two-dimensional electronic spectroscopy and photosynthesis
TRANSCRIPT

Accepted Manuscript
Two-dimensional Electronic Spectroscopy and Photosynthesis: Fundamentals
and Applications to Photosynthetic Light-Harvesting
Gabriela S. Schlau-Cohen, Akihito Ishizaki, Graham R. Fleming
PII: S0301-0104(11)00144-3
DOI: 10.1016/j.chemphys.2011.04.025
Reference: CHEMPH 8200
To appear in: Chemical Physics
Received Date: 4 March 2011
Revised Date: 21 April 2011
Accepted Date: 25 April 2011
Please cite this article as: G.S. Schlau-Cohen, A. Ishizaki, G.R. Fleming, Two-dimensional Electronic Spectroscopy
and Photosynthesis: Fundamentals and Applications to Photosynthetic Light-Harvesting, Chemical Physics (2011),
doi: 10.1016/j.chemphys.2011.04.025
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers
we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and
review of the resulting proof before it is published in its final form. Please note that during the production process
errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.

Tw o-dimensional Electronic Spectroscopy and Photosynthesis: F undamentals and
Applications to Photosynthetic Light-Harv esting
Gabriela S. Schlau-Cohen, Akihito Ishizaki, and Graham R. Fleming∗
Department of Chemistry, University of California, Berkeley, CA 94720, USA and
Physical Biosciences Division, L awrence Berkeley National L aboratory, Berkeley, CA 94720, USA
In natural light harvesting systems, pigment-protein complexes are able to harvest sunlight withnear unity quantum efficiency. These complexes exhibit emergent properties that cannot be simplyextrapolated from knowledge of their component parts. In this Perspective, we focus on how two-dimensional electronic spectroscopy (2DES) can provide an incisive tool to probe the electronic, en-ergetic, and spatial landscapes that must be understood to describe photosynthetic light-harvesting.W e review the theoretical and experimental principles of 2DES, and demonstrate its application tothe study of the Photosystem II supercomplex of green plants. W e illustrate several capabilities of2DES, including monitoring energy transfer pathways, observing excitonic coherence, determiningexcitonic geometry, and informing on the atomic structure.
I. INTR ODUCTION
The development of a unified theoretical descriptionof ultrafast nonlinear spectroscopic measurements byMukamel [1] spurred a large and continuing burst ofexperimental development focused on dynamics of con-densed phase systems. In turn, the results of these ex-periments have stimulated new theoretical approaches,particularly to condensed phase quantum dynamics [2–10]. Nowhere has this synergy been more evident than instudies of photosynthetic light harvesting, where increas-ingly sophisticated ultrafast spectroscopies have beenemployed. These advances have culminated in the useof two-dimensional electronic spectroscopy (2DES) [11–16], following the impressive developments in 2D infraredspectroscopy pioneered by Hochstrasser and coworkers[17]. In this perspective, we describe the theory underly-ing 2DES and its analysis and illustrate the applicationof these ideas to understanding complicated condensedphase systems. W e use the Photosystem II (PSII) su-percomplex of green plants [18, 19], as an example todescribe how 2DES can, by probing structure-functionrelationships, provide an incisive probe of the overall de-sign of this natural device. W e close with some thoughtson directions for future developments in this area.
The PSII supercomplex is a particularly impressivenatural apparatus which functions with remarkable nearunity quantum efficiency at low light levels [22]. Compo-nents varied in size and form combine to harvest sunlightand then use every single photoexcitation to power chem-ical reactions [22, 23]. As shown in the structural modelin Fig. 1a, the PSII supercomplex consists of several as-sociated pigment protein complexes (PPCs), which con-sist of chromophores held in a protein matrix [18, 19].Initial absorption events primarily occur on the periph-ery of the supercomplex in arrays of the major light-harvesting complex of PSII (LHCII). The excitation then
∗Electronic address: [email protected]
a) b)
c)
LHCII
reaction
CP29
center
14,500 15,000
Energy (cm -1)
0.50
0.0015,500
Intensity (au) 1.00
d)LHCIICP29
Linear A bsorption at 77 K
FIG. 1: Structural models of the light-harvesting pigmentprotein complexes from green plants where (a) shows a modelof the Photosystem II supercomplex, (b) a structural modelfrom x-ray crystallography [20] of LHCII, and (c) of the minorcomplex, CP29 [21]. (d) The linear absorption spectra ofLHCII and CP29 at 77 K.
translocates to the reaction center where it drives photo-chemistry. During its migration from the antenna to thereaction center, the excitation travels through so-calledminor complexes, such as chlorophyll-binding protein 29(CP29). Experimental evidence suggests that CP29, aswell as participating in energy transfer processes, playsa role in non-photochemical quenching (NPQ) [24, 25],

2
which is the process by which plants and algae dissi-pate excess energy. The structural models of LHCII andCP29 are shown in Fig. 1b and c, respectively. Thesecomplexes, which complete efficient energy transfer, ex-hibit processes, such as relaxation of the phonon bathand structural fluctuations, which occur on timescalesfrom femtosecond to hours. This combination of dynam-ical timescales and moleclar rearrangements mean thatit is difficult to identify the molecular origin of any givenprocess.
PPCs are systems with emergent properties very differ-ent from those of their individual molecules. Their elec-tronic absorption spectra are congested, generally with-out structure, and result from interactions of the chro-mophores (generally chlorophylls (Chl)) with this pro-tein environment and with neighboring chromophores.The resultant states (often called excitons) are not lo-calized on individual chromophores, interact weakly withtheir environment, exhibit ultrafast electronic relaxationwithin the exciton manifold, and exhibit long-lived elec-tronic quantum coherence [16, 22, 26]. These proper-ties appear to be essential for achieving the near unityquantum efficiency that photosynthetic light harvestingachieves at low light levels [22]. At light levels in ex-cess of that usable for photosynthetic energy conversion,green plants and algae have complex mechanisms for re-ducing the quantum efficiency of energy transfer throughthe antenna to the reaction center [25]. The precise mole-cular mechanisms of this regulatory process is still underachieve investigation, but undoubtedly involves, amongother factors, the dynamic properties listed above.
These dynamic properties often render conventionalspectroscopic techniques such as steady state and timeresolved absorption and emission spectroscopy insuffi-ciently incisive to reveal the microscopic mechanisms,and connections between the spatial, electronic, and dy-namic landscapes which underlie function. 2DES offers asignificantly more detailed window into the microscopicbasis of light harvesting efficiency for the following rea-sons: (1) by spreading out the time-evolution of thespectrum onto two dimensions spectral resolution is en-hanced and pathways of relaxation processes are directlyrevealed, (2) because the spectrum is recorded at the am-plitude, rather than the intensity, level 2DES is directlysensitive to the quantum phase of the system therebyproviding direct evidence of quantum coherence, (3) thepolarizations of the four light pulses involved can be inde-pendently set providing the ability to connect the atomicstructure (determined by x-ray crystallography, for ex-ample) with the electronic structure through the anglesbetween the transition dipole moments. In addition infavorable cases, insight into the unknown atomic struc-ture can be obtained and the uncoupled site energies ofindividual chlorophyll molecules can be estimated.
II. ELECTRONIC ENERGY TRANSFER IN
PHOTOSYNTHETIC PIGMENT-PROTEIN
COMPLEXES
To describe photosynthetic excitation energy transfer(EET), we consider a PPC consisting of N pigments. Inthis Perspective, we restrict the electronic spectrum ofthe mth pigment to the ground state |ϕmg〉 and the firstexcited state |ϕme〉 which corresponds to the Qy transi-tion in a chlorophyll molecule. These states are obtainedby the electronic Schrodinger equation Hel
m(X)|ϕma〉 =Ema(X)|ϕma〉, where Hel
m(X) and Ema(X) are the elec-tronic Hamiltonian and energy of the mth pigment inthe absence of the inter-pigment Coulomb interaction,and depend parametrically on the set of the relevantnuclear coordinates including protein degrees of free-dom, X. Hence, the nuclear dynamics associated withan electronic state |ϕma〉 are described by Hma(X) ≡Ema(X) + (kinetic energy). The normal mode treat-ment is usually assumed for the PPC nuclear dynamics,because anharmonic motion with large amplitudes andlong timescales produces inhomogeneous broadening ontimescales irrelevant to photosynthetic EET. Such slowmotions are, however, often significant for protein func-tionality. Further, it may be assumed that nuclear config-urations for the electronic excited states of pigments arenot greatly different from those for the ground states ow-ing to the absence of large permanent dipoles on the pig-ments. Thus, the nuclear Hamiltonians associated withthe electronic states |ϕmg〉 and |ϕme〉 can be modeled asa set of displaced harmonic oscillators [16, 27],
Hmg(X) = Emg(X0mg) +
∑
ξ
~ωmξ
2(p2
mξ + q2mξ), (2.1)
Hme(X) = Hmg(X) + ~Ωm −∑
ξ
~ωmξdmξqmξ, (2.2)
where qmξ are the dimensionless normal mode coor-dinates introduced around the equilibrium nuclear con-figuration associated with the mth pigment X0
mg, andωmξ and pmξ are the accompanying frequencies andmomenta. In what follows we set Emg(X
0mg) = 0 to sim-
plify our expressions. ~Ωm is the Franck-Condon transi-tion energy, which is also termed the site energy.
In order to characterize the possible states of the wholecomplex of excitable units, product states are introducedas
∏m|ϕmam
〉. This Hartree-like ansatz is reasonableonly if different state vectors |ϕma〉 do not overlap andthe exchange interaction is negligible. For later conve-nience, we order the product state with respect to thenumber of elementary excitations. The overall groundstate with zero excitation reads |0〉 ≡
∏m|ϕmg〉, whereas
the presence of a single excitation at the mth pigment isdescribed by |m〉 ≡ |ϕme〉
∏k( 6=m)|ϕkg〉. Description of
EET on the basis of |m〉 is termed the site representa-
tion. The corresponding expansion of the complete PPC

3
Hamiltonian yields
HPPC = H(0)PPC + H
(1)PPC + · · · , (2.3)
where H(n)PPC (n = 0, 1, . . . ) describes n-excitation man-
ifold comprising n elementary excitations. The Hamil-tonian of the zero-excitation manifold reads
H(0)PPC =
∑
m
Hmg(X)|0〉〈0|, (2.4)
whereas the Hamiltonian of the single-excitation mani-fold takes the form,
H(1)PPC =
∑
m
Hm(X)|m〉〈m| +∑
mn
~Jmn|m〉〈n| (2.5)
with Hm(X) ≡ Hme(X) +∑
k( 6=m) Hkg(X). The elec-
tronic coupling ~Jmn has to be deduced from the inter-pigment Coulomb interaction [28–32]. The electroniccoupling may be also modulated by nuclear motions [33].In the following, however, we assume that nuclear de-pendence of ~Jmn is vanishingly small and employ theCondon-like approximation as usual. Since the inten-sity of sunlight is weak, the single-excitation manifoldis of primary importance under physiological conditions.However, nonlinear spectroscopic techniques such as 2Delectronic photon echo spectroscopy can populate higherexciton manifolds, e.g. the double-excitation manifoldcomprising |mn〉 ≡ (1 − δmn)|ϕme〉|ϕne〉
∏
k( 6=m,n)|ϕkg〉.
The Hamiltonian for the double-excitation manifolds isexpressed as
H(2)PPC =
∑
mn
Hmn(X)|mn〉〈mn|
+∑
mnk
(
~Jmk|mn〉〈kn| + ~Jmk|mn〉〈mk|)
(2.6)
with Hmn(X) ≡ Hme(X) + Hne(X) +∑
k 6=m,n Hkg(X).
From the dynamical point of view, Eqs. (2.1) and (2.2)demonstrate that the electronic energies of the pigmentsexperience modulation by the protein motion. Due to thelarge number of protein degrees of freedom, such dynam-ical modulation can be modeled as random fluctuations.In order to describe fluctuations in electronic states, weintroduce the collective energy gap coordinate [1],
um ≡ Hme(X) − Hmg(X) − ~Ωm = −∑
ξ
~ωξdmξqmξ.
(2.7)
Because qmξ are normal mode coordinates or phonon
modes, the dynamics of um(t) ≡ eiHmgt/~ume−iHmgt/~
can be described as a Gaussian process [34]. There-fore, all environment-induced relaxation processes can bequantified by two-body correlation functions of um(t).Fluctuations in the electronic energy of the mth pigment
Environmental configuration
FIG. 2: Schematic illustration of the electronic ground and
excited states of the mth pigment, |ϕmg〉 and |ϕme〉, af-
fected by nuclear motion of the protein environment. After
electronic excitation in accordance with the vertical Franck-
Condon transition, reorganization takes place from the equi-
librium nuclear configuration with respect to the electronic
ground state |ϕmg〉 to the actual equilibrium configuration in
the excited state |ϕme〉 with dissipation of the reorganization
energy ~λm. This reorganization proceeds on a characteristic
timescale τ rxn
m .
are described by the symmetrized correlation function ofum(t),
Sm(t) ≡1
2〈um(t)um(0) + um(0)um(t)〉mg . (2.8)
where 〈. . . 〉mg denotes averaging over ρeqmg ≡
e−β Hmg/Tr[e−β Hmg ] with β being inverse tempera-ture. After electronic excitation in accordance witha vertical F ranck-Condon transition, reorganizationof the nuclear configuration takes place toward theequilibrium configuration in the excited state withdissipation of the reorganization energy expressed as~λm =
∑ξ ~ωmξd
2mξ/2 on a finite timescale, τ rxn
m . Thisreorganization dynamics can be characterized by theresponse function of um(t),
χm(t) ≡i
~〈[um(t), um(0)]〉mg . (2.9)
The reorganization process, in principle, can be measuredby the time-dependent fluorescence Stokes shift experi-ment [16, 35], where the direct observable quantity is therelaxation function defined by
Γm(t) =
∫∞
t
ds χm(s). (2.10)
The reorganization energy ~λm, which can be measuredvia the Stokes shift magnitude, is given as Γm(0) =2~λm, and the characteristic timescale of the reorgani-zation dynamics is given as τ rxn
m =∫∞
0dt Γm(t)/Γm(0).
When the environmental degrees of freedom can bedescribed classically, the symmetrized correlation, re-sponse, and relaxation functions satisfy the classicalfluctuation-dissipation relation as [34]
Sm(t) =1
βΓm(t), χm(t) = −
d
dtΓm(t). (2.11)

4
We note that the quantum fluctuation-dissipation rela-tion [34] needs to be employed for low-temperature sys-tems where the classical fluctuation-dissipation relationbreaks down. These two-point correlation functions playa significant role in analyses of condensed phase laserspectroscopic signals such as 2D spectra, as will be dis-cussed later.
The electronic coupling ~Jmn, the reorganization en-ergy ~λm, and the characteristic timescale of the reor-ganization dynamics τ rxn
m are three fundamental quan-tities determining the nature of EET in photosyntheticPPCs. Ordinarily, photosynthetic EET is discussed onlyin terms of the mutual relation between magnitudes ofthe electronic coupling and the reorganization energy. Ifthe electronic coupling is small in comparison with thereorganization energy, then the electronic coupling canbe treated perturbatively. This treatment yields Forstertheory [36]. In the opposite limit, it is possible to describethe interaction between electronic excitation and the en-vironment, which is characterized by the reorganizationenergy, in a perturbative manner to derive quantum mas-ter equations such as Redfield theory [37]. However, weshould not overlook that the nature of EET is also depen-dent on the mutual relation between two timescales, τ rxn
m
and J−1mn [16, 33]. The inverse of the electronic coupling,
J−1mn, defines the time an electronic excitation needs to
move from one pigment to another. As was emphasizedin Refs. [6, 16, 38], the relation between J−1
mn and τ rxnm is
more fundamental for the nature of EET than that be-tween ~Jmn and ~λm. In the case of τ rxn
m ¿ J−1mn, it is
difficult to construct a wave function straddling multiplepigments. The environmental reorganization introducesfast equilibration of the environmental configurations,and hence EET occurs after the environmental equili-bration associated with the excited pigment. In this sit-uation, EET is described as a diffusive motion similar toclassical random walk; it follows classical rate laws wherethe transition rate is given by Forster theory. In the op-posite case of J−1
mn ¿ τ rxnm , the excitation can travel al-
most freely from one pigment to others through a photo-synthetic complex as a quantum mechanical wave packetkeeping its phase coherence. Thus, this EET process istermed coherent energy transfer. Obviously, there ex-ists a regime of EET where the two coupling magnitudesand/or the two timescales compete against one another,i.e. λm ∼ Jmn and/or τ rxn
m ∼ J−1mn. This intermediate
regime is typical for photosynthetic EET [11, 16, 31]. Inthis situation, a distinct characterization is not straight-forward because characteristics of both coherent energytransfer and diffusive motion coexist. In order to addressthe issue, two of the authors developed a formally exactsolution from the second-order cumulant time-nonlocalquantum master equation which reduces to the conven-tional Forster and Redfield theories in their respectivelimit of validity [6]. In the regime of coherent wavelikemotion the equation predicts several times longer lifetimeof quantum coherence between electronic excited states ofthe pigments than does the conventional Redfield equa-
tion. Reference [6] demonstrated that quantum coherentmotion can be observed even when reorganization energyis large in comparison to intersite electronic coupling,although this situation has often been regarded as theregime of incoherent hopping diffusive motion. This willbe discussed in more detail in Sec. V C.
One of the viable approaches to observe EET dynam-ics in photosynthetic PPCs is optical spectroscopy. Asmolecular systems become increasingly complex, con-ventional optical spectroscopic techniques such as time-or frequency-resolved absorption spectroscopy or sponta-neous emission spectroscopy become of limited use for ex-traction of direct information on the molecular propertiessuch as site energies, ~Ωm, electronic couplings, ~Jmn,environmental reorganization energies, ~λm, timescalesof environmental reorganization dynamics, τ rxn
m , andthe EET dynamics induced by these quantities in pho-tosynthetic PPCs. This is because conventional one-dimensional (1D) spectroscopies, which project all theinformation onto a single time- or frequency-axis, arenot capable of separating the variety of structural anddynamic contributions present on the 1D axis. In orderto disentangle the 1D signals, one needs to explore mul-tidimensional observables, which project the structuraland dynamic information onto multiple axes. In this re-gard, 2DES, based on the heterodyne-detected four-wavemixing technique, can provide us with far more detailedinformation on photosynthetic PPCs.
III. F OUNDATIONS OF TW O-DIMENSIONAL
ELECTR ONIC SPECTR OSCOPY
A. Nonlinear optical responses
In third-order four-wave mixing techniques, the incom-ing electric field can be written in the form [39, 40],
E(t) =3∑
i=1
EiAi(t − τi)e−iωi(t−τi)+iki·r+iφi + c.c.,
(3.1)
where the laser pulse centered at time τi (τ1 ≤ τ2 ≤τ3) is linearly polarized along the direction characterized
by the unit vector Ei. In Eq. (3.1), ωj , kj , Ai(t) andφi represent the carrier frequency, wavevector, temporalenvelop, and phase of the ith field, respectively. Thethird-order polarization per unit volume, P
(3)(t), can beexpressed as [1]
P(3)(t) = nPPC
∫∞
0
dt3
∫∞
0
dt2
∫∞
0
dt1Φ(t3, t2, t1)
× E(t − t3)E(t − t3 − t2)E(t − t3 − t2 − t1), (3.2)
with nPPC being the number density of PPCs.Φ(t3, t2, t1) is a forth-rank tensor indicating the third-

5
order response function expressed as
Φ(t3, t2, t1) =
(
i
~
)3
× 〈[[[µ(t1 + t2 + t3), µ(t1 + t2)], µ(t1)], µ]ρeqPPC〉, (3.3)
where µ(t) ≡ eiHPPCt/~µe−iHPPCt/~ has been introduced.In what follows we ignore ~
3 in the denominator andnPPC for simplicity of the expression. Due to the three-times commutations, the third-order response functioncontains 23 = 8 contributions (23 = 8 different Liouvillespace pathways),
Φ(t3, t2, t1) = i34
∑
α=1
[Rα(t3, t2, t1) − R∗
α(t3, t2, t1)] (3.4)
with [1]
R1(t3, t2, t1) = F (t1, t2 + t1, t3 + t2 + t1, 0), (3.5a)
R2(t3, t2, t1) = F (0, t2 + t1, t3 + t2 + t1, t1), (3.5b)
R3(t3, t2, t1) = F (0, t1, t3 + t2 + t1, t2 + t1), (3.5c)
R4(t3, t2, t1) = F (t3 + t2 + t1, t2 + t1, t1, 0), (3.5d)
where we have defined the four-point correlation functionof µ(t) as
F (τ4, τ3, τ2, τ1) = 〈µ(τ4)µ(τ3)µ(τ2)µ(τ1)ρeqPPC〉. (3.6)
The macroscopic polarization in Eq. (3.2) generateselectric fields in the phase-matching directions of ks =±k1 ± k2 ± k3,
Esig(t) ∝ iP (3)(t). (3.7)
In the heterodyne detection scheme, an additional elec-tric field called the local oscillator is applied, ELO(t)eiφLO
with φLO being a controllable phase parameter, and thenthe detected signal is given as [1]
S =
∫
∞
−∞
dtE∗
LO(t) · Esig(t). (3.8)
Here we consider the local oscillator to be an impulsivefield E∗
LO(t) ' ELOδ(t − tm), where the pulse centeredat time t = tm is linearly polarized along the directioncharacterized by the unit vector ELO. Thus, the detectedsignal can be a function of a time variable as
S(tm) = ELO · iP (3)(tm). (3.9)
B. 2D electronic photon echo spectroscopy
Photon echo (PE) spectroscopy is an optical coun-terpart of the spin echo in nuclear magnetic resonance(NMR) spectroscopy [35, 41–47]. Originally based onpump-probe, i.e. dynamic hole-burning spectroscopy[17], this is one of the most widely used 2D in-frared/electronic spectroscopic techniques. Using the
above third-order response function theory, one can for-mulate the photon echo polarization and signal elec-tric field. Typically photon echo spectroscopy mea-sures the echo signal field emitted in the direction ofkI = −k1 + k2 + k3 for the incident laser pulses in Eq.(3.1). In what follows time zero is set at the center ofthe third pulse, i.e. τ3 = 0. It is customary to define thecoherence period τ as the separation between the centersof the first two pulses, τ ≡ τ2 − τ1 ≥ 0, and the waitingperiod T as the difference between the second and thirdpulses, T ≡ −τ2 ≥ 0 to ensure pulse ordering.
The expression for the incident laser pulsed in Eq.(3.1) is fairly general for an arbitrary nondegenerate pho-ton echo spectroscopy. Two different frequencies ω1 =ω3 6= ω2 were employed for a two-color electronic coher-ence photon echo experiment [14], while ω1 = ω2 6= ω3
was adopted for the two-color three pulse photon echopeak shift experiment [48–52]. In addition, two-color2D infrared/electronic photon echo spectroscopies wereimplemented [53, 54]. However, most photon echo ex-periments have been performed by using pulses with thesame center frequency ω1 = ω2 = ω3 ≡ ω0 (the degen-erate limit). The degenerate photon echo polarizationPPE is dependent on the controllable parameters (T, τ)in addition to t, and then is given as
PPE(t, T, τ) = e−iω0t+iω0τ
∫
∞
0
dt3
∫
∞
0
dt2
∫
∞
0
dt1
× Φ(t3, t2, t1)E3A3(t − t3)E2A2(t + T − t3 − t2)
× E1A∗
1(t + T + τ − t3 − t2 − t1)eiω0t3−iω0t1 , (3.10)
where the wavevector part, exp[i(k3 + k2 − k1) · r], andthe phase part, exp[i(φ3 + φ2 − φ1)], have been ignoredfor simplicity of the expression. In Sec. IV, the phasepart will be discussed in more detail.
The third-order response function consists of eight dif-ferent contributions as shown in Eqs. (3.4) and (3.5),each of which has a different coherent oscillating pat-tern in time. If the laser frequency ω0 approximatelymatches an electronic transition frequency in the PPCof interest, some of the phase factors originating fromthe response function may cancel with those originatingfrom the laser field. For instance, the coherence oscilla-tion term of R2(t3, t2, t1) is exp(−iΩegt3 + iΩegt1) withΩeg being a characteristic optical transition frequency ofthe PPC. Therefore, this contribution cancels with theelectric field component oscillating as exp(iω0t3 − iω0t1)when ω ' Ωeg. Thus, under the integration in Eq. (3.10)we would have slowly varying terms where oscillatoryfactors canceled and fast oscillating terms where phasefactors added. After the integration, oscillatory termsresult in a much smaller contribution than slowly vary-ing ones, and we can neglect them. This is usually re-ferred to as rotating wave approximation (RWA). In Eq.(3.10) under the RWA, the surviving components amongthe eight different response function contributions areonly R2(t3, t2, t1), R3(t3, t2, t1) and R∗
1(t3, t2, t1), whichare classified to be the rephasing pathways. Thus, the

6
rephasing echo signal SR(t, T, τ) ≡ ELO·iPPE(t, T, τ > 0)can be expressed as
SR(t, T, τ) = [R2(t3, t2, t1) + R3(t3, t2, t1)
− R∗
1(t3, t2, t1)]ELOE3E2E1, (3.11)
where we have employed the impulsive limit of Ai(t −τi) ' δ(t − τi). In the above, the rephasing photon echospectroscopy was discussed, where the echo signal fieldpropagating in the direction of kI = −k1 + k2 + k3 wasmeasured. One can also selectively measure the termsR1, R4, and R∗
2 with coherence oscillation exp(−iΩegt3−iΩegt1) by focusing on the emitted signal in the direc-tion of kII = k1 − k2 + k3 instead. This has beencalled the nonrephasing photon echo (NR-PE) measure-ment. The degenerate nonrephasing photon echo signalSNR(t, T, τ) ≡ ELO · iPNR-PE(t, T, τ > 0) in the impulsivelimit is then given as
SNR(t, T, τ) = [R1(t, T, τ) + R4(t, T, τ)
− R∗
2(t, T, τ)]ELOE3E2E1. (3.12)
Experimentally, one does not detect the polarizationsof Eqs. (3.11) and (3.12) directly as a function of tbut instead with a spectral interferogram as a func-tion of the conjugate frequency ωt, i.e. SR(ωt, T, τ) andSNR(ωt, T, τ). Mathematically, however, there are threepossibilities of double Fourier-Laplace transform of theecho field by choosing a pair of time variables from τ ,T , and t. The most popular choice has been to take τand t variables for the transform, and thus 2D rephasingspectrum and 2D nonrephasing spectrum are given as
SR(ωt, T, ωτ ) =
∫
∞
0
dt eiωtt
∫
∞
0
dτ e−iωτ τSR(t, T, τ),
(3.13)
SNR(ωt, T, ωτ ) =
∫
∞
0
dt eiωtt
∫
∞
0
dτ eiωτ τSNR(t, T, τ),
(3.14)
which allow us to explore electronically excited statesin photosynthetic PPC and their dynamics during thewaiting time T .
In practice, the rephasing and nonrephasing echo sig-nals can be detected in the same direction −k1 +k2 +k3
by exchanging the time-ordering of the first two inci-dent pulses, k1 and k2, in Eq. (3.1). By redefiningT ≡ min(−τ1,−τ2) ≥ 0 and τ ≡ τ2 − τ1, PPE(t, T, τ >0) is the rephasing photon echo polarization whereasPPE(t, T, τ < 0) corresponds to the nonrephasing photonecho polarization. Therefore, a 2D photon echo spectrumcan be obtained as [39, 40, 55]
SPE(ωt, T, ωτ )
=
∫
∞
0
dt eiωtt
∫
∞
−∞
dτ e−iωτ τ ELO · iPPE(t, T, τ) (3.15)
for positive ωt and ωτ . It should be noted that the lower-limit of the τ -integration is −∞ instead of 0. This 2Dphoton echo spectrum is identical to 2D correlation spec-trum defined as the sum of the 2D rephasing and non-rephasing spectra [56, 57],
SPE(ωt, T, ωτ ) = SR(ωt, T, ωτ ) + SNR(ωt, T, ωτ ). (3.16)
C. 2D lineshape of a monomer
In order to clarify the information contained in 2Drephasing, nonrephasing, and photon echo spectra, it isadvisable to begin by examining the 2D lineshapes of amonomer. Because of the Gaussian process of the collec-tive energy gap coordinate in Eq. (2.7), the four-bodycorrelation function given in Eq. (3.6) for the mth pig-ment can be written as [1]
F (τ4, τ3, τ2, τ1)ELOE3E2E1
= e−iΩm(τ4−τ3+τ2−τ1)+fm(τ4,τ3,τ2,τ1), (3.17)
where we have ignored the constant prefactor, and theauxiliary function fm has been introduced as
fm(τ4, τ3, τ2, τ1) = −gm(τ4−τ3)+gm(τ4−τ2)−gm(τ4−τ1)
− gm(τ3 − τ2) + gm(τ3 − τ1) − gm(τ2 − τ1), (3.18)
in terms of the linebroadening function,
gm(t) =1
~2
∫ t
0
ds1
∫ s1
0
ds2
[
Sm(s2) − i~
2χm(s2)
]
.
(3.19)
To calculate the linebroadening function, we employthe overdamped Brownian oscillator model Γm(t) =2~λme−t/τ rxn
m , which has been successfully applied fortheoretical analyses of experimental results [2, 7, 12, 58,59]. From the classical fluctuation-dissipation relationin Eq. (2.11), the symmetrized correlation and responsefunctions are evaluated as Sm(t) = ~
2∆2me−t/τ rxn
m and
χm(t) = (2~λm/τ rxnm )e−t/τ rxn
m , where ~∆m = ~√
2λm/~βstands for the root-mean-square amplitude of the elec-tronic energy fluctuations of the mth pigment. Conse-quently, the linebroadening function can be expressed inthe form,
gm(t) =[
(∆mτ rxnm )2 − iλmτ rxn
m
]
(
e−t/τ rxn
m + t/τ rxnm − 1
)
.
(3.20)
For simplicity, we focus on a situation where τ rxnm is much
faster than the timescale of interest. In this Markov limit,Eq. (3.20) is approximated as a linear function of timevariable,
gm(t) '(
∆2mτ rxn
m − i λm
)
t, (3.21)

7
and thus the rephasing echo signal in the impulsive limitare expressed as
SR(t, T, τ) = 2e−iΩmt−gm(t)eiΩmτ−g∗
m(τ), (3.22)
where the R∗
1 Liouville space pathway does not con-tribute for the case of a two-level transition. It should benoted that t-, T -, and τ -dependences are separable onlywhen the linebroadening function is expressed as a linearfunction of the time variable. This time-domain signalis converted into a complex 2D spectrum by a doubleFourier-Laplace transform with respect to t and τ . Byseparating the absorptive and dispersive contributions,the 2D spectrum is expressed as
SR(ωt, T, ωτ ) = 2[a(ωt)a(ωτ ) + d(ωt)d(ωτ )]
− i 2[d(ωt)a(ωτ ) − a(ωt)d(ωτ )] (3.23)
in terms of the absorption and dispersive components,
a(ω) =∆2
mτ rxnm
(ω − Ω0m)2 + (∆2
mτ rxnm )2
, (3.24)
d(ω) =−ω + Ω0
m
(ω − Ω0m)2 + (∆2
mτ rxnm )2
, (3.25)
respectively, where Ω0m = Ωm −λm has been introduced.
See Fig. 2. The real part of SR(ωt, T, ωτ ) has a ‘mixedphase’ [41] because it consists of a superposition of a pure2D absorptive peak and a pure 2D dispersive peak. This2D lineshape is referred to as phase-twist in the literatureof 2D NMR [60]. It should be noticed that the decayof the dispersive components is much slower than thatof the absorptive components, and thus it is ambiguousto extract accurate information from the phase-twistedlineshapes of 2D rephasing spectra for complex systemssuch as photosynthetic PPCs. It is difficult to distinguishabsorptive features from dispersive contributions becausethe phase-twisted lines sometimes look like absorptive off-diagonal peaks in multi-component 2D spectra, as shownin Fig. 3. Suppression of the dispersive component istherefore desirable for improved resolution. On the otherhand, the nonrephasing echo signal in the impulsive limit,Eq. (3.12), is expressed as
SNR(t, T, τ) = 2e−iΩmt−gm(t)e−iΩmτ−gm(τ), (3.26)
which delivers the 2D nonrephasing spectrum as
SNR(ωt, T, ωτ ) = 2[a(ωt)a(ωτ ) − d(ωt)d(ωτ )]
− 2i [d(ωt)a(ωτ ) + a(ωt)d(ωτ )]. (3.27)
The real part of the 2D nonrephasing spectrum is alsocontaminated by the phase-twisted lineshape due to thedispersive component. Comparing the rephasing andnonrephasing spectra in Eqs. (3.23) and (3.27), how-ever, we can observe that the dispersive components inthe real parts of the two spectra exactly cancel out, andthus we arrive at
SPE(ωt, T, ωτ ) ∝ a(ωt)a(ωτ ) − i d(ωt)a(ωτ ). (3.28)
(a) rephasing (b) nonrephasing (c) photon echo
FIG. 3: Characteristic lineshapes of 2D rephasing, nonrephas-ing, and photon echo spectra for a monomer calculated fromthe real parts of Eqs. (3.23), (3.27), and (3.28), respectiv ely .Normalization of each graph is such that the maximum valueis unity . The contours are plotted at 10% interv als. Red con-tours show positiv e-going peaks, while blue ones are negativ e-going peaks. For the calculations, all-parallel polarizations(ELO = E3 = E2 = E1) were assumed.
In summary, the 2D photon echo spectrum or 2D corre-lation spectrum provides us with a pure absorptive peak[56, 57], in contrast to the 2D rephasing and nonrephas-ing spectra.
IV. EXPERIMENT AL BACK GROUND
In order to describe experimental implementations of2DES, we begin by summarizing the experiment usingthe two apparatuses in our laboratory as an example sys-tem. W e then discuss which experimental considerationsand requirements are more stringent than in conventionalultrafast techniques.
A. Overview of 2D Spectroscopic Measuremen t
As noted above, 2DES is a four-wave-mixing technique,where three interactions between the incident laser fieldswith the sample induce emission of a signal field. Theexperiment can be performed with a variety of pulsegeometries, methods of delay control, and pulse order-ings. Schematics of two apparatuses in our laboratoryare shown in Fig. 4.
The temporal sequence of the incident pulses is shownin Fig. 5a. In order to produce a 2D spectrum, fourpulses impinge on the sample. Pulses 1, 2 and 3 generatethe signal and pulse 4, called the local oscillator (LO), isused for heterodyne detection. As was described in Sec.III, the delay between pulses 1 and 2, τ , is known as thecoherence time, between pulses 2 and 3, T , is known asthe waiting time, and between pulses 3 and the signalemission, t, is known as the detection time. A Fourier-transform across τ and t for a given value of T produces a2D spectrum, which is a correlation map of excitation andemission energies as a function of the time, T , betweenexcitation and emission events.

8
12
34
d ela y 1d ela y 2
1& 2
3& 4
d iffra ctiveop tic (D O )
sa m p le
sp h erica lm irror
sp ectrom eter
1234
Sp ec tro meter
C o mp u ter
Sa mp le
4 -f Pu lse Sh a p er
In p u t Pu lse
D ela y ed a n d Sh a p ed Pu lses
Gra tin g
SLM
F R OG: Pu lse D ia gn o stic s
C L
Ma sk
a)
b)
FIG. 4: Two sample experimental apparatuses for 2D spec-troscopy. a) A fully non-collinear geometry with mechanicaldelays, where τ is controlled by glass wedges on stepper mo-tor stages. Phase-locked pulse pairs are generated by a dif-fractive optic. b) An SLM-based apparatus where delays areintroduced through phase applied by an SLM. Four individualbeams are created by a spatial mask. Phase stability is main-tained because all four beams impinge on the same optics.This figure was reproduced with permission from [26]
The excitation frequency is measured indirectly byscanning in time. Pulses 1 and 2 prepare, or excite, thesystem. When pulse 1 impinges on the sample, the os-cillation of the electric field under the envelope of thelaser pulse induces an oscillation of the transition dipoleswithin the ensemble of molecules in the sample, or a co-herence |g〉〈e|. The frequency of the oscillation of eachmember of the ensemble is proportional to the excitedstate energy, ~Ωα, because every element contributingto the ensemble propagates with its phase factor, eiΩατ .The collective oscillation of the phases of the transitiondipoles is shown in Fig. 5a, bottom. Scanning the co-herence time samples this collective oscillation because itimprints phase onto the signal with increasing τ . Inter-action of the sample with the second laser pulse stops theoscillation between the ground and excited states. Phasecan accumulate during the waiting time, but only fromcoherence pathways, which will be discussed in Sec. V C.After excitation, or interaction with pulses 1 and 2, thesystem evolves until the arrival of pulse 3. The wait-ing time is the period over which the photophysics of
the system are monitored. The arrival of the third pulseagain produces a coherence in the ensemble, |e〉〈g|. Theresulting oscillatory macroscopic polarization within thesample drives the emission of an electric field, the signalfield. Methods of isolating the signal field, which dependson interactions with all three incident beams, from otheremitted electric fields will be discussed in Sec. IV C.
In the experimental implementation of 2DES, the datais collected in the time domain along the τ axis, whichis Fourier-transformed to produce the excitation frequen-cies. In most apparatuses, the Fourier-transform of thet axis into the frequency domain, however, is performedexperimentally by frequency-dispersing the collinear sig-nal and local oscillator and detecting by spectral inter-ferometry. The detected interference, which oscillates inamplitude along τ and exhibits spectral fringes along ωt,is depicted in Fig. 5b. In Fig. 5c, an example of raw2D data is presented in which the oscillations along bothaxes are visible in the intensity of the detected signal.A Fourier-transform across the τ axis generates the 2Dspectrum as shown in Sec. III.
Laser pulses
Sum of oscillating dipoles
a) k1 k2 k3 kL O
τ
b)
τ ωt
c)
T tcoherence
timew aiting
timedetection
time
... −2∆τ −∆τ 0 ∆τ 2∆τ ...
ωt
E sE L
O(τ,
ωt)
FIG. 5: 2D spectroscopy can retrieve the excitation energy ofthe system, usually implemented by sampling the phase evo-lution of the signal through interferometry. a) The incidentlaser pulses and the collective phase of the oscillating dipoles.b) The form of the detected signal. The fringes along the ωt
axis shows interference from heterodyne detection. The oscil-lation for a fixed ωt along the τ axis is the coherent oscillationsof the sum of all dipoles in the system. c) Experimental 2Ddata, where the ωt axis shows fringe pattern and the τ axisshows the oscillation from the phase accumulated as a func-tion of τ . The τ axis is Fourier-transformed to generate a 2Dspectrum.
The example implementations in Fig. 4 use a fully non-collinear geometry [61]. Figure 4a shows an approachwith mechanical delays and pairwise phase stability froma diffractive-optic based approach [61, 62]. The laserbeam is first split by a beam splitter, after which the twopulses are delayed relative to each other by a retroreflec-tor on a delay stage. The two beams are focused onto adiffractive optic optimized for ±1 orders, which generates

9
two pulse pairs. Glass wedges inserted into beams 1 and2 introduce delays with attosecond precision. The signalemerges collinear with the local oscillator and the twobeams are spectrally dispersed and detected on a chargecoupled device in the frequency domain. Figure 4b il-lustrates a pulse-shaper based implementation based onRef. [63]. Delays are introduced by phase applied with areflective 2D spatial light modulator and the four pulsesare selected from an expanded laser beam with a spatialmask. The signal is detected as in Fig. 4a.
B. Phase stability
1. Phase in 2D spectroscopy
2D spectroscopy is a measurement technique that re-cov ers a complex-v alued signal electric field by detect-ing at the amplitude, not intensity, lev el [39]. Deter-mining the phase of the electric field separates the real(absorptiv e) and imaginary (dispersiv e) contributions to2D spectra. In order to prev ent mixing of the real andimaginary components, the two beams used to scan aFourier-transformed axis must be phase stable, i.e. theremust be pairwise phase stability. The measured signal isproportional to the complex-v alued, third order polariza-tion, which depends on the phase of the incident electricfields in the following way,
P(3) ∝ e−iφ1eiφ2eiφ3 (4.1)
where φi is the phase of the ith field as defined in Eq.(3.1). Acquired phase or phase errors, δ φi, in the laserfields which generate the signal become mapped onto themeasured polarization. As this is a heterodyne exper-iment, the relationship between the interferometricallydetected phase and the phase φsig has the following form[62],
∆φdetected = −φ1 + φ2︸ ︷︷ ︸
−∆φ12
+ φ3 − φLO︸ ︷︷ ︸
∆φ3 LO
+φsig (4.2)
which illustrates why pairwise phase stability is sufficientto remov e phase errors. Therefore, any phase differencebetween pulse pairs introduced by interferometer imper-fections in the setup imprints onto the signal, and in thisway mixes the real and imaginary parts of the generated2D spectrum [55].
There are both activ e and passiv e approaches to gen-erate phase stable pulse pairs. Activ e phase stabilizationmethods inv olv e running HeNe tracer beams across eacharm of the 2D spectrometer and continuously correctingfor path length errors [64, 65]. This requires interfer-ometers between each arm. P assiv e phase stabilizationmethods include diffractiv e-optic approaches [61, 62], ap-proaches with conv entional optics where phase errorscancel [66], and fully or partly collinear geometries wherephase cycling isolates the signal [67, 68]. For adaptiv e-optic based apparatuses, using partially or fully collinear
geometries can impede the introduction of phase errors[68]. 2DES apparatuses generally hav e phase stability inthe range of λ/60 to λ/95 ov er many hours [61, 62, 66].
2. Detection of complex signals
Heterodyne Detection. Heterodyne, or interferometric,detection has been described extensiv ely in sev eral ref-erences and is a method for measuring a complex field[1, 69]. In most 2D apparatuses, measurement occurs byspectrally dispersing the collinear signal and local oscilla-tor fields and recording in the frequency domain. The fre-quency domain, heterodyne-detected signal is the squaremodulus of the sum of the two fields,
I(τ0,T0,ωt) = |ELO(ωt)eiωt∆t + Esig(τ0,T0,ωt)|
2
= |ELO(ωt)|2 + E
∗
sig(τ0,T0,ωt)ELO(ωt)eiωt∆t
+ Esig(τ0,T0,ωt)E∗
LO(ωt)e−iωt∆t
+ |Esig(τ0,T0,ωt)|2 (4.3)
One of the non-interferometric components, |ELO|2, can
be measured separately and subtracted. The final term,which scales quadratically with the low amplitude sig-nal field, becomes v anishingly small and so can be ig-nored [40]. The interference fringes are illustrated alongthe ωt axis of Figs. 5b and c. They can be writtenas 2Re[Esig(τ0,T0,ωt)ELO(ωt)] cos(∆φ(ωt)), and so theinterference term oscillates with the phase difference be-tween the two fields. From this information, the totalcharacterization of a field distribution, or the separa-tion of the absorptiv e (real) and dispersiv e (imaginary)components, can be completed using a known referencefield [1]. That is, the frequency domain fringes in thecross term contain information about the phase of thesignal with respect to the local oscillator [69]. The fre-quency of the fringes ∆φ is the sum of acquired phasefrom the time delay, ωt∆t, and from the phase differ-ence, φLO − φsig, between the signal and the local oscil-lator. Because the temporal offset can be independentlymeasured, the phase of the signal can be recov ered.
In a 2D experiment, heterodyne detection offers twoprimary adv antages ov er homodyne, or intensity, detec-tion. The first adv antage is the separation of the real andimaginary components of the emitted field. The second isthat simultaneous detection of two co-propagating fieldsaids in measuring a low amplitude signal. The magni-tude of the cross terms scale linearly with the signal andwith the local oscillator. Because the amplitude of thelocal oscillator can be set to an easily detectable lev el, themixed term essentially enhances a low amplitude signal.
R etrieval of absolute phase. In the case of a per-fectly functioning apparatus, with no phase offset be-tween beams, precise time delays, and no errors in thearms of the interferometer, the heterodyne detected sig-nal has the correct absolute phase [58], meaning no mix-ing of the real and imaginary components.

10
This accuracy and precision is very challenging toachieve experimentally. Both phase jitter underneath thepulse envelope and small temporal distortions manifest asphase errors in the detected signal. As can be seen fromthe form of the electric field given in Eq. (3.1), a timingerror ∆t has a similar effect to a phase error ∆φ, becauseit imprints phase ω∆t. Thus, subwavelength temporalaccuracy is required to maintain phase stability [70]. Forexample, with the 660 nm pulses, as used in the experi-ments described in Sec. V, an optical cycle has a durationof 2.2 fs. Therefore, a 1 fs error produces a phase errorof almost π. This then requires the following temporalcondition for phase stability,
−∆t1 + ∆t2 + ∆t3 − ∆tLO = 0 (4.4)
Experimental apparatuses often contain differences inlengths of the arms of the interferometer, jitter in thearms of the interferometer, small errors in time steps, andindependent fluctuations of the four beam paths whichdestroy phase stability [40]. There are experimental andanalytical methods to correct for constant phase errors,which will be discussed in Sec. IV B 4.
3. R etrieval of Excitation Energies
During the coherence time, the system evolves in a co-herence, so for a given τ0, the system acquires a phaseeiωτ τ0 , as illustrated in Fig. 5a, bottom. Because theexperiment is performed on an inhomogeneously broad-ened ensemble, the slightly different excited state energyof each member produces an oscillation at a slightly dif-ferent frequency, which causes the dephasing in the col-lective oscillation shown in Fig. 5a, bottom. Thus, fora given value ωt, the interference fringes exhibit an os-cillation in amplitude along τ as shown by the dashedline in Fig. 5b, where for each additional increment∆τ = 2π/ωτ , the signal undergoes a full phase rotation.This resultant oscillation in signal amplitude can also beseen along the τ axis in the raw data shown in Fig. 5c. AFourier-transform across the τ axis retrieves frequenciesωτ and generates the 2D spectrum. Because the exci-tation energy is retrieved by controlling the time thatthe dipoles within the sample undergo a collective os-cillation, the measurement requires highly accurate timedelays and phase stable pulses. The phase stability isrequired because an additional phase offset will imprintphase onto the signal similarly to an additional time in-crement.
Time delays In this experiment, the time delay be-tween beams 1 and 2 must be set with higher accuracyand precision than in other techniques. Because scan-ning this time delay constructs a temporal axis which isthen Fourier-transformed to generate frequencies, errorsin temporal sampling produce errors in the frequency do-main information. Sub-wavelength accuracy is requiredto retrieve excitation frequencies without the introduc-tion of Fourier artifacts.
The method by which temporal delays are introducedalso varies among 2D apparatuses. In Fig. 1, the intro-duction of controllable amount of glass using wedges onstepper motors allows < 50 attosecond precision, whichis more precise than with conventional delay methods.In Fig. 1b, the pulses are delayed by the applicationof linear phase using the SLM, which provides 10-50 astemporal resolution [63].
Because of the short period of an optical cycle, recon-structing the frequency information through sampling atthe Nyquist frequency requires a very large number ofdata points. While the precision requirement remains,the number of data points needed can be decreased bytwo approaches, undersampling or sampling in the rotat-ing frame. For the apparatus in Fig. 1a, the τ delayis often sampled at approximately an odd multiple ofthe Nyquist frequency of the excitation frequency. Thefrequency components are limited to those within thebandwidth of the laser, so the detected signal can beunfolded to retrieve the frequency domain information.This method is described in detail in several references[41, 71]. In this way, temporal step size can be increasedby undersampling the time axis and extracting the fre-quency through aliasing methods [41, 71].
The second approach is to detect within the rotatingframe and is the method applied in the apparatus in Fig.4b. We now describe two ways of thinking about apply-ing delays using the rotating wave approximation. First,in the frequency domain, a time delay manifests as aphase factor, eiωt. The additional phase accrued, there-fore, is linear in frequency. Because an SLM directly con-trols the phase of the beams, an SLM can be used to es-sentially off-set the additional phase along the frequencyaxis, which results in terms ei(ω−ωRF)t. The phase off-setalong the frequency axis is the rotating frame frequency,ωRF. Therefore, as additional time delays are applied,the oscillation frequency along the τ axis decreases.
In the second picture, delays introduced in the rotatingframe can be understood as arising from the fact that thephase control introduced by the SLM allows both phaseand time delays to be applied. When the temporal delayis introduced, a phase shift is added as well. The addi-tional phase at each τ is chosen to maintain a constantphase at a selected carrier frequency, and all other fre-quencies components are delayed relative to the referencefrequency. Instead of introducing only delay with eiωτ τ ,both a time and phase component are mapped onto thepulse, ei(ωτ−ωRF)τ . While delays introduced using con-ventional techniques introduce delay eiωτ τ , in rotatingwave detection, at each time step, the addition of a phaseω∆τ−∆φ means that scanning an optical cycle in τ doesnot necessarily correspond to a full phase rotation. In theapparatus in Fig. 4b, the oscillatory cycle is often set to20 fs, which oversamples the oscillations for higher accu-racy.

11
4. Methods to generate correctly phased spectra
2D electronic spectroscopy almost always requires aphase correction factor to account for experimental phaseoffsets, which arise from problems such as unequallengths between the two arms, e.g. φ0 = φ1 − φ2. Inorder to maintain phase stability, however, any phase jit-ter, δφi, must occur on both beams, or during the courseof data collection δφ1 = δφ2 and δφ3 = δφLO. Whilethere are no methods to account for any unequal phasejitter terms, there are several experimental and analyticalmethods to correct for a constant phase difference. Analgorithm-based method has been developed, although ithas yet to be widely implemented [72]. The analyticalmethod does, however, offer the potential for significantexperimental simplification.
In the passively-stabilized, diffractive optic based setupin Fig. 4a, phase offset in 2D data is corrected by com-paring to separately collected pump-probe data [40]. Theprojection-slice theorem of Fourier transforms states thatthe projection in one domain is equivalent to a slice inanother domain. This equivalency gives rise to a rela-tionship between the pump-probe data and the 2D data[41, 71]. The projection along the ωτ axis is related to theslice at τ = 0, which is a pump-probe measurement. Theprojection slice theorem [39, 41] relates the real compo-nent of a projection of the 2D data to spectrally resolvedpump-probe data in the following way,
Spump-prob e(ωt, T0)
= Re
[
ωtEprob e(ωt)
∫∞
−∞
dωτ
2πSPE(ωt, T0, ωτ )
]
(4.5)
where Eprob e(ωt) is the electric field of the probe pulse.Transient absorption experiments are able to recover thecorrect phase because pump-probe experiments involveonly two pulses, where the first two interactions are bothfrom pulse 1 and the third interaction and the local os-cillator are from pulse 2. Therefore, it is an inherentlyphased measurement. By fitting the projection of a 2Dspectrum to the spectrally resolved pump-probe data, theabsolute phase of the measured data can be retrieved. Azeroth and first order phase correction term, eiφ0+iωt∆t,is generally sufficient to match the 2D spectra to thepump-probe data. This approach, however, only workswhen the spectrum (ω1 = ω2, ω3 = ωLO) and the po-
larization (E1 = E2, E3 = ELO) are the same for eachpulse pair.
C. Beam geometry
A 2D spectrum is a sum of the selected energy and mo-mentum conserving pathways, ks = −k1 + k2 + k3. Theexperiment can be performed in fully, partially, or non-collinear beam geometries. In the case of non-collineargeometries, the condition of momentum conservation lim-its the spectrum from the signal detected in the spa-
tially phase-matched direction to a subset of third or-der signals, or a subset of molecular processes [1]. Theselected third-order signal can be extracted through spa-tially windowing and detecting using a box geometry. Inthis geometry, there is background free detection of thegenerated signal. In the case of fully or partially collineargeometries, phase cycling is used to isolate the signal [68].In phase cycling, the signal is isolated by a linear combi-nation formed from selectively multiplying each beam orcombinations of beams by a phase factor, eiπ. For thisprocess, because the signal has phase dependence on allthree excitation pulses, multiplying any odd number ofpulses by the phase factor eiπ flips the phase of the signal.Multiplying an even number of pulses by the phase factordoes not flip the phase of the signal. Subtracting the sumof the odd permutations from the even permutations sep-arates the third order signal, which depends on all threeincident pulses. A phase cycling sequence can also aidin background removal for fully non-collinear geometriesbecause scattered light does not share the third orderphase dependence.
D. Artifacts and spectral phase
In ultrafast experiments, transform-limited compres-sion is often difficult to achieve, particularly for the caseof broad bandwidths. Using laser pulses with differentorders of spectral phase can lead to various artifactswithin a 2D spectrum. Figure 6a-c are experimental, ab-solute value, correlation 2D spectra of the non-resonantresponse of sapphire. Three different spectral phaseswere applied to the four laser pulses using the SLM inthe experimental apparatus from Fig. 4b. Figure 6adisplays the spectrum in the case of almost transform-limited pulses. In this case, basically the spectral profileof the laser pulse is recovered. For the spectrum in Fig.6b, a parabolic phase was applied to the laser pulse. It isimportant to note here that the peak shape generated bythe chirped pulse can easily be confused with correlatedexcited states, or an inhomogeneously broadened peak.For a non-resonant response, however, a correlation isphysically impossible. Cubic spectral phase, as shown inFig. 6c, is characterized by an x-shaped artifact.
For experiments such as pump-probe spectroscopies,after characterizing dispersion in the laser pulses, algo-rithms have been used to remove the effects of spectralphase [73–75]. In 2D spectroscopy, however, algorithmsto remove these artifacts from spectral phase are muchmore complicated [76]. Each peak within the generatedspectrum is a function of four light-matter interactions.The two frequency axes of the spectrum, however, onlyprovide information about the frequency of two of thefour interactions. The two unknown frequencies compli-cate determination of the effect of sub-optimal compres-sion, even for a well-characterized spectral phase.
The spectral phase can change the time delays betweenthe four interactions. Sample Liouville pathways, cor-

12
d)
time
frequ
ency
k1 k2 k3 ksig
a) b) c)transform-limited parabolic spectral phase cubic spectral phase
Ωα
Ωβ
τ T t
R2(αα)
0 00 eα
eα eα
eα 00 0
R2(αβ)
0 00 eα
e β eα
e β 00 0
R2(ααββ)
0 00 eα
eβ eβ
eβ 00 0
eα eα
e)
ω (cm-1)1275011250 12000
ω (cm-1)ω (cm-1)
12750
11250
12000
ω (c
m-1
)
1275011250 12000
12750
11250
12000
ω (c
m-1
)
1275011250 12000
12750
11250
12000
ω (c
m-1
)
FIG. 6: Experimental 2D spectra of the nonresonant responseof sapphire for transform-limited (a), linearly chirped (b), andcubic applied spectral phased (c) pulses. Spectral phase wasapplied to the pulses using the SLM in Fig. 4b. d) A depictionof linearly chirped pulses for a FWM experiment with trajec-tories for three sample Liouville pathways. As seen by the tra-jectories, chirped pulses can result in errors in time spacings.e) Feynman diagrams to illustrate the Liouville pathways in(d).
responding to different molecular processes within thesystem, are shown in Fig. 6d. When all four inter-actions are at the same frequency, such as in the caseof molecular processes corresponding to linear absorp-
tion (R(αα)2 ), there is no effect on the peak. In the case
of population transfer (Fig. 2e), where ω1 = ω2 andω3 = ωLO, the intended and actual waiting times are dif-ferent. Measured population dynamics can, therefore, be
incorrect. Finally, for the case of (R(αβ)2 ), where ω1 6= ω2
and ω3 6= ωLO, the Fourier-transformed time intervalis altered, which leads to incorrect retrieval of frequen-cies. Essentially, these effects can change the time order-ing of the incident pulses and introduce artifacts withinthe Fourier transform. In the case of spectra with wait-ing times after the decay of coherence contributions andmuch longer than the pulse length, chirp effects shouldno longer appear.
V. APPLICATIONS TO PHOTOSYNTHETIC
COMPLEXES
As an example of the utility of 2D electronic spec-troscopy, we discuss how 2D spectroscopy has been ap-plied to disentangling the function of the Photosystem
II (PSII) supercomplex of higher plants. PSII is a so-phisticated natural nano-device, with several associatedPPCs, each containing multiple pigments, with dynam-ics governed by interactions within and between thesePPCs. Understanding the molecular machinery and re-lating structure to function is particularly challenging be-cause of the nonequivalent geometries of the chlorophyllwithin the PPCs and the resultant rugged energy land-scape of plant systems.
A. 2D electronic spectrum of pigment-protein
complexes
2D electronic spectroscopy provides an incisive tool tostudy electronic couplings and electronic energy trans-fer among pigments. In general, optical spectroscopyprovides us with the information projected onto energyeigenstates, and therefore it is common to employ energyeigenstates of electronic excitations, which are termed ex-
citons in the literature. For this purpose, we rewrite thePPC Hamiltonian by substituting Eqs. (2.1)-(2.7) intoEqs. (2.5) and (2.6) as
H(n)PPC = H
(n)site + H
(n)site−en v + Hen v. (5.1)
The first term on the right hand side is the electronicexcitation Hamiltonian comprised of the Franck-Condonenergies and electronic couplings,
H(1)site = ~
∑
m
Ωm|m〉〈m| + ~
∑
mn
Jmn|m〉〈n|, (5.2)
H(2)site = ~
∑
mn
(Ωm + Ωn)|mn〉〈mn|
+ ~
∑
mnk
(
Jmk|mn〉〈kn| + Jmk|mn〉〈mk|)
. (5.3)
The second part in Eq. (5.1) corresponds to the couplingof environmental degrees of freedom to the electronic ex-citations,
H(1)site−en v =
∑
m
um|m〉〈m|, (5.4)
H(2)site−en v =
∑
mn
(um + un)|mn〉〈mn|. (5.5)
The last term in Eq. (5.1) is a set of normal modeHamiltonians, i.e. the phonon Hamiltonian expressed asHen v =
∑
ξ ~ωξ(p2ξ + q2
ξ )/2. To obtain the one- and two-excitation energies and states, the Hamiltonians in Eqs.(5.2) and (5.3) are diagonalized with orthogonal trans-formation operators V and W as
H(1)ex ≡ V −1H
(1)siteV, (5.6)
H(2)ex ≡ W−1H
(2)siteW, (5.7)
where the diagonal elements of H(1)ex and H
(2)ex are the
eigenenergies in the single- and double-excitation man-ifolds. The one- and two-excitation states belonging to

13
the eigenenergies are expressed as
|eα〉 ≡∑
m
V −1αm|m〉; H(1)
ex |eα〉 = ~Ωα|eα〉, (5.8)
|fγ〉 ≡∑
mn
W−1γ(mn)|mn〉; H(2)
ex |fγ〉 = ~Ωγ |fβ〉. (5.9)
We note Roman letters are employed for the site rep-resentation while Greek letters are used for the excitonrepresentation. Once the eigensystems are determined,the exciton transition dipoles can be expressed as linearcombinations of pigments’ transition dipoles,
µα0 = 〈eα|µ|0〉 =∑
m
V −1αmµm0, (5.10)
µγα = 〈fγ |µ|eα〉 =∑
mn
W−1γ(mn)V
−1αmµn0. (5.11)
The excitation-environment coupling Hamiltonian inEqs. (5.4) and (5.5) is also transformed as
H(1)ex−env ≡ V −1H
(1)site−envV, (5.12)
H(2)ex−env ≡ W−1H
(2)site−envW. (5.13)
In this exciton representation, the diagonal matrix ele-ments of the excitation-environment coupling Hamilto-nians are interpreted as fluctuations in the exciton en-ergies, whereas the off-diagonal elements contribute totransition or relaxation processes among excitons. Forexample, the matrix element
〈eα|H(1)ex−env|eβ〉 =
∑
m
umVmαVmβ (5.14)
causes exciton α to relax to relaxation β, |eα〉〈eα| →|eβ〉〈eβ |, in the single-excitation manifold. Here, oneshould not overlook that these eigenstates and eigenen-ergies are obtained via diagonalization of the Hamil-tonian comprised of the Franck-Condon transition en-ergies. However, energies of the actual excitations willdeviate from these values as the environmental reorgani-zation (Stokes shift) takes place [3, 4, 6, 77]. Also, theconcept of the exciton relaxation is useful and intuitivein analyzing ensemble averaged behaviors of PPCs whichare observed by means of condensed phase spectroscopy.Nevertheless, one should be cautious in discussing micro-scopic dynamics in individual PPCs [78]. Although op-tical spectra provide us with the information projectedonto energy eigenstates, this does not necessarily meanthat PPCs are always in their energy eigenstates.
B. Monitoring Pathw ays of Energy Transfer
1. Electronic couplings between pigments
2D electronic spectroscopy is the optical counterpartof 2D NMR spectroscopic methods such as COSY and
Rephasing
Nonrephasing
1 2 3 sig 1 2 3 sig
1 2 3 sig 1 2 3 sig
RephasingRephasing Nonrephasing
FIG. 7: Double-sided Feynman diagrams which represent theLiouville space pathways contributing cross peaks (ωt,ωτ ) =(Ωβ,Ωα) and (Ωγ − Ωα,Ωα) in 2D electronic spectra. Thecorresponding energy-lev el diagrams are also shown, wherethe solid arrows represent interactions of electric fields withk et-states while the dashed arrows represent interactions ofelectric fields with bra-states.
NOESY, which are powerful tools for organic structuralanalysis and structural biology [41]. Electronic cou-plings among different pigments can be identified visu-ally from off-diagonal peaks termed cross peaks. Figure7 presents the double-sided Feynman diagrams produc-ing cross peaks. The Liouville space pathways R3 andR4 contribute to the ground state bleaching (GSB) inthe language of pump-probe spectroscopy, and produce across peak at (ωt,ωτ ) = (Ωβ,Ωα) whose intensity is pro-
portional to 〈(µβ0 ·ELO)(µβ0 ·E3)(µα0 ·E2)(µα0 ·E1)〉ori,where 〈· · · 〉ori denotes averaging over the molecular ori-entations. On the other hand, the pathways R∗
1 and R∗
2
describe the excited state absorption (ESA) and give across peak at (ωt,ωτ ) = (Ωγ − Ωα,Ωα) whose intensity
is ∝ −〈(µγα · ELO)(µγα · E3)(µα0 · E2)(µα0 · E1)〉ori.The ESA is observed as negative-going peaks in the con-ventional 2D electronic spectra with use of all-parallelpolarization, ELO = E3 = E2 = E1. W e note that the

14
Rephasing
Nonrephasing
1 2 3 sig
relaxation
1 2 3 sig
relaxation
1 2 3 sig
relaxation
1 2 3 sig
relaxation
Rephasing Nonrephasing
emergence emergence
FIG. 8: Double-sided Feynman diagrams which represent theexciton relaxation process, |eα〉〈eα| → |eβ〉〈eβ | in the waitingperiod, T . The corresponding energy-level diagrams are alsoshown. 2D electronic spectra show this process as the emer-gence of a cross peak (ωt, ωτ ) = (Ωβ , Ωα) and the decrease ofa peak (ωt, ωτ ) = (Ωγ − Ωβ , Ωα) in magnitude.
sign conventions in 2D spectroscopy are the opposite ofthose used in pump-probe spectroscopy. When we con-sider |eα〉 and |eβ〉 corresponding to electronic excitationson uncoupled pigments |a〉 and |b〉, we find that the crosspeak at (ωt, ωτ ) = (Ωβ = Ωb, Ωα = Ωa) disappears be-cause the GSB pathways and the ESA pathways involv-ing |fγ〉 = |ab〉 destructively interfere and exactly cancelout. Note Ωα = Ωa, Ωβ = Ωb, µα0 = µa0, µβ0 = µb0,and µγα = µb0 from Eq. (5.11). Therefore, the existenceof cross peaks at very short waiting times is definite evi-dence of nonzero electronic couplings.
2. Electronic energy transfer
Spatial transport of electronic excitation energy inphotosynthetic PPCs is observed as stepping down a lad-der of excitonic energies as a function of the waiting pe-riod T , where the electronic excitation is transferred froma higher energy exciton to a lower energy one because ofthe matrix element given in Eq. (5.14). The signature ofsuch relaxation appears in 2D electronic spectra as theemergence of a cross peak (ωt, ωτ ) = (Ωβ , Ωα) betweena lower energy exciton |eβ〉 during the detection periodt and a higher energy exciton |eα〉 during the coherenceperiod τ , as demonstrated with the double-sided Feyn-man diagrams R2 and R1 in Figure 8. Simultaneously,this relaxation process reduces the magnitude of peaksat (ωt, ωτ ) = (Ωγ −Ωβ , Ωα) located around the diagonalline, as shown in the diagram for R∗
1 and R∗
2. Monitor-ing electronic energy migration in photosynthetic PPCswith use of 2D electronic spectra has been extensivelydiscussed [11, 12, 58, 79–83].
3. R elaxation Pathways in LHCII
While off-diagonal peaks at T = 0 indicate coupledexcited states, the appearance of off-diagonal peaks withincreasing waiting time indicates energy transfer. Theappearance and evolution of these peaks can be simul-taneously observed for all states within the spectrum ofthe laser pulse. Although there are other ultrafast tech-niques that also probe femtosecond dynamics, they re-quire either integrating over excitation energy or usingspectrally narrow pulses, which mean a loss of tempo-ral resolution. As described above, in 2D spectroscopy,spectral resolution is maintained across excitation andemission frequencies, with no need to lose temporal res-olution between excitation and emission events. In thecase of closely spaced or overlapping excited states, res-olution across these three dimensions becomes necessaryto elucidate dynamics.
Examination of the major light-harvesting complex ofPhotosystem II (LHCII) by 2D spectroscopy has provedparticularly informative. LHCII is the most abundantpigment-protein complex and binds over 50% of theworld’s chlorophyll, the primary light-harvesting mole-cule. Arrays of LHCII complete the majority of lightabsorption in green plants. The crystal structure modelis shown in Fig. 1a [20]. Each monomer of trimericLHCII contains fourteen chlorophyll found in two spec-tral and structural variants, 8 Chl-a and 6 Chl-b. LHCIIcontains a set of discrete excited states, or excitons, thatexhibit relaxation on multiple timescales. Because thefourteen chlorophyll per monomer give rise to fourteenclosely spaced excited states, the congestion has oftenhindered measurements of the dynamics of individualdonor and acceptor states. While other techniques hadprovided much insight by previously identifying the exis-tence of various relaxation timescales within many spec-

15
tral regions [84–88], 2D experiments improved the tem-poral resolution of observed dynamics, allowed identifi-cation and isolation of intermediates in relaxation path-ways, and surveyed the dynamics throughout the entireQy region [81].
30 fs
70 fs 70 fs
200 fs200 fs
300 fs300 fs
13000 fs 13000 fs
30 fsa)
b)
c)
d)
e)
15000
14700
1550014700
15500
15200
15000
14700
15500
15200
15000
14700
15500
15200
15000
14700
15500
15200
15000
14700
15500
15200
15000
14700
15500
15200
14700
15500
15200
15000
14700
15500
15200
15000
14700
15500
15200
15000
14700
15500
15200
15000
14700
15500
10
0
-10
20
0
-20
30
0
-30
10
0
-10
30
0
-30
10
0
-10
20
0
-20
.001
0
-.001
.002
0
-.002
60
0
-60
3 5
1 2 4
3 5
1 2 4
3 5
1 2 4
3 5
1 2 4
6
FIG. 9: Real, 2D spectra of LHCII for selected waiting timesT = 30, 70, 200, 300, 1.3 fs with relaxation spectra (left) andnon-rephasing spectra (right). Dynamics are highlighted byarrows. This figure was reproduced with permission from [81].
By simultaneously recording all dynamics within theQy region, the 2D spectra map out the relaxation path-ways within LHCII. Real (absorptive) 2D spectra for se-lected time points are shown in Fig. 9, left, and real,non-rephasing spectra are in Fig. 9, right. The spec-tra are divided into three energy regions, indicated bytickmarks: 1) the Chl-a region (14, 700 − 15, 000 cm−1),which contains a low and mid energy band; 2) an inter-mediate region (15, 000 − 15, 200 cm−1), which containshigh energy Chl-a and low energy Chl-b; and 3) the Chl-b region (15, 200 − 15, 500 cm−1). The appearance andevolution of five cross-peaks show energy transfer be-tween these states. Arrows within the 2D spectra high-light these off-diagonal peaks in the non-rephasing spec-tra, because off-diagonal peaks in non-rephasing spectraarise almost entirely from energy transfer. It is impor-
tant to note that the phase-twisted lineshape, or mixedabsorptive and dispersive contributions to peaks in thenon-rephasing spectra, can be interpreted as additionalfeatures. Comparison to the relaxation spectra is, there-fore, important when using the non-rephasing spectra toelucidate the dynamics.
Previously unobserved sub-100 fs dynamics were de-tected within all spectral regions. As seen in Fig. 9a, by30 fs (T = 30 fs) multiple energy transfer processes areevidenced by the appearance of cross-peaks where indi-cated by the white arrows. These peaks grow in for wait-ing times greater than zero, thus showing energy trans-fer. In Fig. 9b, there is increased relative amplitudein the low energy Chl-a band. The change in relativeamplitude of these peaks at 70 fs indicates that thereare multiple processes occurring within the first 100 fs.Rapid relaxation within the Chl-b band is also visible inthe asymmetry of the Chl-b peak in the relaxation spec-trum [81]. That is, there are several faster energy transfersteps within LHCII than found using other experimentaltechniques or predicted by theory.
After the initial population and subsequent depopula-tion of mid-energy Chl-a band, these states again serve asintermediates. In Fig. 9c, in the 200 fs spectrum, there isstill increased relative amplitude in the low energy Chl-aregion. By 300 fs, cross-peaks 3 and 5 again increase inrelative amplitude, as seen in Fig. 9d. The change inwhich states are populated between 200 fs and 300 fs in-dicates that energy transfers into the mid-energy Chl-aband from both the Chl-b and intermediate regions on afew hundred femtosecond timescale.
All picosecond spectra display very similar features,and Fig. 9e is shown as a representative timepoint(T = 13 ps). From 1 ps to 20 ps, when the signal ampli-tude decreases to a level below the noise floor, the spectracontain two notable features. First, the horizontal bandat the energy of the low energy Chl-a indicates most ofthe population across all spectral regions has relaxed intothe low energy Chl-a band. Thus, initial excitations intoChl-b, intermediate, and mid-energy Chl-a regions trans-fer into the low energy Chl-a states. Second, the appear-ance of a new cross-peak shows energy transfer from thelow energy Chl-b into a long-lived intermediate state inthe mid-energy Chl-a region. Excitation remains in theintermediate state, labeled as cross-peak 6, until the sig-nal dies by 20 ps [81]. Therefore, energy transfers out ofthe intermediate state much more slowly than in the caseof other energy transfer steps within LHCII.
After identifying each energy transfer step, under-standing how these processes arise requires elucidatinghow the molecular components give rise to the observedrelaxation pathways. The information provided by 2Dspectroscopy can greatly aid correlating the individualchlorophyll within the x-ray crystallography structurewith each of the observed relaxation steps. Within PPCs,this detailed modeling of relaxation pathways is often dif-ficult. Despite the structural information, as discussed inSec. III, there is little direct information about the de-

16
tails of the pigment Hamilton and pigment-protein inter-action. In combination with theoretical methods, how-ever, the dynamics of each energy transfer step were as-signed to individual chlorophyll, as described in detail inRef. [81]. The interpretation and theoretical results alsoprovided a general picture of how excitation energy flowsthrough the complex. The chlorophyll within LHCII aregrouped into strongly coupled (20 − 100 cm−1) clustersof 2-3 molecules. Because of the strong coupling, andlarge spatial overlap of the excited state wavefunctions,rapid (sub-100 fs) relaxation occurs within these mole-cules. Energy transfer between these clusters, or be-tween excited states mostly localized on more weaklycoupled molecules, occurs in several hundred femtosec-onds. Within the structural model of LHCII, there aretwo chlorophyll well separated from, and thus weaklycoupled to, lower energy chlorophyll. These chlorophyllserve as the sites of long-lived intermediate states.
From this picture of excitation energy flow withinLHCII several fundamental principles behind nature’s de-sign for efficient light-harvesting are observed. First, ex-citation energy eventually relaxes to the low energy Chl-a band, most likely into states on the external edge ofLHCII. The emergent horizontal band which emits fromthe low energy Chl-a region indicates a pooling of en-ergy into these states. The model of delocalized excitedstates creates multiple excited states on an exit site, prox-imal to nearby complexes, thereby aiding migration ofthe excitation out of LHCII and towards the reactioncenter. Second, there is a variety of excitation energytransfer timescales due to a range of couplings. Alongeach relaxation pathway, the combination of timescalesmay aid in unidirectional energy transfer by using cou-pling to overcome energy gaps larger than kBT and byusing small energy gaps to overcome long-lived quantumbeating. Long-lived quantum beating must be overcomebecause perpetual oscillatory motion means there is nopermanent spatial translocation of excitation energy. Inthese systems, the excitation must end up localized onthe reaction center. Coherent interaction, however, playsan important role in how the excitation migrates.
C. Quantum Coherent Electronic Energy T ransfer
1. Quantum Coherence among Excitons
Coherent electronic energy transfer indicates that elec-tronic excitations travel through PPCs as a quantum me-chanical wave packet keeping their phase coherence, aswas discussed in Sec. II. In the exciton representation,this process can be observed as time-evolution of super-position or coherence between different excitons, e.g.
e−iH(1)ex T/~|eα〉〈eβ |e
iH(1)ex T/~ ' e−i(Ωα−Ωβ)T |eα〉〈eβ |.
(5.15)
This quantum coherence manifests itself as oscillationsin the peak amplitudes of 2D photon echo spectra as
Rephasing
Nonrephasing
1 2 3 sig
coherence
coherence
1 2 3 sig
1 2 3 sig 1 2 3 sig
coherence
coherence
RephasingRephasing NonrephasingNonrephasing
beating beating
FIG. 10: Double-sided Feynman diagrams which represent theelectronic quantum coherence |eα〉〈eβ | in the waiting period.The corresponding energy-level diagrams are also shown. Thisprocess is observed as the beating of cross peaks (ωt, ωτ ) =(Ωβ , Ωα) and (Ωγ − Ωα, Ωα) in 2D rephasing spectrum, andas the beating of diagonal peaks (ωt, ωτ ) = (Ωα, Ωα) and(Ωγ − Ωβ , Ωα) in 2D nonrephasing spectrum.
a function of the waiting time T with beat frequenciesequal to the energy differences between the contributingexcitons, Ωα − Ωβ . In congested spectra where excitonsare closely spaced energetically, cross peaks will appearnear their diagonal counterparts. As a result, oscillationsin the amplitude of cross peaks interfere with those of thepeaks directly on the diagonal peaks, making it extremelydifficult to isolate individual coherence contributions inconventional 2D photon echo spectra. As shown in Fig.10, however, the coherence induces oscillatory behaviorof a diagonal peak at (ωt, ωτ ) = (Ωα, Ωα) and a crosspeak located near the diagonal line (Ωγ −Ωα, Ωβ) in 2Dnonrephasing spectra [89], and thus coherence pathwaysassociated with diagonal peaks can be isolated throughanalysis of 2D nonrephasing spectra

17
100
200
300
400
500
600
700
800
900
1000
Bea
t Fre
quen
cy (c
m-1
)
14800 15000 15200
ωt = ωτ (cm-1)
0
100
200
300
400
500
Waiting T im
e (fs)
1450014750
1500015250
15500
ω t = ωτ (
cm-1)
0
Am
plitu
de(a) (b)
FIG. 11: (a) Evolution of the LHCII diagonal, nonrephasingsignal from 2D spectra as a function of waiting time. Beatingdue to electronic coherence is evident in both Chla and b re-gions. Data was acquired at 10 fs intervals, and a cubic splineinterpolation is used for presentation. (b) LHCII coherencepower spectrum for a portion of the Chla region constructedby Fourier transforming along the waiting times (T ) axis in(a). The beat frequency axis begins at 50 cm−1 to remov e theambiguous low frequency components. Contours are placedat 10% interv als and amplitude increases from white to red.This figure is reprinted with permission from Ref. [16]
2. Quantum Coherent Energy T ransfer in LHCII
In recent years Fleming and coworkers have in-vestigated the Fenna-Matthews-Olson (FMO) pigment-protein complex isolated from a green sulfur bacteriumby means of 2D electronic photon echo spectroscopy[58, 59, 79, 80]. One of their experiments succeeded inobserving long-lasting quantum beating providing directevidence for long-lived electronic coherence [80]. The ob-served coherence lasts for time scales similar to the EETtimescales, implying that electronic excitations move co-herently through the FMO protein rather than by inco-herent hopping motion, although it had been generallyassumed that electronic coherence decay so rapidly thatit does not affect the EET [22]. Observation of long-lasting quantum coherence is not unique to the FMOcomplex. Coherent EET dynamics in the reaction centerof a purple bacterium were also revealed by applying thetwo-color electronic coherent photon echo technique [14].Although the electronic quantum coherence in the PPCswas originally observed outside the physiological range oftemperatures, the presence of quantum coherence lastingup to ∼ 300 fs was detected in a conjugated polymers[90], the antenna complexes in marine cryptophyte algae[82] and the FMO complex [91] at physiological tempera-tures. Interestingly, this timescale was consistent with aprevious theoretical prediction [7]. However, the signifi-cance of quantum coherence to electronic energy transferin photosynthetic light harvesting cannot be argued with-out it first being established as a universal phenomenon.If coherence is essential for efficient energy transfer, itshould be present in the large antenna complexes whosesole responsibility is to absorb solar energy and funnel itto the reaction centers. Despite having energy transfer
as its primary function, the FMO complex is not a keyphotosynthetic complex with its analog largely absent inhigher plants. This begs the question as to whether elec-tronic coherence was important enough to survive evo-lution. Recent 2D electronic spectroscopic experimentshave revealed evidence of electronic coherence in LHCIIat cryogenic temperature, 77 K [92].
Figure 11(a) presents the diagonal cut of 2D non-rephasing spectra shown in Fig. 9 as a function ofT . The strong peaks assigned to Chl-a (ω ∼ 14, 500 −
15, 000 cm−1) as well as the weaker peaks from Chl-b(ω ∼ 15, 500 cm−1) demonstrate clear beating through-out the entire 500 fs duration of the experiment. Becausethe excitons responsible for these beating peaks originatefrom individual Chl molecules, the coherent dynamicscan be mapped back onto the site basis. As seen in otherphotosynthetic complexes [14, 80, 82, 91], the coherenceis long-lived in LHCII, with the quantum beating last-ing beyond 500 fs scan of the experiment and beyond thelifetimes of many excitons in the system, suggesting thatthis is a general phenomenon in photosynthetic light har-vesting systems. The observations of long-lived quantumcoherence have led to speculation that coherence maycontribute to optimizing photosynthetic energy transferefficiency. A potential role of quantum coherence is toovercome local energetic traps and aid smooth and robustenergy flow toward targets such as the reaction centersby taking advantage of quantum coherence and the en-ergy landscape of pigments tuned by the protein scaffold[7].
While these studies provide insight into the role ofquantum coherence in photosynthetic EET, questions re-main regarding the underlying mechanisms preservingthe long-lived quantum coherence. In order to elucidatethe origin of the long-lived quantum coherence and its in-terplay with the protein environment, Ishizaki and Flem-ing presented the second-order cumulant time-nonlocalquantum master equation which reduces to the conven-tional Forster and Redfield theories in their respectivelimit of validity [6]. In the regime of coherent wavelikemotion, the equation predicts several times longer life-time of quantum coherence between electronic excitedstates of pigments than the Redfield equation does. Ref-erence [6] also demonstrated that quantum coherent mo-tion can be observed even when reorganization energy islarge in comparison to intersite electronic coupling, al-though this situation is usually regarded as the Forsterregime. The reason for the long-lived quantum coherenceis as follows: In a region of small reorganization energy,the slow fluctuation sustains longer-lived coherent oscilla-tion, whereas the Markov approximation in the Redfieldframework causes infinitely fast fluctuations which thencollapse the quantum coherence. In the region of largereorganization energy, on the other hand, the sluggish re-organization dynamics allows the excitation to stay abovean energy barrier separating two local minima, which cor-respond to the two sites in the adiabatic potential surface,for a prolonged time. The dynamic behavior of the inter-

18
mediate regime, which is typical in photosynthetic EET,can be understood as the combined influence of the slowfluctuation effect in the regime of small reorganizationenergy and the slow dissipation effect in the large reor-ganization energy. In order to visualize this statement, itis helpful to consider the following minimal model insteadof the PPC Hamiltonian in Eqs. (2.1)-(2.5) [6, 16]
HPPC =2
∑
m=0
εm(q)|m〉〈m| + ~J12(|1〉〈2| + |2〉〈1|)
(5.16)
where
ε1(q) = ε0(q) + ~Ω1 − ~ωphdq1, (5.17a)
ε2(q) = ε0(q) + ~Ω2 − ~ωphdq2 (5.17b)
with
ε0(q) =~ωph
2q21 +
~ωph
2q22 . (5.18)
The reorganization energy is expressed as ~λm =~ωphd2/2. The Hamiltonian Eq. (5.16) can be easilydiagonalized, and then adiabatic excitonic potential sur-faces in the single-exciton manifold can be obtained as
E±(q) =ε1(q) + ε2(q)
2±
√
(~J12)2 +
[
ε1(q) − ε2(q)
2
]2
.
(5.19)
In Fig. 12, we draw the adiabatic potential surface forthe lower energy, E−(q), as a function of two phononcoordinates, q1 and q2. The parameters in this modelare chosen to be Ω1 − Ω2 = 100 cm−1, J12 = 100 cm−1,ωph = 53 cm−1, and λ1 = λ2 = 500 cm−1. Since the reor-ganization energy is large compared with the electroniccoupling, we can observe two local minima which repre-sent the two sites, |1〉 = |ϕ1e〉|ϕ2g〉 and |2〉 = |ϕ2e〉|ϕ1g〉.Incoherent hopping EET describes the transition be-tween the local minima. Attention will now be givento the point of origin, which corresponds to the Franck-Condon state. The energy of the point is higher than thebarrier between the minima; therefore, we find that theelectronic excited state is delocalized just after the ex-citation despite being in the incoherent hopping regime,λ > J12. As time increases, the dissipation of reorganiza-tion energy proceeds and the excitation will fall off intoone of the minima and become localized.
Another possible reason for the long-lived quantum co-herence is strong correlation between fluctuations in elec-tronic energies of different pigment, as supported by theexperimental results of Lee, Cheng and Fleming [14]. Ifdifferent pigments share the dynamic effects of their lo-cal environmental, fluctuations in their electronic stateswould be correlated. If fluctuations in electronic energiesof different pigments are correlated or almost synchro-nized, the EET process among the pigments would notexperience any noise or dynamic disturbance. In this
0 4 8-4
-4
0
4
8
low
high
q1
q2
FIG. 12: Adiabatic excitonic potential surface E−(q) givenby Eq. (5.19). The parameters are chosen to be Ω1 − Ω2 =100 cm−1, J12 = 100 cm−1, ωph = 53 cm−1, and λ1 = λ2 =500 cm−1. Contour lines are drawn at 50 cm−1 intervals. Thelocal minimum located around (q1, q2) = (4, 0) corresponds to|1〉 = |ϕ1e〉|ϕ2g〉, whereas that around (q1, q2) = (0, 4) is |2〉 =|ϕ1g〉|ϕ2e〉. The point of origin corresponds to the Franck-Condon state. Figure is reprinted with permission from Ref.[16].
situation, the phase coherence of the quantum mechan-ical wave packet involved in the EET process would bepreserved. In order to obtain further insight into thiseffect, it is also useful to consider the minimal model inEq. (5.16). Here we consider the following instead of Eq.(5.17) [16, 93]:
ε1(q) = ε0(q) + ~Ω1 − ~ωphdq1 − ~ωphζ1dq2, (5.20a)
ε2(q) = ε0(q) + ~Ω2 − ~ωphdq2 − ~ωphζ2dq1, (5.20b)
where the reorganization energy is expressed as ~λm =(1 + ζ2
m)~ωphd2/2, and the collective energy gap coordi-
nates can be expressed as um = −~ωphdqm − ~ωphζmdqn
for m 6= n. As a result, the cross-correlation function ofum(t) and un(t) (m 6= n) is
〈um(t)un(0)〉 = (ζm + ζn)Cqq(t), (5.21)
with Cqq(t) ≡ ~2ωphd2〈qm(t)qm(0). We note that this
cross-correlation function can be positive or negative, asdiscussed in the context of 2D-IR spectroscopy [57, 94];the situations of ζ1 + ζ2 > 0 and ζ1 + ζ2 < 0 corre-spond to the positive and negative correlations, respec-tively. Figure 13 illustrates how the dimer system dis-cussed in Fig. 12 varies for different extremes of envi-ronmental correlation. Similar to Fig. 12, the adiabaticexciton potential surfaces are drawn as a function of twophonon coordinates with minima representing two elec-tronic states. In the case of positive correlation, Fig.13a, the minima move into the same quadrant reducingtheir spacing and hence the energy barrier between them.A Franck-Condon transition to the origin lies above theenergy barrier resulting in a delocalized excited state pro-moting coherence, and hence the EET is robust againstenvironment-induced fluctuations [93]. However, for thenegative correlation case illustrated in Fig. 13b, the min-ima exist in opposite quadrants. The large energy barrier

19
0 4 8-4
-4
0
4
8
0 4 8-4
-4
0
4
8
(a) positive-correlation (b) negative-correlation
low
high
q2
q1 q1
FIG. 13: Adiabatic excitonic potential surfaces for the casesof (a) positive and (b) negative correlated fluctuations. Ineqn (5.20), we have chosen ζ1 = ζ2 = 0.5 for (a) and ζ1 =ζ2 = −0.5 for (b). The other parameters are the same as inFig. 12. Figure is reprinted with permission from Ref. [16].
between them results in a localized state upon excitationto the origin thus eliminating coherence, and hence theEET becomes slower or more inefficient because the EETmust overcome a higher energy barrier between the twostates [93].
In addition to providing insights into the electronic en-ergy transfer in photosynthetic light harvesting, the ob-served quantum beating is useful for identifying the PPCenergy landscape or the exciton energies [92]. Fouriertransforming the diagonal cut of the 2D nonrephasingspectra in Fig. 11(a) with respect to the waiting timeT produces a power spectrum. In Fig. 11(b), a portionof the power spectrum from the Chla region is shown.Peaks appearing at smaller beat frequencies indicate co-herence between excitons that arise from the same chloro-phyll variant. In Fig. 11(b), these low energy beatfrequencies exhibit strong coherence amongst Chla ex-citons. Meanwhile coherences between Chl-a and Chl-bexcitons are evident due to the presence of beat frequen-cies > 500 cm−1. In addition, as each exciton in LHCIIproduces a beating diagonal peak with unique frequencycomponents, the power spectrum allows the electronic en-ergy levels to be deconvolved. Beat frequency peaks alignvertically at positions corresponding to individual exci-ton energies. Analysis of the full power spectrum revealsa relatively evenly-spaced energy landscape in LHCIIwith a high energy Chl-a exciton (ω ∼ 15, 130 cm−1) lo-cated intermediate to the Chl-a and Chl-b bands. Theresulting small energy gap between the Chl-a and Chl-bexcited states and the coherences observed between thesebands facilitate ultrafast relaxation within LHCII [92].
D. P olarization dep endence of 2D sp ectra
1. Orientational Prefactor
As was shown in the above, the 2D rephasing, non-rephasing, and photon echo spectra involve averaging
over molecular orientations. In order to take into consid-eration the orientational average, we assume that the an-gular part of the PPC Hamiltonian, Eq. (2.3) are statis-tically separable from the electronic transition and exci-tation dynamics implicit in the ensemble average. There-fore, individual Liouville space pathways contributing to2D spectra, Eqs.(3.11)-(3.12), can be expressed as
Rα(t, T, τ)ELOE3E2E1 = YoriRα(t, T, τ). (5.22)
The prefactor of the right hand side, Yori, is the so-calledorientational prefactor expressed as
Yori = 〈(µ4 · ELO=4)(µ3 · E3)(µ
2 · E2)(µ1 · E1)〉ori,
(5.23)
with µi being the unit vector characterizing the transi-tion dipole orientation conjugated to the ithe laser pulse.Rα is a scalar part of Rα which describes the electronictransition and excitation dynamics. Interestingly, Eqs.(5.22) and (5.23) indicate that controlling the polariza-tion direction of each one of the excitation laser pulsesand the local oscillator enables us to manipulate individ-ual Liouville space pathways contributing to 2D spectraand to decongest crowded 2D spectra by selecting outcertain Liouville space pathways [58, 59, 95–101].
For evaluation of the orientational factor in Eq. (5.23),we let eX , eY , eZ and ex, ey, ez be the basis v ec-tors of the laboratory-fixed and molecule-fixed coordi-nate systems, respectiv ely. The molecular-fixed v ectorsex, ey, ez can be expressed with the laboratory-fixedbasis eX , eY , eZ with the help of the Euler angles, andthus the following formula is obtained [102]:
〈(eλ1· eI1
)(eλ2· eI2
)(eλ3· eI3
)(eλ4· eI4
)〉ori
=1
30
δλ1λ2δλ3λ4
δλ1λ3δλ2λ4
δλ1λ4δλ2λ3
T
4 −1 −1−1 4 −1−1 −1 4
δI1I2δI3I4
δI1I3δI2I4
δI1I4δI2I3
(5.24)
for λi ∈ x, y , z and Ii ∈ X , Y, Z. Because of µi =∑
λ(µi · eλ)eλ and Ei =∑
I(Ei · eI)eI , Eq. (5.23) canbe recast into
Yori =1
30
[
〈cos θ43 cos θ21〉(5 cos θ43 cos θ21 − Σ)
+〈cos θ42 cos θ31〉(5 cos θ42 cos θ31 − Σ)
+〈cos θ41 cos θ32〉(5 cos θ41 cos θ32 − Σ)]
(5.25)
with Σ ≡ cos θ43 cos θ21 + cos θ42 cos θ31 + cos θ41 cos θ32.In Eq. (5.25), θab is the angle between the ath and bthlaser pulse polarizations, whereas θαβ is the angle be-tween µα and µβ . The av erage ov er the distributions ofPPCs’ static structure 〈· · · 〉 has further been introduced.
A particularly useful choice of the polarization orien-tation is the sequence (π /3 [pulse 1], −π /3 [pulse 2], 0[pulse 3], 0 [pulse LO]), i.e.
θ43 = 0, θ42 = π /3, θ41 = π /3,
θ32 = π /3, θ31 = π /3, θ21 = 2π /3, (5.26)

20
For this sequence, the orientational prefactors for the Li-ouville pathways contributing diagonal peaks vanish, en-abling a closer look at the cross-peaks as the remaining[58, 99]. Therefore, this polarization sequence is termedthe cross-peak-specific sequence. However, we note thatdiagonal peaks in 2D nonrephasing spectra presentingelectronic quantum coherence, i.e. R1 in Fig. 10, do notnecessarily vanish because cos θ43 = cos θ21 = cos θ42 =cos θ31 = µα0 · µβ0 and cos θ41 = cos θ32 = 1.
The polarization sequence (π/4 [pulse 1], −π/4 [pulse2], π/2 [pulse 3], 0 [pulse LO]), i.e.
θ43 = π/2, θ42 = π/4, θ41 = π/4,
θ32 = 3π/4, θ31 = π/4, θ21 = π/2, (5.27)
also eliminates the prefactors for the rephasing Liou-ville pathways contributing diagonal peaks [99]. How-ever, the prefactors for the nonrephasing Liouville path-ways contributing the beating diagonal peaks do not van-ish, e.g. R1 in Fig. 10. Interestingly, this polariza-tion sequence eliminates all response terms except forthose arising from pathways where the system is in acoherence state during the population time, regardlessof whether the pathways contribute to diagonal or crosspeaks [58]. Thus, this polarization sequence may betermed the coherence-specific sequence.
Equation (5.25) leads to the well-known tensor rela-tion,
〈ZZZZ〉 = 〈ZZY Y 〉 + 〈ZY ZY 〉 + 〈ZY Y Z〉, (5.28)
where we have defined 〈I4I3I2I1〉 ≡ 〈(µ4 · eI4)(µ3 ·
eI3)(µ2 · eI2
)(µ1 · eI1)〉ori. Moreover, the orientational
prefactors for the cross-peak-specific polarization and se-quences in Eq. (5.26) and for the coherence-specific se-quence in Eq. (5.27) can be expressed as [96, 99]
3〈ZZY Y 〉 − 〈ZZZZ〉, (5.29)
〈ZY ZY 〉 − 〈ZY Y Z〉, (5.30)
respectively. Equations (5.28) and (5.29) play a role inthe polarization-phased 2D electronic spectroscopy forthe minor antenna complex CP29 of photosystem II [103],as will be discussed below.
2. Isolation of obscured features
The difference in scaling of individual peaks under dif-ferent polarization sequences can decongest crowded 2Dspectra. As seen in Sec. V B 3, the diagonal and off-diagonal regions of 2D spectra often contain multipleoverlapping peaks, where each peak may have contribu-tions from multiple Liouville pathways. Because many ofthe peaks arise from transition dipole moments with dif-ferent relative angles, their amplitudes scale differentlyunder the two polarization sequences. The cross-peak-specific polarization sequence described in the previoussection removes peaks corresponding to linear absorption
that lie along the diagonal. These peaks often have muchlarger amplitude than other pathways and so can domi-nate 2D spectra by obscuring weaker, near-diagonal fea-tures.
This approach also applies to the off-diagonal regionsof 2D spectra. In Fig. 14a and b, a feature that ap-pears to be a shoulder under the all-parallel polarizationhas been isolated into a separate peak under the cross-peak-specific polarization. After isolating this feature,it can then be analyzed to extract information aboutthe dynamics and structural origins of the correspond-ing energy transfer step. As seen in Fig. 14b, the donorstate has an energy of ∼ 15, 400 cm−1 and the acceptorstate has an energy of 15, 176 cm−1 (determined by fit-ting slices from the 2D spectra). As described in detailin Ref. [101], the 2D spectra showed that the acceptorstate was quickly populated, beginning at around 100 fs,and the excitation was observed to remain in this statefor the lifetime of the signal (5 ps). Due to the rela-tively rapid population from other Chl-b states and thevery slow depopulation, the acceptor state was assignedto be localized on Chl-b 605 (where numbering is fromthe structural model from x-ray crystallography) becauseonly this chlorophyll is coupled to other Chl-bs, but veryweakly coupled to the Chl-a band. While modeling basedon the crystal structure had suggested this chlorophyllas the site of a long-lived intermediate state, no directspectroscopic information had characterized the energytransfer processes into and out of the excited state local-ized mostly on Chl-b 605. As this example illustrates,rotation of the polarization of the incident laser beamscan selectively dissect crowded regions of 2D spectra.
3. Elucidating Exciton Geometry
In addition to examining the excited state dynamicsand observing the phenomena behind energy transfer,2D electronic spectroscopy can access information to bet-ter characterize the excited states. More specifically, theability to independently control the polarization of theincident laser pulses provides a means to measure theangle between the transition dipole moments of the ex-cited states that contribute to a given peak [59, 101].
The delocalized excited states, or the excitonic statesobserved spectroscopically, do not have an one-to-onecorrespondence with single chlorophylls. As discussed inSec. II, the excitonic states are delocalized excited statesdetermined by the coupling between chlorophylls and thetransition energies of the uncoupled chlorophylls, whichin turn depend on the surrounding protein environment.Figure 14c and d illustrate the case where each chloro-phyll contains an entirely localized transition (Fig. 14c)and the case where each transition contains contributionsfrom multiple chlorophylls (Fig. 14d). The drastic dif-ferences in position and direction between the two panelsindicate how the excitonic states often have forms whichare not obvious based on the structural model. Describ-

21
ing these states, therefore, requires spectroscopic infor-mation.
The orientational prefactor, or scaling factor, can bemeasured with polarized 2D spectroscopy [101]. EveryLiouville pathway, or molecular process, within a 2Dspectrum is scaled by an orientational prefactor, as dis-cussed in Sec. VD. As described previously, the mag-nitude of the orientational prefactor depends on the dotproduct of the angles between transition dipole momentsand angles between the electric fields of the incident laserpulses averaged over an isotropic ensemble. The orienta-tional prefactor for a peak in each one of a set of spec-tra can be found by amplitude changes in the peak dueto changes in laser pulse polarization. A comparison ofspectra under two known polarization sequences can de-termine the orientational prefactor for a peak in bothspectra [101].
c)
C hlb 605C hlb 606C hlb 607
d)
A cceptorD onor 1D onor 2
400 fs 400 fs
5000 fs5000 fs
a)
15000
14700
15500152001500014700
15500
15200
15000
14700
15500
15200
1500014700 1550015200
7
4
1
13
8
3
b)
ωτ (cm-1)ωτ (cm-1)
ωt (
cm-1
)
ωt (
cm-1
)
FIG. 14: a) Absolute v alue non-rephasing spectrum of LHCIIat T = 400 fs with (0,0,0,0) polarization of the incidentpulses. b) Absolute v alue non-rephasing spectrum of LHCIIT = 400 fs with (π /3,−π /3, 0, 0) polarization of the incidentpulses. c) T ransition dipole moments in the absence of cou-pling. d) T ransition dipole moments where the excitons arechanged by the presence of coupling. e) T ransition dipole mo-ments where the excitons are changed by both the presenceof coupling and different site energies.
This approach was applied to the spectra in Fig. 14aand b. Figure 14 contains absolute value, non-rephasing2D spectra of LHCII under ca) all-parallel (0,0,0,0) andb) cross-peak-specific (π/3,−π/3, 0, 0) polarization con-ditions for T = 400 fs. The spectra are again charac-terized by many of the same features shown in the all-parallel spectra of Fig. 9. The spectra are again dividedinto the Chl-a, intermediate, and Chl-b regions, with thestrongest peaks at the low energy Chl-a, the mid-energyChl-a and the Chl-b energies. While the congestion ofthe spectra means many amplitude changes are indis-cernible under the two polarization conditions, the peakhighlighted by an arrow has much larger amplitude inFig. 14b than in Fig. 14a. Therefore, the orientationalprefactor must be a large value under the cross-peak-
specific polarization sequence and a small one under theall-parallel one, because only where there is a large differ-ence in scaling would the spectra show a visible changein amplitude.
A measured orientational prefactor, for known polar-ization sequences, can be used to estimate the angle be-tween the transition dipole moments of the excitons in-volved in a given peak. Because the scaling factor de-pends on angles between transition dipole moments andon angles between laser pulse polarizations, if the scal-ing can be determined for known polarization sequences,the angles between transition dipole moments can be ex-tracted. In the case of the peak highlighted in Fig. 14aand b, the angle is the one between the donor and accep-tor states for this energy transfer cross-peak. Figure 15dplots the orientational prefactor for the two polarizationsequences shown for an energy transfer peak. Because ofthe increased relative amplitude in Fig. 14b, the anglebetween donor and acceptor states must be close to 90.Only for a small region around 90 would there be anvisible increase in amplitude relative to the many nearbypeaks [101].
4. Determination of site energies
Because the angles between transition dipole momentsdescribed in the previous section provide another observ-able for the system, the information can also access a newparameter of the system. Quantitatively relating struc-ture to function requires a mathematical description ofthe molecular system. Specifically, a model requires de-tailed information about the pigment Hamiltonian andthe pigment-protein coupling.
The diagonal elements of the pigment Hamiltonianvary because of differences in the protein pockets whichsurround the chlorophyll. These values are are the un-coupled transition energies (site energies). The site en-ergies depend on tuning of the local optical and electro-chemical properties by the protein environment througheffects such as electrostatic interactions with amino acidside chains, ligation of the central Mg of the chlorophyll,hydrogen bonding, conformation of the pigment (bowingof the porphyrin ring), and position and relative orien-tation of the chlorophyll to the electric field originatingfrom the dipole of the α-helix backbone.
Neither experimental nor theoretical methods havebeen able to easily access the values of the site energies.As described in Sec. II, the spectroscopic results dependnot only on the pigments themselves, but also on the cou-pling between the pigments and the protein. It is difficultto identify any one parameter of the PPC because of themultiple values which contribute to a spectrum. Also,because site energies differ based on the multiple para-meters described above, and all these values have similarmagnitude, the site energies are difficult to calculate the-oretically. Until now, there have been no spectroscopictechniques to access the site energies, and thus provide a

22
benchmark for the theoretical results.The measured angles between transition dipole mo-
ments can be used to find site energies. The direc-tions of the transition dipole moments depend on siteenergies, electrodynamic coupling between the chloro-phylls, and the position and direction of the transitiondipole moments of the individual chlorophyll. The latertwo parameters can be relatively straightforwardly cal-culated based on the x-ray crystallography structure.Because the site energies depend on many parameterswithin the protein matrix, the molecular structure doesnot help describe the contributions of each molecularcomponent. After having measured the angle betweentransition dipole moments, bounds can be placed on theremaining unknown, the site energies. The angle be-tween donor and acceptor excitons was found to be closeto 90, and the only site energy values which producethis angle are 15, 630 − 15, 710 cm−1 for Chl-b 606 and15, 680 − 15, 760 cm−1 for Chl-b 607 [101]. Figure 14dand e show pictorially why the angle between transitiondipole moments is an accurate method of determiningsite energies. In these two panels, the parameters usedfor coupling and site basis transition dipole moments arethe same, and only the site energies vary. The resul-tant drastic effect in angles between transition dipole mo-ments emphasizes why this angular information providesa measure of the site energy values. The reason why theangle between transition dipole moments is an effectiveway to determine site energies is because the angle be-tween transition dipole moments depends non-linearly onthe site energies. In contrast, excited state energies havea primarily linear dependence on site energies and thusare a much less sensitive measure.
In addition to better describing the system, determina-tion of the site energies provides insight into how the en-ergy landscape is created. Variations in site energies leadto variations in excited state energies. Thus, the site en-ergies are a critical component of how a single molecule,chlorophyll, is used in multiple complexes with differ-ent functions that combine for directional energy trans-fer. The breadth of site energies also serves to broadenthe excited state absorption bands, and therefore allowschlorophyll to absorb across more of the solar spectrum.
E. Comparison to Homology Models
The applications discussed in the preceding sections in-volved extracting information about the functionality ofthe complexes from the spectroscopic results and then re-lating those conclusions to the molecular structure usingmodels based on x-ray crystallography. For many com-plexes within the PSII supercomplex, however, a highresolution structural model has not yet been determined.Additionally, structure determination experiments, suchas x-ray diffraction, are done in conditions which may dif-fer significantly from physiological conditions. Becauseof the precision and breadth of information extracted
14500 15000 15500
14500
15000
15500
ωt (c
m)
ω τ (cm )
-1
-1
T=3.6 ps
0 20 40 60 80
0
0.1
0.2
0.3
relative dipole angle (degrees)
orie
ntat
iona
l fac
tor S
zzzz3S
zzyy -S
zzzz
Re3ZZYY-ZZZZ
ω
ReZZZZ
15000
15500
ωt (c
m)
-1
T=3.6 ps low Chl b15350 cm-1
high Chl b15650 cm-1
14500 15000 15500
14500
15000
15500
ωt (c
m)
ω τ (cm )
-1
-1
ReZZZZT=100 fs
1
0
-114500 15000 15500
14500
)
(cm-1)ω
a) b)
c) d)
FIG. 15: Real, 2D relaxation spectra of CP29 under all-parallel polarization for a) T = 100f s and b) T = 3.6ps.c) The subtraction-generated cross-peak-specific polarizationsequence for T = 3.6ps, with the two cross-peaks discussed inthe text highlighted by arrows. The difference in relativ e am-plitude reflects the difference in angles between the transitiondipole moments for the two donor-acceptor pairs. d) The ori-entational prefactors for the two polarization sequences shown(all-parallel and cross-peak-specific) as a function of the anglebetween transition dipole moments. The v alue of the orienta-tional prefactor is symmetric about 90. Figure is reprintedwith permission [103].
from a 2D experiment, the results can be used to de-termine functionality, and interpret the dynamics withinthe context of spatial structure, without the need forpreviously-acquired high resolution spatial information[83, 103, 104]. In this section, we discuss how these ap-proaches have been applied to the CP29 light-harvestingcomplex to determine the pathways of energy flow andto build a model of spatial positioning of excited states.Understanding CP29 has been challenging because untilthe recent structural model from x-ray crystallography[21], the structural model was based on partial sequencehomology with LHCII [105].
CP29 is one of the three so-called minor complexesthat sits between LHCII, where the majority of light-harvesting occurs, and the PSII core complexes, wherecharge separation tak es place. Experimental evidencesuggests CP29 contributes to light-harvesting and alsoplays a role in photoprotection [24]. According to thismodel, CP29 must absorb and transfer excitation energyand have mechanisms to dissipate excess energy. Basedon the x-ray crystallography information, CP29 contains13 chlorophyll, with 8 Chl-a and 4 Chl-b. As opposedto LHCII, CP29 has a mixed binding site, with a non-specific pock ets that can hold either Chl-a or Chl-b [21].
2DES was used to survey the relaxation dynamics forCP29. 2D spectra are presented in Fig. 15a-c. Similarlyto LHCII, the spectra show excitation of Chl-b, Chl-aand the region between the two bands. However, as seenboth in the 2D spectra of Fig. 15 and the linear absorp-tion spectrum of Fig. 1, CP29 exhibits less collectiveoscillator strength in the Chl-b region than is the case

23
for LHCII, which makes Chl-b to Chl-a energy transfermore challenging to observe. Relaxation from the Chl-bto the Chl-a band is still visible. As the CP29 spectrain Fig. 15a and b illustrate, relaxation occurs from lowenergy and high energy Chl-b states into the Chl-a bandgiving rise to two cross-peaks. Although these peaks ap-pear as a horizontal band in the all parallel spectrum,the additional structure provided by rotation of the po-larization of the laser pulses (Fig. 15c) highlights thattwo features with different relative amplitudes are indeedvisible. These two peaks are indicated with arrows inFig. 15c. Both of these transfer processes occur within3.6 ps. Within CP29, according to the structural model,there are four Chl-b molecules, which are part of threeChl-b/Chl-a clusters of chlorophyll, as shown in Fig. 1c.These two relaxation processes occur within these threeclusters.
Novel polarization experiments were used to generatethe spectrum of CP29 shown in Fig. 15. There are onlythree unique polarization sequences, as described in Sec.V D, and all other polarization sequences are a linearcombination of these three, as described in Eq. (5.25).For this experiment, these four polarization sequenceswere recorded, the three independent polarization se-quences and their linear sum. From the tensor relation ofEq. (5.25), the three unique spectra were normalized rel-ative to each other. This allowed arithmetic-generationof arbitrary polarization combinations. The CP29 polar-ized spectrum in Fig. 15c, generated from a linear com-bination of two of the recorded spectra, is data underthe same polarization sequence as that in Fig. 14b. Thisspectrum illustrates that this approach can be used tocreate spectra under all possible polarization sequences.This technique could be applied, for example, to scanthrough polarizations in order to selectively enhance andsuppress different peaks within congested off-diagonal re-gions, such as shown in Fig. 14a and b. A polarizationscanning approach has also been suggested using beam 3[106, 107].
The two Chl-b to Chl-a peaks can provide structuralinformation. From the two Chl-b to Chl-a peaks, usingthe same approach as described in Sec. V D 3, the an-gles within the heterodimer were determined to be 138
for the low energy Chl-b to Chl-a peak and 53 for thehigh energy Chl-b to Chl-a peak. Thus, there must betransition dipole moments of Chl-b and Chl-a which giverise to observed angles close to these values. With fur-ther analysis, this information will aid in localizing theobserved dynamics to specific pigments within the struc-tural model from x-ray crystallography and constrainingthe parameters of the excited states produced by thosepigments.
Because CP29 sits between LHCII and the reactioncenter, the complex functions as a pathway for energyflow, and when necessary, a site of energy dissipation.The strong Chl-b to Chl-a coupling allows energy trans-fer to the Chl-a states and thus the Chl-b molecules canbroaden absorption for auxiliary light-harvesting. The
Chl-a molecules, therefore, are located within the com-plex along a potential path from LHCII to the reactioncenter. This creates a chain of low energy states, or apath, for excitation energy to flow towards the site ofcharge separation. It is also interesting to note thatneighboring this path are carotenoid molecules, whichare thought to play a role in photoprotection [24, 108].The location of the carotenoids relative to the low energystates suggests that they are well-positioned for whenquenching mechanisms are triggered.
VI. CONCLUSIONS AND OUTLOOK
2DES measures the full, complex third order nonlinearresponse, and contained within this function is the infor-mation that reveals the ways in which molecular machin-ery is designed to give rise to ultrafast dynamics. Thismethod is particularly useful in systems that have struc-tures with multiple components, such as PPCs whichcontain interacting chromophores that in turn interactwith a surrounding protein scaffold to produce migrationof excitation energy through the light-harvesting struc-ture. The spectral and temporal resolution provided by2DES, as well as additional information accessed by con-trol over polarization of the incident beams, provides ameans to dissect congested spectra to elucidate the over-all functionality of PPCs.
The groups of associated PPCs that form a PSII super-complex, exhibit a remarkable ability to harvest sunlightwith near unity quantum efficiency. Understanding therelationship between the molecular structure and light-harvesting functionality of PSII is critical in the contextof our growing need for devices that provide renewableenergy sources. 2DES provides a tool to investigate howmolecular structure produces spatio-temporal dynamicsin a system as sophisticated and varied as the PSII super-complex. In addition to the studies of LHCII and CP29described here, 2DES applied to the PSII reaction cen-ter, which is highlighted in the structural model shownin Fig. 1a, displayed both ultrafast energy transfer, andelectron transfer on a variety of timescales [83]. Thesedifferent spectroscopic features from the PSII reactioncenter, with the addition of electron transfer chains, un-derscore the complexity of the excited state dynamics inthe photosynthetic apparatus.
As advances in 2DES experimental and theoreticaltools continue, these new techniques have the potential tofurther disentangle the structure-function relationshipsthat underlie ultrafast dynamics in natural and artificialsystems. There are several promising new directions forthe method. V ariations in frequency, polarization, or-derings, and number of the incident pulses can accessdifferent information about the sample. Firstly, because2DES is sensitive to couplings, there are means to se-lectively investigate joint functionality of different typesof molecules, which often exhibit well-spaced resonances.Non-degenerate four wave mixing experiments can exam-

24
ine the coupling between these spectrally separated statesto directly probe the interaction between different typesof molecules. For example, chromophore-protein interac-tions can be studied directly in a background free mea-surement with a visible-IR two-color experiment. Sec-ondly, as discussed in Sec. V D, the ability to indepen-dently control the four incident pulses also allows po-larization shaping of the fields. Besides the polarizationschemes discussed in the previous sections, additional po-larization control, which is more challenging to imple-ment in the electronic regime, has been explored theo-retically and experimentally in the IR region [109–111].In addition, fifth order implementations of multidimen-sional spectroscopy offer the potential to directly probehigher order correlation functions, or non-gaussian effectsof the surrounding protein environments. Furthermore,different ordering of the incident pulses has recently beenshown to access electron correlation effects [112]. Finally,one of the most promising areas of development is con-structing an approach to access a microscopic picture ofhow individual systems function.
Experimentally, EET dynamics in photosynthesis havebeen investigated by synchronizing initial electronic exci-tation in the entire ensemble by means of ultrashort laserpulses. Detected signals in such experiments are electricfields generated by macroscopic polarizations defined asthe ensemble average of microscopic dipole moments, asexpressed in Eq. (3.2). For the analyses of the spectra,commonly used theoretical approaches are statistical me-chanical methods based on quantum master equations.In these approaches, the key quantity of interest is thereduced density matrix, i.e. the partial average of thetotal density matrix over the protein environmental de-grees of freedom. It is undeniable that the combinationof condensed phase laser spectroscopy and the reduceddensity matrix approaches have provided much useful in-sight into photosynthetic EET. However, in natural lightharvesting, the initial event is the absorption of one sun-light photon by a single PPC, followed by EET in thePPC independently of the ensemble averaged behavior.Considering the difference between EET dynamics in in-dividual PPCs and their ensemble average means suchensemble approaches might limit our understanding ofthe design principles because the resultant spectra andthe reduced density matrix both involve ensemble aver-ages which may wash out the microscopic details.
Despite the good agreement between the ensemble ex-periments and the ensemble averaged theories [7, 11, 31,79, 80, 91], it is still irresistible to ask questions such as
1) Which have we observed in 2DES, true quantummechanical decoherence or ensemble dephasing which isalso called “fake decoherence”[113]?
2) How does EET proceed in a single PPC? 3) Doesthe initial electronic quantum coherence just decay irre-versibly or can it recur? 4) Can the coherence appear ina PPC excited not by light, but by EET from anotherPPC? 5) How relevant are the laser-based results to pho-tosynthesis in natural sunlight? Providing a precise an-
swer to these questions requires experimental/theoreticalapproaches that look at the dynamics prior to the ensem-ble average. Recently, Ishizaki and Fleming [78] exam-ined such a theoretical approach by employing a mixedquantum/classical simulation which is termed the time-dependent self-consistent field (TDSCF) approach [114–116]. In the approach, only the electronic excitation wastreated quantum mechanically, while the environmentaldegrees of freedom were described as classical variables.Although the combination of two fundamentally differentdescriptions of nature provided by quantum and classicalmechanics might cause serious problems of consistency ingeneral, their simulations [78] showed encouraging agree-ment with results from the more accurate reduced densitymatrix approach [6] with respect to destruction of quan-tum coherent behavior when the ensemble average is con-sidered. This agreement implies the following points asa possible interpretation of the experimentally observedlong-lived electronic quantum coherence: 1) The behav-ior of the reduced density matrix can be interpreted asthe ensemble average of the quantum dynamics in indi-vidual PPCs, i.e. ensemble dephasing or “fake decoher-ence” rather than true decoherence. 2) The quantumcoherent motion may be robust under individual realiza-tions of the environment-induced fluctuations contrary tointuition obtained from the reduced density matrix. 3)The existence of electronic quantum coherence does notdepend on the method of preparing the electronic excita-tion, i.e. whether by laser pulse or natural sunlight pho-ton. 4) Experimentally detected static delocalized states(excitons) in the ensemble-averaged behavior indicatethe existence of wave-like EET dynamics in individualPPCs. Figure 16 presents EET dynamics in a four-sitesystem calculated from the mixed quantum/classical sim-ulation. The excitation Hamiltonian and the excitation-environment coupling parameters are given in the fig-ure caption. Sites 1-2 and sites 3-4 make two stronglycoupled dimers, while the two dimers are weakly cou-pled. This situation is not unusual in photosyntheticcomplexes. Figure 16(A) presents the ensemble averagedbehavior corresponding to the reduced density matrix. InFig. 16(A) we observe quantum coherent wave-like mo-tion between sites 1 and 2 up to 500 fs. However, EET inthe 3-4 dimer and between the dimers do not exhibit anycoherent motion; they are usually regarded as describ-able as incoherent diffusion. On the other hand, Fig.16(B) shows the EET dynamics influenced by a partic-ular realization of the environment-induced fluctuationswithout an ensemble average. Although the ensemble av-eraged behavior exhibits destruction of the wave-like mo-tion, the individual system EET does not. Comparisonbetween Figs. 16(A) and 16(B) clearly demonstrates thequantum coherent effects are washed out in the averageor dephasing mechanism. Interestingly, after the “inco-herent diffusion” from the 1-2 dimer to the 3-4 dimer,wave-like motion is intensified in the 3-4 dimer contraryto the intuition obtained from the reduced density ma-trix.

25
(A)
Po
pu
latio
n o
f S
ite
s
(B)
Time t (ps)
0
0.2
0.4
0.6
0.8
1
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2
0
0.2
0.4
0.6
0.8
1
0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 1.8 2
site 1
site 2
site 3
site 4
FIG. 16: Time evolution of populations in a model four-sitesystem. We set Ω1 = Ω2 = 200 cm, Ω3 = Ω4 = 100 cm,J12 = J34 = 100 cm, J23 = 20 cm; the other excitoniccouplings vanish. The other parameters are λm = 50 cm,τ rxn
m= 100 fs, and T = 300 K. The upper panel (A) presents
the ensemble averaged behavior, whereas the lower panel (B)shows an individual trajectory of the EET dynamics calcu-lated from the mixed quantum/classical simulation withoutthe ensemble average [78].
However, these are theoretical predictions. In order tofully elucidate photosynthetic EET, particularly possibleroles of electronic quantum coherence in photosyntheticlight harvesting, it is important to experimentally over-come the intrinsic coarse-grained nature of the electronicquantum coherence observed for the ensemble of PPCs.For this purpose, it is intriguing to explore a single photo-synthetic PPC or a dilute ensemble far from the thermo-dynamic limit. It is becoming possible to merge ultrafastspectroscopy and single-molecule detection [117–119], forexample, Brinks et al. [119] reported the observation ofvibrational coherence in individual molecules at ambienttemperature by means of the phase-locked spontaneouslight emission technique [120, 121]. Applications of thistechnique to detection of electronic quantum coherence inPPCs would provide further insights into photosyntheticEET and are currently in progress in our laboratory.
The photosynthetic apparatus of plants, algae and bac-teria is a complex system with emergent properties thatcannot be simply extrapolated from the properties of thecomponent parts. This consideration also applies at theindividual PPC level. An essential step on the path toa full understanding of the design principles is the ex-perimental determination of the energetic and electronic
landscapes on which the dynamics play out. Building onthe dramatic increase in knowledge of the spatial land-scapes of PPCs, 2DES has provided fascinating new in-sights into the complementary energetic and electroniclandscapes. With further application of basis set po-larization spectroscopy, multi-color 2DES, use of otherphase matching directions to study electron correlationeffects, and with development of improved methods tostudy the role of the protein motion, a full understand-ing of how these extraordinary examples of nature’s en-gineering may be within reach.
In a monograph entitled What is Life? [122],Schrodinger summarized the emerging molecular aspectsof biology as of 1944 from his perspective. The bookspearheaded the rise of molecular biology and had atremendous impact on the development of 20th-centurybiology. There he stated as follows: “... incredibly smallgroups of atoms, much too small to display exact sta-tistical laws, do play a dominating role in the very or-derly and lawful events within a living organism.” Fur-ther he said, “A single group of atoms existing only inone copy produces orderly events, marvelously tuned inwith each other and with the environment according tomost subtle laws.” These statements are both suggestiveand inspiring toward elucidation of the physical originsof the remarkably high quantum efficiency of photosyn-thetic EET. One needs to explore individual proteins em-bedding pigments not only as randomly fluctuating dis-sipative environments from the statistical point of viewas usual in the literature of photosynthetic EET, butalso as functional environments on the basis of atomic-level understanding. This standpoint should be generallysignificant to understand stochasticity and selectivity inmolecular systems, in particular to elucidate directional-ity dominating biophysical and biochemical events.
Acknowledgments
This work was supported by the Director, Office ofScience, Office of Basic Energy Sciences, of the U.S.Department of Energy under Contract. DE-AC02-05CH11231 and by the Chemical Sciences, Geosciencesand Biosciences Division, Office of Basic Energy Sciences,U.S. Department of Energy under contract DE-AC03-76SF000098. The authors would like to thank JahanM. Dawlaty and Doran Bennett for providing the datashown in Fig. 6. G.S.S.-C. is grateful for an AmericanFellowship from the American Association of UniversityWomen.
[1] S. Mukamel, Principles of Nonlinear Optical Spec-troscopy, Oxford University Press, New York, 1995.
[2] W. M. Zhang, T. Meier, V. Chernyak, S. Mukamel, J.
Chem. Phys. 108 (1998) 7763.[3] M. Yang, G. R. Fleming, Chem. Phys. 182 (2002) 163.[4] S. Jang, Y.-C. Cheng, D. R. Reichman, J. D. Eaves, J.

26
Chem. Phys. 129 (2008) 101104.[5] S. Jang, J. Chem. Phys. 131 (2009) 164101.[6] A. Ishizaki, G. R. Fleming, J. Chem. Phys. 130 (2009)
234111.[7] A. Ishizaki, G. R. Fleming, Proc. Natl. Acad. Sci. USA
106 (2009) 17255.[8] B. Palmieri, D. Abramavicius, S. Mukamel, J. Chem.
Phys. 130 (2009) 204512.[9] G. Tao, W. H. Miller, J. Phys. Chem. Lett. 1 (2010)
891.[10] P. Huo, D. F. Coker, J. Chem. Phys. 133 (2010) 184108.[11] M. Cho, H. M. Vaswani, T. Brixner, J. Stenger, G. R.
Fleming, J. Phys. Chem. B 109 (2005) 10542.[12] D. Zigmantas, E. L. Read, T. Mancal, T. Brixner, A. T.
Gardiner, R. J. Cogdell, G. R. Fleming, Proc. Natl.Acad. Sci. USA 103 (2006) 12672.
[13] R. van Grondelle, V. I. Novoderezhkin, Phys. Chem.Chem. Phys. 8 (2006) 793.
[14] H. Lee, Y.-C. Cheng, G. R. Fleming, Science 316 (2007)1462.
[15] M. Cho, Two-Dimensional Optical Spectroscopy, CRCPress, Boca Raton, 2009.
[16] A. Ishizaki, T. R. Calhoun, G. S. Schlau-Cohen, G. R.Fleming, Phys. Chem. Chem. Phys. 12 (2010) 7319.
[17] P. Hamm, M. Lim, R. M. Hochstrasser, J. Phys. Chem.B 102 (1998) 6123.
[18] J. P. Dekker, E. J. Boekema, Biochim. Biophys. Acta -Bioenergetics 1706 (2005) 12.
[19] J. Nield, J. Barber, Biochim. Biophys. Acta - Bioener-getics 1757 (2006) 353.
[20] Z. F. Liu, H. C. Yan, K. B. Wang, T. Y. Kuang, J. P.Zhang, L. L. Gui, X. M. An, W. R. Chang, Nature 428(2004) 287.
[21] X. Pan, M. Li, T. Wan, L. Wang, C. Jia, Z. Hou,X. Zhao, J. Zhang, W. Chang, Nat. Struct. Mol. Biol.,in press.
[22] H. van Amerongen, L. Valkunas, R. van Grondelle, Pho-tosynthetic Excitons, World Scientific, Singapore, 2000.
[23] H. Kirchhoff, S. Lenhert, C. Buchel, L. Chi, J. Nield,Biochemistry 47 (2008) 431.
[24] N. E. Holt, D. Zigmantas, L. Valkunas, X. P. Li, K. K.Niyogi, G. R. Fleming, Science 307 (2005) 433.
[25] P. Muller, X. P. Li, K. K. Niyogi, Plant Physiol. 125(2001) 1558.
[26] G. S. Schlau-Cohen, J. M. Dawlaty, G. R. Fleming,IEEE-J.S.T.Q.E., in press.
[27] T. Renger, V. May, O. Kuhn, Phys. Rep. 343 (2001)137.
[28] G. D. Scholes, Annu. Rev. Phys. Chem. 54 (2003) 57.[29] G. D. Scholes, G. R. Fleming, Adv. Chem. Phys. 132
(2005) 57.[30] M. E. Madjet, A. Abdurahman, T. Renger, J. Phys.
Chem. B 110 (2006) 17268.[31] J. Adolphs, T. Renger, Biophys. J. 91 (2006) 2778.[32] D. Andrews, C. Curutchet, G. Scholes, Laser Photonics
Rev. 5 (2010) 114.[33] V. May, O. Kuhn, Charge and Energy Transfer Dynam-
ics in Molecular Systems, 2nd Edition, WILEY-VCH,Weinheim, 2004.
[34] R. Kubo, M. Toda, N. Hashitsume, Statistical PhysicsII: Nonequilibrium Statistical Mechanics, Springer-Verlag, New York, 1995.
[35] G. R. Fleming, M. Cho, Annu. Rev. Phys. Chem. 47(1996) 109.
[36] T. Forster, Ann. Phys. 437 (1948) 55.[37] A. G. Redfield, IBM J. Res. Dev. 1 (1957) 19.[38] A. Ishizaki, G. R. Fleming, J. Chem. Phys. 130 (2009)
234110.[39] D. M. Jonas, Annu. Rev. Phys. Chem. 54 (2003) 425.[40] T. Brixner, T. Mancal, I. V. Stiopkin, G. R. Fleming,
J. Chem. Phys. 121 (2004) 4221.[41] R. R. Ernst, G. Bodenhausen, A. Wokaun, Principles of
Nuclear Magnetic Resonance in One and Two Dimen-sions, Clarendon Press, Oxford, 1987.
[42] W. S. Warren, A. H. Zewail, J. Chem. Phys. 75 (1981)5956.
[43] A. M. Weiner, S. D. Silvestri, E. P. Ippen, J. Opt. Soc.Am. B 2 (1985) 654.
[44] G. R. Fleming, Proc. Natl. Acad. Sci. USA 95 (1998)15161.
[45] W. P. de Boeij, M. S. Pshenichnikov, D. A. Wiersma,Annu. Rev. Phys. Chem. 49 (1998) 99.
[46] P. Hamm, M. Lim, R. M. Hochstrasser, Phys. Rev. Lett.81 (1998) 5326.
[47] M. D. Fayer, Annu. Rev. Phys. Chem. 52 (2001) 315.[48] M. Yang, G. R. Fleming, J. Chem. Phys. 110 (1999)
2983.[49] B. S. Prall, D. Y. Parkinson, G. R. Fleming, M. Yang,
N. Ishikawa, J. Chem. Phys. 120 (2004) 2537.[50] T. Mancal, G. R. Fleming, J. Chem. Phys. 121 (2004)
10556.[51] D. Y. Parkinson, H. Lee, G. R. Fleming, J. Phys. Chem.
B 111 (2007) 7449.[52] Y.-C. Cheng, H. Lee, G. R. Fleming, J. Phys. Chem. A
111 (2007) 9499.[53] I. V. Rubtsov, J. Wang, R. M. Hochstrasser, Proc. Natl.
Acad. Sci. USA 100 (2003) 5601.[54] J. A. Myers, K. L. Lewis, P. F. Tekavec, J. P. Ogilvie,
Opt. Express 16 (2008) 17420.[55] J. D. Hybl, A. A. Ferro, D. M. Jonas, J. Chem. Phys.
115 (2001) 6606.[56] M. Khalil, N. Demirdoven, A. Tokmakoff, Phys. Rev.
Lett. 90 (2003) 047401.[57] M. Khalil, N. Demirdoven, A. Tokmakoff, J. Phys.
Chem. A 107 (2003) 5258.[58] E. L. Read, G. S. Engel, T. R. Calhoun, T. Mancal,
T. K. Ahn, R. E. Blankenship, G. R. Fleming, Proc.Natl. Acad. Sci. USA 104 (2007) 14203.
[59] E. L. Read, G. S. Schlau-Cohen, G. S. Engel, J. Wen,R. E. Blankenship, G. R. Fleming, Biophys. J. 95 (2008)847.
[60] G. Bodenhausen, R. Freeman, R. Niedermeyer, D. L.Turner, J. Mag. Reson. 26 (1977) 133.
[61] T. Brixner, I. V. Stiopkin, G. R. Fleming, Opt. Lett. 29(2004) 884.
[62] M. L. Cowan, J. P. Ogilvie, R. J. D. Miller, Chem. Phys.Lett. 386 (2004) 184.
[63] K. Gundogdu, K. W. Stone, D. B. Turner, K. A. Nelson,Chem. Phys. 341 (2007) 89.
[64] L. J. Kaufman, J. Heo, L. D. Ziegler, G. R. Fleming,Phys. Rev. Lett. 88 (2002) 207402.
[65] A. D. Bristow, D. Karaiskaj, X. C. Dai, S. T. Cundiff,Opt. Express 16 (2008) 18017.
[66] U. Selig, F. Langhojer, F. Dimler, T. Lohrig,C. Schwarz, B. Gieseking, T. Brixner, Opt. Lett. 33(2008) 2851.
[67] D. Keusters, H. S. Tan, W. S. Warren, J. Phys. Chem.A 103 (1999) 10369.

27
[68] E. M. Grumstrup, S. H. Shim, M. A. Montgomery, N. H.Damrauer, M. T. Zanni, Opt. Express 15 (2007) 16681.
[69] L. Lepetit, M. Joffre, Opt. Lett. 21 (1996) 564.[70] A. A. Ferro, J. D. Hybl, S. M. G. Faeder, D. M. Jonas,
J. Chem. Phys. 111 (1999) 10934.[71] J. C. Hoch, A. S. Stern, NMR Data Processing, Wiley-
Liss, New York, 1996.[72] J. A. Davis, L. V. Dao, M. T. Do, P. Hannaford, K. A.
Nugent, H. M. Quiney, Phys. Rev. Lett. 100 (2008)227401.
[73] S. A. Kovalenko, N. P. Ernsting, J. Ruthmann, Chem.Phys. Lett. 258 (1996) 445.
[74] S. A. Kovalenko, A. L. Dobryakov, J. Ruthmann, N. P.Ernsting, Phys. Rev. A 59 (1999) 2369.
[75] M. Lorenc, M. Ziolek, R. Naskrecki, J. Karolczak, J. Ku-bicki, A. Maciejewski, Artifacts in femtosecond tran-sient absorption spectroscopy, Applied Physics B-Lasersand Optics 74 (1) (2002) 19.
[76] P. F. Tekavec, J. A. Myers, K. L. M. Lewis, F. D. Fuller,J. P. Ogilvie, Opt. Express 18 (2010) 11015.
[77] W. J. D. Beenken, M. Dahlbom, P. Kjellberg, T. Pul-lerits, J. Chem. Phys. 117 (2002) 5810.
[78] A. Ishizaki, G. R. Fleming, J. Phys. Chem. B, in press.[79] T. Brixner, J. Stenger, H. M. Vaswani, M. Cho, R. E.
Blankenship, G. R. Fleming, Nature 434 (2005) 625.[80] G. S. Engel, T. R. Calhoun, E. L. Read, T.-K. Ahn,
T. Mancal, Y.-C. Cheng, R. E. Blankenship, G. R.Fleming, Nature 446 (2007) 782.
[81] G. S. Schlau-Cohen, T. R. Calhoun, N. S. Ginsberg,E. L. Read, M. Ballottari, R. Bassi, R. van Grondelle,G. R. Fleming, J. Phys. Chem. B 113 (2009) 15352.
[82] E. Collini, C. Y. Wong, K. E. Wilk, P. M. G. Curmi,P. Brumer, G. D. Scholes, Nature 463 (2010) 644.
[83] J. A. Myers, K. L. M. Lewis, F. D. Fuller, P. F. Tekavec,C. F. Yocum, J. P. Ogilvie, J. Phys. Chem. Lett. (2010)2774.
[84] R. Agarwal, B. P. Krueger, G. D. Scholes, M. Yang,J. Yom, L. Mets, G. R. Fleming, J. Phys. Chem. B 104(2000) 2908.
[85] J. M. Salverda, M. Vengris, B. P. Krueger, G. D. Sc-holes, A. R. Czamoleski, V. Novoderezhkin, H. vanAmerongen, R. van Grondelle, Biophys. J. 84 (2003)450.
[86] V. I. Novoderezhkin, M. A. Palacios, H. van Amerongen,R. van Grondelle, J. Phys. Chem. B 109 (2005) 10493.
[87] S. Georgakopoulou, G. van der Zwan, R. Bassi, R. vanGrondelle, H. van Amerongen, R. Croce, Biochemistry46 (2007) 4745.
[88] K. Gibasiewicz, M. Rutkowski, R. Van Grondelle, Pho-tosynthetica 47 (2009) 232.
[89] Y.-C. Cheng, G. R. Fleming, J. Phys. Chem. A 112(2008) 4254.
[90] E. Collini, G. D. Scholes, Science 323 (2009) 369.[91] G. Panitchayangkoon, D. Hayes, K. A. Fransted, J. R.
Caram, E. Harel, J. Wen, R. E. Blankenship, G. S. En-gel, Proc. Natl. Acad. Sci. USA 107 (2010) 12766.
[92] T. R. Calhoun, N. S. Ginsberg, G. S. Schlau-Cohen, Y.-C. Cheng, M. Ballottari, R. Bassi, G. R. Fleming, J.Phys. Chem. B 113 (2009) 16291.
[93] A. Ishizaki, G. R. Fleming, New J. Phys. 12 (2010)055004.
[94] N. Demirdoven, M. Khalil, A. Tokmakoff, Phys. Rev.Lett. 89 (2002) 237401.
[95] S. Woutersen, P. Hamm, J. Phys. Chem. B 104 (2000)
11316.[96] R. M. Hochstrasser, Chem. Phys. 266 (2001) 273.[97] M. T. Zanni, N.-H. Ge, Y. S. Kim, R. M. Hochstrasser,
Proc. Natl. Acad. Sci. USA 98 (2001) 11265.[98] O. Golonzka, A. Tokmakoff, J. Chem. Phys. 115 (2001)
297.[99] J. Dreyer, A. M. Moran, S. Mukamel, Bull. Korean
Chem. Soc. 24 (2003) 1091.[100] D. Abramavicius, D. V. Voronine, S. Mukamel, Biophys.
J. 94 (2008) 3613.[101] G. S. Schlau-Cohen, T. R. Calhoun, N. S. Ginsberg,
M. Ballottari, R. Bassi, G. R. Fleming, Proc. Natl.Acad. Sci. USA 107 (2010) 13276.
[102] D. P. Craig, T. Thirunamachandran, Molecular Quan-tum Electrodynamics, Academic Press Inc., London,1984.
[103] N. S. Ginsberg, J. A. Davis, M. Ballottari, Y.-C. Cheng,R. Bassi, G. R. Fleming, Proc. Natl. Acad. Sci. USA 108(2011) 3848.
[104] E. L. Read, G. S. Schlau-Cohen, G. S. Engel, T. Geor-giou, M. Z. Papiz, G. R. Fleming, J. Phys. Chem. B 113(2009) 6495.
[105] R. Bassi, R. Croce, D. Cugini, D. Sandona, Proc. Natl.Acad. Sci. USA 96 (1999) 10056.
[106] K.-K. Lee, K.-H. Park, S. Park, S.-J. Jeon, M. Cho, J.Phys. Chem. B 114, , in press.
[107] J.-H. Choi, M. Cho, J. Chem. Phys. 133 (2010) 241102.[108] S. Bode, C. C. Quentmeier, P. N. Liao, N. Hafi, T. Bar-
ros, L. Wilk, F. Bittner, P. J. Walla, Proc. Natl. Acad.Sci. USA 106 (2009) 12311.
[109] H. Rhee, Y.-G. June, J.-S. Lee, K.-K. Lee, J.-H. Ha,Z. H. Kim, S.-J. Jeon, M. Cho, Nature 458 (7236) (2009)310.
[110] J.-H. Choi, S. Cheon, H. Lee, M. Cho, Phys. Chem.Chem. Phys. 10 (2008) 3839.
[111] D. V. Voronine, D. Abramavicius, S. Mukamel, Biophys.J. 95 (2008) 4896.
[112] J. Kim, S. Mukamel, G. D. Scholes, Acc. Chem. Res. 42(2009) 1375.
[113] M. A. Schlosshauer, Decoherence and the Quantum-to-Classical Transition, Springer, Berlin, 2007.
[114] P. Ehrenfest, Z. Physik 45 (1927) 455.[115] N. F. Mott, Proc. Cambridge Philos. Soc. 27 (1931) 553.[116] G. D. Billing, The Quantum Classical Theory, Oxford
University Press, New York, 2003.[117] E. M. H. P. van Dijk, J. Hernando, J.-J. Garcia-Lopez,
M. Crego-Calama, D. N. Reinhoudt, L. Kuipers, M. F.Garcia-Parajo, N. F. van Hulst, Phys. Rev. Lett. 94(2005) 078302.
[118] I. Gerhardt, G. Wrigge, G. Zumofen, J. Hwang,A. Renn, V. Sandoghdar, Phys. Rev. A 79 (2009)011402.
[119] D. Brinks, F. D. Stefani, F. Kulzer, R. Hildner, T. H.Taminiau, Y. Avlasevich, K. Mullen, N. F. van Hulst,Nature 465 (2010) 905.
[120] N. F. Scherer, R. J. Carlson, A. Matro, M. Du, A. J.Ruggiero, V. Romero-Rochin, J. A. Cina, G. R. Flem-ing, S. A. Rice, J. Chem. Phys. 95 (1991) 1487.
[121] N. F. Scherer, A. Matro, L. D. Ziegler, M. Du, R. J.Carlson, J. A. Cina, G. R. Fleming, J. Chem. Phys. 96(1992) 4180.
[122] E. Schrodinger, What is Life? The Physical Aspectof the Living Cell, Cambridge University Press, Cam-bridge, 1944.

Gabriela Schlau-Cohen obtained her ScB in Chemical Physics from Brown University in
2003. She then joined the group of Prof. Graham Fleming as a doctoral student at the
University of California, Berkeley and the Lawrence Berkeley National Lab. Her
research interests focus on using non-linear spectroscopy to study photosynthetic energy
transfer, in particular the relationship between the structure and the efficient light-
harvesting functionality of pigment-protein complexes.
Akihito Ishizaki received his PhD in theoretical chemistry from Kyoto University, Japan
in 2008. He then joined the group of Prof. Graham Fleming at the University of
California, Berkeley as a JSPS Postdoctoral Fellow for Research Abroad. He is currently
a postdoctoral fellow at Lawrence Berkeley National Laboratory. His research interests
include open quantum systems, condensed phase chemical dynamics, modeling of optical
responses, and design principles of the primary steps in photosynthesis, especially as they
pertain to quantum coherence.
position which he assumed in April 2009. Fleming served as the Deputy Director of
Lawrence Berkeley National Laboratory from 2005 through 2007. Through joint
appointments as Melvin Calvin Distinguished Professor of Chemistry at UC Berkeley,
and Founding Director of both the Berkeley Lab's Physical Biosciences Division and UC
-shaped the
intersection of physical and biological sciences, while maintaining his own investigations
into ultrafast chemical and biological processes, in particular, the primary steps of
photosynthesis. Throughout his administrative career, Fleming has remained a highly
active scientific researcher. He has authored or co-authored more than 400 publications,
and is widely considered to be one of the world's foremost authorities on ultrafast
processes. In addition to his many other activities, Fleming has given numerous talks
around the world on the inter-relation and inter-complexity of energy, climate and
photosynthesis. In 2007, Fleming led the effort (with co-chair Mark Ratner) to define



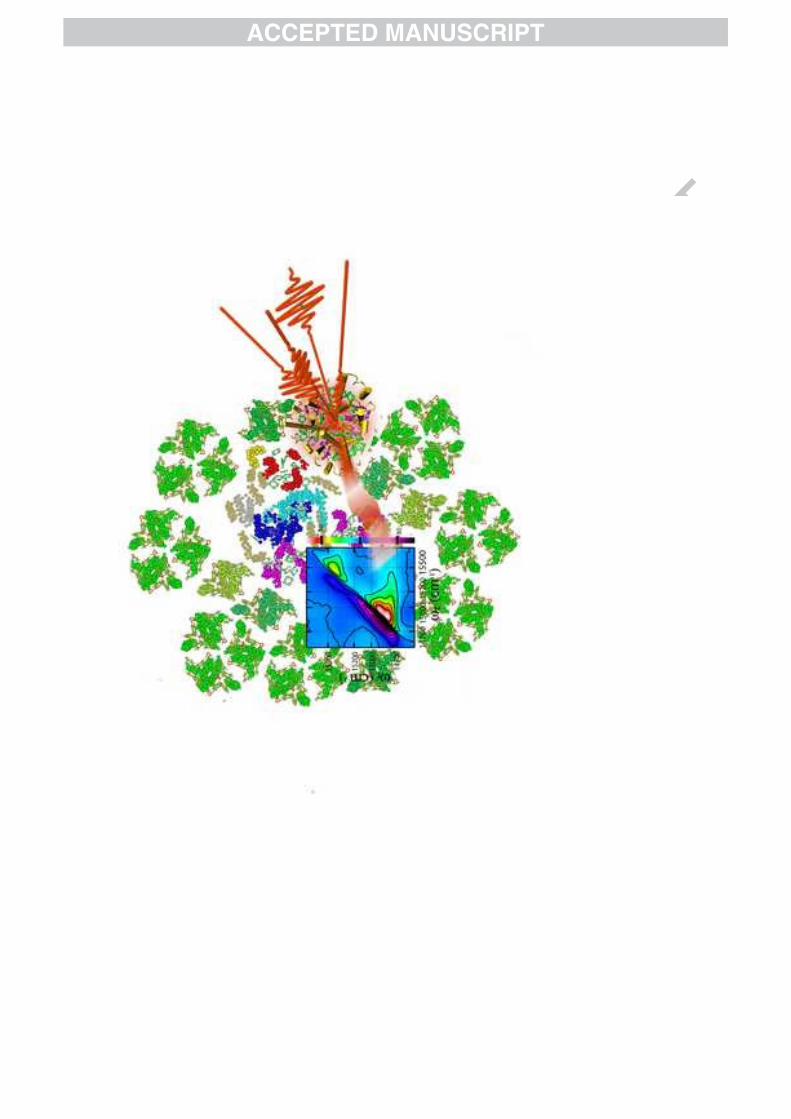

We review the theoretical principles and experimental implementation of 2D
electronic spectroscopy.
2D electronic spectroscopy can be applied to monitor energy transfer
dynamics, observe quantum coherence, determine excited state geometry,
and compare to homology models.
2D electronic spectroscopy reveals structure-function relationships in the
Photosystem II supercomplex.