unidad 3 - ual.dyndns.orgual.dyndns.org/biblioteca/psicologia_del_desarrollo_i/pdf/unidad... · la...
TRANSCRIPT

Unidad 3
• Bases Genéticas y de Desarrollo de la Conducta

Los rasgos dominantes asociados con la presencia de un cromosoma Y adicional incluyen altura inusitada, episodios de agresividad excesiva e inteligencia mínima. Sin que a nadie sorprenda el hecho, en algunos de los más famosos criminales de nuestro tiempo se ha encontrado un cromosoma Y adicional. Un caso entre los primeros de esta índole fue el de Robert Tait, un australiano que en 1962 fue convicto por haber dado muerte con una porra a una anciana de 81 años, en una vicaría donde él había ido en busca de ayuda. En 1965, Daniel Hugon, un establero francés de 31 años de edad, estranguló brutalmente a una prostituta parisiense, aparentemente sin motivo alguno. Durante el curso de su juicio se le halló un cromosoma Y adicional. En abril de 1969, John Farley de seis pies y ocho pulgadas de estatura, y 240 libras de peso, apodado "El alegre gigante verde"; a causa de su extraordinaria dotación física y "Juanote el malo'; por sus arrebatos violentos, confesó haber golpeado, estrangulado, violado y mutilado a una mujer de Queens, Nueva York. Fue defendido sobre la basa de que, debido a la presencia de un cromosoma Y adicional en sus células, él no tenía discernimiento ni control de sus acciones y fue hallado por lo tanto no culpable "en razón de su demencia causada por un desequilibrio cromosómico". (Jarvik y otros, 1973, p. 675)
Estos ejemplos aterradores son una demostración clara de la importancia de los factores genéticos en la conducta, sin hacer mención del carácter excéntrico de nuestro sistema legal. Los individuos que poseen un tipo genético de XYY son del sexo masculino con el sobrante de un cromosoma Y. El tipo genético masculino no normal es "XY" el tipo genético femenino normal, XX. La evidencia actualmente refuerza el concepto de que este cromosoma Y adicional es mucho más común entre hombres convictos de crímenes violentos que entre hombres de la población general.
El conjunto genético ciertamente influye en la conducta como lo indica el síndrome XYY. Más aún, es posible argumentar que la agresión humana puede "residir" en el cromosoma Y. Las mujeres son mucho menos agresivas que los hombres, por lo menos en lo que a violencia física se refiere. Hay dos explicaciones posibles: los hombres son más agresivos y las mujeres lo son menos porque la sociedad enseña estos papeles sociales, y (2) hay una determinación genética inherente a 13 violencia en los hombres. Los datos del síndrome XYY parecen dar cierto apoyo a la hipótesis genética.
La importancia del síndrome se extiende mucho más allá del problema de la violencia considerado en sí mismo. Las anormalidades cromosómicas descubiertas anteriormente se han relacionado con anormalidades físicas o con retardo mental - clasificación burda y global de defectos. La persona que tiene el síndrome de Klinefelter (tipo genético XXY) es de sexo femenino con muchas características masculinas - la "mujer" atleta de aspecto masculino. La persona que tiene el síndrome de Down (mongolismo) posee un cromosoma adicional no ligado al sexo y es retardado mental

además de tener ciertos rasgos y defectos físicos característicos. Se trata de anormalidades físicas e intelectuales relativamente importantes. Sin embargo, el tipo genético XYY produce una anormalidad que es primordialmente conductual - la agresión.
¿Cómo conduce el cromosoma Y a la violencia? Los cromosomas son sim-plemente, después de todo, mezclas de sustancias químicas. El cromosoma Y (masculino), sea dicho de paso, es físicamente mucho más pequeño que el cromosoma X (femenino) Puede describirse como regresivo o como más dominante, dependiendo tal vez de los propios cromosomas de quien opina. De todos modos, la combinación XY produce un individuo de sexo masculino normal y la combinación XX un individuo de sexo femenino normal, con todas las diferencias físicas y conductuales normales. El cromosoma Y adicional exagera varias características masculinas - por ejemplo la altura y la contextura muscular (algunos podrían agregar así mismo, la inteligencia reducida) - además de la agresión. Esto indica que la estructura física y la actividad glandular son en cierta forma controladas por los cromosomas X y Y El desarrollo cerebral mismo - el crecimiento y estructura del sistema nervioso - está bajo el control genético y recibe la influencia de las hormonas. Hay sin embargo todavía, grandes vacíos en la forma como nosotros entendemos que los cromosomas o sea el material genético, controlan el crecimiento y desarrollo y a fin de cuentas la conducta de los seres humanos.
Cada cromosoma está compuesto de una gran cantidad de genes, última unidad estructural de la herencia. Las estructuras y funciones de todos los organismos están determinadas por los genes. Realmente los genes tan sólo determinan las estructuras de las sustancias químicas denominadas proteínas. Estas son de dos tipos: proteínas estructurales y enzimas. Las proteínas estructurales del organismo y las enzimas controlan las reacciones fisicoquímicas del organismo.
Teóricamente todas las funciones biológicas y conductuales pueden retrotraerse a la acción genética. En la práctica sin embargo, al desarrollarse el organismo a partir del momento en que se fertiliza el huevo, los efectos genéticos interactúan con el ambiente. La distinción decisiva se hace entre el genotipo (la estructura genética subyacente de un organismo) y el fenotipo (expresión externa de la acción genética como resultado de la interacción con el ambiente) Entre los animales superiores, toda conducta es fenotípica.
En este capítulo examinaremos los fundamentos de la genética, las bases genéticas de la conducta (materia que es objeto de un nuevo campo fascinante denominado genética conductual), el desarrollo del sistema nervioso y la influencia relativa de la estructura genética y del ambiente en la conducta.
GENÉTICA BÁSICA Las leyes de la herencia
Aunque Darwin postuló que la selección natural operaba por la herencia de las características que demostraban adaptabilidad, el mecanismo era oscuro. Por el tiempo en que Darwin estaba formulando la versión final del Origen de las Especies, un monje

austriaco, Gregor Johan Mendel, experimentaba con la hibridación de los guisantes. Los hallazgos de Mendel publicados en 1866, lo condujeron a las hipótesis de que (1) la herencia se transmite por unidades independientes y heredables; (2) cuando cada padre aporta la misma clase de unidad, la progenie presenta este carácter, pero cuando cada padre suministra una clase diferente de unidad, se forma un híbrido y cuando los híbridos forman sus propias células reproductivas, "se liberan" nuevamente las dos clases diferentes de unidades; y (3) las unidades hereditarias no se afectan por la experiencia de un individuo.
En tales hipótesis formuló Mendel las leyes básicas de la herencia-la noción de los genes-pero por más de tres décadas permaneció desconocido su trabajo. En un lapso de cinco meses durante el año de 1900; tres botánicos europeos redescubrieron el trabajo de Mendel y lo pusieron cara a cara con la biología. El más conocido entre ellos era Hugo De Vries, el cual hizo hincapié en el concepto de las mutaciones como responsables del desarrollo de especies nuevas. Los estudiosos modernos de la genética no insisten en la mutación en la medida de De Vries y sus seguidores. Todos son conscientes sin embargo de que aunque el material genético de una población se halle en proceso continuo de mezcla por razón de los apareamientos, el único camino para que be introduzcan caracteres completamente nuevos es el de la mutación. La base de la herencia
La ciencia de la genética empezó con el redescubrimiento de la obra de Mendel. Se sabía ya que el núcleo de cada célula contiene cierto número de pares de finas estructuras filiformes que, se reproducen por sí mismas cuando se divide la célula y a las cuales se llamó : cromosomas...
Se sabía que las diferentes especies poseen números diferentes de pares-la mosca frutera, 8 cromosomas ó 4 pares estructuralmente similares; el ser humano normal, 46 cromosomas ó 23 pares (Figura 2.1). Se sabía también que cuando se forma un huevo o una célula espermática, no se incorpora en la nueva célula sexual sino un cromosoma de cada par, de tal manera que la fertilización del huevo por parte del espermatozoide produce un individuo nuevo con una complementación total de pares de cromosomas.
En años posteriores se hizo evidente mediante los esfuerzos de hombres tales como Thomas Hunt Morgan y H. J. Muller, que las unidades hereditarias de Mendel, los genes, dependen de los cromosomas. Se descubrió que si por medio de radiación se producen mutaciones de los genes, se forman organismos anormales. Se demostró que los cromosomas constan principalmente de dos sustancias: ácido desoxirribonucleico (DNA) y proteínas. De los dos, el DNA es el más estable, nunca abandona el núcleo y es capaz de reproducirse por sí mismo. Tiene la propiedad fundamental de los genes. Según estudios posteriores, la totalidad del DNA tiene la misma composición química. Consta de un azúcar puro (desoxirribosa), fosfato y cuatro bases: adenina, guanina, timina y citosina (A, G, T, C,).
El siguiente gran paso ocurrió con el descubrimiento de J. D. Watson y F. H. C. Crick respecto de la estructura de la molécula del DNA. Utilizando la cristalografía de

rayos X, Watson y Crick propusieron el modelo de doble hélice (Figura 2.2). Según este modelo, la molécula consta de dos ramales mutuamente enrollados en espiral. El sostén de cada ramal consiste en un arreglo continuo de azúcar-fosfato-azúcar-fosfato. Las bases A, G, T y C están ligadas a dichos sostenes y a su vez se unen entre sí, vinculando en tal forma los dos ramales. Aunque las bases pueden presentarse en cualquier secuencia a lo largo de un ramal, existe una relación fija entre los ramales com-
Figura 2.1 Cromosomas humanos masculinos tomados de un glóbulo blanco. Se usó un
procedimiento que tiñe selectivamente solo los cromosomas de la célula. Los cromosomas aquí mostrados se encontraban en el proceso de división. El homólogo de cada uno de ellos se encuentra localizado en alguna otra parte de la microfotografía.
plementarios. La adenina se une siempre con la timina y la guanina siempre con la citosina. El código genético está contenido en la secuencia de las bases. Dicho código determina las propiedades y efectos de cada gene. La capacidad del DNA para reproducirse por sí mismo depende de la combinación específica de ligaduras de las bases (Figura 2.3). Cuando los ramales se separan durante la división de las células, son capaces de producir copias idénticas de la molécula original DNA seleccionando sus compañeros apropiados.
Los mecanismos de la herencia El DNÁ nunca abandona el núcleo. Los genes ejercen su acción directa deter-
minando las proteínas que deben producirse en los ribosomas (estructuras especializadas que se hallan fuera del núcleo en el citoplasma de la célula). El

mecanismo de traducción del código genético (secuencia de las bases en la molécula DNA) en una proteína implica ácido ribonucléico (RNA), sustancia que en su estructura es similar al DNA pero está presente en toda la célula. Una clase especial de RNA denominada RNA mensajero, transfiere el código del DNA al ribosoma, donde otro RNA el RNA de transferencia, probablemente actúa en la recolección de los aminoácidos particulares requeridos para la elaboración de la proteína codificada por el DNA (Figura
Figura 2.2 El modelo de doble hélice para la duplicación del ADN propuesto por Watson y Crick.
Los dos cabos del ADN se enroscan entre sí formando una doble hélice. Durante la división la molécula se desenrolla, las bases se separan quedando una base de cada par ligada a cada cabo. Tal como se muestra aquí, se forma un cabo complementario, que origina después del fenómeno de duplicación, dos moléculas de ADN. (Tomado de Stent, Gunther S. Molecular Biology of Bacterial Víruses. Copyright ©1963 por W. H. Freeman y Co.)

Figura 2.3 Un segmento de ADN que muestra las combinaciones de enlaces entre las cuatro
bases que lo forman: adenina, guanina, timina y citosina. Como se muestra aquí, la adenina solo se enlaza con la timina y la guanina solo con la citosina. (Segundo dibujo tomado de Lerner, 1. Michael. Heredity, Evolution and Society. Copyright © 1968 por W.H. Freeman y Co.)
2.4). Los tipos de proteínas como también los lugares y oportunidades de su conforma-ción, determinan el sexo de un individuo, el color de su piel e indudablemente indican si se trata del un gusano de la tierra, una mosca frutera, un ratón o un hombre.
Por lo general la formación del RNA ocurre en ocasiones en que las células realizan funciones fisiológicas, pero no mientras se están multiplicando. Durante estos períodos los dos ramales de la hélice ADN están desenrollados. Solamente uno de ellos produce RNA en forma activa. Realiza tal función seleccionando bases complementarias del material celular, pero remplaza la timina (T) por uracila (U). Cuando se termina la cadena de RNA, se desprende del DNA, abandona el núcleo de la célula y se adhiere a los ribosomas (Figura 2.4). Este es el RNA mensajero. El RNA de transferencia lleva aminoácidos a los sitios de los ribosomas, donde tales aminoácidos se recogen en proteínas - en esencia, largas cintas de, aminoácidos enganchados unos con otros. Hay 20 aminoácidos que forman la mayor parte de las proteínas. Hay cuatro bases diferentes de RNA y se requieren tres para especificar un aminoácido. Tomando cada vez tres de las cuatro bases, se llega a un total de 64 aminoacidos-cantidad considerablemente superior a los propios 20 requeridos. Finalmente, las proteínas y enzimas estructurales se forman en los ribosomas.
Los genes de un rasgo particular se presentan en lugares específicos de cromosomas específicos. Los dos cromosomas que forman cada par tienen dentro de sí lugares homólogos. Cuando se forma el huevo o la célula espermática solamente se incluye un cromosoma de cada par y así tan sólo uno de los dos genes presentes en los lugares homólogos del padre queda incluido. Cuando ocurre la fecundación, la célula posee de nuevo cromosomas apareados, pero el 50 por ciento de los genes es de cada uno de los padres. Los diversos genes (secuencias de las bases) que puede haber en un solo lugar cualquiera se denominan alelos. Si el cigote recién formado (huevo fertilizado) hereda el mismo alelo en determinado lugar de los dos cromosomas homó-logos, se dice que el individuo es homocigótico en cuanto a ese gene; si diferentes alelos están presentes en los dos lugares homólogos, el individuo es heterocigótico respecto de ese gene. En el caso más simple, cuando un gene en determinado lugar es

Figura 2.4 Mecanismo de traslación del código genético a una proteína: La información de la
molécula de ADN se transcribe a una molécula de ARN (1) Por un proceso de apareamiento se forma un cabo complementario de ARN (llamado mensajero) (2). El ARN mensajero transfiere el código del ADN a los ribosomas en el citoplasma (3). Los ribosomas son los elementos donde se realiza la síntesis de proteínas, los ARN llamados de transferencia y los aminoácidos ya enlazados (mostrados aquí como números encerrados en rectángulos) se alinean sobre el ARN mensajero en una secuencia particular (4). Estos aminoácidos se combinan formando cadenas de polipéptidos que constituyen las proteínas. (Adaptación de Niremberg, M. W. El código genético: II. Copyright © 19(33 por Scientific American, Inc. Reservados todos los derechos.)
dominante (R) y otro es recesivo (r), el genotipo de cualquier miembro de la población puede ser RR, Rr, o rr. Supóngase que R produce ojos rojos y r ojos grises, en la mosca frutera. Cuando dos miembros heterocigóticos de la población, ambos de ojos rojos se aparean (RrXRr), las proporciones esperadas de los genotipos de la cría serán de 1:2:1 (RR:Rr:rr) y la proporción de fenotipos será de 3:1 (esto es, tres de cada cuatro tendrán los ojos rojos; Figura 2.5). Lo que aquí importa anotar es que se mantiene la variabilidad en la población. Más aún, siendo que uno u otro cromosoma de cualquier par puede ser aportado al cigote y dado que la integración de los cromosomas es aleatoria, se incrementa en gran manera la variabilidad genética a través de la reproducción sexual.
En el último ejemplo, la acción genética dominante es completa. No es posible por el simple examen de los ojos (fenotipo) distinguir la mosca homocigótica de ojos rojos (RR) respecto de la mosca heterocigótica de ojos rojos (Rr). En muchos casos `el predominio es incompleto. Un ejemplo importante desde el punto de vista médico es la

anemia de células falciformes, enfermedad hereditaria más común entre los negros de la costa central de África o entre aquellos cuyos antepasados vinieron de aquella vasta región. El mal se caracteriza por una forma anormal de hoz en las células rojas de la sangre, la cual causa en ellas la formación de un precipitado. Se presenta co-
Figura 2.5 Ejemplo de la acción de los genes dominantes y recesivos en la peterminación del color de :os ojos (fenotipo) en la mosea de la fruta.
Cuando dos moscas son heterócigas por el color de los ojos, la proporción de fenotipos en su descendencia será de 3:1 esto es, tres de cada cuatro tendrán los ojos rojos. Sin embargo la proporción de los genotipos es de 1:2:1 de modo que cuando tres moscas tienen ojos rojos solo una de ellas es homóciga por el color (rojo) de los ojos.
mo resultado de la acción de un solo gene. Un individuo que sea homocigótico respecto de ese gene, padece una enfermedad grave; la mayor parte muere en la niñez, y los que sobreviven son enfermos crónicos de por vida.
En los Estados Unidos solamente un 0,25 por ciento aproximado de la población negra tiene tal enfermedad en la forma homocigótica, pero aproximadamente el 9 por ciento es heterocigótico respecto del gene anormal. Ciertas tribus africanas tienen hasta un 40 por ciento de su población heterocigótica y un 4 por ciento homocigótica. En los individuos heterocigóticos respecto del gene de célula falciforme, el predominio del gene normal es incompleto. En circunstancias ordinarias tales personas se presentan llenas de salud, pero enferman al encontrarse en regiones de altura considerable donde la sangre no recibe oxígeno suficiente. Así pues, los portadores que tienen un gene normal y un gene de célula falciforme ordinariamente no se ven afectados, pero res-ponden en forma anormal al impacto del oxígeno escaso.
Lo que causa desconcierto en tal enfermedad es su gran incidencia en ciertas regiones de África. Todo lo que sabemos acerca de la evolución, indica que tal gene debilitador no debe existir en una proporción tan alta de la población. La razón de su

persistencia se aclaró cuando se observó que las regiones de África donde la anemia de células falciformes es más común, son así mismo las regiones donde la malaria está más extendida y reviste mayor gravedad. La correlación es muy estrecha. En el cinturón central de África, las tribus que viven en regiones ajenas a la malaria no tienen virtualmente anemia de células falciformes. Los individuos heterocigóticos respecto del gene de célula falciforme tienen mejores posibilidades de sobrevivir que las personas normales o los homocigóticos respecto del gene anormal. Un epílogo halagüeño de esta historia lo constituye el hecho de que la incidencia del gene de células falciformes se halla en descenso continuado entre los negros de América donde la anormalidad no tiene valor adaptativo. Este es, en efecto, un ejemplo de la evolución humana en su curso actual.
La variabilidad genética es importante en cualquier población. Es necesaria no sólo para el desarrollo de especies nuevas, sino también para la supervivencia de una especie existente que se enfrenta aun ambiente de continua mutación. La variabilidad de la fusión genética de una población aumenta por razón de procesos tales como la elección aleatoria en la reproducción, las mutaciones (arreglos repetidos de las bases de la molécula DNA), el entrecruzamiento (rompimiento de cromosomas e intercambio de partes entre ares, lo cuales permiten combinaciones nuevas de genes en un cromosoma dado) y la existencia de más de dos alelos para determinado lugar cro-mosómico.
Bases Genéticas De La Conducta
Los genes no actúan directamente sobre la conducta. Intervienen tan sólo para regular la producción de proteínas responsables de la estructura y actividades enzimáticas de un organismo. Sin embargo, desconocemos la base estructural de la conducta en la mayoría de los casos. Por ejemplo, sabemos que un caballo desprovisto de patas no puede correr y conocemos ciertos aspectos de la morfología de los huesos y de los músculos la cual se correlaciona con la mayor velocidad de los caballos. Pero esto no basta; ningún criador de caballos podría ir adelante si basara su negocio exclusivamente en los rasgos anatómicos de las patas de los animales. Otros rasgos-el temperamento, la coordinación neuromuscular, la tenacidad-cuentan mucho en un buen caballo de carreras. En lugar de considerar únicamente las patas, los criadores intentan aparear los animales que han demostrado éxito en el hipódromo. De manera semejante, los investigadores de las bases genéticas de otras conductas realizan sus estudios, sea examinando individuos que ostentan una conducta particular, sea analizando el linaje de individuos que difieren en una conducta dada.
Deben destacarse dos puntos relacionados con el estudio de la genética conductual. Primero, este enfoque no deja de lado los efectos del ambiente en la conducta. Por el contrario, estudia la interacción de los genes y el ambiente tratando de cuantificar la porción de variabilidad observada en un rasgo de determinada población, la cual resulta de las diferencias genéticas entre los individuos y aquélla que es producto de las diferencias ambientales. El porcentaje de variación de un rasgo que se debe a diferencias de genotipo se ha denominado heredabilidad de un rasgo. Esta es una medida o índice numérico que indica la medida en que se hereda un rasgo,

característica o tendencia conductual. Segundo, la gran mayoría de las características conductuales no depende de un solo gene que actúa en una forma totalmente común o completamente particular. Más bien las conductas dependen de muchos lugares donde la contribución de cada gene es pequeña y acumulativa. Estos sistemas se denominan poligénicos.
Estudios experimentales acerca de la manipulación genética Tratándose de animales es posible disponer las condiciones de laboratorio en tal
forma que el ambiente se mantenga razonablemente constante y así mismo pueda practicarse la selección artificial (1) midiendo una característica de conducta particular en una población de laboratorio, (2) seleccionando animales de los extremos superior e inferior y aparcando animales dentro de los extremos (los de puntaje alto con los de puntaje alto y los de puntaje bajo con los de pontaje bajo) y (3) probando las crías y continuando en cada generación subsiguiente el apareamiento entre los extremos. Si en tales condiciones ambientales hay influencia genética en la variabilidad de la conducta, las dos poblaciones diferirán de cada generación sucesiva hasta agotarse la escala de variabilidad genética.
Los primeros estudios que utilizaron la selección natural fueron dirigidos por Robert Tryon en Berkeley. Tryon escogió el aprendizaje de laberinto en las ratas. Después de medir la ejecución de laberinto en una muestra no seleccionada, apareó los ejemplares más rápidos ("brillantes en el laberinto") entre sí, e igualmente los más lentos ("lerdos de laberinto") con los más lentos. Habiéndose cumplido tal procedimiento por ocho generaciones, los dos grupos no presentaron casi ningún traslape (Figura 2.6).
Los experimentos de cría selectiva realizados con ratas y ratones han demostrado que gran cantidad de rasgos conductuales pueden variarse por medio de la manipulación genética. Las características estudiadas incluyen aprendizaje de laberinto, emotividad medida por la tendencia a defecar en una situación de campo abierto, actividad medida por el recorrido realizado en una rueda giratoria, agresividad, proporción de aprendizaje de evitación y susceptibilidad a ataques inducidos por sonidos. Muchos investigadores se han vuelto recientemente hacia la mosca frutera Drosophila para el estudio de las influencias genéticas en la conducta. La Drosophila puede educarse para una conducta foto táctica-acercamiento versus no acercamiento a una fuente de luz-o para una conducta geotáctica-tendencia a subir o bajar en un punto de elección. En un estudio fototáctico realizado en la mosca frutera, Hirsch y Boudreau (1958) cruzaron animales que tendían a acercarse a una fuente de luz (fototaxis positiva) y cruzaron animales que huían de ella. Después de 29 generaciones estimaron que aun en este organismo relativamente simple sólo el 57 por ciento de la variación estaba determinado en forma genética.
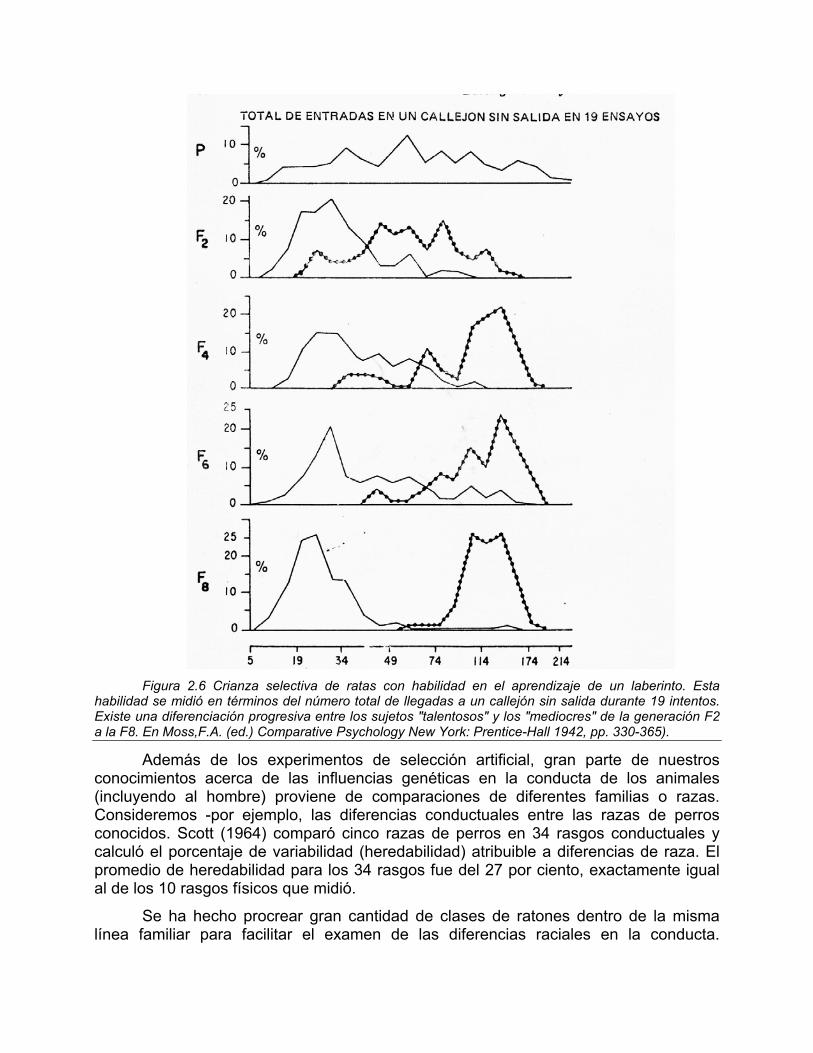
Figura 2.6 Crianza selectiva de ratas con habilidad en el aprendizaje de un laberinto. Esta
habilidad se midió en términos del número total de llegadas a un callejón sin salida durante 19 intentos. Existe una diferenciación progresiva entre los sujetos "talentosos" y los "mediocres" de la generación F2 a la F8. En Moss,F.A. (ed.) Comparative Psychology New York: Prentice-Hall 1942, pp. 330-365).
Además de los experimentos de selección artificial, gran parte de nuestros conocimientos acerca de las influencias genéticas en la conducta de los animales (incluyendo al hombre) proviene de comparaciones de diferentes familias o razas. Consideremos -por ejemplo, las diferencias conductuales entre las razas de perros conocidos. Scott (1964) comparó cinco razas de perros en 34 rasgos conductuales y calculó el porcentaje de variabilidad (heredabilidad) atribuible a diferencias de raza. El promedio de heredabilidad para los 34 rasgos fue del 27 por ciento, exactamente igual al de los 10 rasgos físicos que midió.
Se ha hecho procrear gran cantidad de clases de ratones dentro de la misma línea familiar para facilitar el examen de las diferencias raciales en la conducta.

Aislando una población y apareando continuamente parientes cercanos, puede reducirse radicalmente la variabilidad genética de una población. Mediante el aislamiento y la reproducción libre de mezclas, pueden producirse familias en las cuales los individuos son genéticamente uniformes y diferentes de los de otras familias. Se ha descubierto que las familias de ratones de este tipo difieren en cierta cantidad de rasgos conductuales que incluyen agresividad, actividad locomotora, aprendizaje, preferencia alcohólica, conducta de atesoramiento, cantidad de crías, conducta exploratoria, resistencia a la depresión y conducta de apareamiento. Aunque se hacen muchas críticas al enfoque de la reproducción lineal [dentro de la misma familia] con base principalmente en el hecho de que tal cantidad de homocigóticos producidos nunca podrían presentarse en una población natural próspera, este método nos ha ayudado inmensamente a comprender las influencias genéticas en la conducta.
El grado de actividad locomotora suministra un ejemplo claro de una característica conductual heredada en familias lineales de ratones (Figura 2.7). Se probaron los niveles generales de actividad de seis familias diferentes colocando los animales sobre una reja dividida en cuadros. La actividad se determinó por el número de cuadros cruzados en un período de cinco minutos. Como se observa en la Figura 2.7, las seis familias presentaron grandes diferencias en el grado de actividad locomotora. Resultó igualmente diferente la actividad dentro de una familia en cuanto a la medida de la variabilidad. La familia marcada en la gráfica superior no presenta virtualmente actividad alguna y se observa poca variabilidad. Por el contrario, la familia marcada en la gráfica más baja presenta poca actividad pero una variabilidad considerable. Thiesen (1972) ha demostrado que tales diferencias de familia en la ac-tividad se mantienen en una gran variedad de situaciones de prueba durante años de reproducción. La actividad se presenta como un rasgo conductual estable, genéticamente determinado.
La emotividad parecería ser un aspecto mucho más complejo de la conducta que la actividad. Sin embargo, utilizando como simple indicio de emotividad cierta cantidad de operaciones defecatorias en una situación de prueba en campo abierto, Lindzey y otros (1960) lograron demostrar que la capacidad de respuestas emotivas difiere según las diversas familias de ratones y como tal permanece durante años de procreación. La emotividad, por lo menos en familias lineales de ratones es un rasgo relativamente estable y marcadamente determinado por factores genéticos.
Un ejemplo curioso de reproducción lineal se encuentra en el ratón bailador. Se creía que cl desarrollo de tal familia se remontaba hasta el año 80 A.C. en China, y que probablemente tiene una genealogía continua que se extiende hasta el presente. La conducta de los bailadores es realmente sorprendente: giran con rapidez durante largos períodos sin presentar indicios de vértigo. El defecto genético se expresa claramente en un daño del sistema vestibular y del oído interno.
La susceptibilidad a los ataques audiogénicos es otro ejemplo. Ciertas familias de ratones, frente a determinados sonidos, sufren ataques de efectos similares a los de la epilepsia, que les causan frecuentemente la muerte. No se requiere que el sonido

CUADROS ENTRADAS
Figura 2.7 Grado de actividad locomotriz en seis familias lineales de ratones. La actividad locomotriz se definió como el número de cuadros cruzados durante períodos de cinco minutos. La frecuencia indica el número de animales de cada familia que realizó un número dado de cruces. Nótense las diferencias en el grado de actividad locomotriz entre las distintas familias así como las diferencias en variabilidad de la actividad dentro de las mismas familias. (McClearn, datos sin publicar).
sea fuerte; el retintín de las llaves es un estímulo efectivo para que se produzca el ataque. Este rasgo resulta estar bajo el control de varios genes.
Efectos de la reproducción lineal o selectiva Como se habrá dado cuenta el lector, sin duda alguna, los que padecen ataques,
o los ratones bailadores, no son adaptables. En general, la reproducción lineal tiende a resultados que se manifiestan en defectos tanto físicos como conductuales. Por cada diez crías selectivas de ratones iniciadas por los criadores, mueren por lo menos nueve y los efectos deletéreos de la reproducción lineal son evidentemente más graves aún en el hombre que en el ratón. En general, el grado de parentesco entre las parejas humanas se relaciona directamente con los efectos perjudiciales en los niños. En la población general, la tasa de mortalidad entre los niños de diez años de edad es de 24 por cada 1000. En los niños de matrimonios entre primos es de 81 por cada 1000

(Thiessen, 1972). En un estudio realizado con niños japoneses en edad escolar se descubrió que los niños de primos en primer grado son menos aptos realmente en todas las medidas (se utilizaron 17i) que los niños de control, de padres no relacionados genéticamente (Schull and Neel, 1965). Estos datos aparecen en la Tabla 2.1. En comparación con el grupo de control, los niños empezaban a caminar y hablar en edad más tardía, demostraban menos vitalidad física, alcanzaban puntajes inferiores en pruebas de inteligencia y daban menos rendimiento en la escuela. Causan mayor impresión aún los efectos perjudiciales en los niños producto de los llamados incestos nucleares (hermano con hermana, padre con hija). Adams y Neel (1967) compararon 18 de tales niños con otros de padres no emparentados, iguales en edad, peso, inteligencia y situación socioeconómica. De los niños nacidos de incesto, 5 murieron, 2 eran retardados mentales, 1 tenía el paladar hendido y 3 presentaban inteligencia mínima. Solamente 7 de los 18 parecían moderadamente normales. De los niños de control, 1 presentaba un defecto físico menor y todos los demás eran normales.
El efecto devastador en la viabilidad, salud e inteligencia de los niños causado por el apareamiento entre parientes parece haber sido reconocido por la mayoría de las sociedades primitivas. Indudablemente, ninguna sociedad o cultura de que tengamos conocimiento permite el incesto. Los Faraones del Antiguo Egipto que produjeron una línea notablemente débil de gobernantes por incesto, constituyeron la excepción en su cultura. Lindzey (1967) hace la anotación interesante de que el tabú universal del incesto puede haber sido el resultado de la selección natural al actuar en las culturas humanas primitivas. Todas aquellas tribus que no impidieron el incesto desaparecieron hace mucho tiempo.
Efectos de la hibridación El vigor de los híbridos está al extremo contrario de la reproducción selectiva.
Cuando se juntan dos razas separadas, los hijos tienden a ser más vigorosos que los padres. Este vigor de los híbridos ha sido del conocimiento de los agricultores desde hace mucho tiempo. Todo el grano que se produce hoy es híbrido. Los cereales híbridos son más robustos y echan espigas más largas que las familias que les dieron origen. No hay evidencia clara acerca del vigor híbrido en el hombre. El autor quisiera mencionar los pueblos de Polinesia como ejemplo, pero no existen datos reales. La enfermedad de células falciformes es un ejemplo de matices especiales. En las áreas palúdicas del África los niños heterocigóticos respecto del gene de célula falciforme tienen, en cuanto a la reproducción, una ventaja sobre los individuos homocigóticos y normales. Un caso más desconcertante es el de la enfermedad de Tay-Sachs, forma devastadora de deficiencia mental infantil en el individuo homocigótico respecto del gene apropiado. Se presenta principalmente entre la población judía Ashkenázica. En forma suficientemente extraña, los individuos heterocigóticos respecto del gene presentan una ligera ventaja reproductiva (6 por ciento) sobre los normales (Dobzhansky, 1964).

Tabla 2.1 Efectos de la reproducción lineal sobre la conducta de niños escolares Japoneses
Características Sexo
Promedio de los niños control
Promedios de los hijos de primos en
primer grado
Efecto de la reproducción
lineal Edad en que caminaron (meses) M 14.06 14.19 0.13 F 13.62 14.07 0.45 Edad en que hablaron (meses) M 11.81 12.60 0.79 F 10.38 10.82 0.44 Puntajes de inteligencia Puntaje de realización M 57.37 54.94 2.06 F 55.10 52.52 2.06 Puntaje Verbal M 58.67 55.34 2.76 F 57.01 53.46 2.76 Grado Escolar: Lenguaje M 3.09 2.95 0.10 F 3.28 3.10 0.10
Datos Tomados de Schull, W. J., y Neel, J. V. Los efectos de la reproducción lineal sobre niños Japoneses. Nueva York: Harper & Row Publishers, Inc. (1965).
Estudios en gemelos
La mayor parte de la investigación acerca de las influencias genéticas en la conducta humana ha utilizado gemelos, comparando en general gemelos (esto es, mellizos idénticos) o monocigóticos (MZ) y mellizos o dicigóticos (DZ) del mismo sexo. La razón de tal enfoque consiste en que los gemelos MZ son genéticamente idénticos mientras los mellizos DZ no son en el campo genético más similares que los hermanos comunes. Así pues cualesquiera diferencias entre gemelos MZ deben ser completamente ambientales, mientras las diferencias entre mellizos DZ son tanto hereditarias como ambientales. En la suposición de que los efectos del ambiente sean iguales en los dos grupos, las diferencias entre los gemelos MZ pueden sustraerse de las que existen entre los mellizos DZ, y el resultado debe ser una medida de los efectos de las diferencias genéticas. En esta forma se han realizado numerosos estudios acerca de puntajes en pruebas de inteligencia como también en variables de personalidad. Normalmente los investigadores han encontrado componente genético pequeño pero confiable en las variables de la personalidad y un componente genético mayor en los puntajes de pruebas de inteligencia. En este capítulo volveremos sobre este punto más adelante.

Consecuencias conductuales de los defectos Ja un solo gene Ciertos tipos de deficiencia mental grave resultan de los defectos existentes en
un solo lugar genético. Por ejemplo, una enfermedad conocida como fenilquetonuria (PKU) se caracteriza por un retardo mental grave y la presencia de ácido fenilpirúvico en la orina. La fenilquetonuria resulta de la herencia de un doble gene recesivo. El individuo que es homocigótico respecto del gene produce cantidades insuficientes de una sustancia requerida para la conversión normal de la fenilalanina (aminoácido común presente en muchos alimentos de proteínas) en tirosina. Se cree que el proceso mismo de la enfermedad resulta de los aumentos de niveles en los tejidos y la sangre, del ácido fenilpirúvico y otros subproductos no-normales de la fenilalanina, los cuales actúan como toxinas en el cerebro en desarrollo. Cuando el infante afectado tiene de cuatro a seis meses de edad, ha aparecido el daño cerebral grave e irreversible. Actualmente es posible detectar la fenilquetonuria en asunto de horas después del nacimiento y prevenir la grave deficiencia mental suministrándole al niño una dieta baja en fenilalanina. La mayor parte de los hospitales examinan rutinariamente a cada niño acerca de la fenilquetonuria dentro de los dos días siguientes al nacimiento. Hoy es así mismo posible identificar a los portadores heterocigóticos del gene recesivo y advertir a tales personas mediante consejo genético acerca del peligro para sus hijos antes del nacimiento o aún de la concepción.
Actualmente se sabe que se transmiten genéticamente muchas otras enferme-dades metabólicas que causan deficiencia mental. Dos de ellas, la idiocia amaurótica infantil (enfermedad de Táy-Sachs) y la idiocia amaurótica juvenil resultan de la herencia de un doble gene recesivo de un solo lugar e implican mal funcionamiento del metabolismo lípido. Los efectos de la primera son notorios a los pocos meses de edad; los de la segunda entre los dos y los diez años. La corea de Huntington, caracterizada por deterioro mental progresivo como también por movimientos involuntarios, ilustra el hecho de que la deficiencia mental genéticamente causada no necesariamente aparece al principio de la vida. Esta enfermedad se transmite por un gene dominante y ataca entre los 30 y los 50 años.
Consecuencias conductuales y físicas de las anormalidades cromosómicas
La anormalidad en la complementación cromosómica es responsable de ciertos desórdenes mentales y físicos. El ejemplo más conocido es el síndrome de Down (llamada también mongolismo por tener sus víctimas los ojos sesgados ). Además de los ojos sesgados y otros rasgos morfológicos, las personas afectadas son marcadamente retardadas (el Cl está generalmente entre 20 y 60). El síndrome de Down que es la causa aislada determinable más común del retardo mental, tiene una ocurrencia aproximada de 1 por cada 500 a 600 nacimientos y representa el 10 por ciento aproximado de las personas de reconocido retardo mental en los Estados Unidos. Es causada por una anormalidad cromosómica llamada trisomía -21. A veces en lugar de la complementación general de 46 cromosomas (23 pares), la persona tiene 47, incluyendo un cromosoma adicional del par número 21. A veces, durante la formación de las células sexuales, se han intercambiado partes de los cromosomas 21 y 15 y la persona afectada porta el cromosoma adicional 21 como parte del par número

15. Aunque se desconocen las razones por las cuates se presenta el material genético adicional, se ha demostrado que es u. s probable que el síndrome de Down ocurra en niños de mujeres cuya edad supera el promedio gestatorio.
Los efectos de las anormalidades de número y clase de cromosomas sexuales en la estructura y personalidad humanas han despertado gran interés. Normalmente un individuo de sexo masculino tiene un cromosoma X y un cromosoma Y, mucho más corto que el X. La hembra normal tiene dos cromosomas X. La persona que tiene el síndrome de Klinefelter tiene un cromosoma Y y dos X (genotipo XXY) y es fenotípicamente masculina con cierto grado de feminización, reducido impulso sexual y a veces retardo mental. Se ha demostrado que algunas "mujeres" soviéticas, atletas de habilidades físicas superiores a las de las mujeres normales, tienen el síndrome de Klinefelter, situación que crea problemas tanto de definición como de relaciones in-ternacionales. Aunque se ha informado que muchas de las personas que tienen el síndrome de Klinefelter tienen problemas de personalidad, no es posible desenmarañar la causa y el efecto y los datos no son claros. El síndrome de Klinefelter es desconcertantemente común pues se presenta en 1 de cada 400 nacimientos.
La persona que tiene el síndrome de Turner tiene un cromosoma X normal pero carece del cromosoma Y (genotipo XO). Tal individuo es fenotípicamente femenino, pero es bajo de estatura y tiene características sexuales atenuadas. Se presenta normal en inteligencia general pero presenta una deficiencia específica en discriminaciones espaciales. Tal síndrome no es muy frecuente: 1 caso en 3000 nacimientos.
Tal vez la que causa mayor curiosidad entre las anormalidades cromosómicas sexuales es el síndrome XYY (p.33). Los individuos de genotipo XYY tienden a ser masculinos, de gran estatura, musculados, retardados, de mucha agresividad. El síndrome se encontró inicialmente entre prisioneros y muchos estudios han reducido sus muestras a prisioneros de gran estatura y con antecedentes de episodios violentos--limitación que obviamente introduce un prejuicio de muestreo. El hecho es que las investigaciones realizadas con tales prisioneros revelan en su mayoría una incidencia de genotipo XYY de un 2 a un 12 por ciento. En la población normal la incidencia resulta ser de un 0.1 por ciento aproximado (McClearn, 1970; Jarvik y otros, 1973). A pesar de lo inadecuado de los datos hay cierta base para creer que un cromosoma Y adicional aumenta tanto las características masculinas como las agresivas. No es difícil extrapolar de este síndrome extremo a influencias genéticas mucho más sutiles del cromosoma Y normal en la conducta a evasiva.
En general, los números anormales de cromosomas sexuales dan como resultado anormalidades de orden tanto físico como conductual. Sería sorprendente lo contrario. La investigación que se propone correlacionar la anormalidad cromosómica con la desviación conductual acaba de empezar y hasta el momento se ha ocuparlo tan sólo de adiciones y remociones cromosomas completos. Existen sin duda alguna, muchas anormalidades a nivel de genes individuales que todavía no logran detectarse con el microscopio.

Característicos conductuales poligánicas Cuando una propiedad o característica presenta en una población una dis-
tribución continua o cuantitativa, ello se debe probablemente a ocurrencias que se presentan en más de un lugar genético. Es un problema poligénico. La genética cuantitativa se ocupa del estudio de tales características. El enfoque básico consiste en determinar el grado de relación o correlación con medidas cuantitativas tales como el CI o las pruebas de personalidad para individuos emparentados entre sí o carentes de tal relación. Las comparaciones más importantes han sido las de gemelos (MZ), las de simples mellizos (DZ) y las de individuos sin parentesco. Los resultados de tales comparaciones indican con suma claridad que rasgos tales como la esquizofrenia, la psicosis maníacodepresiva y la inteligencia son altamente heredables (114cClearn and DeFries, 1973).
Esquizofrenia. Según los datos es cada vez más claro que la esquizofrenia es un mal genético. La esquizofrenia es la forma de enfermedad mental de mayor generalización y gravedad. Se presenta en un 1 por ciento aproximado de la población en todos los países y culturas del mundo, sin importar su carácter avanzado o primitivo. La esquizofrenia se caracteriza por varios grados de desorden en los procesos del pensamiento, por alucinaciones y delirios y desintegración de la personalidad. Aunque no hay acuerdo acerca de los criterios exactos de diagnóstico, la característica más ampliamente compartida es la del pensamiento desordenado. Actualmente no tenemos conocimiento de las posibles anormalidades químicas, nerviosas o biológicas que dan como resultado la esquizofrenia; el hecho es que no hay duda de que existen. Aunque los factores ambientales, culturales y sociales pueden ser importantes, particularmente en el desencadenamiento de episodios esquizofrénicos, la enfermedad es fundamentalmente psicobiológica.
La gran mayoría de los estudios de gemelos idénticos versus fraternos ha indicado una concordancia mucho mayor en la esquizofrenia de los gemelos idénticos. Los valores típicos van de 42 a 86 por ciento para gemelos 9 a 15 por ciento para mellizos fraternos ( McClearn, 1970). Así pues, si un gemelo idéntico resulta con esquizofrenia hay por lo menos iguales posibilidades de que en el otro gemelo aparezca también la enfermedad.
Un estudio más convincente de la heredabilidad de la esquizofrenia en niños adoptados, sin parentesco de por medio, fue realizado por Heston (1966). Identificó 47 niños de madres a quienes habían diagnosticado esquizofrenia en un hospital estatal. Los niños fueron separados de sus madres en los primeros días después de nacidos y colocados en hogares adoptivos. Se utilizó cerro control un grupo de niños adoptados a la misma edad, provenientes de madres libres de esquizofrenia. De los 47 niños de madres esquizofrénicas, 5 llegaron a tener la enfermedad y la mitad presentaron claras inhabilidades psicosociales. El grupo de control no presentó esquizofrenia y la incidencia de dificultades psicosociales resultó de un nivel mucho más bajo.
Aunque hemos caracterizado la esquizofrenia como un desorden causado por anormalidades poligénicas, no todas las autoridades en la materia están de acuerdo con tal definición. Algunos consideran que puede explicarse por un solo gene recesivo; otros creen que representa diversas anormalidades de diferentes genes, diagnosticadas

todas como esquizofrenia; algunos también opinan que es el resultado de defectos de dos o más genes. Finalmente, otros persisten en discutir hasta qué punto la esquizofrenia tiene una base claramente genética (ver McClearn and DeFries, 1973, acerca de tales posibilidades).
Inteligencia. Acalorados debates se han llevado a cabo en los últimos años con relación al grado en que se hereda la inteligencia. Buena parte de la emotividad radica en la afirmación de que existen diferencias raciales genéticas en materia de inteligencia. Estos dos problemas deben separarse. Un repaso a los datos puede conducir solamente a la conclusión de que la inteligencia medida por pruebas típicas de inteligencia, está significativamente determinada por factores genéticos. Por el contrario, no estamos en posibilidad actualmente de interpretar los datos relacionados con la raza y la inteligencia.
En cualquier estudio acerca de la base genética de la inteligencia, deben establecerse ciertos puntos que son evidentes. Primero, la inteligencia es una idea-una construcción teórica que tiene casi tantos significados como autoridades han escrito al respecto. Para medir la inteligencia tenemos que medir algún tipo de ejecución. Las medidas de utilización más generalizadas son las pruebas típicas de CI tales como la Stanford-Binet. La inteligencia medida por tal prueba de CI no es ni más ni menos que un puntaje de prueba. No tiene valor independiente, pero sí tiene relaciones y correlaciones con otras clases de ejecución.
Segundo, el puntaje del Cl es un puntaje general de prueba que "per se" no especifica clases determinadas de habilidades. No es necesariamente un pronóstico de éxito en la vida. Muchas personas de éxito no tienen un CI alto y muchos individuos que sí lo tienen, fracasan estruendosamente. Hay sin embargo una gran relación general entre la ejecución de una prueba de Cl y el desempeño en la vida. Quien tiene un nivel de CI muy inferior al normal no logrará en la mayoría de las situaciones lo que consigue una persona normal. La correlación entre el Cl y los grados escolares es de un 0,5 aproximado-relación significativa pero no extraordinariamente estrecha. La relación general entre el CI y el éxito adaptativo aparece indicada en el estudio de largo alcance acerca de niños superdotados que inició Terman en 1921. Hizo el seguimiento de 1500 niños, todos los cuales tenían un CI de 140 o superior. Como grupo estaban definitivamente por encima del dio en salud, desarrollo, éxito literario y científico, situación económica, y no diferían del promedio en cuanto a salud mental o ajuste emocional.
Tercero, el ambiente influye de manera importante en la ejecución de una prueba de CI. Si un individuo se somete a prueba en diversas ocasiones, durante un período de años, no son raras las fluctuaciones de 20 puntos en el CI. Los niños que pasan de un ambiente pobre a otro mejor presentan puntajes superiores de CI. Una parte importante de la ejecución de una prueba de CI depende de la experiencia cultural y del aprendizaje. Muchos puntos de las pruebas típicas de CI suponen familiaridad con la cultura occidental de la clase tedia. Se supone que los niños están familiarizados con cosas tales como el dinero, los almacenes, los carros y las casas. Otro factor de impor-tancia decisiva es la maduración. Los niños que sufren de desnutrición crónica realizan

las pruebas del CI en forma deficiente. Ninguno de estos factores ha de causar sorpresa. La prueba del CI es una medición de ejecución, la cual está bajo el influjo de todos los factores que inciden en el desempeño humano. Hechas todas estas aclaraciones, ¿tiene algo de sorpresa realmente que los puntajes de pruebas de Cl indiquen heredabilidad altamente significativa? El hecho de indicarlo así sugiere que los factores genéticos desempeñan un papel muy importante en la determinación de la ejecución de las pruebas de CI.
En la Tabla 2.2 aparece un resumen de los datos acerca de la inteligencia de gemelos idénticos (MZ) y fraternos (DZ). Los gemelos idénticos que se crían juntos son idénticos en el desempeño escolar y casi idénticos en CI. La correlación de CI sigue siendo alta cuando se crían separadamente (por lo menos a partir de los seis meses de edad), pero el desempeño escolar es irás variable. Hay la crítica común a tales datos de
Tabla 2.2 Correlaciones de fenotipos de fenotipos físicos y conductuales
Datos tomados de Burt (1966) según se citan en McClearn (1970). Reimpreso con autorización, de "Behavioral Genetics" por G.E. McClearn, Ann. Rev. Genetics 4. Copyright © 1970 por Annual Reviews, Inc. Reservados los derechos.
que los gemelos idénticos MZ criados separadamente van a hogares adoptivos de nivel socioeconómico similar. Burt (1966) hizo la evaluación del caso correlacionando la situación ocupacional de las dos familias en que se criaba cada uno de los gemelos idénticos MZ. La correlación resultó ligeramente negativa. Así pues, la alta correlación de CI respecto de gemelos idénticos MZ, criados separadamente, no puede explicarse por niveles socioeconómicos similares de los hogares adoptivos: no había similitud entre los niveles socioeconómicos.
Según trabajos recientes acerca de la base genética de la inteligencia resulta reforzada la heredabilidad de diferentes clases de habilidades. Las habilidades que, definidas ampliamente, parecen presentar una fuerte determinación genética son las que se miden por las subpruebas de información, vocabulario, ordenamiento de: dibujos, símbolos de dígitos de la Escala de inteligencia de Wechsler para niños (WISC), prueba de CI de amplia utilización. Por el contrario, los resultados de las

subpruebas de Memoria y Razonamiento demostraron que tales habilidades tenían poca o ninguna heredabilidad (Vandenberg, 1967, 1968). Estos resultados se basan en un análisis estadístico cuidadoso de la ocurrencia de una diferencia significativa entre gemelos idénticos MZ y mellizos fraternos DZ en las diversas subpruebas.
En cuanto a los factores genéticos subyacentes, el trabajo de Vandenberg sugiere que hay por lo menos cuatro unidades genéticas diferentes las cuales corresponden a las cuatro habilidades heredables. El número de componentes genéticos varía sin embargo con el tipo de prueba que se utilice. Los cálculos del número total de lugares genéticos implicados en la inteligencia, varían de 3 ó 4 a más de 70. Evidentemente el problema no se ha resuelto. Por el contrario, los puntajes extremadamente altos de Cl pueden estar bajo el control de genes aislados y los CI anormalmente bajos pueden así mismo representar acciones de genes independientes, como en el caso de los niños aquejados de fenilquetonuria.
Un problema de interés potencial para la evolución futura del hombre se relaciona con la posibilidad de que la inteligencia esté declinando en la raza humana. Muchos estudios han demostrado que existe una relación inversa significativa entre el CI y el tamaño de la familia. Aparece la conclusión de que continuamente nacen más individuos de Cl inferior, lo cual conduce a calcular que la declinación de CI por generación alcanza un promedio de 2 a 4 puntos. Dado que la humanidad ha existido en la forma del moderno Homo sapiens durante dos mil ;veneraciones por lo menos, o nuestros antepasados remotos eran increíblemente inteligentes o hay algo errado en la inferencia. De hecho, los datos provenientes de pruebas de largo alcance realizadas con niños no indican disminución alguna del CI. La explicación aparece en la Tabla 2.3. Los individuos de bajo Cl tienen en forma significativa menos niños: muchos de CI extremadamente bajo se hallan hospitalizados desde la niñez y no tienen oportunidad alguna de aparearse. Haciendo cuentas, el Cl de la población como uno todo se presenta relativamente estable.
Importancia de la estructura genética para la conducta La estructura genética influye en cierto número de características conductuales
humanas. En el caso de graves perturbaciones mentales tales como la esquizofrenia o el síndrome de Down, la estructura genética parece desempeñar un gran papel. Pero ¿hasta qué punto explica la variabilidad de las características conductuales humanas en la población normal? Obviamente es imposible una respuesta a esta pregunta pero todas las evidencias indican que su papel puede no ser dominante. En la evolución de los mamíferos, especialmente del hombre, la selección natural ha reforzado la plasticidad conductual. El hombre ha sido seleccionado en parte por razón de su habilidad para cambiar rápidamente su conducta según los cambios ambientales de menor cuantía: Más aún, el hombre recuerda la efectividad de los cambios conductuales. En un sentido muy real, la presión' selectiva sobre el hombre se ha ejercido sobre su habilidad para aprender.

Tabla 2.3 Tasas de Reproducción de sujetos con diferente C.I.
C.I. 0-55 56-70 71-85 86-100 101-115 116-130 131+ Número 29 74 208 583 778 269 25 No. niños Porcentaje 1.4 2.5 2.4 2.2 2.3 2.4 3.0
Datos tomados de Higgins et. al. (1962). Reimpreso con autorización, de "Behaviord Genetics" por G. E. McClearn, Ann. Rey. Genetics 4. Copyright © 1970 por Annual Reviews, Inc. Reservados todos los derechos.
DESARROLLO DEL SISTEMA NERVIOSO
El crecimiento, el desarrollo y el avance físicos se hallan bajo fuerte control genético desde el momento en que el huevo es fecundado hasta que muere el organismo. El ambiente desempeña así mismo un papel. Al nacer, crece bruscamente la importancia del ambiente externo. Con anterioridad a tal evento, el ambiente que rodea a la madre tiene poco efecto sobre el feto en desarrollo, mientras no se presente una catástrofe de proporciones considerables. Pero se considera de -gran importancia el ambiente intrauterino del embrión desde el momento de la concepción, aunque actualmente se sabe poco acerca de su influencia en su desarrollo tanto normal como anormal.
Ocho días aproximadamente después de fecundado, el huevo humano se prende a la pared del útero y se divide en dos células. A partir de este momento se acelera la tasa de multiplicación celular. El embrión se divide en tres capas germinales de células-endodermo, ectodermo y mesodermo. Las células del endodermo forman eventualmente órganos internos, las del mesodermo forman los músculos del esqueleto y las del ectodermo se convierten en piel y sistema nervioso.
El ser humano se denomina embrión durante los dos primeros meses y feto desde ese entonces hasta el nacimiento. El corazón empieza a latir a finales del primer mes, el riñón comienza a excretar orina hacia el tercer mes y alrededor del quinto mes se inician movimientos vigorosos del esqueleto-el feto patea, se retuerce y salta. Los pulmones se desarrollan temprano pero están llenos de fluido y carecen esencialmente de función hasta el nacimiento. Un niño prematuro, con ayuda puede sobrevivir si tiene al menos siete meses y un peso aproximado de cinco libras.
Desarrollo embrionario Diferenciación de las células. El crecimiento y diferenciación del sistema nervioso
ilustra el hecho de que en todas las etapas del desarrollo tanto la estructura como la conducta representan una interacción compleja del conjunto genético y el ambiente. Al empezar la vida de un embrión, las células de los tejidos destinados a convertirse en sistema nervioso son de aspecto semejante al de otras células. Luego, durante el proceso por el cual las células embrionarias se dividen en las capas del endodermo,

mesodermo y ectodermo (Figura 2.8) se forma una estructura de tejido mesodérmico en forma de bastón, inmediatamente debajo de una parte del ectodermo y se alarga desde la cabeza casi hasta el otro extremo del embrión. Este bastón axial, denominado notocordio sirve como operador para la diferenciación de las células ectodérmicas de encima en tejido nervioso. Sin el notocordio las células ectodérmicas permanecen sin diferenciarse. Si debajo de otros tejidos se coloca un trozo de notocordio-por ejemplo, debajo de presuntos tejidos estomacales -empieza a formarse un segundo sistema nervioso. A su vez, un presunto tejido nervioso puede convertirse en tejido estomacal si se le trasplanta en forma apropiada. Por lo tanto, el sistema nervioso en su etapa propiamente inicial depende del ambiente.
Figura 2.8 Desarrollo inicial del canal y el tubo neural en el embrión humano. A) El comienzo del canal neural se forma a partir del envolvimiento del tejido ectodérmico. B) Desarrollo posterior del canal neural. C) El canal neural ha comenzado a cerrarse. D) Continuación del cierre del canal neural, a partir de la línea- media en dirección caudal y rostral. E) Vista lateral del embrión mostrando el cierre completo del tubo neural. (Adaptación de Buchanan, A.R. Functional Neuroanatomy. 4a. ed. Filadelfia: Lea ó: Febiger, 1961).
En esta etapa se dice que las células son indeterminadas en cuanto a que se las puede hacer desarrollar en diversas direcciones. Una vez que tales células entran en contacto con el notocordio, ocurre la determinación y se convierten en tejido nervioso aún en el caso de que se les trasplante. El proceso por el cual el notocordio causa la determinación se denomina inducción: el notocordio induce al ectodermo superpuesto a convertirse en tejido nervioso.
La inducción, aunque al respecto es poco lo que entendemos, se presenta como uno de los fenómenos más fundamentales del desarrollo. Actúa siempre para limitar o especificar las oportunidades de desarrollo de un grupo de células mediante su interacción con otro grupo. Al determinarse más y más, las células se diferencian lentamente en tejidos y sistemas orgánicos que constituyen el organismo adulto. La diferenciación es pues todo este proceso durante el cual un grupo relativamente no especializado de células se moldea en corazón, riñón, estómago, músculo o cerebro.
Etapas del desarrollo. En 1842 Karl Ernst von Baer, teórico evolucionista predarwiniano, estableció el principio de que las primeras etapas de los animales superiores se asemejan a las primeras de los inferiores. Dos décadas más tarde, Ernst Haeckel replanteó este principio con el concepto de que un individuo durante su

ontogenia (crecimiento) atraviesa una serie de formas que se asemejan a sus antepasad -)s adultos; de aquí la expresión "la ontogenia recapitula la filogenia". La frase implica que la historia de la evolución (filogenia) puede estudiarse directamente a través del desarrollo de un organismo individual. La evidencia actual indica que von Baer tenía razón hasta cierto punto: los animales del mismo filum se desarrollan generalmente en forma similar hasta el punto en que se apartan según sus propias especializaciones. Las bolsas branquiarias de un embrión de mamífero nunca se desarrollan hasta llegar a ser branquias funcionales como las de un pez adulto, pero claramente se asemejan a las bolsas branquiarias de un embrión de pez. En forma similar, el desarrollo cerebral de un embrión humano y el de un pez, anfibio o pájaro, guardan entre sí notable semejanza hasta un momento en que cada uno empieza a desarrollar sus propias especializaciones.
Las primeras etapas de la formación del sistema nervioso son perfectamente similares en todos los embriones vertebrados (Figura 2.8 y 2.9). Poco después que el ectodermo superior entra en contacto con el notocordio, una placa nerviosa se convierte en rasgo prominente de la superficie superior. Gradualmente se forma un canal longitudinal en la parte media del embrión y se extiende lentamente hacia ambos extremos mientras se elevan los bordes de la placa. Los bordes se elevan progresivamente y el canal se profundiza hasta que se encuentran y funden sus bordes, formando el tubo nervioso (Fig 2.8). El cierre ¿,el tubo, así la formación del canal . parte media del embrión hacia ambos extremos en forma simultánea. Al cerrarse el tuco, ciertos grupos pequeños de células que hay en los bordes no se incorporan en el tubo y se apartan formando así la cresta nerviosa. En sus primeras etapas, el canal nervioso está constituido por una simple capa de células que se dividen rápidamente. Al cerrarse el tubo nervioso, la división celular ocurre todavía primariamente en el espacio interno del tubo, pero un número creciente de células nuevas empieza a emigrar hacia la parte externa del tubo, formando así tres capas diferentes. La capa interna contiene células que están todavía dividiéndose. La capa media consta de cuerpos celulares que están empezando a formar la materia gris o regiones nucleares del sistema nervioso. En la mayoría de los casos tales células nunca vuelven a dividirse; esta región contiene las mismas células que se encuentran en el organismo adulto. La capa externa permanece temporalmente libre de células y sólo la forman las fibras de las células en las dos zonas restantes. Pronto sin embargo, en algunas áreas del cerebro, dicha capa llega a contener células que han continuado su emigración.
Desde que empieza a formarse la placa nerviosa, es más amplio su extremo anterior (frente) y al cerrarse el tubo aparecen tres combas denominadas las tres regiones cerebrales primarias (Figura 2.9). La región más delantera, el prosencéfalo, da origen a la corteza cerebral y a otras regiones cerebrales superiores (Figura 2.9; ver también el Capítulo 3 donde se explican estos términos anatómicos). El mesencéfalo permanece sin dividirse y da origen al mesencéfalo adulto. El romboencéfalo da origen al cerebelo y regiones inferiores del tallo cerebral.
Por medio del crecimiento diferencial en estas regiones, el cerebro adquiere lentamente su forma adulta. Aún en estas primeras etapas, las partes del cerebro no son completamente independientes entre sí y dependen de las estructuras que las rodean para inducir su diferenciación normal. Por ejemplo, si en las primeras etapas se

Figura 2.9 Desarrollo del cerebro humano desde la tercera semana hasta el nacimiento, vista
lateral. A- En el embrión de tres semanas, las tres regiones primarias del cerebro se encuentran ya formadas. B• En el embrión de cuatro semanas el cerebro anterior, el cerebro medio y el posterior han incrementado su desarrollo. C- Embrión de siete semanas; aquí el cerebro anterior ya se ha diferenciado en telencéfalo y diencéfalo y el cerebro posterior en metencéfalo y mielencéfalo. D- En el feto de once semanas el cerebelo está claramente diferenciado. E- El feto de seis meses presenta un alto desarrollo de los hemisferios cerebrales. F- En el niño recién nacido el surco y el giro están ya bien definidos en los hemisferios cerebrales. (Adaptado de Patten, B.M: Human Embriology, 2a ed. New York: McGraw-Hill, 1953).
remueve completamente la parte posterior, se rota 180 grados y se vuelve a colocar en tal forma que la parte anterior quede en contacto con el cordón espinal y el extremo posterior empalme con el mesencéfalo, se desarrolla un cerebro normal. Estas células pueden ser inducidas por su ambiente a formar regiones nucleares completamente dis-tintas que tengan funciones diferentes.
Mientras la división celular está todavía ocurriendo en forma acelerada en la superficie, las neuronas migratorias empiezan a conformarse en grupos,
constituyendo los núcleos del cerebro adulto. Durante esta fase del desarrollo, la diferenciación se está llevando a cabo aun ritmo acelerado en todo el sistema nervioso. No solamente se están formando los núcleos cerebrales; los tractos de fibras están

igualmente comenzando a conectar los núcleos; algunas de las células de la cresta nerviosa se están conformando en ganglios espinales, enviando sus fibras a la médula espinal y hacia fuera, en dirección a la piel; y los tallos ópticos han crecido fuera del diencéfalo para formar las copas ópticas en sus extremos. Estas copas ópticas inducen la formación de una lente a partir del ectodermo superpuesto. El ojo se forma por lo tanto directamente del tejido cerebral.
Regulación del desarrollo. Gran parte de esta diferenciación aparece autocontrolada. Los movimientos musculares empiezan aun faltando la entrada sen-sorial y los núcleos cerebrales han de madurar aislados de sus respectivos órganos sensoriales motores. Sin embargo, después de establecido el modelo básico del sistema nervioso su periferia adquiere cada vez mayor importancia. Por ejemplo, los centros nerviosos se forman con cierto exceso de células en comparación con lo que se observa en el organismo maduro. Por el tiempo en que una región forma conexiones con la periferia (esto es, órganos y receptores que están fuera del cerebro), se degeneran muchas de las células de tal manera que el número final es el normal. Sin embargo, si se remueve el órgano periférico (por ejemplo, el ojo) se degenera un número todavía mayor de células en tal forma que la región cerebral se atrofia (se contrae); a la inversa, si se amplía en alguna forma el órgano periférico más allá de su tamaño ordinario, se degeneran menos células de lo normal. Este efecto se ha demos-trado en el tallo cerebral y médula espinal respecto de los núcleos tanto motores como sensores en muchos vertebrados.
Es interesante anotar que el momento en que la periferia empieza a tener esta influencia en el desarrollo nervioso aparece en correlación con el momento en que los órganos periféricos se hacen funcionales. Esta observación no permite sin embargo la conclusión de que en todos los casos es necesaria la función para el establecimiento de las conexiones nerviosas. Si se hacen crecer partes del sistema nervioso en cultivo de tejidos, se forman conexiones entre muchas neuronas.
El papel de la actividad funcional en los sistemas sensorial y motor en la regulación del desarrollo futuro del sistema nervioso y de la conducta es actualmente uno de los puntos más controvertidos en la neurología del desarrollo: Según unos, existe primordialmente una relación unidireccional en la cual el desarrollo del sistema nervioso no depende de la retroalimentación proveniente del ambiente; es algo preprogramado para desarrollarse de manera autónoma siguiendo un curso determinado. Según otros, se trata de una relación recíproca en la cual el desarrollo del sistema nervioso es modulado continuamente por estímulos ambientales y el desarrollo normal del sistema nervioso depende de esta modulación.
Víctor Hamburger y sus colegas, experimentando con la conducta de embriones de pollo, han destacado los aspectos autónomos del desarrollo de la conducta. Comenzando por la descripción cuidadosa de los tipos y tasas de movimiento en cada etapa del desarrollo anterior al nacimiento, estos autores lograron describir los efectos de las diversas operaciones. Por ejemplo, removiendo parte de la médula espinal o seccionándola cerca del cerebro, encontraron que cada nivel de la médula puede iniciar conductas en las estructuras corporales asociadas. El principal efecto de estas operaciones era el de reducir la cantidad total de actividad sin cambiar sustancialmente el patrón de movimientos. La influencia de la estimulación sensorial de parte de los

músculos y de la piel se estudió en experimentos en los que se hizo una dea-ferentación. La remoción de las entradas sensoriales de la piel y de los músculos no afecta seriamente los patrones de conducta a partir del tercer día embrionario, tiempo en el que se inicia la conducta hasta el día 17, tres días antes del nacimiento. Aunque estos experimentos no logran eliminar los efectos de la estimulación temprana en los patrones de conducta de la incubación o posteriores a ella, ciertamente muestran que buena parte del desarrollo de la conducta puede ocurrir en ausencia de la estimulación sensorial.
Las influencias hormonales en el desarrollo cerebral y conductual son de importancia decisiva. Varios estudios han demostrado que el exceso de la hormona de la tiroides, la tiroxina, estimula en forma anormal la maduración prematura de algunas áreas nerviosas y el desarrollo anticipado de muchos reflejos. Una cantidad muy reducida de tiroxina al principio del desarrollo puede conducir a efectos devastadores como demora en la maduración de muchas conductas reflejas, crecimiento nervioso reducido, patrones anormales de ondas cerebrales y reducida habilidad de aprendizaje. Los efectos de la tiroxina dependen en su mayoría, del suministro adecuado de la hormona durante un período decisivo específico del desarrollo del organismo (en la rata, los primeros 15 días posteriores al nacimiento). Si por este tiempo la hormona no se halla presente, no pueden remediarse las deficiencias. (Esto contrasta con la situación en la edad adulta. Los déficits conductuales producidos por la remoción de la glándula tiroides en un adulto pueden remediarse por medio de prescripción hormonal.)
La presencia de hormonas sexuales masculinas, andrógenos, durante un período decisivo del desarrollo embrionario, influye en los procesos nerviosos que determinan la aparición de características conductuales masculinas o femeninas. Una de las demostraciones más dramáticas fue realizada por Young y sus colaboradores inyectándoles andrógenos a tres monas durante la preñez. En tres de tales casos la cría genéticamente femenina resultó seudohermafrodita (poseían además de los femeninos, órganos sexuales masculinos incompletos). Tales experimentos se comentarán más ampliamente en el Capítulo 9.
Desarrollo postnatal
Al nacer, el niño posee cierto número de conductas reflejas y un cerebro en situación de funcionamiento amplio si no completo. Sin embargo, los reflejos no se hallan aún bajo el control de la corteza cerebral, la más alta región del cerebro. Por ejemplo, el reflejo de prehensión es muy fuerte por la época del nacimiento: un niño recién nacido puede sostener su peso completo, cogido de una barra. Tal reflejo se debilita considerablemente, sin embargo, un mes más tarde. Un reflejo de fuerza semejante se encuentra en el mono recién nacido, pero conserva su fuerza y aparentemente queda bajo control voluntario unos 20 días después del nacimiento. Tal reflejo es decisivo para la supervivencia de los monos, como también sin duda alguna de los primeros antepasados del hombre. Si el recién nacido no logra prenderse del pelo de su madre mientras cuelga de ella, pierde su posición cuando ella se desplaza y al. caer puede perder la vida.

Las ondas cerebrales generadas en la corteza cerebral (Capítulo 4) pueden registrarse desde el cráneo, que cubre el cerebro del recién nacido. De hecho, las ondas cerebrales del feto pueden registrarse aún desde el abdomen de a :padre. Sin embargo, o, la maduración estructural de la corteza cerebral requiere varios años y la corteza cerebral y otras estructuras cerebrales superiores cambian continuamente durante toda la vida del hombre.
CONTROVERSIA DE NATURA Y NURTURA La psicobiología del desarrollo es el estudio del crecimiento y desarrollo del
cerebro y la conducta. Dado que todos los organismos crecen de células individuales, todos los aspectos de la biología y la conducta tienen una historia del desarrollo. Una investigación completa de la psicobiología del desarrollo supone una revisión total de la psicobiología. En parte por esta razón, muchos psicólogos consideran que de suyo el desarrollo no constituye realmente un campo distinto de estudio. Antes bien, si alguien se interesa por determinado fenómeno tal como el aprendizaje instrumental, el desarrollo de la conducta como resultado del crecimiento y de la experiencia es simplemente parte del asunto.
Muchos psicólogos biológicamente orientados sostienen un punto de vista opuesto; tienden a favorecer la noción de que una parte importante del desarrollo conductual es preprogramada, esto es, que depende del crecimiento y desarrollo del sistema nervioso. Determinadas clases de aprendizaje pueden ocurrir solamente cuando los sustratos cerebrales y corporales apropiados se han desarrollado. Viene al caso el siguiente pasaje de Daniel Lehrman:
Un gatito recién nacido puede rehuir un choque eléctrico y puede presentar una respuesta motora a un sonido; pero los intentos de desarrollar una asociación condicionada simple entre el sonido y la retirada al choque no tienen éxito mientras el animalito no haya cumplido dos semanas. Sería sin embargo prematuro concluir de aquí que el gatito no puede aprender gran cosa durante sus primeros 10 días. Mi colega Jay Rosenblatt, al observar los gatitos con sus madres, notó que cuando aquellos tienen 2 ó 3 días de nacidos, cada gatito se alimenta de determinado pezón de la madre y tiene un camino definido para llegar hasta él. En ciertas circunstancias puede demostrarse que cuando el gatito tiene de 7 a 8 días de edad, ha desarrollado lo que tiene visos de derechos de territorialidad en torno al pezón; muestra cierta tendencia a colgarse del pezón de "su pertenencia" cuando un compañero lo provoca, pero suelta el pezón equivocado si se había prendido a este último.
Esa es pues una clase de aprendizaje de cierta importancia y complejidad, que puede observarse en el gatito durante sus primeros días de vida. La diferencia que existe entre el científico que no puede demostrar aprendizaje en un gatito antes de cumplir éste sus diez días y el que puede demostrarlo cuando el animalito tiene tres días de nacido, es, considero yo, la diferencia que existe entre el investigador que se empeña en saber cuándo puede aprender el gatito algo que el experimentador desea que aprenda y aquél que brega por descubrir cuándo puede aprender el gatito algo que el gatito quiere aprender. (Leherman, 1971, p. 465)

Estos conceptos encontrado, acerca de la importancia del crecimiento y desarrollo tienen sus orígenes en la controversia de natura y nurtura. Los psicólogos conductistas tienden a recalcar la importancia de la experiencia pasada y del aprendizaje; los psicobiólogos destacan el papel del crecimiento y el desarrollo genéticamente controlado del organismo. Muchos estudiantes del desarrollo quisieran hoy enterrar el problema de la natura y nurtura. Así, los organizadores de un importante simposio acerca de la psicobiología del desarrollo (Tobach, Aronson, and Shaw, 1971) esperaban evitar "la conceptualización de natura y nurtura, por ser una falsa dicotomía". Se desanimaron sin embargo con "el problema" cuando se desató durante la conferencia una polémica "acerca de la herencia y el ambiente, lo cual no se había pro-gramado".
La razón de que persista el problema de la natura y nurtura es simple.
Es quizá el más fundamental e importante de todos los interrogantes humanos. Los filósofos y eruditos lo han debatido desde los albores de la historia. Constituye una diferencia de marcada realidad. Si la conducta violenta y agresiva de los humanos es primordialmente aprendida a través de la experiencia, puede controlarse por medio de un aprendizaje adecuado. Si es primordialmente genética, el control social debe ser completamente diferente. Los individuos genéticamente violentos deben ser aislados (por ejemplo, en prisiones) de sus eventuales víctimas. Cualquier rehabilitación social bien intencionada será inútil. La violencia no puede eliminarse de la raza humana sino por medio de técnicas genéticas - por medio de apareamiento selectivo (eugenesia) o, por medio de la ingeniería genética (alteración física directa de los genes, del plasma germinal básico en sí mismo).
La enfermedad mental proporciona otra buena explicación al hecho de que no haya todavía desaparecido la controversia acerca de la natura y la nurtura. Actualmente se favorece el concepto de que muchas formas de neurosis, desde condiciones benignas tales como el excesivo rubor hasta temores graves y persistentes que pueden incapacitar totalmente a una persona, son primordialmente objeto de aprendizaje a través de la experiencia. Presumiblemente pueden tratarse mediante la aplicación de principios y técnicas de aprendizaje. Por otra parte, las psicosis graves se presentan como ampliamente genéticas. El tratamiento y prevención de la esquizofrenia no pueden realizarse únicamente por medio de terapias de aprendizaje. Deben utilizarse métodos biológicos, bioquímicos y finalmente genéticos.
Muchos problemas fundamentales de psicología y psicobiología pueden remontarse al campo de la natura y la nurtura en relación con el desarrollo humano. Esto virtualmente se aplica a todos los procesos psicológicos complejos tales como la percepción, la integración sensoriomotora, los procesos del pensamiento y aún el lenguaje. Un concepto extremista presenta el cerebro del infante como una pizarra limpia, una tabula rasa en que la experiencia escribe lo que será la mente humana. En el extremo contrario está el concepto de que la mente humana del adulto viene preprogramada en el cerebro del infante para desenvolverse y desarrollarse con el cree el siento, independientemente de la experiencia. La verdad como toda verdad, se halla

en algún punto medio de los dos extremos. Exploraremos con mayor amplitud estos problemas del desarrollo en los capítulos pertinentes.
La objeción principal a este problema de la natura y la nurtura es la que lo califica de extremada simplificación. Ningún rasgo humano es completamente genético y ninguno es tampoco fruto exclusivo de la experiencia. Los genes y el ambiente interactúan por lo menos desde el momento en que el huevo es fecundado. Los criminales violentos no son siempre violentos; los esquizofrénicos no andan siempre desorientados. La vida es toda ella, una interacción continua entre el plasma germinal y el mundo. La psicobiología moderna se interesa más por investigar todas las fuentes de la conducta que por escoger entre la natura y la nurtura. El análisis, enfocado a la heredabilidad de las características como lo practica la genética conductual contem-poránea, proporciona una imagen más realista de la importancia relativa de las influencias hereditarias y ambientales y de sus interacciones, en el crecimiento y desarrollo de las habilidades, conducta y personalidad del hombre. Nos encontramos sin embargo muy lejos todavía de entender la base genética aún de conductas tan simples y fijas como el patrón de canto del grillo. No somos capaces todavía de describir la cadena de fenómenos que van desde el gene hasta el canto del grillo y eso para no mencionar la que va del gene a las conductas humanas complejas, como la violencia, la esquizofrenia o la inteligencia.
RESUMEN
Los genes son las unidades estructurales de la herencia. Mediante la determi-nación de las estructuras de las proteínas producidas en los ribosomas, los genes determinan las estructuras y funciones de un organismo. Los genes interactúan así mismo con el ambiente. El genotipo de un organismo expresa su estructura genética subyacente. El fenotipo es la expresión externa de la acción del gene, como resultado de la interacción con el ambiente.
Las tres hipótesis formuladas por Mendel constituyen las leyes básicas de la herencia: (1) la herencia se transmite por unidades independientes y heredables (los genes); (2) cuando cada uno de los padres aporta la misma clase de unidad, los hijos manifiestan así mismo esta característica, pero cuando cada padre aporta una clase diferente de unidad, se forma una cría híbrida; y (3) la experiencia de un organismo no afecta los genes.
El DNA es la sustancia fundamental de los genes. Se compone de desoxirribosa, fosfato y cuatro bases: adenina, guanina, timina y citosina. Watson y Crick propusieron el modelo de doble hélice para la estructura del DNA. La molécula está compuesta de dos ramales, envueltos el uno sobre el otro. Las bases adhieren al sostén de cada ramal y luego se ligan entre sí. Aunque las bases pueden presentarse en cualquier secuencia a lo largo de un solo ramal, hay una relación constante entre los dos ramales complementarios de tal manera que la adenina se liga siempre con la timina y la guanina con la citosina. En dicha secuencia de las bases está contenido el código ge-nético. La capacidad del DNA para reproducirse por sí mismo depende de las combinaciones específicas de ligazón entre las bases.

El mecanismo de traducción del código genético en proteína envuelve RNA. El RNA mensajero transmite el código del DNA al ribosoma. En el ribosoma, el RNA de transferencia actúa para reunir los aminoácidos requeridos para la elaboración de las proteínas codificadas por el DNA..
Los genes para un rasgo específico se presentan en lugares específicos de determinados cromosomas. Cada par de cromosomas contiene lugares homólogos. Un cigote se caracteriza como homocigótico si se hereda el mismo alelo en determinado lugar de ambos cromosomas. Se dice que es heterocigótico cuando dos alelos diferentes se hallan en lugares homólogos.
Varios procesos diferentes contribuyen a la variabilidad de la integración genética. La complementación aleatoria durante la reproducción, la mutación, el entrecruzamiento y la existencia de más de dos alelos en determinado lugar cromosómico, sirven todos para aumentar la variabilidad de la integración genética. Dicha variabilidad genética es definitiva para el desarrollo de especies nuevas y para la supervivencia de una especie en un ambiente de mutación continua.
Si los genes no afectan directamente la conducta, sí sirven para regular la producción de proteínas, las cuales son responsables de las actividades estructurales y enzimáticas de un organismo. La primera preocupación de los científicos que trabajan en el campo de la genética conductual es la de la interacción entre los genes y el ambiente. Estos científicos intentan cuantificar la cantidad de variabilidad observada en un rasgo dentro de una población, como resultante de las diferencias genéticas entre los individuos y la cantidad proveniente de las diferencias ambientales. Las conductas son en su mayoría poligénicas; dependen de muchos lugares cromosomáticos, siendo la contribución de cada, gene pequeña y acumulativa.
Con la utilización de animales se han realizado muchos estudios en situaciones de laboratorio donde el ambiente se mantiene relativamente constante. De esta manera es posible practicar la selección artificial y determinar los efectos de la estructura genética en diversas conductas. Los experimentos de apareamiento selectivo han demostrado que es posible hacer variar muchos rasgos conductuales mediante la manipulación genética. Se han establecido así mismo comparaciones entre familias o razas de animales.
Además de las investigaciones experimentales de los efectos de la manipulación genética en la conducta animal, buena paste de la investigación productiva se ha ocupado de las bases genéticas de la deficiencia mental acentuada. La fenilquetonuria, la idiocia amaurática infantil, la idiocia amaurática juvenil y la corea de Huntington constituyen anormalidades que implican un solo lugar genético. La fenilquetonuria, por ejemplo, se debe a la herencia de un doble gene recesivo que causa una deficiencia en la producción de una sustancia que convierte la fenilalanina en tirosina. Los defectos de la complementación cromosómica producen así mismo deficiencias mentales. Quien tiene el síndrome de Down por ejemplo, porta 47 cromosomas en lugar de los 46 ordinarios y sufre en consecuencia un grave retardo mental. Las anormalidades en los números y clases de cromosomas sexuales afectan también la estructura y personalidad del hombre. El síndrome de Klinefelter, el de Turner y el síndrome XYY se consideran como resultados de anormalidades en los cromosomas sexuales.

Las características poligénicas o sean las producidas por acciones de más de un lugar genético, son de importancia en campos tales como el de la esquizofrenia y la inteligencia. Las características poligénicas se han sometido a estudio mediante la determinación del grado de relación, en medidas cuantitativas de ejecución, entre parientes y no parientes. Las comparaciones de gemelos idénticos MZ y fraternos DZ con individuos libres de parentesco han sido de gran ayuda en el estudio del grado de heredabilidad de diversos rasgos. La evidencia actual indica que la esquizofrenia es de carácter genético. La inteligencia medida por las pruebas típicas de CI, parece deter-minarse también en forma significativa por factores genéticos. Sin embargo, factores ambientales como la experiencia cultural, el aprendizaje y la etapa de maduración, afectan todos la ejecución de pruebas de CI.
Aunque el ambiente desempeña un papel importante después del nacimiento del organismo, el crecimiento y el desarrollo físico permanecen bajo un fuerte control genético desde la fecundación del huevo hasta la muerte del organismo. El desarrollo embrionario del sistema nervioso ilustra la interacción de la acción genética y el ambiente en el desarrollo de la estructura y la conducta. Las células del tejido que se convierte en sistema nervioso son originalmente similares a todas las demás células del embrión. Cuando se forman las tres capas germinales (ectodermo, mesodermo y endodermo) desarrolla así mismo el notocordio. Este determina la diferenciación de las células ectodérmicas en tejido nervioso por un proceso denominado inducción, en el cual el desarrollo de un grupo de células actúa para especificar el desarrollo de otro grupo de células por medio de su interacción.
En las primeras etapas del desarrollo cerebral de los vertebrados se forma cierto número de estructuras. Cuando el ectodermo superior entra en contacto con el notocordio, se desarrolla la placa nerviosa a lo largo de la superficie superior. Con la elevación de los bordes de la placa nerviosa, se forma el tubo nervioso y se constituyen tres capas de células. Al cerrarse el tubo nervioso, se desarrollan las tres regiones cerebrales principales (prosencéfalo, mesencéfalo y romboencéfalo). El prosencéfalo se desarrolla formando la corteza cerebral y otras regiones cerebrales superiores. El mesencéfalo permanece sin dividirse y el romboencéfalo se desarrolla formando el cerebelo y el tallo cerebral. Mediante el crecimiento diferencial en estas tres regiones, adquiere el cerebro su forma adulta.
A través de todo el sistema nervioso ocurre también una diferenciación acelerada. Se constituyen los núcleos cerebrales empiezan los tractos de fibras a conectar los núcleos, de las células de la cresta nerviosa se forman los ganglios espinales y del tejido cerebral se forma el ojo. Gran parte de tal diferenciación parece ser autocontrolada. Sin embargo, una vez establecido el patrón básico del sistema nervioso, adquiere mayor importancia su periferia. Es interesante anotar que cuando la periferia comienza a afectar el desarrollo nervioso, se hacen también funcionales los órganos periféricos.
Las influencias hormonales en el desarrollo cerebral y conductual son de suma importancia. Se ha demostrado que una cantidad excesiva de tiroxina estimula de manera anormal la primera maduración de algunas áreas nerviosas y numerosos reflejos. Una cantidad reducida de tiroxina tiene un efecto negativo en el desarrollo cerebral y conductual. El desarrollo normal depende del suministro adecuado de tiroxina

durante un período decisivo del desarrollo. Si la hormona está ausente durante tal período, no pueden remediarse las deficiencias en época posterior.
Una de las perennes controversias en neuropsicología es la de natura versus nurtura. Desde el punto de vista de la nurtura en su forma extremada, el cerebro del infante es una tabula rasa en que la experiencia construye la mente humana. Desde el extremo contrario arguyen los partidarios de la natura que la mente adulta de los humanos está preprogramada en el cerebro del infante para desarrollarse con el crecimiento, en forma independiente de la experiencia. El rígido contraste de estos dos conceptos constituye una simplificación exagerada, dado que ningún rasgo humano es completamente genético o producto exclusivo de la experiencia. Los genes y el ambiente interactúan desde el momento de la concepción.