universidad de cÁdiz facultad de ciencias del mar y
TRANSCRIPT

UNIVERSIDAD DE CÁDIZ
FACULTAD DE CIENCIAS DEL MAR Y AMBIENTALES
DEPARTAMENTO DE BIOMEDICINA, BIOTECNOLOGIA Y SALUD
PÚBLICA, EN EL ÁREA DE GENÉTICA
Optimización de programas de selección en acuicultura
TESIS DOCTORAL
TIAGO FERNANDES FARIAS
Cádiz, 2019


UNIVERSIDAD DE CÁDIZ
FACULTAD DE CIENCIAS DEL MAR Y AMBIENTALES
DEPARTAMENTO DE BIOMEDICINA, BIOTECNOLOGIA Y SALUD
PÚBLICA, EN EL ÁREA DE GENÉTICA
Optimización de programas de selección en acuicultura
Memoria presentada por Don Tiago Fernandes Farias para optar al Grado de Doctor en
Ciencias y Tecnologías Marinas
Fdo.: Tiago Fernandes Farias
Directores:
Dr. Jesús Fernández Martin
Científico Titular de OPIs
Departamento de Mejora Genética Animal
Instituto de Investigación y Tecnología
Agraria y Alimentaria (INIA)
Dr. Miguel Ángel Toro Ibáñez
Catedrático Universidad Politécnica de Madrid
Departamento de Producción Agraria
E.T.S. Ingeniería Agronómica, Alimentaria y
de Biosistemas

RESUMEN
La acuicultura continúa su avance y su desarrollo se consolida por todo el mundo.
A medida que la actividad viene expandiéndose, el espacio para implementación de
nuevos proyectos se torna cada vez más limitante, y por eso, hay una importante
necesidad de proponer estrategias que permitan aumentar su capacidad de biomasa
productiva por área. La mejora genética es una importante herramienta capaz de
contribuir el aumento de la producción y sostenibilidad de la actividad, sin embargo, hay
que tener en cuenta las características biológicas particulares en condiciones de cultivo
de cada especie, con lo cual es importante que estudios sean realizados con el objetivo de
aumentar el conocimiento científico para proveer informaciones importantes para el
desarrollo de técnicas aplicadas en la producción.
En este contexto, el estudio consistió en analizar ciclos reproductivos de las
poblaciones reproductoras de dorada de la población comercial de una empresa, con el
objetivo de desarrollar procedimientos más adecuados para llevar a cabo un programa de
mejora genética en dorada (Sparus aurata). Los datos del trabajo fueron realizados dentro
de las instalaciones de la empresa acuícola Acuicultura ABSA que es el criadero comercial
del Grupo Culmarex, situado en Palma de Mallorca, España. El programa de selección
está basado en el uso de marcadores moleculares para la determinación de las relaciones
parentales y control de la consanguinidad, teniendo la tasa de crecimiento como el
principal carácter a mejorar. La selección se realiza mediante la metodología de
predicción BLUP, utilizando técnicas de optimización, que posibilitan hacer restricciones
al incremento de la consanguinidad, con la finalidad de aumentar la respuesta a la
selección a largo plazo.
Los resultados constataron que se puede conseguir una reducción importante en el
nivel de parentesco de los individuos seleccionados sin perder mucho en valor genético,
fue observado un aumento en el diferencial de selección de 10,5% (I); 23,2% (II) y la
estabilidad de su incremento al 26% (III y IV) para las dos otras situaciones. Por lo tanto,
la metodología de contribuciones óptimas permitió obtener un diferencial de selección
elevado, controlando los niveles de pérdidas de variabilidad. Este resultado es

especialmente importante para la acuicultura, donde una pequeña fracción de
reproductores puede contribuir masivamente en la formación de la descendencia.


Índice
Pag
1. INTRODUCCIÓN ....................................................................................... 01
1.1. LA SITUACIÓN ACTUAL DE LA ACUICULTURA .................... 01
1.1.1. EL DESARROLLO DE LA BIOTECNOLOGÍA EN LA
ACUICULTURA .......................................................................... 03
1.2. LA DORADA (SPARUS AURATA) .................................................. 05
1.2.1. LA CARACTERIZACIÓN DE LA ESPECIE ............................ 05
1.2.2. EL CULTIVO INTENSIVO DE LA DORADA .......................... 08
1.3. LA MEJORA GENÉTICA EN LA ACUICULTURA ...................... 10
1.3.1. LOS CARACTERES CUANTITATIVOS ................................... 11
1.3.2. LA SELECCIÓN ARTIFICIAL ................................................... 13
2. OBJETIVO GENERAL .............................................................................. 16
2.1. OBJETIVOS ESPECÍFICOS .................................................................. 17
3. MATERIAL Y MÉTODOS ........................................................................ 18
3.1. LA EMPRESA .................................................................................. 19
3.2. LOS ANIMALES .............................................................................. 21
3.2.1. LA POBLACIÓN FUNDADORA ............................................... 21
3.2.2. LOS REPRODUCTORES ............................................................ 22
3.2.3. LOS DESCENDIENTES .............................................................. 24
3.2.4. EL MANEJO Y LA TOMA DE MEDIDAS ................................ 26

3.3. LAS CONDICIONES DEL CRIADEO (HATCHERY) .................. 28
3.4. LA EXTRACCIÓN DEL ADN Y LOS ANÁLISIS DE LOS
MICROSATÉLITES ......................................................................... 30
3.5. LA ASIGNACIÓN DEL PARENTESCO ......................................... 32
3.6. ANÁLISIS DE LOS DATOS ............................................................ 33
3.7. LA OPTIMIZACIÓN POR TEMPLADO SIMULADO
(SIMULATED ANNEALING) ............................................................ 34
4. RESULTADOS ............................................................................................ 37
4.1. LA DESCRIPCIÓN ESTADÍSTICA DE LA POBLACIÓN ........... 38
4.2. LA CARACTERIZACIÓN DE LOS LOCI (MICROSATÉLITES).. 41
4.3. LA ASIGNACIÓN DE LA PATERNIDAD (GENEALOGÍA) ....... 43
4.3.1. LAS CONTRIBUCIONES DE LOS REPRODUCTORES ......... 46
4.4. LA ESTIMACIÓN DE LOS PARÁMETROS GENÉTICOS .......... 48
4.4.1. LA HEREDABILIDAD Y CORRELACIONES (GENÉTICAS
Y FENOTÍPICAS) ........................................................................ 49
4.4.2. LAS ESTIMACIONES DEL EFECTO FIJO (FOTOPERIODO) 50
4.5. LA SELECCIÓN POR OPTIMIZACIÓN ........................................ 51
5. DISCUSIÓN ................................................................................................. 54
5.1. LOS PROGRAMAS DE MEJORA GENÉTICA EN DORADA ..... 55
5.2. OPTIMIZACIÓN DE LA SELECCIÓN CON RESTRICCIONES. 64

5.3. CONSIDERACIONES FINALES .................................................... 72
6. CONCLUSIONES ....................................................................................... 75
7. REFERENCIAS BIBLIOGRÁFICAS ....................................................... 77

Lista de Figura
Pag
Figura: 1.01 La evolución de la producción de la acuicultura en España, en
toneladas y por especies, en el periodo 1960-2017 (Apromar,
2018) .............................................................................................. 02
Figura: 1.02 Características morfológicas de un ejemplar adulto de Sparus
aurata ............................................................................................. 06
Figura: 1.03 Los dientes y mandíbulas de un ejemplar adulto de Sparus aurata
(Moretti, et al., 1999) ..................................................................... 06
Figura: 1.04 Distribución de Sparus aurata de acuerdo a la probabilidad de su
ocurrencia (Fish Base, 2018) .......................................................... 07
Figura: 1.05 Esquema representativo del ciclo de vida de la dorada, S. aurata,
en sistema de cultivo intensivo (FAO, 2005) .................................... 10
Figura: 1.06 La selección en la curva de distribución normal, donde, (XP) es el
promedio de la población original, (XS) el promedio de población
seleccionada y (DS) es denominado como el diferencial de
selección ........................................................................................ 13
Figura: 3.01 Localización geográfica de las empresas que componen el
GRUPO CULMAREX (http://www.culmarex.com/granjas-
marinas) .…………………………………………………………………………………………… 19
Figura: 3.02 Una ilustración gráfica del barco Wellboat, utilizado para
transporte y la captura de individuos (Fuente: www.wellboat.cl) .. 20
Figura: 3.03 Vista aérea de las instalaciones de Acuicultura Balear SAU, en
Palma de Mallorca, Islas Baleares (fuente: Google Earth) ............. 21

Figura: 3.04 Los tanques de reproductores de uno de los fotoperiodos.
Natación en círculos y agrupados, movimientos característicos de
desove masal .................................................................................. 23
Figura: 3.05 La mesa de descartes morfológicos ................................................ 25
Figura: 3.06 Tanques circulares de juveniles ...................................................... 26
Figura: 3.07 A: La solución de Eugenol (anestésico)
B: Inmersión de los animales en la solución ................................... 27
Figura: 3.08 La bancada de las mediciones biométricas ..................................... 28
Figura: 3.09 Caja colectora de huevos ................................................................ 29
Figura: 3.10 Flujograma del templado simulado .................................................... 36
Figura: 4.01 Histograma del peso(g) y longitud (cm) en las dos edades medidas 38
Figura: 4.02 La frecuencia relativa en la amplitud alélica de los loci en el
primer multiplex ............................................................................ 42
Figura:4.03 La frecuencia relativa en la amplitud alélica de los loci en el
segunda multiplex .......................................................................... 42
Figura: 4.04 Número de individuos asignados en cada tanque dentro de cada
fotoperiodo. La línea roja es el promedio, y el área roja rayada es
la desviación estándar. La columna azul clara es la segunda
multiplex ........................................................................................ 44
Figura: 4.05 La cantidad de reproductores machos (A) y hembras (B), que
estaban disponibles (inicial) para la reproducción y los que han
contribuido (final) .......................................................................... 47
Figura: 4.06 La contribución de descendientes de cada reproductor macho
(azul) y hembra (rojo). La línea trazada representa el promedio de
la contribución de cada tanque. ...................................................... 48

Figura: 4.07 La optimización de la selección para los diferentes niveles de
restricción para la consanguinidad. Donde el eje Y son los valores
mejorantes estimados (Estimated Breeding Values - EBV´s) y en
el eje X está el parentesco promedio de la población seleccionada
(f) ……………………………………………………………………………………………………… 51
Figura: 5.01 El cronograma paso a paso de los procedimientos realizados en el
programa de mejora por selección aplicados en el cultivo de la
dorada (Sparus aurata). 1) Plantel de reproductores; 2) Toma de
datos (moleculares y biométricos); 3) Construcción de la
genealogía; 4) Análisis de datos (bioinformática y estadística); 5)
Evaluación y selección de los individuos; 6) Clasificación y
reorganización de los reproductores del nuevo plantel. .................. 59
Figura: 5.02 El porcentaje de individuos asignados con la primera ronda de
multiplex, con la segunda, los que no han sido posible asignar y
los análisis ahorrados/evitados utilizándose rondas consecutivas
de multiplexes. ..................................................................................... 61
Figura: 5.03 Diferencial de selección (DS) de cada fotoperiodo (expresado en
porcentaje de la población inicial), en las cuatro situaciones de
parentesco global propuesto. .......................................................... 69
Figura: 5.04 La optimización con restricciones para diferentes cantidades (%)
de individuos seleccionados. .......................................................... 70

Lista de Tablas
Pag
Tabla: 3.01 Número de reproductores que componen cada fotoperiodo. En la
horizontal están los grupos de animales ordenados de acuerdo con
su fecha de nacimiento. En la vertical están los fotoperiodos y las
fechas en que han sido formados. .......................................................... 22
Tabla: 3.02 Número de reproductores (machos y hembras) distribuidos en cada
uno de los dos tanques, que componen cada fotoperiodo .................. 24
Tabla: 3.03 Lista de los loci microsatélites utilizados para la asignación de la
paternidad. ....................................................................................... 31
Tabla: 4.01 El promedio (𝒙), la desviación estándar (DS) y el coeficiente de
variación (CV) de las características fenotípicas medidas. ……..…. 40
Tabla: 4.02 La cantidad de alelos distintos encontrados en cada locus, su
amplitud total (pb) y los alelos más frecuentes (% incidencia) de
los microsatélites en el global de individuos genotipados. ............... 41
Tabla: 4.03 Número de individuos asignados por cada fotoperiodo, utilizando
la 1ª y la 2ª multiplex (en paréntesis el porcentaje de asignación) .. 43
Tabla: 4.04 Número de machos y hembras disponibles y las progenies
asignadas, en los casos en que se han utilizado 4 y 8 microsatélites.. 45
Tabla: 4.05 Correlaciones fenotípicas (debajo de la diagonal; rojo),
correlaciones genéticas ± desviación estándar (arriba de la
diagonal; azul) y heredabilidad ± desviación estándar (diagonal;
negrita) estimados con los tres softwares, (VCE, TM e
REMLF90). ……………………………………………………………………………………….. 49
Tabla: 4.06 Estimación de los efectos de factores fijos de los fotoperiodos para
los diferentes caracteres ................................................................... 50

Tabla: 4.07 Los valores promediados del peso 2 de la población inicial
(genealogía asignada) y el promedio de los seleccionados (5% de
los individuos), aplicando cuatro niveles de restricción propuestas
para parentesco promedio de la población seleccionada (f)................ 53
Tabla: 5.01 Estimación de la heredabilidad de los caracteres medidos, por cada
fotoperiodo separadamente, utilizando los datos del VCE ............... 65

1.
INTRODUCCIÓN

1
1. INTRODUCCIÓN
1.1. LA SITUACIÓN ACTUAL DE LA ACUICULTURA
La acuicultura puede ser definida como la producción en cautividad de animales
(peces, moluscos, crustáceos, reptiles, equinodermos) o plantas que tengan un hábitat
predominantemente acuático, por lo menos en una fase de su vida. Consideramos como
un cultivo cuando es posible almacenarlos y mantenerlos hasta que alcancen las
condiciones necesarias para ser consumidos y/o comercializados. Básicamente es cerrar
el ciclo de vida completo o parcialmente en condiciones controladas, que van desde la
reproducción, alimentación y cosecha en ambientes controlados. Para que sea
considerado un producto de origen acuícola, es necesario que por lo menos en algún
momento de su ciclo de vida, haya habido por algún motivo la intervención humana.
En general, el objetivo de la acuicultura es el aumento de la producción a través de
prácticas como la alimentación artificial, la protección contra predadores, la integración
con otras especies, el control poblacional y la mejora genética. En realidad, la acuicultura
está relacionada con la agricultura y la ganadería, ya que implica la cría y el cultivo y
desarrollo de tecnologías que posibiliten la producción de proteína animal en ambientes
acuáticos. Además, la biomasa productiva por áreas de superficie que se puede alcanzar
en estos cultivos es mucho mayor que los cultivos de animales terrestres, indicando que
la producción de organismos acuáticos tiene un importante protagonismo para suplir esa
creciente demanda por productos pesqueros. Por lo tanto, el desarrollo de la acuicultura
tiene una importante relevancia como motor en el desarrollo económico capaz de
proporcionar seguridad alimentaria a nivel mundial.
La acuicultura se mantiene como uno de los sectores de la producción de alimentos
que más ha crecido en los últimos años. De acuerdo con el último informe Apromar
(2018), En 2016, España fue el estado miembro de la Unión Europea con una mayor
cosecha de acuicultura (283.831 t). Para 2017 las cifras de cosecha de acuicultura en
España son por un total 345.635 toneladas y valor de primera venta en 452,6 millones de
euros. La principal especie producida en España ha sido el mejillón (273.517 t), seguida
por la lubina (21.269 t), la trucha arco iris (17.948 t) y la dorada (13.643 t). (Figura: 1.01)

2
Figura: 1.01 La evolución de la producción de la acuicultura en España, en
toneladas y por especies, en el periodo 1960-2017 (Apromar, 2018)
El desarrollo de la acuicultura continúa su avance y consolidación por todo el
mundo, y eso se debe en gran parte a los importantes esfuerzos que vienen siendo
realizados con el objetivo de proponer alternativas que sean capaces de elaborar sistemas
de producción cada vez más eficientes. En las últimas décadas ya se han logrado
resultados importantes basadas en la elaboración de piensos cada vez más nutritivos y
específicos, y nuevas técnicas de manejo, en el control de enfermedades y más
recientemente el aprovechamiento de los recursos genéticos.
A medida que la actividad viene expandiéndose, el espacio para implementación de
nuevos proyectos acuícolas se torna cada vez más limitante, por eso la necesidad de
proponer estrategias que permitan aumentar su capacidad de biomasa productiva
ocupando el mismo espacio. Por lo tanto, para que la actividad pueda seguir creciendo a
ritmo acelerado, aún falta superar una serie de grandes desafíos, y para aumentar su
eficiencia y productividad, el sector debe dirigir sus esfuerzos hacia iniciativas de
investigación e innovación capaces de ampliar los conocimientos necesarios.
La producción mundial viene experimentando una nueva revolución industrial,
donde la biotecnología está actuando en prácticamente todos los procesos productivos,
elaborando alternativas capaces de reducir costos y desarrollando productos cada vez más
eficientes, aproximando cada vez más los intereses del sector productivo a los científicos.

3
Y la mejora genética es considerada una importante herramienta tecnológica, capaz de
contribuir en gran medida con la eficiencia de la producción acuícola y en el aumento de
la sostenibilidad de la actividad. A continuación, vamos ver cómo el desarrollo de la
biotecnología viene beneficiando los índices de producción en la acuicultura.
1.1.1. EL DESARROLLO DE LA BIOTECNOLOGÍA EN LA
ACUICULTURA
La acuicultura es una antigua forma de cultivo que data de 2000 años en China y
en el Imperio Romano (Balon, 1995; Dunham, et al., 2001). Sin embargo, solo en las
últimas décadas la acuicultura se ha convertido en una práctica global a nivel mundial.
La producción ha crecido dramáticamente desde principios de los años ochenta, y será
cada vez más importante a medida que aumente la demanda de productos pesqueros, sin
perjudicar a las poblaciones naturales. Actualmente, la acuicultura ya representa más del
50% del consumo mundial de pescado (FAO, 2009).
La investigación genética y su aplicación han tenido un papel significativo en el
desarrollo de la acuicultura, y el impacto será cada vez más importante a medida que va
siendo desarrollada la biotecnología aplicada a la acuicultura. Aunque sea una actividad
reciente, ya presenta resultados relevantes en varias especies acuícolas, y la expectativa
es que sea posible lograr mejoras aún mucho mayores. (Dunham, et al., 2001).
Importantes avances empezaron a surgir en las últimas décadas, trayendo consigo
importantes progresos en el área de la biología molecular, juntamente al desarrollo de la
informática, están se convirtiendo en herramientas de enorme potencial en la
caracterización de los recursos genéticos. El desarrollo de los microsatélites para especies
con importancia en acuicultura ha crecido enormemente en los últimos años dada su
utilidad para la identificación y diferenciación de stocks, control de la endogamia y grado
de parentesco, mapas de ligamiento, programas de selección genética asistida por
marcadores o para manipulaciones genéticas relacionadas con poliploidías o ginogénesis
(Martínez y Figueras, 2007)

4
Actualmente, la acuicultura viene experimentando progresos en la mejora de la
producción para una multitud de características, incluyendo tasa de crecimiento,
eficiencia de conversión alimenticia, aspectos reproductivos, resistencia a enfermedades,
tolerancia a la baja calidad del agua, tolerancia al frio, forma del cuerpo, rendimiento de
la canal y calidad de la carne. (Dunham, 2011)
La combinación de programas de mejoramiento genético, ingeniería tradicional,
biotecnológica e ingeniería genética, resulta en la utilización de mejores genotipos para
la acuicultura y en el manejo del plantel de reproductores. Y por ello, se están
implementando comercialmente una variedad de técnicas genéticas, que incluyen la
selección, los sistemas de apareamiento, la hibridación interespecífica, la reversión sexual
y la poliploidía, para mejorar la producción acuícola de peces y mariscos.
Para el desarrollo de una acuicultura sostenible, hay que tener en cuenta que cada
especie tiene una estructuración genética singular debido a la combinación de factores
biológicos, oceanográficos, ambientales y/o antropogénicos lo que implica que es
fundamental conocer las características particulares de cada especie para que puedan
reproducirse en cautividad. La dorada (Sparus aurata), por ejemplo, es una especie muy
apropiada para los cultivos en la región mediterránea. Debido a su buen precio de
mercado, alta tasa de supervivencia y hábitos alimentarios, viene experimentando una
rápida transformación en la industria. El éxito en la producción de esta especie, se debe
en parte a la viabilidad de los cultivos de alta productividad alcanzados en las jaulas
marinas. Ese resultado ha sido logrado con el aumento del conocimiento científico
ocurrido en las últimas décadas, que ha facilitado el desarrollo de técnicas eficientes
relacionadas al manejo, nutrición y genética, A continuación, vamos a profundizar en las
características biológicas de esta especie, así como los cultivos intensivos a gran escala,
que facilita su producción en los países del Mediterráneo.

5
1.2. LA DORADA (Sparus aurata)
1.2.1. LA CARACTERIZACIÓN DE LA ESPECIE
La dorada (Sparus aurata, Linnaeus, 1758) es un pez perciforme que pertenece a la
familia de los espáridos. Los miembros de la familia Sparidae contiene un gran número
de especies de varios géneros, dispersos por los mares tropicales y templados. Solamente
en el Mediterráneo se encuentran once géneros de la familia, representados por: Dentex,
Sparus, Diplodus, Pagellus, Pagrus, Lithognatus, Spondyliosoma, Oblada, Crenidens
Boops y Sarpa.
La clasificación taxonómica de la dorada es:
Reino: Animalia (195.185 especies)
Filo: Chordata (22.199 especies)
Clase: Actinopterygii (16.719 especies)
Orden: Perciformes (8.164 especies)
Familia: Sparidae (124 especies)
Género: Sparus (2 especies)
Especie: Aurata
Es un pez comprimido lateralmente, con el perfil cefálico convexo y bien armado
de dientes. La cabeza es relativamente grande y compacta con el perfil del rostro curvado
convexamente. El hocico y el preopérculo no presentan escamas, y las mejillas son
escamosas. Tiene una coloración gris plateado, más oscuro en el dorso y bastante clara
en la zona ventral. La aleta dorsal es de color gris azulada y presenta una línea media
negra. La caudal es grisácea con los bordes negros. Presenta líneas longitudinales oscuras
a menudo presentes sobre los costados del cuerpo. Además, tiene dos características que
ayudan a su identificación sin lugar a dudas. La primera es una mancha negra en el origen
de la línea lateral, extendiéndose sobre el margen superior del opérculo donde está
bordeada por un área rojiza. Y la otra es la que determina su denominación comercial,
que es la banda frontal dorada entre los ojos bordeada por dos áreas oscuras. (Figura:
1.02)

6
Figura: 1.02 Características morfológicas de un ejemplar adulto de Sparus aurata
Las características relacionadas a su hábitat alimentario también son muy
particulares de la especie. Tiene la boca grande, con labios bastante gruesos y carnosos.
La mandíbula superior es ligeramente más larga que la inferior dando la forma de una
boca baja y levemente oblicua que, sin embargo, no se extiende más allá de la mitad del
ojo. Ambas mandíbulas muestran 4 a 6 dientes tipo caninos localizados en la parte
anterior, y a continuación 2 a 4 series de molares pequeños pero muy potentes, que son
capaces de triturar los caparazones de los moluscos de los que se alimentan
frecuentemente. Pueden aprovechar una amplia gama de alimentos naturales, pero como
en la mayoría de los esparídos, son preferentemente carnívoros y depredadores de
especies zoobentónicas (desde bivalvos y gasterópodos hasta algas macrofitas o
anélidos), crustáceos, cefalópodos e incluso pequeños peces. (Figura: 1.03)
Figura: 1.03 Los dientes y mandíbulas de un ejemplar adulto de Sparus
aurata (Moretti, et al., 1999).

7
La dorada es un pez hermafrodita protándrico, es decir, que a lo largo de su vida
actúa primero como macho y luego como hembra, aunque hay registros de individuos que
permanecieron machos durante toda su vida. En el primer año de vida suelen ser
inmaduros, aunque algunos pueden llegar a madurar como machos, pero en la mayoría de
los individuos, la primera maduración sexual suele ocurrir durante el segundo año de vida.
Tiene bastante sensibilidad a los cambios de temperatura, dejando de alimentarse si
la temperatura baja de 12 – 13°C, siendo letal en el orden de 5 – 7°C. Por el contrario,
tiene el crecimiento muy rápido a temperaturas de 25 – 26°C y soporta valores extremos
hasta 32 – 33°C. Son animales eurihalinos y son capaces de vivir en un amplio rango de
concentración de sales sin que afecte a su metabolismo (3 - 70‰), dándole la posibilidad
de penetrar en zonas de estuarios y lagunas costeras con una amplia variación de
salinidad. Debido a esta característica, se encuentran en diversas zonas del ambiente
acuático y su presencia es muy común en todo Mediterráneo, distribuyéndose por las
costas del atlántico oriental, desde Guinea y Senegal (incluyendo también las islas
Canarias) hasta el Cantábrico y el sur de las Islas Británicas. (Figura 1:04)
Figura: 1.04 Distribución de Sparus aurata de acuerdo a la probabilidad de su
ocurrencia (Fish Base, 2018)

8
1.2.2. EL CULTIVO INTENSIVO DE LA DORADA
Su cultivo en la región mediterránea está experimentado una transformación desde
ser una industria con alto margen de beneficio y baja producción a una de bajos márgenes
y alta productividad. El rápido desarrollo de la producción en jaulas marinas ha llevado a
una disminución de los precios, cultivándose una gran cantidad de peces para reducir los
costos por unidad de producción. Generalmente, cada piscifactoría tiene su propia unidad
de reproducción, donde mantienen los reproductores de varios grupos de edad, los machos
más jóvenes y las hembras mayores.
Los reproductores pueden ser manipulados ambientalmente para extender o
modificar el tiempo de reproducción. Los peces se mantienen en tanques equipados con
un sistema computarizados que controla sistemáticamente la temperatura del agua y la
intensidad de luz. La maduración sexual se obtiene exponiendo a los reproductores a
condiciones de fotoperiodo y temperatura del agua que ocurren durante el periodo natural
de desove. El desove de las hembras se puede obtener por inoculación (5 – 20 mg/kg) de
GnRHA (D-Ala6; Pro9Net-mGnRH) (Moretti, et al., 1999).
En el momento de la madurez sexual se desarrolla la parte ventral de la gónada,
originando testículos funcionales. El control de la proporción de sexos en los tanques de
desove es un factor muy importante y se necesita tomar precauciones porque el cambio
de sexo está determinado por factores sociales. La presencia de machos jóvenes al final
del periodo de desove, por ejemplo, aumenta el número de peces más viejos que se
convierten en hembras. Por otro lado, la presencia de hembras muy viejas reduce el
cambio de sexo en peces más jóvenes. No obstante, esta proporción dependerá también
de otros factores externos que no están todavía aclarados. El tema del cambio del sexo es
muy importante, y merece ser abordado en un estudio detallado.
Hay dos sistemas principales de cría de larvas de dorada llamados de pequeña
escala y de gran escala. El sistema de crianza de pequeña escala (< 10m3) se caracteriza
por un máximo control de los parámetros ambientales y está concebido para producir un
gran número de juveniles (150 – 250/litro). La técnica de gran escala (~200m3) simula un
ecosistema natural. Esta técnica garantiza mucha mejor calidad de las larvas que el

9
sistema de pequeña escala, pero produce mucho menos juveniles con una capacidad
máxima de 10/litro (Moretti, et al., 1999).
La dorada puede ser cultivada de varias maneras, en estanques y lagunas costeras,
con métodos extensivos y semi-intensivo, o en instalaciones con base en tierra y en jaulas
marinas de alta densidad, que son los sistemas de cultivo intensivo. Sin embargo, en los
sistemas intensivos normalmente sigue a otras fases de reproducción, cría de larvas y pre-
engorde. Las fases intensivas de pre-engorde y engorde de dorada pueden realizarse en
instalaciones con base en tierra, con tanques rectangulares de cemento que varían en
tamaño (200 – 3.000m3) de acuerdo con el tamaño de los peces y las demandas de
producción. El engorde también puede hacerse en jaulas marinas, tanto en sitios
protegidos y semi-expuestos (jaulas flotantes) como en sitios totalmente expuestos (jaulas
semi-sumergibles o sumergibles).
El engorde en jaulas marinas es simple y económico; es el sistema de engorde
normalmente usado en la cuenca mediterránea. Aunque las densidades (10 – 15kg/m3)
son más bajas que en los tanques, hay grandes ventajas que hacen al cultivo en jaulas más
rentable. Por ejemplo, no hay costes de energía para bombeo, aireación o tratamiento de
agua post-crianza. Sin embargo, en la crianza en jaulas no es posible el control de la
temperatura, lo que resulta en un período de crianza más largo para alcanzar el tamaño de
mercado, o en la necesidad de sembrar juveniles más grandes. En promedio, las doradas
más grandes y pre-engordadas (10g) alcanzan el primer tamaño comercial (350 – 400g)
alrededor del año, mientras que juveniles más pequeños (5g) alcanzan el mismo tamaño
en alrededor 16 meses (Cataudella y Bronzi, 2001). (Figura: 1.05)

10
Figura: 1.05 Esquema representativo del ciclo de vida de la dorada, S. aurata, en
sistema de cultivo intensivo (FAO, 2005).
1.3. LA MEJORA GENÉTICA EN LA ACUICULTURA
Existe la creencia generalizada de que con la mejora genética de los organismos
acuáticos se pueden alcanzar resultados mucho mejores de los que se han obtenido en los
programas de mejora genética de organismos terrestres. Tal especulación se debe a la
existencia de algunas características reproductivas favorables de estos animales, entre
ellas podemos citar, la mayor variabilidad genética en comparación con vertebrados
homeotermos, la alta fecundidad en peces e invertebrados que permite una selección más
intensa, determinación sexual más plástica, así como la fecundación externa y escasa
presencia de mecanismo de aislamiento reproductivo de estos organismos, permiten
cruces inter específicos que a menudo presentan progenies viables.

11
Sin embargo, la aplicación de los principios de la genética cuantitativa en la
acuicultura es un fenómeno relativamente reciente y el sector aún no ha aprovechado
plenamente las tecnologías disponibles para aumentar la producción, principalmente
cuando se compara con el éxito obtenido en otros sectores productores de alimentos, sean
de origen animal y/o vegetal. Solamente en las últimas décadas se ha empezado a
reconocer la importancia que tiene el mejoramiento genético en el desarrollo de la
acuicultura y que es posible lograr ganancias genéticas muy significativas mediante la
aplicación de programas bien planificados, impulsados por los avances en la biología
molecular. Pero incluso con los modernos métodos moleculares aplicados a la mejora
animal y vegetal, la genética cuantitativa continúa desempeñando un papel fundamental
en el desarrollo de estos programas.
La genética cuantitativa, o genética biométrica, estudia los organismos por medio
de sus características métricas a nivel poblacional, una vez que la segregación de los genes
involucrados no puede ser seguida individualmente, como es el caso de la genética
cualitativa. Por lo tanto, cualquier atributo que varíe continuamente, y que puede ser
medido, podría ser estudiado en principio, como una característica métrica. Cuando ese
proceso tiene como objetivo seleccionar características deseables de interés, esta acción
es denominada de selección artificial, que será el próximo tema abordado en este trabajo.
1.3.1. LOS CARACTERES CUANTITATIVOS
La mayoría de los rasgos de interés económico de las plantas cultivadas y los
animales domésticos, entre los que desde luego se incluyen peces, crustáceos, moluscos
y otras especies implicadas en acuicultura, son caracteres cuantitativos que muestran una
variación hereditaria poblacional debida a la segregación de varios loci, cuya expresión
puede ser modificada por acción del medio. Esta influencia ambiental implica que la
correspondencia entre fenotipo y genotipo no sea una relación perfecta, de manera que
dos individuos con genotipos idénticos pueden tener fenotipos distintos, y, por otra parte,
que la igualdad fenotípica no lleva forzosamente consigo la genética.
La teoría estadística que estudia la variación a continua fue desarrollada por Ronald
Fisher, Sewall Wright y John Haldane, y se basa en utilizar modelos estadísticos que

12
permitan predecir el comportamiento genético de poblaciones, para las características que
son determinadas por muchos genes (poligénicas). Conforme a lo propuesto por Fisher
(1918), la varianza genética tiene tres componentes principales: la varianza genética
aditiva, la desviación dominante y la desviación epistática o de las interacciones entre
genes. Siempre que un heterocigoto se asemeje más a uno de los homocigotos que al otro,
surgirá una varianza debida a dominancia. Por eso que, el fenotipo en los caracteres
cuantitativos, no sólo depende directamente de los alelos que lleva el individuo, sino que
también se ve afectado por las interacciones de un alelo sobre otro (dominancia) del
mismo locus y por el efecto de un gen sobre la expresión de otro (epistasia).
Los estudios genéticos de una característica métrica se realizan a nivel poblacional.
Las estadísticas usadas para describir la variación biológica incluyen, las medidas de
tendencias central (promedio), las medidas de dispersión (varianza, amplitud, desvío
estándar) y las medidas de relación (correlación, covarianza), que describen cómo/cuánto
una variable puede ser modificar por la influencia de la otra variable. El estudio de la
variación fenotípica y la descomposición en sus componentes permiten determinar las
propiedades genéticas de la población. En esta tarea puede ser muy útil el estudio del
grado de semejanza entre los parientes.
Sin embargo, además de los factores de naturaleza genética, los caracteres
cuantitativos también están fuertemente influenciados por los factores externos, con lo
cual presentan distintas respuestas cuando se mantienen en condiciones ambientales
diferentes. Así que, la expresión del valor fenotípico (P) es la suma de los efectos
genéticos (G), ambientales (E) y la interacción de ambos tipos de factores (GxE). En este
sentido, la variación fenotípica puede ser representada por:
P = G + E + GxE
Esta fórmula es la base de la genética cuantitativa y su utilización es fundamental
para implementar a los programas de selección. A continuación, vamos describir los
conceptos básicos de cómo actúa la selección artificial.

13
1.3.2. LA SELECCIÓN ARTIFICIAL
En una población en equilibrio Hardy-Weinberg, la frecuencia relativa de los alelos
y las proporciones genotípicas se mantienen estables a lo largo del tiempo. Desde el punto
de vista del mejoramiento genético, la población no está ni progresando ni retrocediendo
genéticamente, y, para modificar esta estabilidad es necesario el empleo de fuerzas
capaces de alterar las frecuencias génicas. Una de estas fuerzas es la selección artificial,
que es un proceso que induce a una reproducción orientada que tiene el objetivo de
concentrar el patrimonio genético deseable en la población.
La selección es la técnica más utilizada en el mejoramiento de los caracteres
cuantitativos, y su aplicación permite modificar la composición del acervo genético de
una población, de manera que los valores promedios de éstos vayan cambiando en el
sentido deseado a medida que transcurren las generaciones. La mejora genética por medio
de la selección artificial consiste en seleccionar en una población los mejores individuos
que irán a generar la nueva población, o sea, la población seleccionada o población
mejorada. Admitiendo que las características siguen un patrón de distribución normal, la
selección de fenotipos superiores puede ser representada gráficamente como la Figura:
1:06.
Figura: 1.06 La selección en la curva de distribución normal, donde, (XP) es el
promedio de la población original, (XS) el promedio de población
seleccionada y (DS) es denominado como el diferencial de selección

14
Considerando que la selección es realizada a partir de la evaluación fenotípica, el
diferencial de selección (DS) dependerá, a su vez, de la intensidad de selección (i) que
dependerá de la proporción de individuos seleccionados (b).
La respuesta a la selección es medida por la formula denominada como “la ecuación
fundamental del mejoramiento genético por medio de la selección”, que es la más
importante y sencilla aplicación de la genética cuantitativa a la mejora genética animal
(Lopes, 2005). Es la ecuación que va predecir la eficiencia de la selección, o sea, predice
cuánto los descendientes serán genéticamente superiores en comparación al promedio de
la generación de sus padres. Pero, en los programas de mejora genética, el objetivo es
siempre maximizar la ganancia genética por unidad de tiempo. Así que, para predecir la
ganancia genética, se debe llevar en consideración también el tiempo necesario, o sea, el
intervalo de cada generación. Por lo tanto, la ganancia genética esperada por unidad de
tiempo puede ser dado por:
∆G = i·h2·σP / t
Donde, ∆G= ganancia genética; i= intensidad de selección; h2= heredabilidad; σP=
variación fenotípica; t= intervalo generacional.
Cuando examinamos la ecuación, vemos que la respuesta a la selección podrá ser
mejorada cuando aumentamos la intensidad de selección (menor proporción de
individuos) o cuando disminuimos el intervalo de generacional. Sin embargo, existe un
importante conflicto de intereses, puesto que cuando se aumenta la intensidad de
selección, consecuentemente aumenta también la consanguinidad, siendo así, necesario
encontrar un equilibrio para mantener un progreso genético rentable a lo largo de las
generaciones. Durante los últimos años se han propuesto muchas estrategias para
controlar la consanguinidad en programas de selección (Toro y Pérez-Enciso, 1990;
Meuwissen, 1997; Fernández y Toro, 1999; Villanueva, et al., 2004). El objetivo es
reducir la tasa de consanguinidad manteniendo el mismo (o superior) nivel de respuesta
o bien incrementar la respuesta, pero manteniendo un nivel prefijado de la
consanguinidad. Las estrategias propuestas se basan en: a) modificar el número de
individuos seleccionados y su contribución a la siguiente generación; b) modificar el
método de estimación del valor mejorante; c) implementar un adecuado sistema de
apareamiento (Fernández, et al., 2004).

15
Basándose en tales estrategias, el principal objetivo del trabajo es lograr la
implementación de un programa de mejora genética en dorada, mediante estrategias que
utilizan los marcadores moleculares y las informaciones del valor genético para predecir
qué individuos deben ser seleccionados para la formación de la siguiente generación, para
aumentar la probabilidad de generar descendientes superiores manteniendo controlado el
incremento de la consanguinidad. Todo ello en condiciones productivas de una empresa
de dorada.

16
2.
OBJETIVO
GENERAL

17
2. OBJETIVO GENERAL
El objetivo principal de este proyecto fue el de desarrollar los procedimientos más
adecuados para llevar a cabo un programa de mejora genética en dorada. Se trata de
mejorar la producción de dorada en la empresa Aquacultura Balear, S.A.U. (ABSA) que
es el criadero comercial del Grupo Culmarex, situado en Palma de Mallorca, España. La
finalidad es desarrollar un programa de selección con la tasa de crecimiento como
principal carácter a mejorar que esté basado en el uso de marcadores moleculares para la
determinación de las relaciones genealógicas y el control de la consanguinidad. Se tendrá
en cuenta la necesidad de reducir los costes implicados para que la estrategia sea viable a
nivel comercial en diferentes condiciones de cultivo.
2.1. OBJETIVOS ESPECÍFICOS
Evaluar la diversidad genética, el parentesco y las contribuciones relativas de
todos los reproductores del stock inicial.
Evaluación mediante la relación coste/beneficios de las diferentes alternativas
para el diseño del plan de selección.
Estimar el parentesco genealógico usando las informaciones moleculares.
Estimar los parámetros genéticos (heredabilidad y correlaciones).
Estimación de los valores mejorantes de los individuos mediante metodología
BLUP.
Optimizar la selección por medio de una metodología que permitan mantener el
incremento de la consanguinidad a niveles aceptables.

18
3.
MATERIAL
&
MÉTODOS
19
3. MATERIAL Y MÉTODOS
3.1. LA EMPRESA
El Grupo Culmarex (www.culmarex.com) es una empresa que fue constituida en
1986 por un grupo de inversores de Águilas, la cual tras diferentes etapas fue comprada
en su totalidad en junio de 2011 por Cook Aquacultura Inc., compañía canadiense
considerado una de las mayores industrias salmoneras en el mundo. Es un grupo de
granjas marinas y es el productor más importante de dorada y lubina en España, y una de
las mayores piscifactorías en mar abierto.
Actualmente, el grupo está compuesto por 11 empresas: ABSA, Basademar,
Bersolaz, Blue & Green, Culmaréx, Gramabasa, Lebeche Carboneras, Lebeche Gorguel,
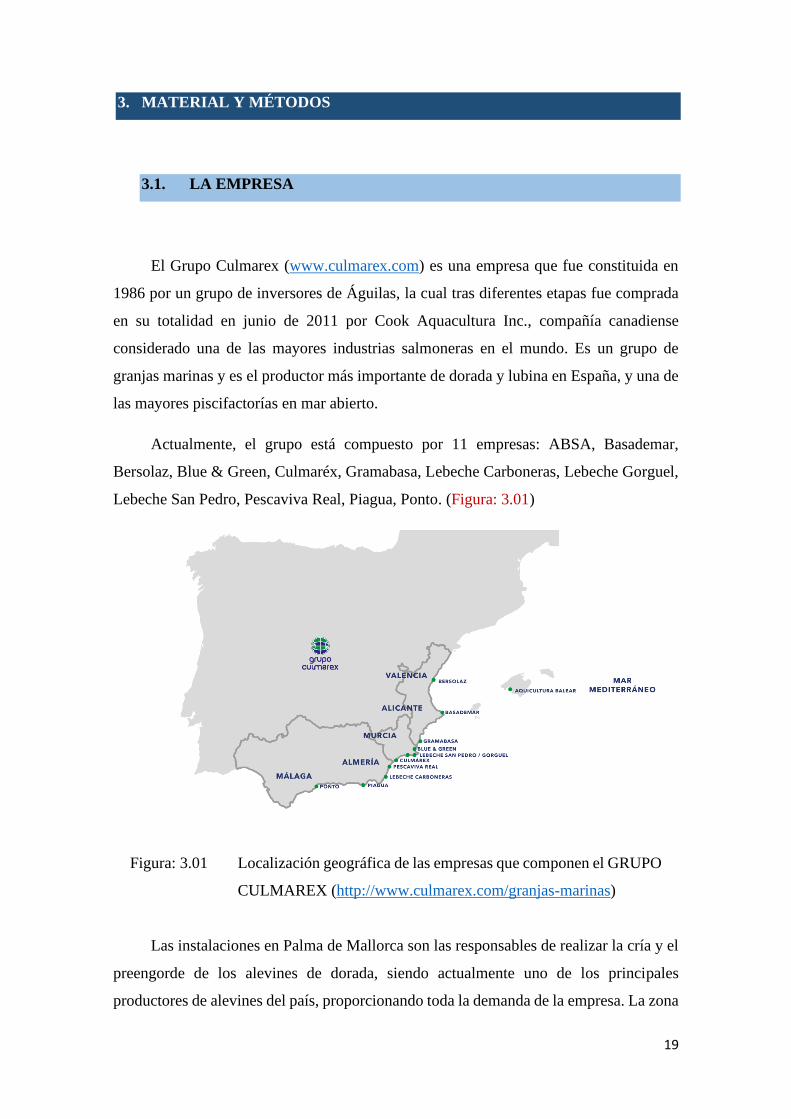
Lebeche San Pedro, Pescaviva Real, Piagua, Ponto. (Figura: 3.01)
Figura: 3.01 Localización geográfica de las empresas que componen el GRUPO
CULMAREX (http://www.culmarex.com/granjas-marinas)
Las instalaciones en Palma de Mallorca son las responsables de realizar la cría y el
preengorde de los alevines de dorada, siendo actualmente uno de los principales
productores de alevines del país, proporcionando toda la demanda de la empresa. La zona

20
se destaca por la contrastada calidad de su agua produciendo individuos de calidad
superior y sanidad garantizada. Además, tiene una ubicación muy estratégica en el
Mediterráneo que facilita el transporte, y aseguran una entrega rápida y eficaz de los
alevines en las jaulas de engorde, por medio del barco “wellboat” (Figura: 3.02).
Figura: 3.02 Una ilustración gráfica del barco Wellboat, utilizados para transporte y la
captura de individuos (Fuente: www.wellboat.cl)
Los datos presentados en este trabajo han sido realizados por medio de un contrato
predoctoral asociado al proyecto de investigación titulado “Programas de selección
genética para la mejora de la producción en dorada y lubina en la planta de ABSA, S.A.U.
(CDTI) (ref: 2011/CE035)”, financiado a través del Departamento de Genética de la
Facultad de Veterinaria de la Universidad de Santiago de Compostela. El estudio
consistió en analizar ciclos reproductivos de las poblaciones reproductoras de dorada,
Sparus aurata en el criadero ABSA, que tienen ya implantado un programa de
mejoramiento genético desde finales del año de 2007.
Los experimentos fueron realizados dentro de las instalaciones de la empresa
acuícola Acuicultura Balear ABSA, que está situado en Islas Baleares, Mallorca, España.
Es la compañía integrada en el grupo Culmarex responsable de la crianza de juveniles

21
para las otras empresas, centrando su actividad en la explotación del criadero y el pre-
engorde de alevines de dorada y lubina. (Figura: 3.03)
Figura: 3.03 Vista aérea de las instalaciones de Acuicultura Balear SAU, en Palma
de Mallorca, Islas Baleares (fuente: Google Earth)
3.2. LOS ANIMALES
3.2.1. LA POBLACIÓN FUNDADORA
La población inicial estaba formada por 643 individuos. Los animales fueron
clasificados en 6 grupos de edad (A – F), correspondiendo a un intervalo de nacimiento
entre 2002 hasta 2007, de acuerdo con la Tabla 3.01.

22
Tabla: 3.01 Número de reproductores que componen cada fotoperiodo. En la horizontal
están los grupos de animales ordenados de acuerdo con su fecha de nacimiento.
En la vertical están los fotoperiodos y las fechas en que han sido formados.
N.º Ind (edad) A(Mar/02) B(Dec/02) C(Abr/05) D(Oct/05) E(May/06) F(Mar/07) Total
Inicial 153 161 151 80 42 56 643
CD4 (Dec/07) 32 28 13 - - - 73
CD5 (Abr/08) 30 23 38 - - - 91
CD1 (May/09) 22 32 18 - - 29 101
CD2 (Ago/09) 28 29 23 - - 26 106
CD3 (Dec/09) 7 9 44 22 42 - 124
Total 119 121 136 22 42 55 495
Descartados 34 40 15 58 0 1 148
Todos los individuos analizados han permanecidos en condiciones ambientales
similares entre los tanques (ej. periodo de luz, temperatura del agua, proporción sexual,
densidad poblacional, dieta, salinidad, etc.), manteniéndolas constantes durante todo el
periodo.
3.2.2. LOS REPRODUCTORES
Los reproductores de la población fundadora fueron distribuidos y almacenados en
sus fotoperiodos de acuerdo con la Tabla 3.01. El ciclo anual de reproducción estaba
formado por cinco ciclos de fotoperiodos, cada uno constituido por dos tanques de 10m3.
Los tanques eran mantenidos en una estructura cubierta, cerrada y aislado de
contaminaciones externas. En estas condiciones, los reproductores eran mantenidos bajo
un riguroso control sobre variaciones de las condiciones ambientales, principalmente la

23
temperatura y fotoperiodo, parámetros relevantes para realizar la inducción de las puestas
masales. (Figura: 3.04).
Figura: 3.04 Los tanques de reproductores de uno de los fotoperiodos. Natación en
círculos y agrupados, movimientos característicos de desove masal.
En cada tanque de reproducción, los animales fueron distribuidos en una proporción
sexual de aproximadamente 20 machos (peso medio = 2,5 Kg) y 30 hembras (peso
medio= 2,7 Kg), con el objetivo de mantener la densidad media dentro del rango de
aproximadamente 11 a 14 Kg/m3.
Fueron utilizados un total de 495 animales para formación de la población F0. El
número de reproductores y su distribución por tanque y fotoperiodo se describen en la
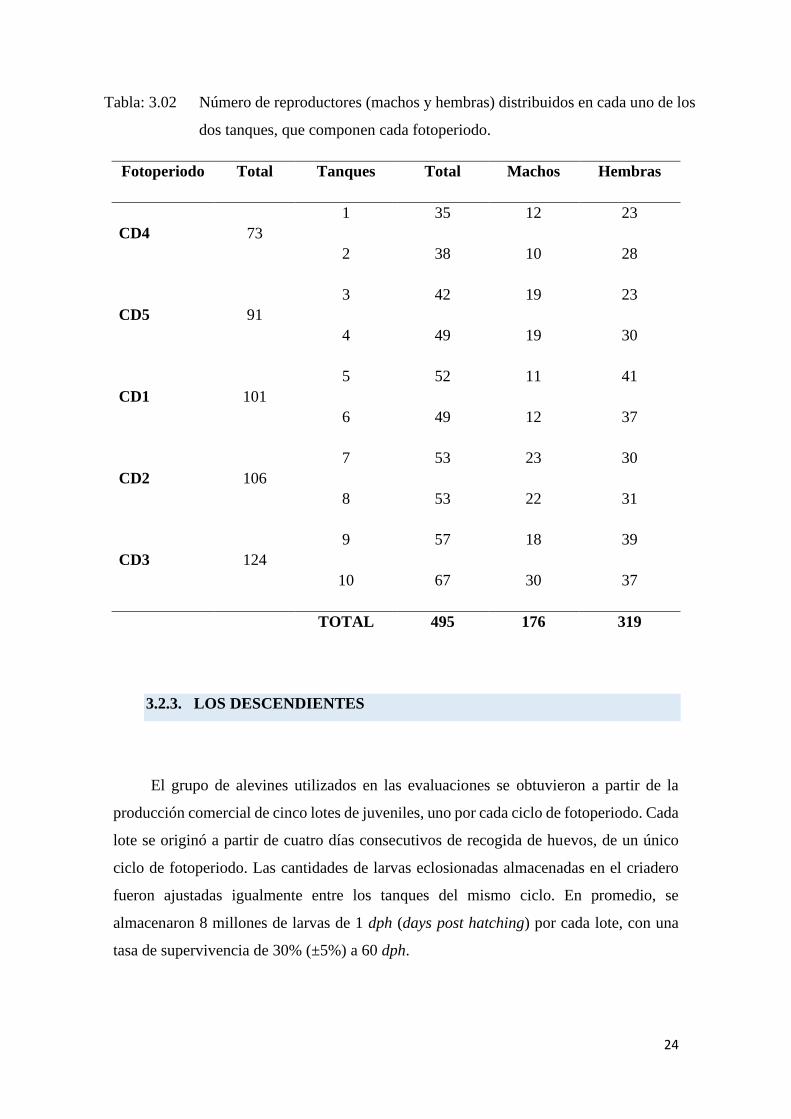
Tabla: 3.02.
24
Tabla: 3.02 Número de reproductores (machos y hembras) distribuidos en cada uno de los
dos tanques, que componen cada fotoperiodo.
Fotoperiodo Total Tanques Total Machos Hembras
CD4 73
1 35 12 23
2 38 10 28
CD5 91
3 42 19 23
4 49 19 30
CD1 101
5 52 11 41
6 49 12 37
CD2 106
7 53 23 30
8 53 22 31
CD3 124
9 57 18 39
10 67 30 37
TOTAL 495 176 319
3.2.3. LOS DESCENDIENTES
El grupo de alevines utilizados en las evaluaciones se obtuvieron a partir de la
producción comercial de cinco lotes de juveniles, uno por cada ciclo de fotoperiodo. Cada
lote se originó a partir de cuatro días consecutivos de recogida de huevos, de un único
ciclo de fotoperiodo. Las cantidades de larvas eclosionadas almacenadas en el criadero
fueron ajustadas igualmente entre los tanques del mismo ciclo. En promedio, se
almacenaron 8 millones de larvas de 1 dph (days post hatching) por cada lote, con una
tasa de supervivencia de 30% (±5%) a 60 dph.

25
Cuando alcanzaban aproximadamente los 130 dph, los individuos eran evaluados
mediante un proceso de selección manual, en el que se analizaban las características
externas del animal, descartando aquellos que presentaban características indeseadas o
una anomalía aparente. (Figura 3.05)
Figura: 3.05 La mesa de descartes morfológicos
De los animales no descartados, eran entonces seleccionados aleatoriamente 1.500
alevines y transferidos a tanques de 20m³, donde serían criados conjuntamente hasta
llegar a la edad de aproximadamente los 500dph. (Figura: 3.06)

26
Figura: 3.06 Tanques circulares de juveniles
Durante la fase de crecimiento, los juveniles fueron alimentados diariamente hasta
la saciedad con una dieta comercial peletizada (utilizándose tanto alimentadores
automáticos como alimentación manual). La temperatura del agua se mantuvo constante
durante todo el experimento 20°C (± 1°C).
3.2.4. EL MANEJO Y LA TOMA DE MEDIDAS
Para el marcaje de los peces y la toma de medidas, el procedimiento se inicia con
la preparación de una solución de anestésico de eugenol con concentración
aproximadamente 30mg/L, diluida en alcohol (70%). Los animales son retirados de los
tanques y son introducidos en un cubo conteniendo la solución. Se quedan allí hasta que
se les note los efectos anestésicos, caracterizados por la natación desorientada con el
vientre hacia arriba (Figura: 3.07). Una vez anestesiados, los individuos son retirados del
cubo y se realizan los procedimientos necesarios para la marcación con microchip,

27
biometrías y colecta del material biológico. Terminado el manejo, los animales vuelven
a un cubo, adaptado con una tela, que tiene en un sistema de renovación de agua continua
y permanecen allí hasta que se noten haber recuperado su movilidad nuevamente.
Figura: 3.07 A: La solución de Eugenol (anestésico)
B: Inmersión de los animales en la solución
Todos los individuos incluidos en este trabajo (tanto los reproductores como los
juveniles) fueron etiquetados físicamente mediante la inyección de un transponder interno
(pit-tag) colocado debajo de la aleta dorsal posibilitando la identificación individual de
cada animal. La marcación se realizaba cuando los individuos alcanzaban
aproximadamente los 100g, y durante ese proceso, se realizaron dos medidas,
aproximadamente a los 350 y 500 dph, para colectar las informaciones fenotípicas
necesarias para el proceso de estimación y predicción genética.
Se colectaron datos biométricos del peso (g) y de la longitud estándar (cm), que es
la medida comprendida entre el inicio de la cavidad bucal hasta la última vértebra de la
aleta caudal. Las medidas del peso fueron hechas con ayuda de una balanza comercial, y
la longitud con una cinta métrica con ayuda de una mesa luminosa, que facilitaba la
precisión de la lectura (Figura: 3.08).

28
Figura: 3.08 La bancada de las mediciones biométricas
3.3. LAS CONDICIONES DEL CRIADERO (HATCHERY)
La reproducción de los peces en cautividad suele seguir las siguientes etapas, la
preparación e inducción de los peces reproductores (maduración sexual), el desove y
recogida de los huevos, la incubación en condiciones controladas y la cría inicial de las
larvas. En 2011, la fecundidad media de los reproductores en ABSA alcanzó la cifra de
9,9 Kg de huevos por hembra en cada tanque (es decir: 3,7 kg de huevos/kg de hembras).
Dentro de cada fotoperiodo, las tasas de fecundación estaban por encima del 90%
mientras que el promedio de las tasas de eclosión de los lotes comerciales era del 86%.
La maduración sexual se obtuvo exponiendo a los reproductores a condiciones de
fotoperiodo y temperatura del agua que mimetizan el periodo natural del desove. Por ello,
los tanques del criadero se mantuvieron bajo dos regímenes de luz y temperatura del agua,
que fueron controlados cuidadosamente durante todo el periodo. El desove natural fue
desencadenado por la reducción de la temperatura del agua hasta 17°C, con un
fotoperiodo 09:45 HL (hours of light) / 14:15 HD (hours of darkness). Bajo estas

29
condiciones, el periodo de desove duró cerca de cuatro meses y medio, con una media de
2,5 kg de huevos viables (cerca de 3 millones) recogidos, diariamente, en el pico del
desove (entre 11:30 y 13:30 HL). Los fotoperiodos son organizados de forma
sincronizada, para que las puestas sean continuas durante todo el año.
Los huevos ya fecundados son retirados de la colectora de huevos (Figura: 3.09) y
llevados a las instalaciones de cultivo larvario (hatchery), donde permanecen hasta el
agotamiento del saco vitelino y empezar a recibir alimentos vivos (microalgas, artemia y
rotíferos) hasta alcanzar aproximadamente 1 gramo. En promedio, durante este periodo
se almacenaron aproximadamente 2 millones de larvas por día, con una tasa de
supervivencia próximo al 30% (± 5%) a los 60 dph (days-post-hatching).
Figura: 3.09 Caja colectora de huevos
Los reproductores fueron alimentados manualmente, todos los días, hasta la
saciedad. Las tasas de alimentación diaria variaban según el patrón estacional que
dependía principalmente de la etapa de maduración sexual de los reproductores, que iba

30
de 0,6% la biomasa del tanque (fuera de la temporada de desove) hasta el 0,2% (durante
la temporada de desove). Los peces se alimentaron con una dieta comercial peletizada,
siendo la dieta complementada, en el periodo de desove, con la aportación de calamares
congelados, tres veces por semana.
3.4. LA EXTRACCIÓN DEL ADN Y LOS ANÁLISIS DE LOS
MICROSATÉLITES
Los análisis de ADN han sido realizados todos por el grupo de investigación
ACUIGEN (http://www.acuigen.es/en) de la Facultad de Veterinaria de la USC del
campus de Lugo, que estaban encargados de los procedimientos de biología molecular
del proyecto.
El ADN genómico se obtuvo de un pequeño trozo de la aleta caudal de todos los
animales utilizados en este estudio. La extracción del ADN se realizó utilizando el kit
TissueDNAKit EZ96® (OmegaBio -tek).
Se genotiparon un total de ocho microsatélites distribuidos en dos multiplex de
cuatro loci cada uno. El primer multiplex estaba constituido por los loci Sal15, Sal19,
SaGT32 y SaGT41a, mientras que el segundo lo formaban PbMS16, PbMS2, Sal14 y
SauG46INRA (Batargias, et al., 1999; Stockley, et al., 2000; Launey, et al., 2003; Brown,
et al., 2005a,b).
Estos loci se seleccionaros a partir de la herramienta de microsatélites desarrollada
anteriormente por Castro, et al., 2007 (SAGT41a, PbMS16 y PbMS2), y de la
información disponible sobre los mapas de ligamiento y diversidad genética del mapa
híbrido de radiación de dorada y nuevos marcadores (Franch, et al., 2006; Sarropoulou,
et al., 2007; Sal15, Sal19, SaGT32, Sal14 y SauG46INRA), sustituyendo algunos loci
estrechamente ligados por otros independientes y altamente polimórficos. (Tabla: 3.03)

31
Tabla: 3.03 Lista de los loci microsatélites utilizados para la asignación de la
paternidad.
Microsatélites Secuencia de los Cebadores
Sal15 F: ACACTGTCTTTCTGTCCCTCACAC
R: GAGTAACACAGCCTCAGTTGAAGC
Sal19 F: ATTCTTCACAGGCCCAACACAAA
R: GAAAACACCGGCCCAGTACGA
SaGT32 F: GAGCAGACACCAGTGCATG
R: GATAATGGCCAAAAGTCACTG
SaGT41a F: TCAAAGACAGATGGAGCTGG
R: GTCACATCAGTCTGCACTTG
PbMS16 F: AAACAGTCGGGACACAAAGC
R: GGACAGGGGCCTAGTTTATC
PbMS2 F: CTCGCGGTAGTTGTTCACAGAGC
R: GCAACAATGGAGGGTTCGTCATC
Sal14 F: TGCCAGAATGAATACCAACTGGTG
R: ATAATCAAAGTACCCCTGCATGTC
SauG46INRA F: GTGAACACCTGCCAGACG
R: GCATCGAGGTCAAGTACCTG
Los genotipos se obtuvieron por electroforesis capilar en un secuenciar automático
ABI3730x1 DNA Analyzer (Applied Biosystems), y la caracterización de las series
alélicas se realizó utilizando el programa GeneMapper v.4.0 (Applied Biosystems).
Ambas reacciones multiplex se realizaron en 10 μl de volumen total incluyendo 5
μl de Kit GoTaq® Hot Start Colorless Master Master Mix (www.promega.com), 1 μl de
mezcla de cebadores, 4 μl de agua MQ y 1 μ del ADN. La reacción de PCR incluía una
primera etapa de desnaturalización a 95°C durante 2 minutos, y 28 ciclos de
desnaturalización del ADN a 94°C durante 30 segundos, renaturalización durante 1
minuto a 57ªC, y extensión durante 1 minuto a 72ªC, un paso de extensión final se realizó
a 72°C durante 5 minutos.
El segundo multiplex se utilizó solo en los casos en que los resultados obtenidos
con el primero eran relativamente insuficientes, es decir, con proporción baja de progenie
asignada. Por lo tanto, todos los individuos no asignados en la primera ronda se analizaron
adicionalmente utilizando el segundo multiplex.

32
3.5. LA ASIGNACIÓN DEL PARENTESCO
Normalmente, cuando se eligen microsatélites para la asignación familiar, lo más
importante es el número de alelos que tienen y la heterocigosidad (Ho, He). También es
importante elegir microsatélites de diferentes grupos de ligamiento, para tener mayor
representación de todos los cromosomas. Si los microsatélites son del mismo grupo de
ligamiento pueden dar informaciones similares, y estará teniendo información sobre la
misma zona genómica y duplicarse la información sin mayor ventaja de asignación.
Para la asignación del parentesco molecular hemos utilizado el software FAP,
(Familiar Asignation Program), que es un programa de ordenador que se ejecuta en DOS.
Este paquete se refiere a la cuestión de asignar padres potenciales a cada juvenil problema,
por medio de exclusión mendeliana basado en el genotipo (Taggart, 2007). El programa
también ofrece herramientas útiles para identificar loci problemáticos y errores de
genotipado durante la asignación real.
La asignación se realizó para cada grupo de fotoperiodo, considerando todos los
posibles cruces de pares candidatos en cada uno de los dos tanques reproductores. No se
permitió ninguna incompatibilidad para determinar la exclusión, y solamente los
individuos asignados a un solo par de padres se mantuvieron para el análisis y se
convirtieron en los candidatos para la selección. Las probabilidades teóricas de exclusión
se dar por la enumeración completa de todos los genotipos (padres e hijos), y la formación
de la familia son realizados a partir de los posibles emparejamientos dado por las
frecuencias alélicas de cada loci, compartidos por descendencia.
Con base en estas cuentas directas de genotipos, el software utiliza de un algoritmo
de pesquisa comparativa que calcular la probabilidad de que individuos tengan
descendencia compartida a partir de los dados genotípicos de los padres e hijos.
La reconstrucción del pedigrí también permitió determinar el número de
reproductores que realmente aportan descendencia a la siguiente generación y las
contribuciones exactas de cada uno de ellos. A partir de esta información, se estimó el
tamaño efectivo de la población (Ne) como 4N/ (2 + Vk), donde N es el número total de
padres disponibles (machos y hembras) y Vk la varianza del número de descendientes
asignados a cada uno de ellos. El Ne esperado suponiendo contribuciones al azar

33
(distribución Poisson), se calculó como (4NS x ND) / (NS + ND), siendo NS y ND el número
disponible de machos y hembras, respectivamente, en el tanque donde se realizó el
apareamiento.
Para explicar las diferencias en el porcentaje de asignación de paternidad entre los
fotoperiodos, se estimó la relación genética de los reproductores en cada tanque utilizando
el parentesco molecular, mediante el método de similitud alélica (Nejati-Javaremi, et al.,
1997). También se calculó la consanguinidad molecular de cada individuo, es decir, la
homocigosidad observada para los marcadores.
3.6. ANÁLISIS DE LOS DATOS
Las estimaciones de la heredabilidad y correlaciones se obtuvieron a partir de una
muestra de 6.285 individuos originados de los cinco grupos reproductores de
fotoperiodos, usando únicamente los individuos asignados con éxito a un solo par de
padres con el programa FAP. Dado que no había información previa sobre el pedigrí, los
hermanos, medios hermanos y parejas no relacionadas eran las únicas posibilidades para
cualquier par de peces.
Para la estimación se utilizaron tres paquetes de software (VCE, REMLF90 y TM).
Los programas VCE 6.0 (Kovac, et al., 2002) y los REMLf90 (Misztal, et al., 2002)
proporcionan estimaciones REML de los componentes de la varianza. Por otro lado, TM
(Bayesian-based software, Legarra, et al., 2008) utiliza métodos de muestreo MCMC y
Gibbs con a priori uniformes para efectos fijos y componentes de varianza y proporciona
distribuciones posteriores para los parámetros de interés.
Los datos fueron analizados utilizando la metodología de los modelos lineales
mixtos. Los componentes de la varianza de todos los caracteres considerados en este
estudio fueron estimados por máxima verosimilitud restringida (REML) y por inferencia
bayesiana utilizando métodos MCMC. Los valores mejorantes para cada individuo
(EBVs) fueron estimados por el procedimiento BLUP (mejor predictor lineal insesgado).
Los valores fueron estimados utilizándose del siguiente modelo lineal animal:
y =Xβ + Zu+ e

34
En términos matriciales, el modelo puede ser descrito como:
donde y es el vector de valores fenotípicos para el carácter, X es la matriz de
incidencia (de valores 0 y 1) y β el vector de efectos fijos (media, fotoperiodos y tanque),
Z es la matriz de incidencia (de valores 0 y 1) asociadas al vector u, que son los valores
mejorantes, o sea, el efecto aleatorio animal, y e el vector de los residuos. Los valores
mejorantes y los errores residuales se distribuyen con una normal N y desviación estándar
(0, σu2) y N (0, σe
2), respectivamente.
3.7. LA OPTIMIZACIÓN POR TEMPLADO SIMULADO (SIMULATED
ANNEALING)
La selección mediante la metodología BLUP es actualmente la más utilizada en los
programas de mejora genética, sin embargo, dado que las estimaciones informaciones
referentes a todos los parientes, su aplicación suele inducir mayores tasas de
consanguinidad que la selección basada únicamente en la información fenotípica
individual. Para evitar la endogamia, es conveniente hacer la selección artificial con
restricciones al incremento de la consanguinidad, a fin de aumentar la respuesta a
selección a largo plazo. Por estos motivos, hemos utilizado técnicas de optimización, que
posibilitan hacer la elección de los mejores individuos, manteniendo controlado en el
incremento en los valores de endogamia.
El hecho de que la relación gráfica existente entre los valores genéticos estimados
(BLUP) y el coeficiente de endogamia (F) no sea lineal, pone de manifiesto que, es
atractivo, desde un punto de vista práctico, buscar una solución en la que se mantenga
una respuesta razonable en la búsqueda por mejores resultados genéticos, sin generar una

35
consanguinidad excesiva. La aplicación del método de contribuciones óptimas (optimal
contributions – OC; Meuwissen, et al., 1997) se presenta como una importante
herramienta para maximizar la ganancia genética, combinando el equilibrio entre el
mérito genético y la consanguinidad de la población.
La optimización consiste en buscar una solución o un conjunto de soluciones
óptimas para una determinada función o un conjunto de funciones, determinando los
valores que deben tomar las variables para maximizar/minimizar la función objetivo
óptima, satisfaciendo al mismo tiempo las restricciones impuestas. Aunque existan otros
métodos para maximizar la respuesta por medio de la optimización (Meuwissen y
Sonesson, 1998; Grundy, et al., 2000; Chakraborty, et al., 2002; Villanueva, et al., 2002;
Tang y Li, 2006; Li, et al., 2006; Li, et al., 2008; Kinghorn, 2010; Sonesson, et al., 2010;
Nishio, et al., 2010) hemos utilizado de un algoritmo probabilístico denominado de
templado simulado (simulated annealing), que utiliza fundamentos análogos de la
termodinámica para simular un proceso probabilístico semejante al calentamiento de
moléculas y su consecutivo enfriamiento. En este proceso, cuando un sólido es enfriado
bruscamente, sus átomos formaran una estructura irregular y de alta energía. Mientras
que, con enfriamiento lento, los átomos encontraran tiempo suficiente para organizarse
en una estructura regular (cristal) que supone la disposición de energía mínima.
En el caso del simulated annealing el método consiste en una búsqueda “ciega” por
el conjunto de soluciones factibles. En una interacción del proceso de enfriamiento, una
solución actual es modificada ligeramente para producir una nueva alternativa que puede
reemplazarla o no. Si la nueva solución (x´) tiene mejor valor (Z) que la anterior, la
reemplaza (x=x´), si no, se utiliza la fórmula para decidir la probabilidad de aceptarla es:
P(∆Z) = e ∆Z/T
En la práctica, se escoge un número aleatorio (n) uniformemente distribuido en el
intervalo (0,1). Ese número se compara con P(∆Z). Si n < P(∆Z), x’ reemplaza a x como
solución actual. Si n ≥ P(∆Z), x se usa de nuevo como paso inicial de una próxima
iteración. Al principio, T es un valor alto (fundición), y luego va disminuyendo
(enfriamiento), es decir, cada vez es menor la probabilidad de que la nueva alternativa
reemplace a la anterior si es peor. El proceso puede ser descrito por el flujograma a la
Figura: 3.10.

36
Figura: 3.10 Flujograma del templado simulado
El proceso se repite iterativamente hasta que se considere que llegó al óptimo. La
eficacia de este algoritmo (la obtención de una solución óptima) va a depender de los
valores utilizados en una serie de parámetros que dirigen su funcionamiento. Los
parámetros son la temperatura inicial y temperatura final o punto de parada, la tasa de
reducción progresiva de temperatura y por el número de iteraciones para seleccionar el
nuevo estado a cada valor de temperatura. La calibración de estos valores aumenta la
probabilidad de localizar una solución óptima global. Es por eso importante realizar un
estudio previo para determinar los parámetros más adecuados.

37
4.
RESULTADOS

38
4. RESULTADOS
4.1. LA DESCRIPCIÓN ESTADÍSTICA DE LA POBLACIÓN
La Figura: 4.01 muestra el gráfico de distribución de frecuencias (histograma) para
las características fenotípicas medidas para el peso (g) y la longitud (cm), obtenidas tanto
en la primera como en la segunda medición para la población de los cinco grupos de
fotoperiodos estudiados. En general, la primera medición fue realizada cuando los
individuos tenían 346,8 ± 6,1 días de edad, y en este momento pesaban cerca de 159,5 ±
35,3 g, y la longitud del cuerpo estaba cerca de 18,4 ± 1,4 cm. En un intervalo de
aproximadamente 5 meses fue realizada la segunda medición, cuando los individuos
tenían una edad promedio de 497,7 ± 7,4 días, con el peso medio ahora de 397,9 ± 63,9 g
y la longitud en 24,6 ± 1,2 cm.
Figura: 4.01 Histograma del peso(g) y longitud (cm) en las dos edades medidas
Evaluando las poblaciones de los fotoperiodos separadamente, se observa que, en
la primera medida, la desviación estándar (y el coeficiente de variación) para todas las
variables estudiadas fue mayor en los grupos de fotoperiodo CD4 y CD5, ya que los datos
de campo fueron recolectados durante en un intervalo de días mayor, marcando en la
primera medición periodos de hasta 12 días (CD4) y de 19 días (CD5). Mientras que en
los grupos de fotoperiodos CD1, CD2 y CD3, los datos de campo se recogieron en un

39
intervalo de 2 días, con excepción de la segunda medición para el CD1 que se realizó en
un intervalo de 12 días.
El fotoperiodo CD4 fue el que presentó la primera medición a la edad más tardía
(352 días) pero con intervalo de apenas 143 días hasta la segunda medición. Sin embargo,
fue en este fotoperiodo que se registró el peso más grande para la segunda medición
(670,0 g), así como también los mayores valores de la desviación estándar (68,98) y en el
coeficiente de variación (18,03).
El fotoperiodo CD5 fue en el que se realizaron las primeras mediciones a la edad
más temprana (340 días), que puede ser uno de los motivos por lo cual han presentado
valores más bajos para el peso (132,9 g) y la longitud (16,9 cm). Sin embargo, para estas
mismas características fueron las que presentaron los mayores valores del coeficiente de
variación para el peso (21,72) y longitud (6,75) en la primera medición. Mientras que, en
la segunda medición, estos animales han crecido bastante bien, y presentaron uno de los
mejores resultados. Estos resultados van de acuerdo con el hecho de que, en este
fotoperiodo, fueron los que presentaron mayores valores para la ganancia, tanto del peso
(1,79 g/día) cuanto para la longitud (0,05 cm/día), aunque también hay que considerar
que el intervalo de días entre las mediciones también fue uno de los mayores
(aproximadamente 158 días).
Mientras que el fotoperiodo CD1 fue el que presentó resultados más bajos para la
ganancia diaria de peso (1,38 g) y longitud (0,03 cm), ha presentado uno de los mayores
coeficientes de variación para la ganancia del peso (18,97).
El fotoperiodo CD2 ha sido el que tuvo la segunda medición a la edad más tardía
(506 días) y también con el mayor intervalo entre las mediciones (162 días), y
aparentemente fue también la que presentó mejor homogeneidad entre todos los
fotoperiodos, presentando los menores valores tanto para la desviación estándar como
para el coeficiente de variación para casi todas las medidas biométricas tomadas, tanto en
la primera como en la segunda medición.
El fotoperiodo CD3 ha sido aquel que presentó menores variaciones durante las dos
mediciones, así como el menor intervalo entre las medidas (139 días). Sin embargo, fue
el fotoperiodo que presentó los mayores resultados promediados, tanto para el peso como
la longitud en las dos mediciones realizadas.
40
Los coeficientes de variación fueron mayores para el peso que para la longitud para
ambas medidas independientemente de la edad, de 22,11 y 7,83 para la edad 1, y de 16,07
y 5,07 para la edad 2, respectivamente. La media, la desviación estándar y el coeficiente
de variación de las medidas de peso y longitud en las dos edades consideradas, y las
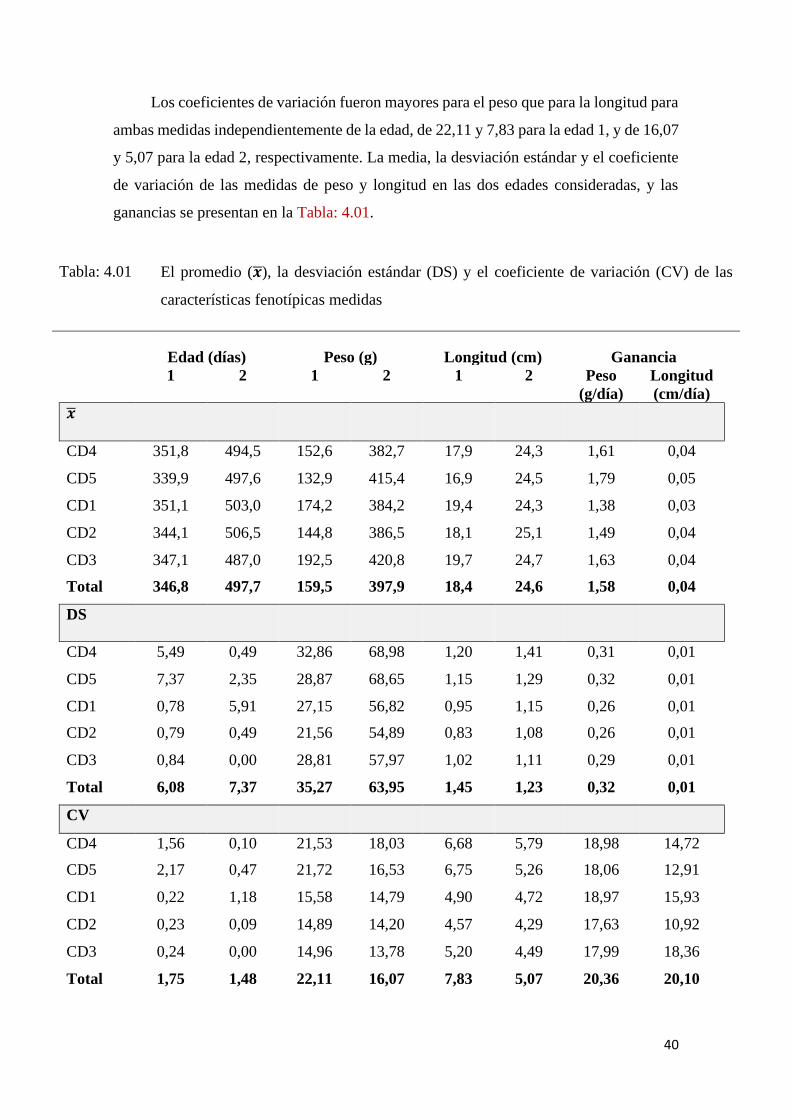
ganancias se presentan en la Tabla: 4.01.
Tabla: 4.01 El promedio (𝒙), la desviación estándar (DS) y el coeficiente de variación (CV) de las
características fenotípicas medidas
Edad (días) Peso (g) Longitud (cm) Ganancia
1 2 1 2 1 2 Peso
(g/día)
Longitud
(cm/día)
𝒙
CD4 351,8 494,5 152,6 382,7 17,9 24,3 1,61 0,04
CD5 339,9 497,6 132,9 415,4 16,9 24,5 1,79 0,05
CD1 351,1 503,0 174,2 384,2 19,4 24,3 1,38 0,03
CD2 344,1 506,5 144,8 386,5 18,1 25,1 1,49 0,04
CD3 347,1 487,0 192,5 420,8 19,7 24,7 1,63 0,04
Total 346,8 497,7 159,5 397,9 18,4 24,6 1,58 0,04
DS
CD4 5,49 0,49 32,86 68,98 1,20 1,41 0,31 0,01
CD5 7,37 2,35 28,87 68,65 1,15 1,29 0,32 0,01
CD1 0,78 5,91 27,15 56,82 0,95 1,15 0,26 0,01
CD2 0,79 0,49 21,56 54,89 0,83 1,08 0,26 0,01
CD3 0,84 0,00 28,81 57,97 1,02 1,11 0,29 0,01
Total 6,08 7,37 35,27 63,95 1,45 1,23 0,32 0,01
CV
CD4 1,56 0,10 21,53 18,03 6,68 5,79 18,98 14,72
CD5 2,17 0,47 21,72 16,53 6,75 5,26 18,06 12,91
CD1 0,22 1,18 15,58 14,79 4,90 4,72 18,97 15,93
CD2 0,23 0,09 14,89 14,20 4,57 4,29 17,63 10,92
CD3 0,24 0,00 14,96 13,78 5,20 4,49 17,99 18,36
Total 1,75 1,48 22,11 16,07 7,83 5,07 20,36 20,10
41
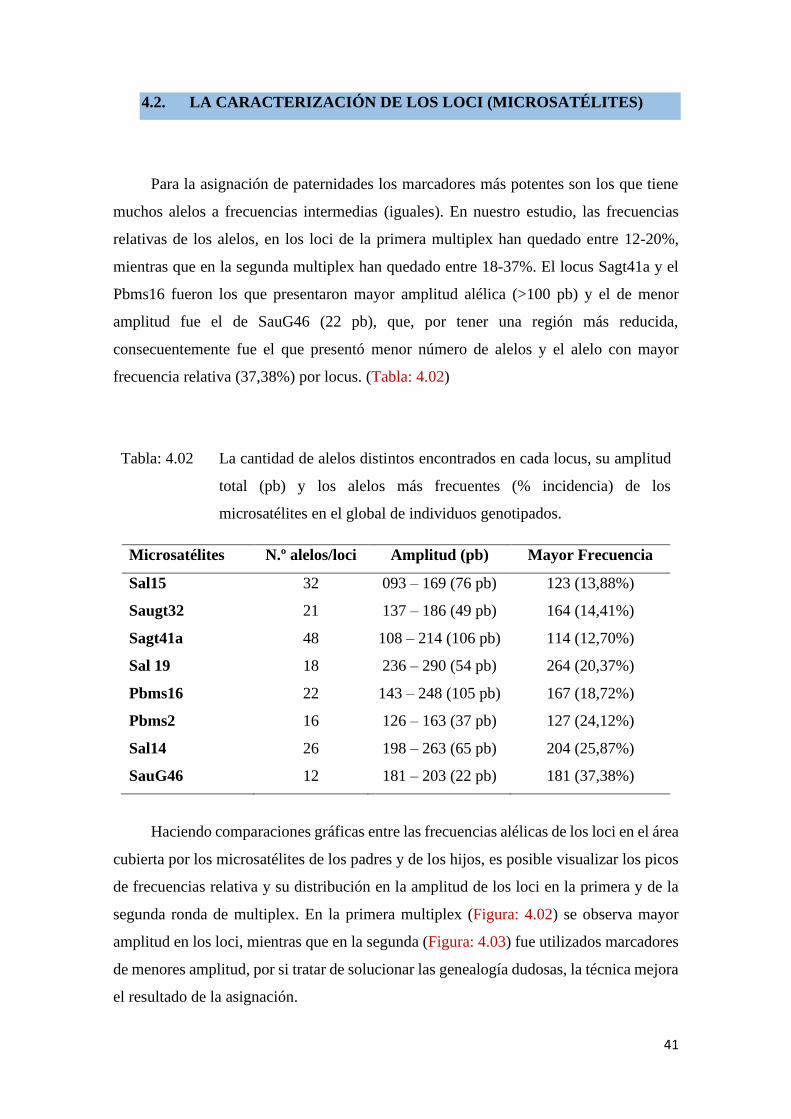
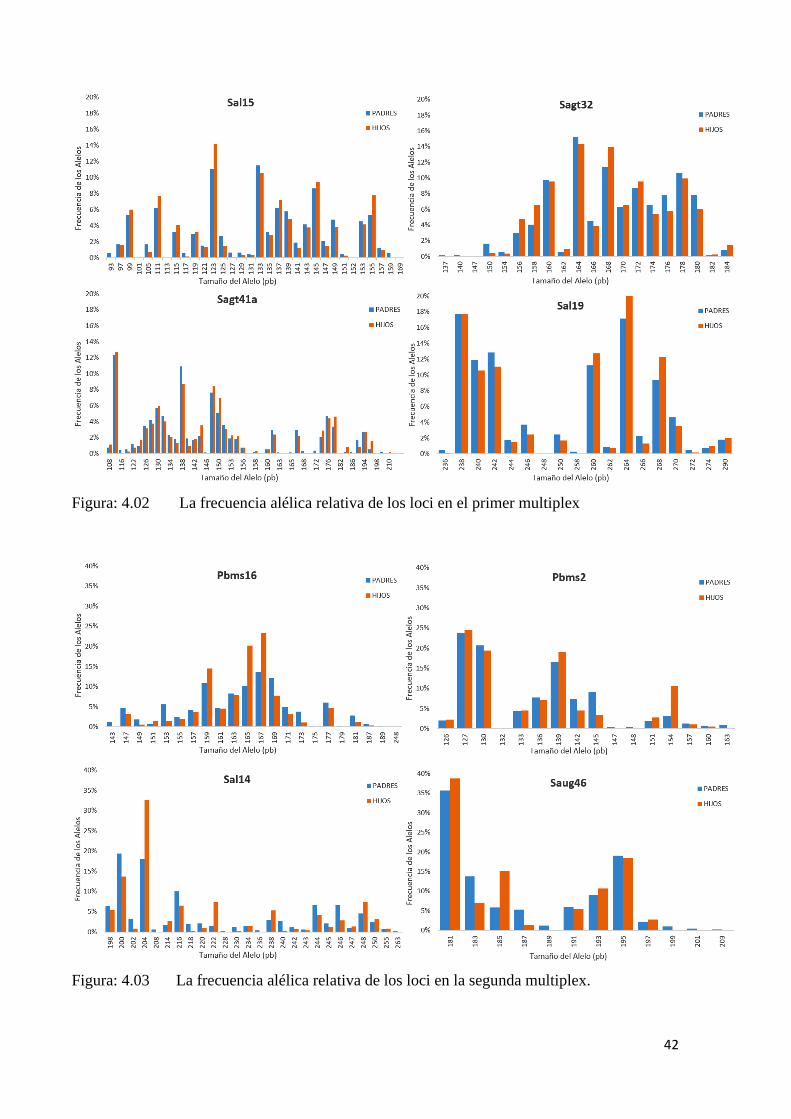
4.2. LA CARACTERIZACIÓN DE LOS LOCI (MICROSATÉLITES)
Para la asignación de paternidades los marcadores más potentes son los que tiene
muchos alelos a frecuencias intermedias (iguales). En nuestro estudio, las frecuencias
relativas de los alelos, en los loci de la primera multiplex han quedado entre 12-20%,
mientras que en la segunda multiplex han quedado entre 18-37%. El locus Sagt41a y el
Pbms16 fueron los que presentaron mayor amplitud alélica (>100 pb) y el de menor
amplitud fue el de SauG46 (22 pb), que, por tener una región más reducida,
consecuentemente fue el que presentó menor número de alelos y el alelo con mayor
frecuencia relativa (37,38%) por locus. (Tabla: 4.02)
Tabla: 4.02 La cantidad de alelos distintos encontrados en cada locus, su amplitud
total (pb) y los alelos más frecuentes (% incidencia) de los
microsatélites en el global de individuos genotipados.
Microsatélites N.º alelos/loci Amplitud (pb) Mayor Frecuencia
Sal15 32 093 – 169 (76 pb) 123 (13,88%)
Saugt32 21 137 – 186 (49 pb) 164 (14,41%)
Sagt41a 48 108 – 214 (106 pb) 114 (12,70%)
Sal 19 18 236 – 290 (54 pb) 264 (20,37%)
Pbms16 22 143 – 248 (105 pb) 167 (18,72%)
Pbms2 16 126 – 163 (37 pb) 127 (24,12%)
Sal14 26 198 – 263 (65 pb) 204 (25,87%)
SauG46 12 181 – 203 (22 pb) 181 (37,38%)
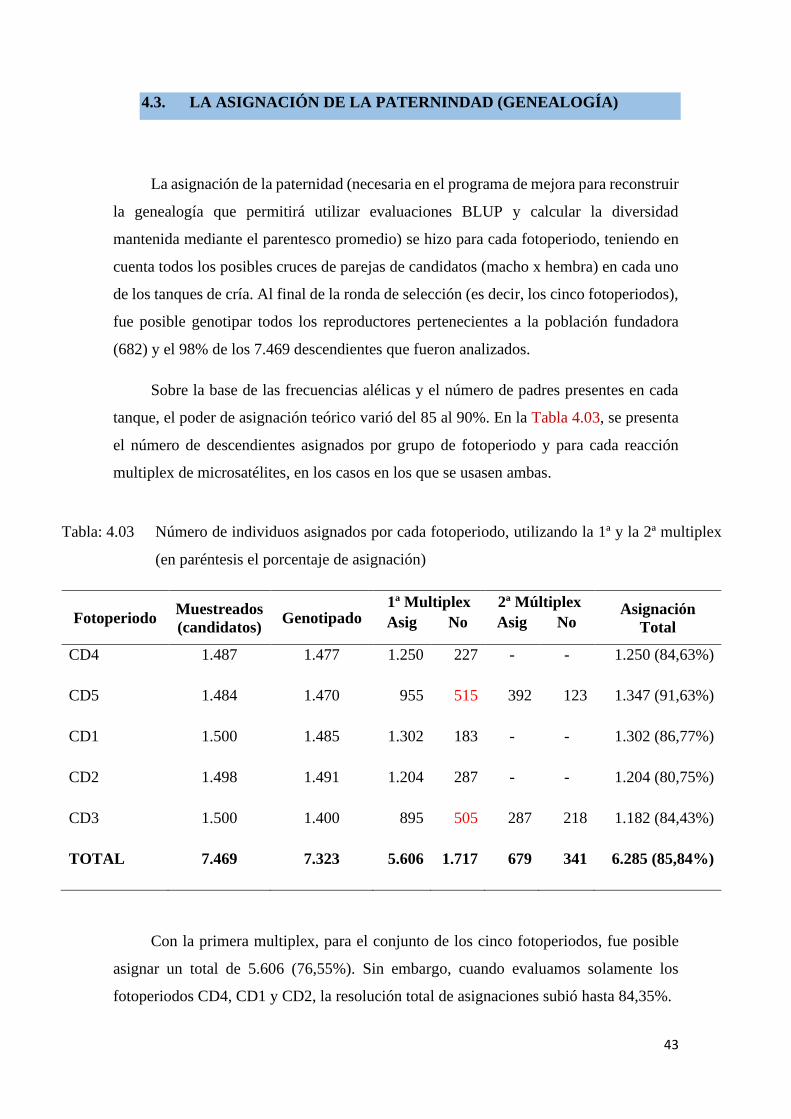
Haciendo comparaciones gráficas entre las frecuencias alélicas de los loci en el área
cubierta por los microsatélites de los padres y de los hijos, es posible visualizar los picos
de frecuencias relativa y su distribución en la amplitud de los loci en la primera y de la
segunda ronda de multiplex. En la primera multiplex (Figura: 4.02) se observa mayor
amplitud en los loci, mientras que en la segunda (Figura: 4.03) fue utilizados marcadores
de menores amplitud, por si tratar de solucionar las genealogía dudosas, la técnica mejora
el resultado de la asignación.
42
Figura: 4.02 La frecuencia alélica relativa de los loci en el primer multiplex
Figura: 4.03 La frecuencia alélica relativa de los loci en la segunda multiplex.
43
4.3. LA ASIGNACIÓN DE LA PATERNINDAD (GENEALOGÍA)
La asignación de la paternidad (necesaria en el programa de mejora para reconstruir
la genealogía que permitirá utilizar evaluaciones BLUP y calcular la diversidad
mantenida mediante el parentesco promedio) se hizo para cada fotoperiodo, teniendo en
cuenta todos los posibles cruces de parejas de candidatos (macho x hembra) en cada uno
de los tanques de cría. Al final de la ronda de selección (es decir, los cinco fotoperiodos),
fue posible genotipar todos los reproductores pertenecientes a la población fundadora
(682) y el 98% de los 7.469 descendientes que fueron analizados.
Sobre la base de las frecuencias alélicas y el número de padres presentes en cada
tanque, el poder de asignación teórico varió del 85 al 90%. En la Tabla 4.03, se presenta
el número de descendientes asignados por grupo de fotoperiodo y para cada reacción
multiplex de microsatélites, en los casos en los que se usasen ambas.
Tabla: 4.03 Número de individuos asignados por cada fotoperiodo, utilizando la 1ª y la 2ª multiplex
(en paréntesis el porcentaje de asignación)
Fotoperiodo Muestreados
(candidatos) Genotipado
1ª Multiplex 2ª Múltiplex
Asignación
Total Asig No Asig No
CD4 1.487 1.477 1.250 227 - - 1.250 (84,63%)
CD5 1.484 1.470 955 515 392 123 1.347 (91,63%)
CD1 1.500 1.485 1.302 183 - - 1.302 (86,77%)
CD2 1.498 1.491 1.204 287 - - 1.204 (80,75%)
CD3 1.500 1.400 895 505 287 218 1.182 (84,43%)
TOTAL 7.469 7.323 5.606 1.717 679 341 6.285 (85,84%)
Con la primera multiplex, para el conjunto de los cinco fotoperiodos, fue posible
asignar un total de 5.606 (76,55%). Sin embargo, cuando evaluamos solamente los
fotoperiodos CD4, CD1 y CD2, la resolución total de asignaciones subió hasta 84,35%.

44
Esta diferencia se debe a que los fotoperiodos CD5 y CD3 han presentado
resultados de asignación muy baja, del 64,96% y 63,93% respectivamente. Del total de
2.870 individuos disponibles en estos fotoperiodos solo 1.850 han sido asignados a una
única pareja de padres. Los otros 1.020 individuos restantes, que presentaron parentesco
dudoso (que fueron asignados a más de un padre o una madre) pasaron un nuevo análisis
con la segunda multiplex. El resultado obtenido permitió la asignación de un conjunto
extra de 679 individuos (66,57%), siendo 392 para el CD5 y 287 para el CD3. En total,
con las dos multiplexes, fue posible reconstruir la genealogía de 6.285 descendientes
(85,84%) de los 7.323 que fueron genotipados correctamente.
De un modo general, la potencia de asignación fue más o menos la misma para
todos los fotoperiodos. Con excepción del fotoperiodo CD4, que presentó mayor
diferencia entre los tanques. Los fotoperiodos CD5 y CD3 una vez que se añadieron los
asignados en la segunda multiplex han alcanzado el mismo patrón (Figura 4:04).
Figura: 4.04 Número de individuos asignados en cada tanque dentro de cada
fotoperiodo. La línea roja es el promedio, y el área roja rayada es la
desviación estándar. La columna azul clara es la segunda multiplex.
Tratando de determinar las causas de la menor precisión de la asignación en los dos
fotoperiodos mencionados, se calculó el coeficiente de parentesco molecular y el

45
coeficiente de endogamia (homocigosis observada) para los reproductores de los dos
tanques. El parentesco en los dos tanques del fotoperiodo CD5 y los otros dos del CD3
fue hasta un 15% más alto que la promedio para los otros tanques y la endogamia un 40%
más alto. Este hecho puede estar detrás del peor desempeño al reconstruir las familias
observado en esos tanques.
En los casos en los que se aplicaron las dos multiplexes (CD5 y CD3), podemos
calcular la ganancia obtenida aumentando el número de marcadores en términos del
número extra de reproductores con descendencia asignada al usar la segunda (Tabla: 4.04)
Tabla: 4.04 Número de machos y hembras disponibles y las progenies asignadas, en los
casos en que se han utilizado 4 y 8 microsatélites
Fotoperiodo Tanque Machos Hembras Machos Asignados Hembras Asignadas
4 micros 8 micros 4 micros 8 micros
CD5 3 19 23 14 14 15 15
4 19 30 13 17 21 21
CD3 9 18 39 12 12 17 17
10 30 37 17 20 30 30
Con el aumento del número de microsatélites genotipados, se pudieron asignar 318
descendientes nuevos en el tanque 4 y 227 individuos en el tanque 10 suponiendo un
aumento de 4 y 3 machos representados, respectivamente. Sin embargo, en los otros dos
tanques que hubo nuevos individuos asignados con la segunda multiplex (74 en el tanque
3 y 60 individuos para el tanque 9, como muestra la Figura 4:04), estos individuos
provenían de los mismos reproductores que ya tenían contribución con la primera
multiplex.
Por lo tanto, los nuevos individuos asignados no aumentan significativamente el
número de familias de medios hermanos (HS) ya que apenas la contribución de una
pequeña cantidad de nuevos machos fue detectada. Los valores de Ne obtenidos de la
varianza de las contribuciones pasaron de 21,94 a 22,51, de 18,54 a 18,30, de 6,84 a 16,81
y de 34,36 a 45,21 para los tanques 3, 4, 9 y 10 respectivamente. También es notable que

46
el aumento en familias de hermanos (FS) al comparar la asignación con 4 y 8
microsatélites es mayor que el número de nuevas familias HS detectadas. Esta es otra
indicación de que el número limitado de reproductores que contribuyen no es una
consecuencia del proceso de asignación, sino que refleja el hecho biológico de que no
todos los padres se reproducen o de que en el proceso de muestreo desde la puesta hasta
la selección de los individuos candidatos (los genotipados y medidos) hay un sesgo que
hace que no todos los reproductores estén representados.
4.3.1. LAS CONTRIBUCIONES DE LOS REPRODUCTORES
Al reconstruir la genealogía, hemos podido identificar las contribuciones
individuales de cada reproductor y su participación efectiva en el grupo de candidatos. El
resultado mostró que no todos los reproductores han contribuido para la formación de los
candidatos que han sido asignados, y que hubo diferencias significativas en la
comparación entre machos y hembras, fotoperiodo e incluso entre tanques. Una
distribución desequilibrada de la contribución supone una reducción del tamaño efectivo
esperado de la población.
En relación a la participación, los machos han estado mejor representados. De los
178 posibles padres, un 79,21% ha participado para la formación de la progenie. El
fotoperiodo CD4 fue el que presentó mayores índices (95,45%), mientras que el CD3 fue
el peor con 66,67% en la participación de machos en sus dos tanques. En las hembras, de
las 319 disponibles para la reproducción, solamente 215 han contribuido a la
descendencia (67,40%). El fotoperiodo CD4 obtuvo nuevamente el mejor índice como
fotoperiodo (86,27%) y uno de sus tanques consiguió el mayor porcentaje de hembras
representadas (91,30%). Por el contrario, los peores resultados promedio han sido
observados en el fotoperiodo CD1 (58,97%), aunque el tanque con porcentaje más bajo
fue el 9 que pertenecía al fotoperiodo CD3 (apenas un 43,59% de las hembras
disponibles).
En general, el promedio de individuos disponibles en cada tanque y los que
realmente contribuyen, fue de 17,8 ± 5,9 y 14,1 ± 3,7 respectivamente para machos, y de

47
31,9 ± 6,1 y 21,5 ± 4,1 en las hembras, lo que significó una reducción de
aproximadamente 18% y 32%, respectivamente (Figura: 4.05).
Figura: 4.05 La cantidad de reproductores machos (A) y hembras (B), que estaban disponibles
(inicial) para la reproducción y los que han contribuido (final).
Teniendo en cuenta sólo los descendientes que fueron asignados, la contribución
media general de los reproductores para todos los fotoperiodos, fue de 44,57 ± 47,87 para
los machos (oscilando desde 32 hasta 95 individuos) y para las hembras de 29,23 ± 31,37
(con contribuciones que iban desde 19 hasta los 41 descendientes).
La mayor variación en las contribuciones masculinas individuales se observó en el
tanque 2, (CD4), donde se constató que un único reproductor fue responsable por producir
345 individuos, representando el 40% de toda la progenie asignada en este tanque,
mientras el que menos ha contribuido ha dejado apenas 20 individuos, generando una
contribución promedio de aproximadamente 95 ± 32 individuos. Mientras que, el tanque
10 (CD3), fue el que registró mayor participación de los reproductores con promedio de
contribuciones de 32 ± 5 individuos, siendo que el macho que más contribuyo no ha
sobrepasado los 14% en la representatividad de la progenie. Para las hembras, las
contribuciones fueron más uniformes cuando consideramos todos los fotoperiodos,
excepto por los tanques 3 (CD5) y el tanque 8 (CD2), que presentaron una única hembra
contribuyendo con más del 25% de la progenie, con un promedio de 41 ± 10 (CD5) y 37
± 9 (CD2), respectivamente para los tanques 3 y 8. Mientras que el tanque 1 (CD4) ha
sido el que tuvo la contribución más equilibrada, con promedio de 19 ± 3 individuos.
(Figura 4:06).
A B
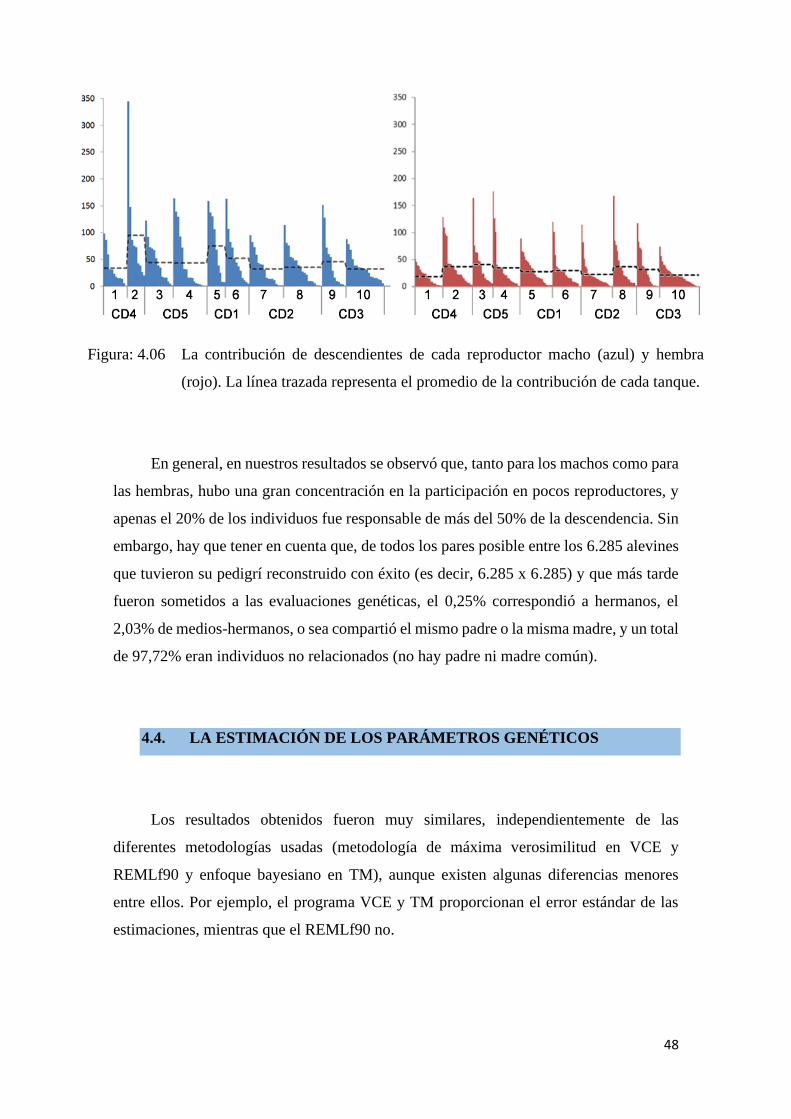
48
Figura: 4.06 La contribución de descendientes de cada reproductor macho (azul) y hembra
(rojo). La línea trazada representa el promedio de la contribución de cada tanque.
En general, en nuestros resultados se observó que, tanto para los machos como para
las hembras, hubo una gran concentración en la participación en pocos reproductores, y
apenas el 20% de los individuos fue responsable de más del 50% de la descendencia. Sin
embargo, hay que tener en cuenta que, de todos los pares posible entre los 6.285 alevines
que tuvieron su pedigrí reconstruido con éxito (es decir, 6.285 x 6.285) y que más tarde
fueron sometidos a las evaluaciones genéticas, el 0,25% correspondió a hermanos, el
2,03% de medios-hermanos, o sea compartió el mismo padre o la misma madre, y un total
de 97,72% eran individuos no relacionados (no hay padre ni madre común).
4.4. LA ESTIMACIÓN DE LOS PARÁMETROS GENÉTICOS
Los resultados obtenidos fueron muy similares, independientemente de las
diferentes metodologías usadas (metodología de máxima verosimilitud en VCE y
REMLf90 y enfoque bayesiano en TM), aunque existen algunas diferencias menores
entre ellos. Por ejemplo, el programa VCE y TM proporcionan el error estándar de las
estimaciones, mientras que el REMLf90 no.
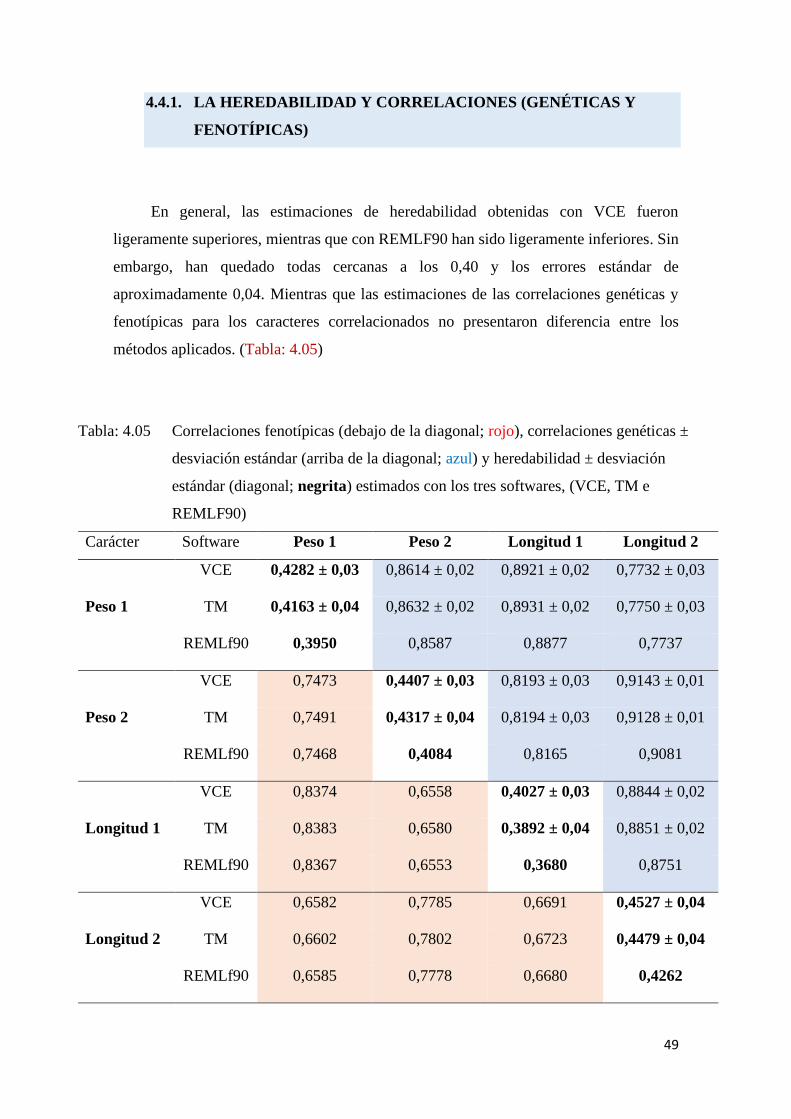
49
4.4.1. LA HEREDABILIDAD Y CORRELACIONES (GENÉTICAS Y
FENOTÍPICAS)
En general, las estimaciones de heredabilidad obtenidas con VCE fueron
ligeramente superiores, mientras que con REMLF90 han sido ligeramente inferiores. Sin
embargo, han quedado todas cercanas a los 0,40 y los errores estándar de
aproximadamente 0,04. Mientras que las estimaciones de las correlaciones genéticas y
fenotípicas para los caracteres correlacionados no presentaron diferencia entre los
métodos aplicados. (Tabla: 4.05)
Tabla: 4.05 Correlaciones fenotípicas (debajo de la diagonal; rojo), correlaciones genéticas ±
desviación estándar (arriba de la diagonal; azul) y heredabilidad ± desviación
estándar (diagonal; negrita) estimados con los tres softwares, (VCE, TM e
REMLF90)
Carácter Software Peso 1 Peso 2 Longitud 1 Longitud 2
Peso 1
VCE 0,4282 ± 0,03 0,8614 ± 0,02 0,8921 ± 0,02 0,7732 ± 0,03
TM 0,4163 ± 0,04 0,8632 ± 0,02 0,8931 ± 0,02 0,7750 ± 0,03
REMLf90 0,3950 0,8587 0,8877 0,7737
Peso 2
VCE 0,7473 0,4407 ± 0,03 0,8193 ± 0,03 0,9143 ± 0,01
TM 0,7491 0,4317 ± 0,04 0,8194 ± 0,03 0,9128 ± 0,01
REMLf90 0,7468 0,4084 0,8165 0,9081
Longitud 1
VCE 0,8374 0,6558 0,4027 ± 0,03 0,8844 ± 0,02
TM 0,8383 0,6580 0,3892 ± 0,04 0,8851 ± 0,02
REMLf90 0,8367 0,6553 0,3680 0,8751
Longitud 2
VCE 0,6582 0,7785 0,6691 0,4527 ± 0,04
TM 0,6602 0,7802 0,6723 0,4479 ± 0,04
REMLf90 0,6585 0,7778 0,6680 0,4262

50
Las heredabilidades para el carácter del peso en la primera y la segunda medida
fueron similares, mientras que para la longitud difirieron más entre las dos edades. Siendo
el valor estimado más bajo observado para la longitud a la primera edad con el REMLF90
(0,36) y el valor más alto para la longitud a la segunda edad con el VCE (0,45). Sin
embargo, ninguna de estas diferencias ha sido significativas.
Las correlaciones genéticas más altas fueron entre el peso y la longitud a la misma
edad (0,89 para la primera edad y 0,91 para la segunda edad) y la más baja entre el peso
y la longitud en diferentes edades (0,77 entre el peso a la primera edad y la longitud a la
segunda, y de 0,81 entre la longitud a la primera edad y el peso a la segunda edad).
Las correlaciones fenotípicas siguieron el mismo patrón, máximo entre peso y
longitud a la misma edad (0,83 para la primera edad y 0,77 para la segunda edad) y
mínimo entre diferentes rasgos en diferentes edades (0,66 entre el peso a la primera edad
y la segunda edad y 0,65 entre la longitud a la primera edad y el peso a la segunda edad).
4.4.2. LAS ESTIMACIONES DEL EFECTO FIJO (FOTOPERIODO)
En la Tabla 4.06 se muestran las estimaciones del efecto de fotoperiodo obtenidas
con el software VCE 6.0. De acuerdo con los resultados estimados, se hace notar que
existe un efecto fijo significativo entre los ciclos que se debe a las condiciones específicas
de cada fotoperiodo. Existe un buen acuerdo entre estas estimaciones y el promedio de
los valores fenotípicos de los correspondientes fotoperiodos, aunque las diferencias se
magnifican.
Tabla: 4.06 Estimación de los efectos de factores fijos de los fotoperiodos para los
diferentes caracteres
Fotoperiodo Peso 1 (g) Peso 2 (g) Longitud 1 (cm) Longitud 2 (cm)
CD4 -43,33 -45,67 -1,84 -0,5
CD5 -58,41 -5,03 -2,83 -0,39
CD1 -17,70 -30,52 -0,37 -0,40
CD2 -49,91 -43,22 -1,76 0,13
CD3 0,00 0,00 0,00 0,00

51
4.5. LA SELECCIÓN POR OPTIMIZACIÓN
Para realizar la selección, se decidió utilizar los valores del peso en la segunda
medida, que, además de presentar mayor variancia genética y fenotípica, son también los
más cercanos a la talla comercial. Pero, para que no se incremente demasiado la
consanguinidad, hay que elegir los seleccionados no solo por su valor mejorante (EBV´s)
sino que se impone una restricción sobre el parentesco promedio de los seleccionados.
Para eso, hemos seleccionado aproximadamente el 5% de los candidatos disponibles en
cada fotoperiodo, que es el número de individuos que se ha calculado que se necesitan
para hacer la reposición de los reproductores. Pero se pone un valor máximo de parentesco
entre los seleccionados (consanguinidad en la descendencia) que no se quiere pasar. En
la práctica se generan varias soluciones, desde aquella en la que el parentesco es el
mínimo posible (I) y se va aumentando hasta aquella en la que no hay restricciones (IV).
Con el resultado de todas las soluciones, se traza una línea de tendencia con la que se
busca una solución que equilibre la respuesta esperada con el parentesco promedio de los
seleccionados. En la Figura: 4.07 se muestra el resultado gráfico de las soluciones
obtenidas para cada fotoperiodo, desde el valor mejorante inicial (sin selección), hasta la
máxima respuestas esperada (IV).
Figura: 4.07 La optimización de la selección para los diferentes niveles de restricción para
la consanguinidad. Donde el eje Y son los valores mejorantes estimados
(Estimated Breeding Values - EBV´s) y en el eje X está el parentesco
promedio de la población seleccionada (f)

52
En el gráfico, se observa que los valores genéticos estimados por BLUP van
creciendo a medida que se va dando restricciones menos estrictas/exigentes en el
parentesco medio de los seleccionados. Sin embargo, el aumento es muy bajo para valores
de parentesco más elevados. Por tanto, se puede conseguir una reducción importante en
el nivel de parentesco de los seleccionados sin perder mucho en valor genético respecto
del máximo posible que corresponde a no hacer restricción.
En los fotoperiodos CD1 y CD3, se observó que, en la situación inicial la población
presentaba un valor promedio del BLUP muy reducido, y con una ligera aplicación de la
restricción eligiendo individuos en la situación I ya ha sido suficiente para producir un
aumento significativo en el promedio de la población, que había iniciado con valores
próximos a cero hacia valores superiores a dos docenas. En estos casos, la curva ya
empieza a linealizar incluso antes de restringir hacia los medios-hermanos, y pone de
manifiesto que, en estas poblaciones, no es adecuado relajar las restricciones.
Comparando con el peso promedio de la población en la situación inicial (población
total disponible para la selección, o sea, los con genealogía asignados) y en las cuatro
diferentes situaciones de restricciones para el parentesco medio global, hemos constatado
que el diferencial de selección corresponde a un aumento del peso de los seleccionados
de aproximadamente 10,5% en la situación I, y pasó a más del doble cuando fueron
seleccionados los individuos en la situación II, alcanzando el 23,2% en el incremento del
peso promedio de la población inicial. Mientras que el intervalo comprendido entre la
situación III y la situación IV, el porcentaje de incremento peso promedio de la población
seleccionada tiende a estabilizar cerca de los 26%. Y en el caso del fotoperiodo CD1 y
CD3 el límite de aumento en el peso promedio de la población ya está prácticamente
alcanzado (Tabla: 4.07)
La alta tasa reproductiva de la dorada (en general de las especies de acuicultura)
permite que se pueda seleccionar un porcentaje muy pequeño de reproductores respecto
del total de evaluados. De esa manera el diferencial de selección es considerable incluso
en los casos en los que la restricción en parentesco es máxima. Así el peso en los
seleccionados en el escenario I es entre un 10% y un 15% (según fotoperiodos) más alto
que en el conjunto de los candidatos. En el escenario sin restricciones este incremento
puede llegar al 36% (CD4). Esta circunstancia, unida a la moderada heredabilidad para el
peso, hace que la respuesta esperada a la selección sea elevada.
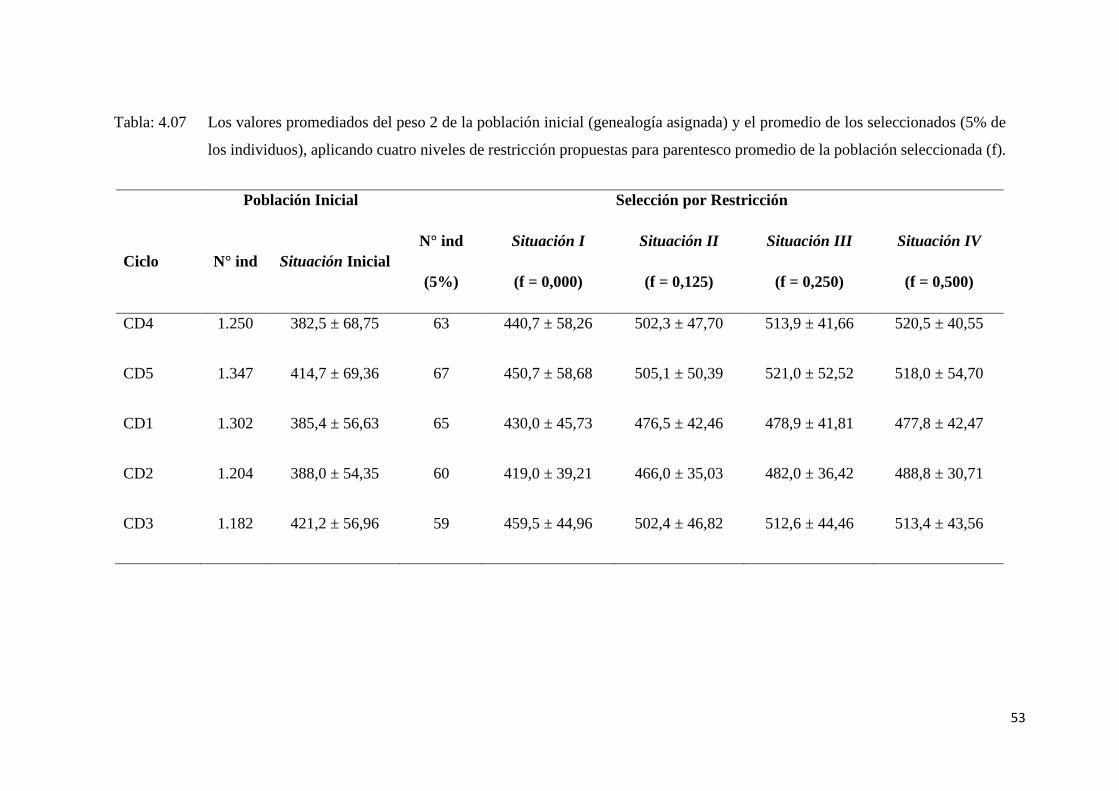
53
Tabla: 4.07 Los valores promediados del peso 2 de la población inicial (genealogía asignada) y el promedio de los seleccionados (5% de
los individuos), aplicando cuatro niveles de restricción propuestas para parentesco promedio de la población seleccionada (f).
Población Inicial Selección por Restricción
Ciclo N° ind Situación Inicial
N° ind
(5%)
Situación I
(f = 0,000)
Situación II
(f = 0,125)
Situación III
(f = 0,250)
Situación IV
(f = 0,500)
CD4 1.250 382,5 ± 68,75 63 440,7 ± 58,26 502,3 ± 47,70 513,9 ± 41,66 520,5 ± 40,55
CD5 1.347 414,7 ± 69,36 67 450,7 ± 58,68 505,1 ± 50,39 521,0 ± 52,52 518,0 ± 54,70
CD1 1.302 385,4 ± 56,63 65 430,0 ± 45,73 476,5 ± 42,46 478,9 ± 41,81 477,8 ± 42,47
CD2 1.204 388,0 ± 54,35 60 419,0 ± 39,21 466,0 ± 35,03 482,0 ± 36,42 488,8 ± 30,71
CD3 1.182 421,2 ± 56,96 59 459,5 ± 44,96 502,4 ± 46,82 512,6 ± 44,46 513,4 ± 43,56

54
5.
DISCUSIÓN

55
5. DISCUSIÓN
5.1. LOS PROGRAMAS DE MEJORA GENÉTICA EN DORADA
La dorada es actualmente uno de los peces cultivados más importantes en Europa,
especialmente en la región del mediterráneo, y es por eso que hay una creciente demanda
de las empresas para implementar programas de mejora genética buscando mejorar su
producción. La industria acuícola está interesada en caracteres comunes para la mayoría
de las especies cultivadas y comparte una serie de objetivos de selección, entre los que se
incluyen tanto los que influyen sobre los costes de producción y beneficio comercial, así
como aquellos que condicionan las preferencias del consumidor.
En este sentido, el objetivo de la mejora en peces suele ser para el crecimiento y el
tamaño corporal, la edad a la madurez sexual, la resistencia a enfermedades, la calidad de
la carne y el índice de conversión de alimento en peso (Toro y López-Fanjul, 2007).
Aunque desde el punto de vista económico, el interés para el incremento de la aptitud
cárnica son los más relevantes y donde normalmente se obtienen los mejores resultados.
Para poder implementar un programa de mejora genética en dorada sería necesario,
tener en cuenta las condiciones industriales e intereses de producción en cada una de sus
fases para la definición de los caracteres objeto de la selección, la estima de parámetros
genéticos y la cuantificación de la interacción genotipo x ambiente (GxA). Las industrias
criadoras de dorada realizan la reproducción utilizando un elevado número de animales,
realizando el desove por medio de puestas masales. En este sentido, la determinación de
las relaciones familiares solamente es posible con la utilización conjunta de marcadores
físicos y moleculares (Wesmajervi, et al., 2006). La utilización de los marcadores
moleculares asociados a las marcaciones físicas (PIT-Tag) junto con aplicaciones
bioinformáticas se utilizan ampliamente para la reconstrucción de las relaciones
familiares en diversos organismos acuáticos (Fernández y Toro, 2006). En dorada, estas
herramientas podrían favorecer las condiciones necesarias para ejecutar programas de
mejora genética por selección de esta especie.
Si la obtención y mantenimiento de estas informaciones en organismos acuáticos
ya es una tarea complicada, el problema se torna más difícil aun en el caso de la

56
reproducción de la dorada, donde las hembras desovan en tandas intermitentes, pudiendo
poner de 20.000 – 80.000 huevos al día durante un periodo de hasta 4 meses seguidos
(Ortega, 2008). En ABSA, las siembras se realizan con intervalos de 5 días
aproximadamente para maximizar la contribución de los individuos. Con este volumen
de huevos producidos por medio de puestas masales, recolectados durante días
consecutivos, sin que sean mantenida las familias separadas hasta la talla de marcaje,
mantener la información genealógica sin la utilización de marcadores moleculares es una
tarea prácticamente imposible.
Además, la problemática de mantener el control del plantel reproductor se agrava
más aún cuando se trata de una especie que cambia del sexo, y puede contribuir una
generación como macho y en la siguiente como hembra. En dorada, durante el primer año
de vida suelen ser inmaduros, y la primera maduración sexual suele ocurrir durante el
segundo año de vida, cuando se desarrolla la parte ventral de la gónada originando
testículos funcionales. Al final de esta primera estación reproductora, todos los individuos
inician un proceso de reversión sexual, que se concluye en aproximadamente un 80% de
la población que a partir del tercer año de vida se comportarán como hembras. El 20%
restante interrumpe los procesos de reversión y permanece como macho. La maduración
de la gónada es progresiva, y esto se traduce en una puesta secuenciada que realiza
durante 2-4 meses, a razón de unos miles de huevos al día (Ortega, 2008).
Sin embargo, en nuestros datos hubo registro de algunos casos de machos con más
de 6 años, y nunca han cambiado de sexo, así como individuos que ya habían cambiado
hacia hembra con edades inferiores a los 2 años. Aparentemente no se constató ninguna
relación de reversión con el peso o con la longitud, aunque en los periodos reproductivos
los peces suelen reducir su ritmo de crecimiento, puesto que dirigen sus recursos hacia el
desarrollo de los aparatos reproductivos.
Con el advenimiento de la tecnología del ADN recombinante, se han desarrollado
toda una gama de marcadores moleculares que combinan la especificidad de la reacción
en cadena de la polimerasa (PCR), con la capacidad analítica de los secuenciadores
automáticos para la separación de los fragmentos. Los marcadores genéticos tienen una
variedad de usos de interés para la acuicultura, y su aplicación viene posibilitando el
desarrollo de diversas investigaciones sobre cómo aprovechar mejor los recursos
genéticos de los organismos acuáticos, como, por ejemplo: caracterizar la variabilidad

57
genética de poblaciones (Li, et al., 2017), identificar especies y cepas (Briñez, et al.,
2011), asignar paternidad y parentesco (Martínez y Fernández, 2008), apoyar la
trazabilidad (Parreño-Marchante, et al., 2014), identificar loci asociados a caracteres
cuantitativos (QTL) (Laghari, et al., 2014) así como realizar selección asistida por
marcadores (MAS) (Yue, 2014). Avances tecnológicos más recientes utilizando chips de
alta densidad, está posibilitando realizar el mapeamiento de los SNPs (Single Nucleotide
Polymorphism), utilizados en conjunto con plataformas modernas de genotipado, son
capaces de procesar muchas amuestras para muchos marcadores en un único análisis,
proporcionando la inclusión de la información genómica en programas cría selectiva en
acuicultura, su aplicación es denominada de selección genómica (Zenger, et al., 2019).
Estos avances aumentarán la eficiencia de los programas de cría selectiva, que ahora
pueden identificar los animales individualmente, incluso cuando son mantenidos todos
juntos, en el mismo tanque durante todo su desarrollo. Desde el punto de vista genético,
esta estrategia presenta la ventaja de minimizar las fuentes de parecido por ambiente
común entre los miembros de la misma familia (Herbinger, et al., 1999), lo que
proporciona mayor precisión en las estimas tanto de los parámetros genéticos como de
los valores mejorantes.
Obtener estas informaciones son muy importantes, principalmente cuando se trata
de una especie que tiene tanta oscilación en las condiciones de cultivo, desde la eclosión
hasta el periodo de cosecha. Los alevines son pre-engordados y engordados
fundamentalmente en jaulas situadas en el Mar Mediterráneo o en el océano Atlántico, y
en coordenadas geográficas muy diferentes, como es el caso de Canarias donde las
temperaturas son más elevadas y estables a lo largo de todo el año frente al Mediterráneo.
Sin embargo, también se engordan alevines de dorada con el sistema de esteros o tanques
en tierra, propios del sur de España. Todas estas variaciones existentes en los sistemas de
producción y la localización de las instalaciones de dorada resultan en diferentes
caracteres de interés para los piscicultores. Por lo que es de utilidad estimar el
comportamiento productivo de las poblaciones considerando las interacciones GxA que
pueden limitar o incluso perjudicar el progreso genético (Saillant, et al., 2006; Dupont-
Nivet, et al., 2008).
En acuicultura, la utilización de sistemas de marcación física aún tiene algunas
limitaciones de tamaño del individuo, lo que implica que los grupos familiares tienen que

58
ser criados por separado hasta que los individuos son lo suficientemente grandes para ser
etiquetados. Esto requiere costosas instalaciones de multi-tanques, y pueden también
presentar efectos ambientales sistemáticos comunes a hermanos. Además, para algunas
especies, no hay métodos de etiquetado físico fiable disponibles
El desarrollo de la marcación física individual por medio los microchips, asociados
a las técnicas de marcadores moleculares, han posibilitado trazar perfiles de identificación
molecular de la familia, representando un hito en la cría selectiva de las especies
acuáticas. Esta técnica permite que diferentes grupos de familiares se mantengan en un
tanque común desde la fertilización, reduciendo sustancialmente los costes implicados en
la cría y eliminando los problemas relacionados con los efectos ambientales comunes.
También permite trabajar con un mayor número de familias y por lo tanto facilita el uso
de intensidades de selección más altas sin la rápida acumulación de endogamia.
De acuerdo con (Toro y López, 2007), los principios generales para si establecer un
programa de mejora de los rasgos productivos en organismos acuáticos debe ser dado por
i) la definición del objetivo de mejora en el contexto de un determinado sistema de
producción; ii) la elección de la población base y, si procede, el sistema de cruzamiento;
iii) el desarrollo de un sistema de evaluación y selección; iv) la transmisión del progreso
genético obtenido en el núcleo mejorado a las poblaciones comerciales. En este sentido,
los procedimientos realizados en este proyecto para llevar a cabo los principios generales
citados arriba, fueron realizados de acuerdo con el siguiente cronograma. (Figura: 5.01)

59
Figura 5.01 El cronograma paso a paso de los procedimientos realizados en el
programa de mejora por selección aplicados en el cultivo de la dorada
(Sparus aurata). 1) Plantel de reproductores; 2) Toma de datos
(moleculares y biométricos); 3) Construcción de la genealogía; 4)
Análisis de datos (bioinformática y estadística); 5) Evaluación y
selección de los individuos; 6) Clasificación y reorganización de los
reproductores del nuevo plantel.
Paralelamente a las ventajas que proporcionan los marcadores físicos y
moleculares, el desarrollo de métodos estadísticos específicos ha dado lugar a una enorme
profusión de trabajos empíricos y teóricos en este campo, constituyendo un área muy
activa de investigación (Sonesson y Meuwissen, 2009; Dong, et al., 2016; Shepherd, et
al., 2010; Neves, et al., 2012). Utilizando la estadística y la computación para determinar
las relaciones familiares, se pueden procesar de forma relativamente rápida un gran
número de muestras, considerando que la asignación de los padres está basada en el poder
de exclusión en un pequeño número de loci (Marshall, et al., 1998).

60
Una parte sustancial de los desarrollos estadísticos asociados al estudio de las
relaciones de parentesco, surge inicialmente para la resolución de problemas relacionados
con la biología evolutiva y la ecología molecular. Más que la propia identificación del
parentesco, las cuestiones que se plantean son la estimación del flujo genético o el censo
efectivo poblacional, los sistemas de apareamiento y cuidados de la prole, etc. (Wilson y
Ferguson, 2002; Jones y Ardren, 2003). Por otra parte, el estudio de las relaciones de
parentesco ha tenido aplicación en mayor medida en la piscicultura, que sigue modelos
intensivos de explotación, y menos en la producción de crustáceos, y especialmente
moluscos, con modelos extensivos o semiextensivos.
A primera vista, la asignación por exclusión de paternidad es la solución más
atractiva y simple para la asignación de parentesco. El potencial de exclusión de un locus
puede ser definido como, la capacidad para excluir un falso padre tomado al azar de la
población de referencia (normalmente la población de posibles progenitores), cuando el
otro progenitor es conocido o desconocido. El análisis depende en esencia del cálculo de
la proporción de genotipos que son incompatibles con el padre verdadero. El potencial de
exclusión será igual a la suma de las frecuencias de todos los genotipos incompatibles,
promediado sobre todos los posibles padres (Chakraborty, et al., 1988; Sancristobal y
Chevalet, 1997).
Obviamente la capacidad de exclusión dependerá del número de candidatos
putativos, del número de loci y de su polimorfismo (Wilson y Ferguson, 2002; Jones y
Ardren, 2003). Cuantos más candidatos, mayor número de loci serán necesarios para la
exclusión de todos los falsos padres. Sin embargo, dependiendo del escenario, es
frecuente que con el número de loci manejado no podamos excluir todos los falsos
candidatos, y que, en consecuencia, tengamos más de dos padres posibles para algunos
descendientes (Wilson y Ferguson, 2002), o sea, paternidad incierta. Considerando que
los costos de los microsatélites nos fuerzan a limitar el número de loci utilizados, para su
aplicabilidad a nivel industrial a larga escala, se hace necesario la utilización de
estrategias que puedan disminuir los costes operacionales y maximizar el resultado de
asignaciones esperado, reduciendo así la los parentescos dudosos.
En este sentido, el desarrollo de PCR múltiplex son de gran utilidad ya que permiten
reducir los costes económicos por reacción y optimizar los análisis de reconstrucción de
genealogía y variabilidad genética de las poblaciones de cultivo (Neff, et al., 2000;
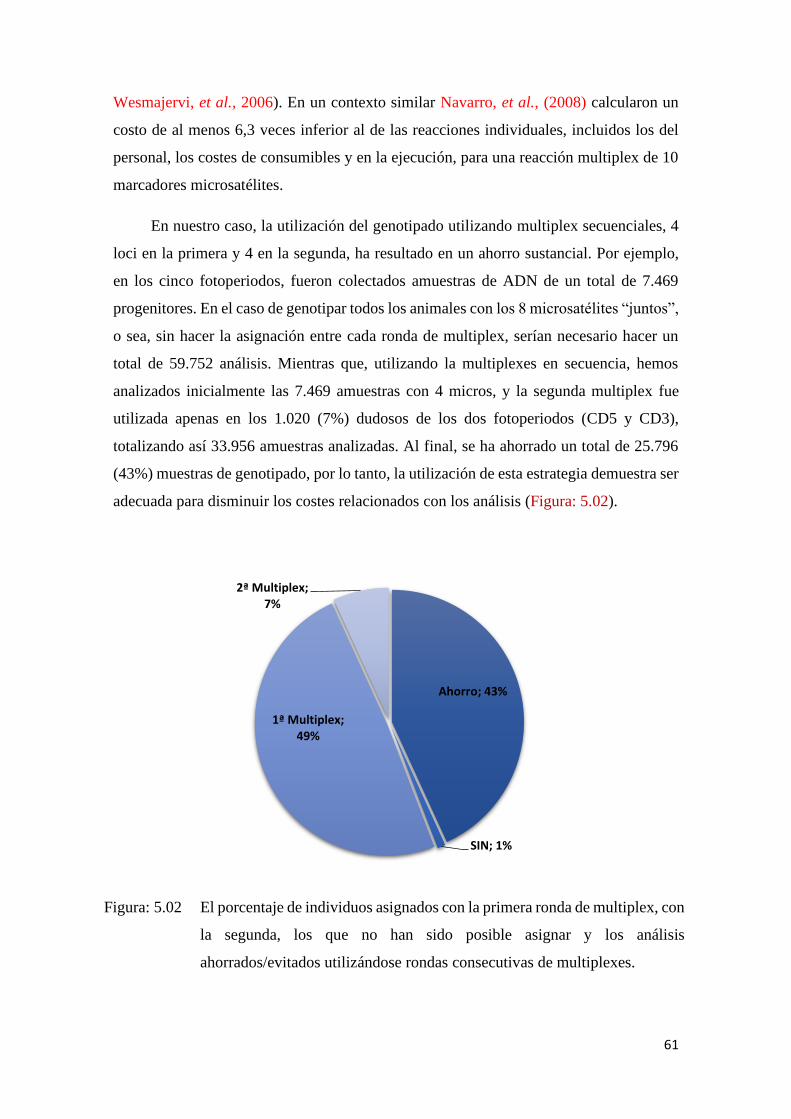
61
Wesmajervi, et al., 2006). En un contexto similar Navarro, et al., (2008) calcularon un
costo de al menos 6,3 veces inferior al de las reacciones individuales, incluidos los del
personal, los costes de consumibles y en la ejecución, para una reacción multiplex de 10
marcadores microsatélites.
En nuestro caso, la utilización del genotipado utilizando multiplex secuenciales, 4
loci en la primera y 4 en la segunda, ha resultado en un ahorro sustancial. Por ejemplo,
en los cinco fotoperiodos, fueron colectados amuestras de ADN de un total de 7.469
progenitores. En el caso de genotipar todos los animales con los 8 microsatélites “juntos”,
o sea, sin hacer la asignación entre cada ronda de multiplex, serían necesario hacer un
total de 59.752 análisis. Mientras que, utilizando la multiplexes en secuencia, hemos
analizados inicialmente las 7.469 amuestras con 4 micros, y la segunda multiplex fue
utilizada apenas en los 1.020 (7%) dudosos de los dos fotoperiodos (CD5 y CD3),
totalizando así 33.956 amuestras analizadas. Al final, se ha ahorrado un total de 25.796
(43%) muestras de genotipado, por lo tanto, la utilización de esta estrategia demuestra ser
adecuada para disminuir los costes relacionados con los análisis (Figura: 5.02).
Figura: 5.02 El porcentaje de individuos asignados con la primera ronda de multiplex, con
la segunda, los que no han sido posible asignar y los análisis
ahorrados/evitados utilizándose rondas consecutivas de multiplexes.
Ahorro; 43%
SIN; 1%
1ª Multiplex; 49%
2ª Multiplex; 7%

62
En estudios sobre el poder de resolución de los microsatélites para la dorada, Castro,
et al., (2007), utilizando 11 microsatélites a través de reacciones individuales, y Navarro,
et al., (2009b), utilizando 9 microsatélites en una misma ronda de multiplexes, han
logrado el 100% de asignaciones. Sin embargo, en condiciones industriales, la necesidad
de obtener mejores resultados de la genealogía está directamente relacionado a los costes
para su obtención.
En nuestro trabajo, la utilización de la segunda multiplex fue eficiente para
identificar la paternidad del animal, pero de acuerdo con los datos presentados en la tabla
4:03, vemos que solamente hemos logrado aumentar una pequeña cantidad de machos
reproductores, mientras que la cantidad de hembras sigue igual. Y, aunque la segunda
multiplex haya proporcionado información para identificar 679 individuos más, el
resultado no ha sido efectivo para la formación de nuevas familias. Por lo tanto, los
nuevos individuos asignados no aumentaron el número de familias.
Por otro lado, el principio de exclusión es válido siempre que no existan errores de
genotipado o alelos nulos, con lo cual no es realista. El porcentaje de errores de
genotipado se sitúa en torno al 2% para microsatélites (Estoup, et al., 1998; Castro, et al.,
2006) y la proporción de alelos nulos es variable, pero puede afectar a cerca del 10% de
loci microsatélites, y sus frecuencias, aunque suelen estar por debajo del 10%, pueden
alcanzar valores del 20% o superiores (Dakin y Avise, 2004). Ambas circunstancias
conducirían a falsas exclusiones o también, aunque menos frecuentemente, a inclusiones
erróneas. Además, cuantos más loci manejemos, mayor será la probabilidad de error y,
en consecuencia, se incrementarán las falsas exclusiones. Una posibilidad para evitar tal
problemática, es introducir cierta flexibilidad en el sistema permitiendo un número
determinado de incompatibilidades (Vandeputte, et al., 2006; Taggart, 2007).
Obviamente, esta solución disminuye el potencial de exclusión, por lo que es esencial la
evaluación del mínimo número de loci para asignar correctamente la mayor proporción
posible de candidatos (Slavov, et al., 2005).
Otra información importante, paralela a la asignación, es la información sobre la
contribución de los individuos en las especies de desove masal (Waldbieser y Wolters,
1999; Magoulas, 1998) que son fundamentales para la correcta gestión de reproductores.
La participación efectiva del mayor número de reproductores para la formación de la
nueva generación es importante para aumentar el número de familias formadas, una vez

63
que, una mala distribución en la contribución acarrea una reducción del tamaño efectivo
esperado de la población.
En nuestro trabajo se observó que el 20% de los individuos que contribuyen a la
descendencia fueron responsables de más del 50% de la misma. Haciendo comparaciones
entre sexos, vemos que un mayor porcentaje de machos ha participado (80% de los
reproductores disponibles en los tanques) mientras que la participación de las hembras
fue menor (67%). Esto puede agravarse en el caso de una especie que posee
hermafroditismo funcional, como es el caso de la dorada, que pueden contribuir con su
material genético en sus primeros años como macho y los siguientes como hembra. Los
cambios de sexo dentro de un plantel de reproductores, pueden causar grandes problemas
dentro de un programa de cría selectiva, caso de que no sean identificados los cambios
con precisión y a tiempo. Este es un aspecto fundamental para tomar las decisiones
relacionadas a la reorganización y gestión del plantel de reproductores.

64
5.2. OPTIMIZACIÓN DE LA SELECCIÓN CON RESTRICCIONES
El objetivo de cualquier programa de mejora es el incremento simultáneo de uno o
varios caracteres de interés comercial, ponderando cada uno de ellos por su respectivo
valor comercial en las condiciones de cultivo. Las técnicas de mejoramiento genético
aplicadas a las especies zootécnicas están basadas en el conocimiento de los parámetros
genéticos (heredabilidades y correlaciones genéticas). Es importante obtener estas
estimaciones con la mejor precisión posible, principalmente cuando son datos obtenidos
en condiciones industriales que, por motivos obvios, deben minimizar las posibles fuentes
de errores que puedan influir directamente en la interpretación de los datos.
La industria de la dorada está principalmente interesada en seleccionar rasgos que
son comunes a la mayoría de las especies de peces de cultivo comercial y que incluyen
aquellos que determinan los costes de producción y/o afectan las preferencias del
consumidor (Navarro, et al., 2009b; Chavanne, et al., 2016). Knibb, et al., (1997)
estimaron las heredabilidades de rasgos de crecimiento en dorada basadas en los datos
tomados al peso de sacrificio después de una generación de selección divergente, es decir,
una línea seleccionada para el aumento de peso y la otra seleccionada para menor peso.
Ambas líneas se compararon con una línea de control para evitar los problemas de
respuesta asimétrica a la selección en direcciones opuestas. Los valores estimados fueron
de 0,51 ± 0,13 para la línea ascendente y 0,29 ± 0,04 para la descendente. Navarro, et al.,
(2009a) estimaron las heredabilidades para caracteres de crecimiento a diferentes edades,
también bajo condiciones industriales, encontrando valores de 0,31 y 0,27 para peso y
longitud a 330 días y de 0,34 y 0,33 para peso y longitud a 509 días. En nuestro estudio,
se encontraron valores de heredabilidad más altos (alrededor de 0,40) y podrían deberse
a las diferentes condiciones de manejo o a la mayor variabilidad genética de nuestra
población. En cualquier caso, la heredabilidad de ambos estudios muestra que la dorada
es una especie con un componente genético aditivo significativo para los caracteres de
crecimiento, un hecho económico relevante para la industria. Antonello, et al., (2009) y
Chavanne, et al., (2014) también obtuvieron estimaciones de heredabilidad para el
crecimiento en el mismo rango, de 0,38 y 0,40, respectivamente. Como ya demostraron
Chavanne, et al., (2014) estas estimaciones deberían utilizarse para mejorar la
productividad de la población.

65
En el presente trabajo hemos presentado resultados detallados sobre 10 tanques de
desove masal, que proporcionan errores estándar muy bajos para las estimaciones y
permiten identificar y corregir posibles sobreestimaciones. Como, por ejemplo, cuando
analizamos los datos de heredabilidad de cada fotoperiodo por separado con los obtenidos
general, utilizando los resultados del VCE 6.0 (Tabla: 5.01),
Tabla: 5.01 Estimación de la heredabilidad de los caracteres medidos, por cada fotoperiodo
separadamente, utilizando los datos del VCE
Fotoperiodo Peso 1(g) Peso 2(g) Longitud 1(cm) Longitud 2(cm)
CD4 0,6284 ± 0,08 0,5921 ± 0,08 0,5247 ± 0,09 0,5647 ± 0,08
CD5 0,2040 ± 0,05 0,3428 ± 0,07 0,2621 ± 0,06 0,4191 ± 0,08
CD1 0,2560 ± 0,06 0,3925 ± 0,07 0,3751 ± 0,07 0,4401 ± 0,08
CD2 0,5273 ± 0,07 0,5424 ± 0,09 0,5440 ± 0,08 0,5063 ± 0,08
CD3 0,5126 ± 0,08 0,3705 ± 0,06 0,3312 ± 0,07 0,3340 ± 0,06
Todos Fotoperiodos 0,4282 ± 0,03 0,4407 ± 0,03 0,4027 ± 0,03 0,4527 ± 0,04
Vemos que cuando se realizan las estimaciones por separado, los valores del error
típico son más altos y las heredabilidades presenta mucha oscilación. Como se observa
en el fotoperiodo CD4 y CD5, que fueron formados por prácticamente los mismos grupos
de reproductores (Tabla: 3.01), con una pequeña diferencia en la formación de los ciclos,
y sometidos a las mismas condiciones del cultivo (manejo, alimentación, etc.) Mientras
que, los valores estimados con todos los fotoperiodos a la vez han presentado estimas del
error más bajas, y además de acercarse al valor promediado a de los 5 fotoperiodos. En
esta ocasión, el error puede ser corregido basándose en los valores obtenidos en los otros
fotoperiodos. Tal disparidad puede haber sido consecuencia del gran intervalo de días que
transcurrió entre la primera y segunda medida, que se refleja en los resultados de la
desviación estándar y el coeficiente de variación presentados en la Tabla: 4.01.

66
Fernandes, et al., (2017), en la publicación referente a los datos de este proyecto,
para los caracteres peso y longitud en ambas edades (350 y 500 días aproximadamente)
mostraron valores de heredabilidad cercanos a 0,40 y errores estándar cerca del 0,03.
Estos valores son más altos que los encontrados por Navarro, et al., (2009b)
probablemente debido a las condiciones más uniformes de manejo. Además de esto, la
correlación entre ambos rasgos es alta, hasta un valor de 0,91 entre el peso y la longitud
en la segunda edad, y, por lo tanto, cualquiera de estos rasgos podría ser usado como
objetivo criterio de selección. Navarro, et al., (2009b) sugirieron que la longitud podría
ser un rasgo preferible para la selección porque es más fácil de medir en las instalaciones.
Sin embargo, de acuerdo con la opinión de la empresa, el objetivo principal era la
ganancia del peso, porque es la condición que produce mayores beneficios económicos,
una vez que los peces son comercializados por su peso y no por su talla.
Por otro lado, la correlación genética encontrada entre los mismos rasgos medidos
a diferentes edades sugiere que, en este caso, sería posible seleccionar reproductores
futuros en función de las medidas en la primera edad, que podría ser de relevancia
económica para la empresa, tanto por disminuir el intervalo entre generaciones, así como
los costes de mantenimiento de reproductores.
A pesar de las diferencias biológicas existentes entre las especies, la mayoría de los
programas de mejora genética para peces comparten los objetivos de mejoramiento
relacionados con el crecimiento, la presencia de deformidades, la calidad de la carne y la
resistencia a las enfermedades. Por ejemplo, Vandeputte, et al., (2004) estimaron la
heredabilidad para algunas características relacionadas con el crecimiento en juveniles de
la carpa (Cyprinus carpio) mediante cruces factoriales completos y el uso de marcadores
moleculares. Los estimados de heredabilidad fueron de 0,33, tanto para el peso como para
la longitud y 0,37 para el factor de condición (k). Concluyeron que el mejoramiento
mediante procesos de selección puede ser exitoso en los juveniles de la carpa, pero que
sería conveniente hacer estas estimaciones a la edad comercial.
Vandeputte, et al., (2008) también utilizaron marcadores microsatélites para
establecer el parentesco en Cyprinus carpio y estimaron la heredabilidad en
características relacionadas con el crecimiento, señalando valores de 0,21 a 0,33 para la
longitud y de 0,31 a 0,44 para el peso. Los autores, concluyeron que, aunque la selección
para el crecimiento parece posible para la carpa común, este sería un proceso a largo plazo

67
antes de ser apreciable. Kocour, et al., (2007) utilizando pedigrí molecular con
microsatélites, encontraron para la misma especie en tallas de mercado valores de
heredabilidad altos (>0,5) en la longitud estándar, peso del cuerpo, porcentaje de grasas
y tamaño relativo de la cabeza, mientras que para la altura y ancho relativos del cuerpo
presentaron valores medios de heredabilidad (0,2 – 0,5).
Pante, et al., (2002) estimaron los componentes de la varianza dominante y aditiva
para el peso del cuerpo a la edad de cosecha durante 6 generaciones, en la trucha arcoíris
(Oncorhynchus mykiss), lo cual, ha permitido confirmar la presencia de una variación
genética dominante significativa para estas características en la especie. En el salmón
coho (Oncorhynchus kisutch), Neira, et al., (2006), realizaron selección para el peso a la
edad de cosecha en dos poblaciones durante 4 generaciones, encontrando valores de
heredabilidad muy parecidos (0,40 – 0,44) en ambas poblaciones.
En otros grupos de peces y con otras características la selección también ha sido
aplicada con éxito. Charo-karisa, et al., (2005) trabajando con la tilapia del Nilo
(Oreochromis niloticus) investigaron la tolerancia al frío de los juveniles de esa especie
obteniendo estimas de heredabilidad de 0,33. Saillant, et al., (2006) caracterizaron 3-6
loci microsatélites en la lubina (Dicentrarchus labrax) en un programa de selección
familiar y estimaron la heredabilidad y las interacciones genotipo x ambiente para el peso
del cuerpo utilizando exposición a diferentes temperaturas y densidades de cultivo. Los
autores demostraron un importante componente genético aditivo y la presencia de efectos
genotipo x ambiente. En el bacalao (Gadus morhua), Odegard, et al., (2010) estudiaron
la heredabilidad para la resistencia a la necrosis nerviosa viral en un programa de
selección familiar, y encontraron valores elevados, aun cuando los autores señalan que la
estructura de datos no fue la óptima para distinguir los efectos ambientales y los efectos
genéticos aditivos.
Los ejemplos son menos abundantes en crustáceos y moluscos y entre ellos
podemos mencionar, los trabajos de Benzie, et al., (1997) y Kenway, et al., (2006) en
Penaeus monodon, que obtuvieron valores bajos de heredabilidad lo que indica una gran
variación producto de una fuente control ambiental en el crecimiento de este camarón.
Hetzel, et al., (2000) encontraron una heredabilidad moderada en Penaeus japonicus. En
moluscos, He, et al., (2008) estudiaron, en la ostra perla Pinctada fucata, la selección
para el alto de la concha y encontraron un promedio de heredabilidad de 0,71 y una

68
respuesta a la selección para el crecimiento del alto de la concha marcadamente efectiva
en la segunda generación. Deng, et al., (2009) analizaron la talla de la ostra perla china
Pictada martensii y encontraron heredabilidades entre 0,22 y 0,64 indicando una
variabilidad genética considerable y postularon que la selección masal hasta la segunda
generación habría sido efectiva. Liang, et al., (2009) estimaron la heredabilidad en la
vieira japonesa Patinopecten yessoensis señalando valores de 0,47 para la respuesta a la
selección y 0,27 para la heredabilidad realizada, sugiriendo que la selección masal para
crecimiento en esta especie será efectiva.
Por lo tanto, identificar y elegir correctamente los individuos formadores de la
siguiente generación no es una tarea tan sencilla, una vez que se deberá mantener un
equilibrio entre el potencial genético y los niveles de endogamia, y en la realización de
esa tarea las opciones pueden ser muy específica y limitadas. El sistema BLUP se ha
convertido en el método de referencia para la predicción del valor genético de los
animales porque asegura una evaluación precisa del valor genético pero su uso debe ser
cuidadoso ya que provoca una reducción rápida del tamaño efectivo de la población. Por
eso, se necesita controlar nivel de variabilidad en las poblaciones bajo la selección
mediante el uso equilibrado y dirigido en la elección de los reproductores, evitando
además los apareamientos consanguíneos. En este sentido la optimización de la respuesta
con una restricción en la consanguinidad podría garantizar el éxito de la selección a lo
largo de los ciclos.
La optimización empleada en este trabajo ha demostrado eficiencia a la hora de
seleccionar individuos con superioridades frente a los evaluados. El diferencial de
selección global para el peso en la segunda medición fue de 41,6g (10,5%) en la situación
I, de 92,1g (23,2%) en la situación II, de 103,3g (26%) en la situación III y de 105,4 g
(26,6%) en la situación IV. (Figura: 5.03)

69
Figura: 5.03 Diferencial de selección (DS) de cada fotoperiodo (expresado en
porcentaje de la población inicial), en las cuatro situaciones de
parentesco global propuesto.
Sin embargo, de todas las variables que un criador de animal puede manipular, el
tamaño de la población es el que tiene más amplia gama de consecuencias. Eso ocurre
porque cada población va presentar un límite sobre el máximo valor genético alcanzable
cuando es sometida a un proceso de restricción de consanguinidad. Cuando hacemos
simulaciones de diferentes intensidades de selección (%) aplicadas en las poblaciones de
los cinco fotoperiodos, lo que se observa es que, la curva de tendencia entre los valores
genéticos estimados (estimated breeding value – ebv) y el promedio del parentesco
alcanza un valor asintótico cada vez en niveles de parentesco más bajos a medida que
vamos aumentando el porcentaje de animales seleccionados. Por lo tanto, de acuerdo con
los gráficos, en ninguno de los fotoperiodos sería ventajoso sobrepasar el 8% de
individuos seleccionados, cantidad a partir de la cual la curva empieza a linealizar
precozmente. Sobre todo, en los fotoperiodos CD1 y CD3, donde ya se nota este patrón
de linealización incluso con el 16% de los individuos seleccionados. (Figura: 5.04)
0,000
0,125
0,250
0,500
0
5
10
15
20
25
30
35
CD4CD5
CD1CD2
CD3
Par
en
tesc
o G
lob
al
DS
(%)
Fotoperiodo

70
Figura: 5.04 La optimización con restricciones para diferentes cantidades (%) de individuos seleccionados

71
Nuestros resultados demuestran que el método de selección no sólo es apropiado,
sino también muy eficiente para mantener la varianza genética en los nuevos
reproductores, garantizando tanto la mínima consanguinidad como la mínima pérdida de
diversidad genética. En el gráfico, se observa que el 4% sería el que podría ofrecer una
mejor respuesta sin aumentar demasiadamente el parentesco de la población. En nuestro
trabajo, fue utilizado una tasa de 5% y con la restricción de la situación II (f = 0,125) el
diferencial de selección ha sido superior a los 20%. Estos valores están de acuerdo con
el trabajo de Toro y Pérez-Enciso (1990), donde realizaron simulaciones en un ejemplo
sencillo de una población conformada por 8 machos y 8 hembras seleccionadas de 32
animales evaluados en cada sexo practicando cinco generaciones de selección se
concluyó que utilizar el 5% por generación es una buena alternativa para que se garanticen
la endogamia por debajo de un nivel aceptable sin pérdidas significativas en la respuesta.
Por estos motivos, las tasas de renovación y la reorganización de los reproductores
es uno de los criterios más importantes para establecer el ritmo al que la mejora genética
puede avanzar dentro de la población. En dorada, la principal limitación es el
mantenimiento del cociente sexual en el plantel de reproductores, una vez que el cambio
del sexo complica el registro de los datos, puesto que un individuo que había sido
catalogado como un macho en su último registro, puede haber cambiado hacia hembra en
el año siguiente. Por lo tanto, las hembras son el sexo más importante para el proceso de
gestión, pues es el sexo que se mantiene dentro de los criaderos ya que los machos se
convierten en hembras, y su conservación sólo podrá ser a través de la criopreservación
de espermatozoides, una técnica que se ha establecido con éxito en dorada (Suquet, et al.,
2000), mientras que la conservación de óvulos femenina aún no es un logro real (Cerda,
2002).

72
5.3. CONSIDERACIONES FINALES
Según el informe de las Naciones Unidas (FAO, 2016), se prevé que la población
mundial actual, que es de aproximadamente 7.400 millones de personas, alcance los 8.500
millones en 2030 y los 9.700 millones en 2050 y que ese crecimiento se produzca,
principalmente, en países en desarrollo. Por lo tanto, garantizar la seguridad alimentaria
y nutricional suficiente para esa población en crecimiento es un desafío enorme. Frente a
esa creciente demanda de alimentos y el problema de la sobrepesca de los stocks en la
naturaleza, la FAO considera la acuicultura como la única alternativa posible para suplir
esa demanda
Para que podamos aumentar los índices productivos, es fundamental que se
intensifiquen los estudios sobre las especies cultivadas a fin de obtener mejores
resultados, y en estas condiciones, el establecimiento de programas de mejora genética
surge como una importante herramienta con gran potencial. Aún falta mucho para que la
acuicultura pueda alcanzar el nivel en que se encuentra la ganadería, donde la producción
está basada casi exclusivamente en el cultivo de poblaciones mejoradas genéticamente,
pero no cabe duda de que el aumento consistente en la demanda por el producto acuícola
motivará a los productores a mejorar la eficiencia de sus productos por medio de las
técnicas de la mejora genética.
Sin embargo, uno de los principales problemas para poner en marcha programas de
mejora genética en especies acuáticas está en la dificultad en mantener los registros de
identificación individual y controlar las informaciones de pedigrí, principalmente porque
son animales que poseen fecundación y desarrollo externos, alta prolificidad y reducidos
tamaños en las fases iniciales de vida, además de tener poca o casi ninguna diferenciación
fenotípica entre sexos. Por lo tanto, los altos costes para obtener estas informaciones
hacen muchas veces inviable la ejecución de proyectos de investigación y su aplicabilidad
en el sector productivo. En un estudio analizando los costes-beneficios de los programas
de cría selectiva en acuicultura, se estimaron en € 149.556/año para los programas con
núcleos y € 127.845/año para los programas sin núcleos de multiplicadores (Janssen, et
al., 2018).

73
Con lo cual, la implantación de programas de mejora genética en acuicultura queda
restringida a las grandes empresas, que vienen cada vez más apostando en mejorar su
material genético, produciendo animales con calidad muy superior a los demás y
diferenciando su producto. A largo plazo, las empresas que no poseen capacidad de
inversión financiera o que no desean utilizar los recursos genéticos para mejorar su
productividad, irán perdiendo cada vez más competitividad en el mercado, y
consecuentemente, facilitando el monopolio de las grandes empresas en el sector, como
ya ha acontecido con otros ejemplos del sector alimenticio.
Por eso, el desarrollo de marcadores, físicos y moleculares, eficientes y de bajo
costo podrían proporcionar las condiciones necesarias para que sean ampliamente
utilizadas en un número mayor de programas de mejora genética, y consecuentemente
posibilitando que los pequeños y medios productores puedan gestionar mejor sus
criaderos en el sentido de aprovechar sus recursos genéticos, principalmente para la su
difusión en los países en desarrollos,
Para eso, es necesario mayores inversiones y apoyo del gobierno hacia el sector
investigador y productivo, para que podamos caminar hacia una acuicultura más
competitiva y rentable. La utilización de multiplexes secuenciales supone un importante
ahorro en los gastos de análisis moleculares. Para obtener buenos resultados, es
importante utilizar microsatélites específicos y preferentemente que tengan una amplitud
alélica suficiente para caracterizar y distinguir con seguridad las repeticiones de cada uno
de ellos. Los SNPs se han convertido en una alternativa eficiente al uso de los
microsatélites.
Los problemas generados por la endogamia son más serios en los cultivos de peces
de lo que son en el ganado doméstico. Debido a su alta fecundidad es muy probable que
un número muy pequeño de individuos puede hacer una gran contribución a la
composición genética de la generación siguiente principalmente en los cultivos de
especies con características de desove masal. No cabe duda de que la tasa de endogamia
puede aumentar rápidamente, aunque tengamos un alto número de reproductores. Por ello
es importante que la mayor cantidad de reproductores esté contribuyendo efectivamente
para la formación de las nuevas generaciones, preferiblemente en proporciones iguales.
En nuestro estudio, más del 50% de la descendencia identificada provenía de apenas el
20% de los reproductores. En este caso, es aconsejable proponer correcciones en los ciclos

74
futuros, en el sentido de buscar una participación más homogénea del plantel, caso
contrario, resultará en pérdida de la variabilidad genética que inducirá a una disminución
de la varianza aditiva para el carácter objeto del mejoramiento genético (Gjerde, et al.,
1983).
En una selección ineficiente y desequilibrada, el aumento de la endogamia podría
llegar a niveles que causen problemas en la tasa de crecimiento y otros caracteres
productivos que podrán disminuir significativamente (Kincaid, 1976, 1983; Gjerde, et al.,
1983; Sbordoni, et al., 1986; Su, et al., 1996).
En resumen, los programas de cría selectiva deben ser diseñados para maximizar el
progreso genético, evitando la pérdida de la diversidad genética. Esta preocupación debe
de ser mayor en el diseño y ejecución de programas de cría selectiva en organismos
acuáticos, que en los programas equivalentes en animales domésticos. Por lo tanto,
utilizar la optimización de la selección aplicando restricciones a la tasa de endogamia es
muy importante en los programas de cría selectiva en peces. Sin embargo, la relativa falta
de recursos (instalaciones, económicos y humanos) es uno de los desafíos que enfrenta el
futuro desarrollo de la genética acuícola, dado que sus resultados son a largo plazo y que
pueden presentan unas características reproductivas peculiares (fecundación, ciclo
generación, desove masal, hermafroditismo, etc.). Por ello pueden pasar algunos años
hasta que logre generar los beneficios financieros que compensen la inversión monetaria
aplicada.
En los cultivos de los organismos acuáticos, es posible encontrar una gran variedad
de diferentes procedimientos de gestión y manejo productivo, incluso cuando se trata del
cultivo de una misma especie dentro de la misma región como consecuencia de una
variedad de efectos ambientales. Por lo tanto, creemos que la popularización de las
técnicas de la mejora genética en la acuicultura posibilitará la ejecución de varios
programas independientes, que consecuentemente, estará beneficiando todo el sector
científico y productivo, generando una mayor cantidad de datos y estimaciones cada vez
más precisas, obteniendo resultados más efectivos con menores costes. De este modo, el
mercado se podrá beneficiar de los procedimientos de gestión óptimos y de la difusión de
las acciones de la mejora.

75
6.
CONCLUSIONES

76
6. CONCLUSIONES
• Con el resultado final de 86,84% de asignación obtenidas en este trabajo, podemos
concluir que los microsatélites son herramientas de gran importancia para la
estimación del parentesco molecular en organismos acuáticos. El potencial de
resolución de estos marcadores moleculares está directamente relacionado al
polimorfismo, o sea que cuantos más alelos más difíciles es que los descendientes
tengan genotipos compatibles, y por lo tanto más fácil la asignación.
• En condiciones industriales, es importante equilibrar los costes con los beneficios
extra que se obtienen de la implantación de diferentes tecnologías. En este sentido,
la utilización de multiplex secuenciales proporcionó un ahorro del 43% en los
costos con análisis moleculares. La adición de nuevos marcadores, no
necesariamente significará un aumento en el número de familias formadas, ya que
tiene mayor influencia el hecho de que no todos los padres se reproducen.
• En general, los valores obtenidos en las estimas de heredabilidad encontradas
fueron de h2 >0,43, y de rA >0,85 para las correlaciones genéticas y de rP >0,72 para
las correlaciones fenotípicas. Estos valores indican que la implementación de
programes de mejora genética para un carácter simple tiene gran potencial en la
producción de dorada, influyendo en varias características del crecimiento.
• La utilización de la metodología de Contribuciones óptimas (OC, por sus siglas en
inglés) permitió obtener diferenciales de selección elevados, pero controlando los
niveles de pérdida de variabilidad. Esto es especialmente importante en acuicultura,
donde una pequeña fracción de reproductores pueden contribuir masivamente en la
formación de la descendencia.

77
7. REFERENCIAS BIBLIOGRÁFICAS
ANTONELLO, J., MASSAULT, C., FRANCH, R., HALEY, C., PELLIZZARI,
C., BOVO, G., PATARNELLO, T., KONING, D., BARGELLONI, L. (2009).
“Estimates of heritability and genetic correlation for body length and resistance to
fish pasteurellosis in the gilthead sea bream (Sparus aurata L.)”. Aquaculture;
298:29-35.
APROMAR. (2018). “Informes anuales: La acuicultura en España”. 93 pg.
(http://www.apromar.es/content/informes-anuales). [26 de febrero de 2018].
BALON, E.K. (1995). “Origin and domestication of the wild carp, Cyprinus carpio:
from Roman gourmets to the swimming flowers”. Aquaculture; 129:3-48.
BATARGIAS, C., DERMITZAKIS, E., MAGOULAS, A., ZOUROS, E. (1999).
“Characterization of six polymorphic microsatellite markers in gilthead seabream,
Sparus aurata (Linnaeus 1758)”. Molecular Ecology Notes; 8:897-899.
BENZIE, K., KENWAY, M., TROTT, L. (1997). “Estimates for the heritability of
size in juveniles Penaeus monodon prawns for hals-sib matings”. Aquaculture;
152:49-53.
BRIÑEZ, R.B., CARABALLO, O.X., SALAZAR, V.M. (2011). “Genetic
diversity of six populations of red hybrid tilapia, using microsatellites genetic
markers.” Revista MVZ Córdoba; 16(2):2491-2498.
BROWN, C.R., TSALAVOUTA, M., TERZOGLOU, V., MAGOULAS, A.,
MCANDREW, B.J. (2005b). “Additional microsatellites for Sparus aurata and
cross-species amplification within the Sparidae family”. Molecular Ecology
Notes; 5:605-607.
BROWN, C.R., WOOLLIAMS, J.A., MCANDREW, B.J., (2005a). “Factors
influencing effective population size in commercial populations of gilthead
seabream, Sparus aurata”. Aquaculture; 247:219-225.

78
CASTRO, J., PINO, A., HERMIDA, M., BOUZA, C., CHAVARRÍAS, D.,
MERINO, P., SÁNCHEZ, L., MARTÍNEZ, P. (2007). “A microsatellite marker
tool for parentage assessment in gilthead seabream (Sparus aurata)”.
Aquaculture; 272:210-216.
CASTRO, J., PINO, A., HERMIDA, M., BOUZA, C., RIAZA, A., FERREIRO,
I., SÁNCHEZ, L., MARTÍNEZ, P. (2006). “A microsatellite marker tool for
parentage analysis in Senegal sole (Solea senegalensis): Genotyping errors, null
alleles and conformance to theoretical assumptions”. Aquaculture; 261:1194-
1203.
CATAUDELLA, S., BRONZI, P. (2001). “Acquacoltura responsabile”. Unimar-
Uniprom, Rome, Italy: 683 pp.
CERDA, J. (2002). “Mecanismos fisiológicos durante la hidratación del huevo de
teleósteos: hacia el desarrollo de nuevos métodos de criopreservación”. Boletín
del Instituto Español de Oceanografía; 18:145-152.
CHAKRABORTY, R., MEAGHER, T.R., SMOUSE, P.E. (1988). “Parentage
analysis with genetic markers in natural populations. I. The expected proportion
of offspring with unambiguous paternity”. Genetics; 118:527-536.
CHAKRABORTY, R., MOREAU, L., DEKKERS, J.C.M. (2002). “A method to
optimize selection on multiple identified quantitative trait loci”. Genetics
Selection Evolution; 34:145–170.
CHARO-KARISA, H., REZK, M., BOVENHUIS, H., KOMEN, H. (2005).
“Heritability of cold tolerance in Nile tilapia Oreochromis niloticus juveniles”.
Aquaculture; 249:115-123.
CHAVANNE, H., JANSSEN, K., HOFHERR, J., AQUATRACE
CONSORTIUM, CONTINI, F., HAFFRAY, P., KOMEN, H., NIELSEN,
E.E., BARGELLONI, L. (2016). “A comprehensive survey on selective
breeding programs and seed market in the European aquaculture fish industry”.
Aquaculture International; 24:1287-1307.

79
CHAVANNE, H., PARATI, K., CAMBULI, C., CAPOFERRI, R., JIMENEZ,
C.A., GALLI, A. (2014). “Microsatellites markers to depict the reproductive and
genetic patterns of farmed gilthead sea bream (Sparus aurata): illustration by a
case study on mass spawning”. Aquaculture Research; 45:577-590.
DAKIN, E.E., AVISE, Y.J.C. (2004). “Microsatellite null alleles in parentage
analysis”. Heredity; 93:504-509.
DENG, Y., FU, S., DU, X., WANG, Q. (2009). “Realized heritability and genetic
estimates of larval shell length in the chineses pearl oyster Pinctada martensii as
three different salinities”. North American Journal Aquaculture; 71:302-306.
DONG, L., XIAO, S., WANG, Q., WANG, Z. (2016). “Comparative analysis of
the GBLUP, emBayesB, and GWAS algorithms to predict genetic values in large
yellow croaker (Larimichthys crocea).” BMC Genomics; 17:460.
DUNHAM, R.A. (2011). “Aquaculture and Fisheries Biotechnology and Genetics,
2nd edition”. CAB International, Wallingford, UK. 504 pg.
DUNHAM, R.A., MAJUMDAR, K., HALLERMAN, E., BARTLEY, D., MAIR,
G., HULATA, G., LIU, Z., PONGTHANA, N., BAKOS, J., PENMAN, D.,
GUPTA, M., ROTHLISBERG, P. AND HOERSTGEN-SCHWARK, G.
(2001). “Review of the status of aquaculture genetics”. In: Subasinghe, Technical
Proceedings of the Conference on Aquaculture in the Third Millennium, Bangkok,
Thailand, 20–25 February 2000. NACA, Bangkok and FAO, Rome, pp. 129-157.
DUPONT-NIVET, M., VANDEPUTTE, M., VERGNET, A., MERDY, O.,
HAFFRAY, P., CHAVANNE, H., CHATAIN, B. (2008). “Heritabilities and
GxE interactions for growth in the European Sea bass (Dicentrarchus labrax L.)
using a marker-based pedigree”. Aquaculture; 275:81-87.
ESTOUP, A., GHARBI, K., SANCRISTOBAL, M., CHEVALET, C.,
HAFFRAY, P., GUYOMARD, R. (1998). “Parentage assignment using
microsatellites in turbot (Scophthalmus maximus) and rainbow trout
(Oncorhynchus mykiss) hatchery populations”. Canadian Journal of Fisheries and
Aquatic Sciences; 55:715-725.

80
FAO (2005). “Programa de información de especies acuáticas”. Sparus aurata.
Programa de información de especies acuáticas. Texto de Colloca, F.; Cerasi, S.
In: Departamento de Pesca y Acuicultura de la FAO [en línea]. Roma.
Actualizado 8 February 2005. [Citado 22 October 2013].
(http://www.fao.org/fishery/culturedspecies/Sparus_aurata/es#tcNA002B).
FAO (2009). “The State of World Fisheries and Aquaculture 2008”. Food and
Agriculture Organization of the United Nations, Rome.
FAO (2016). “El estado mundial de la pesca y la acuicultura”. Contribución a la
seguridad alimentaria y la nutrición para todos. Roma. 224 pp.
FERNANDES, T., HERLIN, M., BELLUGA, M.D.L., BALLÓN G.,
MARTINEZ P., TORO MIGUEL, FERNÁNDEZ J. (2017). “Estimation of
genetic parameters for growth traits in a hatchery population of gilthead sea bream
(Sparus aurata L.)”. Aquaculture International; 25: 499-514.
FERNÁNDEZ, J., TORO, M. A. (2006). “A new method to estimate relatedness
from molecular markers”. Molecular Ecology; 15:1657-1667.
FERNÁNDEZ, J., TORO, M.A. (1999). “The use of mathematical programming
to control inbreeding in selection schemes”. Journal of Animal Breeding and
Genetics; 116:447-466.
FERNÁNDEZ, J., VEGA, L., TORO, M.A. (2004). “Control de la
consanguinidad en programas de selección en acuicultura”. ITEA. Producción
animal; 100(3): 188-191.
FISH BASE (2018). “Computer Generated Native Distribution Map for Sparus
aurata (Gilthead seabream)”. www.aquamaps.org, version of Aug. 2013. Web.
Accessed 22 Oct. 2018. (http://www.fishbase.org/summary/Sparus-aurata.html).
FISHER, R.A. (1918). “The causes of human variability”. Eugen. Rev; 10:213-220.
FRANCH, R., LOURO, B., TSALAVOUTA, M., CHATZIPLIS, D.,
TSIGENOPOULOS, COSTAS S.; SARROPOULOU, E., ANTONELLO, J.,
MAGOULAS, A., MYLONAS, C., BABBUCCI, M., PATARNELLO, T.,

81
POWER, D.M., KOTOULAS, G., BARGELLONI, L. (2006). “A Genetic
Linkage Map of the Hermaphrodite Teleost Fish Sparus aurata L.” Genetic
Society of America; 174:851-861.
GJERDE, B., GUNNES, K., GJEDREM, T. (1983). “Effect of inbreeding on
survival and growth in rainbow trout.” Aquaculture; 34:327-332.
GRUNDY, B., VILLANUEVA, B., WOOLLIAMS, J.A. (2000). “Dynamic
selection for maximizing response with constrained inbreeding in schemes with
overlapping generations”. Animal Scienc Journal; 70:373–382.
HE, M., GUAN, Y., YUAN, P., ZHANG, H. (2008). “Realized heritability and
response to selection for shell height int the pearl oyster Pinctada fucata
(Gould)”. Aquaculture Resource; 39:801-805.
HERBINGER, C.M., O’REILLY, P.T., DOYLE, R.W., WRIGHT, J.M.,
O’FLYNN, F. (1999). “Early growth performance of Atlantic salmon full-sib
families reared in single family tanks versus in mixed family tanks”. Aquaculture;
173:105-116.
HETZEL, D., CROCOS, P., DAVIS, G., MOORE, S., PRESTON, N. (2000).
“Response to selection and heritabiliy for growth in the Kirima prawn, Penaeus
japonicus”. Aquaculture; 181:215-223.
JANSSEN, K., SAATKAMP, H., KOMEN, H. (2018). “Cost-benefit analysis of
aquaculture breeding programs”. Genetic Selection Evolution; 50:2.
JONES, A.G., ARDREN, W.R. (2003). “Methods of parentage analysis in natural
populations”. Molecular Ecology; 12:2511-2523.
KENWAY, M., MACBETH, M., SALMON, C., MCPHEE, K., BENZIE, K.,
WILSON, K., KNIBB, W. (2006). “Heritability and genetic correlations of
growth and survival in black tiger prawn Penaeus monodon reared in tanks”.
Journal Aquaculture; 259:138-145.
KINCAID, H.L. (1976). “Effects of inbreeding on rainbow trout populations.”
Transactions of the American Fisheries Society; 105:273-280.

82
KINCAID, H.L. (1983). “Inbreeding in fish populations used for aquaculture”.
Aquaculture; 33:215–227.
KINGHORN, B.P. (2010). “An algorithm for efficient constrained mate selection”.
Genetics Selection Evolution; 43:4.
KNIBB, W., GORSHKOVA, G., GORSHKOV, S. (1997). “Selection for growth
in the gilthead seabream, Sparus aurata L”. Israeli Journal of Aquaculture –
Barmidgeh; 49:57-66.
KOCOUR, M., MAUGER, S., RODINA, M., GELA, D., LINHART, O.,
VANDEPUTTE, M. (2007). “Heritability estimates for processing and quality
traits in common carp (Cyprinus carpio) using a molecular pedigree”.
Aquaculture; 270:43-50.
KOVAČ, M., GROENEVELD, E., GARCÍA-CORTÉS, L.A., (2002). “VCE-5:
A package for the optimization of dispersion parameters”. Proceedings of the 7th
World Congress on Genetics Applied to Livestock Production, 20-23 August
2002, Montpellier, France. (ftp://ftp.tzv.fal.de/pub/vce6/).
LAGHARI, M.Y., LASHARI, P., ZHANG, Y., SUN, X. (2014). “Identification of
Quantitative Trait Loci (QTLs) in Aquaculture Species”. Reviews in Fisheries
Science & Aquaculture; 22(3):221-238.
LAUNEY, S., KRIEG, F., HAFFRAY, P., BRUANT, J.S., VANNIERS, A.,
GUYOMARD, R. (2003). “Twelve new microsatellite markers for gilthead
seabream (Sparus aurata): Characterization, polymorphism and linkage”.
Molecular Ecology; 3:457-459.
LEGARRA, A., VARONA, L., DE MATURANA E.L., (2008). “Program TM.”
http://snp.toulouse.inra.fr/*alegarra.
LI, L., LIN, H., TANG, W., LIU, D., BAO, B., YANG, J. (2017). “Population
genetic structure in wild and aquaculture populations of Hemibarbus maculates
inferred from microsatellites markers”. Aquaculture and Fisheries; 2:78-83.

83
LI, Y., VAN DER WERF, H.J., KINGHORN, P.B. (2006). “Optimization of a
crossing system using mate selection”. Genetics Selection Evolution; 38:147–165.
LI, Y., VAN DER WERF, H.J., KINGHORN, P.B. (2008). “Optimal utilization
of non-additive quantitative trait locus in animal breeding programs”. Journal of
Animal Breeding and Genetics; 125:342–350.
LIANG, J., ZHNAG, G., ZHENG, H. (2009). “Divergent selection and realized
heritability for growth in the Japanese scallop Patinopecten yessoensis Jay”.
Aquaculture Resource; 41:1315-1321.
LOPES, P.S. (2005). “Teoria do melhoramento animal”. – Belo Horizonte:
FEPMVZ-Editora, 118p.: Inclui bibliografia ISBN: 85-87144-21-9.
MAGOULAS, A. (1998). “Application of molecular markers to aquaculture and
broodstock management with special emphasis on microsatellite DNA”. In,
Bartley, D and Basurco, B (Eds) Genetics and Breeding of Mediterranean
Aquaculture Species. Proceedings of the seminar of the CIHEAM network on
technology of aquaculture in the Mediterranean (TECAM), April 1997, CIHEAM,
Zaragoza, Spain. pp153-168, Cahiers Options Mediterrennes, vol 34.
MARSHALL, T.C., SLATE, J., KRUUK, E.B., PEMBERTON, J.M. (1998).
“Statistical confidence for likelihood-based paternity inference in natural
populations”. Molecular Ecology; 7:639-655.
MARTÍNEZ, P., FERNÁNDEZ, J. (2008). “Estimating parentage relationships
using molecular markers in aquaculture.” Aquaculture Research Trends; 10:59-
112.
MARTÍNEZ, P.P., FIGUERAS, A.H. (2007). “Libro Genética y Genómica en
Acuicultura”.: Observatorio Español de Acuicultura. Editorial CSIC- OESA,
Madrid. ISBN 978-84-00-08553-7. 890p
MEUWISSEN, T.H.E. (1997). “Maximizing the response of selection with
predefined rate of inbreeding.” Journal of Animal Science; 75:934–940.

84
MEUWISSEN, T.H.E., SONESSON, A.K. (1998). “Maximizing the response of
selection with a predefined rate of inbreeding: overlapping generations.” Journal
of Animal Science; 76:2575–2583.
MISZTAL, I., TSURUTA, S., STRABEL, T., AUVRAY, B., DRUET, T., LEE,
D.H. (2002). “BLUPF90 Andre lated programs (BGF90).” In: Proceedings of the
7th world congress on genetics applied to livestock production, Montpellier,
France, 20–23 Aug 2002.
MORETTI, A., PEDINI FERNANDEZ-CRIADO, M., CITTOLIN, G.,
GUIDASTRI, R. (1999). “Manual on Hatchery Production of Seabass and
Gilthead Seabream”, Volume 1. FAO, Rome 194 p.
NAVARRO, A., BADILLA, R., ZAMORANO, M.J., PASAMONTES, V.,
HILDEBRANDT, S., SÁNCHEZ, J.J., AFONSO, J.M. (2008). “Development
of two new microsatellite multiplex PCRs for three sparid species: gilthead
seabream (Sparus auratus L.), red porgy (Pagrus pagrus L.) and red banded
seabream (P. auriga, Valenciennes, 1843) and their application to paternity
studies”. Aquaculture; 285:30–37.
NAVARRO, A., ZAMORANO, M.J., HILDEBRANDT, S., GINÉS, R.,
AGUILERA, C., AFONSO, J.M. (2009b). “Estimates of heritabilities and
genetic correlations for growth and carcass traits in gilthead seabream (Sparus
aurata L.), under industrial conditions”. Aquaculture; 289:225-230.
NAVARRO, A., ZAMORANO, M.J., HILDEBRANDT, S., GINÉS, R.,
AGUILERA, C., AFONSO, J.M. (2009a). “Estimates of heritabilities and
genetic correlations for body composition traits and G×E interactions, in gilthead
seabream (Sparus auratus L.)”. Aquaculture; 295:183-187.
NEFF, B.D., FU, P., GROSS, M.R. (2000). “Microsatellite multiplexing in fish”.
Transactions of the American Fisheries; 129:584-593.
NEIRA, R., DÍAZ, N., GALL, G., GALLARDO, J., LLHORENTE, J., ALERT,
A. (2006). “Genetic improvement in coho salmon (Oncorhynchus kisutch):
Selection response for early spawning date”. Aquaculture; 257:1-8.

85
NEJATI-JAVAREMI, A., SMITH, C., GIBSON, J.P. (1997). “Effect of total
allelic relationship on accuracy of evaluation and response to selection”. Journal
of Animal Science; 75:1738-1745.
NEVES, H. H., CARVALHEIRO, R., QUEIROZ, S.A. (2012). “A comparison of
statistical methods for genomic selection in a mice population”. BMC Genetic;
13:100.
NISHIO, M., KAHI, A.K., HIROOKA, H. (2010). “Optimization of mate
selection based on genotypic information with overlapping generations”. Journal
of Animal Breeding and Genetics; 127:34–41.
ODEGARD, J., SOMMER, A., PRAEBEL, A. (2010). “Heritability of resistance
to viral nervous necrosis in Atlantic cod (Gadus morhua)”. Aquaculture; 200:59-
64.
ORTEGA, A. (2008). “Cultivo de la Dorada (Sparus aurata)”. Colección
Cuadernos de Acuicultura, nº 1. FOESA, CSIC y MARM. Madrid, España. 44
páginas.
PANTE, M.A., GJERDE, B.J., MCMILLAN, I., MISZTAL, I. (2002).
“Estimation of additive and dominance genetic variances for body weight at
harbes in rainbow trout Oncorhynchys mykiss”. Aquaculture; 204:383-392.
PARREÑO-MARCHANTE, A., ALVAREZ-MELCON, A., TREBAR, M.,
FILIPPIN, P. (2014). “Advanced traceability system in aquaculture supply
chain.” Journal of Food Engineering; 122:99-109.
SAILLANT, E., DUPONT-NIVET, M., HAFFRAY, P., CHATAIN, B. (2006).
“Estimates of heritability and genotype-enviromental interactions for body weight
in sea bass (Dicentrarchus labrax L.) raised under communal rearing conditions”.
Aquaculture; 254:139-147.
SANCRISTOBAL, M., CHEVALET, Y.C. (1997). “Error tolerant parent
identification from a finite set of individuals”. Genetics Research Cambridge;
70:53-62.

86
SARROPOULOU, E., FRANCH, R., LOURO, B., POWER, D. M.,
BARGELLONI, L., MAGOULAS, A., SENGER, F., TSALAVOUTA, M.,
PATARNELLO, T., GALIBERT, F., KOTOULAS, G., AND GEISLER, R.
(2007). “A gene-based radiation hybrid map of the gilthead sea bream Sparus
aurata refines and exploits conserved synteny with Tetraodonnigro viridis”. BMC
Genomics; 8:44.
SBORDONI, V., DE MATTHAEIS, M., COBOLLI SBORDONI, M., LA
ROSA, G., MATTOCCIA, M. (1986). “Bottleneck effects and the depression of
genetic variability in hatchery stocks of Penaeus japonicus (Crustacea,
Decapoda)”. Aquaculture; 57:239–251.
SHEPHERD, R.K., MEUWISSEN, T.H.E., WOOLIAMS, J.A. (2010).
“Genomic selection and complex trait prediction using a fast EM algorithm
applied to genome-wide markers”. BMC Bioinfomatics; 11:529.
SLAVOV, G.T., HOWE, G.T., GYAOUROVA, A.V., BIRKES, D.S., ADAMS,
W.T. (2005). “Estimating pollen flow using SSR markers and paternity exclusion:
accounting for mistyping”. Molecular Ecology; 4:3109-3121.
SONESSON, A.K., MEUWISSEN, T.H.E. (2009) “Testing strategies for genomic
selection in aquaculture breeding programs”. Genetic Selection Evolution; 41:37.
SONESSON, A.K., WOOLLIAMS, J.A., MEUWISSEN, T.H.E. (2010).
“Maximizing genetic gain whilst controlling rates of genomic inbreeding using
genomic optimum contribution selection”. Proc. 9th World Cong. Genet. Appl.
Livest. Leipzig, Germany.
STOCKLEY, B.M., ROGERS, A.D., IYENGAR, A., MENEZES, G., SANTOS,
R., LONG, A. (2000). “Ten microsatellite loci isolated and developed for the
black spot seabream, Pagellus bogaraveo (Brunnich, 1768)”. Molecular Ecology;
9:993-1011.
SU, G.S., LILJEDAHL, L.E., GALL, G.A.E. (1996). “Effects of inbreeding on
growth and reproduction traits in rainbow trout (Oncorhynchus mykiss)”.
Aquaculture; 142:139-148.

87
SUQUET, M., DREANNO, C., FAUVEL, C., COSSON, J., BILLARD, R.
(2000). “Cryopreservation of sperm in marine fish”. Aquaculture Research; 31
(3):231–243.
TAGGART, J.B. (2007). “FAP: an exclusion-based parental assignment program
with enhanced predictive functions”. Molecular Ecology Notes; 7:412-415.
TANG, G., LI, X. (2006). “Optimizing selection on multiple identified quantitative
trait loci in population with overlapping generations”. Journal of Genetics and
Genomics; 33:429–440
TORO, M.A., LÓPEZ-FANJUL, C. (2007). “Diseño de programas de mejora
genética en acuicultura”. En Martínez, P. y Figueras, A. (coord.) Genética y
genómica en acuicultura, págs. 185-211, Editorial C. S. I. C., Madrid.
TORO, M.A., PÉREZ- ENCISO, M. (1990). “Optimization of response under
restricted inbreeding”. Genetic Selection Evolution; 22:93-107.
VANDEPUTTE, M., KOCOUR, M., MAUGER, S., DUPONT-NIVET, M., DE
GUERRY, D., RODINA, M., GELAD, D., VALLOD, D., CHEVASSUS, B.,
LINHART, O. (2004). “Heritability estimates for growth-relates traits using
microsatellite parentage assignment in juvenile common carp (Cyprinus carpio)”.
Aquaculture; 235(4):223-236.
VANDEPUTTE, M., KOCOUR, M., MAUGER, S., RODINA, M., LAUNAY,
A. (2008). “Genetic variation for growth and two summers of age in the common
carp Cyprinus carpio: heritability estimates and response to selection”.
Aquaculture; 277(2):7-13.
VANDEPUTTE, M., MAUGER, S., DUPONT-NIVET, M. (2006). “An
evaluation of allowing for mismatches as a way to manage genotyping errors in
parentage assignment by exclusion”. Molecular Ecology Notes; 6:265-267.
VILLANUEVA, B., PONG-WONG, R., WOOLLIAMS, J. A., AVENDAÑO, S.
(2004). “Managing genetic resources in selected and conserved populations”. In:
Farm Animal Genetic Resources; 113:132.

88
VILLANUEVA, B., PONG-WONG, R., WOOLLLIAMS, J.A. (2002). “Marker
assisted selection with optimized contribution of candidates to selection”.
Genetics Selection Evolution; 34:679–703.
WALDBIESER, G.C., WOLTERS, W.R. (1999). “Application of polymorphic
microsatellite loci in a channel catfish, Ictalurus punctatus, breeding program”.
Journal of the World Aquaculture Society; 30:256-262.
WESMAJERVI, M.S., WESTGAARD, J.I., DELGHANDI, M. (2006).
“Evaluation of a novel pentaplex microsatellite marker system for paternity
studies in Atlantic cod (Gadus morhua L.)”. Aquaculture Resource; 37:1195-
1201.
WILSON, A.J., FERGUSON, M.M. (2002). “Molecular pedigree analysis in
natural populations of fishes: approaches, applications, and practical
considerations”. Canadian Journal of Fisheries and Aquatic Sciences; 59:1696-
1707.
YUE, G.H. (2014). “Recent advances of genome mapping and marker-assisted
selection in aquaculture”. Fish and Fisheries; 15:376–96.
ZENGER, K.R., KHATKAR, M.S., JONES, D.B., KHALILISAMANI, N.,
JERRY, D.R., RAADSMA, H.W. (2019). “Genomic selection in aquaculture:
Application, limitations and opportunities with special reference to marine shrimp
and pearl oysters”. Frontiers in Genetics; 9: 693.