi confirmado que as antocianinas podem inibir e prevenir diabetes e
câncer, além de combater o estresse oxidativo (SANTOS et al., 2008).
A cada ano um número maior de pessoas estão procurando produtos que contenham não
apenas sabor agradável ou que sejam de origem vegetal, mas também que possam contribuir para
melhoria de saúde e essa conscientização pode levar a surgimentos de produtos os quais dantes
não teriam aceitação. Essa perspectiva não se restringe apenas às antocianinas, mas também a
outros compostos bioativos como as isoflavonas encontradas em várias espécies vegetais,
notadamente em soja (Glycine max (L) Merril). A soja é, dentre as espécies vegetais estudadas a
que comporta maiores quantidades de isoflavonas (CARRÃO PANIZZIS et al., 2009).

23
O uso da soja pode combater e prevenir os efeitos da menopausa evitando a reposição
hormonal (IMAI, 2015). Os grãos de soja são apontados como os principais reservatórios de
isoflavonas (CARRÃO-PANIZZI et al., 2009). Além de sua atividade como fitoestrógeno, as
isoflavonas têm sido confirmadas como coadjuvantes no combate ao estresse oxidativo evitando
doenças cardíacas, câncer e outras patologias (ROSSI et al., 2004).
O efeito do uso da soja como fonte de fitoestrógenos tem sido questionado (SILVA et
al., 2009). Provavelmentes isto se deve ao fato de que para as isoflavonas serem absorvidas pelo
trato intestinal é mister que o mesmo esteja em homeostase (OLIVEIRA; BASTOS, 2011), caso
contrário sua ingesta se torna inócua. A situação de disbiose impede a absorção eficaz de
compostos bioativos. O uso de isoflavonas com probióticos, como o kefir, tem perspectiva de
causar o restabelecimento da homeostase e a consequente absorção de compostos benéficos ao
hospedeiro (YANG, 2015; COPPA et al., 2006).

24
2 REVISÃO BIBLIOGRÁFICA
2.1. KEFIR
O kefir é uma bebida feita a partir da fermentação do leite vegetal ou animal, em
temperatura ambiente, entre 10°C e 25°C, por colônias de Lactobacilos e leveduras, conhecidas
como “grãos de kefir”.
A palavra “kefir” deriva do turco keif, que pode ser traduzida como “bom sentimento”
ou “sentir-se bem” (OTLES; CAGINDI, 2003). Originário do Cáucaso foi introduzido no resto
do mundo, apenas, no início do século XX. Portanto ele é obtido através da incubação do
substrato com os grãos de kefir que são gelatinosos com massa irregular branco ou levemente
amarelos de tamanho variável oscilando entre vários milímetros a 3 cm de diâmetro. Sua forma é
similar à pipoca ou mesmo à couve em flor (LA RIVIERE et al., 1967; MARSHALL, 1993).
O kefir pode ser feito com água ou com todo o tipo de leite de origem animal ou vegetal
como, por exemplo, leite de coco, de soja, de arroz, de aveia, etc. A vantagem do kefir em água,
quando comparado ao de leite, é que pode tomar-se em grande quantidadee passa mais
rapidamente para a corrente sanguínea, se converte em uma bebida de consistência cremosa,
espessa e uniforme com um leve sabor ácido e um aroma moderado de levedura fresca (como a
cerveja). O kefir pode ser bebido puro ou, se não apreciar a acidez, pode-se adicionar um pouco
de adoçante ou mesmo misturar nas sopas, nos sumos de fruta ou batidos. Tem, igualmente, uma
efervescência natural, de gosto “carbonatado” que lhe confere sabor único e o seu odor agradável
muito característico (ANSELMO et al., 2010).
Os grãos de kefir são constituídos por leveduras homofermentativas como
Kluyveromyces marxianus; e leveduras heterofermentativas como Saccharomyces omnisporus,
Saccharomyces cerevisae e Saccharomyces exiguus, Lactobacillus casei, Bifidobaterium sp e
Streptococcus salivarius subsp thermophilus, entre outros (POGAČIĆ et al., 2013). Interessante
ressaltar que o kefir cultivado com leite de soja contém valores mais altos de bactérias ácido-
lácticas do que o kefir obtido a partir do leite de origem animal.
Observou-se que os Lactococcus e leveduras predominam na região mais externa dos
grãos de kefir, enquanto que os Lactobacillus se concentram em seu interior onde também
aparecem mais leveduras (WANG et al., 2012). Não se compreende ainda exatamente o

25
mecanismo de formação dos grãos de kefir. Wang et al., (2012) sugerem que inicialmente o
Lactobacillus kefiranofaciens e Saccharomyces turicensis começam a se autoagregarem e a
coagregarem formando pequenos grânulos. Essa agregação é facilitada pela queda do pH.
Microrganismos produtores de biofilme (Lactobacillus kefiri, Kluyveromyces marxianus HY1 e
Pichia fermentans HY) então aderem a superfície destes pequenos grânulos devido às
propriedades da superfície de suas células e de sua forte capacidade de agregação. Origina-se
assim um biofilme ainda bem fino. Após isto as leveduras e os Lactobacillus continuam se
agregando e se fundindo com o biofilme dos grânulos e, em consequência, torna-se uma
microcolônia tridimensional. Ocorre um aumento de densidade ainda mais acentuado pelo
acréscimo de células e componentes do meio de cultura formando assim o grão de kefir (WANG
et al., 2012, PRADO et al., 2015). É importante ressaltar que ocorre concomitantemente a síntese
de um exopolissacarídeo, o kefirano, secretado notadamente por Lactobacillus kefiranofaciens, o
qual contribui para a formação da matriz exopolissacarídea, principal componente dos biofilmes
de kefir e onde as células se fixam.
2.1.1. Efeito probiótico do kefir
O kefir é efetivo contra uma variedade de doenças (HOSONO et al., 1990). Vários
estudos têm investigado sua atividade antitumoral (FURUKAWA et al., 1991; ZAMBERI et al.,
2016) e atividade antimicrobiana in vitro contra uma extensa variedade de bactérias gram-
positiva e gram-negativas e contra alguns fungos (CEVIKBAS et al., 1994; ZACCONI et al.,
1995; BOURRIE et al., 2016).
Matsuu et al. (2003), comprovaram que o kefir pode proteger células troncos da
apoptose após o tratamento por radiação. Além de seu efeito desintoxicante, atua como
regenerador da microbiota intestinal benéfica e estimula as defesas naturais (SPINLER et al.,
2016) possibilitando o tratamento de pessoas debilitadas.
Contitui um sistema biológico que não tem sido plenamente compreendido,
provavelmente devido a sua complexidade e, embora haja muitos estudos descrevendo os grãos
de diferentes origens, eles apenas parcialmente analisam o sistema (BESHKOVA et al. 2003).

26
2.1.2 Compostos voláteis do meio de cultura
O sabor único do kefir resulta da presença de vários flavorizantes naturais que são
produzidos durante sua fermentação (BESHKOVA et al., 2003). O kefir produzido de culturas
puras não teve a mesma qualidade e aceitação.
Duitschaever et al. (1987) comprovaram que apenas 40% das pessoas apreciam o sabor
natural do kefir ao provarem a primeira vez. Adicionando flavorizante ou modificando o processo
fermentativo através da inserção de leveduras com lactobacilos houve um aumento da aceitação
do kefir quando comparado ao kefir tradicional (DUITSCHAEVER et al., 1987; MUIR et al.,
1999).
A importância do aroma no sabor de frutas e bebidas destiladas é advinda do fato que a
sensação de sabor em grande parte é proveniente do olfato. O aroma de um alimento pode ser
explicado pela ocorrência de compostos químicos cuja principal característica é a volatilidade, a
qual permite que tais compostos sejam percebidos pelos receptores nasais, tanto durante a
degustação do alimento (detecção retronasal), como pelo odor exalado a distância (NÓBREGA,
2003). Inclusive a qualidade e a quantidade do aroma retronasal liberado durante o consumo de
alimento afeta o tempo de exposição dos receptores nasais para estímulo do aroma e isto pode
influenciar nas características hedônicas e nutricionais e consequentemene o consumo desses
alimentos (MESUROLLE et al., 2013). Portanto, cumpre ressaltar que não apenas o tipo de
composto volátil altera a qualidade e o sabor das substâncias, mas também as características de
odor dos compostos voláteis dependem de suas concentrações. Compostos com grande
quantidade podem se tornar intragáveis enquanto que em pequena quantidade são apetecíveis,
assim concentrações muito elevadas podem modificar características de odor consideradas
"agradáveis" para "extremamente desagradáveis", condição essa igualmente observada em alguns
compostos voláteis sulfurados (NÓBREGA, 2003). O diacetil ou 2,3-Butanodiona, por exemplo,
é um composto aromático cujo aroma é desagradável no leite fermentado (KANEKO et al.,
2014). Em seu trabalho Trelea et al. (2008) através de auxílio de modelagem computadorizada
obteve uma formulação permitindo o cálculo da intensidade do aroma retronasal como uma
função da transferência das propriedades voláteis dos compostos aromáticos presentes em
matrizes alimentares e as características anatomofisiológicas dos consumidores.
Em se tratando de produtos fermentados o seu sabor e aroma provêm de sua produção de

27
ácidos voláteis e não voláteis e de compostos carbonilas (TAMIME; ROBINSON, 1999;
MARSHAL et al., 1984; FERNANDEZ-GARCIA; MCGREGOR, 1994).
Os principais compostos que influenciam no sabor e aroma do kefir são o ácido
indolacético, a acetona, acetaldeído, etanol, propanona e 2-butanona (IMHOF et al., 1995;
DEGORCE-DUMAS et al., 1986; OTT et al., 1997; KNEIFEL et al., 1992). Foi observado que
na armazenagem o acetaldeído aumenta e a acetona decresce. O acetaldeído e a acetona têm
recebido particular atenção com relação ao seu papel no sabor durante a fermentação do kefir. Há
um aumento em sua concentração durante a manufatura do kefir.
A adição de frutose aumenta a produção de vários flavorizantes voláteis sem aumentar o
tempo necessário para sua fermentação.
Além de se provar através do paladar, milhares de compostos voláteis podem ser
detectados e diferenciados, em menor ou maior escala, pelo olfato. Isso depende de diversos
fatores, dentre os quais, a estrutura química do composto volátil e a interação deste com a matriz
alimentícia. Tais fatores, por sua vez, são responsáveis por características importantes dos
compostos, a exemplo o valor do limiar de odor (limiar de detecção ou "threshold"), que pode ser
definido como a concentração mínima pela qual um composto pode ser detectado pelo sentido do
olfato (BELITZ; GROSCH, 1999).
Muitos compostos voláteis são detectados através de cromatografia gasosa e dentre
estes, uma parcela significativa possui grande impacto na qualidade do aroma das frutas e bebidas
alcoólicas, tais como ésteres, álcoois e compostos sulfurados. Os ésteres são dos mais
importantes, pois causam grande impacto tanto no sabor de frutas como de bebidas fermentadas.
Também têm sua importância os aldeídos, álcoois superiores, entre outros (BESHKOVA et al.,
2003)
Os ésteres geralmente possuem valores de limiar de odor relativamente baixos,
assumindo características de aroma mais marcantes.
A principal origem dos ésteres, nas bebidas alcoólicas, está no metabolismo secundário
intracelular das leveduras durante a fermentação alcoólica. Dentre os ésteres existentes, os tipos
etílicos de ácidos graxos e acetatos são considerados importantes em bebidas alcoólicas, devido
às suas concentrações serem relativamente elevadas, às suas características de aroma serem
agradáveis e aos seus valores de limiar de odor serem relativamente baixos (BELITZ; GROSCH,
1999).

28
2.1.3 Processos fermentativos do kefir e produção de etanol
O kefir é um probiótico constituído por uma biomassa complexa com bactérias ácido
lácticas e ácido acéticas que vivem em simbiose em prol do hospedeiro originando um produto
fermentado que realiza a síntese e melhor absorção de proteínas, vitaminas e minerais
(FARNWORTH, 2005). As bactérias do ácido lático se concentram na superfície dos grãos
enquanto que as leveduras predominam no centro (LIN et al., 1999). Há uma relação simbiótica
entre os microrganismos presentes nos grãos de kefir onde as bactérias e leveduras vivem e
compartilham seus bioprodutos como fonte de energia e fatores de crescimento e é justamente
essa associação de microrganismos a responsável pela fermentação lática e alcoólica
(WITTHUHN et al., 2005; WANG et al., 2012; HAMET et al., 2013).
Há vários parâmetros que influenciam a produção de etanol com o uso do complexo
microbiano do kefir tais como pH do meio de cultura, concentração do inóculo e do substrato, a
composição deste substrato, lavagem dos grãos e o tipo do leite. Para Sanchez et al. (2004) a
temperatura é uma das variáveis mais importantes. A produção de etanol pelo kefir geralmente se
dá em torno de 2% (ZAJŠEK; GORŠEK, 2010; SALOM et al., 2010). No entanto a produção de
etanol pode ser induzida através de controle na concentração da fonte de carbono (sacarose) e da
temperatura podendo-se obter etanol até a 14,5 °GL (DORNELLES et al., 2006). Observou-se
que a produção do etanol inicia-se em torno de 5 horas após a inoculação dos grãos de kefir
(GUZEL-SEYDIM et al., 2005).
Para Dornelles et al. (2006), o aumento na concentração dos grãos de kefir não alterou a
concentração do etanol, mas, alterações na concentração de sacarose surtiram este efeito,
enquanto Guzel-Seydim et al. (2005) concluiram que na armazenagem do kefir o etanol não
volatiza, ocorrendo pelo contrário um ligeiro aumento em sua concentração.
2.1.4 A associação simbiótica do kefir
O kefir exerce sua ação probiótica no nosso organismo provavelmente através da
competição por sítio de adesão. Isto foi proposto por Santos et al. (2003) trabalhando com duas
cepas de microrganismos isolados do kefir, Lactobacillus acidophilus CYC 10051 e
Lactobacillus kefiranofaciens (CYC 10058) verificaram que tinham capacidade de adesão às

29
células do cólon humano, bem como de inibir a aderência da Salmonella typhimurium, além de
atividade antimicrobiana contra várias estirpes patogênicas.
A estabilidade da balança populacional nos grãos de kefir tem sido atribuída à interação
simbiótica entre as leveduras e as bactérias ácido-lácticas (ADACHI et al., 1990). Enquanto as
leveduras provêm as vitaminas e aminoácido e fatores de crescimento para as bactérias o produto
final da fermentação das bactérias são o substrato das leveduras como exemplo mencionado da
lactose e do ácido láctico que pode ser metabolizado por Saccharomyces cerevisae. O ácido lático
é um inibidor do crescimento do L. kefiranofaciens e ao ser consumido pela levedura proporciona
condições para o crescimento da bactéria (CHEIRSILP et al., 2003), isto parece explicar por que
Mitsue et al. (1999) reportaram que algumas leveduras habilitam a produção de kefirano por L.
kefiranofaciens.
Pode-se afirmar com relação ao kefir que se trata de um sistema biológico que não tem
sido plenamente compreendido ainda que haja muitos estudos descrevendo diferentes fontes de
grãos estes são estudos que analisam os sistemas apenas parcialmente falhando em analisá-lo
como um todo (CHEIRSILP et al., 2003).
O alto teor de açúcar e a baixa concentração de aminoácidos encontrados no kefir de
água deveria resultar em um habitat em carência, no entanto o que se obtém é uma bebida
fermentada com uma comunidade microbiana bastante diversificada, mas em equilíbrio e
estabilidade (LAUREYS; DE VUYST, 2014). Estabelece-se, portanto um nicho ecológico onde
apenas organismos muito bem adaptados a formar consórcios com outros conseguem crescer e se
prover nutrientes essenciais (LAUREYS; DE VUYST, 2014). No kefir metabolizado em água
ocorre sinergia entre leveduras e bactéria ácido-lácticas notadamente com lactobacilos (STADIE
et al., 2013).
O contato e a adesão na interação fungo-bactéria é fundamental para o desenvolvimento
de biofilmes polimicrobianos (GRANILLO et al., 2015). O cultivo de ambos em consórcio em
kefir mostrou crescimento na produção de células de ambos evidenciando o mutualismo que
ocorre entre estes seres. Os Lactobacillus provêm a acidificação do meio, condição
imprescindível para o desenvolvimento das leveduras que por sua vez dispõem nutrientes
essenciais para as bactérias. No caso específico do Lactobacillus hordei as leveduras contribuem
nesta interação trófica ao prover aminoácidos e vitamina B6 já o Lactobacillus nagelii é
beneficiado pelos aminoácidos produzidos pela levedura. Quando cultivado juntos o

30
Zygotorulaspora florentina é induzido a liberar arginina que é essencial para o Lactobacillus
nagelii (STADIE et al., 2013).
Ainda sobre a ação do kefir através da competição por sítio de adesão e por nutrientes,
sabe-se que este é o modo mediante o qual o kefir compete com os agentes patogênicos e assim
impede o seu estabelecimento e desenvolvimento. O concurso pelo sítio com a consequente
exclusão dos seus concorrentes pode explicar por que os probióticos devem ter administração
contínua com doses significativas para ser eficaz (SAAD, 2006). Além disso, o complexo
microbiano do kefir produz ácido lático, antibióticos e antibactericidas que inibem o
desenvolvimento de microrganismos patogênicos (LIU et al., 2002).
A ação antipatogênica do kefir é efetiva contra Salmonella, Helicobacter, Shigella,
Staphylococcus, Escherichia coli, Enterobacter aerogenes, Proteus vulgaris, Bacillus subtilis,
Micrococcus luteus, Listeria monocytogenes, Streptococcus pyrogenes, (LOPITZ et al., 2006),
Streptococcus faecalis KR6, Fusarium graminearu CZ1 (ISMAIEL et al., 2011), e contra
Candida albicans (RODRIGUES et al., 2005).
Também pode prevenir diarreia e enterocolite causada por Clostridium difficile (BOLLA
et al., 2013).
Mas seus efeitos benéficos se estendem para outros aspectos igualmente importantes. O
kefir tem se revelado eficaz em inibir a formação de esporos e a produção de aflatoxina B1.
Sintetizada e secretada por Aspergillus flavus, se trata de um composto tóxico formado tanto em
campo de cultivo como quando o amendoim é armazenado. Portanto o kefir aparece como uma
promissora alternativa para reforçar a segurança alimentar oferecendo proteção contra
intoxicação por aflatoxina (ISMAIEL et al., 2011).
Além disso, a sua ação probiótica provoca modulação na resposta imune do hospedeiro,
estimulando a resposta imunitária específica.
No organismo, os probióticos ativam os macrófagos os quais acionam a síntese e
excreção de citocinas as quais vão causar o aumento da atividade dos linfócitos T Killer, as
células assassinas naturais (FEDORAK; MADSEN 2004; RASTALL et al., 2005; SAAD, 2006).
Mendes et al. (2013) identificaram os passos mediante os quais as leveduras interagem
de maneira simbiótica com bactérias ácido láticas analisando apenas duas espécies de
microrganismos do kefir crescendo em co-cultivo: Lactobacillus delbrueckii subsp. Bulgaricus e
Saccharomyces cerevisae. Primeiro o Lactobacillus delbrueckii hidrolisa a lactose, procedimento

31
este que não pode ser efetivado pelo Saccharomyces cerevisae. A lactose é, portanto, hidrolisada
em galactose e glicose. Sequentemente a galactose é metabolizada pela levedura provendo assim
uma fonte de carbono, haja vista que a galactose não pode ser metabolizada pelo Lactobacillus
delbrueckii. O Lactobacillus cresce apenas na presença de aumento de concentração de CO2 e
neste cultivo anaeróbico a levedura através da fermentação alcoólica fornece o CO2. A levedura
provê também aminoácidos para a bactéria inclusive alanina. Concomitantemente a quelação do
ferro pelos Lactobacillos a partir do lactato se torna uma importante fonte para a levedura.
Rodrigues et al. (2005), analisaram a atividade antimicrobiana e cicatrizante do kefir e
da sua fração de carboidratos contra várias bactérias e contra Candida albicans. Feridas induzidas
em ratos Wistar foram inoculadas com patógenos dentre os quais Staphylococcus aureus. O
ferimento foi tratado com um gel de kefir a 70% e observou-se que tanto o kefir como sua Fração
e Carboidratos do Kefir (FCK) mostraram atividade contra todos os organismos testados sendo
mais efetiva a atividade contra Streptococcus pyogenes. Constatou-se um efeito protetor sobre a
pele do tecido habilitando a cura do ferimento em apenas sete dias comparado com o controle
positivo. Em um trabalho avaliativo das propriedades de uma fração isolada de carboidratos do
kefir em açúcar (FCK), Moreira et al. (2008), demostraram sua atividade anti-inflamatória
abrindo perspectivas na aplicação de agentes anti-inflamatórios oriundos de carboidratos
poliméricos.
2.1.5 O kefirano
Há três importantes componentes nos grãos de kefir: as leveduras, as bactérias ácido-
láticas e o kefirano (BESHKOVA et al., 2003). Estes três componentes conferem ao kefir a
qualidade de uma comunidade simbiótica que comporta propriedades peculiares. O kefirano é um
polissacarídeo sintetizado pelos grãos de kefir composto por partes iguais de galactose e glicose
(YEESANG et al., 2008) em unidades repetitivas de hexa e heptassacarídeos (FARNWORTH,
2005; GUZEL-SEYDIM et al., 2005). Ele é usado pelos microrganismos para emaranhar-se
dentro dos grãos de kefir (YEESANG et al., 2008). Um complexo e fino biofilme pode ser
observado no exterior dos grãos enquanto no interior o material fica comprimido. Este filme é
composto por uma microbiota predominantemente em forma de limão ou por um longo filamento
de células crescendo em associação com bactérias cocos e bacilos (LOPITZ-OTSOA et al., 2006)

32
Até pelo menos o início deste século pouco havia sido estudado sobre as melhores
condições para formação do kefirano no próprio kefir. As informações sobre alterações que
ocorrem ao transcorrer do processo fermentativo eram então escassas e se limitavam apenas ao
que se refere à formação de ácidos orgânicos e substâncias flavorizantes (GUZEL-SEYDIM et
al., 2000). O kefirano não está presente apenas nos grãos, mas também no meio de cultura, bem
como nos produtos fermentados (leite e soro) (RIMADA; ABRAHAM, 2003).
Sabe-se que apenas os grãos podem ser inoculados para produzir mais kefir (SIMOVA
et al., 2002), não se utiliza o meio de cultura do kefir pois, mesmo a microbiota do kefir estando
em equilíbrio a estrutura e a quantidade das espécies e de vários grupos de microrganismos
mudam significativamente ao longo das vias bioquímicas no meio de cultura de kefir (SIMOVA
et al., 2002). Isto se deve ao fato que o meio é alterado pela atuação de vários microrganismos.
No kefir as alterações da população ocorrem também em função de outros fatores como origem,
diferente metodologia ou substrato (PINTADO et al., 1996), contudo, há uma microbiota
permanente nos kefir. Fontain et al. (2003) afirmam que quanto maiores os grãos de kefir
utilizados mais baixo o nível de espécies de micróbios pode ser encontrado nos mesmos.
Após a retirada dos grãos este meio ainda há grãos invisíveis, indistinguíveis mesclados
ao meio e eles também produzem e são consituídos por kefirano. Estes grãos invisíveis do meio
contêm alta concentração de cocos e lactobacilos e é isto, justamente o que caracteriza o kefir
como um probiótico (BESHKOVA et al., 2003), pois são ingeridos quando a pessoa consome o
kefir.
O kefirano é produzido por lactobacilos em especial o Lactobacillus kefiranofaciens o
qual atua após 48 horas de cultivo. Antes disso predominam os lactococos e depois deles os
lactobacilos se tornam então a espécie dominante (FONTAIN et al., 2003).
Nos primeiros estágios da produção de kefir a combinação de microrganismos não se
formou ainda para uma produção de kefirano em concentração suficiente para reter os grãos
estritamente coesos (FONTAIN et al., 2003). Isto torna fácil que sejam removidos dos grãos e
disponibilizados para isolamento (FONTAIN et al., 2003) o que proporciona a condição ideal
para que seja efetivado o isolamento de bactérias que produzem o kefirano.
Sintetizado por Lactobacillus como Lactobacillus rhamnosus, L. kefirand e L.
kefiranofasciens, o kefirano tem sido utilizado pelas indústrias (DUBOC; MOLLET, 2001;
MICHELI et al., 1999) primariamente para produzir uma tradicional bebida carbonatada

33
levemente alcóolica em leite fermentado a qual melhora a viscosidade e a viscoelasticidade dos
géis de leite com propriedade que preveni e cura doenças (REHM, 2009).
As vias metabólicas que levam à produção de kefirano não são completamente
compreendidas, sabe-se que esse polissacarídeo é composto por unidades repetitivas de hexa e
heptassacarídeos constituídos principalmente de glicose e galactose, com ramificações
estruturadas (FARNWORTH, 2005; GUZEL-SEYDIM et al., 2005) as quais são representadas na
Figura 1.
Figura 1 - Estrutura do o kefirano, polissacarídeo encontrado nos grãos de kefir.
Fonte: Próprio autor
Desde o início do seu isolamento, tem sido relatado que o kefirano pode ser produzido
por uma variedade de bactérias isoladas dos grãos de kefir obtido de diferentes fontes de
carboidratos como açúcar mascavo, leite de origem animal e de origem vegetal, entre outras
(FARNWORTH, 2005). Sabe-se também que este exopolissacarídeo não está presente apenas nos
grãos, mas também nos produtos fermentados (leite e soro) (RIMADA; ABRAHAM, 2003).
Há relatos que o kefirano possui atividade antibacteriana e antitumoral, modula o
sistema imunológico do intestino e protegem as células epiteliais contra Bacillus cereus
(MAEDA et al., 2004; RODRIGUES et al., 2005; VINDEROLA et al., 2006; MEDRANO et al.,
2007).
Mais de 100 espécies de bactérias e leveduras fazem parte da complexa microbiota do
kefir (TAKIZAWA et al., 1998). Já em 1990 Adachi et al. presumiram que haveria uma interação
entre estes seres e classificou os microrganismos em 4 grupos: bactérias ácido lácticas

34
homofermentativas, bactérias ácido lácticas heterofermentativas, leveduras assimiladoras de
lactose e leveduras não assimiladoras de lactose. As bactérias ácido-lácticas homofermentativas
quebram a molécula de lactose em galactose e glicose proporcionando para as leveduras não
assimiladoras de lactose a galactose que pode ser metabolizado por elas.
As bactérias ácido-láticas homofermentativas produtoras de kefirano podem ser
estimuladas por CO2, ou por etanol ou talvez por algum metabólito desconhecido produzido pelo
metabolismo dos outros microrganismos presentes nos grãos de kefir. Tem havido uma busca
crescente em produzir kefirano e por isso há muito interesse em se compreender as complexas
interações que ocorrem no ecossistema do kefir (MITSUE et al., 1999). Contudo o mecanismo de
produção de kefirano em culturas mistas não foi totalmente esclarecido, pelo menos, até o
momento,
O consumo do CO2 por Saccharomyces cerevisae pode habilitar produção de kefirano. O
ácido lático inibe o L. kefiranofasciens tanto no que se refere ao crescimento como na produção
de kefirano e como Saccharomyces cerevisae consegue metabolizar o ácido lático conclui-se que
pode ser habilitado para incrementar a produção de kefirano. Muitos meios já foram utilizados
para reduzir o ácido láctico e assim acionar maior produção de kefirano, mas o uso do
Saccharomyces cerevisae que consome o ácido lático do meio provou ser prático e viável
(CHEIRSILP et al., 2003).
O gênero Lactobacillus é predominante no kefir (PINTADO et al., 1996; ANGULO et
al., 1993; WITTHUHN, 2004), isto evidencia a importância deste grupo na produção do kefir e
do kefirano. São bactérias deste gênero as responsáveis pelo kefirano e consequentemente pelos
grãos de kefir onde ocorre a incrustação das bactérias e leveduras (LA RIVIERE et al., 1967;
MARSHALL et al., 1984). Várias bactérias podem sintetizar kefirano e dentre estas a mais
conhecida é o Lactobacillus kefiranofaciens descoberto por Fujisawa et al. (1988) que o
identificaram e provaram ser o principal produtor de kefirano. O Lactobacillus kefiranofaciens é
inclusive o microrganismo predominante no kefir compondo 65% da sua microbiota (TOBA et
al.,1987)). Alguns pesquisadores buscaram a produção desse exopolissacarídeo isolando esta
espécie e buscando substratos alternativos com o objetivo tanto de potencializar sua produção
como encontrar alternativas mais econômicas, por exemplo o uso do sago (YEESANG et al.,
2008). No entanto, a produção de kefirano a partir do uso do complexo microbiano do kefir pode
ser mais prática pela facilidade de se obter o produto e pela ação simbiótica que torna o kefir

35
melhor produtor do que a bactéria isolada, pois a ação sinergética do kefir é potencializada com a
presença de todos os seus componentes (TOBA et al., 1987). Isto pôde ser comprovado por
Frengova et al. (2002), os quais avaliando a produção de exopolissacarídeos (o kefirano) por
bactérias ácido-lácticas isoladas de grãos de kefir selecionaram uma cepa de Lactobacillus
bulgaricus que sobressaiu dentre as 40 cepas de bactérias lácticas isoladas pela alta atividade
exopolissacarídica e verificaram que na associação dessa cepa com outras bactérias lácticas e
levedura (simulando uma cultura iniciadora de kefir) , a produção de kefirano foi de 1,7 vezes
maior que a do L. bulgaricus isolado. (FRENGOVA et. al. 2002; OTTOGALLI et al., 1973).
Além disso, no kefir há outras bactérias que também produzem kefirano como o L.
kefiranofaciens
La Riviere et al. (1967) descobriram bactérias heterofermentativas formadoras de
cápsulas as quais foram isoladas dos grãos de kefir. Para Toba et al. (1987) isto foi decorrente de
purificação incompleta, mas o L. kefiranofaciens pode produzir kefirano a partir de
monossacarídeos (CHEIRSILP et al., 2003) além de se encontrar no meio bactérias
heterofermentativas com esta capacidade. Em 1994 Takizawa et al. anunciaram duas novas
espécies, L. kefirgranum e L. parakefir, dentre os lactobacilos da flora do kefir. Takizawa et al.
(1998) confirmaram que o Lactobacillus kefiranofaciens é o principal produtor de kefirano, mas
encontraram como predominante na microflora o Lactobacillus kefirgranum que não produz o
kefirano. Observaram que ambos estão interligados nos grãos de kefir.
2.1.5.1 Kefirano: possíveis aplicações
O kefirano é primariamente utilizado para produzir uma tradicional bebida carbonada
levemente alcóolica em leite fermentado a qual melhora a viscosidade e a viscoelasticidade dos
géis de leite com propriedade que preveni e cura doenças (REHM, 2009).
O kefirano é também emulsificador e agente quelante e tem a propriedade de substituir
gorduras (CHEIRSILP et al., 2003). Tem atividade antitumoral (SHIOMI et al., 1982). Sua
atividade antibacteriana tem sido comprovada, bem como sua atuação como modulador do
Sistema Imunológico (MAEDA et al., 2004; RODRIGUES et al., 2005; VINDEROLA et al.,
2006; MEDRANO et al., 2007). Por estas razões há interesse que se descubra meios de produção
em massa de kefirano (YOKOI; WATANABE, 1992; MITSUE et al., 1998).

36
Há também interesse industrial com respeito à possível utilização do kefirano como
agente de viscosidade de produtos lácteos e também como aumentar a estabilidade destes
produtos (ABRAHAM; RIMADA, 2006). O kefirano tem sido utilizado na indústria alimentícia
como aditivo alimentar para produtos fermentados realçando as propriedades reológicas dos géis
de leites fermentados aumentando sua viscosidade aparente e a estabilidade desses produtos
durante seu tempo de armazenamento (WANG et. al., 2008).
2.1.5.2 Utilização do kefirano como biopolímero
Há um aumento da demanda por polímeros naturais, pois exercem aplicações tanto para
a medicina como industrial (WANG et al., 2010) e por isso polímeros produzidos por
microrganismos têm sido intensamente pesquisados.
Os microrganismos sintetizam dois tipos de polissacarídeos, um deles pode ser agregado
aos grãos e é chamado de capsular (CPS) e um exopolissacarídeo que pode formar um filme no
meio e é chamado de EPS (SUTHERLAND, 2001). Sabe-se que muitos microrganismos podem
produzir exopolissacarídeos.
Apesar de vários estudos reportando o uso de polissacarídeos de diferentes fontes para
preparar filme e embalagens, relativamente pouca atenção se dava a exopolissacarídeos
produzidos por microrganismos devido a sua baixa produção (PAUL et al., 1986). O kefir, no
entanto produz quantidade expressiva de kefirano que pode fornecer material para biofilme
formando géis e soluções de baixa viscosidade (GHAMSELOU et al., 2011). Esse polissacarídeo
sintetizado pelo kefir, tem várias importantes vantagens sobre outros polissacarídeos, pois além
de proteção contra estresse mecânico tem capacidade de servir como controlador da umidade, de
oxigênio, de dióxido de carbono e de umidade (GONTARD et al, 1993), e da volatilização de
compostos flavorizantes. Industrialmente o kefirano pode ser utilizado também como agente
quelante e texturizante. Uma alta produção destes polissacarídeos tem sido alcançada facilmente
isolada dos grãos, portanto os exopolissacarídeos do kefir são uma alternativa para embalagem de
alimentos.
O aumento na demanda por polímeros naturais para várias aplicações tem conduzido ao
desenvolvimento da produção de exopolissacarídeos produzidos por microrganismos com várias
aplicações tanto industriais como medicinais (GHASEMLOU et al., 2011).

37
Além das supracitadas o kefirano tem outras importantes vantagens com relação a outros
polissacarídeos, tais como propriedades antibacterianas, antifúngicas e antitumorais além de ser
antimutagênico (MAEDA et al., 2004; MUROFUSHI et al., 1983). Estas macromoléculas
apresentam reologia pseudoplástica e alta viscosidade, características estas que oferecem a
possibilidade de serem utilizadas como emulsificantes, viscosificante, floculadoras e agentes
quelantes (WANG et al., 2010; KODALI et al., 2009, DE VUYST et al., 2001) e, em alguns
casos podem também ter forte resistência a alteração de pH e temperatura (KIM et al., 2000;
KNOSHAUG et al., 2011).
Há um imenso potencial na utilização de biopolímeros como o kefirano em aplicações
farmacêuticas e medicinais. Estas moléculas bioativas contribuem em diferentes atividades
fisiológicas humanas como antitumorais, anti-inflamatórias, agentes antivirais. Elas induzem a
síntese de interferon e a atividade citocina, inibem formação de coágulos pela aglutinação de
plaquetas (CALAZANS et al., 1997; ARENA et al., 2006).
Estes biofilmes agem como uma barreira controlando umidade oxigênio, dióxido de
carbono, lipídeos e componentes flavorizantes com a vantagem de serem biodegradáveis
(GONTARD et al., 1993). Outro fato que deve ser levado em consideração é poder satisfazer os
clientes que estão cada vez mais exigentes por produtos de alta qualidade e que não firam o meio
ambiente, pois embalagens produzidas com biofilmes além de biodegradáveis proporcionam o
aumento do prazo de validade dos produtos (GHASEMLOUA et al., 2011).
2.1.5.3 Fatores para a produção de kefirano
Cheirsilp et al. (2003) confirmaram que ao contrário do que se afirmava antes o estresse,
o CO2 e o O2 não estimulam a produção de kefirano. Ainda Cheirsilp et al. (2003) afirmam que é
fato bem estabelecido que o ácido lático inibe a produção de kefirano por inibir o crescimento de
seu principal produtor o L. kefiranofaciens e assim sugerem que a remoção do ácido lático agiliza
a produção de kefirano. Vários métodos de remoção de ácido lático do meio de cultura para
habilitar a produção de kefirano foram desenvolvidos (HAYAKAWA et al.,1990; OHARA et al.,
1993).
No entanto sabe-se que o L. kefiranofaciens consegue metabolizar a sacarose e induzir a
produção de kefirano mesmo na ausência de lactose (WITTHUHN, 2004) e consequentemente

38
não produzindo ácido lático em grande quantidade que é o produto final da metabolização da
lactose.
2.2 BIOFILMES MICROBIANOS
Um biofilme é um agregado de células microbianas (bactérias, fungos, algas e
protozoários), irreversivelmente associadas com uma superfície (seja de materiais sólidos ou
interfaces), ligada em uma matriz de polissacarídeo primário, formando comunidades biológicas
com um elevado grau de organização, onde as bactérias formam comunidades estruturadas,
coordenadas e funcionais. Portanto os biofilmes são superfícies que comportam alto grau de
organização estrutural associados a colônias de bactérias mantidas coesas através de polímero
secretado por suas células o qual é denominado de substância polimérica extracelular (SPE)
(WATNICK; KOLTER, 2000; DONLAN, 2002; ZHANG et al., 2015). Esta matriz consiste em
uma junção de compostos poliméricos de polissacarídeos primários os quais formam a Substância
Polímera Extracelular (SPE). São grupos de células em uma interface cimentada juntas através de
polissacarídeos, proteínas DNA e lipídeos (HONG et al., 2012). Os biofilmes conferem
resistência ao stress, proteção contra impactos químicos e mecânicos e disponibilizam os
nutrientes, por isso são fundamentais para sobrevivência dos microrganismos. Muitas bactérias
são altamente habilitadas para viver em biofilmes os quais mantêm as células agregadas através
da sua matriz polimérica extracelular. Com os biofilmes elas obtêm a proteção contra o estresse
além de habilitar sua logística de nutrientes (NADELL et al., 2015).
Os microrganismos vivem insertos nestes biopolímeros e apenas com raras exceções
vivem isoladamente. Estas células nos biofilmes são cimentadas no local pelo polímero secretado
pela matriz polimérica consistindo de polissacarídeos, proteínas e formam uma barreira protetora
que confere resistência a defesas do hospedeiro durante a infecção e tolerância a vários agentes
antimicrobianos tais como desinfetantes e antibióticos (HONG et al., 2012).
O biofilme é essencial para a sobrevivência dos microrganismos nele agregado
(FLEMMING; WINGENDER, 2001a; SUTHERLAND, 2001; VAN HULLEBUSCH, 2003; VU
et al., 2009).
Materiais de origem não celular como cristais de minerais, partículas de corrosão, argila,
silte podem também ser encontrados na matriz do biofilme. Até partículas do sangue a depender

39
do local onde o biofilme se desenvolveu podem ser encontrados na matriz do biofilme.
Os organismos associados em biofilme diferem dos planctônicos (de vida livre em
suspensão) no que se refere aos genes que são transcritos (DONLAN, 2002). Biofilmes podem se
formar numa ampla variedade de superfícies incluindo tecidos vivos, instrumentos médicos, em
água potável ou industrial ou em sistemas aquáticos naturais (FLEMING; WINGENDER,
2001b). O biofilme em um sistema de água é altamente complexo contendo produtos de corrosão,
argila, diatomáceas e bactérias filamentosas, já o biofilme sobre aparelhos médicos se apresenta
composto de uma simples película cobrindo o aparelho médico juntamente com a matriz da
Substância Polimérica Extracelular (SPE) (DONLAN, 2002). Os biofilmes têm sido relacionados
a muitas doenças infecciosas e inflamatórias bem como à biocorrosão e a bioadesão em diversas
áreas.
Os biofilmes podem também ser utilizados em aplicações benéficas tais como
biorremediação além de reterem potencial para serem utilizados em transformações químicas em
biorrefinarias (HONG et al., 2012). Por estas aplicações as culturas mixadas (consórcios)
comparadas com monoculturas, têm a vantagem de serem poderosas para cumprirem mais
complexas transformações (por exemplo, aquelas que requerem múltiplos passos), além de que
são mais resistentes ao stress ambiental. Por esta razão consórcios têm sido considerados como
uma opção supraexcelente em biologia sintética (BRENNER et al., 2008). Contudo, há
dificuldades no sentido de se controlar a formação de biofilme em consórcio (HONG et al.,
2012). Alterações nas condições ambientais (nível de nutrientes, por exemplo, ou depleção de
oxigênio) podem ser o gatilho de remoção de biofilmes.
A substância polimérica extracelular (SPS) que compõe os biofilmes constituem o
espaço intercelular dos agregados microbianos e formam as estruturas e a arquitetura da matriz de
biofilme (VU et al., 2009).
A formação do biofilme protege a comunidade do microrganismo do estresse do
ambiente quer seja este natural como industrial (FLEMMING; WINGENDER, 2001b; AHIMOU
et al., 2007.
A síntese de biofilmes nos mais diversos ambientes sugere que estes microrganismos
têm a capacidade de responder às mudanças de condições do ambiente alterando a sua SPE e
habilidade de adesão dependendo a superfície a qual estão aderidos (AHIMOU et al., 2007). Em
superfícies sólidas a colonização microbiana pode ser afetada por vários parâmetros, numa

40
superfície cheia de reentrâncias, por exemplo, há mais facilidade de formação de biofilmes por
prover mais áreas de contato.
A colonização em superfícies sólidas pode ser afetada por diversos parâmetros, por
exemplo, o grau de colonização de certas superfícies aumenta com a rugosidade da superfície,
pois esta oferece reentrâncias onde a colônia pode se fixar em uma área relativamente protegida
reduzindo forças e tendo provida uma superfície maior para adesão (DONLAN, 2002).
Os microrganismos aderem mais rapidamente a superfícies hidrofóbicas e não polares do
que a superfícies polares e hidrofílicas.
A presença de flagelos e o grau de produção de SPE são fatores que afetam
consideravelmente a taxa e o grau de anexação das células microbianas a diferentes superfícies
(DONLAN, 2002).
A constituição e a quantidade do biofilme depende do material disponível, do nitrogênio
ou do oxigênio disponibilizado, do tipo de microrganismo, temperatura, idade do microrganismo,
pH e da disponibilidade de nutrientes. Há biofilmes compostos por ácidos nucleicos e proteínas,
já os biofilmes produzidos por kefir são polissacarídeos.
O kefir atua como probiótico e impede que ocorra nos biofilmes a proliferação de seres
patogênicos cujo comportamento é individualizado com relação aos seres que compõem os
probióticos. Somente seres com capacidade especial conseguem formar consórcios com outros
seres (LAUREYS; DE VUYST, 2014). Além de organismos probióticos, repelirem
contaminantes através da competição por sítio e por nutrientes o kefir secreta biocidas que
repelem os seres indesejáveis (KOJIĆ et al., 2007; LENGKEY et al., 2013).
Nas bactérias gram-negativas alguns dos polissacarídeos são neutros ou polianiônicos. A
presença de ácido urônico ou piruvato cetal-ligado habilita suas propriedades aniônicas, e então
permite a associação de cátions divalentes tais como cálcio e magnésio os quais aumentam a
força de ligação no biofilme desenvolvido (DONLAN, 2002; SUTHERLAND, 2001).
Há três tipos de forças envolvidas no processo de formação de biofilmes, são as
interações eletrostáticas, as pontes de hidrogênio e as forças de dispersão de London (MAYER et
al, 1999). Estas forças associadas contribuem para a estabilidade da matriz do biofilme
(FLEMMING; WINGENDER, 2001b).
Outro fator que contribui para a variação da capacidade de aderir do biofilme é a
diferente composição do biofilme que influencia a extensão mediante a qual o microrganismo

41
possa aderir tanto a superfícies hidrofílicas como a hidrofóbicas. O crescimento do biofilme
resulta de um processo complexo que envolve transporte de moléculas orgânicas e inorgânicas e
células microbianas para a superfície e subsequente adsorção a superfície e finalmente adesão
definitiva, corroborada pela produção de SPE (VAN HULLEBUSCH, 2003; ROMANI et al.,
2008).
As células em um biofilme são envolvidas com uma matriz composta basicamente de
substância polissacarídeo extracelular (SPE) que provê proteção à bactéria contra uma variedade
de causadores de estresse ambiental tais como radiação UV, pH, choque osmótico e dissecação
(FLEMMING, 1993; WATERS; BASSLER, 2005; RUIZ et al., 2004).
Devido à complexidade, a formação de biofilme é regulada em diferentes vias por
diversos mecanismos, dentre este a mais estudada é a formação de biofilme e diferenciação pela
regulação Quórum Sensing (QS).
O Quórum Sensing (QS) permite que bactéria mantenha comunicação célula a célula e
regula a expressão de genes em resposta a densidade (HOOSHANGI, S.; BENTLEY, 2008). Em
uma determinada densidade de população os genes envolvidos na diferenciação e maturação do
biofilme são ativados.
Desta forma seres considerados tradicionalmente como unicelulares passam a agir em
grupo como um único ser multicelular, tendo sua ação intensificada (RUMJNEK et al., 2004).
Estas células associadas a biofilmes apresentam expressão ao gene específico que muitas vezes é
controlada pelo Sistema de Quórum Sensing para permitir seu aumento em resistência
(SHIMADA et al., 2012).
As diferenças que existem no sistema de Quórum Sensing são devidas ao fato que cada
sistema necessita ser otimizado para promover a sobrevivência no nicho especializado no qual
uma espécie particular reside. É por isso que em cada Sistema de Quórum Sensing vamos
encontrar o tipo de sinal específico, receptor, mecanismo de transdução e alvos refletindo uma
única espécie particular de bactéria. A habilidade em se comunicar é fundamental às bactérias
(WATERS; BASSLER, 2005).
O primeiro sistema de Quórum Sensing foi detectado na bactéria Vibrio fischeri, e é
considerado um paradigma do que ocorre com as bactérias gram-negativas com relação a via de
Quórum Sensing (NEALSON; HASTINGS, 1979).
Seres abissais bioluminiscentes do gênero Eurpyma scolopes têm seus órgãos

42
bioluminiscentes colonizados por bactérias Vibrios fischeri. Nestes órgãos a bactéria cresce e ao
atingir determinada densidade induz expressão dos genes requeridos para bioluminescência. A
lula (Eurpyma scolopes) pode usar esta biolumenescência para mascarar sua sombra e desta
forma evitar a predação, enquanto a bactéria se beneficia porque este órgão é rico em nutrientes e
permite sua proliferação em número que seria inatingível no ambiente marinho (NEALSON;
HASTINGS, 1979).
Quórum Sensing (QS) explica também porque em alguns casos não há formação de
biofilme, pois não havendo a densidade populacional suficiente os genes para formação de
biofilmes não são ativados.
Não obstante saber-se que os microrganismos existem predominantemente em
comunidades multicelulares dentro de biofilmes na maioria dos ambientes, este sistema complexo
tem que ser estudado ainda para se compreender melhor a complexidade das interações dentro
dos biofilmes bem com sua função na anexação e na proliferação bacteriana.
2.2.1 Aplicações de biofilmes
As Substâncias Poliméricas Extracelular (SPE) estão relacionadas com consequências
tanto benéficas como deletérias dos agregados, tais como, formação de biofilmes, floculação e
sedimentação biológica. Em bioincrustação estas substâncias aumentam a resistência à fricção,
alteram as propriedades da superfície tais como a hidrofobicidade, aspereza, textura, cor, entre
outras. Em biocorrosão de metais elas estão envolvidas através da sua propriedade de se ligar a
íons de metal provocando ou acelerando processos corrosivos. Em biodecomposição de rochas
elas contribuem por sua propriedade de dissolução de metais.
As SPE absorvem substâncias poluentes tais como metais e moléculas orgânicas. Tanto
podem lixiviar águas de esgoto como podem ser usadas para purificar a água devido a suas
propriedades de absorção. Em biotecnologia pode ser explorada sua capacidade de alterar a
viscosidade de alimentos, tintas e óleos hidráulicos. Devido suas propriedades hidratantes podem
ser usadas na indústria de cosméticos e na indústria farmacêutica. Possíveis aplicações como
biosurfactante e como cola biológica são também previstas. As EPSs são um interessante
componente dos biofilmes e como tal contêm um imenso potencial de aplicações nas mais
diversas áreas (FLEMMING; WINGENDER, 2001).

43
2.2.2 Biofilmes e biorremediação
Interações físicas e fisiológicas entre os microrganismos em um biofilme disponibilizam
os nutrientes ao mesmo tempo em que remove potenciais metabólitos tóxicos (O'TOOLE, 2000).
As biomembranas têm despertado interesse intensificado em várias áreas como indústria
alimentícia, farmacêutica, embalagens e, mais recentemente, biorremediação. Isto se deve a sua
diversidade estrutural e suas propriedades reológicas peculiares (FIALHO, 2008). Estudos
recentes sugerem que biorremediação através de biofilme pode ser uma alternativa proficiente
para remediação em relação a se fazer uso de microrganismos planctônicos (SINGH et al., 2006).
Sua aplicação em biorremediação de efluentes criados pela mineração tem sido um
campo muito promissor (GAYLARD et al., 2005; PEREIRA; FREITAS, 2012).
A utilização de biofilme tem sido considerada a melhor alternativa para biorremediação
do que aplicação de bactérias planctônicas, visto que células formadoras de biofilmes têm maior
possibilidade de adaptação a diferentes ambientes (COSTERTON et al., 2003; SINGH et al.,
2003).
A propriedade dos biofilmes de remover metais pesados do ambiente se deve a sua
capacidade de floculação que lhe confere habilidade de se ligar aos íons de metais pesados das
soluções (PAUL; PAUL, 2008).
As bactérias redutoras de sulfato (BRS) são encontradas frequentemente em águas
contaminadas com metais pesados. São altamente eficientes na degradação anaeróbica de muito
poluentes orgânicos e na precipitação de metais pesados de águas de esgoto (SINGH;
CAMEOTRA, 2004).
2.2.1.2 Biorremediação de pesticidas
A biorremediação tem sido geralmente considerada um método menos dispendioso e
mais eficiente para desintoxicação de vários compostos xenobióticos e poluentes (VERHAGEN
et al., 2011). Por ser um processo natural promove um tratamento adequado ao meio, seu custo é
relativamente baixo quando comparado a outras alternativas convencionais de tratamento de
resíduos sólidos. Não obstante, para se obter elevado rendimento no processo, é necessário se

44
determinar quais são as condições que favorecem a atividade microbiana, como por exemplo:
meio anóxico, teor de nutrientes elevado, tempo de retenção, atividade enzimática, temperatura,
pH, e inóculo aclimatado ao meio.
O uso de sistemas de biorremediação têm sido propostos para tratar a fonte da
contaminação por pesticidas (DE WILDE et al., 2007), porquanto os mesmos apresentam
resistência à hidrólise e oxidação. Assim a degradação bacteriana é naturalmente a via dominante
de eliminação de pesticidas no ambiente (WOLFE et al., 1978). A capacidade degradação de
pesticidas destes sistemas pode ser habilitada através da utilização de bactérias especializadas em
degradação de pesticidas isoladamente ou em consórcio com outras (VERHAGEN et al., 2011).
Têm sido empreendidos para isso monoculturas e culturas mistas capazes de degradar pesticidas
(VERHAGEN et al., 2011).
Em muitas estratégias de isolamento o enriquecimento é processado em meio líquido
agitado, o que favorece à bactéria crescer bem em suspensão. Entretanto na natureza, muitas
bactérias crescem agregadas cada uma a outra e à superfície (COSTERTON et al., 2007). Uma
vez que biofilmes são o modo predominante de vida das bactérias (PARSEK; FUQUA, 2004;
MOONS et al., 2009) o uso de meio liquido com agitação pode ser representar um obstáculo e
isto é um importante fator a ser considerado porque bactérias que se proliferam em biofilme
crescem muito pouco em suspensão e podem ser ineficientes quando este meio de seleção é usado
(VERHAGEN et al., 2011).
2.2.3 Forças de adesão envolvidas em síntese de biofilmes
Muitos problemas causados por biofilmes advêm do fato de que eles têm alta resiliência
além de extraordinária capacidade de conter forças mecânicas. Permanece ainda enigmático
como as propriedades destas forças mecânicas dos biofilmes habilitam a bactéria a sobreviver em
diferentes ambientes químicos (GRUMBEIN et al., 2014).
Devido a estes fatos é importante a compreensão de como se formam e que forças estão
envolvidas neste processo. A compreensão das forças envolvidas na adesão do microrganismo é
de fundamental importância para áreas como microbiologia objetivando elucidar as funções
celulares, mas também é de interesse da medicina no que concerne às infecções e ferimentos bem

45
como para a biotecnologia no estudo da agregação das células. (DUFRÊNE, 2015). Hoje já se
sabe que a maioria das infecções são causadas por biofilmes cujo perfil de crescimento habilita a
patogenicidade da bactéria. Uma vez agregados aos biofilmes as bactérias ficam englobadas por
uma substância protetora que é a matriz da substância polimérica extracelular (SPE)
(HARAPANAHALLI et al., 2015).
Entre as várias estratégias utilizadas para estudo dos biofilmes tem se destacado o uso da
Microscopia de Força Atômica (AFM) (DUFRÊNE, 2015).
Há, portanto poderosas forças que atuam na formação dos biofilmes e na adesão das
bactérias aos mesmos. A adesão não específica de bactérias é o primeiro passo para a formação
de biofilmes em superfícies abióticas. As forças que atuam nesse processo são as Forças de Van
der Walls além de forças eletrostáticas. Interações hidrofóbicas de curto espectro desempenham
importante papel. São forças tanto que atraem como repelem a bactéria individualmente desta
superfície. À distância de 50 nm forças atrativas da superfície já são detectadas o que indica uma
larga faixa de forças, mas há evidências de que as proteínas da parede celular das bactérias são as
responsáveis pela adesão interagindo com forças hidrofóbicas da superfície (THEWES et al.,
2014). Experimentos demonstram que as células que não compõem os biofilmes são excluídas e
mesmo quando crescem próximo ao biofilme são removidas por forças. Estas mesmas forças
protegem que adentrem nos biofilmes outras células (THEWES et al., 2014). A proteína RbmA
enlaça as células mães com as células filhas ligadas na matriz conferindo uma proteção às células
que produzem o biofilme enquanto outras são exclusas ou repelidas (THEWES et al., 2014).
Kannan et al. (2014) estudaram as forças de adesão de biofilmes formados por
Pseudomonas aeruginosa em poros de silicone os quais danificam aparelhos semicondutores e
implantes. Fazendo uso de microscopia de força atômica (AFM) constataram que ocorre uma
variação da rugosidade nestes biofilmes a qual foi quantificada. Os resultados mostraram um
aumento na força de adesão com o aumento da rugosidade na superfície concluindo, portanto que
a força de adesão do biofilme é afetada pela rugosidade.
Há também evidências de que os biofilmes podem utilizar a capacidade de absorção de
certos metais em forma iônica como zinco, cobre, ferro e alumínio contidos na matriz do biofilme
para evitar erosão por forças. Curiosamente muito destes íons metálicos são tóxicos para as
formas planctônicas das bactérias componentes do biofilme, mas eles têm sua toxidade suprimida
na matriz dos biofilmes (o que sugere também a importância de biofilmes para biorremediação).

46
Assim os biofilmes claramente cumprem funções a destacar quais sejam a regulação das
propriedades mecânicas dos biofilmes e a formação de uma barreira seletiva para conter
substâncias tóxicas (GRUMBEIN et al., 2014).
A resistência das bactérias a antibióticos diferentemente da resistência convencional
(genética) disponibiliza também de um mecanismo através da formação de biofilmes os quais
reduzem a suscetibilidade a antibióticos e contribuem para a persistência das infecções
(STEWART, 2002). As bactérias crescem agregadas em biofilmes os quais habilitam sua
proteção contra estresse tanto físico como químico (TAYLOR et al., 2014; ZHANG et al., 2015)
além de facilitar a disponibilidade de nutrientes conferem também habilidade para sobreviver em
condições ambientais adversas. Os biofilmes conferem resistência a antibióticos bem como
dificultam o cuidado com ferimentos, pois são recalcitrantes à remoção bem como ao
debridamento dos mesmos (BIRKENHAUER et al., 2014; TAYLOR et al., 2014).
Foi descoberto que muitas feridas se tornam crônicas devido à formação de biofilmes os
quais impedem a entrada de antibióticos e facilitam a bactéria criar resistência aos mesmos uma
vez que a dosagem aplicada é insuficiente para exterminá-las, mas suficiente para criar
resistência. Os biofilmes são recalcitrantes à remoção através da formação de uma matriz
protetora que protege os organismos residentes de forças inimigas. (BIRKENHAUER et al.,
2014.).
Esta proteção pode ser mecânica, ou seja, o biofilme constitui uma barreira que impede a
penetração do antibiótico, mas também se caracteriza pela diferenciação dos microrganismos que
compõem os biofilmes que se tornam mais aptos e mais fortes quando em biofilmes. Diante
destes fatos é notória a necessidade de se utilizar novas estratégias que envolvam o combate à
formação de biofilmes antes de se utilizar antibióticos. Os campos de combates são escrutinados
a partir do conhecimento das características dos biofilmes, como aderência à superfícies,
formação de matriz extracelular, quórum sensing, regulação de maturação dos biofilmes e
dispersão, estes são os alvos a serem explorados no desenvolvimento de tratamento específico
contra biofilmes (TAYLOR et al., 2014). Em outra estratégia genes responsáveis pela formação
de biofilmes estão sendo alvos de quimioterapia, pois entende-se que ao desabar a resistência dos
biofilmes habilita-se o antibiótico para atuar nas infecções que são refratárias ao tratamento
convencional (STEWART, 2002).

47
Recentemente tem sido comprovado que os probióticos inibem a formação de biofilmes
por microrganismos patogênicos, que consiste no primeiro passo para a instalação da infecção
(FERNANDEZ et al., 2015; VIDYALAXME et al., 2014).
Além de mecanismos de competição os probióticos engatilham as células do Sistema
Imunológico do hospedeiro e também secretam biocidas que atacam possíveis invasores
(FEDORAK; MADSEN, 2004; RASTALl et al., 2005; SAAD, 2006).
2.2.4 Propriedades reológicas dos biopolímeros
Para entender a microestrutura destes filmes é necessário o estudo de suas propriedades
reológicas. De fato, o conhecimento destas propriedades é de grande utilidade para proporcionar
uma mais ampla compreensão das propriedades dos biofilmes (XU et al., 2004).
A alta viscosidade de um filme pode, por exemplo, formar uma solução que retém
bolhas de ar tornando difícil sua remoção e oculta moldes em suas finas camadas (CUQ et al.,
1993), por isso soluções formadoras de filmes de baixa viscosidade têm atraído os pesquisadores
(MORAES et al., 2009). Para que se forme uma camada de mesmo nível sobre uma superfície
sólida a viscosidade da solução formadora do filme deve evitar formar depressões por efeito de
gravidade e permitir que a capilaridade seja uniforme no filme (PERESSINI et al., 2003). O
conhecimento das propriedades reológicas que são sensíveis a variações da estrutura molecular é
útil no desenvolvimento de relações função-estrutura para sistemas de soluções polissacarídeas
(XU et al., 2004). Por isso a caracterização das soluções formadoras de filme contribui para a
compreensão das diferenças nas propriedades finais do filme (XU et al., 2004).
Não obstante o kefirano ser sugerido para formação de embalagens o fato de permitir a
passagem de umidade é uma característica indesejável para este objetivo e por isso tem se
procurado impermeabilizadores como glicerol para proporcionar a retenção do líquido e tornar a
membrana viável para este uso. O uso destes impermeabilizadores podem causar alterações nas
propriedades reológicas do biofilme, alterações estas que podem ser estudadas através de várias
estratégias tecnológicas tais como Espectroscopia de Infravermelho por Transformação de
Fourier (FT-IR), Difração de Raio X (XRD), Microscopia de Tunelamento (STM) e Microscopia
de Força Atômica (AFM) (GHASEMLOU et al., 2011).
O uso de Microscopia de Força Atômica (AFM) tem sido uma das ferramentas para este

48
estudo uma vez que se alcançou um grande avanço estes últimos anos concomitantemente com a
Microscopia Eletrônica de Varredura (MEV). Ghasemlou et al. (2011), fez uso destas tecnologias
para analisar as alterações reológicas causadas pela utilização de impermeabilizadores glicerol e
sorbitol sobre o kefirano.
Kodali et al. (2009) constataram a produção de uma substância polimérica extracelular
(SPE) por B. coagulans o qual é um heteropolímero composto de galactose, manose e fucose,
glicose e glicosamina. Com significativa atividade emulsificante em óleos vegetais. A fucose foi
pela primeira vez identificada como componente de um exopolissacarídeo. Sua estrutura
caracterizada pela repetição das suas unidades de açúcares unidas através de ligações glicosídicas
constituem os blocos de construção que formarão o esqueleto da estrutura molecular do biofilme
de exopolissacarídeo (SPE) (KODALI et al., 2009).
A presença de numerosos grupos hidroxilas na estrutura, característica comum aos
açúcares, viabilizam que ocorram as ligações de hidrogênio, formando assim cadeias tanto
lineares como helicoidais num entrelaçamento bi e tridimensional estabelecendo uma estrutura
em rede (CHOWDHURY et al, 2011).
2.2.5 Métodos físicos para estudo dos biofilmes
2.2.5.1 Utilização de microscopia de força atômica (AFM)
A microscopia de Força Atômica ou “Atomic Force Microscopy” (AFM), foi
desenvolvida em 1986 por Gerd Binnig, Calvin Quate e Christoph Gerbver. A AFM permite
obter imagens reais, em três dimensões, da topografia das superfícies, com uma resolução
espacial que se aproxima das dimensões atômicas (BAKER et al., 1997).
A AFM traduz em imagens as propriedades interfaciais dos eletrodos e possibilita a
observação direta da arquitetura da superfície. Diante destes fatos esta técnica pode trazer
informações importantes sobre a morfologia da superfície dos eletrodos modificados com
moléculas biológicas (CHIORCEA PAQUIM et al., 2004; OLIVEIRA BRETT et al., 2005).
Uma pequena ponta de quartzo acoplada a uma consola que fica fixada a um suporte
escaneia a superfície da amostra nas três dimensões do espaço; semelhante a uma agulha de toca-
discos de vinil. Esse consola, chamado de cantlever, é uma haste flexível que retém em sua

49
extremidade a agulha com dimensões de poucos micra. A imagem é obtida através de um sistema
de posicionamento com cerâmicas piezoelétricas, capazes de realizar movimentos nas três
direções (xyz), com precisão de ângstrons (Å).
Durante a varredura da amostra há interações entre o cantilever e os átomos da amostra.
Estas interações podem ser atrativas ou repulsivas sendo que as atrativas são devido às forças de
Van der Walls e ocorre entre distâncias maiores. Quando aproximamos a agulha da amostra
ocorre a repulsão entre os orbitais eletrônicos dos átomos da superfície da amostra e os da ponta
do microscópio de força atômica. Forças de atração podem ter origem tanto em fatores físicos,
como a capilaridade, ou químicos como a afinidade entre o cantilever e a amostra. A força de
repulsão se deve a interação coulombiana (GIESSIBL, 2003).
São as forças resultantes destas interações que fazem com que o cantilever (de 100 a
200 mm de comprimento) se aproxime ou se afaste gerando deflexões. Na cantilever ou consola é
fixado um pequeno elemento piezelétrico (modulador piezo) para conduzir o feixe de laser na sua
frequência de ressonância e então conecta-se a amostra ao piezoelétrico tridimensional, o scanner
x,y,z. Na parte superior da haste existe uma superfície espelhada, que reflete a luz de um feixe de
laser, que em seguida passa através de uma lente e incide sobre um fotodetector (matriz de
fotodiodos) cujo sinal de saída é recolhido por um amplificador diferencial, que mede as
variações de posição e da intensidade da luz produzidas pelas deflexões da consola. Assim os
movimentos nanométricos podem ser detectados, gerando sinais elétricos que são armazenados e
processados por um computador e convertidos em imagem topográficas, bi ou tridimensionais da
superfície do material em resolução atômica (RUBIO-SIERRA et al., 2005) conforme está
demostrado na figura 2.
Durante esta varredura, é utilizado um sistema de alinhamento, onde um feixe de laser
incide sobre o cantilever e refletindo em um sensor, de quatro quadrantes, fornece informação de
posição para o sistema de realimentação e controle. Este corrige a posição do cantilever de forma
a manter o contato com a amostra, durante a varredura e permitir a obtenção da imagem
(GIESSIBL, 2003).

50
Figura 2 - Diagrama de funcionamento do microscópio de varredura de força
Fonte: Herman et al. (1997)
A imagem obtida na AFM é resultante da convolução da topografia real da amostra com
a forma da agulha do cantilever. Este é uma das principais fontes de artefatos de imagem nesta
técnica.
As diferentes técnicas fornecem diversas possibilidades para gerar imagens de diferentes
tipos de amostras e para gerar uma ampla gama de informações. Os modos de fazer as imagens,
também chamados modos de varredura ou de operação, referem-se fundamentalmente à distância
mantida entre a sonda (ponteira) e a amostra, no momento da varredura, e às formas de
movimentar a ponteira sobre a superfície a ser estudada. Há um contínuo de modos possíveis de
fazer imagens, devido às diferentes interações em função da distância entre a ponteira e a
amostra, assim como ao esquema de detecção utilizado. A escolha do modo apropriado depende
da aplicação específica que se deseja fazer e de vários fatores os quais condicionam a obtenção
da imagem de diferentes modos, como o tipo de amostra e as condições da mesma, o tipo de
cantilever utilizado e o tipo de varredura entre outros. No caso da amostra é importante se ela é
rígida ou não, pois isto vai determinar se a varredura foi de contato utilizando o modo contato,
contato intermitente ou não contato. Geralmente se faz as imagens de amostras de filmes finos
nos modos de contato intermitente ou não-contato, pois assim as interações do cantilever com a
amostra são menos severas, o que diminui a possibilidade de danos tanto na amostra como na

51
agulha (GIESSIBL, 2003).
2.2.5.2 Microscopia Eletrônica de Varredura (MEV)
O microscópio eletrônico de varredura (MEV) é um equipamento capaz de produzir
imagens de alta ampliação (até 300.000 x) e resolução e é considerado um dos melhores
dispositivos para se analisar as características microestruturais de materiais sólidos. Foi
idealizado primeiramente por Knoll (1935) que descreveu o conceito do MEV. Em 1938 Von
Ardenne construiu um microscópio eletrônico de varredura e transmissão (STEM) adaptando
bobinas de varredura a um microscópio eletrônico de transmissão. O princípio de funcionamento
do MEV consiste na emissão de feixes de elétrons por um filamento capilar de tungstênio
(eletrodo negativo), mediante a aplicação de uma diferença de potencial que pode variar de 0,5 a
30 KV. Essa variação de voltagem permite a variação da aceleração dos elétrons, e também
provoca o aquecimento do filamento. A parte positiva em relação ao filamento do microscópio
(eletrodo positivo) atrai fortemente os elétrons gerados, resultando numa aceleração em direção
ao eletrodo positivo. A correção do percurso dos feixes é realizada pelas lentes condensadoras
que alinham os feixes em direção à abertura da objetiva. A objetiva ajusta o foco dos feixes de
elétrons antes dos elétrons atingirem a amostra examinada.
A microscopia eletrônica de varredura (MEV) tem sido muito utilizada no estudo dos
biofilmes. Sousa et al. (2015) reportam sobre o uso da técnica para se estudar a estrutura de
biofilmes de Nontuberculous mycobacteria (NTM). Esta bactéria tem a capacidade de agregar
biofilmes em diferentes superfícies. Os biofilmes podem atuar como reservatório ambiental. Esta
possibilidade de agregar biofilmes em diferentes superfícies foi correlacionada com a capacidade
de se dispersar sobre o meio sólido (SOUSA et al., 2015).
A microscopia eletrônica de varredura (MEV) foi utilizada para avaliar a presença de
microrganismos aderidos à superfície de mini-implantes que falhavam devido à perda de
estabilidade (FERREIRA et al., 2015) bem como a de adesão e formação de biofilmes por
Streptococcus mutans e Candida albicans (LAVIN et al., 2015).
Microrganismos de diferentes espécies interagem em vários nichos ecológicos
eventualmente causando infecção. Durante este processo de infecção pode ser formado biofilmes
com apenas uma espécie de microrganismo ou por multiespécies. Através de técnicas de

52
microscopia incluindo microscopia eletrônica de varredura (MEV), Granillo et al. (2015)
estudaram biofilmes formados por Aspergillus fumigatus e Staphyloccocus aureus que são agente
etiológicos responsáveis por causar ceratite. A adesão entre bactéria e fungos é fundamental para
a formação de biofilmes polimicrobianos (Frey-Klett et al., 2011).
2.3 COMPOSTOS FENÓLICOS E SUA IMPORTÂNCIA
Nos últimos anos os efeitos de compostos fenólicos têm sido constantemente estudados
devido suas propriedades antioxidantes com ação preventiva e terapêutica contra várias doenças
crônico-degenerativas tais como a hipercolesterolemia, câncer dos seios e da próstata e doenças
cardiovasculares (CROZIER et al., 2009; GARCIA-LAFUENTE et al., 2009). Dentre os
compostos fenólicos se destacam os flavonóides dos quais fazem parte as isoflavonas.
Os efeitos biológicos dos compostos fenólicos dependem da sua origem e da quantidade
consumida na dieta, pois há uma correlação positiva entre o consumo e seus efeitos benéficos à
saúde dependendo obviamente de sua biodisponibilidade. A biodisponibilidade ocorre em função
da absorção intestinal, da bioconversão por colônias da microbiota, da dispersão pelos tecidos do
organismo e, finalmente, da eliminação.
A capacidade dos flavonoides sequestrar espécies reativas de oxigênio e nitrogênio é
decorrente da presença de uma orto-hidroxilação no anel B da molécula de flavonóides (figura 3),
do número de hidroxilas livres e uma dupla ligação na posição C2-C3 ou a presença de uma
hidroxila no carbono C3 (BURDA; OLESZEK, 2001). Mas, não se sabe exatamente de que
maneira os compostos fenólicos atuam na atividade antioxidante, há possibilidade que a mesma
seja decorrente da ação sinergética entre diversos compostos fenólicos e não advinda apenas de
um único componente (ARNOUS et al., 2001; LEE et al., 2003). Há, entretanto, evidências
comprobatórias de que a atividade antioxidante dos compostos fenólicos cresce com o aumento
no número de grupamentos hidroxila e diminuição de grupos glicosilados (FUKUMOTO;
MAZZA, 2000) e que antioxidantes fenólicos funcionam como sequestradores de radicais e
algumas vezes como quelantes de metais (AMES et al., 1993; RICE-EVANS et al., 1996).
Os compostos fenólicos são produzidos pelo metabolismo secundário das plantas e são
encontrados principalmente nas frutas. Recentemente as frutas têm sido consideradas alimentos
funcionais devido ao seu baixo conteúdo calórico em comparação com o uso de carboidratos

53
industriais, aliada às suas propriedades como antioxidantes além de conterem importantes
compostos nutricionais tais vitaminas, fenóis, minerais e fibras os quais numa ação sinergética
desempenham importante papel como preventivos de estresse oxidativo (ORAZEM et al., 2011).
Há fortes evidências obtidas em estudos recentes relacionando a alta incidência de câncer
com deficiência no consumo regular de frutas e verduras (KOBORI, 2003). Não apenas o
aparecimento de câncer de pulmão e de estômago dentre outros, mas também doenças
cardiovasculares, cataratas e disfunções do sistema imunológico e doenças neurovegetativas se
relacionam com deficiências nutricionais marcadamente de frutas e vegetais (VATTEM et al.,
2005), por isso estudos epidemiológicos têm apresentado fortes evidências de que o aumento no
consumo de frutas está diretamente ligado à diminuição de riscos de várias doenças vegetativas
como arteriosclerose, desordens do coração e do cérebro e diferentes tipos de câncer (HEGEDÚS
et al., 2010).
As principais substâncias advindas dos vegetais e responsáveis pelo efeito protetor são os
carotenoides, os flavonoides os isoflavonoides e ácidos fenólicos (BOYER; LIU 2004;
GRAMZA; KORCKZAC, 2005). Os flavonoides previnem o câncer e o mecanismo através do
qual este efeito protetor é exercido ocorre através da sua ação interferindo em passos que
conduzem a formação de tumores, como também protegem o DNA de danos oxidativos e desta
forma inibem a ativação do câncer (GALATI; O’BRIEN, 2004).
As antocianinas suprimem a proliferação de células oncogênicas, portanto ajudam a
evitar câncer além de diabetes e problemas cardiovasculares e neurológicos (KONCZAK;
ZHANG, 2004; LILA, 2004). O valor do uso tradicional de antocianina para reduzir a
hipertensão tem sido confirmado cientificamente (KONCZAK; ZHANG, 2004; LILA, 2004).
Assim também o efeito benéfico de seu uso para combater infecção do trato urinário tem sido
confirmado pelo poder anti-inflamatório e antimicrobiano das antocianinas (KONCZAK;
ZHANG, 2004).
O uso de fitoquímicos é uma das armas que podem ser empregadas tanto para evitar
como combater o estresse oxidativo. Muitas plantas têm sido descobertas no sentido de se evitar
formação de radicais livres ou de combater o estresse oxidativo (IVORRA et al., 2015).
O corpo usa de defesas antioxidantes para manter o processo oxidativo dentro dos
limites fisiológicos do contrário acontecem com os danos oxidativos uma ampliação e
consequentes danos sistêmicos

54
De maneira inevitável os processos metabólicos que nos permitem viver geram também
radicais livres os quais se produzidos em grande quantidade sem serem inativados podem trazer
danos às células. Os mecanismos de geração de radicais livres ocorrem, sobretudo, nas
mitocôndrias, membranas celulares e no citoplasma (BARBOSA et al., 2010). Os radicais livres
são átomos ou moléculas ou mesmo fragmentos de molécula que contêm elétrons não carregados.
O organismo produz substâncias antioxidantes que conseguem inibir e reduzir lesões causadas
por radicais livres. Além disso, o uso de substâncias naturais antioxidantes trazem benefícios
incontestáveis na preservação da saúde.
Pode ser considerado um radical livre qualquer átomo, molécula com elétrons
desemparelhados na camada de valência podendo se ligar ou atrair elétrons de outra molécula e
com isso desestabilizando genes, proteínas ou mesmo células. Podem por exemplo despertar um
oncogene ou causar envelhecimento precoce. Exemplos de radicais livres são: oxigênio
molecular (O2), radical hidroxiol (OH-), ânion superóxido (O-), radical peroxil (ROO), radical
alcoxil (RO) e óxido nítrico (NO) (WISEMAN, 2005).
Os radicais livres são moléculas altamente instáveis, com meia-vida muito curta, pois
são moléculas muito reativas podendo sequestrar elétrons de moléculas importantes. Podem ser
muito prejudiciais para o cumprimento e manutenção das funções fisiológicas vitais,
paradoxalmente as mesmas que dão origem aos mesmos.
Há mecanismos de defesa antioxidante criados pelo organismo para limitar ou impedir
os danos a nível intracelular. São substâncias denominadas antioxidantes que inibem ou reduzem
as lesões causadas pelos radicais livres protegendo a célula contra efeitos dos mesmos (SOARES,
2002).
O estresse oxidativo é causado por uma produção excessiva de radicais livres e podem
causar dano e morte celular. O cérebro é particularmente suscetível ao estresse oxidativo, pois
consome em torno de 20 % de todo oxigênio absorvido (LAU et al., 2005) e 25% de toda glicose
obtida na nutrição. Para lidar com isso geralmente durante o estresse ocorre um aumento de
defesas antioxidantes enzimáticas.
O estresse oxidativo pode ter várias causas dentre as principais consumo de substâncias
lesivas como álcool, tabagismo e drogas, exercício pesado, preocupação excessiva, reação
depressiva diante de eventos conturbados peculiares da existência, etc.

55
2.4 FLAVONÓIDES
Flavonoides são substâncias de baixo peso molecular produzido pelo metabolismo
secundário das plantas que exercem efeito protetor antioxidante ou proteção aos raios UV. São
encontrados em bebidas como o chá verde, em frutas, em vinhos, em sementes como soja além de
plantas de uso tradicional como gingko biloba e ginseng. Mais de 4000 flavonoides de diferentes
ocorrências naturais já foram descritos na literatura (KING; YOUNG, 1999; GALATI;
O’BRIEN, 2004; HOUSTON, 2005).
Os flavonoides desempenham um papel fundamental na proteção contra agentes
oxidantes, como raios ultravioletas, poluição ambiental, substâncias químicas presentes nos
alimentos, entre outros. Atuam também como agentes terapêuticos em um elevado número de
patologias, tais como aterosclerose e câncer. Dado que não podem ser sintetizados pelo nosso
organismo, sendo representativos da parte não energética da dieta humana, são obtidos através da
ingestão de alimentos que os contenham ou de suplementos nutritivos (GONÇALVES, 2008;
ALVES et al., 2012).
Dentre os compostos fenólicos presentes em frutas e vegetais, os mais comumente
encontrados são os flavonoides. As propriedades benéficas destes compostos podem ser
atribuídas à sua capacidade de sequestrar os radicais livres, podendo inibir os processos da
oxidação em certos sistemas.
Compostos fenólicos são responsáveis por propriedades terapêuticas de grande interesse
para confrontar os problemas advindos da vida moderna, propriedades como oxidação de
gorduras e diminuição de problemas cardiovasculares, aumento de consumo energético e redução
de risco de incidência de câncer (TREVISANATO; KIM, 2000; ALVES et al., 2012). Estes
efeitos são advindos da sua capacidade de inativar os radicais livres ao reagir com eles formando
radicais estáveis (ALVES et al., 2012). A estrutura química dos compostos fenólicos contém pelo
menos um anel aromático com grupamento hidroxila que lhe confere o poder de neutralização de
radicais (GIADA; MANCINI-FILHO, 2006).
Há interesse em se encontrar substâncias antioxidantes naturais para nutrição ou
aplicação farmacológica. Ao transgredir as normas para uma vida saudável através do
sedentarismo e da alimentação desregrada o homem procura na natureza compostos que lhe
permita conviver com seus vícios sem necessitar de mudanças radicais em seu modo errôneo de

56
se alimentar. Com efeito, o uso de produtos naturais poderá vir a contribuir para melhoria da
qualidade de vida e precaução contra os principais problemas de saúde, no entanto
concomitantemente se torna necessário combater o sedentarismo e o uso indiscriminado de
produtos lesivos ao organismo tais como uso excessivo de frituras, consumo de álcool,
tabagismo, etc. Em 1997, Decker já advertia que não obstante os flavonóides comportarem
propriedades benéficas advindas de sua capacidade de sequestrar os radicais livres, podendo
inibir os processos da oxidação em certos sistemas, isso não significa que eles possam proteger as
células e os tecidos de todos os tipos de danos oxidativos.
O uso de antioxidantes artificiais tem sido restringido em virtude de sua toxidade.
Busca-se, portanto, encontrar substâncias antioxidantes naturais (DEGÁSPARI;
WASZCZYNSKYJ, 2004). Embora o uso de frutas e verduras tenha seu uso consagrado pelo
conteúdo de vitaminas, fibras e minerais é o seu poder antioxidante encontrado em outros
nutrientes como compostos fenólicos, fenóis e flavonoides dentre os quais as antocianinas, que
faz com que haja um crescente aumento de interesse e de consumo dos mesmos (SILVA, 2007).
Os flavonoides participam da regulação de atividade enzimáticas para produção de
substâncias responsáveis pelo flavor. É importante ressaltar que o sabor adstringente de frutas é
proveniente da ação de flavonoides precipitando proteínas que não se dissolvem na boca
(SKREDE; WROLSTAD, 2002; GRAMZA; KORCZAK, 2005). Há, portanto uma correlação
entre o sabor adstringente (responsável pela sensação de “amarrar a boca” bem como a sensação
de sabor amargo) com o conteúdo de compostos fenólicos (SLIMESTAD et al., 2009, BRAVO et
al., 1998). Os mesmos compostos fenólicos desempenham papel importante no mecanismo
natural de defesa e nas propriedades benéficas dos frutos (SLIMESTAD et al., 2009).
Ao lado de tudo isto os flavonoides são de particular interesse devido suas potentes
propriedades antioxidantes além de exercerem importantes funções em muitos processos que
previnem diabetes e disfunção erétil (CASSIDY et al., 2016). Além disso, os compostos fenólicos
(antocianinas e flavonoides entre outros) dão aos frutos qualidades desejáveis tais como cor e
propriedades antioxidantes (BRAVO et al., 1998).
A estrutura química dos flavonoides apresenta um anel denominado grupo flavilíco
(figura 3). A capacidade, ou seja, a propriedade dos flavonoides sequestrar espécies reativas de
oxigênio e nitrogênio é devida à presença de uma orto-hidroxilação no anel B da molécula de
flavonóide, do número de hidroxilas livres e uma dupla ligação na posição na posição C2-C3 ou a

57
presença de uma hidroxila no carbono C3 (BURDA; OLESZEK,2001). Mas, não se sabe
exatamente de que maneira os compostos fenólicos corroboram na atividade antioxidante, há
possibilidade que a mesma seja decorrente da ação sinergética entre diversos compostos fenólicos
e não advinda apenas de um único componente (ARNOUS et al., 2001; LEE et al., 2003)
Figura 3 - Estrutura do núcleo flavílico
Fonte: Próprio autor
Sabe-se, entretanto que a atividade antioxidante dos compostos fenólicos cresce com o
aumento no número de grupamentos hidroxila e diminuição de grupos glicosilados
(FUKUMOTO; MAZZA, 2000).
Antioxidantes fenólicos funcionam como sequestradores de radicais e algumas vezes
como quelantes de metais (AMES et al., 1993; RICE-EVANS et al., 1996).
2.5 ANTOCIANINAS
Para classificação dos flavonoides se utilizam critérios baseados no número e na posição
dos grupos hidroxila e metoxila no núcleo flavílico. Em consonância com estes dados são
encontradas predominantemente 6 tipos de antocianidinas: pelargonidina, cianidina, delfinidina,
petunidina, peonidina e malvidina (Fig.4).

58
Figura 4 – Antocianidinas encontradas em frutas
Fonte: Próprio autor.
As antocianinas fazem parte do grupo dos flavonoides e têm a mesma origem
biossintética, portanto compartilham características próprias dos mesmos como, por exemplo, o
esqueleto básico composto por C6-C3-C6. Diferem dos flavonoides por serem glicolisadas e
também pela coloração mais forte. Além disso, as antocianinas têm um poder maior de oxidação
(PIMENTEL et al., 2005; MALCHER, 2011). Quando as antocianidinas apresentam uma ou mais
unidades de açúcar anexadas ao núcleo flavílico são denominadas antocianinas.
Mais de 20 antocianinas já foram identificadas e dentre estas as seis que tem sido
utilizada na indústria e também com potencial farmacológico são a perlargonidina; cianidina;
delfinidina; pertunidina; malvidina e peonidina (GRISEBACH, 1982; MALCHER, 2011).
O que caracteriza a antocianina é o núcleo básico composto pela estrutura de íons 4-
hidroxiflavilum, portanto as antocianinas são compostas por três partes, aglicona (chamada de
antocianidina), um açúcar e um grupo acil nem sempre presente (AKWIE, 2000).

59
As antocianinas são reconhecidas por suas propriedades farmacológicas e medicinais,
bem como por ser anticarcinogênica, anti-inflamatória e antimicrobiana, prevenindo a oxidação
de proteínas de baixa densidade (LDL), o chamado colesterol ruim, enfermidades
cardiovasculares e doenças neurológicas (KUSKOSKI et al., 2006; BRAVO, 1998).
As antocianinas têm efeito antioxidante e atuam inibindo os efeitos desencadeados por
radicais livres. O combate aos processos oxidativos impede que ocorra danos ao DNA e às
macromoléculas, danos estes que com o tempo têm efeitos cumulativos que podem desencadear
doenças como o câncer, cardiopatias e catarata (SANTOS et al., 2008). O alto teor de
antocianinas com seu consequente poder antioxidante tem despertado interesse crescente no
estudo do açaí (Euterpe oleracea Mart.). Certamente este fato está relacionado com a
comprovação de que o fruto do açaí (Euterpe oleracea Mart.) apresenta efeito antimutagênico
(GALOTTA & BOAVENTURA, 2005). As polpas de açaí (Euterpe oleracea Mart.)
apresentaram elevados valor no parâmetro atividade antioxidante, sendo, portanto, consideradas
uma grande fonte de antioxidantes (SANTOS et al, 2008).
As principais antocianidinas encontradas nos alimentos são mostradas na figura 5.
Figura 5 – Princípais antocianidinas presentes nos alimentos
Fonte: Próprio Autor
As principais antocianidinas encontradas no açaí (Euterpe oleracea Mart.) foram
cianidina 3-glicocida, cianidina 3-sambubiocida cionidina 3-rutinocida e peonidina 3-rutinocida
(MULABAGAL et al., 2012).
A diferenciação das antocianinas pode ser feita através da aferição do número dos
grupos hidroxilas, do grau de metoxilação, do número de açúcares com suas ligações químicas e
finalmente através da identificação da natureza do grupo alifático (FRANCIS,1992; MALCHER,
2011).
Antocianidinas R1 R2
Perlagonidina H H
Cianidina OH H
Delfinidina OH OH
Peonidina OCH3 H
Petunidina OCH3 OH

60
A cor característica das antocianinas é consequência da presença do açúcar, de ácidos,
grupos metóxilo e de hidroxila que além da cor conferem estabilidade. Varia de vermelho a azul
com várias tonalidades A alteração da cor é um indício da degradação da molécula sendo a cor
marrom indesejada. Esta alteração constitui um indício de degradação e pode ser causada por
fatores como pH, concentração da solução, presença de copgmentação, luz e oxigênio.
Com relação ao açúcar presente nas antocianinas os mais frequentemente encontrados
são a glicose, xilose, arabinose, rammose e galactose (TERCI, 2004).
Concernente à estabilidade os açúcares conferem estabilidade às antocianinas, mas não é
tão importante a natureza do açúcar e sim sua posição na molécula que vai exercer significativa
influência na reatividade da antocianina (RIBEIRO; SERAVALLI, 2004), sendo mais estáveis
quando glicosadas na posição C-3 (JURDI, 1964).
As antocianinas são muito instáveis sendo que os principais fatores que afetam sua
estabilidade são alterações de pH e temperatura. O pH altera a estabilidade, na faixa de 1 a 3 as
antocianinas adquirem a cor vermelha e na faixa de 4 a 6,5 a cor é púrpura, uma característica
bem perceptível no açaí (Euterpe oleracea Mart.) cuja faixa de pH fica em tono de 4 a 5
(GRISEBACH, 1982).
Com relação à temperatura sabe-se que temperaturas elevadas degradam as antocianinas
(GRISEBACH, 1982). Temperaturas mais baixas retêm o pigmento cuja destruição se evidencia
na alteração da cor de vermelha para marrom. A cor também sofre alterações de acordo com o
processamento e estocagem do material (GRISEBACH, 1982).
Outro fator importante é a luminosidade e a presença de oxigênio que juntos causam
alteração da cor das antocianinas as tornando escuras (CARDOSO, 1997; RIBEIRO, 2004). Em
atmosfera inerte não há alteração da cor mesmo com exposição à luz.
Algumas enzimas como a glicosídeas, polifenoloxidases e peroxidases degradam as
antocianinas provocando escurecimento e perdas da cor. Isto ocorre durante o processamento do
suco de frutas (CARDOSO, 1997).
Para sua preservação as antocianinas não podem ser estocadas em recipiente de ferro, de
alumínio ou de latão, pois estes provocam alterações nas mesmas.
As antocianinas podem ser extraídas com etanol ou metanol, pois são solúveis em
solventes polares. Elas são encontradas predominantemente nas folhas e nos frutos

61
principalmente na casca dos frutos e esta extração deve ser feita em meio ácido objetivando desta
forma evitar a oxidação do pigmento.
As antocianinas nos frutos são localizadas principalmente na casca, porém também são
encontradas em menores quantidades nas polpas. Foram descobertos fatores reguladores que
promovem o acúmulo de antocianinas. Há proteínas que interagem com as proteínas reguladoras
das vias de síntese das antocianinas. Há uma interação entre fatores que acumulam antocianinas e
genes promotores de sua biossíntese (AN et al., 2012)
Pesquisas têm confirmado a eficácia do uso tradicional de antocianinas para combater
acidez estomacal (ALVAREZ-SUAREZ et al., 2011) e combate de inflamação (BASTOS et al.,
2009; ALMEIDA et al., 2014). De acordo com Almeida et al. (2014) estas moléculas são
promissoras moduladoras de inflamação. Têm sido confirmadas mais propriedades dantes
desconhecidas como anticancerígeno, prevenção de oxidação proteínas de baixa densidade (LDL)
combatendo o colesterol ruim, enfermidades cardiovasculares e doenças neurológicas
(KUSKOSKI et al., 2006; BRAVO, 1998). Recentemente tem sido confirmado que antocianinas
podem evitar a elevação da glicemia do sangue, portanto podendo ser útil para tratamento e
prevenção de diabetes (ALZAID et al., 2013). Sabe-se que os polifenóis interagem diretamente
com o transporte da glicose regulando a taxa de absorção de glicose. Este mecanismo foi
confirmado além de se confirmar que os flavonoides podem modular a glicemia através de
decréscimo da expressão gênica relacionada com o transporte de glicose (ALZAID et al., 2013).
2.6. O GÊNERO EUTERPE
Há três espécies do gênero Euterpe conhecidas no Brasil. Entre estas há o palmito
(Euterpe edulis), encontrado na mata atlântica, tem sido levado à beira da extinção devido a
redução da floresta atlântica e sua exploração de maneira insustentável. No médio rio Solimões
encontramos a espécie Euterpe precatoria e em toda a Amazônia em grandes populações, nas
bordas do rio ou em áreas periodicamente inundadas encontramos o E. oleracea (PESSOA et al.,
2010).

62
2.6.1 O açaizeiro (Euterpe oleracea Mart.)
O açaizeiro (Euterpe oleracea Mart.) é encontrado em várzeas ao longo de rios e
igarapés, nas baixadas e áreas úmidas, por toda extensão da Amazônia (MALCHER, 2011). A
palmeira do açaí (Euterpe oleracea Mart.) fornece os frutos e o palmito cuja exploração resulta
em uma economia que mantém tanto a população como contribui para a sustentabilidade da
floresta causando impacto altamente positivo na economia local (QUEIROZ; MOCHIUTTI,
2001). Tendo como centro de origem, portanto o estuário do Amazonas o açaizeiro (Euterpe
oleraccea Mart.) é uma espécie frutífera com bastante diversidade genética. Além de ser
encontrado em toda região norte do Brasil também é comum em outros países que compõem a
Amazônia, quais sejam Venezuela, Colômbia, Equador, Suriname e Guiana e também é
encontrado na América Central (Panamá) (OLIVEIRA; FARIAS NETO, 2006; NOGUEIRA et
al., 2006).
Sabe-se que o açaizeiro floresce e frutifica durante o ano todo com dois períodos de pico
bem distintos fornecendo duas safras, sendo o primeiro de janeiro a maio e o segundo de
setembro a dezembro (CALZAVARA, 1972; OLIVEIRA; FERNANDES, 1993, MALCHER,
2011).
O que caracteriza o açaizeiro é o fato desta palmeira apresentar grande número de
estirpes (de 4 até 25) e assim uma touceira composta de perfilhos. Pode atingir até 35 metros de
altura e o tronco mede de 8 a 25 cm de diâmetro. A copa do açaizeiro é formada por 8 até 15
folhas de até 2 metros de comprimento com cor verde oliva, composta por 40 a 80 folíolos
(DONADIO et al., 2002).
O nativo da Amazônia costumava obter o extrato do açaí (Euterpe oleracea Mart.) com
o uso de peneiras e até das mãos. Há alguns anos advieram as batedeiras que são máquinas
fabricadas com esta função específica. O alto conteúdo de fibras obriga o uso de muita água
obtendo ainda assim uma polpa muito viscosa e muito apreciada pela população. Sendo
consumido com carne assada e farinha constitui em muitos lugares a principal refeição do dia.
Muitos estudos sobre ação benéfica das frutas tiveram intenso aumento objetivando
analisar sua ação antioxidante (SCHAUSS et al., 2006). O açaí (Euterpe oleracea Mart.) teve um
aumento de consumo tanto nacional como internacional o que tem induzido estudos no sentido de

63
se conhecer suas propriedades nutritivas e fotoquímicas (MALCHER, 2011). O açaí (Euterpe
oleracea Mart.) é rico em compostos fenólicos além de antocianinas (PEIXOTO et al., 2016).
As florestas de açaí (Euterpe oleracea Mart.) são abundantes por toda a planície
inundadas da Amazônia, pois é nativo nesta região. Pode ser extraído de maneira sustentável
Os frutos do açaí (Euterpe oleracea Mart.) são arredondados com diâmetro oscilando
entre 1 e 2 cm. Seu pericarpo é de aproximadamente 1 mm de cor pardo-violácea, contendo uma
polpa oleaginosa e comestível, a semente possui o endocarpo duro e fibroso (SANTOS et al.,
2008; PESSOA et al., 2010). Esse fruto arredondado do açaí (Euterpe oleracea Mart.) apresenta
o mesocarpo formado por epiderme, parênquima externo, esclerênquima externo, parênquima
armazenador e parênquima interno. A epiderme e o parênquima externo ou parênquima
antocianínico são ricos em antocianinas. O parênquima armazenador é rico em lipídeos e o
interno comporta os sistemas monosteles e vascular (PESSOA et al., 2010).
Devido ao seu elevado teor de antocianinas o açaí (Euterpe oleracea Mart.) pode ser
considerado um alimento nutracêutico e são justamente estas mesmas antocianinas que atribuem
ao açaí (Euterpe oleracea Mart.) cor avermelhada ou roxa (IADEROZA et al., 1992; OZELA et
al., 1997). A figura 6 mostra o mapa conceitual com as propriedades do açaí, seus principais
compostos bioativos e o direcionamento para incorporá-los em biofilmes de kefir.
Genero EuterpeTres especies
EmulsaopH
Propriedades reológicas
Radicais livresEstresse oxidativo
Antioxidante
BIOFILMESAntocianinas
Compostos Fenólicos
PROPRIEDADES
Flavonóides
Euterpe oleraceaAcai
Kefir
Figura 6- Mapa conceitual apresentando as propriedades e os principais compostos bioativos do extrato de açaí com
a incorporaração de antocianinas do açaí (Euterpe oleracea Mart.) em biofilme de kefir

64
2.6.1.1 Classificação de extrato de açaí (Euterpe oleracea Mart.) de acordo com o pH
De acordo com o pH, os alimentos são classificados como de baixa acidez (pH > 4,50),
ácidos (pH de 4,00 a 4,50) e muito ácidos (pH < 4,00). Essa classificação se baseia no pH
mínimo para a multiplicação e produção de toxina do Clostridium botulinum (pH = 4,5) e no pH
mínimo para a multiplicação da grande maioria das bactérias (pH = 4,00). O pH do suco obtido
com a polpa do açaí (Euterpe oleracea Mart.) é em torno de 4,5 (SANTOS et al., 2008),
As amostras de kefir em açaí (Euterpe oleracea Mart.) são considerados de alta acidez,
estando, portanto, fora da faixa de risco para a multiplicação e produção da toxina do C.
botulinum.
2.6.1.2 Açaí (Euterpe oleracea Mart.) e fotoproteção
Recentemente mais consumidores estão procurando por produtos que contenham
compostos naturais que absorvam as ondas UV devido aos benefícios que tais produtos
comportam. Dentre estes produtos tem se descoberto que extratos de óleos vegetais podem conter
a absorção UV além de fornecer fotoproteção. Há um interesse crescente pela alta capacidade
antioxidante do açaí (Euterpe oleracea Mart.) a qual é consequência da alta quantidade de
compostos polifenólicos presentes no mesmo. Convém ressaltar que as antocianinas são
predominantemente polifenóis, mas há outros compostos que não são antocianinas no açaí
(Euterpe oleracea Mart.) que são também polifenóis e ácidos fenólicos também presentes no
extrato de açaí (Euterpe oleracea Mart.). Mais de 90% do total de compostos fenólicos do açaí
(Euterpe oleracea Mart.) são antocianinas. As antocianinas pertencem à classe dos polifenóis e
podem servir como antioxidante causando a fotoproteção através da eliminação de espécies
reativas do oxigênio durante o estresse fotoxidativo (FERRARI & ROCHA-FILHO, 2011).
2.6.1.3 Componentes do extrato de açaí (Euterpe oleracea MART.)
O extrato de açaí (Euterpe oleracea Mart.) é riquíssimo em antocianinas. Fonte de
lipídeos, proteínas, fibra e minerais como Mn, Cu e B bem como vitaminas. O elevado valor

65
energético do açaí (Euterpe oleracea Mart.) é devido a sua riqueza em lipídeos. Os seus
compostos fenólicos explicam sua capacidade antioxidante (SANTOS et al., 2008) e tem em sua
composição propriedades energéticas, plásticas, minerais, essenciais para nosso metabolismo
(MALCHER, 2011).
Além disto, outro parâmetro importante com relação ao açaí (Euterpe oleracea Mart.) é
obtido através da identificação e quantificação das antocianinas responsáveis pelas propriedades
fotoquímicas do açaí (Euterpe oleracea Mart.) (GALORRI et al., 2004).
O extrato de açaí (EHEEo) apresenta propriedades nutricionais importantes além de se
tratar de um fruto com alto valor energético (cerca de 247 kcal/100g de polpa). Para FREIRE et
al. (2000) o valor calórico elevado do açaí (Euterpe oleracea Mart.) é devido a grande
quantidade de lipídeos sobretudo ácidos graxos como óleo palmítico e linoleico semelhante à
composição do óleo de oliva (ROGEZ, 2000). O açaí (Euterpe oleracea Mart.) é rico em fibra
alimentar insolúvel.
A polpa de açaí (Euterpe oleracea Mart.) é considerada como um alimento nutracêutico,
devido ao seu elevado teor de antocianinas, responsáveis pela cor avermelhada do fruto
(IADEROZA et al., 1992; OZELA et al., 1997).
2.7 ISOFLAVONAS PRESENTES EM GÉRMEN DE SOJA (Glycine max (L.) Merril)
As isoflavonas compreendem as agliconas daidzeína, genisteína e gliciteína, os
respectivos b-glicosídios e os conjugados malonil-glicosídios e acetil-glicosídios. As isoflavonas
são compostos fenólicos bioativos cujo consumo pode reduzir os riscos de doenças
cardiovasculares, de alguns tipos de câncer e de diabetes (BARNES et al., 1999; MANDARINO,
2010; LAUDANNA, 2006; IMAI, 2015). Dentre as isoflavonas, especificamente a genisteína e a
daidzeína desempenham um papel importante na prevenção de doenças crônicas, tais como,
osteoporose, doenças do coração, câncer e diabetes (ESTEVES; MONTEIRO, 2001)
As isoflavonas também reduzem risco de osteoporose, pois têm atividade
antirreabsorção do osso além de exercerem efeitos sobre a diferenciação e proliferação dos
osteoblastos (YU et al., 2015). Estudos epidemiológicos ratificam que o consumo de produtos
derivados da soja reduz efetivamente o risco de câncer dos seios e de próstata (VARINSKA et
al., 2015).

66
Recentemente foi confirmado que a genisteína pode suprimir o crescimento de células
cancerígenas. A genisteína tanto inibiu como induziu a fragmentação do DNA de células
cancerígenas do sarcoma do útero (YEH et al., 2015), ela é inibidora de várias vias que
desenvolvem o processo da carcinogenese (VARINSKA et al., 2015). Convém ressaltar que a
genisteína é também uma potente inibidora da angiogênese (VARINSKA et al., 2015) cujo
descontrole é um dos passos fundamentais que conduzem o crescimento do câncer, sua invasão e
metástase.
Além de exercerem um potente efeito antioxidante as isoflavonas podem exercer no
organismo humano efeito hormonal (IMAI, 2015). São fitoestrógenos ativos com reconhecida
capacidade hipocolesterolêmica (ROSSI et al., 2004). Pessoas obesas contêm no sangue
marcadores de inflamação, citocinas pró-inflamatórias e mediadores de inflamação. As plantas
desenvolveram ao logo de seu processo evolucionário compostos que podem ser utilizados no
combate à inflamação e infecções, dentre eles, as isoflavonas. O consumo da soja desempenha
um papel na modulação de marcadores de inflamação através dos seus compostos bioativos,
isoflavonas, saponinas e flavonoides (TEZUKA; IMAI, 2015).
Dentre todos os grãos já estudados, o grão de soja é o que possui os maiores teores de
isoflavonas, as quais estão relacionadas a fatores genéticos e ambientais. O teor de isoflavonas
em cultivares de soja realizado no Brasil variam entre 12 e 461 mg.100 g –1, de acordo com a
relação da formação de isoflavonas no grão de soja com efeitos genéticos e com o clima da região
em que a soja foi cultivada (CARRÃO-PANIZZI et al., 2009a).
Os teores de isoflavonas na soja também variam conforme a parte morfológica da qual
são extraídas (cotilédone, hipocótilo e casca). Estudos mostraram que a maior concentração de
isoflavonas se encontra no hipocótilo, com valores de 10 a 20 vezes maiores do que nos grãos
inteiros (TSUKAMOTO et al., 1995). Kudou et al. (1991) observaram que o hipocótilo apresenta
cinco a seis vezes mais isoflavonas que os cotilédones, sendo que a glicitina e seus derivados
somente ocorrem no hipocótilo (42,84% do total de isoflavonas deveu-se à presença das formas
glicitinas). Também são encontradas isoflavonas na raiz da soja e são justamente as agliconas
daidzeína e genisteína as responsáveis pela atração de bactérias nitrificadoras à raiz. Na ausência
de nitrogênio há um aumento na produção dessas isoflavonas (SUGIYAMA et al., 2015).
Além do fator genético a concentração de isoflavonas nos grãos de soja é fortemente
relacionada por condições do ambiente sendo um dos mais relevantes a temperatura

67
especialmente durante o desenvolvimento dos grãos., Quando a soja é cultivada em regiões cujas
temperaturas oscilam em torno de 20oC, a concentração média de isoflavonas é de 147,8mg/100g
(FT-Abyara) e 180,1mg/100g (IAS 5), Já quando a soja é cultivada em regiões mais quente por
exemplo com temperatura média de 25oC as isoflavonas apresentam de 73,5mg/100g até
85,5mg/100g dependendo da espécie (CARRÃO-PANIZZI et al., 2002).
Processos fermentativos podem alterar os níveis e formas das isoflavonas no produto
final, reduzindo em até 13 vezes a quantidade de isoflavonas (ROSSI et al., 2004). A ação
hidrolítica da -glicosidase própria de determinadas bactérias láticas pode fazer com que as
agliconas (genisteína e daidzeína) fiquem em níveis mais altos (MATSUDA et al., 1992; WANG;
MURPHY, 1994).
Rossi et al. (2004) constataram que o processo fermentativo de fabricação de iogurte com
leite de soja causou uma degradação isoflavonas fazendo que o produto final apresentasse uma
concentração 13 vezes menor do que no grão “in natura”.
A concentração dessas formas depende do processamento da soja, pois ocorre a conversão
de uma forma em outra decorrente do tratamento recebido para obtenção de seus variados
produtos. Ocorre também a perda das isoflavonas no processo, por exemplo, quando submetida
ao cozimento às perdas chegam a 50% (WANG, 1995), pois a solubilidade das isoflavonas
aumenta com a temperatura. Nos processos em que a soja permanece de molho em água, em
temperatura ambiente por tempos prolongados a perda das isoflavonas fica em torno de 11%
(WANG, 1995).
Fatores como a variedade do grão, a região e época de cultivo, as condições climáticas e,
inclusive, as condições de processamento alteram a concentração. Além disso, o tempo e
temperaturas empregados nos tratamentos térmicos podem alterar tanto as concentrações quanto
às formas de isoflavonas na soja (ELDRIDGE et al., 1983; PARK et al., 2002).
Ainda Rossi et al. (2004) constataram que após a fermentação com Enterococcus faecium
e Lactobacillus jugurti as formas agliconas apareceram em concentrações bem menores. O sabor
amargo e adstringente normalmente verificados nos derivados de soja que causa uma menor
aceitação da soja e de seus produtos advém justamente destas agliconas (ARAÜJO et al., 1997;
AUSSENAC et al., 1998).

68
REFERÊNCIAS
ABRAHAM, A. G.; RIMADA, P. S. Kefiran improves rheological properties of glucono-δ-
lactone induced skim milk gels. International Dairy Journal. v. 16, p. 33-39, 2006.
ADACHI, S.; ITOH, T.; TOBA, T.; ARIHARA, K.; MUKAI, T. Ecology of lactic acid bacteria
with special reference to kefir-granule formation by Lactobacillus kefiranofaciens. Biseibutsu, v.
6, p.15-25, 1990.
AHIMOU, F.; SEMMENS, M. J.; HAUGSTAD, G.; NOVAK, P. J. Effect of protein,
polysaccharide, and oxygen concentration profiles on biofilm cohesiveness. Appl. Environ.
Microbiol, v. 73, p. 2905-2910, 2007.
AKWIE, S. N. L. T. Cinética de Amadurecimento, Extração e Estabilidade das Antocianinas
do Açaí. 2000. Dissertação (Mestrado em Engenharia química) - Universidade Federal do Pará,
Belém.
ALMEIDA M.C.; OYAMA L. M.; OLIVEIRA J. L.; CARVALHO G. M.; ROSSO V. V.;
SOUSA M. A. L.; NASCIMENTO C. M.; PISANI, L. P. 2014. Jussara (Euterpe edulis Mart.)
supplementation during pregnancy and lactation modulates the gene and protein expression of
inflammation biomarkers induced by trans-fatty acids in the colon of offspring. Mediators
Inflamm., v. 2014, Article ID 987927, 11 pages
ALVAREZ-SUAREZ J. M.; DEKANSKI D.; RISTIĆ S.; RADONJIĆ, N. V.; PETRONIJEVIĆ
N. D.; GIAMPIERI, F.; ASTOLFI P.; GONZÁLEZ-PARAMÁS, A. M.; SANTOS-BUELGA C.;
TULIPANI S.; QUILES J. L. ; MEZZETTI B; BATTINO M. Strawberry polyphenols attenuate
ethanol-induced gastric lesions in rats by activation of antioxidant enzymes and attenuation of
MDA increase. PLoS One, v. 6, n. 10, p. 258-278, 2011.
ALVES, A. P. C.; CORRÊA, A. D.; MARQUES, T. R.; RAMOS, V. O.; SIMÃO, A. A.; LAGE,
F. F. Compostos Fenólicos e Potencial Antioxidante de Diferentes Extratos de Farinha de
Cascas de Jabuticaba. 52° Congresso Brasileiro de Química 14-18/10/2012 Recife, Pe
ALZAID, F.; CHEUNG, H. M.; PREEDY, V. R; SHARP, P. A. Regulation of glucose
transporter expression in human intestinal Caco-2 cells following exposure to an anthocyanin-
rich berry extract PLoS One, v. 8, n. 11, p. 78932, 2013.
AMES, B. N; SHIGENAGA, M. K.; HAGEN, T. M. Oxidants, Antioxidants and the
degenerative diseases of aging. Proceedings of the National Academy of Sciences., v. 90, n.17,
p.7915-7922, 1993.

69
AN, X. H.; TIAN, Y. ; CHEN, K. Q.; WANG, X. F.; HAO, Y. J. The apple WD40 protein
MdTTG1 interacts with bHLH but not MYB proteins to regulate anthocyanin accumulation. J
Plant Physiol., v. 169, n. 7, p. 710-717, 2012.
ANGULO, L.; LOPEZ, E.; LEMA, C. Microflora present in kefir grains of the Galician Region
(North-West of Spain). Journal of Dairy Research, v. 60, p. 263-267, 1993.
ANSELMO, R. J.; VIORA, S. S.; OJEDA, P. A.; LAUSADA, L. I., Efecto Antagónico del Kefir
sobre Endosporas y Células Vegetativas de Bacillus Cereus y Clostridium Perfringe., Inf.
tecnol., v. 21 n. 4, p. 131-138, 2010.
ARAÜJO, J. M. A.; CARLOS, J. C. S.; SEDYAMA, C. S. Isoflavonas em grãos de soja:
importância da atividade de B-glicosidase na formação do sabor amargo e
adstringente. Ciênc.Tecnol.Alim., v. 17, p. 137-141, 1997.
ARENA, A.; MAUGERI, T.L.; PAVONE, B.; IANNELLO D.; GUGLIANDOLO, C.;
BISIGNANO, G. An exopolysaccharide produced by Geobacillus thermodenitrificans strain B3-
72: Antiviral activity on immunocompetent cells. International Immunopharmacol. v. 6, p. 8–
13, 2006.
ARNOUS, A.; MAKRIS, D. P.; KEFALAS, P. Effect of principal polyphenolic components in
relation to antioxidant characteristics of aged red wines. Journal of Agricultural and Food
Chemistry., v. 49, p. 5736-5742, 2001.
ASHFORD, M.; FELL, J. T.; ATTWOOD, D.; SHARMA, H.; WOODHEAD, P. J. Studies on
pectin formulations for colonic drug delivery. J. Control. Release, v. 30, p. 225-232, 1994
AUSSENAC, T. et al. Quantification of isoflavones bycapillary zone electrophoresis in soybean
seeds: effects of variety and environment. Am.J. Clin. Nutr., v. 68, p. 1 480-1 485, 1998.
BAKER, A. A.; HELBERT, W.; SUGIYAMA, J.; MILES M. J. High-Resolution Atomic Force
Microscopy of Native Valonia Cellulose in Microcrystals, Struct Biol., v. 119, n. 2, p. 129-38,
1997.
BARBOSA, K. B. F. Estresse oxidativo: conceito, implicações e fatores modulatórios. Rev.
Nutr., v. 23, n. 4, p. 629-643, 2010.

70
BARNES, S.; KIM, H.; XU, J. Soy in the prevention and treatament of chronic diseases. In:
CONGRESSO BRASILEIRO DE SOJA, 1999, Londrina. Anais... Londrina: Embrapa CNPSo,
1999, p. 295-310. (Embrapa CNPSo. Documentos, n. 124).
BASTOS, D. H. M.; ROGERO, M. M.; ARÊAS, J. A. G. Mecanismo de ação de compostos
bioativos dos alimentos no contexto de processos inflamatórios relacionados à obesidade. Arq.
Bras. Endocrionol. Metab., v. 53, n. 5, p. 646-656, 2009.
BELITZ, H. D.; GROSCH, W. Aroma Substances. In: BELITZ, H-D.; GROSCH, W. (Eds) Food
Chemistry. Berlin; Springer-Verlag, Cap.5, p.319-377. 1999.
BENTLEY, W. E.; HOOSHANGI, S. From unicelular properties to multicelular behavior:
bacteria quorum sensing circuitry and applications. Curr Opin Biotechnol., v. 19 p. 550-555,
2008.
BESHKOVA, D. M.; SIMOVA, E. D.; FRENGOVA, G. I., SIMOV, Z. I.; DIMITROV, Z. P.
Production of volatile aroma compounds by kefir starter cultures. International Dairy Journal,
v. 13, p. 529-535, 2003.
BIRKENHAUER, E.; NEETHIRAJAN, S.; WEESE, J. S. Collagen and hyaluronan at wound
sites influence early polymicrobial biofilm adhesive events. BMC Microbiol., v. 14, p. 191,
2014.
BOLLA, P. A.; CARASI, P.; BOLLA, Mde. L.; DE ANTONI, G. L.; SERRADELL, Mde. L.
Protective effect of a mixture of kefir-isolated lactic acid bacteria and yeasts in a hamster model
of Clostridium difficile infection. Anaerobe, v. 21, p. 28- 33, 2013.
BOURRIE, B. C.; WILLING, B. P.; COTTER, P. D. The Microbiota and Health Promoting
Characteristics of the Fermented Beverage Kefir. Front Microbiol, v. 7, 647, 2016.
BOYER, J.; LIU, R. H. Apple phytochemicals and their health benefits. Nutrition Journal, v. 3,
n. 5, p. 1-15, 2004
BRAVO I. Polyphenols: Chemistry, dietary source, metabolism, and nutritional significance.
Nutr. Rev., v. 56, p. 317–333, 1998.
BRENNER, K.; YOU, L.; ARNOLD, F. H. Engineering microbial consortia: a new frontier in
synthetic biology, Trends Biotechnol. v. 26, p. 483 – 489, 2008.

71
BURDA, S.; OLESZEK, W. Antioxidant and antiradical activities of flavonoids. Journal of
Agricultural and Food Chemistry, v. 49, p. 2774-2779, 2001.
CALAZANS, G. M. T.; LOPES, C. E.; LIMA, R. M. O. C.; FRANÇA, F. P. Antitumuor
activities of levans produced by Zymomonas mobilis strains. Biotechnol. Lett., v. 19, p. 19-21,
1997.
CALZAVARA, B. B. G. As possibilidades do açaizeiro no estuário amazônico. Boletim da
Faculdade de Ciências Agrárias do Pará. Belém: FCAP, 1972. 103p. Campinas,SP. v. 15,n. 1, p.
71-81.jan./abr. 2002.
CARDOSO, S. M. Obtenção de antocianinas a partir do repolho vermelho (Brassica
oleracea Var. Capitata). Centro de Ciências da Saúde, UFPA, Belém, 1997, 24p
CARRÃO-PANIZZI, M. C.; BERHOW, M.; MANDARINO, J. M. G.; OLIVEIRA, M. C. N.
Enviromental and genetic variation of isoflvones content of soybean seeds grown in Brazil.
Pesquisa Agropecuária Brasileira, v. 44, n. 11, p. 1444-1451, 2009
CASSIDY, A.; FRANZ, M.; RIMM, E. B. Dietary flavonoid intake and incidence of erectile
dysfunction. Am J Clin Nutr, v. 103, n. 2, p. 534-541, 2016.
CHEIRSILP, B., SHIMIZU, H., SHIOYA, S. Modelling and optimization of environmental
conditions for kefiran production by Lactobacillus kefiranofaciens. Appl. Microbiol.Biotechnol.
v. 57, p. 639-646, 2001.
CHOWDHURY, S. R.; BASAKA, R. K.; SENB, R; ADHIKARI, B. Characterization and
emulsifying property of a carbohydrate polymer produced by Bacillus pumilus UW-02 isolated
from waste water irrigated agricultural soil. International Journal of Biological
Macromolecules, v. 48, p. 705–712, 2011.
COPPA, G. V.; ZAMPINI, L.; GALEAZZI, T,; FACINELLI B.; FERRANTE, L.; CAPRETTI,
R.; ORAZIO, G. Human milk oligosaccharides inhibit the adhesion to Caco-2 cells of diarrheal
pathogens: Escherichia coli, Vibrio cholerae, and Salmonella fyris. Pediatr Res, v. 59, n. 3, p.
377-382, 2006.
COSTA, P., LOBO, J. M. S. Formas farmacêuticas de liberação modificada. Rev. Port. Farm.,
v. 59, n. 4, p. 181-190, 1999.
COSTERTON, J. W.; STEWART, S.; GREENBERG, E. P. Bacterial biofilms: A common cause
of persistent infections. Science, v. 284, p. 1318-1322, 1999.

72
COSTERTON, W.; VEEH, R.; SHIRTLIFF, M.; PASMORE, M.; POST, C.; EHRLICH, G. The
application of biofilm science to the study and control of chronic bacterial infections. J. Clin.
Invest., v.112, p. 1466-1477, 2003.
CROZIER, A.; JAGANATH, I. B.; CLIFFORD, M. N. Dietary phenolics: chemistry,
bioavailability and effects on health. Nat Prod Rep, v. 26, n. 8, p. 1001-1043, 2009.
CUQ, B.; AYMARD, C.; CUQ, J. L.; S. GUILBERT. Edible Packaging Films Based on Fish
Myofibrillar Proteins: Formulation and Functional Properties. Journal Food Science, v. 60, p.
1369–1374, 1995.
CEVIKBAS, A., E. YEMNI, E.; EZZEDENN, F. W.; YARDIMICI, T. Antitumoural,
antibacterial and antifungal activities of kefir and kefir grain. Phytother. Res., v. 8, n. 78-82.,
1994.
CHIORCEA PAQUIM, A. M.; DICULESCU, V. C.; ORETSKAYA, T. S.; OLIVEIRA BRETT.
A. M. AFM and electroanalytical studies of synthetic oligonucleotide hybridization. Biosens
Bioelectron. v. 20, n. 5, p. 933-44, 2004.
DAVEY, M. E., O'TOOLE, G. A. Microbial biofilms: from ecology to molecular genetics.
Microbiology and Molecular Biology Reviews, v. 64, n. 847, 2000.
DE VUYST, L.; DE VIN, F.; VANINGELGEM, F.; DEGEEST, B. Recent developments in the
biosynthesis and applications of heteropolysaccharides from lactic acid bacteria. International
Dairy Journal., v.11, p. 687–707, 2001.
DE WILDE, T.; SPANOGHE, P.; DEBAER, C.; RYCKEBOER, J.; SPRINGAEL, D.; JAEKEN,
P. Overview of on-farm bioremediation systems to reduce the occurrence of point source
contamination. Pest Management Science, v. 63, p. 111-128, 2007.
DECKER, E. A. Phenolics: prooxidants or antioxidants? Nutr Rev. v. 55, p. 396-398, 1997.
DEGÁSPARI, C. H.; WASZCZYNSKYJ, N. Antioxidants Properties of Phenolic Compounds.
Revista Visão Acadêmica, v. 5, n. 1, 2004.
DEGORCE-DUMAS, R.; GOURSAUD, J.; LEVEAU, J. Y. Analyse des composes volatils du
yaourt par chromatographie en phase gazeuse-espace de tete (headspace). Industries
Alimentaires et Agricoles, v. 9, p. 805–808, 1986.
DONADIO, L.; C.; MÔRO, F. V.; SERVIDONE, A. A. Frutas brasileiras. Jaboticabal, 288p.
2002.
DONLAN, R. M. Biofilms: Microbial life on surfaces. Emerging Infect. Dis. v. 8, p. 881-890,
2002.
DORNELLES, A. S.; RODRIGUES, S. Fermentação Alcoólica de Caldo de Cana utilizando
grãos de Kefir. Ciência Agronômica, v. 37, n. 3, p. 386-390, 2006.

73
DRESSMAN, J. B., AMIDON, G. L., REPPAS, C., SHAH, V. P. Dissolution testing as a
prognostic tool for oral drug absorption: immediate release dosage forms. Pharm. Res., v. 15, n.
1, p. 11-22, 1998.
DUBOC, P.; MOLLET, B. Applications of exopolysaccharides. International Dairy Journal, v.
11, p. 759–768, 2001.
DUFRÊNE, Y. F. Sticky microbes: forces in microbial cell adhesion Trends Microbiol, v. 23, n.
6, p. 376-382, 2015.
DUITSCHAEVER, C. L.; KEMP, N.; EMMONS, D. Pure culture formulation and procedure for
the production of kefir. Milchwissenschaft, v. 42, p. 80-82, 1987.
ELDRIDGE, A. C.; KWOLEK, W. F. Soybean isoflavones: effects of environment and variety
on composition. J. Agric. Food Chem., v. 31, p. 394-396, 1983.
ESTEVES, E. A.; MONTEIRO, J. B. R. Efeitos benéficos das isoflavonas de soja em doencas
crônicas Rev. Nutr., v. 14, n. 1, p. 43-52, 2001.
FARNWORTH, E. R. Kefir – a complex probiotic. Food Science and Technology Bulletin:
Functional Foods, v. 2, p. 1-17, 2005.
FEDORAK, R. N.; MADSEN, K. L. Probiotics and prebiotics in gastrointestinal disorders.
Current Opinion Gastroenterology, v. 20, p. 146-155, 2004.
FERNÁNDEZ, C. E.; GIACAMAN, R. A.; TENUTA, L. M.; CURY, J. A. Effect of the
Probiotic Lactobacillus rhamnosus LB21 on the Cariogenicity of Streptococcus mutans UA159 in
a Dual-Species Biofilm Model. Caries Res., v. 49 n. 6, p. 583-590, 2015.
FERNANDEZ-GARCIA, E.; MCGREGOR, J. U. Determination of organic acids during the
fermentation and cold storage of yogurt. Journal of Dairy Science, v. 77, p. 2934–2939, 1994.
FERRARI, M.; ROCHA-FILHO, P. A. Multiple emulsions containing amazon oil: açaí oil
(Euterpe oleracea). Revista Brasileira de Farmacognosia, v. 21, n. 4, p. 737-743, 2011.
FERREIRA, A. L. A., MATSUBARA, L. S. Radicais livres: conceitos, doenças relacionadas,
sistema de defesa e estresse oxidativo Rev. Assoc. Med. Bras., v. 43 n. 1, p. 61-68, 1997.
FERREIRA, A. A.; TETTE, P. A. S.; MENDONÇA, R. C. S.; SOARES, A. S.; CARVALHO,
M. M. Detection of exopolysaccharide production and biofilm-related genes in Staphylococcus
spp. isolated from a poultry processing plant. Food Sci. Technol, v. 34, n. 4, p. 710-716, 2014.
FERREIRA, N. O.; ANDRUCIOLI, M. C.; NELSON-FILHO, P.; ZANELLA, E. P.;
CONSOLARO, A.; ROMANO, F. L.; MATSUMOTO, M. A. Bacterial biofilm on successful and
failed orthodontic mini-implants-a scanning electron microscopy study. Microsc. Res. Tech., v.
78, n. 12, p. 1112-1116, 2015.
FIALHO, A.M.; MOREIRA, L.M.; GRANJA, A.T.; POPESCU, A.O.; HOFFMANN, K.; SÁ-
CORREIA, I. Occurrence, production, and applications of gellan: current state and perspectives.
Appl. Microbiol. Biotechnol, v. 79, n. 6, p. 889-900, 2008.

74
FLEMMING, H. C.; WINGENDER, J. Relevance of microbial extracellular polymeric
substances (EPSs) – Part I: Structural and ecological aspects. Water Sci. Technol., v. 43, p.1-8,
2001.
FLEMMING, H. C.; WINGENDER, J. Relevance of microbial extracellular polymeric
substances (EPSs) – Part II: Technical aspects. Water Sci. Technol. v. 43, p. 9-16, 2001.
FLEMMING, H. C. Biofilms and environmental protection. Water Science and Technology, v.
27, p. 1-10, 1993.
FONTÁN, M. C. G.; MARTÍNEZ, S.; FRANCO, I.; CARBALHO, J. Microbiological and
chemical changes during the manufacture of kefir made from cows’ milk, using a commercial
starter culture. International Dairy Journal, v. 16, p. 762-767, 2006.
FRANCIS, F. J. Analysis of anthocyanins. In: MARKAKIS, P. (ed). Anthocyanins as food
colors. New York: Academic Press, 1982, p. 181-207
FREIRE, A. C.; PODCZECK, F.; SOUSA, J.; VEIGA, F. Liberação específica de fármacos no
cólon por via oral. II - Tipos de sistemas utilizados. Revista Brasileira de Ciências
Farmacêuticas v. 42, n. 3, 2006.
FRENGOVA, G. I.; SIMOVA, E. D.; BESHKOVA, D. M.; SIMOV, Z. I. Exopolysaccharides
produced by lactic acid bacteria of kefir grains. Zeitschrift für Naturforschung v. 57, p. 805–
810, 2002.
FREY-KLETT, P.; BURLINSON, P.; DEVEAU, A.; BARRET, M.; TARKKA, M.;
SARNIGUET, A. Bacterial-fungal interactions: hyphens between agricultural, clinical,
environmental, and food microbiologists. Microbiol Mol Biol v.75, p. 583–609, 2011.
FUJISAWA, T.; ADACHI, S.; TOBA, T.; ARIHARA, K.; MITSUOKA, T. Lactobacillus
kefirarnofaciens sp. nov., isolated from kefir grains. Int. J. Syst. Bacterial., v. 38, p. 12-14,
1988.
FUKUMOTO, L. R.; MAZZA, G. Assessing Antioxidant and Prooxidant Activities of Phenolic
Compounds. J. Agric. Food Chem. v. 48, n. 8, p. 3587-3604, 2000.
FURIGA, A.; ROQUES, C.; BADET, C. Preventive effects of an original combination of grape
seed polyphenols with amine fluoride on dental biofilm formation and oxidative damage by oral
bacteria. J. Appl. Microbiol, v. 116, p. 761–771, 2014.
FURUKAWA, N.; MATSUOKA, A.; TAKAHASHI, T.; YAMANAKA, Y. Effects of fermented
milk on the delayed-type hypersensitivity response and survival day in mice bearing Meth-A.
Anim. Sci. Tec., v. 62, p. 579-585, 1991.
GALATI, G.; O’ BRIEN, P. J. Potenctial toxicity of flavonoids and other dietary phenolics:
significance for their chemo preventive and anticancer properties. Free Radical Biology &
Medicine, v. 37, n. 3, p.287-303, 2004.

75
GALLORI, S.; BILIA, A. R..; BERGONZI, M. C.; BARBOSA,W. L. R..; VINCIERI, F. F.
Polyphenolic constituints of fruit pulp of Euterpe oleracea Mart. Chomatographia, v. 59, p.
739-743, 2004.
GALOTTA, A. L.; Q. A.; BOAVENTURA, M. A. D. Quim. Nova, v. 28, p. 610, 2005.
GARCÍA-LAFUENTE, A.; GUILLAMÓN, E.; VILLARES, A.; ROSTAGNO, M. A.;
MARTÍNEZ, J. A. Flavonoids as anti-inflammatory agents: implications in cancer and
cardiovascular disease. Inflamm Res., v. 58, p. 537–552, 2009.
GARROTE, G. L.; ABRAHAM, A. G.; DE ANTONI, G. L. Chemical and microbiological
characterization of kefir grains. Journal of Dairy Research, v. 68, p. 639-652, 2001.
GAYLARD, C. C.; BELLINASO, M. L.; MANFIO, G. P. Aspectos biológicos e técnicas da
biorremediação de xenobióticos. Biotecnologia, Ciência e Desenvolvimento, Brasília, v. 8, n. 34,
2005.
GHASEMLOU, M; KHODAIYANA, F.; OROMIEHIEB, A.; Rheological and structural
characterization of film-forming solutions an biodegradable edible film made from kefiran as
affected by various plasticizer types. International Journal of Biological Macromolecules, v.
49, p. 814–821, 2011.
.GHOSH, C.; MANJUNATH,G. B.; KONAI, M. M.; DIVAKARA S. S. M. UPPU, D. S. S.;
HOQUE, J.; PARAMANANDHAM, K.; SHOME, B. R.; HALDAR J. Aryl-Alkyl-Lysines:
Agents That Kill Planktonic Cells, Persister Cells, Biofilms of MRSA and Protect Mice from
Skin-Infection PLoS One, v. 10, n. 12, DOI: 10.1371/journal.pone.0144094, 2015.
,GIADA, M. L. R.; MANCINI FILHO, J. Importância dos compostos fenólicos da dieta na
promoção da saúde humana. Publication UEPG Ciências Biológicas e da Saúde, v. 12, n. 4, p.
7-15, 2006.
GIESSIBL, F. J. Advances in atomic force microscopy. Rev. Mod. Phys., v. 75, p. 949, 2003.
GONÇALVES, A. E. S. Avaliação da capacidade antioxidante de frutas e polpas de frutas
nativas e determinação dos teores de flavonóides e vitamina c. 2008. 88 f. Dissertação
(Programa de Pós Graduação em Ciências dos Alimentos - Universidade de São Paulo: Faculdade
de ciências farmacêuticas. São Paulo. 2008.
GONTARD, N.; GUILBERT, S.; CUQ J.-L. Water and Glycerol as Plasticizers Affect
Mechanical and Water Vapor Barrier Properties of an Edible Wheat Gluten Film. Journal of
Food Science, v. 58, n. 1, p. 206-211, 1993.
GORBUSHINA A. A.; BROUGHTON, W. J. Microbiology of the atmosphere-rock interface.
how biological interactions and physical stresses modulate a sophisticated microbial ecosystem.
Annu. Rev. Microbiology, v. 63, n. 431–450, 2009.
GRAMZA, A.; KORCZAK, J. Tea constituents (Camelia sinensisL.) as antioxidant in lipid
systems. Trends in Food Science & Technology, v. 16, p. 351-358, 2005.

76
GRANILLO, A. R.; CANALES, M. G. M.; ESPÍNDOLA, M. E. S.; RIVERA, M. A. M.,
LUCIO, V. M. B.; TOVAR, A. V. R. Antibiosis interaction of Staphylococccus
aureus on Aspergillus fumigatus assessed in vitro by mixed biofilm formation BMC Microbiol.
v. 15, n. 33, 2015.
GRISEBACH, H. Biosynthesis of Anthocyanins. In: Markakis, P. (ed). Anthocyanins as Food
Color. New York: Academic Press, 1982, p. 69-90.
GRUMBEIN, S.; OPITZ, M.; LIELEG, O. Selected metal ions protect Bacillus subtilis biofilms
from erosion. Metallomics v. 6, n. 8, p. 1441-50, 2014.
GUZEL-SEYDIM, Z.; WYFFELS, J.; SEYDIM, A. C.; GREENE, A.; K. Turkish kefir and
kefigrains: microbial enumeration and electron microscopic observation. International Journal
of Dairy Technology, v. 58, p. 25-29, 2005.
GUZEL-SEYDIM, Z. B.; SEYDIM, A. C.; GREENE, A. K.; BODINE, A. B. Determination of
organic acids and volatile flavor substances in kefir during fermentation. Journal of Food
Composition and Analysis, v. 13, p. 35–43, 2000.
GUZEL-SEYDIM, Z.; SEYDIM, A. C.; GREENE, A. K. Organic acids and volatile flavor
components evolved during refrigerated storage of kefir. Journal of Dairy Science, v. 83, p.
275–277, 2000.
HAMET, M. F.; LONDERO, A.; MEDRANO, M.; VERCAMMEN, E.; VAN HOORDE, K.;
GARROTE, G. L. et al. Application of culture-dependent and culture-independent methods for
the identification of Lactobacillus kefiranofaciens in microbial consortia present in kefir
grains. Food Microbiology, v. 36, p. 327–334, 2013.
HANSON-RESEARCH-CORPORATION. Dissolution: Past, Present & Future. 2. ed.
Chatsworth: Hanson Research, 1996. p. 2-12.
HARAPANAHALLI, A. K.; CHEN, Y.; LI, J.; BUSSCHER, H. J.; VAN DER MEI, H. C.
Influence of Adhesion Force on icaA and cidA Gene Expression and Production of Matrix
Components in Staphylococcus aureus Biofilms. Appl Environ Microbiol, v. 81, n.10 p. 3369-
3378, 2015
HAYAKAWA, K., SANSAWA, H., NAGAMUNE, T., Endo, I. High density culture of
Lactobacillus casei by cross-flow culture method based on kinetic properties of the
microorganisms. J. Ferment. Bioengeering, v. 70, p. 589-593, 1990.
HEGEDÚS, A.; ENGEL, R.; ABRANKÓ, L.; BALOGH, E.; BLÁZOVICS, A.; HERMÁN, R.;
HALÁSZ, J.; ERCISLI, S.; PEDRYC, A.; STEFANOVITS-BANYAI, E. Antioxidant and
antiradical capacities in apricot (Prunus armeniaca L.) fruits: Variations from genotypes, years,
and analytical methods. Journal Food Science, v. 75, p. 722–730, 2010.
HERRMANN, P. S. P.; SILVA, M. A. P.; BERNARDES FO, R.; JOB, A. E.; COLNAGO, L. A.;
FROMMER, J. E.; MATTOSO, L. H. C. Microscopia de Varredura por Força: uma Ferramenta

77
Poderosa no Estudo de polímeros. Polímeros: Ciência e Tecnologia, ano VII, n. 4, p, 51-61,
1997.
HONG, S. H.; HEGDE, M.; KIM, J.; WANG, X.; JAYARAMAN, A.; WOOD, T.K. Synthetic
quorum-sensing circuit to control consortial biofilm formation and dispersal in a microfluidic
device. Nature Communications, v.3, p. 613, 2012
HOOSHANGI, S.; BENTLEY, W. E. From unicellular properties to multicellular behavior:
bacteria quorum sensing circuitry and applications. Current Opin. Biotechnology, v. 19, p. 550-
555, 2008.
HOSONO, A., TANABE, T.; OTANI, H. Binding properties of lactic acid bacteria isolated from
kefir milk with mutagenic amino acid pyrolyzates. Milchwiss, v. 45, p. 647-651, 1990.
HOUSTON, M. C. Nutraceuticals, vitamins, antioxidants, and minerals in the prevention and
treatment of hypertension. Progress in Cardiovascular Diseases, v. 47, n. 6, p. 396-449, 2005.
IADEROZA, M.; BALDINI, I. S. D.; BOVI, M. L. A.; Anthocyanins from fruits of açaí (Euterpe
oleracea Mart.) and juçara (Euterpe edulis Mart). Tropical Science, v.32, p. 41-46, 1992,
IMAI, S. Soybean and processed soy foods ingredients, and their role in cardiometabolic risk
prevention. Recent Pat Food Nutr Agric. v. 7, n. 2, p. 75-82, 2015.
IMHOF, R.; BOSSET, J. O. Relationships between microorganisms and formation of aroma
compounds in fermented dairy products. Zeitschrift Lebensmitel Unters Forschung, v.198, p.
267–276, 1994.
IMHOF, R., GLATTLI, H., BOSSET, J. O. Volatile organic compounds produced by
thermophilic and mesophilic mixed strain dairy starter cultures. Lebensmitel-Wissenschaft und-
Technologie, v. 27, p. 442–449, 1994.
IMHOF, R., GLATTLI, H., BOSSET, J. O. Volatile organic compounds produced by
thermophilic and mesophilic single strain dairy starter cultures. Lebensmitel-Wissenschaft und-
Technologie, v. 28, p. 78–86, 1995.
ISMAIEL, A. A.; GHALY, M. F.; EL-NAGGAR, A. K. Milk kefir: ultrastructure, antimicrobial
activity and efficacy on aflatoxin B1 production by Aspergillus flavus. Curr. Microbiology, v.
62, p. 1602–1609, 2011.
IVORRA, M.; ISSEKUTZ, A.; CORTIJO, J.; MORCILLO, E.; BLAZQUEZ, M.; SANZ M.; et
al. Effect of boldine, secoboldi-ne, and boldine methine on angiotensin II-induced neurtrophil
recruitment in vivo. Journal Leukocytes Biology, v. 78, p. 696-704, 2005.
JURD, L. Reactions Involued in Sulfite Bleacling of Antliocyanins. Juvenal. Food Science, v. 29
p.16-19, 1964
KANNAN, A.; KARUMANCHI, S. L.; KRISHNA, V.; THIRUVENGADAM, K.;
RAMALINGAM, S.; GAUTAM, P. Nanoscale investigation on Pseudomonas aeruginosa

78
biofilm formed on porous silicon using atomic force microscopy. Scanning, v. 36, n. 5, p. 551-
553, 2014.
KANDLER, O.; KUNATH, P. Lactobacillus kefir sp. nov., a component of the microflora of
kefir. Systematic Applied Microbiology, v. 4, p. 286-294, 1983.
KANEKO, D.; IGARASHI, T.; AOYAMA, K. Reduction of the off-flavor volatile generated by
the yogurt starter culture including Streptococcus thermophilus and Lactobacillus delbrueckii
subsp. bulgaricus in soymilk. J Agric Food Chem., v. 62, p. 1658-1663, 2014.
KIM, J.; CHUN, J.; HAN, H. U. Leuconostoc kimchii sp nov., a new species from kimchi.
International Journal of Systematic and Evolutionary Microbiology, v. 50, p. 1915-1919,
2000.
KING, A.; YOUNG, G. Characteristics and occurrence of phenolic phytochemicals. Journal of
American Diet. Association, v.99, p.213-218, 1999.
KNEIFEL, W.; ULBERTH, F.; ERHARD, F.; JAROS, D. Aroma profiles and sensory properties
of yogurt and yogurt-relatedproducts I. Screening of commercially available starter cultures.
Milchwissenchaft, v. 47, p. 362–365, 1992.
KNOLL M Aufladepotentiel und Sekundaremission elektronenbestrahter. Korper. Z tech. Phys.,
v. 16, p. 467-475, 1935.
KNOSHAUG, E. P.; AHLGRENT, J. A.; TREMPY, J. E. Growth associated polysaccharide
expression in Lactococcus lactis subsp cremoris ropy. Dairy Science, v. 83, p. 633–640, 2000.
KOBORI, M. In vitro-screening for cancer-supressive effect of food components, JARQ, v.37,
n.3, p. 159-165, 2003.
KODALI, V. P.; DAS, S.; SEN, R. K., An exopolysaccharide from a probiotic: Biosynthesis
dynamics, composition and emulsifying activity. Food Research International, v. 42, p. 695–
699, 2009.
KOJIĆ, M.; LOZO, J.; BEGOVIĆ, J.; JOVČIĆ, B.; TOPISIROVIĆ, L. J. Characterization of
Lactococci isolated from homemade kefir. Arch Biol Sci, v. 59, p. 13-22, 2007.
KONCZAK, I.; ZHANG, W. Anthocyanins-more than nature’s colours. Journal of Biomedicine
and Biotechnology., v. 5, p. 239-240, 2004.
KUDOU, S.; FLEURY, Y.; WELTI, D.; MAGNOLATO, D.; UCHIDA, T.; KITAMURA, K.;
OKUBO, K. Malonyl isoflvone glycosides in soybean seeds (Glycine max Merrill). Agricultural
and Biological Chemistry, v. 55, n. 9, p. 2227-2233, 1991
KUSKOSKI, E. M.; ASUERO, A. G.; MORALES, M. T.; FETT, R. Frutos Tropicais silvestres e
polpas congeladas: atividade antioxidante, polifenóis e antocianinas. Ciência Rural, v.36, n.4,
p.1283-1287, 2006.

79
LA RIVIERE, J. W. M.; KOOIMAN, P.; SCHMIDT, K. Kefiran, a novel polysaccharide
produced in the kefir grain by Lactobacillus brevis. Archives Microbiology, v. 59, p. 269-
278, 1967.
LAU, F. C.; SHUKITT-HALE, B.; JOSEPH,J. A. The beneficial effects of fruit polyphenolson
brain aging. Neurobiology of Aging., v. 26, p. 128-132. 2005.
LAUDANNA, E. Propriedades funcionais da soja. Informe Agropecuário, v. 27, n. 230, p. 15-
18, 2006.
LAUREYS, D.; DE VUYST L. Microbial species diversity, community dynamics, and
metabolite kinetics of water kefir fermentation. Appl. Environ. Microbiol., v. 80, p. 2564-72,
2014.
LAVIN, P.; DE SARAVIA, S. G.; GUIAMET, P. Scopulariopsis sp. and Fusarium sp. in the
Documentary Heritage: Evaluation of Their Biodeterioration Ability and Antifungal Effect of
Two Essential Oils. Microb. Ecol., v. 71, n. 3. p. 628-633, 2015.
LEE, K. W.; KIM, Y. J.; LEE, H. J.; LEE, C. Y. Cocoa has more phenolic phytochemicals and a
higher antioxidant capacity than teas and red wine. Journal of Agricultural and Food
Chemistry. v. 51, p. 7292-7295, 2003.
LILA, M. A. Anthocyanins and human health: an in vitro investigative approach. Journal of
Biomedicine and Biotechnology, v. 5, p. 306-313, 2004.
LIN, C. W.; CHEN, H. L.; LIU, J. R. Identification and Characterisation of lactic acid bacteria
and yeasts isolated from kefir grains in Taiwan. Australian journal of dairy technology, v. 54,
n. 1, p. 14-18, 1999.
LIU, J. R.; WANG S. Y.; LIN Y. Y.; LIN C. W. Antitumor activity of milk, kefir and soya milk
kefir in tumor bearing mice. Nutr. Cancer, v. 44, p. 183–187, 2002.
LOPITZ-OTSOA, F.; REMENTERIA, A; ELGUEZABAL, N.; GARAIZAR, J. Kefir: A
symbiotic yeasts-bacteria community with alleged healthy capabilities. Rev Iberoam Micol., v.
23, p. 67-74, 2006.
MACHERAS, P.; REPPAS, C.; DRESSMAN, J. B. Biopharmaceutics of orally administered
dosage forms. Chiester: Taylor & Francis, 1995. p. 75-91.
MAEDA, H.; ZHU, X. ; OMURA, K. ; SUZUKI, S.; KITAMURA, S. Effects of an
exopolysaccharide (kefiran) on lipids, blood pressure, blood glucose, and constipation.
Biofactors, v. 22, p. 97–200, 2004.
MAEDA, H.; ZHU, X.; OMURA, K.; SUZUKI, S.; KITAMURA, S. Structural characterization
and biological activities of an exopolysaccharide kefiran produced by Lactobacillus
kefiranofaciens WT-2B(T). J Agric Food Chem., v. 52, p. 5533-5538, 2004.

80
MALCHER, E. S. L. T; Influência da sazonalidade sobre a composição química e ativida
antioxidante do açaí (Euterpe oleracea Mart.). 2011. 189 f. Tese (Programa de Pós-Graduação
em Biodiversidade Tropical) Universidade Federal do Amapá, Macapá, 2011.
MANADAS, R.; PINA, M. E.; VEIGA, F. A dissolução in vitro na previsão da absorção oral de
fármacos em formas farmacêuticas de liberação modificada. Revista Brasileira de Ciências
Farmacêuticas., v. 38, p. 375-399, 2002.
MANDARINO, J. M. G. Compostos Antinutricionais da Soja: Caracterização e
Propriedades Funcionais. In: COSTA, N. M. B.; ROSA, C. O. B. Alimentos funcionais:
componentes bioativos e efeitos. Rio de Janeiro: Rubio, 2010. p. 177-192.
MARCHINI, F. B.; MARTINS, D. M. F. S.; TEVES, D. C.; SIMÕES, M. J. Efeito do óleo de
rosa mosqueta na cicatrização de feridas abertas. Revista Paulista de Medicina, v. 106, p. 356,
1998.
MARSHALL, V. M. Starter cultures for milk fermentation and their characteristics. Journal of
the Society of Dairy Technology, v. 46, p. 49-56, 1993.
MARSHALL, V. M.; COLE, W. M.; BROOKER, B. E. Observations on the structure of kefir
grains and the distribution of the microbiota. Journal of Applied Bacteriology, v. 57, p. 491-
497, 1984.
MATIJASIK, B.B.; ROGELSJ, I.; Lactobacillus K7, a probiotic strain. Food Technol
Biotechnol., v. 38, n. 1, p. 13–21, 2000.
MATSUDA, S. et al. Hydrolysis of isoflavones in soybean cooked syrup by Lactobacillus casei
subsp.rhaminosus IFO 3425. J. Ferm .Bioeng, v.74, p.301-304, 1992.
MATSUU, M.; SHICHIJO, K.; OKAICH, K. The Protective Effect of Fermented Milk Kefir on
Radiation-induced Apoptosis Res., 4 in colonic Crypt Cells of Rats. J. Radiat., v. 4, p. 111-115,
2003.
MAYER, C.; MORITZ, R.; KIRSCHNER, C.; BORCHARD, W.; MAIBAUM, R.;
WINGENDER, J.; FLEMMING, H.-C. The role of intermolecular interactions: studies on model
systems for bacterial biofilms. Int. J. Biol. Macromol., v. 26, p. 3-16, 1999.
MEDRANO, K.; PÉREZ, P. F.; ABRAHAM, A. G. Kefiran antagonizes cytopathic effects of
Bacillus cereus extracellular factors. International Journal of Food Microbiology, v. 122, p. 1–
7, 2008.
MENDES, F.; SIEUWERTS, S.; DE HULSTER, E.; ALMERING, M. J.; LUTTIK, M. A. et al.
Transcriptome-based characterization of interactions between Saccharomyces cerevisiae and
Lactobacillus delbrueckii subsp. bulgaricus in lactose-grown chemostat cocultures. Appl.
Environ. Microbiol., v. 79, p. 5949–5961, 2013.
MESUROLLE, J.; SAINT-EVE, A.; DÉLÉRIS, I.; SOUCHON, I. Impact of fruit piece structure
in yogurts on the dynamics of aroma release and sensory perception. Molecules, v. 18, n. 5, p.
6035-56, 2013.

81
MICHELI, L.; UCCELLETTI, D., PALLESCHI, C.; CRESCENZI, V. Isolation and
characterisation of a ropy Lactobacillus strain producing the exopolysaccharide kefiran. Appl.
Microbiol. Biotechnol. v. 53, p. 69–74, 1999.
MITHANI, S. D.; BAKATSELOU, V.; TENHOOR, C. N.; DRESSMAN, J. B. Estimation of the
increase in solubility of drugs as a function of bile salt concentration. Pharm. Res., v. 13, p. 163-
167, 1996.
MOONS, P.; MICHIELS, C. W.; AERTSEN, A. Bacterial interactions in biofilms. Critical
Reviews in Microbiology, v. 35, p. 157-168, 2009.
MORAES, I. C. F.; CARVALHO, R. A.; BITTANTE, A. Q. B.; SOLORZA-FERIA J.;
SOBRAL, P. J. A., Film forming solutions based on gelatin and poly(vinyl alcohol) blends:
Thermal and rheological characterizations. Journal Food Eng. v. 95, p. 588–596, 2009.
MUIR, D. D.; TAMIME, A. Y.; WSZOLEK, M. Comparison of the sensory profiles of kefir,
buttermilk and yogurt. International Journal of Dairy Technology, v. 52, n. 4, p. 129–134,
1999.
MULABAGAL, V.; KELLER, W. J.; CALDERÓN, A. I. Quantitative analysis of anthocyanins
in Euterpe oleracea (açaí) dietary supplement raw materials and capsules by Q-TOF liquid
chromatography/mass spectrometry. Pharm Biology, v. 50, n. 10, p. 1289-1296. 2012.
MUROFUSHI, M.; SHIOMI M.; AIBARA. K. Effect of orally administered polysaccharide from
kefir on delayed-type hypersensitivity and tumor growth in mice. Japan Journal Medicine
Science Biology v. 36, p. 49–53, 1983.
NADELL, C. D.; DRESCHER, K.; WINGREEN, N. S.; BASSLER, B. L. Extracellular matrix
structure governs invasion resistance in bacterial biofilms. ISME Journal Advance, v. 55, n. 11,
p. 396-407, 1997.
NEALSON K. H., HASTINGS J. W. Bacterial bioluminescence: its control and ecological
significance. Microbiology Review, v. 43, p. 496–518, 1979.
NÓBREGA, I. C. C. Análise dos compostos voláteis da aguardente de cana por concentração
dinâmica do "headspace" e cromatografia gasosa-espectrometria de massas. Ciênc. Tecnol.
Aliment., v. 23, n. 2, 2003.
NOGUEIRA, O. L.; FIGUERÊDO, F. J. C.; MULLER, A. A. Sistema de Produção do Açaí
(Euterpe oleracea Mart.). Embrapa Amazônia Oriental; Sistemas de Produção, n.4, Belém, PA,
2006.
OH, H. G ;NALLAMSHETTY, S.; RHEE, E. J. Increased Risk of Progression of Coronary
Artery Calcification in Male Subjects with High Baseline Waist-to-Height Ratio: The Kangbuk
Samsung Health Study. Diabetes Metab J., v. 40, n. 1, p. 54-61, 2016.
OHARA, H., HIYAMA, K., YOSHIDA, T. Lactic acid production by filter-bed-type reactor. J.
Ferment. Bioengineering, v. 76, p. 73-75. 1993.

82
OLIVEIRA D. M.; BASTOS, D. H. M. Biodisponibilidade de ácidos fenólicos Quim. Nova,
Vol. 34, No. 6, 1051-1056, 2011
OLIVEIRA, A. C.; VALENTIM, B. I.; GOULART, M. O. F.; SILVA, C. A.; BECHARA, E. J.
H.; TREVISAN, M. T. S. Fontes vegetais naturais de antioxidantes. Quím. Nova, v. 32, n. 3, p.
689-702 2009.
OLIVEIRA BRETT, A. M.; CHIORCEA PAQUIM, A. M.; DICULESCU, V.; ORETSKAYA,
T. S. Synthetic oligonucleotides: AFM characterisation and electroanalytical studies.
Bioelectrochemistry, v. 67, n.2, p. 181-90, 2005.
OLIVEIRA, M. S. P.; FERNANDES, T. S. D. Aspectos da Floração do Açaizeiro (Euterpe
olerácea Mart.) nas condições de Belém-PA. In: Congresso Nacional de Botânica, 44, São
Paulo, 1993. Resumos, São Luís: SBB, 1993. p.159.
OLIVEIRA, M. S. P.; FARIAS NETO, J. T. Variabilidade entre progênies de açaizeiro para
caracteres de germinação. In: Contribuição ao desenvolvimento da fruticultura na Amazônia.
Belém, PA, 2006, p.91-97.
ORAZEM, P.; STAMPAR, F.; HUDINA, M. Quality analysis of “Redhaven” peach fruit grafted
on 11 rootstocks of different genetic origin in a replant soil. Food Chem. v. 124, p. 1691–1698,
2011.
OTLES, S.; CAGINDI, O. Kefir: A Probiotic Dairy-Composition, Nutritional and Therapeutic
Aspects. Pakistan Journal of Nutrition, v. 2, n. 2, p. 54-59, 2003
OTT, A.; FAY, L. B.; CHAINTREAU, A. Determination and origin of the aroma impact
compounds of yogurt flavor. Journal of Agriculture and Food Chemistry, v. 45, p. 850–858,
1997.
OTTOGALLI, G.; GALLI, A.; RESMINI, P.; VOLONTERIO, G. Composizione microbiologica,
chimica ed ultrastructura dei granuli di Kefir. Ann. Microbiol. Enzymology, v. 23, p. 109-121,
1973.
OZELA, E. F.; STRINGHETA, P. C.; LIMA, A. A. S.; FARIAS, M. L. T. S. Estudo comparativo
do teor de antocianinas presente no açaí (Euterpe oleracea Mart.) nos períodos de safra e
entresafra. Livro de Programas e Resumo III Simpósio Americano de Ciências de
Alimentos. Campinas, FEA, p.55, 1999.
PARK, Y. K.; AGUIAR, C. L.; ALENCAR, S. M.; MASCARENHA, S. A.; SCAMPARINI, A.
R. P. Conversão de malonil-glicosil isoflavonas em isoflavonas glicosiladas presentes em alguns
cultivares de soja brasileira. Ciênc.Tecnol.Alim., v. 22, n.1 p. 30-35, 2002.
PARSEK, M. R.; FUQUA, C. Biofilms 2003: Emerging themes and challenges in studies of
surface-associated microbial life. Journal of Bacteriology, v. 186, p. 4427-4440, 2004.
PAUL, F.; MORIN, A.; MONSAN, P. Microbial polysaccharides with actual potential industrial
applications. Biotechnol. Adv., v. 4, p. 245–259, 1986.

83
PAUL, A.; PAUL, A. K. Microbial extracellular polymeric substances: central elements in heavy
metal bioremediation. Indian J. Microbiology, v. 48, p. 49-64, 2008.
PEETERS, R.; KINGET, R. Film-forming polymers for colonic drug delivery: I. Synthesis and
physical and chemical properties of methyl derivates of Eudragit S. Int. J. Pharm., v. 94, p. 125-
134, 1993.
PEIXOTO, H.; ROXO, M.; KRSTIN, S; RÖHRIG, T. RICHLING, E.; WINK, M. An
Anthocyanin-Rich Extract of Acai (Euterpe precatoria Mart.) Increases Stress Resistance and
Retards Aging-Related Markers in Caenorhabditis elegans. Journal Agric Food Chem, v. 64, n.
6, p. 1283-1290, 2016
PEREIRA, A. R. B.; FREITAS, D. A. F. Uso de Microorganismos Para a Biorremediação de
Ambientes Impactados. Rev. Elet. em Gestão, Educação e Tecnologia Ambiental, v. 6, n. 6, p.
975 – 1006, 2012.
PERESSINI, D.; BRAVIN, B.; LAPASIN, R.; RIZZOTTI, C.; SENSIDONI, A. Starch-
methylcellulose based edible films: rheological properties of film-forming dispersions. Journal
of Food Engineering, v. 59, p. 25-32, 2003.
PESSOA, J. D. C.; ARDUIN, M.; MARTINS, M. A.; CARVALHO, J. E. U. Characterization of
açaí (Euterpe oleracea Mart.) fruits and its processing residues. Braz. Arch. Biol. Technology,
v. 53, n. 6, 2010.
PIMENTEL, C. V. M. B.; FRANCKI, V. M.; GOLLUCKE, A. P. B. Alimentos funcionais:
Introdução às principais substâncias bioativas em alimentos. 1º edição, São Paulo, Editora
Varela, 2005.
PINTADO, M. E., LOPES DA SILVA, J. A., FERNANDES, P. B., MALCATA, F. X., HOGG,
T. A. Microbiological and rheological studies on Portuguese Kefir grains. International Journal
of Food Science and Technology, v. 31, p. 15–26.1996.
POGAČIĆ, T.; ŠINKO, S.; ZAMBERLIN, S.; SAMARŽIJA, D. Microbiota of kefir grains,
Mljekarstvo, v. 63, n. 1, p. 3-14, 2013.
POLI, A.; ANZELMO, G.; NICOLAUS, B. Bacterial Exopolysaccharides from Extreme Marine
Habitats: Production, Characterization and Biological Activities. Mar Drugs, v. 8, n. 6, n. 1779–
1802, 2010.
PRADO, M. R.; BLANDÓN, L. M.; VANDENBERGHE, L. P. S.; RODRIGUES, C.;
CASTRO, G. R.; SOCCOL, V. T.; SOCCOL, C. R. Milk kefir: composition, microbial cultures,
biological activities, and related products. Front Microbiol, v. 6, p. 1177-1187, 2015.
PREECHAGOON, D.; UDOMPRATEEP, A.; MANWIWATTANAGUL, G. Improved
dissolution rate of poorly soluble drug by incorporation of buffers. Drug Dev. Ind. Pharm. v.
26, n. 8, p. 891-894, 2000.

84
QUEIROZ, J, A, L.; MOCHIUTTI, S. Manejo de mínimo impacto para produção de frutos em
açaizais nativos no estuário Amazônico. Comunicado Técnico-Embrapa –AP. Novembro,
2001; p:1-5.
RASTALL, R. A.; GIBSON, G. R.; GILL, H. S.; GUARNER F.; KLAENHAMMER, T. R.;
POT, B.; REID, G.; ROWLAND, I. R.; SANDERS, M. E. Modulation of the microbial ecology
of the human colon by probiotics, prebiotics and symbiotic to enhance human health: An
overview of enabling science and potential applications. FEMS Microbiology Ecology, v. 52, p.
145-152, 2005.
REHM, B. Microbial exopolysaccharides: Variety and potential applications. In Microbial
production of biopolymers and polymer precursors: applications and perspectives; Caister
Academic: Norfolk, UK, 2009; p. 229-254.
RIBEIRO, E. P.; SERAVALLI, E. A. G. Química de Alimentos. São Paulo: Editor Edgar
Blücher: Instituto Mauá de Tecnologia, 2004, 184p.
RICE-EVANS, C. A.; MILLER, N. J.; PAGANGA, G. Iron-mediated free radical effects on
erythrocytes: the role of desferrioxamine. Free Radic. Biol. Med. v. 20, n.7, p. 933-956, 1996.
RODRIGUES, K. L.; CAPUTO, L. R.; CARVALHO, J. C. T.; EVANGELISTA, J.;
SCHNEEDORF, J. M. Antimicrobial and healing activity of kefir and kefiran extract. Int J
Antimicrob Agents, v. 25, n. 5, p. 404-408, 2005.
ROGEZ, H. Açaí: Preparo, Composição e Melhoramento da Conservação. Belém: Edufpa,
2000, p.313.
ROMANI, A. M.; FUND, K.; ARTIGAS, J.; SCHWARTZ, T.; SABATER, S.; OBST, U.
Relevance of polymeric matrix enzymes during biofilm formation. Microb. Ecol., v. 56, p. 427-
436, 2008.
ROSSI, E. A.; ROSIER, I.; DÂMASO, A. R.; CARLOS, I. Z.; VENDRAMINI, R. C.;
ABDALLA, D. S. P.; TALARICO, V. H.; MINTO, D. F. Determinação de isoflavonas nas
diversas etapas do processamento do “iogurte” de soja. Alim. Nutr., v. 15, p. 93-99, 2004.
RUBIO-SIERRA,, F. J.; HECKL, W. M.; STARK, R. W. Nanomanipulation by Atomic Force
Microscopy. Advanced Engineering Materials, v. 7, n. 4, p. 193–196, 2005.
RUIZ, L. M.; VALENZUELA, S.; CASTRO, M.; GONZALEZ, A.; FREZZA, M.; SOULÈRE,
L.; ROHWERDER, T.; QUENEAU, Y.; DOUTHEAU, A.; SAND, W.; RUMJANEK, N. G.;
FONSECA, M. C. C.; XAVIER, G. R. Quorum sensing em sistemas agrícolas. Revista
Biotecnologia Ciência e Desenvolvimento, v. 33, p. 35-50, 2004.
SAAD, S. M. I. Probióticos e prebióticos: o estado da arte. Revista Brasileira de Ciências
Farmacêuticas 42: 1-16, 2006.
SALOM, C. M.; SILVA, W. J.; CALLEGARI, F. L.; ROCHA, A. V.; BORGES, M.;
JERÔNIMO, M. Desenvolvimento, aceitabilidade e intenção de compra de bebida láctea com

85
adição de leite fermentado de kefir e polpa de manga. III Seminário de Iniciação Científica e
Inovação Tecnológica, 15/10/2010.
SANCHEZ, S.; BRAVO, V.; MOYA, A. J.; CASTRO, E.; CAMACHO, F., Influence of
temperature on the fermentation of D-xyklose by Pachysolem tannophilus to produce ethanol and
xylitol. Proc. Biochem., v. 39, p. 673-679, 2004.
SANTOS, A.; SAN MOURO, M.; SANCHEZ, A.; TORRES, J. M.; MARQUINA, D. The
antimicrobial properties of different strains of Lactobacillus spp. isolated from kefir. Systematic
and Applied Microbiology, v. 26, p. 434-437, 2003.
SANTOS, G. M.; MAIA, G. A.; SOUSA, P. H. M.; COSTA, J. M. C.; FIGUEIREDO, R. W.;
PRADO, G. M. Correlação entre atividade antioxidante e compostos bioativos de polpas
comerciais de açaí (Euterpe oleracea Mart.) Archivos Latinoamericanos de Nutrición, ALAN,
v.58, n.2, 2008.
SHAUSS, A. G.; WU, W.; PRIOR, R. L.. Phytochemical and nutrient of the freezedried
Amazonian pal berry, Euterpe oleracea Mart. Journal of Agriculture and food chem, 54, 8598-
8603, 2006.
SHIMADA, K.; ITOH, Y.; WASHIO, K.; MORIKAWA, M. Efficacy of forming biofilms by
naphthalene degrading Pseudomonas stutzeriT102 toward bioremediation technology and its
molecular mechanisms. Chemosphere, v. 87, p. 226–233, 2012.
SHIOMI, M.; SASAKI, K.; MUROFUSHI, M.; AIBARA, K. Antitumor acitivity in mice of
orally administered polysaccharide from kefir grain. Jpn. J. Med. Sci. Biol., v. 35, p.75-80,
1982.
SHUAI, S; YUE, S.; HUANG, Q.; WANG, W.; YANG, J.; LAN, K.; YE, L. Preparation,
characterization and in vitro/vivo evaluation of tectorigenin solid dispersion with improved
dissolution and bioavailability. Eur J Drug Metab Pharmacokinet, 2015 IN PRESS
SILVA, C. R.; LIMA, S. M. R.; ALVES, D.L.; AOKI, T. Análise crítica dos efeitos da glycine
max nas doenças cardiovasculares e seus fatores de risco / Critical analysis on the effects of
glycine max in cardiovascular diseases and its risk factors. Femina. v. 37, n.2, p. :107-113, 2009.
SILVA, R. S. Potencial antioxidante correlacionado com fenóis totais e antocianinas de
cultivares de amora-preta, mirtilo, morango e pêssego – 2007. 58f. Dissertação (Programa de
Pós-Graduação em Ciência e Tecnologia Agroindustrial) - Faculdade de Agronomia Eliseu
Maciel. Universidade Federal de Pelotas. - Pelotas, 2007.
SIMOVA, E.; BESHKOVA, D.; ANGELOV, A.; HRISTOZOVA, T. S., FRENGOVA, G.;
SPASOV, Z. Lactic acid bacteria and yeasts in ke¢r grains and ke¢r made from them. J. Ind.
Microbiol. Biotechnol., v. 28, p. 1-6, 2002.
SINGH, P.; CAMEOTRA, S. S. Enhancement of metal bioremediation by use of microbial
surfactants. Biochem. Biophys. Res. Commun, v. 319, 291-297, 2004.

86
SINGH, R.; PAUL, D.; JAIN, R. K. Biofilms: implications in bioremediation. Trends Microbiol
v. 14, p. 389-397, 2006.
SKREDE, G.; WROLSTAD, R. E. Flavonoids from berries and grapes. Functional Foods:
Biochemical and Processing Aspects. Chapter 3, v.2, CRC Press LLC.2002. Disponível online
em: < http://lab.ac.ntu.edu.tw/cereal/Course_Download/>.
SLIMESTAD, R.; VANGDAL, E.; BREDE, C. Analysis of phenolic compounds in six
norwegian plum cultivars (Prunus domestica L.). J. Agric. Food Chem., v. 57, p. 11370–11375,
2009.
SOARES, S.E. Ácidos fenólicos como antioxidantes. Revista de Nutrição. v.15, n. 1, p. 71-81,
2002.
SOUSA, S.; BANDEIRA, M.; CARVALHO, P. A.; DUARTE, A.; JORDAO, L. Nontuberculous
mycobacteria pathogenesis and biofilm assembly. Int. J. Mycobacteriol. v. 4, n. 1, p. 36-43,
2015.
SPINLER, J. K.; ROSS, C. L.; SAVIDGE, T. C. Probiotics as adjunctive therapy for preventing
Clostridium difficile infection - What are we waiting for?. Anaerobe, 2016. IN PRESS.
STADIE, J.; GULITZ, A.; EHRMANN, M. A.; VOGEL, R. F. Metabolic activity and symbiotic
interactions of lactic acid bacteria and yeasts isolated from water kefir. Food Microbiol, v. 35, p.
92-98, 2013.
STEWART, P. S. Mechanisms of antibiotic resistance in bacterial biofilms. International
Journal of Medical Microbiology, v. 292, n. 2, p. 107–113, 2002.
SUGIYAMA, A.; YAMAZAKI, Y.; YAMASHITA, K.; TAKAHASHI, S.; NAKAYAMA, T.;
YAZAKI, K. Developmental and nutritional regulation of isoflavone secretion from soybean
roots. Bioscience, Biotechnology and Biochemistry, v. 80, n. 1, p. 89-94, 2015.
SULLIVAN, A.; NORD, C. E. Probiotics in human infections. Journal Antimicrob Chemother,
v. 50, p. 625–627; 2000.
SUTHERLAND, I. W. Biofilm exopolysaccharides: a strong and sticky framework.
Microbiology, v. 147, p. 3-9, 2001.
TAKIZAWA, S.; KOJIMA, S.; TAMURA, S.; FUJIMAGA, S.; BENNO, Y.; NAKASE, T.
Lactobacillus kefirgranum sp. nov. and Lactobacillus parakefir sp. nov., two new species from
kefir grains. Int. J. Syst. Bacteriol., v. 44, p. 435-439, 1994.
TAKIZAWA, S.; KOJIMA, S.; TAMURA, S.; FUJINAGA, S.; BENNO, Y.; NAKASE, T. The
composition of the Lactobacillus flora in kefir grains. Systematic and Applied Microbiology, v.
21, p. 121–127, 1998.

87
TAMIME, A. Y.; ROBINSON, R. K. Biochemistry of fermentation. In A. Y. Tamime, & R. K.
Robinson (Eds.), Yoghurt science and technology (2nd ed.) (pp. 432–475). Cambridge, UK,
1999.
TAYLOR, P. K.; YEUNG, A. T.; HANCOCK, R. E. Antibiotic resistance in Pseudomonas
aeruginosa biofilms: towards the development of novel anti-biofilm therapies. J Biotechnol, v
191, p. 121-130, 2014.
TERCI, D. B. L. Aplicações analíticas e didáticas de antocianinas extraídas de frutas. 213f, tese
(Doutorado em Química e Educação) – Departamento de Química Analítica, Universidade de
Campinas, São Paulo, 2004.
TEZUKA, H.; IMAI, S. Immunomodulatory Effects of Soybeans and Processed Soy Food
Compounds. Recent Pat Food Nutr Agric, v. 7, n. 2, p. 92-99, 2015.
THEWES, N.; LOSKILL, P.; JUNG, P.; PEISKER, H.; BISCHOFF, M.; HERRMANN, M.;
JACOBS, K. Hydrophobic interaction governs unspecific adhesion of staphylococci: a single cell
force spectroscopy study. Beilstein J Nanotechnol, v. 5, p. 1501-12, 2014.
TOBA, T.; ARIHARA, K.; ADACHI, S. Comparative study of polysaccharides from kefir
grains, an encapsulated homofermentative Lactobacillus species and Lactobacillus kefir.
Milchwissenschaften, v. 42, p. 565-568, 1987.
TRELEA, I. C.; ATLAN, S.; DÉLÉRIS, I.; SAINT-EVE, A.; MARIN, M.; SOUCHON, I.
Mechanistic mathematical model for in vivo aroma release during eating of semiliquid foods.
Chem Senses. v. 33, n. 2, p. 181-92, 2008.
TREVISANATO, S. I.; KIM, Y. I. Tea and Health. Nutrition Reviews, v. 58, n. 1, p.1-10, 2000.
TSUKAMOTO, C.; SHIMADA, S.; IGITA, K.; KUDOU, S.; KOKUBUN, M.; KUBO, K.;
KITAMURA, K. Factors Affecting Isoflvone Content in Soybean Seeds: Changes in Isoflvones,
Saponins, and Composition of Fatty Acids at Different Temperatures during Seed Development.
Journal of Agricultural and Food Chemistry, v. 43, n. 5, p. 1184-1192, 1995.
UI, J. H.; QIAN, Y.; MIAO, W. J.; XIA, Q. Preparation and in vitro evaluation of gastroretentive
dosage form containing puerarin-HP-beta-CD inclusion complex. Zhongguo Zhong Yao Za
Zhi, v. 33, p. 1960-1963, 2008.
VAN HULLEBUSCH, E. D.; ZANDVOORT, M. H.; LENS, N. L. Metal immobilisation by
biofilms: mechanisms and analytical tools. Rev. Environ. Sci. Biotechnol, v. 2, p. 9-33, 2003.
VAN HULLEBUSCH, E. D.; ZANDVOORT, M. H.; LENS, P. N. L. Metal immobilisation by
biofilms: mechanisms and analytical tools. Rev. Environ. Sci. Biotechnol., v. 2, p. 9-33, 2003.

88
VARINSKA, L.; GAL, P.; MOJZISOVA, G.; MIROSSAY, L.; MOJZIS, J. Soy and breast
cancer: focus on angiogenesis. Int J Mol Sci, v. 16, p. 11728-49, 2015.
VATTEM, D. A.; GHAEDIAN, R.; SHETTY, K. Enhancing health benefits of berries through
phenolic antioxidant enrichment: focus on cranberries. Asia Pac Journal Clinical Nutrition,
v.14, n.12, p.120-130, 2005.
VERHAGEN, P.; DE GELDER, L.; HOEFMAN, S.; DE VOS, P.; BOON, N. Planktonic versus
biofilm catabolic communities: importance of the biofilm for species selection and pesticide
degradation. Appl. Environ. Microbiol, v. 10, p. 1128, 2011.
VIDIGAL, P. G.; SVIDZINSKI, T. I. E. Leveduras nos tratos urinário e respiratório: infecção
fúngica ou não? J Bras Patol Med Lab., v. 45, n. 1, p. 55-64, 2009.
VIDYALAXME, B.; ROVETTO, A.; GRAU, R.; AGRAWAL, R. Synergistic effects of
probiotic Leuconostoc mesenteroides and Bacillus subtilis in malted ragi (Eleucine corocana)
food for antagonistic activity against V. cholerae and other beneficial properties. J Food Sci
Technol, v. 51, n. 11, p 3072-3082, 2014.
VINDEROLA, G.; DUARTE, J.; THANGAVEL, D.; FARNWORTH, E.; PERDIGON, G.;
MATAR, C. Immunomodulating capacity of kefir. Journal of Dairy Research. v. 75, p.175-202,
2006.
VU, B.; CHEN, M.; CRAWFORD, R. J.; IVANOVA E. P. Bacterial Extracellular
Polysaccharides Involved in Biofilm Formation. Molecules, v. 14, p. 2535-2554, 2009.
WANG, H. J. Quantification of potentially anticarcinogenic isoflavones in soy foods and the
effect of processing on the composition of isoflavones. Dissertation Abstracts Intern., B, v. 55,
n. 7, 128p. 1995.
WANG, H. J.; MURPHY, P. A. Isoflavone content in commercial soybean foods. J. Agric.Food
Chem, v. 42, p.1666-1673, 1994.
WANG, S. Y.; CHEN, K. N.; LO, Y. M.; CHIANG, M. L.; CHEN, H. C.; LIU, J. R. et al.
Investigation of microorganisms involved in biosynthesis of the kefir grain. Food Microbiology,
v. 32, p. 274–285, 2012.

89
WANG, Y.; AHMED, Z.; FENG, W.; LI, C.; SONG, S. Physicochemical properties of
exopolysaccharide produced by Lactobacillus kefiranofaciens ZW3 isolated from Tibet kefir.
International Journal of Biological Macromolecules, v. 43, p. 283-288, 2008.
WANG, Y; LI, C; LIU, P; AHMED, Z; XIAO, P; BAI, X. Physical characterization of
exopolysaccharide produced by Lactobacillus plantarum KF5 isolated from Tibet Kefir.
Carbohydr. Polym., v. 82, p. 895–903, 2010.
WATERS, C. M.; BASSLER, B. L. Quorum sensing: cell-to-cell communication in bacteria.
Annu. Rev. Cell Dev. Biol., v. 21, p. 319-346, 2005.
WATNICK, P., KOLTER, R. Biofilms, city of microbes. J. Bacteriol., v. 13, p. 20–26, 2000.
WILWERTH, M. W.; SILVA, C. V. N.; SOUZA, G. B.; BERNARDI, A. C. C.; LOPES, C. H.;
BORGES, M. T. M. R.; VERRUMA-BERNARDI, M. R. Determinação de minerais e metais
pesados em açúcar mascavo, 8a Jornada Científica UEFSCCA, 5 a 9 de outubro de 2009
WISEMAN, A. Functional-food protected by biomonitoring of reactive oxygen species (ROS).
Trends in Food Science & Technology. v.16, p.166-168, 2005.
WITTHUHN, R. C.; SCHOEMAN, T.; BRITZ, T. J. Isolation and characterisation of the
microbial population of different South African Kefir grains. International Journal of Dairy
Technology, v. 57, p. 33–37, 2004.
WITTHUHN. R. C.; SCHOEMAN, T.; BRITZ, T. J. Characterisation of the microbial population
at different stages of Kefir production and Kefir grain mass cultivation. International Dairy
Journal, v. 15, p. 383–389, 2005.
WOLFE, N. L.; ZEPP, R. G.; PARIS, D. F. Carbaryl, propham and chlorpropham - Comparison
of rates of hydrolysis and photolysis with rate of biolysis. Water Research, v. 12, p. 565-571,
1978.
XU, X.; LIU, W.; ZHANG, L. Rheological behavior of Aeromonas gum in aqueous solutions.
Food Hydrocolloids, v. 20, n.5, p. 723-729, 2006.
YEESANG C.; CHANTHACHUM, S.; CHEIRSILP, B. Sago starch as a low-cost carbon source
for exopolysaccharide production by Lactobacillus kefiranofaciens. World J Microbiol
Biotechnol, v.24, p.1195–1201, 2008.

90
YEH, C. C.; FAN, Y.; JIANG, L.; YANG, Y. L.; HE, B.; YOU, L.; MANN, M. Genistein
Suppresses Growth of Human Uterine Sarcoma Cell Lines via Multiple Mechanisms. Anticancer
Res, v. 35, n. 6, p. 3167-3173, 2015.
YOKOI, H.; WATANABE, T. 1992. Optimum culture conditions for production of kefiran by
Lactobacillus sp. KPB-167B isolated from kefir grains. J. Ferment. Bioeng., v. 74, p. 327-329,
1992.
YU, F.; LIU, Z.; TONG, Z.; ZHAO, Z.; LIANG, H. Soybean isoflavone treatment induces
osteoblast differentiation and proliferation by regulating analysis of Wnt/β-catenin pathway.
Gene, v. 573, n. 2, p. 273-277, 2015.
ZACCONI, C.; SCOLARI, G.; VESCOVO, M.; SARRA, P. G. Competitive exclusion of
Campylobacter jejuni by kefir fermented milk. Annals of Microbiology, v. 53, p. 179-187, 2003.
ZAJŠEK, K.; GORŠEK, A. Modeling of batch kefir fermentation kinetics for ethanol production
by mixed natural microflora. Food and Bioproducts, v. 88, p. 55-60, 2010.
ZAMBERI, N. R.; ABU, N.; MOHAMED, N. E.; NORDIN, N.; KEONG, Y. S.; BEH, B. K.;
ZAKARIA, Z. A.; NIK ABDUL RAHMAN, N. M.; ALITHEEN, N. B. The Antimetastatic and
Antiangiogenesis Effects of Kefir Water on Murine Breast Cancer Cells. Integr Cancer Ther,
2016. IN PRESS.
ZHANG, W.; DAI, W.; TSAI, S. M.; ZEHNDER, S. M.; SARNTINORANONT, M.;
ANGELINI, T. E. Surface indentation and fluid intake generated by the polymer matrix of
Bacillus subtilis biofilms. Soft Matter, v. 11, n. 18, p. 3612-3617, 2015.
ZHANG, Z.; KOU, X.; FUGAL, K.; MCLAUGHLIN, J. Comparison of HPLC Methods for
Determination of Anthocyanins and Anthocyanidins in Bilberry Extracts. J. Agric. Food
Chem., v. 52, n. 4, pp 688–691, 2004.
ZHOU, J.; LIU, X.; JIANG, H.; DONG, M. Analysis of the microflora in Tibetan kefir grains
using denaturing gradient gel electrophoresis. Food Microbiol, v. 26:770–5, 2009.

91
3 OBJETIVOS
3.1 GERAL
Padronizar a produção de biofilmes de kefir tendo como substrato açúcar mascavo, açúcar branco
e aferir suas interações com extrato de açaí (Euterpe oleracea Mart.) e com extrato de gérmen de
soja (Glycine max (L.) Merril) e realizar a determinação de parâmetros de obtenção do meio de
cultura com kefir e açúcar mascavo e com açúcar branco em interação com extrato de açaí
(Euterpe oleracea Mart.) e com extrato de gérmen soja (Glycine max (L.) Merril).
3.2 ESPECÍFICOS
Aferir a viabilidade do uso de açúcar branco para formar biofilmes com kefir e aferir o pH
e temperatura para formação destes biofilmes
Estabelecer o pH e temperatura para produção de biofilmes de kefir tendo como substrato
açúcar mascavo e produção de biofilmes com açúcar mascavo associado ao extrato de açaí
(Euterpe oleracea Mart.) e para biofilmes de kefir com açúcar mascavo associado com
extrato de gérmen de soja (Glycine max (L.) Merril).
Determinar os compostos voláteis majoritários sintetizados em meio de cultura de kefir
com açúcar mascavo.
Proceder a análise da rugosidade dos biofilmes bem como a estrutura dos biofilmes de
kefir cultivado em substrato açúcar mascavo e em biofilme de kefir com com extrato de
açaí (Euterpe oleracea Mart.) bem como, em kefir cultivado com gérmen de soja (Glycine
max (L.) Merril) utilizando Microscopia de Força Atômica (AFM) e microscopia
eletrônica de varredura MEV;
Extração e quantificação das antocianinas que foram incorporadas biofilme de kefir com
com extrato de açaí (Euterpe oleracea Mart.) e presentes no meio de cultura.
Extração e quantificação as isoflavonas que foram incorporadas aos biofilmes com gérmen
de soja (Glycine max (L.) Merril) bem como das isoflavonas presentes no respectivo meio
de cultura.
Avaliação do perfil de dissolução de antocianinas em biofilmes de extrato de açaí em

92
condições de simulação dermatológica;
Avaliação da estabilidade de antocianinas incorporadas em biofilmes de extrato de açaí
(Euterpe oleracea Mart.)
Avaliação do perfil de dissolução de isoflavonas de biofilmes de kefir com gérmen de soja
(Glycine max (L.) Merril) em simulação do sistema gastrintestinal;
Análise da rugosidade de biofilmes de kefir cultivado em açúcar mascavo e com gérmen
de soja (Glycine max (L.) Merril) utilizando Microscopia de Força Atômica (AFM);
Determinar a composição de macro e microelementos de meio de cultura e de biofilmes
formados com kefir e açúcar mascavo, em biofilmes com extrato de açaí (Euterpe oleracea
Mart.) e com extrato de gérmen de soja (Glycine max (L.) Merril).

93
CAPÍTULO 1: ESTUDO DA OBTENÇÃO DE BIOFILMES DE KEFIR EM
DIFERENTES SUBSTRATOS

94
Estudos da viabilidade de obtenção e padronização de biofilmes de kefir
Antônio Ferreira de Oliveira1,2, Cleydson Breno Rodrigues dos Santos2, Adriana Maciel
Ferreira2, Roberto Messias Bezerra2, Robert Ronald Magña Zamora3, Rodrigo Alves
Soares Cruz4, Jesus Rafael Rodriguez Amado2 and José Carlos Tavares Carvalho1,2*
1Programa de Pós-graduação em Biodiversidade Tropical, Departamento de Ciências Biológicas e da Saúde,
Universidade Federal do Amapá *2Laboratorio de Pesquisa em Fármacos, Curso de Farmácia, Departamento de Ciências Biológicas e da Saúde,
Universidade Federal do Amapá, Rodovia Juscelino Kubitisheck, km 02, CEP 68902-290, Macapá, Amapá, Brasil 3Laboratorio de Nanotecnologia de Materiais, Curso de Física, Departamento de Ciências Exatas e Naturais,
Universidade Federal do Amapá 4Laboratorio de Nanobiotecnologia Fitofarmacêutica, Curso de Farmácia, Departamento de Ciências Biológicas e
da Saúde, Universidade Federal do Amapá
*Corresponding to author: email [email protected]
RESUMO
O kefir é uma bebida obtida através da fermentação por Lactobacillus e leveduras, sendo
considerado um probiótico. A formação de biofilmes a partir de grãos de kefir (KG) foi
padronizada utilizando soluções de açúcar branco (AB) e mascavo (AM) como meio de cultura.
Diferentes concentrações de substrato (AB e AM) e do inóculo (KG) foram avaliadas para a
determinação da concentração necessária para a formação de biofilmes. Após 20 dias os
biofilmes formados na superfície do meio de cultura foram analisados por microscopia óptica
(OM), microscopia de força atômica (AFM), e microscopia eletrônica de varredura (SEM). Os
macro e microelementos foram quantificados por espectrofotometria de absorção atômica (AAS).
A concentração de KG necessária para a formação de biofilme em ambos substratos foi de 20 g/L
e a concentração de AM e AB para formação de biofilme foi de 40 g/L e 20 g/L, respectivamente.
Com relação a determinação de macro e microelementos a concentração de Fe+2 foi de 2,725
mg/kg em AM e 1,570 mg/kg em AB. A concentração de Ca+2 foi maior nos biofilmes formados
em AB (2,150 mg/kg) do que nos formados em AM (0,550 mg/kg). Também foram encontrados
Mg+2 (1,589 mg/kg em AM e 1,862 mg/kg em AB), K+ (2,405 mg/kg em AM e 2,645 mg/kg em
AB) e Zn+2 (0,276 mg/kg para o AM, e 0,849 mg/kg em AB). A análise dos compostos voláteis
do meio de cultura do kefir por cromatografia gasosa acoplada a espectrometria de massa (CG-
EM) mostrou os seguintes compostos voláteis majoritários: ácido hexadecanoico, eicosano, 2
metil-eicosano. A microscopia óptica a 400 x revelou as estruturas de formação de biofilmes e a
fotomicrografia eletrônica de varredura (SEM) mostrou a coadesão de bactérias às hifas de
fungos filamentosos. As imagens de AFM mostraram que a rugosidade (RMS) da superfície do
biofilme é dependente das concentrações de AM e KG. A partir dos resultados obtidos pode-se
afirmar que pelas técnicas físico-quimicas aplicadas foi possível padronizar a obtenção, e
caracterizar os biofilmes de kefir, servindo como meio veiculação de elementos nutritivos ou
funcionais.
Palavras-chave: Biofilmes. Kefir. Lactobacillus. Leveduras. Probiótico.

95
ABSTRACT
Kefir a beverage produced through fermentation by Lactobacillus and yeast is considering a
probiotic. The occurrence of biofilm formation from kefir grains was determined using white and
brown sugar solution as culture media. Differents concentrations of substrate and inoculum
(grains of kefir) were tested in order to determine optimal concentration for biofilm formation.
After 20 days kefir grains and biofilms formed on the surface of culture media were weighed to
determine growth rate. The biofilms were analyzed for microelements and structure using optical
microscopy and Atomic Force Microscopy (AFM) and scanning electron microscopy (SEM). The
macro- and microelements were quantified by atomic absorption spectrophotometry (AAS) The
best substrate concentration for biofilm formation was 40 gl-1 with brown sugar (BS) and 20 gl-1
with white sugar (WS). Regarding the macro and microelements, the concentration of Fe+ 2 was 2,725
mg/kg in BS and 1,570 mg/kg in WS. The concentration of Ca2+ was higher in biofilms with WS (2,150
mg/kg) than that formed with BS (0.550 mg/kg). Also found Mg + 2 (1,589 mg / kg in BS and 1,862
mg / kg in WS), K + (2,405 mg / kg in BS and 2,645 mg / kg WS) and Zn + 2 (0.276 mg / kg for
the BS and 0.849 mg / kg WS). The analysis of volatile compounds of kefir culture medium by
gas chromatography-mass spectrometry (GC-MS) showed the following volatile compounds
majority: hexadecanoic acid, eicosane, 2-methyl-eicosane. The AFM images showed that the
surface roughness (RMS) of the surface of the biofilm is dependent on the concentrations of
sugar brown sugar (BS) and kefir grains (KG). Photomicrography by scanning electron microscopy
(SEM) showed the adhesion of bacteria to the hyphae of filamentous fungi, while AFM images showed
the surface roughness (RMS). Thus, physical and chemical techniques were applied to standardize the
production and characterization of kefir biofilms. From the results it can be said that the physical-
chemical techniques applied was possible to standardize and characterize kefir biofilms, serving
as a means placement nutritious or functional elements
.Key Words: Biofilms, kefir, Lactobacillus, yeast, probiotic, Atomic Force Microscopy
AFM

96
1 INTRODUÇÃO
Na História da Microbiologia os microrganismos foram primeiramente descritos como
sendo unicelulares. Analisando o seu metabolismo a partir dessa perspectiva foram descritos com
base em suas características em meio de cultura nutricionalmente rico. Na natureza, no entanto,
em geral não há meio com abundância de nutrientes, portanto estes seres têm desenvolvido
estratégias que lhes permitem comunicar-se uns com os outros e agir como um ser multicelular.
Isto ocorre através da formação de biofilmes, que cumprem várias funções que lhes permite
sobreviver em diversos habitats da biosfera (FLEMMING; WINGENDER 2001a;
SUTHERLAND, 2001; VAN HULLEBUSCH, 2003; VU et al., 2009).
Nos últimos anos tem sido descrito que na natureza microrganismos existem
predominantemente como biofilmes e através desta estratégia adquirem alta tolerância a estresse
biológico, químico e físico (GORBUSHINA; BROUGHTON 2009) além de protegerem as
bactérias de bacteriófagos, amebas, vários biocidas e antibióticos (COSTERTON et al., 1999,
CERQUEIRA et al., 2013).
Kefir é uma bebida obtida através da fermentação do leite ou suco de frutas ou água com
açúcar, e é metabolizada por Lactobacillus e leveduras que formam os grãos de kefir. Os KG se
assemelham a couve em flor, medindo de 3 mm até a 1 cm. São de forma irregular, branco ou
amarelo (LA RIVIERE et al., 1967). Atua como probiótico e impede a proliferação de seres
patogênicos cujo comportamento é individualizado com relação aos seres probióticos. Somente
seres com capacidade especial conseguem formar consórcios com outros seres (LAUREYS; DE
VUYST, 2014). Organismos probióticos, com efeito, conseguem evitar contaminantes através da
competição por sítio e por nutrientes. Além disso o kefir secreta biocidas que repelem os seres
indesejáveis.
O alto teor de açúcar e a baixa concentração de aminoácidos encontrada no kefir
metabolizado em água deveria resultar em um habitat carente, no entanto o que se obtém é uma
bebida fermentada com uma comunidade microbiana bastante diversificada, e estável (STADIE
et al., 2013). Estabelece-se um nicho ecológico onde apenas organismos muito bem adaptados em
associação com outros conseguem crescer e se prover de nutrientes essenciais. No kefir obtido
com água e açúcar ocorre um sinergismo entre leveduras e bactéria ácido-lácticas notadamente o
Lactobacillus (STADIE et al., 2013). O cultivo de ambos em consórcio no kefir mostrou

97
crescimento na produção de células evidenciando o mutualismo que ocorre entre esses seres. Os
Lactobacillus produzem a acidificação do meio, que é condição imprescindível para o
desenvolvimento das leveduras, que por sua vez disponibiliza nutrientes essenciais para as
bactérias (STADIE et al., 2013).
O kefir compete com patógenos pelo sítio de adesão e por nutrientes impedindo o seu
estabelecimento. Essa competição por sítio com a consequente exclusão dos seus concorrentes
pode explicar por que os probióticos devem ter administração contínua com doses significativas
para ser eficaz. Além disso, a sua ação probiótica provoca modulação na resposta imune do
hospedeiro, estimulando a resposta imunitária específica. Os probióticos ativam os macrófagos os
quais acionam a síntese e excreção de citocinas as quais vão causar o aumento da atividade dos
linfócitos T killer (FEDORAK; MADSEN, 2004; RASTALL et al., 2005; SAAD, 2006).
Além de inibir seres microrganismos patogênicos, recentemente foi descoberto que
compostos voláteis do kefir podem contribuir para repelir competidores. De acordo com
Aunsbjerg et al. (2015) a importância dos compostos voláteis tem sido subestimada pois além da
influência sobre o sabor eles ajudam no controle de fungos. Os seguintes compostos voláteis
foram encontrados no kefir: propanol, acetaldeído, acetona, aldeído, butanonas, diacetil (2,3-
Butanodiona), etanol, cetona, pentanol e propionato (Gorner et al., 1972). Otles e Cagindi (2003)
encontraram no kefir, principalmente diacetil (2,3-Butanodiona) e acetaldeido, e Ott et al. (2000)
descreveram que os precursores durante e fermentação do iogurte foram diacetonas, 2,3-
butanodiona (diacetil) e 2,3-pentanediona (2 acetolactato e 2-acetohidroxibutirato).
O objetivo deste estudo foi padronizar a obtenção de biofilmes de kefir em substrato de
açúcar (AB e AM) como meio de cultura, para a formação de biofilmes.

98
2 MATERIAL E MÉTODOS
Os grãos de kefir foram obtidos no Laboratório de Pesquisa em Fármacos, do
Departamento de Ciências Biológicas e da Saúde, da Universidade Federal do Amapá, Amapá,
Brasil.
2.1 Seleção da concentração de grãos de kefir
Todos os recipientes utilizados no processo, foram previamente esterilizados, em
seguida 1000 mL das soluções de AB e AM (40 g/L) foram vertidas nos frascos, e esterilizados a
121°C durante 20 minutos em autoclave (Phoenix, Brasil). Nas soluções, foram adicionados em
câmara de fluxo laminar 0.25, 0.5, 1, 2, 5, 10, 20, 40, 60 e 80 g/L de KG e armazenados a 25°C
durante 20 dias. Após este período, os biofilmes formados foram retirados e pesados. Foram
então aferidos o volume e o pH das soluções do meio.
A concentração de grãos de kefir (KG) selecionada, foi aquela que formou biofilmes
com maior biomassa e consistência. Foi considerado consistente o biofilme que quando retirado
do meio não perdeu a integridade, ou seja, não se fragmentou.
2.2 Determinação da concentração de substrato para a formação de biofilme
Os KG (40 g/L) foram adicionados nas soluções de AB e AM nas seguintes
concentrações: 0.25, 0.5, 1, 2, 5, 10, 20, 40, 60, e 80 g/L. O experimento foi realizado em
triplicata, aferindo o pH do meio no vigésimo dia do cultivo (Modelo 255 HI, Hanna Instruments
EUA).
2.3 Controle da temperatura para produção de biofilme de kefir
Para a padronização da temperatura necessária para produção de biofilme de kefir, o
meio foi cultivado em 15°C, 30°C e 38°C em estufa, e também à temperatura ambiente 25 ± 2°C.
A concentração dos grãos de kefir utilizada foi de 20 g/L, que foi inoculado em solução de AM a
40 g/L. O experimento foi realizado em triplicata, e após 20 dias foram realizadas as mensurações

99
da biomassa.
2.4 Doseamento de macro e microelementos nos grãos de kefir e biofilme por
espectrometria de absorção atômica
Foram utilizadas 100g das amostras de biofilmes e 100g de KG. Para a quantificação dos
macro e microelementos seguiu-se o método descrito por Salazar et al. (2011). As análises foram
realizadas utilizando espectrofotômetro de absorção atômica (AAS Shimadzu modelo 6300).
2.5 Análise dos compostos voláteis do meio de cultura do kefir por cromatografia gasosa
acoplada a espectrometria de massa (CG-EM)
O meio de cultura composto por solução de AM (40 g/L) e grãos de kefir (20 g/L) foi
incubado por 20 dias, filtrado e submetido à hidrodestilação em um aparelho do tipo Clevenger
contendo 1 mL de hexano no tubo coletor. Após 3 horas de hidrodestilação a fase superior
hexânica contendo as substâncias voláteis foi coletada em um frasco de vidro, seca com sulfato
de sódio anidro e analisada por cromatografia em fase gasosa.
A análise por cromatografia em fase gasosa (CG) da solução hexânica foi realizada em
aparelho GC-2010 (SHIMADZU). A amostra injetada na coluna DB-1MS (30 m de
comprimento, 0,25 mm de diâmetro interno e 0,25 μm de espessura do filme), com detecção de
ionização por chama (DIC). As condições da CG foram: hélio como gás de arraste, fluxo com
taxa de 1mL/min e injeção de split com taxa de 1:50 e temperaturas de 270° no injetor, 290°C no
detector e de 60°C a 290°C na coluna, com taxa de aquecimento de 1°C/min.
A solução hexânica das substâncias voláteis foi analisada por cromatografia em fase
gasosa acoplada à espectrometria de massas (CG-EM) em aparelho GCMS-QP2010
(SHIMADZU), utilizando detector por ionização eletrônica. A solução hexânica foi injetada na
coluna ZB-5MS (30m de comprimento, 0,25mm de diâmetro interno e 0,25μm de espessura do
filme), diretamente acoplada ao espectrômetro de massa. As condições da CG foram: hélio como
gás de arraste, fluxo com taxa de 1mL/min e injeção de split com taxa de 1:40. As temperaturas
de análise estão na tabela 3. As condições da EM foram: voltagem de ionização de 70 eV e taxa
de varredura 1scan/s.

100
O cálculo da percentagem de cada componente na amostra foi realizado com base na
área relativa de cada sinal em comparação com a área total no experimento de CG-DIC. A
estrutura química de cada constituinte foi determinada pela comparação dos espectros de massa
com o banco de dados do aparelho (biblioteca NIST) e pela comparação dos índices aritméticos
calculados para cada sinal com os descritos na literatura (ADAMS, 2007). O índice aritmético
foi determinado pela injeção, nas mesmas condições de uma solução contendo uma serie
homóloga de n-alcanos C-C (VAN DEN DOOL & KRATZ, 1963).
2.6 Análise estrutural de biofilmes de kefir por microscopia óptica e por AFM
As microfotografias dos biofilmes foram realizadas utilizando microscópio óptico
(Olympus, EUA) com resolução de 400x, sendo analisadas as características das estruturas
microscópicas externas do biofilme.
A morfologia da superfície dos biofilmes foi analisada por AFM (Easyscan2 AFM/STM,
Nanosurf Co.) mantendo-se a umidade relativa do ambiente em torno de 51%. A varredura
(tamanho de digitalização) foi de 30 mm x 30 mm. As imagens foram obtidas em contact mode.
Foi utilizado cantilever de forma retangular com constante de mola 0,77 nN/m e com força
constante de 0,2 N/m e frequência de ressonância 13 KHz. Para estas medições os biofilmes
foram desidratados e eletrodepositados em lâminas com filmes ouro e após 03/10 ciclos de
digitalização foram analisadas por voltametria cíclica.
Imagens de diferentes regiões dos biofilmes foram examinadas e analisadas no software
AFM para calcular os valores da rugosidade (RMS). Os parâmetros utilizados nesta análise foram
a rugosidade média (Rm) e seu valor eficaz (RMS), calculados usando as seguintes equações
(KHULBE et al., 2008):
Rm =
Onde Rm é a rugosidade média e Z (x, y) é a função da altura da área.
Onde RMS é expresso como a raiz quadrada de rugosidade média.

101
32.7 Análise estrutural de biofilmes de kefir por microscopia eletrônica de varredura
(SEM)
As amostras de biofilme de kefir (secos) foram introduzidas diretamente no deben
acessory do microscópio eletrônico de varredura (TM3030Plus – Tabletop microscope - Hitachi)
e as imagens foram captadas através do software TM3030Plus.
2.8 Análise estatística
Para análise estatística dos resultados foi utilizado o software StatGraphics Centurium v.
XVI (StatEasy Co. MA, USA). Foi empregada ANOVA seguida do teste de Tukey, e as médias
de peso dos biofilmes foram analisadas através do teste t de Student, e resultados com p < 0.05
foram consideradas significativas.

102
3. RESULTADOS
3.1 Seleção da concentração de grãos de kefir
Foram observadas diferenças entre as massas dos biofilmes quando correlacionadas com
as respectivas concentrações de grãos de kefir utilizadas no experimento tanto com uso de
solução de AB como na solução de AM. Houve diferença estatística significante na produção de
biofilme utilizando-se AM como substrato (ANOVA, F = 196,39, p < 0,0001). Foi observado
aumento na produção de biofilme de forma concentração-dependente de KG, atingindo o máximo
em 20 g/L (12,07±0,25 g). Acima desta concentração, não houve diferença estatística entre a
massa de biofilme (Tabela 1).
Semelhantemente houve diferença estatística significante entre a massa de biofilme
formada utilizando-se AB como substrato (ANOVA, F = 269,55, p < 0,0001), e não houve
diferença significativa entre a massa de biofilme produzida nas concentrações acima de 10 g/L
(Tabela 2).
Os biofilmes obtidos com grãos de kefir na concentração de 20 g/L apresentaram
consistência para ambos substratos.
Tabela 1. Formação de biofilmes em diferentes concentrações de grãos de kefir e substratos
(açúcar mascavo e açúcar branco).
No CK
(g/L) Açúcar mascavo Açúcar branco
Peso do Biofilme (g) pH Massa do Biofilme
(g) pH
Média ± DP Média ± DP Média ± DP Média ± DP
1 0.25 1,13±0,14ª 2,88±0,01 1,21±0,35 3,24±0,08
2 0.5 2,25±0,53a 2,87±0,11 2,44±0,27 3,08±0,04
3 1.0 4,65±0,96b 2,80±0,03 5,92±0,45 2,99±0,01
4 2.0 5,60±0,44b 2,64±0 ,05 7,43±0,47 2,84±0,08
5 5.0 8,92±0,45c 2,91±0,03 9,19±0,55 2,71±0,02
6 10.0 9,90±0,65c 2,92±0,01 17,66±0,58x 2,65±0,08

103
7 20.0 12,07±0,25d 2,89±0,03 16,27±0,29x 2,55±0,02
8 40.0 11,84±0,54d 2,79±0,03 16,63±0,73x 2,62±0,02
9 60.0 11,96±0,36d 2,66±0,04 17,59±0,90x 2,60±0,02
10 80.0 11,79±0,48d 2,63±0,02 17,26±1,48x 2,63±0,01
Letras sobrescritas diferentes indicam diferença estatística (Teste de Tukey)
CK, concentração de massa do kefir, n=3
3.2 Determinação da concentração de substrato para a formação de biofilme
Os biofilmes foram formados em diferentes concentrações de AB e AM com alterações
do pH das soluções. Com as diferentes concentrações de AM foram observadas diferenças
significativas entre as massas de biofilmes obtidos (ANOVA, F = 19,78, p < 0,0001). A
concentração necessária de açúcar mascavo para a formação de biofilme foi de 10 g/L (9,64 ±
0,51g). A massa do biofilme obtido nesta concentração não foi significativamente diferente da
dos biofilmes obtidos em concentrações mais elevadas de AM (Tabela 2).
Houve diferenças estatísticas quando utilizou-se as diferentes concentrações de AB
como substrato (ANOVA, F = 40,01 e valor de p < 0,0001), e a melhor concentração para a
formação de biofilme foi a de 10 g/L (16,07 ± 2,54 g) (Tabela 2).
Um dos parâmetros mais importantes a considerar para a formação de biofilmes
consistentes é a concentração do substrato (açúcar). Houve mudança significativa na formação do
biofilme com o aumento da concentração de açúcar tanto AM como AB (Tabela 2).
Table 2. Formação de biofilmes em função da concentração de açúcar branco e mascavo.
No CS
(g/L) Açúcar Macavo Açúcar Branco
Peso do Biofilme (g) pH Peso do Biofilme
(g) pH
Média ± DP Média ± DP Média ± DP Média ± DP
1 0.25 1,06±0,27a 3,28±0,01 1,07±0,33a 3,03±0,04
2 0.5 2,09±0,51b 3,17±0,07 2,51±0,34a 3,10±0,02
3 1.0 4,95±1,04c 3,02±0,03 6,44±1,05b 2,95±0,05
4 2.0 5,95±0,56d 2,93±0,07 8,43±1,09b 2,95±0,06

104
5 5.0 7,67±0,89e 2,91±0,02 9,19±0,45b 2,83±0,03
6 10.0 9,64±0,51f 2,84±0,09 16,07±2,54c 2,88±0,02
7 20.0 11,62±0,92f 2,83±0,03 13,11±2,27c 2,84±0,03
8 40.0 11,69±0,76f 2,72±0,03 16,22±0,99c 2,73±0,04
9 60.0 11,96±0,36f 2,81±0,04 16,89±2,02c 2,66±0,06
10 80.0 11,78±0,45f 2,84±0,08 16,74±1,16c 2,62±0,10
CS concentração do substrato, n=3
Diferentes letras sobrescritas indicam diferença estatística (Teste de Tukey HSD)
Observou-se semelhança no crescimento do biofilme em soluções de AM e AB. No
entanto, a fim de se obter biofilmes consistentes foi necessário 40 g/L de AM enquanto apenas 20
g/L de AB foram necessários. Os biofilmes formados em AB foram mais consistentes o que os
biofilmes formados em AM, e a concentração necessária de grãos de kefir para a formação de
biofilme foi de 20 g/L em ambos os açúcares, e se formaram primeiramente no fundo do frasco e
através do processo de expansão se propagaram para a superfície (Figura 2). A presença de
substâncias aglutinantes mantém a integridade do biofilme, mesmo após a sua remoção do meio,
e são estas substâncias que atuam na formação de estruturas no fundo do frasco (Figura 3).
Tanto nos experimentos para determinação da concentração de grãos de kefir como na
determinação da concentração do substrato para a formação de biofilme, o pH da solução
remanescente foi inferior a 3,30 (Tabela 2 e Tabela 3), após 12 meses verificou-se que
permaneceu inalterado, e consequentemente o meio continuou a apresentar resistência à
contaminação, mesmo depois desse período.

105
Figura 1 - Biofilme de kefir obtido em açúcar mascavo 40g/L, e grãos de kefir na concentração de 20
g/L.
Figura 2 - Biofilme de kefir (20 g/L) em açúcar mascavo (40 g/L). Observa-se os biofilmes formados
no fundo do frasco se expandindo para a superfície.
Figura 3 - Meio de cultura de kefir (20 g/L) em açúcar mascavo (40 g/L). a) Material aglutinado no
fundo do béquer e b) Grãos de kefir adesivados por substância aglutinante.
3.3 Controle da temperatura para produção de biofilme de kefir
A maior formação de b’omassa dos biofilmes foi na temperatura de 30°C, mas os
biofilmes também se formaram à temperatura ambiente (25±2°C). Na temperatura de 16°C não

106
houve formação de biofilmes, mas pôde-se observar fragmentos de biofilmes os quais ficaram
dispersos no meio, e a 38°C não constatou-se formação de biofilmes.
3.4 Doseamento de macro e microelementos nos grãos de kefir e biofilme por
espectrometria de absorção atômica
Os biofilmes formados em solução de AM apresentaram níveis relativamente mais altos
de Fe+2 (2,725 mg/kg) em comparação com os biofilmes formados em AB (1,570 mg/kg) (Tabela
3). O Ca+2, pelo contrário, foi maior nos biofilmes formados em AB (2,150 mg/kg) do que nos
biofilmes formados em AM (0,550 mg/kg). Os seguintes macro e microelementos, também foram
encontrados nos biofilmes: Mg+2 (1,589 mg/kg em AM e 1,862 mg/kg em AB), K+ (2,405 mg/kg
em AM e 2,645 mg/kg em AB) e Zn+2 (0,276 mg/kg para o AM e 0,849 mg/kg em AB).
Tabela 3. Concentração de macro e microelementos presentes em grãos de kefir e em biofilmes e
obtidos em açúcar mascavo e açúcar branco.
Elemento Grãos de kefir
mg/kg Açúcar branco
mg/kg Açúcar mascavo
mg/kg
Ca+2 0.519 2.150 0.550
K+ 2.655 2.645 2.405
Mg+2 1.524 1.862 1.589
Cu+2 0.048 0.095 0.070
Fe+2 2.575 1.570 2.725
Zn+2 0.245 0.849 0.276
Mn+2 0.203 0.955 0.150
3.5 Análise dos compostos voláteis do meio de cultura do kefir por cromatografia gasosa
acoplada a espectrometria de massa (CG-EM)
Os compostos majoritários detectados no meio de cultura foram: ácido hexadecanóico,
hexatriacotano, e tetratetracotano (Tabela 4).

107
Tabela 4. Compostos voláteis majoritários encontrados no meio de cultura do kefir
Composto Volátil Tempo de retenção
(min.)
Área %
Ácido Undecanóico 28,853 0,57
Ácido Tetradecanóico 35,550 1,03
Ácido Hexadecanóico 42,217 28,47
Ácido Hexanedióico 53,875 0,75
Eicosano 54,250 0,61
2-metil-eicosano 59,200 1,20
Hexatriacontano 61,558 1,37
Tetratetracontano 63,858 1,12
Ácido nonanóico 17200 0,38
Butanediol 15,043 0,13
3.6 Análise estrutural de biofilmes de kefir por microscopia óptica e por AFM
Os biofilmes formados em 20 dias utilizando 20 g/L de grãos de kefir e 40 g/L de AM
(Figura 1) foram visualizadas por microscopia óptica a 400× (Figura 3). Observou-se a estrutura
de formação de biofilmes e a uniformidade com ausência de porosidade. As microfotografias dos
biofilmes mostraram que as estruturas se fundiram formando os biofilmes, e a análise por AFM,
mostrou picos relacionados as variações na rugosidade da superfície dos biofilmes (Figuras 4 e
5).
Na análise do biofilme obtidos com diferentes concentrações de AM por AFM, revelou
diferenças nos parâmetros de rugosidade (RMS) com concentrações crescentes do açúcar (Figura
4). A rugosidade dos biofilmes cresceu com o aumento da concentração de grãos de kefir. Na
concentração de 40 g/L a característica reológica rugosidade foi a melhor visualizada. As
imagens de AFM mostraram que a rugosidade da superfície do biofilme apresentou correlação
com a alteração da concentração de grãos de kefir (Figuras 4). Constata-se o aumento de
rugosidade (z= 410 µm, 510 µm e 563 µm, respectivamente) à proporção que se aumenta a
concentração de grãos de kefir no meio.

108
Figura 4 - Foto de biofilme de kefir (20g/L) com açúcar mascavo (40 g/L) através de microscopia óptica
com resolução 400x mostrando a estrutura de formação de biofilmes e a ausência de porosidade do
biofilme.
Figura 5 - Imagem topográfica de biofilmes de kefir em açúcar mascavo (40 g/L) através de microscopia
de força atômica (AFM). Concentração de GK a) 20 g/Lb) 60 g/L e c) 80 gl1
Rugosidade
Rugosidade

109
Figura 6 - Imagem topográfica de biofilme de kefir com açúcar mascavo obtida através de microscopia de
varredura de força (AFM). Concentração AM 40 g/L e GK 20 g/L. Observa-se uniformidade do tecido do
biofilme e a rugosidade.
3.7 Análise estrutural de biofilmes de kefir por microscopia eletrônica de varredura (SEM)
Na análise dos biofilmes por SEM, observou-se várias camadas sobrepostas em imagem
de perfil (Figura 6) e a composição de microrganismos, com coagregação de bactérias e fungos
filamentosos (Figura 7).
Figura 7 - Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (20 g/L) em açúcar
mascavo (40 g/L) visualizado de perfil onde se observa as camadas de exopolissacarídeo que se
sobrepõem formando o biofilme. Dados de magnificação são mostrados na figura.
Figura 8 - Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (20 g/L) em açúcar
mascavo (40 g/L) onde se observa as bactérias adesivada às hifas de fungos. Magnificação de a) 800x
b)2000x e c) 3000x.

110
4 DISCUSSÃO
O experimento com açúcar branco, ou seja, industrial se restringiu a se demostrar a
possibilidade de se cultivar kefir e formar biofilmes com o mesmo. Uma vez que comumente se
utiliza açúcar mascavo no cultivo do kefir os experimentos seguintes se concentraram apenas em
resultados obtidos com a utilização de açúcar mascavo.
Geralmente se pensa que o açúcar branco é inapropriado para se cultivar o kefir por
conter alguma possível substância de uso industrial que lhe limitaria o crescimento e impediria o
processo de fermentação. Também se acredita que como o açúcar mascavo é mais rico em
nutrientes é mais viável para cultivo do kefir, pois o meio de cultura com açúcar branco se torna
pobre nutricionalmente devido ao processo industrial ao que o açúcar é submetido que lhe
destitui de minerais e compostos encontrados no açúcar mascavo. Com efeito, Laurey e De Vust
(2014) afirmam que o alto teor de açúcar e a baixa concentração de aminoácidos encontrados no
kefir de água deveria resultar em um habitat em carência, no entanto o que se obtém é uma
bebida fermentada com uma comunidade microbiana bastante diversificada em equilíbrio e
estabilidade (LAUREYS; DE VUYST, 2014) , isto provavelmente decorre do fato que,
conforme Stadie et al. (2013) asseveram, no kefir metabolizado em água ocorre s sinergia entre
leveduras e bactéria ácido-lácticas notadamente com lactobacilos (STADIE et al., 2013).
As leveduras fornecem os aminoácido e vitaminas ao kefir. Por isso mesmo um meio
pobre em nutriente não consiste em obstáculo para desenvolvimento do mesmo.
Os parâmetros mais importantes para formação de biofilmes de kefir foram a
concentração do inóculo, a temperatura e a concentração de substrato. De acordo com Heydorn et
al. (2000) e Stoodley et al. (2002) o aumento de células em um biofilme faz parte de seu próprio
processo de maturação, podendo acontecer até 10 dias após a adesão inicial. Os biofilmes de kefir
se formaram após 5 dias, mas apenas com 20 dias apresentaram-se consistentes. O aumento de
células ocorre em decorrência da divisão celular e da coadesão de outras células presentes no
sistema em seu estado planctônico por isso a concentração do inoculo é muito importante. A
concentração necessária de grãos de kefir para a formação de biofilmes foi de 20 g/L.
Quando se utilizou quantidade de inoculo insuficiente (abaixo de 20 g/L) ocorreu a

111
formação apenas de fragmentos de biofilmes, tanto com o uso de solução de AB como de AM.
Em relação à variação de concentração de grão de kefir para a preparação do kefir,
Garrote et al. (2001) demonstraram que 10 g/L de grãos de kefir resulta em um produto viscoso e
não muito ácido e a concentração recomendada de grãos de kefir utilizados para obter uma bebida
ácida com baixa viscosidade e alta efervescência é de 100 g/L. Para Sarkar (2008) 5% de grãos
de kefir resultou na produção de etanol e ácidos voláteis. Em AM, 20 g/L de grãos de kefir
mostrou ser o ideal para a formação de biofilmes.
A temperatura ideal para a formação de biofilmes foi de 30°C, mas os biofilmes também
se formaram à temperatura ambiente (25 ± 2°C). Na temperatura de 38°C não se formaram os
biofilmes enquanto que à temperatura de 16°C apenas fragmentos de biofilmes se formaram.
Asadishad et al. (2014) observaram que temperaturas abaixo de 10°C paralisam a produção de
biofilmes por Bacillus subtilis. Já os biofilmes formados por Listeria sp são mais resistentes na
temperatura de 15°C do que a 37°C (SHIMAMURA et al., 2015).
Para Perni et al. (2006) um dos parâmetros mais importante para a formação de biofilme
é a concentração do substrato. Apesar da concentração de 10 g/L ter sido estatisticamente
considerada a ideal para formação de biofilmes, mas a consistência somente foi alcançada quando
se fez uso de 20 g/L para AB e 40 g/L para o AM.
A concentração de açúcar é importante, pois fornece os hidrogênios necessários para a
formação de pontes de hidrogênio que resultará em biofilmes consistentes. De acordo com
Chowdhury et al. (2011) a presença de numerosos grupos hidroxilas na estrutura que é
característica comum dos açúcares, viabilizam que ocorram as ligações de pontes de hidrogênio
intermolecularmente em grande quantidade. Em concentrações de substrato abaixo de 10 g/L
ocorre a formação apenas de fragmentos de biofilmes e em concentração acima de 10 g/L mas
abaixo de 40 g/L para o AM o biofilme se forma, mas, sem consistência e se fragmenta
facilmente.
Há três tipos de forças envolvidas no processo de formação de biofilmes, as interações
eletrostáticas, as pontes de hidrogênio e as forças de dispersão de London (MAYER et al., 1999).
Estas forças associadas contribuem para a estabilidade da matriz do biofilme (FLEMMING;
WINGENDER, 2001b). Além dessas forças atuarem na formação dos biofilmes ocorre
concomitantemente a ação de substâncias aglutinantes que unem os fragmentos do biofilme.
Mesmo após a ebulição do meio a temperatura de 100°C estas substâncias ainda atuam fazendo a

112
junção dos fragmentos dos biofilmes.
O crescimento do biofilme resulta de um processo complexo que envolve transporte de
moléculas orgânicas e inorgânicas e células microbianas para a superfície e subsequente adsorção
a superfície e finalmente adesão definitiva, corroborada pela produção de exopolissacarídeos
(SPE) (FLEMMING, 1993; WATERS; BASSLER, 2005; RUIZ et al., 2004). Outro fator que
contribui para a variação da capacidade de adesão do biofilme é a diferente composição do
biofilme que influencia a extensão mediante a qual o microrganismo possa aderir tanto a
superfícies hidrofílicas como a hidrofóbicas (VAN HULLEBUSCH, 2003; ROMANI et al.,
2008).
O AB formou os biofilmes utilizando concentração menor (20 g/L) do que a do AM (40
g/L), provavelmente devido ao fato de que o mesmo contém mais sacarose que o açúcar mascavo.
A consistência dos biofilmes aumenta ao transcorrer do tempo. A formação de biofilmes
iniciou-se apenas após cinco dias da inoculação, isto provavelmente ocorreu porque os
microrganismos iniciam os processos que levam à formação de biofilme somente após três dias
após a inoculação, quando também o pH decai através da ação de bactérias ácido lácticas (por
exemplo, Leuconostoc spp.). Por isso foi possível observar a formação inicial de estruturas de
biofilme somente com 5 dias.
Tanto com o aumento da concentração de grãos de kefir, como do substrato para a
formação de biofilme, o pH da solução remanescente decai para menos de 3,0 (Tabelas 1 e 2). Os
grãos de kefir contêm várias espécies de Lactobacillus, Lactococcos, Leuconostoc além de
bactérias secretoras de ácido acético e leveduras. Estes microrganismos secretam ácido láctico,
antibióticos e bactericidas que inibem o crescimento de microrganismos indesejáveis e
patogênicos (ABE et al., 2013; LEITE et al., 2013; LENGKEY et al., 2013). Uma das causas de
resistência ao crescimento de outros microrganismos é a produção de ácidos com a consequente
queda do pH (Tabela 1 e 2). De acordo com Abe et al. (2013) o pH baixo indica resistência à
contaminação, e para Mangia et al. (2015) indica forte fermentação. À medida que aumentou-se a
concentração de grãos de kefir o pH do meio decaiu até 2,8 (Tabela 2). O pH do meio caiu tanto
em solução de AM, como em solução de AB. O aumento da concentração do substrato (AM) foi
acompanhado por decaimento do pH, bem mais acentuado do que com a variação de
concentração de KG, o que sugere o aumento da atividade dos microrganismos sobre o substrato,
resultando em produtos finais que tornam o meio acidificado como os ácidos lático e acético. O

113
meio de cultura com kefir após 18 meses permaneceu com o pH inalterado (2,81) e isento de
contaminantes.
Wang et al. (2010) relataram um exopolissacarídeo de Lactobacillus plantarum
composto por manose, glicose e galactose com capacidade de formar biofilmes com o tempo de
30 horas em pH 6,5. Para Lengkey et al. (2013) o aumento da concentração de KG de 5% a 25%
causou decaimento do pH, e para Irygoyen (2005) o kefir fez baixar o pH do leite até 4,0. O pH
do meio com kefir se estabilizou abaixo de 3,0 indicando forte fermentação.
Os macroelementos que foram previamente descritos em kefir por Liut Kevicius e
Sarkinas (2004) foram K+, Ca+2, e Mg+2, enquanto os microelementos foram Cu+2, Zn+2, Fe+2,
Mn+2 e Co+2. Relataram também grande quantidade de Ca+2 e de Mg+2 em kefir.
No biofilme de kefir formado em meio com AB foi encontrado Ca+2, mais do que no
formado em meio com AM e nos KG, provavelmente devido ao tratamento químico que o AB é
submetido industrialmente. Já o Fe+2 foi encontrado tanto nos KG, como nos biofilmes formados
em meio com AM em quantidade maior do que nos biofilmes formados em meio com AB. O
Zn+2 foi encontrado em maior concentração nos biofilmes formados com AB do que nos
biofilmes formados com AM (Tabela 3).
Os biofilmes podem utilizar a capacidade de absorção de certos metais em forma iônica
como Zn+2, Cu+2, Fe+2 e Al+3 contidos em sua matriz para evitar erosão por forças (GRUMBEIN
et al., 2014). Alguns desses íons metálicos são tóxicos para as formas planctônicas das bactérias
componentes do biofilme, mas eles têm sua toxicidade suprimida na matriz dos biofilmes
(GRUMBEIN et al., 2014), o que ratifica a viabilidade de biofilmes para biorremediação. Assim
além dos biofilmes exercerem a regulação das propriedades mecânicas eles formam uma barreira
seletiva para conter substâncias tóxicas (GRUMBEIN et al., 2014).
Os principais compostos voláteis encontrados no meio contendo kefir por Beshkova et
al. (2003) foram acetaldeido, acetona, etil-acetato, 2-butanona, diacetil (2,3-Butanodiona) e
etanol. Os compostos voláteis podem variar em quantidade dependendo do substrato, Cais-
Sokolinska et al. (2015) observaram que em leite de cabra a presença de ácidos graxos poli-
insaturados e o cultivo do kefir influenciou na produção de 2,3-butanediona tendo havido
também aumento de ácido lático e etanol. Mangia et al. (2014) encontraram os seguintes
compostos voláteis em Morocan, uma bebida fermentada de leite, etanol, acetaldeido, diacetil
(2,3-Butanodiona) e acetoina. Enquanto que os principais compostos voláteis encontrados no

114
iogurte foram ácido lático, acetaldeído, diacetil (2,3-Butanodiona), acetoina, acetona, e 2-
butanona (CHENG, 2010), os quais contribuem para o aroma peculiar do iogurte bem como para
seu sabor. Os principais compostos voláteis encontrados no meio contendo kefir neste estudo
foram propanol, eicosano, ácido acético e pentanol.
A microscopia de força atômica (AFM) pode ser usada para avaliar a progressão
morfológica dos biofilmes (PEREIRA et al., 2015), bem como proporcionar o entendimento do
mecanismo biofísico da organização estrutural das cápsulas de polissacarídeos na formação de
biofilmes (WANG et al., 2015). O parâmetro estudado através desta metodologia neste estudo foi
a rugosidade. Os picos no AFM mostram variações na rugosidade da superfície dos biofilmes.
Nos biofilmes formados utilizando diferentes concentrações de AM (Figura 3), revelou
diferenças nos parâmetros de rugosidade (RMS) com concentrações crescentes do açúcar (Figura
3 e 4). A rugosidade dos biofilmes cresceu com o aumento da concentração de KG. O aumento da
rugosidade causa aumento na capacidade de incorporar substancias no biofilme o que pode ser
usado tanto para substâncias deletérias à saúde como metais pesados (biorremediação), como
para substâncias com propriedades terapêuticas.
A microscopia eletrônica de varredura tem sido utilizada para estudos da estrutura de
biofilmes (FERREIRA et al., 2015; LAVIN et al., 2015; SOUSA et al., 2015). A análise do perfil
dos biofilmes por SEM, mostrou a sobreposição de estruturas na formação de biofilmes (Figura
5), e a coadesão de bactérias com hifas de fungos (Figura 6), confirmando os estudos de Stadie et
al. (2013) sobre a interação simbiótica entre ambos.
Os biofilmes formados principalmente na parte inferior do frasco se expandem para a
superfície (Figura 7). A presença de substâncias aglutinantes mantém a integridade do biofilme,
mesmo após a sua remoção do meio e são estas substâncias que atuam na formação de estruturas
inicialmente no fundo do frasco (Figura 8).
Biofilmes de kefir são descritos apenas como constituintes dos KG (LOPTIZ-OTSOA et
al., 2006). Não existem relatos anteriores sobre a formação de biofilme extracelular de kefir na
superfície do meio de cultura, no entanto Furukawa et al. (2011) relataram a descoberta de
biofilmes de Lactobacillus plantarum e Saccharomyces cerevisiae em consórcio com outras
leveduras, enquanto Dertli et al. (2013) relataram dois exopolissacárideos secretados por
Lactobacillus johnsonii e Wang et al. (2010) relataram um exopolissacarídeo de Lactobacillus
plantarum composto por manose, glicose e galactose com capacidade de formar biofilmes.

115
A formação de biofilme é um processo que ocorre de forma ubíqua (SINGH et al.,
2006). A preparação do kefir para formação de biofilmes constitui um processo relativamente
fácil, já que não são necessários dispositivos especiais, entretanto, apesar de que o kefir inibe o
crescimento de contaminantes, deve-se ter preocupações em relação a contaminação.

116
5 CONCLUSÃO
As técnicas aplicadas permitiram a padronização do processo de obtenção e
caracterização do biofilme de kefir. Observou-se que a consistência do biofilme é dependente do
tempo de cultivo e a temperatura ótima é de 30oC. A concentração de KG determinada para a
formação de biofilmes foi de 20g/L para os substratos AB e AM, sendo que a concentração de
AB foi menor que do AM. Os compostos voláteis majoritários presentes no meio de cultivo do
kefir são das classes dos ácidos graxos e hidrocarbonetos alifáticos.
A partir das análises dos micro e macroelementos encontrados nos KG e nos biofilmes,
pode-se inferir que tanto os KG como os biofilmes incorporam esses elementos, e através da
análise por AFM observou-se que a rugosidade do biofilme é dependente da concentração de GK,
e por SEM foi identificada a coadesão das bactérias aos fungos filamentosos, demonstrando a
importância dessa interação para o desenvolvimento dos biofilmes.
AGRADECIMENTOS
Os autores agradecem a CAPES, pelo auxílio à pesquisa do Programa Pró-Amazônia - CAPES -
nº 3292/2013 - AUXPE, e CNPq - Proc. 402332/2013-0 – Biotec, pelo suporte financeiro à
pesquisa.

117
REFERÊNCIAS
ABE, A.; FURUKAWA, S.; WATANABE, S.; MORINAGA, Y. Yeasts and lactic acid bacteria
mixed-specie biofilm formation is a promising cell immobilization technology for ethanol
fermentation. Appl Biochem Biotechnol., v. 171, n. 1, p. 72-79, 2013.
ADAMS, R.P. (2007): Identification of essential oils components by gas chromatography/ mass
spectroscopy. Allured Publishing Corporation. Carol Stream, IL, USA.
ASADISHAD, B.; OLSSON, A. L.; DUSANE, D. H.; GHOSHAL, S.; TUFENKJI, N. Transport,
motility, biofilm forming potential and survival of Bacillus subtilis exposed to cold temperature
and freeze-thaw. Water Res., v. 58, p. 239-247, 2014.
AUNSBJERG, S. D.; HONORÉ, A. H.; MARCUSSEN, J.; EBRAHIMI, P.; VOGENSEN, F. K.;
BENFELDT, C.; SKOV, T.; KNØCHEL, Contribution of volatiles to the antifungal effect of
Lactobacillus paracasei in defined medium and yogurt. Int J Food Microbiol., v. 194, p. 46-53,
2015.
BESHKOVA, D. M.; SIMOVA, E. D.; FRENGOVA, G. I., SIMOV, Z. I.; DIMITROV, Z. P.
Production of volatile aroma compounds by kefir starter cultures. International Dairy Journal,
v. 13, p. 529-535, 2003.
CERQUEIRA, L.; OLIVEIRA, J. A.; NICOLAU, A.; AZEVEDO, N. F., VIEIRA, M. J.
Biofilm formation with7 mixed cultures of Pseudomonas aeruginosa/Escherichia coli on silicone
using artificial urine to mimic urinary catheters. Biofouling, v. 29, p. 829-840, 2013.
CHOWDHURY, S. R.; BASAKA, R. K.; SENB, R; ADHIKARI, B. Characterization and
emulsifying property of a carbohydrate polymer produced by Bacillus pumilus UW-02 isolated
from waste water irrigated agricultural soil. International Journal of Biological
Macromolecules, v. 48, p. 705–712, 2011.
CHENG, H. Volatile flavor compounds in yogurt: a review. Crit Rev Food Sci Nutr, v. 50, p.
938-950, 2010.
COSTERTON, J. W.; STEWART, S.; GREENBERG, E. P. Bacterial biofilms: A common cause
of persistent infections. Science v. 284, p. 1318-1322, 1999.
FEDORAK, R. N.; MADSEN, K. L. Probiotics and prebiotics in gastrointestinal disorders.
Current Opinion Gastroenterology v. 20, p. 146-155, 2004.
FLEMMING, H. C.; WINGENDER, J. Relevance of microbial extracellular polymeric
substances (EPSs) – Part I: Structural and ecological aspects. Water Sci. Technol. v. 43, p.1-8,
2001.

118
FLEMMING, H. C.; WINGENDER, J. Relevance of microbial extracellular polymeric
substances (EPSs) – Part II: Technical aspects. Water Sci. Technol. v. 43, p. 9-16, 2001.
FURUKAWA, S.; NOJIMA, N.; YOSHIDA, K.; HIRAYAMA, S.; OGIHARA, H.;
MORINAGA, Y. The importance of inter-species cell-cell co-aggregation between Lactobacillus
plantarum ML11-11 and Saccharomyces cerevisiae BY4741 in mixed-species biofilm formation.
Biosci Biotechnol Biochem v. 75, p.1430-1434, 2011.
GARROTE, G. L.; ABRAHAM, A. G.; De ANTONI, G. L. Chemical and microbiological
characterization of kefir grains. Journal of Dairy Research v. 68, p. 639-652, 2001.
GORBUSHINA, A. A., BROUGHTON, W. J. Microbiology of the atmosphere-rock interface.
how biological interactions and physical stresses modulate a sophisticated microbial ecosystem.
Annu. Rev. Microbiol. v. 63, p. 431–450, 2009.
GRUMBEIN, S.; OPITZ, M.; LIELEG, O. Selected metal ions protect Bacillus subtilis biofilms
from erosion. Metallomics, v. 6, p. 1441-1450, 2014.
HEYDORN, A.; NIELSEN, A.T.; HENTZER, M.; STERNBERG, C.; GIVSKOV, M.
Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology,
v.146, p.395–407, 2000.
KOJIĆ, M.; LOZO, J.; BEGOVIĆ, J.; JOVČIĆ, B.; TOPISIROVIĆ, L. J. Characterization of
Lactococci isolated from homemade kefir. Arch Biol Sci, v. 59, p. 13-22, 2007.
KHULBE, K.C.; FENG, C.; MATSUURA, T. Synthetic Polymeric Membranes:
Characterization by Atomic Force Microscopy, 1st ed., Springer, 2008.
LA RIVIERE, J. W. M.; KOOIMAN, P.; SCHMIDT, K. Kefiran, a novel polysaccharide
produced in the kefir grain by Lactobacillus brevis. Arch. Microbiol., v. 59, p. 269-278, 1967.
LAUREYS, D.; DE VUYST, L. Microbial species diversity, community dynamics, and
metabolite kinetics of water kefir fermentation. Appl Environ Microbiol v. 80, p. 2564-72,
2014.
LEITE, A.M.; LEITE, D.C.; DEL AGUILA, E.M.; ALVARES, T.S.; PEIXOTO, R.S.; MIGUEL,
M. A.; SILVA, J.T.; PASCHOALIN, V.M. Microbiological and chemical characteristics of
Brazilian kefir during fermentation and storage processes. J Dairy Sci, v. 96, p. 4149-4159,
2013.
LENGKEY, H. A. W.; SIWI, J. A.; BALIA, R. L. The effect of various starter dosages on kefir
quality. Lucrări Ştiinţifice-Seria Zootehnie, v. 59, p. 113-116, 2013.
LIUT KEVICIUS, J. V. A.; SARKINAS, A. Studies on the growth conditions and composition of
kefir grains – as a food and forage biomass. Dairy Science Abstracts, v. 66, p. 903, 2004.
LOPITZ-OTSOA, F.; REMENTERIA, A; ELGUEZABAL, N.; Garaizar, J. Kefir: A symbiotic
yeasts-bacteria community with alleged healthy capabilities. Rev Iberoam Micol. v. 23, p. 67-

119
74, 2006.
MANGIA, N. P.; GARAU, G.; MURGIA, M. A.; BENNANI, A. ;DEIANA, P. Influence of
autochthonous lactic acid bacteri and enzymatic yeast extracts on the microbiological,
biochemical and sensorial properties of Lben generic products. J Dairy Res, v. 81, p. 193-201,
2014.
MAYER, C.; MORITZ, R.; KIRSCHNER, C.; BORCHARD, W.; MAIBAUM, R.;
WINGENDER, J.; FLEMMING, H.-C. The role of intermolecular interactions: studies on model
systems for bacterial biofilms. Int. J. Biol. Macromol., v. 26, p. 3-16, 1999.
OTLES, S.; CAGINDI, O. Kefir: A Probiotic Dairy-Composition, Nutritional and Therapeutic
Aspects Pakistan Journal of Nutrition v. 2 p. 54-59, 2003
PERNI, S.; JORDAN, S. J.; ANDREW, P. W.; SHAMA, G. Biofilm development by Listeria
innocua in turbulent flow regimes. Food Control, v.1 n 7, p. 875–883, 2006.
PEREIRA, F. D.; BONATTO, C. C.; LOPES, C. A.; PEREIRA, A. L.; SILVA, L. P. Use of
MALDI-TOF mass spectrometry to analyze the molecular profile of Pseudomonas aeruginosa
biofilms grown on glass and plastic surfaces. Microb Pathog, v. 86, p. 32-37, 2015.
RAJESH, P.S.; RAI, V.R. Use of aiiA gene amplification for AHL-lactonase production from
endophytic bacterium Enterobacter species. Int J Biol Macromol, v. 72, p. 1009 -1013, 2015.
RASTALL, R. A.; GIBSON, G. R.; GILL, H. S. GUARNER, F.; KLAENHAMMER T. R.; POT,
B.; Reid, G.; Rowland, I. R.; Sanders, M. E.; Modulation of the microbial ecology of the human
colon by probiotics, prebiotics and symbiotic to enhance human health: An overview of enabling
science and potential applications. FEMS Microbiology Ecology, v. 52, p. 145-152, 2005.
ROMANI, A. M.; FUND, K.; ARTIGAS, J.; SCHWARTZ, T.; SABATER, S.; OBST, U.
Relevance of polymeric matrix enzymes during biofilm formation. Microb. Ecol., v. 56, p. 427-
436, 2008.
SAAD, S. M. I. Probióticos e prebióticos: o estado da arte. Revista Brasileira de Ciências
Farmacêuticas, v. 42, p. 1-16, 2006.
SALAZAR, R.F.S.; ALCÂNTARA, M. A. K; IZÁRIO FILHO, H. J. Evaluation of Sample
Preparation of Nickel. R. Bras. Ci.Solo, v. 35, p. 241-248, 2011.
SARKAR, S. Biotechnological innovations in kefir production: a review. British Food Journal,
v. 110, p. 283-295, 2008.
SCARDINO, A. J.; DE NYS R. (2011) Mini review: Biomimetic models and bioinspired
surfaces for fouling control. Biofouling v. 27, p. 73-86, 2011.
SHIMAMURA, N.; IRIE, F.; YAMAKAWA, T.; KIKUCHI, M.; SAWAI, J.; HEATED
SCALLOP-SHELL. Powder Treatment for Deactivation and Removal of Listeria sp. Biofilm

120
Formed at a Low Temperature Biocontrol. Science, v.20, p. 153-157, 2015.
SINGH, R.; PAUL, D.; JAIN, R. K. Biofilms: implications in bioremediation. Trends Microbiol
v. 14, p. 389-397, 2006.
STADIE J.; GULITZ A.; EHRMANN M. A.; VOGEL R. F. Metabolic activity and symbiotic
interactions of lactic acid bacteria and yeasts isolated from water kefir. Food Microbiol, v. 35, p.
92-98, 2013.
STATGRAPHICS CENTURION XVI SOFTWARE (Statpoint Technologies, Inc, Warrenton, VA,
USA).
STOODLEY, P.; SAUER, K.; DAVIES, D. G.; COSTERTON, J. W. Biofilms as complex
differentiated communities. Annual Review Microbiology, v. 56, p. 187-209, 2002.
SUTHERLAND, I.W. Biofilm exopolysaccharides: a strong and sticky framework.
Microbiology, v. 147, p. 3-9, 2001.
VAN HULLEBUSCH, E. D.; ZANDVOORT, M. H.; LENS, N. L. Metal immobilisation by
biofilms: mechanisms and analytical tools. Rev. Environ. Sci. Biotechnol, v. 2, p. 9-33, 2003.
VU, B.; CHEN, M.; CRAWFORD, R. J.; IVANOVA, E. P. Bacterial Extracellular
Polysaccharides Involved in Biofilm Formation. Molecules, v.14, p.2535-2554, 2009.
WANG, Y; LI, C; LIU, P; AHMED, Z; XIAO, P; BAI, X. Physical characterization of
exopolysaccharide produced by Lactobacillus plantarum KF5 isolated from Tibet Kefir.
Carbohydr. Polym., v. 82, p. 895–903, 2010.
WATERS, C.M.; BASSLER, B.L. Quorum sensing: cell-to-cell communication in bacteria.
Annu. Rev. Cell Dev. Biol., v. 21, p. 319-346, 2005.
VAN DEN DOOL, H. & KRATZ, P.D. A generalization of the retention index system including
linear temperature programmed gas-liquid partition chromatography. Journal of Chromatography.
v. 11, p. 463-471, 1963.

121
CAPÍTULO 2: ESTUDO DO BIOFILME DE KEFIR ASSOCIADO AO
EXTRATO HIDROETANOLICO DE Euterpe oleracea Mart. (AÇAÍ). UM
POSSÍVEL PROBIÓTICO AMAZÔNICO

122
Estudo de biofilme de Kefir associado ao extrato hidroetanólico de Euterpe oleracea Mart.
(açaí). Um possível probiótico da Amazônia
Antonio Ferreira de Oliveira1,2, Larissa Daniele Machado Goés2, Fernanda Borges de
Almeida2,3, Daniele da Cruz de Assis2, Adriana Ferreira Maciel2, Jonatas Lobato Duarte2,3,
Caio Pinho Fernandes1,3, Roberto Messias Bezerra2, Robert Saraiva de Matos4, Robert
Magña Zamora2, José Carlos Tavares Carvalho1,2*
1Programa de Pós-graduação em Biodiversidade Tropical, Departamento de Ciências Biológicas e da Saúde,
Universidade Federal do Amapá 2Laboratório de Pesquisa em Fármacos, Curso de Farmácia, Departamento de Ciências Biológicas e da Saúde,
Universidade Federal do Amapá 3Laboratório de Nanobiotecnologia Fitofarmacêutica, Curso de Farmácia, Departamento de Ciências Biológicas e
da Saúde, Universidade Federal do Amapá 4Programa de Pós-graduação em Ciências Farmacêuticas, Departamento de Ciências Biológicas e da Saúde,
Universidade Federal do Amapá
*Corresponding author:[email protected]
RESUMO
Kefir é um complexo microbiano capaz de produzir biofilmes. O objetivo deste estudo foi obter
biofilmes associados ao extrato hidroetanólico de Euterpe oleracea Mart. (BEHEEo) como
método de recuperação de antocianinas em biofilme de kefir. Assim, cinética de formação do
BEHEEo foi efetivada, em diferentes concentrações do extrato aquoso de Euterpe oleracea Mart.
(EHEEo), de grãos de kefir (GK) e de açúcar mascavo (AM); a determinação da dosagem de
macro e microelementos de BEHEEo, a análise das características reológicas e estruturais por
microscopia de força atómica (AFM), e antocianinas em BEHEEo. A melhor concentração de
AM para formar BEHEEo foi de 40 g/L com 100 ml de EHEEo (12,62 ± 0,1626 g). A liberação
das antocianinas do BEHEEo ocorreu a partir de 5 minutos (4,3 ± 0,6 mg/100 g) com pico
máximo em 60 minutos (20,8 ± 0,3). A concentração de Mg++, Fe++ e Ca++ foi de 0,1808 mg/kg,
0,1740 mg/kg e 0.089 mg/kg, respectivamente, enquanto a concentração de Zn++ foi 0,533 mg/kg.
A análise AFM revelou diferenças de rugosidade dependente da concentração de GK, pode-se
observar a presença de Lactobacillus e leveduras. A concentração de antocianinas em BEHEEo
de 24 meses foi de 26 mg/100g. Portanto, infere-se que o BEHEEo incorpora as antocianinas
EHEEo e tem potencial para aplicações terapêuticas em várias patologias, em que é necessário o
processo antioxidante.
Palavras-chave: Biofilme. Kefir. Açaí. Euterpe oleracea Mart. Antocianinas, AFM.
IMPORTANTE : NÃO SE COLOCA ABREVIAÇÃO MESMO EM RESUMO???

123
ABSTRACT
Kefir is a microbial complex capable of producing biofilms. The objective of this study was to
obtain biofilms associated with hydroethanolic extract of Euterpe oleracea Mart. (BEHEEo) as
fixing method of recovery anthocyanins in kefir biofilm. Thus, BEHEEo formation kinetics was
made, at different concentrations of aqueous extract of Euterpe oleracea Mart. (EHEEo), of kefir
grains (GK) and of brown sugar (BS); the determination of macro and microelements of
BEHEEo, the analysis of the rheological and structural characteristics by atomic force
microscopy (AFM), and anthocyanins dosage in BEHEEo. The best concentration of BS to form
BEHEEo was 40 g/L with 100 ml of EHEEo (12.62 ± 0.1626 g). The release of the BEHEEo
anthocyanins occurred from 5 minutes (4.3 ± 0.6 mg/100 g) with maximum peak at 60 minutes
(20.8 ± 0.3). The concentration of magnesium, iron and calcium was 0.1808 mg/kg, 0.1740
mg/kg and 0.089 mg/kg, respectively, indicating the concentration of Zn++ which was 0.533
mg/kg. The AFM analysis revealed diferences of roughness dependent on the concentration of
GK, with presence of Lactobacillus and yeast. The concentration of anthocyanins in BEHEEo
two years after the acquisition was 26 mg/100g. Therefore, it is suggested that the BEHEEo
incorporated the EHEEo anthocyanins and has potential for therapeutic applications in several
pathologies wherein it is necessary the antioxidative processes.
Key words: Biofilm. Kefir. Açaí. Euterpe oleracea Mart.. Anthocyanins. AFM.

124
1 INTRODUÇÃO
O kefir é um soro é produzido a partir da fermentação de bebidas fermentadas por grãos
de kefir (LAUREYS; DE VUYS, 2014). O crescimento e a função de bactérias no interior de uma
população são cruciais para a sua sobrevivência (DAVEY; O'TOOLE, 2000). No kefir há uma
predominância de Lactobacillus os quais têm alta capacidade catabólica e são complementadas
por leveduras com alta capacidade biossintética.
Ao se cultivar o kefir em água com açúcar durante vinte dias, ocorre a formação de
biofilmes com possíveis propriedades terapêuticas contra organismos patogénicos.
Biofilmes se formam em superfícies e contêm elevado nível de organização estrutural,
sendo, em geral, associados com colónias de bactérias mantidas juntas por um polímero secretado
pelas suas células, que é chamado substância polimérica extracelular (SPE) (WATNICK;
KOLTER, 2000; DONLAN, 2002, ZHANG et al., 2015).
Os biofilmes conferem resistência ao estresse, proteção contra impactos químicos e
mecânicos e fornecem nutrientes, por isso eles são essenciais para a sobrevivência de
microrganismos (FLEMMING; WINGENDER, 2001a; SUTHERLAND, 2001; VAN
HULLEBUSCH, 2003; VU et al., 2009). Biofilmes repelem microrganismos competidores, por
outro lado têm capacidade de absorver substâncias. Estas substâncias podem ser metais pesados
por isso biofilmes podem ser utilizado em biorremediação. No entanto, os filmes também podem
absorver substâncias terapêuticas, e têm sido demonstrado por Rodrigues et al. (2005) que o kefir
pode ser aplicado no tratamento de feridas e queimaduras entre outras aplicações.
Dentre as muitas estratégias utilizadas para o estudo de biofilmes as que têm sido mais
realçadas são utilização de microscopia de força atómica o (AFM) pois permite medir as forças
de interação entre as células e as interações célula-célula (DUFRÊNE, 2015).
O microscópio eletrônico de varredura (MEV) é um equipamento capaz de produzir
imagens de alta ampliação (até 300.000x), sendo utilizado para estudos de estrutura dos
biofilmes, para avaliar a presença de microrganismos aderidos à superfície e estudar a capacidade
de adesão e formação de biofilmes (FERREIRA et al., 2015; LAVIN et al., 2015; GRANILLO et
al., 2015; SOUSA et al., 2015).

125
Uma das substâncias cujo estudo tem sido intensificado nos últimos anos são as
antocianinas, devido às suas propriedades de proteção e cura. As antocianinas têm um papel
importante em funções fisiológicas relacionadas com a saúde humana (LEE et al., 2013).
Estudos demonstraram a ação das antocianinas em câncer através da inibição de
metástase e angiogénese, além de reconstituir a matriz extracelular (LEE et al., 2013; DING et
al., 2006; BAGCHI et al., 2004), com efeito as antocianinas têm ação comprovada contra o
câncer do cólon (YUN et al., 2010).
O valor da utilização tradicional de antocianinas no controle da hipertensão tem sido
confirmado cientificamente, bem como o seu efeito sobre infecção do trato urinário com seu
efeito anti-inflamatório e antimicrobiano. As antocianinas podem suprimir a proliferação celular
oncogénica além de atuar na prevenção do câncer, e da diabetes e atuar no combate aos
problemas cardiovasculares e neurológicos (KONCZAK; ZHANG, 2004).
As antocianinas têm comprovado efeito antioxidante e assim atuam na inibição dos
efeitos provocados pelos radicais livres. Ao prevenir processos oxidativos evitam danos ao ADN
e às macromoléculas, os quais poderiam provocar doenças tais como o câncer, doenças do
coração e catarata (AUBERVAL et al., 2015).
O alto conteúdo de antocianinas com a seu consequente poder antioxidante, tem
despertado crescente interesse no estudo do açaí (Euterpe oleracea Mart.). Este fato está
relacionado com o efeito antimutagênico de açaí (Euterpe oleracea Mart.) (GALOTTA;
BOAVENTURA, 2005). Seus frutos têm alto valor nutritivo além de ser considerado uma grande
fonte de antioxidantes (SANTOS et al., 2008).
O Euterpe oleracea Mart. é encontrada em zonas úmidas ao longo de rios e córregos,
nas terras baixas e zonas úmidas, ao longo de toda extensão da Amazônia (MALCHER, 2011).
Devido ao seu alto teor de antocianinas o açaí (Euterpe oleracea Mart.) pode ser considerado um
alimento funcional, e são justamente estes componentes ativos que atribuem a estes frutos uma
peculiar cor púrpura avermelhada (IADEROZA et al., 1992; OZELA et al., 1997).
Os frutos açaí têm um alto teor de antocianinas, lípidos, proteínas, fibras e minerais tais
como o Mn++, Cu++ e B++ e vitaminas. O alto valor energético dos frutos provém da sua riqueza
em lipídios. Seus compostos fenólicos explicam a sua capacidade antioxidante (SANTOS et al.,
2008; MALCHER, 2011).

126
O objetivo deste estudo foi obter biofilmes kefir associados com EHEEo como meio de
fixação e uso de antocianinas.

127
2 MATERIAL E MÉTODOS
Os grãos de kefir foram obtidos no Laboratório de Pesquisa em Fármacos da
Universidade Federal do Amapá, Amapá, Brasil. Os frutos maduros de Euterpe oleracea Mart.
foram coletadas na Chácara "Recanto Santa Clara", (localizado em -0.96039 N; -51.268450 O) na
cidade de Mazagão, Amapá, Brasil.
2.1 OBTENÇÃO DE EXTRATO HIDROETENÓLICOS DE Euterpe oleracea Mart.(EHEEO)
Foram utilizados 500g da fruta madura de Euterpe oleracea Mart. colocados à
temperatura ambiente (23°C) em etanol a 98°GL acidificado com 2% de ácido fórmico durante 2
horas. Após este procedimento, a solução de extração foi filtrada e a massa descartada. O EHEEo
foi concentrado em rotoevaporador (Hikal, Índia), e a determinação de antocianinas foi realizada
com base no método AOAC (2005-02).
2.2 OBTENÇÃO DE BIOFILMES ASSOCIADOS AO EXTRATO HIDROETANÓLICO DE
Euterpe Oleracea MART. (BEHEEo)
Para obter o biofilme associados ao extrato hidroetanólico de Euterpe oleracea Mart.
(BEHEEo), frascos com capacidade de 3,6 L foram assepsiados e em seguida colocado numa
capela de fluxo laminar sob a radiação UV ultravioleta, durante duas horas. Uma solução de 400
mL açúcar mascavo (AM) (40 g/L) com GK (0,25, 0,5, 1, 2, 5, 10, 20, 40, 60 e 80 g/L) foram
adicionados a 100 ml de EHEEo e acondicionadas a 25°C durante 20 dias. Após este tempo, os
BEHEEo foram removidos e pesados. O volume e o pH da solução remanescente foram medidos
e considerou-se a melhor concentração de GK para a formação BEHEEo a que resultou maior
biomassa com consistência, ou seja, o biofilme que pôde ser separado sem perda de integridade.

128
2.3 DETERMINAÇÃO DA CONCENTRAÇÃO DE SUBSTRATO COM EHEEo PARA
FORMAR BIOFILMES
Foram inoculados 40 g/L de GK em diferentes concentrações de solução AM (0,25, 0,5,
1, 2, 5, 10, 20, 40, 60 e 80 g / l) com 100 ml da EHEEo, com um volume final de 500 ml. O
experimento foi realizado em triplicata. Após 20 dias pH, volume e peso de BEHEEo foram
medidos, sendo considerado o melhor tratamento a que apresentou a maior biomassa BEHEEo
com consistência.
2.4 DETERMINAÇÃO DA CONCENTRAÇÃO DE EHEEo OBJETIVANDO FORMAÇÃO
DE BIOFILME
As concentrações de 10, 25, 50, e 100 mL de EHEEo foram inoculados completando o
volume para 500 ml com solução AM (40 g/L) com grãos de kefir (20 g/L). Após 20 dias, os
grãos de kefir e os BEHEEo formados foram pesados. O pH, a temperatura e o volume foram
medidos e foi realizada a dosagem de antocianinas, tanto no BEHEEo como no meio de cultura,
com base no método da AOAC (2005-02). Foi considerada a melhor concentração a que
apresentou a maior concentração de antocianinas.
2.5 EXTRAÇÃO E QUANTIFICAÇÃO DE ANTOCIANINAS INCORPORADAS EM
BIOFILMES
A quantificação de antocianinas foi realizada através do método da AOAC (2005-02).
GK foi inoculado (20 g/L) em 400 ml de solução de AM 40 g/L com 100 ml de EHEEo. Após 20
dias o BEHEEo foi removido, pesado numa balança analítica e, após desidratado, 1 g BEHEEo
foi colocado em 100 ml de água destilada pH 6.5 e deixado repousar durante 24 horas sempre
protegido da luz, à temperatura ambiente (25°C). Em seguida foram retirados 10 ml desta
amostra para doseamento de antocianina para ensaio de diluição.
Também foi retirado 10 ml do meio de cultura bem como 1 g do EHEEo para
doseamento. Ambas as amostras foram filtradas em papel de filtro Whatmam papel nº 3.
Porttanto foram quantificadas as antocinqaina em uma amostra do extrato puro de açaí
(EHEEo) (1 g), uma amostra do meio de cultura com kefir com açaí (10 mL) e uma amostra do

129
extrato aquoso do biofilme de açaí (10 ml). Ambas amostras foram diluídas em tampão (KCl 0,03
M, pH 1), e em tampão de acetato de sódio (0,4 M, pH 4,5), e completado o volume para 50 mL.
Em seguida, as leituras foram realizadas em Shimadzu UVMini 1240 (Shimadzu Corporation,
Quioto, Japão) em um comprimento de onda de 520 nm e 700 nm, respectivamente. Para calcular
a concentração de antocianina, foi empregue a seguinte fórmula:
TAC = A / e x l x MW x DF x M / W x 100%, onde:
TAC = teores de antocianinas totais em%
A = (A520 nm - A700 nm) pH1.0 - (A520 nm - A700 nm) pH4,5
MW (peso molecular) = 1-449.2mol
DF = factor de diluição
W = peso da amostra em mg
L = percurso óptico
= 26.900 M coeficiente de extinção em L mol -1 cm -1 para cyd-3-glu
A antocianina dos frutos E. oleracea é expressa como cianidina-3-glucose (INÁCIO et al.,
2013) e este método baseia-se no fato de que as antocianinas monoméricas sofrem alteração
estrutural reversível em função do pH.
2.6 PERFIL DA LIBERAÇÃO E DISSOLUÇÃO DE ANTOCIANINAS DE BEHEEo
Neste ensaio foi utilizada o Dissolutor Modelo 299-6 (Nova Ética, Ltda. São Paulo,
Brasil) com velocidade de rotação de 75 rpm. Em cada vaso de o equipamento foi colocado 1 g
de BEHEEo e 100 mL de água destilada, pH 7,0 a qual foi utilizado como solvente. A
temperatura foi mantida a 37±1°C. As amostras foram coletadas a 5, 10, 30 e 60 minutos
respectivamente. Após o final do processo, as amostras foram filtradas através de papel Whatman
n° 1, e, em seguida, realizada a determinação de antocianinas utilizando o método AOAC (2005-
02).

130
2.7 DOSAGEM DE MACRO E MICROELEMENTOS DE BEHEEO
Foram utilizados 100 g das amostras BEHEEo para a determinação de macro e
microelementos, usando o método de calcinação descrito por Salazar et al. (2011). A análise foi
feita por espectrofotometria de absorção atômica modelo 6300 Shimadzu AAS.
2.8 ANÁLISE ESTRUTURAL BEHEEo POR MICROSCOPIA DE FORÇA ATÔMICA (AFM)
A morfologia da superfície de BEHEE foi analisada por microscopia de força atômica
(AFM Easyscan2/STM, Nanosurf; EUA). A umidade relativa do ar durante as medições foi
mantida em 51%. A varredura foi de 30 mm x 30 mm, e as imagens foram obtidas no modo de
contato. O cantilever tem forma retangular com constante de mola de 0,77 nN/m e força
constante de 0,2 N/m, frequência de ressonância de 13 KHz. Para a análise de BEHEEo estes
foram desidratados e eletrodepositados em lâminas com filmes de ouro depois de 03/10 ciclos de
varrimento e analisados por voltametria cíclica no dispositivo.
Vinte imagens de diferentes regiões foram examinadas e analisadas, e os valores de
rugosidade (RMS) foram calculados. O parâmetro estudado foi a rugosidade média (Rm) e o seu
valor eficaz (RMS), definidos pelas equações descritas por Khulbe et al. (2008):
dxdyyxZRm ),(
Onde Rm é a media de rugosidade e Z (x,y) é o perfil vertical da área
E RMS expressa a média da raiz quadrada da rugosidade

131
2.9 ANÁLISE DE BIOFILMES DE KEFIR ATRAVÉS DE MICROSCÓPIO ELETRÔNICA
DE VARREDURA (MEV)
A Microscopia Eletrônica de Varredura (MEV) foi utilizada para se analisar a estrutura e
a composição microbiológica dos biofilmes de kefir com açaí. Foi utilizado o Microscópio
Eletrônica de Varredura TM3030Plus do Laboratório de Pesquisa em Fármacos da UNIFAP.
2.10 ANÁLISE ESTATÍSTICA
Para análise estatística dos resultados do software Statgraphics Centurion XVI
(STATPOINT Technologies, Inc, Warrenton, VA, EUA) foi usado. Foi utilizada ANOVA
seguido pelo teste de Tukey, e o peso médio de BEHEEo foram analisados utilizando o teste t de
Student, e os resultados com p <0,05 foram considerados significativos.

132
3 RESULTADOS
3.1 OBTENÇÃO DE BEHEEO
Constatou-se a formação de BEHEEo cuja biomassa foi dependente do aumento da
concentração da GK (Figura 1).
Observou-se a variação do pH, que era 4,42 no início, e depois decresceu de acordo com
a concentração de GK (Tabela 1).
A ANOVA mostrou a diferença entre os tratamentos. O teste de Tukey identificou que
acima de 10 g/L (13,10±0,30 g) não ocorreu nenhum aumento significativo na biomassa. No
entanto, somente com 40 g/L (14,52±0,45 g) foi obtido a consistência de BEHEEo, que em
concentrações mais baixas fragmenta-se facilmente.
Um parâmetro importante para a obtenção de BEHEEo é a consistência. A consistência
é aqui simplesmente definida como a formação de BEHEEo coesivo nas suas estruturas.
BEHEEo sem consistência fragmenta-se facilmente e se dispersa no meio. A consistência foi
obtida usando apenas concentrações iguais ou superiores a 40 g/L (14,52±0,45 g). O pH decaiu
de 4,42 para 3,46 (Tabela 1) proporcionalmente ao aumento da concentração de GK.
Tabela 1 - Formação de BEHEEo associado ao EHEEo em diferentes concentrações de GK em AM (40
g/L)
Conc. of GK (g) Cresc. Médias (g) pH Volume
0,25 2,35±0,24 4,42±0,01 480
0,5 2,57±0,27 4,61±0,01 482
1,0 6,50±0,20 3,98±0,07 470
2,0 6,20±0,22 3,95±0,04 480
5,0 8,95±0,20 3,76±0,02 475
10,0 13,10±0,30 3,79±0,01 465
20,0 13,45±0,33 3,65±0,01 465
40,0 14,51±0,45 3,66±0,04 445
60,0 13,33±0,23 3,45±0,05 450
80,0 11,52±0,33 3,46±0,04 430

133
Os números representam a media ± o desvio padrão de n = 3.
A figura 1 mostra biofilems formados com 20 g/L de grão de kefir, 40 g/L de açúcar
mascavo e 100 mL de extrato hidroetanólicos de açaí (EHEEo)
a) b)
Figura 1 – Biofilmes de kefir com extrato hidroetanólico de Euterpe oleracea (BEHEEo)
formado com grão de kefir (GK) 20 g/L, açúcar mascavo, 40 g/L e extrato de açaí (100 mL). Biofilme
retirado do meio de cultura (1a) e biofilme liofilizado (1b).
3.2 DETERMINAÇÃO DA CONCENTRAÇÃO DE SUBSTRATO COM EHEEo PARA
FORMAR BIOFILMES
O aumento da concentração de AM com EHEEo resultou em BEHEEo coesa. Sem a presença de
AM e apenas EHEEo, o BEHEEo formado mostrou nenhuma coesão. Com o AM o BEHEEo a
partir de 20 g/L (12,44 ± 0,332 g) já apresenta consistência (Tabela 2) e as antocianinas foram
incorporados a eles (Tabela 3).
A ANOVA mostrou diferenças significativas entre os tratamentos com diferentes
concentrações de AM e através do teste de Tukey foi mostrado que acima de 10 g/L
(12,68±0,3272 g) não há diferenças significativas nos tratamentos. Mas, para obter BEHEEo
consistentes foi necessária concentração de 40 g/L (12,62±0.1626 g) (Tabela 2).
A formação de BEHEEo estabilizou-se com cerca de 10 g/L e o pH caiu de 4,61 para
2,72 (Tabela 2).
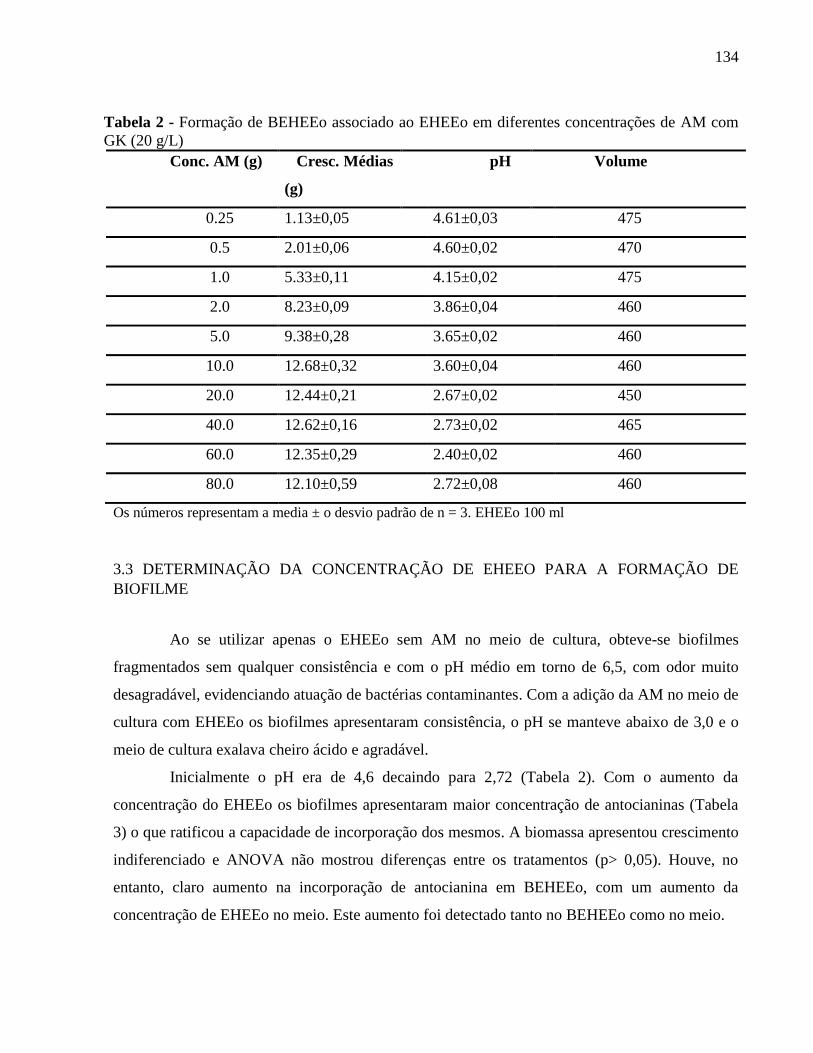
134
Tabela 2 - Formação de BEHEEo associado ao EHEEo em diferentes concentrações de AM com
GK (20 g/L)
Conc. AM (g) Cresc. Médias
(g)
pH Volume
0.25 1.13±0,05 4.61±0,03 475
0.5 2.01±0,06 4.60±0,02 470
1.0 5.33±0,11 4.15±0,02 475
2.0 8.23±0,09 3.86±0,04 460
5.0 9.38±0,28 3.65±0,02 460
10.0 12.68±0,32 3.60±0,04 460
20.0 12.44±0,21 2.67±0,02 450
40.0 12.62±0,16 2.73±0,02 465
60.0 12.35±0,29 2.40±0,02 460
80.0 12.10±0,59 2.72±0,08 460
Os números representam a media ± o desvio padrão de n = 3. EHEEo 100 ml
3.3 DETERMINAÇÃO DA CONCENTRAÇÃO DE EHEEO PARA A FORMAÇÃO DE
BIOFILME
Ao se utilizar apenas o EHEEo sem AM no meio de cultura, obteve-se biofilmes
fragmentados sem qualquer consistência e com o pH médio em torno de 6,5, com odor muito
desagradável, evidenciando atuação de bactérias contaminantes. Com a adição da AM no meio de
cultura com EHEEo os biofilmes apresentaram consistência, o pH se manteve abaixo de 3,0 e o
meio de cultura exalava cheiro ácido e agradável.
Inicialmente o pH era de 4,6 decaindo para 2,72 (Tabela 2). Com o aumento da
concentração do EHEEo os biofilmes apresentaram maior concentração de antocianinas (Tabela
3) o que ratificou a capacidade de incorporação dos mesmos. A biomassa apresentou crescimento
indiferenciado e ANOVA não mostrou diferenças entre os tratamentos (p> 0,05). Houve, no
entanto, claro aumento na incorporação de antocianina em BEHEEo, com um aumento da
concentração de EHEEo no meio. Este aumento foi detectado tanto no BEHEEo como no meio.

135
Tabela 3 - Concentração de antocianinas no BEHEEo formado com grãos de kefir GK (20 g/L) com AM
(40 g/L) em diferentes concentrações de EHEEo.
Conc. de
EHEEo (g/L)
Antociani
nas mg/100g
pH
10 0.00±0,00 2.23±0,02
25 4.93±0,40 2.32±0,01
50 13.10±0,26 3.13±0,01
100 18.06±0,21 3.89±0,02
Os números representam a media ± o desvio padrão de n = 3.
3.4 PERFIL DA LIBERAÇÃO E DISSOLUÇÃO DE ANTOCIANINAS DE BEHEEO
O perfil de dissolução em meio aquoso, mostraram que a liberação das antocianinas do
BEHEEo ocorreu a partir de 5 minutos atingindo o pico em 60 minutos (Tabela 4).
Tabela 4 - Perfil de liberação e dissolução de antocianina do BEHEEo
Tempo
(Minutos)
Antocianinas mg/100g Porcentagem (%)
5 4,3±0,6 0,0040
15 3,8±0,8 0,0038
30 14,5±0,5 0,0145
60 20,8±0,3 0,0208
Os números representam a media ± o desvio padrão de n = 3.
3.5 EXTRAÇÃO E DOSAGEM DE ANTOCIANINA DO BEHEEo
O meio de cultura de dois meses após o início da cultura mostrou ainda antocianinas
(19,32 ± 0,5 mg/100 g).

136
Tabela 5 – Quantificação da concentração de antocianinas in BEHEEo em diferentes períodos.
Produto Concentração
mg/100g
Tempo
EHEEo 110,66±0,57 instantâneo
BEHEEo (12 meses) 34,5±0,70 12 meses
BEHEEo (24 meses) 33,87 ±0,57 24 meses
Os números representam a media ± o desvio padrão de n = 3.
3.6 DOSAGEM DE MACRO E MICROELEMENTOS DE BEHEEO
A concentração de Mg++, Fe++ e Ca++ foi de 0,1808 mg/kg, 0,1740 mg/kg e 0,089 mg/kg,
respectivamente. A concentração de Zn++ foi 0,533 mg/kg (Tabela 6).
Tabela 6 - Concentração de macro e microelementos do EHEEo e do BEHEEo.
Macro e microelementos AM
(mg/kg)
EHEEo
(mg/kg)
BEHEEo
(mg/kg)
Mg++ 0,1856 0,1823 0,1808
Zn++ 0,5740 0,2438 0,5330
Fe++ 0,2496 0,2446 0,089
Cu++ 0,0269 0,0034 0
Ca++ 0,1836 0,1572 0,1384
3.7 COMPOSTOS VOLÁTEIS DO EHEEO
Os principais compostos voláteis encontrados no kefir cultivado com EHEEo foram
ácidos carboxílicos (Tabela 7). Alguns destes compostos são precursores da formação de outros
após armazenamento.

137
Tabela 7 – Compostos voláteis majoritários encontrado no kefir com EHEEo.
Composto Tempo de retenção
(min.)
Área %
2-butanol 3,248 0,23
Ácido pentanóico 4,008 0,10
Ácido acético 8,742 0,12
Ácido propanodióico 13,200 0,10
Butanediol 15,043 0,13
Butanol 15,300 0,13
Ácido nonanóico 17200 0,38
Ácido decanóico 21,333 0,57
Ácido dodecanóico 28,700 2,65
Ácido Tetradecanóico 35,625 3,21
Ácido heptadecano 36,883 0,54
Ác. Benzenodicarboxílico 38,608 0,46
Eicosano 40,100 0,38
Ácido Hexadecanóico 40,858 0,44
Kaur-16-ene 43,917 0,52
Ácido Heptadecanóico 44,950 0,17,
Ácido Hexadecano 46,117 0,21
Ácido octadecanóico 47,842 1,20
Ácido 2-pentenóico 51,358 0,82
Ác 8-metil-pentadecano 56,783 0,28
Nonadecano 61,558 0,66
Tetratetracontano 63,867 0,60
3.8 ANÁLISE ESTRUTURAL BEHEEO POR MICROSCOPIA DE FORÇA ATÔMICA
Na análise de BEHEEo por AFM, obteve imagens em 3D com diferentes concentrações
de EHEEo (Figura 2) que revelaram diferentes configurações de rugosidade (RMS), dependente
da concentração (Figura 2b e 2d). Há um aumento da rugosidade dependente da concentração de

138
EHEEo no meio de cultura. Com concentrações inferiores (20 g/L), foi revelada a presença
predominante de Lactobacillus (Figuras 2a) e em concentrações mais elevadas (60 g/L) prevalece
a presença de leveduras (Figura 2c). Estes resultados estão de acordo com Stadie et al. (2013) que
demonstraram que Lactobacillus metabolizam o meio de cultura e assim o disponibiliza para as
leveduras.
Figura 2 - Imagem gráfica através de microscopia de força atômica (AFM) de biofilme de kefir a)
Imagem gráfica da superfície do biofilme quando utilizado 20 g/L de grãos de kefir e 40 g/L açúcar
mascavo. b) Imagem topográfica do biofilme quando utilizado 20 g/L de grãos de kefir e 40 g/L
açúcar mascavo. c) Imagem gráfica da superfície do biofilme quando utilizado 60 g/L de grãos de
kefir e 40 g/L açúcar mascavo. d) Imagem topográfica do biofilme quando utilizado 60 g/L de
grãos de kefir e 40 g/L açúcar mascavo.Em todo este experimento ao meio de cultura foi
adicionado 100 g EHEEo.

139
3.9 ANÁLISE ESTRUTURAL BEHEEO POR MICROSCOPIA ELETRÔNICA DE
VARREDURA (MEV)
Figura 3 - Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (20 g/L) com açúcar
mascavo (40 g/L) e açaí (100 g/L) (BEHEEo) obtida com resolução 2500x onde se observa a presença de
emaranhado de hifas.
3.10 DETERMINAÇÃO DA TEMPERATURA NA FORMAÇÃO DE BEHEEo
Em relação à formação BEHEEo a temperatura é um dos parâmetros mais importantes.
Com o meio de cultura cultivado à temperatura ambiente (25 ± 2°C) foi obtido com um BEHEEo
biomassa significativa e consistente ao passo que à temperatura de 15°C observou-se a formação
de estruturas sem consistência, fragmentadas e dispersas no meio. A temperatura ótima foi de
30°C e não houve formação a 38°C.

140
Tabela 8 – Efeito da temperatura sobre a síntese de BEHEEo
Temp. (°C) Biomassa (g)
15 0
25 21,46±0,50
30 27,53±0,92
38 0
Os números representam as médias ± desvio padrão de n=3

141
4 DISCUSSÃO
Na formação do biofilme é crucial a adesão inicial das células, e a característica da
superfície de tem uma grande influência nesta fase (PEREIRA et al., 2015). Por conseguinte, a
formação de biofilmes depende da variação da concentração de substâncias e das condições de
fluxo na superfície e também da turbulência meio de cultura de (PERNI et al., 2006).
Os três fatores mais importantes para formar a BEHEEo foram: tempo, temperatura e
concentração de substrato (Tabelas 8 e 2). Biofilmes podem ser formar em apenas 24 horas
(ALIMOVA et al., 2006), e de acordo com Heydorn et al. (2000) e Stoodley et al. (2002) o
crescimento de células em um biofilme é parte do seu próprio processo de maturação, que ocorre
até 10 dias após a adesão inicial. Este aumento é um resultado tanto da divisão celular como da
co-adesão de outras células presentes no sistema em estado planctônico. A formação BEHEEo só
ocorreu depois de 20 dias, embora já pudesse ser observada a formação de estruturas do biofilme
a partir do quinto dia. Estas estruturas são agregadas por substância aglutinante secretada no meio
de modo que formam o biofilme.
O BEHEEo formou-se à temperatura ambiente (cerca de 25°C). À 15°C, os biofilmes
não se formaram, mas observou-se fragmentos que ficaram dispersas no meio. Asadishad et al.
(2010) relataram que a baixas temperaturas (inferiores a 10°C) ocorre a paralisação da produção
de biofilmes por Bacillus subtilis. Os biofilmes formados por Listeria sp. são mais resistentes à
temperatura de 15°C do que a 37°C (SHIMAMURA et al., 2015). Já o BEHEEo não se formou a
37°C. A temperatura mais expressiva com relação à formação de BEHEEo foi de 30°C (Tabela
7).
No que diz respeito à concentração de substrato (AM), pode-se observar que em baixas
concentrações as estruturas dos biofilmes são formadas, mas não se fundem permanecendo
dispersas no meio e em concentrações superiores a 20 g/L o biofilme se forma, mas não apresenta
coesão, sendo fragmentado à menor agitação do meio. A partir da concentração de 40 g/L, a
consistência foi alcançada, a qual coincide com o aumento da rugosidade biofilmes. O
crescimento da concentração de substrato coincidiu proporcionalmente com o aumento de
microrganismos presentes no biofilme. Este é um fator muito importante para a adsorção de
moléculas presentes no biofilme, que precede a adesão de bactérias (RUBIO et al., 2006).
Quando o substrato foi insuficiente ou quando se utilizou apenas a EHEEo como substrato sem a

142
presença de AM, os biofilmes que se formaram mostraram-se sem coesão e por isso apenas
fragmentos de biofilmes se formaram, os quais permaneceram dispersos no meio.
O aumento da concentração de substrato (AM) foi acompanhado pelo decaimento do pH
de maneira muito mais significativa do que a variação de pH com o aumento da concentração de
grãos de GK, o que sugere a ocorrência do aumento da atividade de microrganismos sobre o
substrato, resultando em produtos finais que tornam o meio acidificado pela presença de ácido
láctico e ácido acético. O meio de cultura com kefir após 18 meses permaneceu com pH
inalterado (2,81) e livre de contaminantes. De acordo com a Abe et al. (2013) o baixo pH
constitui uma proteção contra a contaminação. Embora o pH baixo ou muito acima da
neutralidade afete diretamente o desenvolvimento de biofilme por interferir com a força próton-
motora, que é a força usada pela bactéria para gerar o gradiente eletroquímico (PEREIRA, 2001),
o pH baixo não interferiu na formação da BEHEEo.
Os microrganismos presentes no kefir são incorporados no biofilme (Figura 3). Assim,
eles assegurar um ambiente livre de contaminantes. O pH cai até que um nível de proteção contra
contaminantes. Probióticos como kefir são organismos que vivem em consórcio e repelem
organismos patogênicos (SANTOS et al., 2003).
As antocianinas incorporadas no BEHEEo e permaneceram estáveis 24 meses após a
formação. Estas antocianinas foram liberadas do BEHEEo em meio aquoso em pH 7,0, sem
perder as suas características. Este fato é importante porque as antocianinas do açaí (Euterpe
oleracea Mart.) inevitavelmente sofrem degradação a partir da colheita até chegar ao
consumidor. Rogez et al. (2012), concluíram que a cinética de degradação de antocianinas se
efetiva com uma semi-vida de 50 horas, ocorrendo, concomitantemente, com a infecção por
micro-organismos tais como fungos, bactérias mesofílicas e coliformes fecais.
A liberação das antocianinas do BEHEEo ocorreu a partir de 5 minutos, atingindo o pico
aos 60 minutos. Esta cinética da liberação dependente do tempo é um fator importante para a
aplicação de BEHEEo, uma vez que o aumento gradual na liberação antocianina sugere que
também pode haver um aumento da eficiência ao longo do tempo.
No meio de cultura do kefir as antocianinas permaneceram estáveis durante pelo menos
dois meses. Após esse período, elas começaram a ser degradada, juntamente com o meio.
Bobbio et al. (2000) descreveram dois tipos de antocianinas em EHEEo, a cianidina-3-
arabinoside e a cianidina-3-arabinoside-arabinosil as quais considerou como as duas principais

143
antocianinas do E. oleracea tendo encontrado na casaca das suas frutas o teor de 50,00 ±
5nmg/100g. Eles concluíram que a fruta como um todo deve ter 263 mg/ 100g de casca, pois a
casca corresponde a apenas 19% do fruto. No entanto as antocianinas do E. oleracea estão
concentradas apenas na casca.
Neste estudo encontrou-se 110 mg/100g de antocianina no EHEEo. No BEHEEo
verificou-se 36 mg/100g. Cumpre ressaltar que a dosagem foi realizada em BEHEEo com seis
meses de incorporação.
O meio de cultura sem AM contendo apenas o EHEEo como substrato apresentou
biofilmes com baixa coesão, fragmentados e com pH de 6,7, sugerindo ações de diferentes tipos
de bactérias.
Os perfis de liberação e dissolução de BEHEEo, sintetizado em menos de 12 meses
liberou 34,5±0,70 mg/100 g de cianidina-3-glucose (Tabela 5), enquanto BEHEEo de 24 meses
liberou 33,87 ±0,57 mg de cianidina-3-glucose.
Com relação à presença de macro e microelementos, pode-se frisar a presença de 0,553
mg/kg de zinco no BEHEEo e de 0,243 mg/kg no EHEEoe 0,574. mg/kg no açúcar mascavo.
Rogez (2000) encontraram 7 mg/100g no extrato de açaí (Euterpe oleracea Martius) e Menezes
et al. (2008) encontraram 2,82 mg/100g no extrato de açaí liofilizado (Euterpe oleracea Martius).
Pode-se infeir que o zinco incorporado no biofilme foi, portanto predominantemente advindo do
açúcar mascavo
Os biofilmes podem utilizar a capacidade de absorção de alguns metais na sua forma
iónica, tais como o zinco, o cobre, o ferro e alumínio contido na sua matriz a fim de evitar forças
de erosão (GRUMBEIN et al., 2014). Estes metais iónicos são tóxicos para as formas
planctônicas de bactérias, componentes do biofilme, mas a sua toxicidade é suprimida na matriz
do biofilme (GRUMBEIN et al., 2014).
A análise por AFM pode ser usado para avaliar a evolução morfológica de biofilmes
(CHATTERJEE et al., 2014, PEREIRA et al., 2015), bem como proporcionar uma compreensão
dos mecanismos biofísicos do impacto da estrutura organizacional da cápsula de polissacarídeo
na formação de biofilme (WANG et al., 2015).
O AFM faz medidas nano mecânicos que fornecem dados que mostram como a
organização capsular afeta a adesão das células e, consequentemente, a formação de biofilmes
(WANG et al., 2015). Neste estudo, através de AFM, foi registada a presença de Lactobacillus

144
em grande quantidade nos biofilmes (Figuras 2a), especialmente em concentrações mais baixas
de EHEEo 10 g/L. Em concentrações mais altas (60 g/L) verificou-se também a presença
predominante de levedura (Figura 2c). O Lactobacillus acidifica o meio e assim proporciona
condições ótimas para o metabolismo das leveduras (STADIE et al., 2013) que se proliferam em
BEHEEo com o aumento da concentração de EHEEo a 60 g/L. No kefir, as leveduras fornecer os
aminoácidos essenciais e vitaminas, como ácido fólico para os lactobacilos os quais reduzem o
pH criando condições ideais para a levedura, numa relação mutualística esclarecida por Stadie et
al. (2013).
Sabe-se que o substrato influencia a formação de biofilme e também que a extensão da
colonização microbiana aumenta com o aumento da rugosidade da superfície (CHARACKLIS et
al., 1984). Portanto o aumento na concentração do substrato causa um crescimento do biofilme
com o consequente aumento da rugosidade (VIANA, 2009). Com a análise por AFM é possível
detectar estas alterações da rugosidade da superfície do biofilme (LÓPEZ-JIMÉNEZ et al., 2015).
A variação da concentração de EHEEo no meio de cultura foi acompanhada por alterações na
rugosidade do BEHEEo (Figuras 3b, 2d). O aumento da rugosidade constitui um fator muito
importante para indicar maior capacidade de retenção de células e incorporação de substâncias
como antocianinas.

145
5 CONCLUSÃO
O BEHEEo formado reteve e preservou as antocianinas de EHEEo. A análise por AFM
confirmaram o aumento da rugosidade da BEHEEo dependente da concentração de EHEEo, e
mostrou Lactobacillus e leveduras de maneira profícua nestes biofilmes.
O BEHEEo reteve antocianinas de EHEEo por períodos relativamente longos, e com as
condições in vitro (pH 7,0), as antocianinas de BEHEEo foram liberadas.
6 AGRADECIMENTOS
Autores gostariam de agradecer a CAPES (nº 3292/2013 AUXPE) e CNPq Proc. 402332 / 2013-0 (Biotec)
pelo apoio financeiro.

146
REFERÊNCIAS
ABE, A.; FURUKAWA, S.; WATANABE, S.; MORINAGA, Y. Yeasts and lactic acid bacteria
mixed-specie biofilm formation is a promising cell immobilization technology for ethanol
fermentation. Appl Biochem Biotechnol. v. 171, n. 1, p. 72-79, 2013.
ALIMOVA, A.; ROBERTS M.; KATS, A.; RUDOLPH, E.; STEINER, J. C.; ALFANO, R. R.;
GOTTLIEB, P. Effects of Smectite Clay on Biofilm formation by microorganisms. Biofilms, v
3, p 43-47, 2006.
AOAC. (2006). Official methods of analysis of the Association of Official Analytical
Chemists. Arlington: Ed. Patrícia Cuniff. <http://www.aoac.org/omarev1/2005_02.pdf>.
BAGCHI, D. et al. Anti-angiogenic, antioxidant, and anti-carcinogenic properties of a novel
anthocyanin-rich berry extract formula. Biochemistry, v. 69, p. 75-80, 2004.
BOBBIO, F. O.; DRUZIAN, J. I.; ABRÃO, P. A.; BOBBIO, P. A.; FADELLI, S. Identificação e
quantificação das antocianinas do fruto do açaizeiro (Euterpe oleracea Mart.) Ciênc. Tecnol.
Aliment., v. 20 n.3, p. 388-390, 2000.
CHARACKLIS, W. G. Biofilm development: a process analysis. Microbial Adhesion and
Aggregation, New York, 1984.
DAVEY M. E., O'TOOLE G. A. Microbial biofilms: from ecology to molecular genetics.
Microbiology and Molecular Biology Reviews, v. 64, p. 847, 2000.
DING M.; FENG R.; WANG S. Y., BOWMAN L.; LU Y.; QIAN Y.; CASTRANOVA V.;
JIANG B. H.; SHI X. 2006 Cyanidin-3-glucoside, a natural product derived from blackberry,
exhibits chemopreventive and chemotherapeutic activity. J. Biol. Chem., v. 281, p. 17359-
17368, 2006.
DONLAN, R.M. Biofilms: Microbial life on surfaces. Emerging Infect Dis, v. 8, p. 881-890,
2002.
DUFRÊNE, Y. F. Sticky microbes: forces in microbial cell adhesion. Trends Microbiol, v.23, n.
6, p. 376-382, 2015.
FLEMMING, H. C., WINGENDER J. Relevance of microbial extracellular polymeric substances
(EPS) – Part I: Structural and ecological aspects. Water Sci Technol, v. 43, p. 1-8, 2001.

147
GALOTTA, A. L. Q. A.; BOAVENTURA, M. A. D. Quim. Nova, v. 28, p. 610, 2005.
HEYDORN, A.; NIELSEN, A. T.; HENTZER, M.; STERNBERG, C.; GIVSKOV, M.
Quantification of biofilm structures by the novel computer program COMSTAT.
Microbiology, v.146, p.395–407, 2000.
IADEROZA, M.; BALDINI, I. S. D.; BOVI, M. L. A.; Anthocyanins from fruits of açaí (Euterpe
oleracea Mart.) and juçara (Euterpe edulis Mart.). Tropical Science, v. 32, p. 41-46, 1992.
INÁCIO, M. R. C.; DE LIMA, K. M. G.; LOPES, V. G.; PESSOA, J. D. C.; DE ALMEIDA
TEIXEIRA, G. H. Total anthocyanin content determination in intact açaí (Euterpe oleracea
Mart.) and palmitero-juçara (Euterpe edulis Mart.) fruit using near infrared spectroscopy (NIR)
and multivariate calibration. Food Chem, v. 136, p. 1160–1164, 2013.
KHULBE K. C.; FENG C.; MATSUURA T. Synthetic Polymeric Membranes:
Characterization by Atomic Force Microscopy, 1st ed., Springer, 2008.
KONCZAK, I.; ZHANG, W. Anthocyanins-more than nature’s colours. Journal of Biomedicine
and Biotechnology. v.5, p.239-240, 2004.
LAUREYS, D.; DE VUYST L. Microbial species diversity, community dynamics, and
metabolite kinetics of water kefir fermentation. Appl. Environ. Microbiol. v. 80, p. 2564-72,
2014.
LEE S. J.; HONG S.; YOO S. H.; KIM G. W. Cyanidin-3-O-sambubioside from Acanthopanax
sessiliflorus fruit inhibits metastasis by downregulating MMP-9 in breast cancer cells MDA-MB-
231. Planta Med, v. 79, p. 1636-1640, 2013.
LILA, M.A. Anthocyanins and human health: an in vitro investigative approach. Journal of
Biomedicine and Biotechnology, v. 5, p. 306-313, 2004.
LÓPEZ-JIMÉNEZ, L.; ARNABAT-DOMÍNGUEZ, J.; VIÑAS, M.; VINUESA, T. Atomic force
microscopy visualization of injuries in Enterococcus faecalis surface caused by Er,Cr:YSGG and
diode lasers. Med Oral Patol Oral Cir Bucal v. 20, p. 45-51, 2015.
MALCHER, E. S. L. T; Influência da sazonalidade sobre a composição química e atividade
antioxidante do açaí (Euterpe oleracea Mart.), Tese apresentada como requisito parcial para
obtenção de título de doutor ao programa de pós graduação em Biodiversidade tropical da

148
Universidade Federal do Amapá. Orientador: Prof. . Dr. José Carlos Tavares Carvalho, pag. ___,
20011.
OZELA, E. F.; STRINGHETA, P. C.; LIMA, A. A. S.; FARIAS, M. L. T. S. Estudo comparativo
do teor de antocianinas presente no açaí (Euterpe oleracea Mart.) nos períodos de safra e
entresafra. Livro de Programas e Resumo III Simpósio Americano de Ciências de
Alimentos. Campinas, FEA, p.55, 1999.
PERNI, S.; JORDAN, S. J.; ANDREW, P. W.; SHAMA, G. Biofilm development by Listeria
innocua in turbulent flow regimes. Food Control, v.1, p. 875–883, 2006.
PEREIRA, M. O. B. O. Comparação da eficácia de dois biocidas (carbamato e glutaraldeido)
em sistemas de biofilme. 2001. 234 p. Tese (Doutorado) Universidade do Minho, Braga –
Portugal.
PEREIRA, F. D.; BONATTO, C. C.; LOPES, C. A.; PEREIRA, A. L.; SILVA, L.P. Use of
MALDI-TOF mass spectrometry to analyze the molecular profile of Pseudomonas aeruginosa
biofilms grown on glass and plastic surfaces. Microb Pathog v. 86, p. 32-37, 2015.
RODRIGUES, K. L; CAPUTO, L. R.;CARVALHO, J. C. T.; EVANGELISTA, J.
SCHNEEDORF, J. M. Antimicrobial and healing activity of kefir and kefiran extract. Int J
Antimicrob Agents. v. 25, p. 404-408, 2005.
ROGEZ, H. Açaí: Preparo, Composição e Melhoramento da Conservação. Belém: Edufpa,
2000, p.313.
ROGEZ, H.; AKWIE, S. N.; MOURA, F. G.; LARONDELLE, Y. Kinetic modeling of
anthocyanin degradation and microorganism growth during postharvest storage of açai fruits
(Euterpe oleracea). J Food Sci, v. 77, p. 300-306, 2012.
RUBIO, C.; OTT, C.; AMIEL, C.; MORAL, I. D.; TRAVERT, J.; MARIEY, L.
Sulfato/thiosulfato reducing bacteria characterization by FT-IR spectroscopy: A new
approach to biocorrosion control. Journal of Microbiological Methods, v. 64, p. 287-
296, 2006.
SALAZAR, R.F.S.; ALCÂNTARA, M. A. K; IZÁRIO FILHO, H. J. Evaluation of Sample
Preparation of Nickel. R. Bras. Ci.Solo, v. 35, p. 241-248, 2011.

149
SANTOS, A.; SAN MOURO, M.; SANCHEZ, A.; TORRES, J. M.; MARQUINA, D. The
antimicrobial properties of different strains of Lactobacillus spp. isolated from kefir. Systematic
and Applied Microbiology. v. 26, p. 434-437, 2003.
SANTOS, G. M.; MAIA, G.A.; SOUSA, P. H. M.; COSTA, J. M. C.; FIGUEIREDO, R. W.;
PRADO, G. M. Correlação entre atividade antioxidante e compostos bioativos de polpas
comerciais de açaí (Euterpe oleracea Mart.). Archivos Latinoamericanos de Nutrición,
ALAN, v.58, n. 2, 2008.
SHIMAMURA, N.; IRIE, F.; YAMAKAWA, T.; KIKUCHI, M.; SAWAI, J. Heated scallop-
shell powder treatment for deactivation and removal of Listeria sp. biofilm formed at a low
temperature. Biocontrol Sci, v.20, p. 153-157, 2015.
STADIE J.; GULITZ A.; EHRMANN M. A.; VOGEL R. F. Metabolic activity and symbiotic
interactions of lactic acid bacteria and yeasts isolated from water kefir. Food Microbiol, v. 35, p.
92-98, 2013.
STOODLEY, P.; SAUER, K.; DAVIES, D. G.; COSTERTON, J. W. Biofilms as
complex differentiated communities. Annual Review Microbiology, v. 56, p. 187-209,
2002.
STATGRAPHICS CENTURION XVI SOFTWARE (Statpoint Technologies, Inc, Warrenton, VA,
USA).
SUTHERLAND I.W. Biofilm exopolysaccharides: a strong and sticky framework.
Microbiology, v. 147, p. 3-9, 2001.
VAN HULLEBUSCH, E. D.; ZANDVOORT, M. H.; LENS, N. L. Metal immobilisation by
biofilms: mechanisms and analytical tools. Rev Environ Sci Biotechnol v. 2, p. 9-33, 2003.
VIANA, M. G. Avaliação de produtos naturais sobre biofilmes formados em
sistema dinâmico. 2009, 90 p. Dissertação (Mestrado em Ciência e Engenharia de
Petróleo) - Centro de Ciências Exatas e da Terra, Universidade Federal do Rio Grande
do Norte, Natal, 2009.
VU, B.; CHEN, M.; CRAWFORD, R. J.; IVANOVA, E. P. Bacterial Extracellular
Polysaccharides Involved in Biofilm Formation. Molecules, v.14, p.2535-2554, 2009.
WALTON, M. C. et al. Anthocyanins absorption and antioxidant status in pigs. Journal of
Agricultural and Food Chemistry, v. 54, n. 20, p. 7940-7946, 2006.

150
WANG, H.; WILKSCH, J.J.; STRUGNELL, R.A.; GEE, M. L. Role of Capsular Polysaccharides
in Biofilm Formation: An AFM Nanomechanics Study. ACS Appl Mater Interfaces, v. 7, n. 23,
p. 13007-13013, 2015.
WATNICK, P.; KOLTER, R. Biofilms, city of microbes. J Bacteriol, v. 13, p. 20–26, 2000.
YUN, J. W.; LEE, W. S.; KIM, M. J.; LU J. N.; KANG, M. H.; KIM, H.G.; KIM, D. C.; CHOI,
E. J.; CHOI, J. Y.; LEE, Y. K.; RYU, C. H.; KIM, G.; CHOI, Y. H.; PARK, O. J; SHIN, S. C.
Characterization of a profile of the anthocyanins isolated from Vitis coignetiae Pulliat and their
anti-invasive activity on HT-29 human colon cancer cells. Food Chem Toxicol, v. 48, p. 903-
909, 2010.
ZHANG, W.; DAI, W.; TSAI, S. M.; ZEHNDER, S. M.; Sarntinoranont, M.; angelini, T. E.
Surface indentation and fluid intake generated by the polymer matrix of Bacillus subtilis
biofilms. Soft Matter, v. 11, p. 3612-3617, 2015.

151
CAPÍTULO 3: ESTUDO DO BIOFILME DE KEFIR ASSOCIADO AO GÉRMEN DE
SOJA (Glycine max L. Merril)

152
ESTUDO DO BIOFILME DE KEFIR ASSOCIADOAO GÉRMEN DE SOJA (Glycine max
(L.) Merril)
Antonio Ferreira de Oliveiraa,b, Adriana Maciel Ferreiraa , Adriana Valente dos Santosa, Mariana
Baia Goes , Helison de Oliveira Carvalho, Roberto Messias Bezerrad; Robert Maguiña Zamorad,
e José Carlos Tavares Carvalhoa,b*
aPrograma de Pós-graduação em Biodiversidade Tropical, Departamento de Ciências Biológicas e da Saúde,
Universidade Federal do Amapá bLaboratório de Pesquisa em Fármacos, Curso de Farmácia, Departamento de Ciências Biológicas e da Saúde,
Universidade Federal do Amapá cLaboratório de Nanobiotecnologia Fitofarmacêutica, Curso de Farmácia, Departamento de Ciências Biológicas e
da Saúde, Universidade Federal do Amapá dPrograma de Pós-graduação em Ciências Farmacêuticas, Departamento de Ciências Biológicas e da Saúde,
Universidade Federal do Amapá
*Corresponding author:[email protected]
*Corresponding author: email: [email protected]
RESUMO
As isoflavonas são compostos fenólicos bioativos cujo consumo pode reduzir os riscos de
doenças. Dentre todos os grãos já estudados, o grão de soja é o que possui os maiores teores de
isoflavonas. O kefir é um complexo microbiano formado por Lactobacillus e leveduras em
simbiose que restaura a homeostase intestinal, sendo que o kefir pode formar biofilmes que são
constituídos por uma matriz extracelular onde os microrganismos se alojam e através dele
adquirem características peculiares. Este estudo teve como objetivo a formação de biofilmes com
kefir associado ao extrato de gérmen de soja com a finalidade de se obter a incorporação de
isoflavonas do gérmen de soja nos mesmos. Diferentes concentrações de grãos de kefir e de
gérmen de soja foram cultivados em meio de cultura com solução de açúcar mascavo (40g/L) por
20 dias quando os biofilmes foram retirados e foi aferido o pH do meio de cultura e a biomassa.
As isoflavonas foram extraídas do meio de cultura e quantificadas através de cromatografia
líquida de alta eficiência (CLAE). A quantidade de isoflavonas totais encontradas em extrato
puro de gérmen de soja foram 12 µg/L para gliciteína e 18 µg/L para genisteína. No meio de
cultura encontrou-se com 20 dias 19.59±3.57 µg/L de gliciteina e 23.86±2.21 µg/L de genisteina.
A melhor concentração de grãos de kefir com vista a extração de isoflavonas foi 40 g/L com
11,67 µg/L de gliciteína e 17,78 µg/L de genisteína. O processo fermentativo não afetou a
concentração das isoflavonas. Portanto, foi obtido meio de cultura com kefir e isoflavonas bem
como biofilmes de kefir com gérmen de soja. A fusão das propriedades do kefir com as
isoflavonas abre um potencial de aplicação terapeutica considerável.
Palavras-chave: Biofilmes. Kefir. Gérmen de Soja. Glycine max (L.) Merril. Isoflavonas

153
ABSTRACT
Isoflavones are bioactive phenolic compounds whose consumption can reduce the risk of
diseases. Among all the grains of beans that have been studied, soy is the one with the highest
levels of isoflavones. Kefir is a microbial complex consisting of Lactobacillus and yeast in
symbiosis that restores homeostasis intestine. Kefir can form biofilms who consist of an
extracellular matrix where microorganisms are housed and through acquire peculiar
characteristics. The aim of this study was to biofilm formation with kefir grains in the presence of
soy extract for performing the incorporation of isoflavones. Different concentrations kefir grains
and soy germ were growned in culture medium with brown sugar solution (40g/L) for 20 days
when biofilms were removed and the pH of the culture medium and biomass were measured.
Isoflavones of the medium of cuiture were extracted and quantified by high-performance liquid
chromatography (HPLC). The amount of total isoflavones found in soy germ extract were 12
µg/L for glycitein and 18 µg/L of genistein. The culture medium after 20 day was found
19.59±3.57 µg/L for glycitein and 23.86±2.21 µg/L of genistein. The best concentration of kefir
grains in order to isoflavone extraction was 40 g/L being found 11.67 µg/l glycitein and 17.78
µg/L of genistein. The fermentation process did not affected the concentration of isoflavones.
Thus, it was confirmed the incorporation of isoflavones in the biofilm with kefir. The merger of
kefir property with isoflavones opens a significant therapeutic application potential.
Keywords: Biofilms. Kefir. Soy Germ. Glycine max L. Merrill. Isoflavones

154
1 INTRODUÇÃO
Nos últimos anos os efeitos de compostos fenólicos têm sido constantemente estudados
devido suas propriedades antioxidantes com ação preventiva e terapêutica contra várias doenças
crônico-degenerativas tais como a hipercolesterolemia, câncer dos seios e da próstata e doenças
cardiovasculares (CROZIER et al., 2009; GARCÍA-LAFUENTE et al., 2009). Dentre os
compostos fenólicos se destacam os flavonóides dos quais fazem parte as isoflavonas.
As isoflavonas compreendem as agliconas daidzeína, genisteína e gliciteína, os
respectivos b-glicosídios e os conjugados malonil-glicosídios e acetil-glicosídios. As isoflavonas
são compostos fenólicos bioativos cujo consumo pode reduzir os riscos de doenças
cardiovasculares, de alguns tipos de câncer, e de diabetes (BARNES et al., 1999; MANDARINO,
2010; LAUDANNA, 2006; IMAI, 2015). As isoflavonas também reduzem risco de osteoporose,
de câncer dos seios e de próstata (YU et al., 2015; VARINSKA et al., 2015).
Além de exercerem um potente efeito antioxidante as isoflavonas podem exercer no
organismo humano um efeito hormonal (IMAI, 2015) como fitoestrógenos ativos com
reconhecida capacidade hipocolesterolêmica (ROSSI et al., 2004).
Dentre todos os grãos já estudados, o grão de soja é o que possui os maiores teores de
isoflavonas, as quais estão relacionadas a fatores genéticos e ambientais (CARRÃO-PANIZZI et
al., 2009).
Processos fermentativos podem alterar os níveis e formas das isoflavonas nos produtos
acabados, reduzindo em até 13 vezes a quantidade de isoflavonas (ROSSI et al., 2004). A ação
hidrolítica da glicosidase própria de determinadas bactérias láticas pode fazer com que as
agliconas (genisteína e daidzeína) fiquem em níveis mais altos (MATSUDA et al., 1992; WANG;
MURPHY, 1994).
O kefir é um probiótico constituído por uma biomassa complexa com bactérias ácido
lácticas e ácido acéticas que vivem em simbiose em prol do hospedeiro originando um produto
fermentado que realiza a síntese e melhor absorção de proteínas, vitaminas e minerais
(FARNWORTH, 2005).
O kefir em água com açúcar constitui em um soro produzido a partir de fermentação de
bebidas fermentadas pelos grãos de kefir (LAUREY; DE VUYST, 2014).
O kefir tem possibilidade de participar em processos de formação de biofilmes.

155
Biofilmes são superfícies com alto grau de organização estrutural associados a colônias de
bactérias mantidas coesas através de um polímero secretado por suas células o qual é denominado
de substância polimérica extracelular (SPE) (ZHANG et al., 2015). Os biofilmes conferem
resistência ao stress, proteção contra impactos químicos e mecânicos e disponibilizam os
nutrientes, por isso são fundamentais para sobrevivência dos microrganismos (DAVEY;
O'TOOLE, 2000; GORBUSHINA; BROUGHTON, 2009).
O objetivo deste estudo foi a formação de biofilmes com kefir na presença de extrato de
germem de soja. Objetivou-se determinar a concetração necessária de gérmen de soja, de grãos
de kefir e para a produção de biofilmes, extrair e quantificar as isoflavonas que foram
incorporadas nos biofilmes e no meio de cultura com kefir

156
2 MATERIAL E MÉTODOS
2.1 DETERMINAÇÃO DA CONCENTRAÇÃO DE GÉRMEN DE SOJA NECESSÁRIA
PARA A FORMAÇÃO DE BIOFILMES COM KEFIR E AM
Para a obtenção dos biofilmes com gérmen de soja foram utilizados frascos com
capacidade de 3,6 L rinsados com etanol a 70% e colocados em cabine de fluxo laminar sob
radiação ultravioleta UV por duas horas. A uma solução de açúcar mascavo (AM) (40 g/L) com
20 g/L de grãos de kefir GK foram adicionadas diferentes concentrações de gérmen de soja (0, 1,
2, 5, 10, 20 g/L). Os frascos foram então acondicionadas à temperatura ambiente 25±2 °C por 20
dias. Após este tempo os biofilmes formados foram retirados e pesados. O volume e o pH da
solução remanescente foram medidos. O doseamento de isoflavonas foi realizado tanto no meio
como nos biofilmes. Foi considerada a melhor concentração de gérmen de soja a que apresentou
biomassa com consistência, ou seja, aquela que pôde ser separado do meio sem perda da
integridade com a maior concentração de isoflavonas.
Padrões de gliciteína e genisteína (Sigma Aldrich) foram utilizados na concentração de
0,0125mg/mL. Para preparao da solução padrão as isoflavonas foram extraídas do gérmen de soja
20 g/L com a utilização de etanol 70% por duas horas.
2.2 DETERMINAÇÃO DA CONCENTRAÇÃO DE GRÃOS DE KEFIR NECESSÁRIA PARA
A FORMAÇÃO DE BIOFILMES COM GÉRMEN DE SOJA
Foram preparadas soluções de açúcar mascavo 40 g/L esterilizados a 121 °C durante 20
minutos em autoclave (Phoenix, Brasil). Foram adicionados em câmara de fluxo laminar 20 g/L
de gérmen de soja com as concentrações de 5, 10, 20, 40, 60 e 80 g/L de grãos de kefir nos
respectivos frascos. Os frascos foram armazenados a 25°C durante 20 dias. Após este período, os
biofilmes formados foram separados e pesados. Foi realizada o doseamento das isoflavonas tanto
no meio como nos biofilmes. Também foram aferidos o volume e o pH das soluções restantes. Os
biofilmes formados foran analisados por microscopia de força atômica (AFM) e por microscopia
eletrônica de varredura (MEV). O parâmetro analisado através da microscopia de força atômica
(AFM) foi a rugosidade.

157
2.3 DETERMINAÇÃO DA CONCENTRAÇÃO DE ISOFLAVONAS NO MEIO DE
CULTURA E NOS BIOFILMES
A extração das isoflavonas foi realizada de acordo com a metodologia de Carrão-Panizzi
et al. (2002). Foram retiradas 100 ml de cada amostra do meio de cultura com solução de açúcar e
kefir com gérmen de soja e filtrado em papel Whatman com microporos com 8 de diâmetro.
Em seguida 1,5mL foi filtrado através de membrana com poros de 0,45 µm, da marca
“Millipore”. Para injeção no cromatógrafo líquido, foram utilizados 20 µL do extrato filtrado. A
separação e a quantificação das isoflavonas foram realizadas de acordo com modificações na
metodologia preconizada por Berhow (2002). As análises foram realizadas em cromatógrafo
líquido de alta performance-HPLC (Shimadzu corporation) equipado com auto-injetor, detector
de arranjo diodos com varredura de 190 a 500 nm.
Condições cromatográficas: cromatogramas obtidos em 254 nm, com temperatura do
forno mantida em 30 ºC, coluna de fase reversa Shim-pack VP-ODS (150 x 4.6 mm; 5 µm),
volume de injeção de 10 µL, utilizando como fase (A) água acidificada a 0,1 % com acido
acético, e como fase (B) Metanol, proporção (60:40) com vazão de 1 mL/min. A quantificação
das isoflavonas gliciteína e genisteína foram realizadas por comparação das áreas dos picos
obtidas a partir do preparo de soluções padrões contendo 10 µg/mL de gliciteína e 15 µg/mL de
genisteína diluídas em Metanol, posteriormente filtradas em membrana filtrante com poros de
0,45 µm (Millepore®) (César et al., 2007).
Os teores de isoflavonas determinados foram expressos em mg/100 g de amostra em
base seca. As isoflavonas separadas e quantificadas foram agliconas (gliciteína e genisteína
(COWARD et al., 1993). Os valores de isoflavonas foram apresentados em equivalentes
agliconas (GÓES-FAVONI et al., 2010).
2.4 DETERMINAÇÃO DE PERÍODO DE INCORPORAÇÃO DE ISOFLAVONAS
Os teores de isoflavonas foram quantificados no meio de cultura com 5, 10, 15 e 20 dias.
Foram efetivados o doseamento para gliciteína e genisteína.

158
2.5 DOSAGEM DE MACRO E MICROELEMENTOS DE BIOFILMES DE KEFIR COM
GÉRMEN DE SOJA
Foram utilizados 100 g gérmen de soja e 100 g de biofilmes formado com 20g/l de
gérmen de soja e 40 g/L de grãos de kefir para determinar as concentrações de macro e
microelementos através do processo de calcinação descrita por Salazar et al. (2011). A análise foi
levada a cabo num espectrofotômetro de absorção atómica (AAS Shimadzu modelo 6300). Estas
análises foram realizadas no Laboratório de Absorção Atômica e Bioprospecção (LAAB) da
Universidade Federal do Amapá, UNIFAP - Amapá - Brasil.
2.6 ANÁLISE ESTRUTURAL DE BIOFILMES DE KEFIR COM GÉRMEN DE SOJA
ATRAVÉS DE MICROSCOPIA DE FORÇA ATÔMICA (AFM)
A morfologia da superfície dos biofilmes formado com gérmen de soja foi analisada por
um Microscópio de Força Atômica (AFM), Easyscan2 AFM/STM da Empresa Nanosurf no
Laboratório de Ciências dos Materiais (LABMAT), Departamento de Física da UNIFAP. A
umidade relativa durante as medidas foi mantida a 51%.
A varredura (tamanho de digitalização) foi de 30 mm X 30 mm. As imagens foram
obtidas em contact mode (modo de contato). O cantilever utilizado tem forma retangular com
uma constante de mola de 0,77 nN/m e com uma força constante 0,2 N/m e frequência de
ressonância de 13 KHz. Para estas medições os biofilmes foram desidratados e eletrodepositados
em lâminas com filmes ouro e após 03/10 ciclos de digitalização foram analisadas por
voltametria cíclica.
Imagens de diferentes regiões dos biofilmes foram examinadas e analisadas com o
software AFM para calcular valores da rugosidade (RMS). Vários parâmetros de rugosidade
podem ser medidos através de AFM. Os parâmetros utilizados neste trabalho são a média de
rugosidade (Rm) e seu valor eficaz (RMS), calculados usando as seguintes equações (KHULBE
et al., 2008):
Onde Rm é a rugosidade média e Z (x, y) é a função da altura da área.

159
Onde RMS é expresso como a raiz quadrada de rugosidade média.
2.7 ANÁLISE DE BIOFILMES DE KEFIR COM GÉRMEN DE SOJA ATRAVÉS DE
MICROSCÓPIO ELETRÔNICA DE VARREDURA (MEV)
A Microscopia Eletrônica de Varredura (MEV) foi utilizada para se analisar a estrutura e
a composição microbiológica dos biofilmes de kefir com gérmen de soja. Foi utilizado o
Microscópio Eletrônica de Varredura TM3030Plus do Laboratório de Pesquisa em Fármacos da
UNIFAP. do Laboratório de Pesquisa em Fármacos da Universidade Federal do Amapá
(UNIFAP), Macapá, Amapá, Brasil.
2.8 ANÁLISE ESTATÍSTICA
Para análise estatística dos resultados foi utilizado o software Statgraphics Centurion
XVI (STATPOINT Technologies, Inc, Warrenton, VA, EUA). Foi utilizada ANOVA seguido
pelo teste de Tukey, e o peso médio dos biofilmes de kefir com gérmen de soja foram analisados
utilizando o teste t de Student, e os resultados com p <0,05 foram considerados significativos.
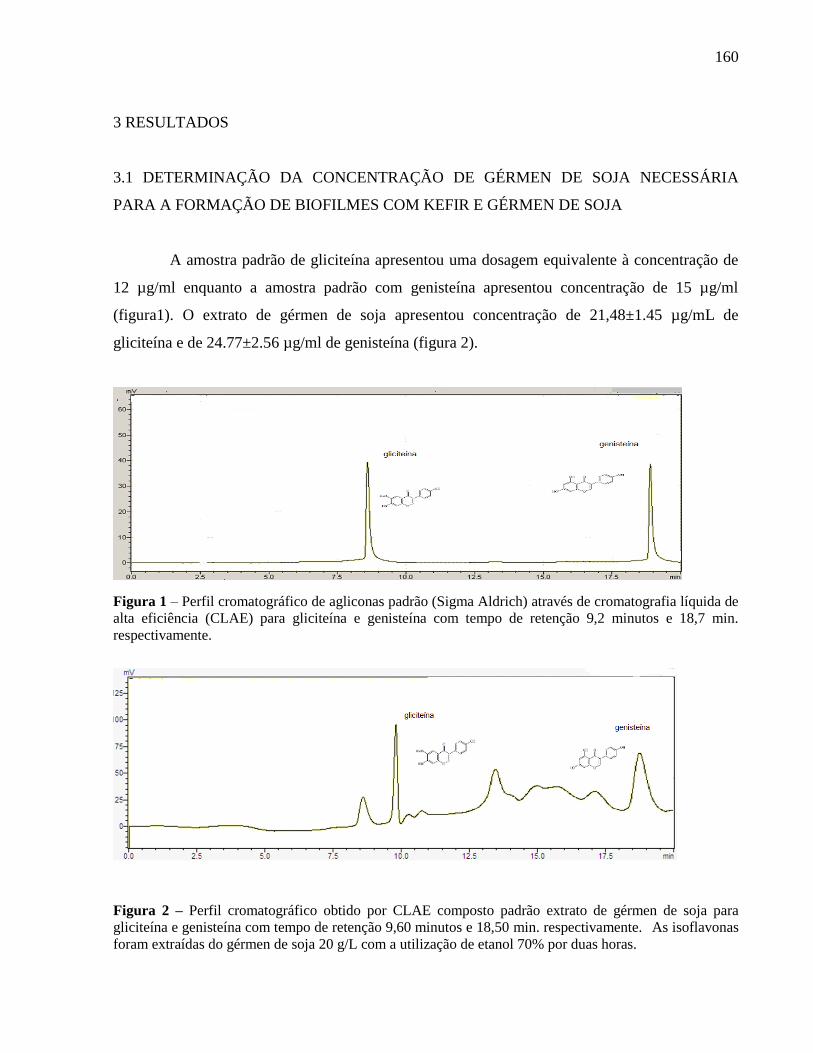
160
3 RESULTADOS
3.1 DETERMINAÇÃO DA CONCENTRAÇÃO DE GÉRMEN DE SOJA NECESSÁRIA
PARA A FORMAÇÃO DE BIOFILMES COM KEFIR E GÉRMEN DE SOJA
A amostra padrão de gliciteína apresentou uma dosagem equivalente à concentração de
12 µg/ml enquanto a amostra padrão com genisteína apresentou concentração de 15 µg/ml
(figura1). O extrato de gérmen de soja apresentou concentração de 21,48±1.45 µg/mL de
gliciteína e de 24.77±2.56 µg/ml de genisteína (figura 2).
Figura 1 – Perfil cromatográfico de agliconas padrão (Sigma Aldrich) através de cromatografia líquida de
alta eficiência (CLAE) para gliciteína e genisteína com tempo de retenção 9,2 minutos e 18,7 min.
respectivamente.
Figura 2 – Perfil cromatográfico obtido por CLAE composto padrão extrato de gérmen de soja para
gliciteína e genisteína com tempo de retenção 9,60 minutos e 18,50 min. respectivamente. As isoflavonas
foram extraídas do gérmen de soja 20 g/L com a utilização de etanol 70% por duas horas.

161
O aumento da concentração de gérmen de soja no meio de cultura coincidiu com o
aumento concomitantemente do conteúdo de isoflavonas quantificadas no mesmo. Assim
também, ocorreu que o crescimento da biomassa do biofilme foi maior com aumento de gérmen
de soja no meio (Tabela 1) mas não coincidiu exatamente com a incorporação de isoflavonas no
meio. A concentração de gérmen de soja de 10 g/L foi a que apresentou maior incorporação de
isoflavonas (39,63±0,64 µg/L de gliciteína e 42,24±0,4 µg/L de genisteína) (Tabela 1). O pH se
manteve constante e baixo entr 2,22 e 2,91 com 5g/L de gérmen de soja. (Tabela 1).
Também através da análise estatística constatou-se diferenças significante com relação à
incorporaçaõ de isoflavonas utilizando-se concentrações diferentes de gérmen de soja (ANOVA,
F = 207,39, p < 0,0001). Foi observado o aumento de incorporação de isoflavonas no meio de
cultura em concentrações crescentes de gérmen de soja atingindo o pico em 10 g/L (39,63±0,64
µg/L de gliciteína e 42,24±0,4 µg/L de genisteína). Houve diferença estatística entre presença de
isoflavonas apresentada nas concentrações de 20 mg/L (19,80±0,36 µg/L de gliciteína e
23,59±0,23 µg/L de genisteína) e a encontrada quando se utilizou a concentração de 10 mg/L
(Tabela 1).
Tabela 1 - Formação do biofilme com GK (20 g/L) e AM (40 g/L) em diferentes concentrações de
gérmen de soja (GS).
Conc. de GS (g) Cresc.(g)
média
isoflavonas pH Volume
gliciteína genisteína
0 6,75±0,14 0 2,28±0,02 480
1 10,06±0,26 7.65±0,20 9.57±0,10 2,29±0,01 485
2,0 17,30±1,02 11,54±0,10 10,41±0,06 2,40±0,02 480
5,0 7,63±0,46 17.50±0,07 13.75±0,09 2,91±0,02 475
10,0 16,52±0,95 39,63±0,64 42.24±0,40 2,33±0,01 465
20.0 46,37±0,4 19.80±0,36 23.59±0,23 2,43±0,02 465

162
Figura 3 – Perfil cromatográfico do meio de cultura com kefir 20 g/L com açúcar mascavo (40 g/L) mais
extrato de gérmen de soja a 10 g/L obtido através de cromatografia líquida de alta eficiência (CLAE) para
gliciteína e genisteína com tempo de retenção 9,8 minutos e 19,8 min. respectivamente.
3.2 DETERMINAÇÃO DA CONCENTRAÇÃO DE GRÃOS DE KEFIR NECESSÁRIA PARA
A FORMAÇÃO DE BIOFILMES COM GÉRMEN DE SOJA
Houveram diferenças estatísticas quando se utilizou diferentes concentrações de grãos
de kefir em meio de cultura com açúcar mascavo e gérmen de soja (ANOVA, F = 40,01 e valor
de p < 0,0001) e a melhor concentração objetivando formação de biofilme foi de 40 g/L
(44,49±2,09 g) (Tabela 2). Com relação à incorporação de isoflavonas a concentração de 60 g/L
foi a maior (32,65±3,98 µg/L de gliciteína e 39,89±3,25 µg/L de genisteína) (Tabela 2)
Tabela 2 - Formação do biofilme com gérmen de soja GS (40 g/L) e açúcar mascavo AM (40 g/L) em
diferentes concentrações de grãos de kefir (GK)
Conc. de GK
(g)
Cresc.(g)
média
isoflavonas pH Volume
gliciteína genisteína
10 23,87±0,15 8,76±0,67 9,44±1,43 4,03±0,01 485
20 19,48±0,35 10,35±0,54 11,78±2,55 3,76±0,02 480
40 44,49±2,09 17.48±2.41 14,09±1,45 3,80±0,15 475
60 32,85±0,95 32,65±3,98 39,89±3.25 3,80±0,01 465
80 32,55±0,70 19.59±3.57 22,78±8.21 3,77±0,0 465
100 34,39±0,45 22,34±1,54 23,45±2,23 3,49±0,02 471

163
Figura 4 – Foto de biofilmes de kefir (20 g/L) com açúcar mascavo (40 g/L) com gérmen de soja a) 20
g/L, b) 40 g/L, c) 80 g/L e d) 100 g/L
3.3 DETERMINAÇÃO DE PERÍODO NECESSÁRIO PARA INCORPORAÇÃO DE
ISOFLAVONAS
Constatou-se a presença de isoflavonas no meio de cultura com 5 dias (5,47±0.52 µg/L de
gliciteína e 7,4±0,49 µg/L de genisteína) atingindo o pico com 20 dias (Tabela 3). Além disso
com 5 dias ainda não houve formação de biofilmes, o que foi constatado de maneira perceptível
com 20 dias (19,59±3,57µg/L de gliciteína e 23,86±2,21µg/L de genisteína) (Tabela 3).
Tabela 3 – Determinação de período de tempo necessário para formação de biofilme e incorporação de
isoflavonas do biofilme de kefir associado ao germen de soja
Tempo
(Dias)
Biomassa (g) pH Vol (mL) Isoflavonas (µg/g)
gliciteina genisteina
5 2,95±0,24 3,66 204±3,51 5,47±0.52 7,4±0,49
15 5,81±0,23 3,57 185±1,67 17,51±0,47 18,56±0,63
20 10,54±1,10 3,49 188±1,27 19.59±3.57 23.86±2.21
Os números representam a média ± o desvio padrão

164
Figura 5 – Perfil cromatográfico obtido por CLAE de meio de cultura com kefir 20 g/L com extrato de
gérmen de soja 20 g/L cultivado por 5 dias para gliciteína e genisteína com tempo de retenção 9,60
minutos e 18,50 min. respectivamente.
3.4 DOSAGEM DE MACRO E MICROELEMENTOS DE BIOFILME COM GÉRMEN DE
SOJA
Quanto a composição de macro e microelementos foram encontrados no extrato de gérmen de
soja e nos biofilmes de kefir com gérmen de soja os seguintes componentes: Fe++ 0,916 mg/kg no
extrato do gérmen de soja e 2,54 mg/kg em biofilmes de kefir com gérmen, Mg++ 0,567 mg/kg m
gérmen de soja e 0,689 mg/kg em biofilme de kefir com extrato de gérmen de soja, Ca++ 0,836
mg/kg, em extrato de gérmen de soja e 0,740 mg/kg em biofilme de kefir com gérmen de soja. A
concentração de Zn++ foi 0,533 mg/kg em extrato de gérmen de soja e 0,654 mg/kg em biofilme
de kefir com extrato de gérmen de soja (Tabela 5). Não foi encontrado cobre.
Tabela 4 - Concentração de macro e microelementos do biofilme de kefir com gérmen de soja
Macro e microelementos GS
(µg/kg)
Biofilme com GS
(g/kg)
Mg++ 2,027 1,9514
Zn++ 0,1015 0,0845
Fe++ 0,1715 0,1689
Cu++ 0,1562 0,1264
Ca++ 0,1280 0,1660

165
3.5 ANÁLISE ESTRUTURAL BIOFILMES DE KEFIR E GÉRMEN DE SOJA POR
MICROSCOPIA DE FORÇA ATÔMICA E MICROSCOPIA ELETRÔNICA DE
VARREDURA (MEV)
Na análise de biofilmes por AFM, obteve imagens com diferentes concentrações de
gérmen de soja, que revelaram as configurações de rugosidade (RMS), de acordo com a
concentração (Figuras 6, 7 e 8). A rugosidade permaneceu sem alteração significativa não
dependendo, portanto da concentração de gérmen de soja no meio de cultura. A imagem de perfil
obtida em microscopia eletrônica de varredura (MEV) mostra a estrutura de formação destes
biofilmes com camadas se sobrepondo Figura 9).
,
Figura 6 – Imagem gráfica obtida através de microscopia de força atômica (AFM) mostrando a
rugosidade (0,287 µm) de biofilme de kefir formado 2 g/L de gérmen de soja. Observa-se a presença de
Lactobacillus
Figura 7 – Imagem gráfica de biofilme de kefir com gérmen de soja (10g/L) obtida com microscopia de
força atômica (AFM). O aumento da concentração não alterou a rugosidade (0, 283 µm).

166
Figura 8 – Imagem gráfica de biofilme de kefir com gérmen de soja (80g/L) obtida com microscopia de
força atômica (AFM). A rugosidade RMS (0,167 µm) naõ foi alterada significativamente com o aumento
da concentração de gérmen de soja..
Figura 9 - Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (20 g/L) com açúcar
mascavo (40 g/L) e gérmen de soja (20g/L) visualizada de perfil onde se observa as camadas que formam
os biofilmes se sobrepondo

167
Figura 10 - Fotomicrografia eletrônica de varredura (MEV) de biofilme de kefir (40 g/L) com açúcar
mascavo (40 g/L) com gérmen de soja (20 g/L) onde se visualiza hifa com moléculas adesivadas à mesma.

168
4 DISCUSSÃO
O processo fermentativo com kefir não resultou em diminuição na quantidade de
isoflavonas em relação à extração realizada com o extrato de gérmen de soja em procedimento
normal (com etanol a 70%). Enquanto o extrato de gérmen de soja apresentou a concentração de
21,48±1,45 µg/mL de gliciteína e de 24,77±2,56 µg/mL de genisteína (figura 2) a extração
realizada através do processo fermentativo com extrato de gérmens de sjoa e kefir resultou no seu
pico na concentração de 10 g/L para gliciteína 39,63±0,64 µg/L e para ginesteína 42,24±0,40
µg/L. Portanto, embora ROSSI et al. (2004) tenham reportado que o processo fermentativo
causou redução na quantidade de isoflavonas, o processo fermentativo com kefir não causou a
degradação das mesmas.
Para Xu et al. (2015) utilizando Lactobacillus plantarum CECT 748 T o conteúdo das
isoflavonas foi reduzido no processo fermentativos (de 3477 g/g para 794,84 g/g), tendo
ocorrido uma conversão das isoflavonas que passaram da forma glicosada para agliconas em
produtos fermentados de soja.
Com a utilização do kefir o processo fermentativo foi eficiente em extrair as isoflavonas
sem causar degradação das mesmas o que está de acordo com os dados obtidos por PURI et al.
(2015) que constataram que a fermentação de leite de soja com diferentes microrganismos pode
habilitar com mais eficiência o valor nutricional do que a fermentação utilizando monocultura
devido à correlação positiva entre o nível de enzimas e o conteúdo de isoflavonas.
O processo fermentativo pode alterar a biodisponibilidade das isoflavonas e também
pode converter a isoflavonas glicosadas em agliconas (YOO et al., 2015; XU et al., 2015),
compostos que efetivamente interessam para uso humano pois normalmente o organismo não
consegue absorver isoflavonas em formas glicosadas (OH, 2014).
É comum no processo fermentativo a redução de isoflavonas glicosadas por que o
radical glicosídeo é metabolizado pelos microrganismos (LANDETE et al., 2015). Por isso,
geralmente, no processo fermentativo ocorre um aumento de agliconas, o que é importante
considerando-se que as agliconas têm sido revelados como principais compostos da soja que
comportam propriedade terapêuticas (QUEIROZ, 2014). Tem sido confirmado que a genisteína
combate o câncer, além de assumir função de fitoestrógeno, enquanto que a gliciteína tem
despertado crescente interesse científico em função de suas atraentes propriedades biológicas

169
(ROH, 2014).
O uso do kefir ocasionou um efeito protetor no meio de cultura, impedindo a
desnaturação das proteínas da soja e, consequentemente, que ocorresse a putrefação. Quando o
processo fermentativo foi paralisado por algum fator (temperatura abaixo de 20°C, por exemplo)
ocorreu desnaturação protéica do extrato de gérmen de soja com consequente aumento do pH
(oscilando entre 6,5 a 7,5) e alteração de cor e odor do meio. Constatou-se também que, nestas
condições, as isoflavonas foram degradadas.
Houve diferença estatistica com relação à concentração de grãos de kefir utilizada
(p<0,01). O Teste Tukey confirmou que a melhor concentração de grãos para obtenção de
biomassa foi 40 g/L. As concentrações de 60 a 100 g/L naõ apresentaram diferenças estatísticas
quanto a formação de biomassa. A extração de isoflavonas que apresentou mais elevadas
dosagens foi obtida utilizando-se concentração de 60 g/L (32,65±3,98 µg/L de gliciteína e
39,89±3,25 µg/L de genisteína). O pH baixou de 6,5 no início do experimento para 3,49 na
concentraçãode 100g/L de grãos de kefir.
Utilizando a cultura de kefir para fermentação de leite de soja, Bau et al. (2015)
observaram que o pH baixou para 4,9 e ocorreu aumento de bactérias ácido láticas e
bioconversão de formas glicosadas em agliconas. Para Chun et al. (2008) utilizando co cultura de
Lactobacillus paraplantaruam, Enterococcus faecium e Weissela sp em fermentação de leite de
soja o pH baixou até 3,8 e, em 12 horas apenas, ocorreu a bioconversão de formas glicosadas de
isoflavonas para agliconas.
O tempo para incorporação de isoflavonas do meio de cultura foi de 20 dias obtendo
19,59±3,57 µg/L de gliciteína e 23,86±2,21 µg/L de genisteína (Tabela 3, figura 3). Com 5 dias
houve extração de 5,47±0.52 µg/L de gliciteína e 7,4±0,49 µg/L de genisteína (Tabela 3 figura 3).
Chun et al. (2006) estudaram o processo fermentativo com leite de soja após apenas 12 horas.
Bau et al. (2015) usou a cultura do kefir para fermentar o leite de soja por 30 horas resultando em
0,28 µg/g de gliciteína e 1,67 µg/g de genisteína. Anderson et al (1998) confirmaram que doses
mais baixas de genisteína agem similarmente a estrógenos com um efeito benéfico ao tecido
ósseo, mas em doses elevadas, podem exercer efeitos potencialmente adversos às funções
celulares dos tecidos ósseos.
Com relação à composição de macro e microelementos foi encontrado Fe++ 0,916 mg/kg
no extrato do gérmen e bem mais em biofilmes de kefir com gérmen de soja (2,54 mg/kg)

170
(Tabela4) decorrente, provavelmente do fato que o kefir foi cultivado em açúcar mascavo que é
rico em ferro.
Constaou-se também a incorporação de Mg++, tendo sido encontrado 0,567 mg/kg em
gérmen de soja e 0,689 mg/kg em biofilme de kefir associado ao extrato de soja. Em biofilmes de
kefir formado apenas em cultivo com açúcar mascavo a quantidade de Mg++ encontrada foi
inferior que em biofilmes de kefir associado ao extrato de gérmen desoja, o que sugere que o
magnésio foi incorporado em biofilmes proveniene tanto do açúcar mascavo como do extrato de
soja. Vale ressaltar que o magnésio tem grande importância em várias reações de anabolismo.
De igual maneira constaotuse que o Zn++ foi encontrado tanto no extrato de soja (0,533
mg/kg) como nos biofilmes de kefir associado ao extrato de gérmen de soja (0,654 mg/kg)
(Tabela4). A presença de zinco é de relevante importância nos biofilmes de kefir associado ao
gérmen de soja pois sabe-se que o Zn++ aumenta o efeito anabólico no metabolismo ósseo quando
adicionado a isoflavonas, notadamente, genisteína (YAMAGUCHI; GAO, 1998).
Ibramugy e Chibale (2013) encontraram os seguintes macroelementos nos grãos de soja:
fósforo, 0,52 mg/100g, ferro, 16,02 mg/ 100g, potássio, 1671,86 mg/100g, sódio, 67,16 mg/100g
e apenas traços de cálcio, paradoxalmente os resultados mostraram que a soja é uma significativa
fonte de cálcio para o organismo tendo sido encontrado no extrato e no gérmen de soja (0,836
mg/kg) como nos biofilmes (0,740 mg/kg) (Tabela4). Sgarbieri et al. (1981) em 100 g de amostra
em base seca encontraram 226 mg de cálcio. Este elemento é fundamental em prevenção e
tratamento de osteoporose, pois uma uma dieta provendo o cálcio, com uso de alimentos
contendo isoflavonas da soja e ômega 3, vitaminas D e K com atividade física regular e na
ausência do tabagismo mantém a saúde óssea e retarda ou mesmo evita a osteoporose. (ISHIMI,
2015; PAWLOWSKI et al., 2015). Há confirmação que o consumo de soja devido a sua alta
quantidade de isoflavonas evita a osteoporose através da inibição da reversão do cálcio. De
acordo com Zheng et al. (2016) as isoflavonas da soja têm efeitos sobre os ossos afetando a
modulação da remodelagem dos ossos evitando a perda óssea.
Através da análise de biofilmes por AFM, as imagens obtidas de biofilmes obtidos com
diferentes concentrações de gérmen de soja revelaram que as configurações de rugosidade (RMS)
foram dependentes da concentração (Figuras 6, 7 e 8). A rugosidade permaneceu sem alteração
significativa não dependendo, portanto da concentração de gérmen de soja no meio de cultura
(Figuras 6, 7 e 8). Com o uso de extrato de açaí (EHEEo) houve a relação dependente da

171
concentração do mesmo com a rugosidade. Sugere-se portanto que a concentração de açaí altera o
biofilme no parâmetro rugosidade devido ao açaí (Euterpe oleraçae Martius) ser rico em
compostos energéticos que podem afetam positivamente a rugosidade ou seja a estutura do
biofilme. Já na soja (Glycyne max (L) Merril predomina proteínas que não são utilizados como
fonte de carbono.
Condições de estresse, alteração de pH e disbiose dificultam a absorção intestinal de
compostos e de drogas. O sistema de liberação e absorção pode ser mantido salutar através da
ingestão e constante reposição de probióticos que mantêm as colônias da microbiota. Portando a
ingestão concomitante de probióticos com drogas pode ser uma eficaz estratégia para se
conseguir uma microbiota saudável em homeostase, e assim, se obter eficiente absorção de
fármacos (PRUDHVIRAJ et al., 2015) e também de compostos fenólicos cujos efeitos biológicos
dependem do seu mecanismo de ação no corpo que é afetado pela bioconversão pela microbiota
do cólon (BREYNAERT et al., 2010). Os biofilmes de kefir incorporaram isoflavonas
apresentando as condições para manter a homeostase intestinal para viabilizar a absorção eficaz
das isoflavonas.

172
5. CONCLUSÃO
Os biofilmes de kefir incorporaram isoflavonas e o processo fermentativo com os
grãos de kefir não reduziu a quantidades de isoflavonas. A melhor concentração de grãos de
kefir para produção de biomassa foi de 40 g/L, mas para a incorporação de agliconas a melhor
concentração foi de 60g/L. Com relação ao tempo de cultivo para a maior incorporação de
isoflavona nos biofilmes foram necessários 20 dias de cultivo. Nos biofilmes ocorreu a
incorporação de magnésio e zinco em maior quantidade do que a encontrada no gérmen de
soja. Não ocorreu alteração de rugosidade com o aumento da concentração de gérmen de
soja.

173
CONSIDERAÇÕES FINAIS
O estudo de kefir, evidentemente não é consumado com os resultados apresentados nesta
tese, porque trata-se de um tema de grande amplitude e relevância na área dos probióticos. Aqui
estudou-se a otimização da produção de biofilmes do kefir, como meio principal para
incorporação e veiculação de compostos bioativos de origem vegetal.
Constatou-se neste estudo a obtenção de biofilmes de kefir com consistência quando se
utilizou 20 g/L de grão de kefir com solução de açúcar mascavo 40 g/L. Já com solução de açúcar
branco foi necessário apenas soluções com 20 g/L. Com referência aos parâmetros cinéticos, a
temperatura ótima para obtenção dos biofilmes foi de 30°C e o pH estabilizou-se em torno de 2,8.
Em relação aos compostos voláteis majoritários presentes no meio de cultivo do kefir, por
cromatografia a gás detectou-se a predominância de compostos da classe dos ácidos carboxílicos,
ácidos graxos e hidrocarbonetos alifáticos e confirmou-se por espectrometria de absorção
atômica, que os biofilmes incorporaram macro e microelementos, notadamente o cálcio em
biofilmes sintetizados com solução de açúcar branco e o ferro em biofilmes sintetizados em
solução de açúcar mascavo. Na análise por AFM esta revelou que a rugosidade do biofilme é
dependente da concentração de grão de kefir.
Obteve-se biofilmes com extrato hidroetanólico de Euterpe oleracea (BEHEEo), os quais
preservaram as antocianinas do extrato hidroetanólico de açaí (EHEEo), e a análise por AFM
destes biofilmes confirmou que ocorre aumento da rugosidade do BEHEEo dependente da
concentração de EHEEo, concomitantemente mostrando Lactobacillus e leveduras de maneira
profícua nestes biofilmes.
Confirmou-se que os BEHEEo incorporaram e retiveram as antocianinas do EHEEo por
períodos relativamente longos (26 meses) e que em condições in vitro (pH 7,0, temperatura 38°C)
as antocianinas foram liberadas do BEHEEo.
Através da análise por microscopia eletrônica de varredura (SEM) do BEHEEo foi
possível constatar a coadesão das bactérias aos fungos filamentosos, evidenciando a importância
dessa interação para o desenvolvimento dos biofilmes.
Concernente à utilização de gérmen de soja no meio de cultura com kefir ocorreu a
formação de biofilmes os quais incorporaram isoflavonas (genisteína e gliciteína) e o processo
fermentativo não causou a redução de sua quantidade. Constatou-se que a concentração de grãos

174
de kefir necessária para obtenção de biofilmes com gérmen de soja foi de 40 g/L com relação à
biomassa, porém com 60 g/L de gérmen de soja obteve-se a maior quantidade de isoflavonas
incorporadas nos biofilmes. Foram necessários 20 dias para maior incorporação de isoflavonas do
gérmen de soja no meio de cultivo com kefir. A rugosidade dos biofilmes não foi dependente da
concentração de gérmen de soja.
Portanto, em decorrência da análise dos resultados obtidos conclui-se que através da
aplicação dos parâmetros estabelecidos neste estudo, biofilmes de kefir podem ser produzidos e
que a incorporação de compostos bioativos nestes biofilmes se torna factível, obtendo-se um
probiótico Amazônico com imenso potencial de aplicação terapêutica.

175
REFERÊNCIAS
ARAÜJO, J. M. A.; CARLOS, J. C. S.; SEDYAMA, C. S. Isoflavonas em grãos de soja:
importância da atividade de B-glicosidase na formação do sabor amargo e
adstringente. Ciênc. Tecnol. Alim. v. 17, p. 137-141, 1997.
AUSSENAC, T. et al. Quantification of isoflavones bycapillary zone electrophoresis in soybean
seeds: effects of variety and environment. Am. J. Clin. Nutr. v. 68, p. 1 480S-1 485S, 1998.
ANDERSON, J. J. B., GARNER, S. C. Phytoestrogens and human function. Nutrition Today, v.
32, n. 6, p. 232-239, 1997.
ASHFORD, M.; FELL, J. T.; ATTWOOD, D.; SHARMA, H.; WOODHEAD, P. J. Studies on
pectin formulations for colonic drug delivery. J. Control. Release, v. 30, p. 225-232, 1994
BARNES, S.; KIM, H.; XU, J. Soy in the prevention and treatament of chronic diseases. In:
CONGRESSO BRASILEIRO DE SOJA, 1999, Londrina. Anais... Londrina: Embrapa CNPSo,
1999, p. 295-310. (Embrapa CNPSo. Documentos, n. 124).
BAÚ, T. R.; GARCIA, S.; IDA, E. I. Changes in soymilk during fermentation with kefir culture:
oligosaccharides hydrolysis and isoflavone aglycone production. Int J Food Sci Nutr., v. 66, n 8,
p. 845-850, 2015.
BREYNAERT, A.; BOSSCHER, D.; KAHNT, A.; CLAEYS, M.; COS, P.; PIETERS, L.;
HERMANS, N. Development and Validation of an in vitro experimental GastroIntestinal
Dialysis Model with Colon Phase to Study the Availability and Colonic Metabolisation of
Polyphenolic Compounds. Planta Med. v. 81, n. 12-13, p.1075-83, 2015.
BURDA, S.; OLESZEK, W. Antioxidant and antiradical activities of flavonoids, Journal of
Agricultural and Food Chemistry, v.49, p.2774-2779, 2001.
CARRÃO-PANIZZI, M. C.; BERHOW, M.; MANDARINO, J. M. G.; OLIVEIRA, M. C. N.
Enviromental and genetic variation of isoflvones content of soybean seeds grown in Brazil.
Pesquisa Agropecuária Brasileira, Brasília, v. 44, n. 11, p. 1444-1451, 2009ª
CARRÃO-PANIZZI, M. C.; FAVONI, S. P. G.; KIKUCHI, A. Extraction time for isoflvone
determination. Brazilian Archives of Biology and Technology, v. 45, n. 4, p. 515-518, 2002.
CÉSAR, I. C.; BRAGA, F.C., VIANNA-SOARES, C. D.; NUNAN, E. A.; BARBOSA, T. A. F.;
MOREIRA-CAMPOS, L. M. M. Determinação de daidzeína, genisteína e gliciteína em cápsulas
de isoflavonas por cromatografia em camada delgada (CCD) e cromatografia líquida de alta

176
eficiência (CLAE). Brazilian Journal of Pharmacognosy, v. 17, n. 4, p. 616-625, 2007.
CHUN, J.; JEONG, W. J.; KIM, J. S.; LIM, J.; PARK, C. S.; KWON, D. Y.; CHOI, I.; KIM, J.
H. Hydrolysis of isoflavone glucosides in soymilk fermented with single or mixed cultures of
Lactobacillus paraplantarum KM, Weissella sp. 33, and Enterococcus faecium 35 isolated from
humans. Journal of Microbiology & Biotechnology, v. 18, n. 3, p. 573-578, 2008.
COSTA, P., LOBO, J. M. S. Formas farmacêuticas de liberação modificada. Rev. Port. Farm.,
Lisboa, v. 59, n. 4, p. 181-190, 1999.
COWARD, L.; BARNES, N. C.; SETCHELL, K. D. R.; BARNES S. Genistein, Daidzein, and
their b-glucoside conjugates: Antitumor isoflavones in soybean foods from American and Asian
diets. Journal of Agricultural and Food Chemistry, v. 41, p. 1961-1967, 1993.
CROZIER, A.; JAGANATH, I. B.; CLIFFORD, M. N. Dietary phenolics: chemistry,
bioavailability and effects on health. Nat Prod Rep, v. 26, n. 8, p. 1001-1043, 2009.
DAVEY, M. E., O'TOOLE, G. A. Microbial biofilms: from ecology to molecular genetics.
Microbiology and Molecular Biology Reviews, v. 64, n. 847, 2000.
DRESSMAN, J. B., AMIDON, G. L., REPPAS, C., SHAH, V. P. Dissolution testing as a
prognostic tool for oral drug absorption: immediate release dosage forms. Pharm. Res., New
York, v. 15, n. 1, p. 11-22, 1998.
ELDRIDGE, A.C.; KWOLEK, W.F. Soybean isoflavones: effects of envirnment and variety on
composition. J. Agric. Food Chem., v. 31, p. 394-396, 1983.
FARNWORTH, E. R. Kefir – a complex probiotic. Food Science and Technology Bulletin:
Functional Foods, v. 2, p. 1-17, 2005.
FREIRE, A. C.; PODCZECK, F.; SOUSA, J.; VEIGA, F. Liberação específica de fármacos no
cólon por via oral. II - Tipos de sistemas utilizados. Revista Brasileira de Ciências
Farmacêuticas v. 42, n. 3, 2006.
GALATI, G.; O’ BRIEN, P. J. Potenctial toxicity of flavonoids and other dietary phenolics:
significance for their chemo preventive and anticancer properties. Free Radical Biology &
Medicine. v. 37, n. 3, p. 287-303, 2004.

177
GARCÍA-LAFUENTE, A.; GUILLAMÓN, E.; VILLARES, A.; ROSTAGNO, M. A.;
MARTÍNEZ, J. A. Flavonoids as anti-inflammatory agents: implications in cancer and
cardiovascular disease. Inflamm Res., v. 58, p. 537–552, 2009.
GÓES-FAVONI, S. P.; CARRÃO-PANIZZI, M. C.; BELÉIA, A. D. P. Changes of isoflvone in
soybean cotyledons soaked in different volumes of water. Food Chemistry, Oxford, v. 119, p.
1605-1612, 2010.
GORBUSHINA, A.A., BROUGHTON, W.J. Microbiology of the atmosphere-rock interface.
how biological interactions and physical stresses modulate a sophisticated microbial ecosystem.
Annu. Rev. Microbiol. v. 63, p. 431–450, 2009.
HANSON-RESEARCH-CORPORATION. Dissolution: Past, Present & Future. 2. ed.
Chatsworth: Hanson Research, 1996. p. 2-12.
HOUSTON, M.C. Nutraceuticals, vitamins, antioxidants, and minerals in the prevention and
treatment of hypertension. Progress in Cardiovascular Diseases, v.47, n.6, p.396-449, 2005.
IBRAMUGY, A. D.; CHIBALE, M. A. avaliação físico-química dos grãos de soja Laborat.
Nacion. Higien. Aliment. Aguas, p. 6, 2013 Acessado dia 24-01-2016, 20:00 hs ver HTLM
IMAI, S. Soybean and processed soy foods ingredients, and their role in cardiometabolic risk
prevention. Recent Pat Food Nutr Agric v. 7, n. 2, p. 75-92, 2015.
ISHIMI, Y. Osteoporosis and Lifestyle. Journal Nutr Sci Vitaminol., v. 61, p. 139-141, 2015.
KING, A.; YOUNG, G. Characteristics and occurrence of phenolic phytochemicals. Journal of
American Diet. Association, v.99, p.213-218, 1999.
KUDOU, S.; FLEURY, Y.; WELTI, D.; MAGNOLATO, D.; UCHIDA, T.; KITAMURA, K.;
OKUBO, K. Malonyl isoflvone glycosides in soybean seeds (Glycine maxMerrill). Agricultural
and Biological Chemistry, v. 55, n. 9, p. 2227-2233, 1991
LAUDANNA, E. Propriedades funcionais da soja. Informe Agropecuário, v. 27, n. 230, p. 15-
18, 2006.
LANDETE, M. J.; HERNÁNDEZ, T.; ROBREDO, S.; DUEÑAS, M.; DE LAS RIVAS, B.;
ESTRELLA, I.; MUÑOZ, R. Effect of soaking and fermentation on content of phenolic
compounds of soybean (Glycine max cv. Merit) and mung beans (Vigna radiata [L] Wilczek).
Int J Food Sci Nutr, v. 66, p. 203-209, 2015.

178
LAUREYS, D.; DE VUYST L. Microbial species diversity, community dynamics, and
metabolite kinetics of water kefir fermentation. Appl. Environ. Microbiol., v. 80, p. 2564-2572,
2014.
LIN, C. W.; CHEN, H. L.; LIU, J. R. Identification and Characterisation of lactic acid bacteria
and yeasts isolated from kefir grains in Taiwan. Australian journal of dairy technology, v. 54,
n. 1, p. 14-18, 1999.
MACHERAS, P.; REPPAS, C.; DRESSMAN, J. B. Biopharmaceutics of orally administered
dosage forms. Chiester: Taylor & Francis, 1995. p.75-91.
MANADAS, R.; PINA, M. E.; VEIGA, F. A dissolução in vitro na previsão da absorção oral de
fármacos em formas farmacêuticas de liberação modificada. Revista Brasileira de Ciências
Farmacêuticas Brazilian Journal of Pharmaceutical Sciences, v. 38, p. 375-399, 2002.
MANDARINO, J. M. G. Compostos Antinutricionais da Soja: Caracterização e
Propriedades Funcionais. In: COSTA, N. M. B.; ROSA, C. O. B. Alimentos funcionais:
componentes bioativos e efeitos. Rio de Janeiro: Rubio, 2010. p. 177-192.
MATSUDA, S. et al. Hydrolysis of isoflavones in soybean cooked syrup by Lactobacillus casei
subsp.rhaminosus IFO 3425. J. Ferm .Bioeng. , v.74, p.301-304, 1992.
MITHANI, S. D.; BAKATSELOU, V.; TENHOOR, C. N.; DRESSMAN, J. B. Estimation of the
increase in solubility of drugs as a function of bile salt concentration. Pharm. Res., v. 13, p. 163-
167, 1996.
NADELL, C. D.; DRESCHER, K.; WINGREEN, N. S.; BASSLER, B. L. Extracellular matrix
structure governs invasion resistance in bacterial biofilms. ISME Journal Advance, v. 55, n. 11,
p. 396-407, 1997.
OH, H. G.; NALLAMSHETTY, S.; RHEE, E. J. Increased Risk of Progression of Coronary
Artery Calcification in Male Subjects with High Baseline Waist-to-Height Ratio: The Kangbuk
Samsung Health Study. Diabetes Metab J., v. 40, n. 1, p. 54-61, 2016.
PARK, Y. K.; AGUIAR, C. L.; ALENCAR, S. M.; MASCARENHA, S. A.; SCAMPARINI, A.
R. P. Conversão de malonil-glicosil isoflavonas em isoflavonas glicosiladas presentes em alguns
cultivares de soja brasileira. Ciênc. Tecnol. Alim., v. 22, n.1 p. 30-35, 2002.
PAWLOWSKI, J. W.; MARTIN, B. R.; MCCABE, G. P.; MCCABE, L.; JACKSON, G. S.;
PEACOCK, M.; BARNES, S.; WEAVER, C. M. Impact of equol-producing capacity and soy-
isoflavone profiles of supplements on bone calcium retention in postmenopausal women: a
randomized crossover. Am. J. Clin. Nutr., v. 102, n. 2, p. 695-703, 2015.

179
PREECHAGOON, D.; UDOMPRATEEP, A.; MANWIWATTANAGUL, G. Improved
dissolution rate of poorly soluble drug by incorporation of buffers. Drug Dev. Ind. Pharm., v.
26, n. 8, p. 891-894, 2000.
PEETERS, R.; KINGET, R. Film-forming polymers for colonic drug delivery: I. Synthesis and
physical and chemical properties of methyl derivates of Eudragit S. Int. J. Pharm., v. 94, p. 125-
134, 1993.
PRUDHVIRAJ, G.; VAIDYA, Y.; SINGH, S. K.; YADAV, A. K.; KAUR, P.; GULATI, M.;
GOWTHAMARAJAN, K. Effect of co-administration of probiotics with polysaccharide based
colon targeted delivery systems to optimize site specific drug release. Eur J Pharm Biopharm,
v. 97, p. 164-172, 2015
PURI, A.; MIR, S. R.; PANDA, B. P. Effect of sequential bio-processing conditions on the
content and composition of vitamin K2 and isoflavones in fermented soy food, J Food Sci
Technol. v. 52, n. 12, p. 8228-8235, 2015.
QUEIRÓS, L. D. Biotransformação de compostos fenólicos do extrato de soja para obtenção
de produto rico em compostos bioativos, 2014. 84 p. Dissertação (mestrado) – Universidade
Estadual de Campinas, (UNICAMP) Faculdade de Engenharia de Alimentos. Campinas, 2014.
ROH, C. Microbial transformation of bioactive compounds and production of ortho-
dihydroxyisoflavones and glycitein from natural fermented soybean paste. Biomolecules, v. 4, p.
1093-1101, 2014.
ROSSI, E. A.; ROSIER, I.; DÂMASO, A. R.; CARLOS, I. Z.; VENDRAMINI, R. C.;
ABDALLA, D. S. P.; TALARICO, V. H.; MINTO, D. F. Determinação de isoflavonas nas
diversas etapas do processamento do “iogurte” de soja. Alim. Nutr., v. 1 5, p. 93-99, 2004.
SGARBIERI, V. C.; GARRUTI, E. C.; GUZMÁN, E. C. Soybeans as an extender of common
beans. JAOCS, v.58, n. 3, p. 522-526, 1981.
SHUAI, S; YUE, S.; HUANG, Q.; WANG, W.; YANG, J.; LAN, K.; YE, L. Preparation,
characterization and in vitro/vivo evaluation of tectorigenin solid dispersion with improved
dissolution and bioavailability. Eur J Drug Metab Pharmacokinet, 2015 IN PRESS [Epub ahead
of print]
STATGRAPHICS CENTURION XVI SOFTWARE (Statpoint Technologies, Inc, Warrenton, VA,
USA).
SUGIYAMA, A.; YAMAZAKI, Y.; YAMASHITA, K.; TAKAHASHI, S.; NAKAYAMA, T.;

180
YAZAKI, K. Developmental and nutritional regulation of isoflavone secretion from soybean
roots. Biosci. Biotechnol. Biochem., v.80, n. 1, p. 89-94, 2015.
TEZUKA, H.; IMAI, S. Immunomodulatory Effects of Soybeans and Processed Soy Food
Compounds. Recent Pat Food Nutr Agric, v. 7, n. 2, p. 92-99, 2015.
TREVISANATO, S. I.; KIM, Y. I. Tea and Health. Nutrition Reviews, v.58, p.1-11, 2000.
TSUKAMOTO, C.; SHIMADA, S.; IGITA, K.; KUDOU, S.; KOKUBUN, M.; KUBO, K.;
KITAMURA, K. Factors Affecting Isoflvone Content in Soybean Seeds: Changes in Isoflvones,
Saponins, and Composition of Fatty Acids at Different Temperatures during Seed Development.
Journal of Agricultural and Food Chemistry, Washington, v. 43, n. 5, p. 1184-1192, 1995.
http://dx.doi.org/10.1021/jf00053a0120, 2000.
UI, J. H.; QIAN, Y.; MIAO, W. J.; XIA, Q. Preparation and in vitro evaluation of gastroretentive
dosage form containing puerarin-HP-beta-CD inclusion complex. Zhongguo Zhong Yao Za
Zhi, v. 33, p. 1960-1963, 2008.
VARINSKA, L.; GAL, P.; MOJZISOVA, G.; MIROSSAY, L.; MOJZIS, J. Soy and breast
cancer: focus on angiogenesis. Int J Mol Sci, v. 16, p. 11728-11749, 2015.
WANG, H. J.; MURPHY, P. A. Isoflavone content in commercial soybean foods. J. Agric.Food
Chem., v. 42, p.1666-1673, 1994.
WANG, H. J. Quantification of potentially anticarcinogenic isoflavones in soy foods and the
effect of processing on the composition of isoflavones. Dissertation Abstracts Intern., B, v. 55,
n. 7, 128p. 1995.
XU, L.; DU, B.; XU, B. A systematic, comparative study on the beneficial health components
and antioxidant activities of commercially fermented soy products marketed in China. Food
Chem, v. 174, p. 202-13, 2015.
YAMAGUCHI, M.; GAO, Y. H. Anabolic effect of genistein and genistin on bone metabolism
in the femoral-metaphyseal tissues of elderly rats: the genistein effect is enhanced by zinc. Mol
Cell Biochem., v. 178, n. 1-2, p.377-382.
YEH, C. C.; FAN, Y.; JIANG, L.; YANG, Y. L.; HE, B.; YOU, L.; MANN, M. Genistein
Suppresses Growth of Human Uterine Sarcoma Cell Lines via Multiple Mechanisms. Anticancer
Res, v. 35, n. 6, p. 3167-3173, 2015.

181
YOO, H. W.; CHANG, M. J.; KIM, S. H. Fermented soybeans by Rhizopus oligosporus reduce
femoral bone loss in ovariectomized rats. Nutr Res Pract, v. 8, p. 539-543, 2015.
YU, F.; LIU, Z.; TONG, Z.; ZHAO, Z.; LIANG, H. Soybean isoflavone treatment induces
osteoblast differentiation and proliferation by regulating analysis of Wnt/β-catenin pathway.
Gene, v. 573, n. 2, p. 273-277, 2015.
ZHANG, W.; DAI, W.; TSAI, S. M.; ZEHNDER, S. M.; SARNTINORANONT, M.;
ANGELINI, T. E. Surface indentation and fluid intake generated by the polymer matrix of
Bacillus subtilis biofilms. Soft Matter, v. 11, n. 18, p. 3612-3617, 2015.
ZHENG, X.; LEE, S. K.; CHUN, O. K. Soy Isoflavones and Osteoporotic Bone Loss: A Review
with an Emphasis on Modulation of Bone Remodeling. Journal Med Food, v. 19, n. 1, p. 1-14,
2016.