transmission electron microscopy protocols for … · transmission electron microscopy protocols...
TRANSCRIPT

Transmission electron microscopy protocols for capsule visualisation in pathogenic respiratory and meningeal bacteria
M. Birkhead*,1 K. Ganesh1, K.M. Ndlangisa1 and H. J. Koornhof1,2 1 National Institute for Communicable Diseases, Private Bag X4, Sandringham, 2131, South Africa 2 Emeritus Professor, Department of Clinical Microbiology and Infectious Diseases, Faculty of Health Sciences, University
of the Witwatersrand, Johannesburg, South Africa
The human nasopharynx has a commensal bacterial flora which may become pathogenic (invasive), causing pneumonia, sepsis, meningitis and otitis media. A major virulence factor in this pathogenesis is the presence of a polysaccharide bacterial capsule, the production of which is controlled by a number of interacting variables (including temperature and capsule gene presence/expression/phase variation). Capsule composition is the basis of routine microbiological typing as well as the target of current pneumococcal vaccines. However, there is growing evidence that strains lacking a capsule (non-typeable strains) may also cause disease. Several different transmission electron microscopy protocols were tested on control cultures (known phenotypes) in order to visualise bacterial capsular material, ultimately for the phenotypic description of routinely grown, invasive, non-typeable isolates from the South African national laboratory-based, surveillance programme. Of these protocols, the most successful involved the use of ruthenium red, L-lysine acetate and an osmotically-adjusted buffer, though for some Gram-negative bacterial cells (those lacking sialic acid/heteropolymeric polysaccharides in their cell walls), it proved necessary to use a similar protocol but with the ruthenium red being replaced with another cationic dye, namely alcian blue (pyridine variant). The pyridine variant of alcian blue stained better than alcian blue 8XG, albeit somewhat indiscriminately (in that it was not specific for capsular polysaccharides). Identification and characterisation of the capsular status of these sorts of Gram-negative cells therefore need careful analysis of phenotypic information, and correlation with both genotypic data and environmental variables.
Keywords: transmission electron microscopy; bacterial capsules; ruthenium red; alcian blue; L-lysine acetate
1. Introduction
Invasive bacterial pathogens are disease-causing microorganisms transposed to sites of the host’s body that are usually sterile – such as blood, or the meningeal layer (meninges) covering the brain and spinal cord. Classically, invasive potential as a manifestation of virulence through immune evasion, has been linked to the presence of a polysaccharide capsule [1-6], although more recently, virulence through immune evasion has been shown to be mediated by non-capsular antigens [7] conferring invasive potential to non-encapsulated isolates [8-10]. It is the capsule that is used routinely for typing of pneumo- and meningo-coccal bacteria for diagnostic and surveillance purposes [11-13], and as capsular polysaccharides elicit antigenic responses in the human host that confer type-specific immunity, they have been used successfully as vaccine targets and epidemiological markers [7,14,15]. The bacterial capsule lies external to the cell wall and is visible at the electron microscope level as a delicate network of fine, anastomosing polysaccharide fibrils [16,17], the production of which is dependent upon the interaction of both environmental variables [18-25] and genetic traits [9,26-34]. Given that extracellular, capsular material is composed of highly hydrated polymers of monosaccharides or heteropolysaccharides [6,19,35], visualisation of this fragile substance using transmission electron microscopy (TEM) is difficult. Electron microscopists have addressed this challenge with a variety of fixation/immuno-electron microscopy protocols [33, 36-41], with the majority of TEM studies involving the use of cationic dyes in conjunction with fixatives, in order to simultaneously stain and stabilise the anionic capsular material. The most frequently used dyes have been ruthenium red and alcian blue, with or without en bloc staining with tannic acid, uranyl acetate or lanthanum [16,42-49], but even after careful TEM processing with a cationic dye, the fragile nature of the capsule can result in the apparent absence of capsular material in up to 70% of encapsulated cells [50]. Ruthenium red molecules act as both catalysts for the reduction of osmium tetroxide [51], and as a fixative for the highly acidic, sialylated lipo-oligosaccharides / lipopolysaccharides of the outer wall layer of Neisseria, which then react with the osmium [52]. However, use of ruthenium red+osmium tetroxide did not necessarily demonstrate capsule presence in Neisseria meningitidis serogroup W [53] or the enteric pathogen Campylobacter jejuni [19], but the addition of a diamine such as L-lysine acetate, to the ruthenium red-buffered fixative, produced improved capsular imaging of a variety of bacteria, for both transmission and scanning electron microscopy [35,42,49]. This variation in ruthenium red capsular staining success for TEM may be due to differences in bacterial cell wall composition, as this is the basis for differential staining of Gram-positive and Gram-negative bacteria at the light microscope level [54]. In Gram-positive cells, lipo- and wall-teichoic acids (which are anionic glycopolymers), stabilise the cell walls by attracting cations such as magnesium and sodium [55]. Similarly, it was shown that capsule is the major determinant of zeta potential (net surface charge that mediates the electrostatic interaction between particles), and so, surface charges vary according to serotype [24]. The Gram-negative wall consists of an inner membrane,
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
628
___________________________________________________________________________________________

peptidoglycan layer and an outer membrane which has phospholipids on its inner surface and lipid A (hydrophobic anchor for lipopolysaccharide) on the outer surface [56]. The lipopolysaccharides have a surface-exposed oligosaccharide chain, which may be linked further to a longer O antigen chain composed of repeating sugar subunits [57]. There is good correlation between the strength of the anionic charge, intensity of capsular staining, invasive ability, and the amount of surface-exposed sialic acid and lipo-oligosaccharides, with incorporation of sialic acid into surface structures also protecting the pathogen against innate and adaptive immune responses [34, 57-59]. For TEM, alcian blue has been used for capsular staining of Gram-negative bacteria with more success than ruthenium red [16,19,44,50,53], although there are few observations made on Gram-positive bacterial capsules stained with alcian blue [18] given the relative success of ruthenium red staining. The use of alcian blue in aldehyde fixatives is known to precipitate mucopolysaccharide-protein complexes and acid mucopolysaccharides resulting in electron dense staining of material [48], with different formulations of alcian blue providing different staining intensities [60]. In terms of interpretation of bacterial capsule staining, it is relevant to note that alcian blue staining of Columbia agar alone occurs, while staining of Escherichia coli pili, C-polysaccharides and mucoid-antigen, has also been described [18,19]. In this study, a number of different preparative protocols were assessed for staining of Gram-positive and Gram-negative bacteria, ultimately for describing the ultrastructure of selected South African bacterial isolates causing invasive disease.
2. Materials and Methods
2.1 Isolates
Known encapsulated cultures included: Haemophilus influenzae type b (ATCC®-33533 TM); Neisseria meningitidis serogroups A (ATCC®-13077 TM), B (ATCC®-13090 TM), W (ATCC®-35559 TM), Y (ATCC®-35561 TM) Bordetella pertussis (ATCC®-9797 TM); Streptococcus pneumoniae 19F (ATCC® BAA-657 TM). Additional serotypes of S. pneumoniae (serotypes 1, 8, 18C), collected through GERMS-SA (Group for Enteric, Respiratory and Meningeal Disease Surveillance in South Africa) which is a national, laboratory-based surveillance system for invasive pneumococcal disease in South Africa [12], were included. S. pneumoniae R6 (ATCC® BAA-255 TM) was used as a non-encapsulated control for Gram-positive cells, while Neisseria lactamica (ATCC®-23970 TM) was used as a non-encapsulated control for Gram-negative isolates. All isolates were grown routinely at 37°C on agar plates (Diagnostic Media Products, Johannesburg, South Africa) prior to processing for electron microscopy: isolates of N. meningitidis, N. lactamica and S. pneumoniae on 5% horse blood agar plates for 24 hours; H. influenzae on chocolate agar for 24 hours; B. pertussis for 48 hours on charcoal agar. Selection and development of an acceptable TEM protocol for capsular staining was performed using at least one encapsulated and one non-encapsulated control isolate per protocol. Given the difficulty in capsule visualisation in Gram negative bacteria, initial emphasis (Protocols 2.2.1 and 2.2.2 below) was on an encapsulated N. meningitidis serogroup, W, as this is currently the most commonly identified meningococcal serogroup in South Africa [61]. Not all bacterial isolates were processed using the individual protocols described below.
2.2 Electron Microscopy
For all protocols, bacterial colonies were fixed in situ to avoid loss of capsular material [44]. As it would prove too costly to process streaked agar plates (9cm diameter) by flooding, glass polytop vials were used to punch out agar discs on which the exponentially growing colonies occurred. The agar+colony discs were processed in the vials until ethanol dehydration, at which point the colonies were gently scraped from the agar into 1.5ml Eppendorf safe-lockTM centrifuge tubes, and centrifuged for twenty minutes at 500 xg (4°C) between each subsequent stage of dehydration and infiltration. For all protocols, a Leica EM UC6 with a diamond knife was used for ultramicrotomy, and 70 nm sections on 0.25% formvar-coated, copper slot-grids were viewed at 80kV on a BioTwin Spirit transmission electron microscope (FEI Company, USA) fitted with a Quemesa CCD camera (Olympus, Germany). Only illustrative examples of selected isolates processed according to the various preparative protocols, are presented.
2.2.1 TEM protocol 1: simultaneous glutaraldehyde-osmium tetroxide fixation
The method of Franke et al. [62] was included as one of the earliest techniques considered to be superior for enhanced staining of polysaccharide-containing structures. All steps were carried out on ice to prevent precipitation of reduced osmium. A 4% concentration of glutaraldehyde stock solution was selected for primary fixation of one hour. Further processing was according to the published protocol, with the epoxy resin of choice being a low viscosity, epoxy resin (Agar Scientific, United Kingdom). Grids were double-stained with uranyl acetate and lead citrate [63].
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
629
___________________________________________________________________________________________

2.2.2 TEM protocol 2: pre-incubation with a cationic dye and en bloc staining
In the protocol described by Hayat [63], which is specifically for the extracellular matrix of bacteria, colonies are incubated at room temperature in 0.1 M cacodylate buffered-0.15% ruthenium red for one hour, prior to fixation with cacodylate-buffered 3.6% glutaraldehyde containing 0.15% ruthenium red [63]. Following buffer rinses and post-fixation (1% osmium tetroxide with 0.15% ruthenium red), cells are encapsulated in 2% agar and stained en bloc in 0.5% uranyl acetate. Two deviations in the present study, from the original protocol were: the bacterial cells were re-suspended in uranyl acetate (not embedded in agar); pre-incubation alone was investigated (no uranyl acetate en bloc staining before dehydration and embedding, as this was considered unnecessary in subsequent publications [43]). An additional variation was to use alcian blue instead of ruthenium red, as Hendley et al. [44] had found alcian blue (in cacodylate buffer of pH 7.2) to provide more consistent staining than ruthenium red (in cacodylate buffer of pH 6.5), for the capsules of N. meningitidis. Pre-incubation with 1.5% alcian blue (pyridine variant, Sigma-Aldrich, South Africa, CAS 123439-83-8) in 0.1 M cacodylate buffer pH 7.05 with 0.1% acetic acid) was followed with and without en bloc uranyl acetate staining. All variations concluded with routine ethanol dehydration (30%, 50%, 70%, 90%, 100%), embedding in Agar Scientific low viscosity resin and double-staining with uranyl acetate and lead citrate.
2.2.3 TEM protocol 3: cationic dye and L-lysine acetate
This protocol was based on those developed by Jacques and Graham [49], Erlandsen et al. [35] and Hammerschmidt et al. [42]. As in Protocol 2.2.2 above, the present study additionally substituted ruthenium red with alcian blue (0.15% with 0.1% acetic acid in 0.1 M sodium cacodylate buffer pH 7.15). Two different forms of alcian blue, viz. alcian blue 8GX (Fluka CAS 75881-23-1) and the pyridine variant (Sigma-Aldrich CAS 123439-83-8), were tested. The use of 0.1 M sodium cacodylate buffer that had not been osmotically adjusted (as in Erlandsen et al. [35]) was also investigated, and as there are published images showing that omission of a diamine such as L-lysine acetate, resulted in lack of capsular preservation [53], inclusion of the diamine was unquestioned. However, as there were other minor differences from the Hammerschmidt et al. [42] method, a detailed protocol follows:
• 0.1 M sodium cacodylate buffer was prepared from deionised, distilled water with 0.09 M sucrose, 0.01 M CaCl2.2H2O, and 0.01 M MgCl2.6H2O. Buffer used in conjunction with 0.075% ruthenium red had a pH of 6.9, whilst the pH of buffer used with either of the alcian blue dyes was 7.15;
• Primary fixative consisted of buffer containing 2% methanol-free formaldehyde, 2.5% glutaraldehyde, and either 0.075% ruthenium red or 0.15% alcian blue.
• Agar discs with small colonies, were fixed for 20 minutes on ice, with primary fixative to which was added 1.55% L-lysine acetate;
• This was followed by primary fixative alone (no L-lysine acetate) for 3 hours, on ice in a refrigerator; • Three rinses of 20 minutes each, with chilled buffer containing the appropriate dye, ensuring that no
fixative was trapped beneath the agar disc in the polytop vial; • Post-fixation in 1% osmium tetroxide in dyed buffer, for one hour on ice; • Five rinses with chilled buffer with the appropriate dye, for 20 minutes each; • Dehydration in a chilled, graded ethanol series (10%, 20%, 35%, 50%, 70%, 90%, 100% x 4) at 30 minute
intervals. After the initial (10%) ethanol dehydration step, bacterial colonies were gently scraped off the agar discs, as the agar discs shrunk with increasing ethanol concentrations, resulting in the detachment of colonies which were then potentially lost during subsequent pipetting. Subsequent dehydration steps therefore included a 15 minute centrifugation at 500 xg, 4ºC. Samples were left in 70% ethanol overnight (on ice in a refrigerator), with dehydration continuing the following day;
• After the final 100% ethanol rinse, cells (refrigerated on ice) were infiltrated with 1:1 ethanol:London Resin White (medium) for 4 hours, 1:2 ethanol:resin overnight; three changes of pure resin at 12 hour intervals.
• Adequate pelleting of the cells after dehydration and during resin infiltration required increases in the time and speed of centrifugation, but this never exceeded a maximum of 40 minutes at 3000 xg;
• After the final centrifugation, the pelleted cells were transferred into gelatine capsules, topped up with fresh resin, and polymerised for 48 hours at 62ºC;
• Sections were stained for 5 minutes in saturated, aqueous uranyl acetate, being rinsed subsequently in distilled water and air-dried before viewing.
Where appropriate, further reference to the variations described for Protocol 3 above, will be abbreviated to: RR+L for ruthenium red and L-lysine; ABPV+L for alcian blue pyridine variant with L-lysine acetate, and AB8XG+L for alcian blue 8XG with L-lysine acetate.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
630
___________________________________________________________________________________________

2.2.4 TEM protocol 4: Negative staining for surface detail corroboration
During evaluation of TEM protocols 2 and 3, a number of Gram-negative isolates were seen to possess non-capsular, filamentous structures/regularly spaced buttons of osmiophilic material, when sectioned and stained. In order to elucidate these structures, living colonies from the same plate used for resin embedding, were mixed with a drop of phosphate buffered saline, adsorbed onto 0.3% formvar-coated, 300 mesh copper grids for 5 minutes, negatively stained for 5 seconds in 2% phosphotungstic acid (pH 6.8), rinsed in double distilled water, and air-dried before viewing.
3. Results
3.1 TEM protocol 1: simultaneous glutaraldehyde-osmium tetroxide fixation
Combined glutaraldehyde-osmium fixation on ice, failed to preserve any capsular material for staining in any encapsulated, Gram-negative isolate (Fig. 1a).
3.2 TEM protocol 2: pre-incubation with a cationic dye and en bloc staining
Pre-incubation of N. meningitidis serogroup W cells with ruthenium red, resulted in a fine stippling along the outer wall layer. At higher magnification, the stippling could be discerned as fairly regularly-spaced, osmiophilic buttons (Fig.1b). As expected, uranyl acetate en bloc staining enhanced ultrastructural contrast. Pre-incubation of these encapsulated Gram-negative cells with alcian blue (pyridine variant), resulted in discrete osmiophilic deposits around the cell walls, and depending on the plane of section, visible as finger-like processes extending outwards from the cells (Fig. 1c). The high concentration of alcian blue used for pre-incubation studies (1.5%) caused an intracellular, granular precipitate to be observed in many of the cells (Fig. 1c). Uranyl acetate en bloc staining improved the definition of the constituent thread-like structure of the projections (Fig. 1d), which were tentatively identified as clumped pili or fimbriae, on the basis of their appearance in negatively-stained cells from the same plates as were used for the sectioned material (Fig. 1e). More importantly, some of the cells pre-incubated with alcian blue and stained en bloc with uranyl acetate, had a fragile (discontinuous), crystalline crust (Fig. 1f) – although more than 95% of the cells showed no such detail.
3.3 TEM protocol 3: cationic dye and L-lysine acetate
Cells processed with RR+L, but without the use of an osmotically-adjusted buffer, suffered plasmolysis (Fig. 1g). In most of the Gram-negative, encapsulated isolates processed in this study, the RR+L protocol resulted in capsular staining, although this staining varied between genera and meningococcal serogroups (Figs. 1h-j). However, there was little evidence of capsule in cells of N. meningitidis serogroup A or Bordetella pertussis when stained with RR+L, especially when compared to the non-encapsulated N. lactamica control (Figs. 2a, 2d, 2f). The alternative use of alcian blue improved the visualisation of capsule in encapsulated Gram-negative cells in which ruthenium red failed to stain the capsular material (Figs. 2c, 2e), with ABPV+L staining more successfully than the AB8XG+L (Figs. 2b, 2c). However, staining with ABPV+L was not specific for capsular material and appeared indiscriminate in all Gram-negative isolates including the non-encapsulated control (Fig. 2g), so that osmiophilic material was found scattered throughout the sample. Additionally, adjacent cells within a sample could be differentially stained (Figs. 3a-c). Similarly, staining of the avirulent, laboratory-adapted R6 strain of S. pneumoniae (the Gram-positive, non-encapsulated control for the present study) with either ABPV+L or AB8XG+L, resulted in nonspecific staining suggestive of capsular material which was not evident in the cells stained with RR+L (Figs. 3d-f). Encapsulated cells of S. pneumoniae were consistently well stained with the RR+L protocol, though as observed in the Gram-negative encapsulated cells, there was variability in the appearance of the capsular material between different serotypes (Figs. 4a-e). There was also variability in the capsular material staining of cells from the same colonies in the same resin section (Figs. 4f-k). AB8XG+L staining of S. pneumoniae encapsulated cells resembled the RR+L staining more closely albeit with extracellular polysaccharides seemingly associated with other external cellular structures (Fig. 4l), but once again, the ABPV+L combination stained all polysaccharides indiscriminately (Figs. 4m, 4n).
3.4 TEM protocol 4: negative staining
Negative staining of whole cells revealed the presence of pili extending from the bacterial cell surface (Fig. 1e), although a clearly defined, capsular layer was not unequivocally present.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
631
___________________________________________________________________________________________

Fig. 1 Staining of encapsulated Gram-negative cells (Neisseria meningitidis serogroup W Figs. a-h; N. meningitidis serogroup B Fig. i; and Haemophilus influenzae type b Fig. j): (a) Protocol 1 (combined glutaraldehyde-osmium tetroxide) in which no capsule is evident; (b) Protocol 2 (pre-incubation with ruthenium red, ‘en bloc’ uranyl acetate staining) revealing osmiophilic nodules (arrows); (c) Protocol 2 (pre-incubation with alcian blue pyridine variant) revealing finger-like osmiophilic protrusions (arrows) from the cell surface, and the alcian blue, granular precipitate (arrowheads); (d) Protocol 2 (pre-incubation with alcian blue pyridine variant, ‘en bloc’ uranyl acetate staining) showing the twisted, fibrillar nature (arrows) of the finger-like protrusions. (e) Protocol 4 for negative staining of whole cells to show numerous pili and fimbriae extending from the cell periphery; (f) Protocol 2 (pre-incubation with alcian blue pyridine variant, ‘en bloc’ uranyl acetate staining) infrequently had a disrupted crystalline capsular layer, clearly different from the presumptive pili (arrows); (g) Plasmolysis of apparently non-encapsulated cells of N. meningitidis serogroup W, when processed according to Protocol 3 RR+L, but without the use of an osmotically adjusted buffer; (h) Protocol 3 RR+L (with osmotically adjusted buffer) produced a dense, crystalline capsular layer; (i) In serogroup B, Protocol 3 RR+L (with osmotically adjusted buffer) produced an osmiophilic, less crystalline, capsular layer with less distinct outer definition; (j) In a different genus (Haemophilus), Protocol 3 RR+L (osmotically adjusted buffer) produced a thick crystalline capsular layer. Scale bars: a, b, e, f, h, i, j = 200 nm; c, g = 300 nm; d = 25 nm.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
632
___________________________________________________________________________________________

Fig. 2 Gram-negative cells (Neisseria meningitidis serogroup A Figs. a-c; Bordetella pertussis Figs. d, e; N. lactamica Figs. f, g) processed with dye variations of Protocol 3: (a) Encapsulated cells with homopolymeric capsules lacking sialic acid, fail to demonstrate the capsular layer that is present, when stained with RR+L; (b) Encapsulated cells with homopolymeric capsules lacking sialic acid, show some capsular staining with AB8XG+L; (c) Encapsulated cells with homopolymeric capsules lacking sialic acid, show an extensive layer of osmiophilic exo-polysaccharides around each cell, with ABPV+L staining; (d) RR+L staining of the encapsulated cells of B. pertussis fails to demonstrate capsular material; (e) A distinct capsular layer is apparent in ABPV+L staining of cells from the same culture as in figure d; (f) The Gram-negative, non-encapsulated control cells showed no extracellular polysaccharide staining with RR+L; (g) The same culture processed in figure f, but stained with ABPV+L, shows staining of extracellular material trapped between cells, but no capsular layer around any of the cells. Scale bars all = 200 nm.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
633
___________________________________________________________________________________________
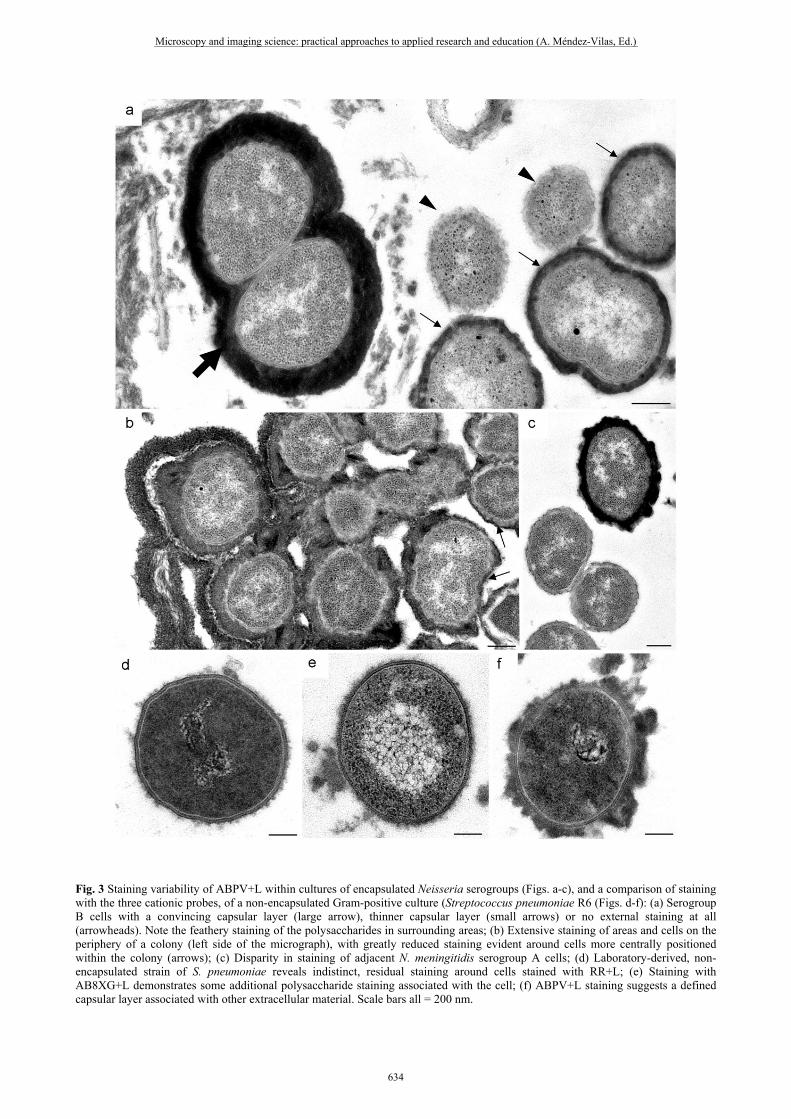
Fig. 3 Staining variability of ABPV+L within cultures of encapsulated Neisseria serogroups (Figs. a-c), and a comparison of staining with the three cationic probes, of a non-encapsulated Gram-positive culture (Streptococcus pneumoniae R6 (Figs. d-f): (a) Serogroup B cells with a convincing capsular layer (large arrow), thinner capsular layer (small arrows) or no external staining at all (arrowheads). Note the feathery staining of the polysaccharides in surrounding areas; (b) Extensive staining of areas and cells on the periphery of a colony (left side of the micrograph), with greatly reduced staining evident around cells more centrally positioned within the colony (arrows); (c) Disparity in staining of adjacent N. meningitidis serogroup A cells; (d) Laboratory-derived, non-encapsulated strain of S. pneumoniae reveals indistinct, residual staining around cells stained with RR+L; (e) Staining with AB8XG+L demonstrates some additional polysaccharide staining associated with the cell; (f) ABPV+L staining suggests a defined capsular layer associated with other extracellular material. Scale bars all = 200 nm.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
634
___________________________________________________________________________________________

Fig. 4 Staining variation between (Figs. a-e) and within (Figs. e-k) encapsulated Streptococcus pneumoniae serotypes using Protocol 3: RR+L (Figs. a-k); AB8XG (Fig. l) or ABPV+L (Figs. m, n): (a, b) Different isolates of serotype 1 both with densely crystalline capsules; (c) Serotype 18C with an exquisitely delicate, fibrous capsular layer; (d) Serotype 8 with an extensive capsular network of less dense, chunky, crystalline material; (e) Serotype 19F with a radiating filigree of capsular material; (f) Serotype 19F with a more compact, thicker capsular layer than in Fig. e; (g - k) Cells of serotype 18C from the same culture, with (g) Capsular material being loosely fibrillar between cells (arrows), and more compacted into a discrete layer around terminal cells in the chain (arrowheads); (h) densely-staining polysaccharides surrounding a cell; (i) Tangential section through a cell, showing polysaccharide clumping associated with fibrillar material (arrows) extending from the cell wall; (j) Transverse section through apical region of terminal cell showing fine, reticulate capsular filigree; (k) Extracellular polysaccharides (arrows), more compacted than in the previous image, but which appear to be connected to the cell - whether or not this is capsular material lifting from the cell surface, is uncertain; (l, m, n) Serotype 19F stained with: (l) AB8XG+L; (m) ABPV+L, showing many concentric layers of different densities around the cell, as well as extraneous staining (arrows); (n) ABPV+L with good capsular staining, but also indiscriminate staining of clumps of pili (arrows) and other adherent polysaccharides. Scale bars: a, b, c, d, e, f, m, n = 200 nm; g, h, i, j, k = 100 nm.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
635
___________________________________________________________________________________________

4. Discussion
Protocols 1 and 2 failed to produce appropriate capsular staining, and although capsule was evident in a few Gram-negative, encapsulated cells pre-incubated with alcian blue and subsequently stained with uranyl acetate, this was the exception rather than the norm, and would certainly not be adequate for describing/verifying the capsular status of routinely-grown, surveillance isolates of N. meningitidis and S. pneumoniae. On the basis of the images in Hammerschmidt et al. [42], it was expected that Protocol 3 incorporating the diamine L-lysine acetate and a cationic dye (ruthenium red), in a slightly acidic, osmotically-adjusted buffer, would result in good staining of the capsules, particularly the Gram-positive S. pneumoniae cells. This was indeed the case, and results of the present study, particularly for the Gram-positive bacteria, support those of Hammerschmidt et al. [42]. However, the substitution of one cationic dye (ruthenium red), with another (alcian blue), led to non-specific staining that would complicate the phenotypic description of new isolates, so the use of alcian blue, particularly the pyridine variant, is not recommended for Gram-positive cells. This non-specific staining may result from the planar shape and relatively high molecular weight of the alcian blue molecules, and the reason for the more generalised staining with the pyridine variant (as opposed to the 8XG alcian blue), is probably due to differences in the ring structures of the two dyes. This is corroborated by the fact that another alcian blue variant (tetrakis (methylpyridimium) chloride alcian blue CAS119851-27-3) which was recommended by Redfern et al. [60] for intense staining of cartilage, has a similar ring structure to that of the pyridine variant (closed ring without a sulphur atom). As all isolates were grown and processed in the same ways, and controls stained as expected, the absence of capsule in an isolate could not be attributed to the fixation protocol or scraping of colonies from the agar prior to dehydration. However, there was rarely complete uniformity between cells within a sample, which corroborates previous studies in which groups of encapsulated cells surrounded by naked ones were described [44], while variation in capsular staining/preservation was attributed both to the age of the cells and to their position in the colony – peripheral cells being better preserved than those centrally within colonies [18, 19, 64]. It is recommended, therefore, that any surveillance isolate to be investigated, should not have been subjected to prolonged storage or multiple passages, and that a number of grids sectioned from different blocks of the same specimen are viewed and the results compared with genome sequencing data, before a final conclusion on capsular status can be made with conviction. In addition to the age/position-related capsule variation (clearly evident between cells of both N. meningitidis and S. pneumoniae isolates), the variability in capsular appearance between different serotypes/groups processed according to protocol 3 above, could be attributable at least partially to the composition of specific bacterial walls and capsules: of the Gram-negative cells investigated, H. influenzae type b and N. meningitidis W both have dense, crystalline capsular staining with RR+L – and both have polysaccharide capsules composed of heteropolymers (sialic acid-containing disaccharides (hexosyl-sugars) in N. meningitidis W [65] or linear acids of ribose, ribitol (a five-carbon sugar alcohol) and phosphate linked by phosphodiester bonds in H. influenzae type b [6]). In contrast, RR+L staining of homopolymeric polysaccharides is reduced (as evident in N. meningitidis serogroup B, constituted by (α2→8)-linked-N-acetyl-neuraminic acid), whilst the non-sialylated homopolymers show virtually no ruthenium red-stained capsular material at all (serogroup A having a non-sialic acid capsule of homopolymers of N-acetyl-D-mannosamine -1 – P).
5. Conclusions
Recommendations from this study include the use of the RR+L staining protocol first. This will enable visualisation of capsule in Gram-positive cells, but if it fails to show capsule in Gram-negative surveillance isolates (which potentially may be encapsulated), then the ABPV+L protocol should be followed. However, given the non-specific staining that may result from this method, the importance of concurrent processing of positive and negative control cultures, as well as the viewing of representative grids from at least 3 different portions of each specimen cannot be overemphasized. As a number of environmental variables are known to affect capsular expression, the application of any staining protocol for capsular visualisation will not necessarily yield conclusive results for surveillance isolates if these are grown routinely, as currently occurs in diagnostic laboratories. It is therefore recommended that for a complete synthesis of genotypic and ultrastructural characterisation, fresh isolates grown under different environmental conditions should also be ultrastructurally investigated.
Acknowledgements The authors appreciate the knowledge and time given in illuminating discussion and commentary on the text, of Sheilagh B Smit and Leonard H Damelin; and the assistance of staff from the Centre for Respiratory and Meningeal Diseases. This research was funded by the National Institute for Communicable Diseases, a Division of the National Health Laboratory Service. Monica Birkhead thanks her mentor, Micky, for being her microscopy muse.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
636
___________________________________________________________________________________________

References [1] Tzeng Y-L, Stephens DS. Epidemiology and pathogenesis of Neisseria meningitidis. Microbes and Infection 2000;2:687-700. [2] Chung JY, Wilkie I, Boyce JD, Townsend KM, Frost AJ, Ghoddusi M, Adler B. Role of capsule in the pathogenesis of fowl
cholera caused by Pasteurella multicida serogroup A. Infection and Immunity 2001;69(4):2487-2492. [3] Joyce EA, Chan K, Salama NR, Falkow S. Redefining bacterial populations: a post-genomic reformation. Nature Reviews
Genetics 2002;3:462-73. [4] Apicella MA. Neisseria meningitidis. In: Mandell GL, Bennett JE, Dolin RD, editors. Principles and practice of infectious
diseases. 7th ed. Philadelphia: Churchill Livingstone, Elsevier; 2010. p.2737-52. [5] Hill DJ, Griffiths NJ, Borodina E, Virji M. Cellular and molecular biology of Neisseria meningitidis colonisation and invasive
disease. Clinical Science 2010;118:547-64. doi:10.1042/CS20090513. [6] Winn W, Allen SD, Janda WM, Koneman EW, Procop GW, Schrekenberger PC, Woods GL. Koneman’s colour atlas and
textbook of diagnostic microbiology. 6th ed. Baltimore:Lippincott Williams and Wilkins; 2006. [7] Lamelas A, Harris SR, Röltgen K, Dangy J-P, Hauser J, Kingsley RA et al. Emergence of a new epidemic Neisseria
meningitidis serogroup A clone in the African meningitis belt: high-resolution picture of genomic changes that mediate immune evasion. mBio 5(5):e01974-14.doi:10.1128/mBio.01974-14.
[8] Johswich KO, Zhou J, Law DKS, St. Michael F, McCaw SE, Jamieson FB, et al. Invasive potential of nonencapsulated disease isolates of Neisseria meningitidis. Infection and Immunity 2012;80(7):2346-53.
[9] Park IH, Geno KA, Sherwood LK, Nahm MH, Beall B. Population-based analysis of invasive nontypeable pneumococci reveals that most have defective capsule synthesis genes. PLoS ONE 2014;9(5):e97825. doi:10.1371/journal.pone.0097825
[10] Keller LE, Robinson DA, McDaniel LS. Nonencapsulated Streptococcus pneumoniae: emergence and pathogenesis. mBio 2016;7(2):e01792-15.
[11] Austrian R. The quellung reaction, a neglected microbiologic technique. Mount Sinai Journal of Medicine 1976;43(6):699-709. [12] Huebner RE, Klugman KP, Matai U, Eggers R, Hussey G. Laboratory surveillance for Haemophilus influenzae type b,
meningococcal, and pneumococcal disease. Haemophilus Surveillance Working Group. South African Medical Journal 1999;89:924-925.
[13] Yazdankhah SP and Caugant DA. Neisseria meningitidis: an overview of the carriage state. Journal of Medical Microbiology 2004;53:821-32.
[14] Harrison OB, Claus H, Jiang Y, Bennett JS, Bratcher HB, Jolley KA, et al. Description and nomenclature of Neisseria meningitidis capsule locus. Emerging Infectious Diseases 2013;19(4):566-73.
[15] Koornhof HJ, Madhi SA, Feldman C, von Gottberg A, Klugman KP. A century of South African battles against the pneumococcus –‘the Captain of Death.’ South African Journal of Epidemiology and Infection 2009;24(4):7-19.
[16] Cassone A, Garaci E. The capsular network of Klebsiella pneumoniae. Canadian Journal of Microbiology 1977;23:684-9. [17] Erlandsen SL, Lei M, Martine-Lecave I, Dunny G, Wells CL. High resolution cryoFESEM of microbial surfaces. Microscopy
and Microanalysis 2003;9:273-278. [18] Reimann K, Heise H, Blom J. Attempts to demonstrate a polysaccharide capsule in Neisseria gonorrhoea. Acta Pathologica,
Microbiologica et Immunologica Scandinavica 1988;96(8):735-40. [19] Karlyshev AV, McCrossan MV, Wren BW. Demonstration of polysaccharide capsule in Campylobacter jejuni using electron
microscopy. Infection and Immunity 2001;69(9):5921-3. [20] Allegrucci M, Sauer K. Characterization of colony morphology variants isolated from Streptococcus pneumoniae biofilms.
Journal of Bacteriology 2007;189(5):2030-8. doi:10.1128/JB.01369-06. [21] Hathaway LJ, Brugger SD, Morand B, Bangert M, Rotzetter JU, Hauser C, et al. Capsule type of Streptococcus pneumoniae
determines growth phenotype. PLoS Pathogens 2012;8(3): e1002574. [22] Kang SO, Wright JO, Tesorero, Lee H, Beall B, Cho KH. Thermoregulation of capsule production by Streptococcus pyogenes.
PLoS ONE 2012;7(5): e37367. [23] Van Dyk JS, Low Ah Kee N, Frost CL, Pletschke BI. Extracellular polysaccharide production in Bacillus licheniformis SVD1
and its immunomodulatory effect. BioResources 2012;7(4):4976-93. [24] Li Y, Weinberger DM, Thompson CM, Trzcinski K, Lipsitch M. Surface charge of Streptococcus pneumoniae predicts serotype
distribution. Infection and Immunity 2013;81(12):4519-24. [25] Loh E, Kugelberg E, Tracy A, Zhang Q, Gollan B, Ewles H et al. Temperature triggers immune evasion by Neisseria
meningitidis. Nature 2013;502:237-40. [26] Swartley JS, Marfin AA, Edupugant S, Liu L-J, Cieslak P, Perkins B, et al. Capsule switching of Neisseria meningitidis.
Proceedings of the National Academy of Sciences USA 1977;94: 271-6. [27] Dolan-Livengood JM, Miller YK, Martin LE, Urwin R, Stephens DS. Genetic basis for nongroupable Neisseria meningitidis.
Journal of Infectious Diseases 2003;187:1616-28. [28] Hammerschmidt S, Hilse R, van Putten JP, Gerardy-Schahn R, Unkmeir A, Frosch M. Modulation of cell surface sialic acid
expression in Neisseria meningitidis via a transposable genetic element. European Molecular Biology Organization Journal 1996;15(1):192-8.
[29] Hammerschmidt S, Müller A, Sillmann H, Mühlenhoff M, Borrow R, Fox A et al. Capsule phase variation in Neisseria meningitidis serogroup B by slipped-strand mispairing in the polysialyltransferase gene (said): correlation with bacterial invasion and the outbreak of meningococcal disease. Molecular Microbiology 1996:20(6):1211-20.
[30] Spinosa MR, Progida C, Talà A, Cogli L, Alifano P, Bucci C. The Neisseria meningitidis capsule is important for intracellular survival in human cells. Infection and Immunity 2007;75(7):3594-3603.
[31] Uria MJ, Zhang Q, Li Y, Chan A, Exley RM, Gollan B et al. A generic mechanism in Neisseria meningitidis for enhanced resistance against bactericidal antibodies. Journal of Experimental Medicine 2008;205(6):1423-34.
[32] Salter SJ, Hinds J, Gould KA, Lambertsen L, Hanage WP, Antonio M et al. Variation at the capsule locus, cps, of mistyped and non-typable Streptococcus pneumoniae isolates. Microbiology 2012;158:1560-0. doi 10.1099/mic.0.056580-0
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
637
___________________________________________________________________________________________

[33] Melchiorre S, Camilli R, Pietrantoni A, Moschioni M, Berti F, Del Grosso M et al. Point mutations in wchA are responsible for the non-typeability of two invasive Streptococcus pneumoniae isolates. Microbiology 2012;158:338-44.
[34] Apicella MA. Nontypeable Haemophilus influenzae: the role of N-acetyl-5-neuraminic acid in biology. Frontiers in Cellular and Infection Microbiology 2012;2 doi:10.3389/fcimb.2012.00019
[35] Erlandsen SL, Kristich CJ, Dunny GM, Wells CL. High-resolution visualisation of the microbial glycocalyx with low-voltage scanning electron microscopy: dependence on cationic dyes. Journal of Histochemistry and Cytochemistry 2004;52(11):1427-35.
[36] Tzeng Y-L, Datta AK, Strole CA, Lobritz MA, Carlson RW, Stephens DS. Translocation and surface expression of lipidated serogroup B capsular polysaccharide in Neisseria meningitidis. Infection and Immunity 2005;73(3):1491-1505.
[37] Nevot M, Deroncele V, Lopez-Iglesias C, Bozal N, Guinea J, Mercade E. Ultrastructural analysis of the extracellular matter secreted by the psychrotolerant bacterium Pseudoalteromonas antartica NF3. Microbial Ecology 2006;51:501-7.
[38] Sorensen UBS, Blom J, Birch-Andersen A, Henrichsen J. Ultrastructural localisation of capsules, cell wall polysaccharide, cell wall proteins and F antigen in pneumococci. Infection and Immunity 1988;56(8):1890-6.
[39] Jacques M, Gottschalk M. Use of monoclonal antibodies to visualise capsular material of bacterial pathogens by conventional electron microscopy. Microscopy and Microanalysis 1997;3:234-8.
[40] Doern GV, Buckmire FLA. Ultrastructural characterisation of capsulated Haemophilus influenzae type b and two spontaneous nontypable mutants. Journal of Bacteriology 1976;127(1):523-35.
[41] Reese S, Guggenheim B. A novel TEM contrasting technique for extracellular polysaccharides in In Vitro biofilms. Microscopy Research and Technique 2007;70:816-22.
[42] Hammerschmidt S, Wolff S, Hocke A, Rosseau S, Müller E, Rohde M. Illustration of pneumococcal polysaccharide capsule during adherence and invasion of epithelial cells. Infection and Immunity 2005;73(8):4653-67.
[43] Graham L, Orenstein JM. Processing tissue and cells for transmission electron microscopy in diagnostic pathology and research. Nature Protocols 2007;2(10):2439-50.
[44] Hendley JO, Powell KR, Salomonsky NL, Rodewald RR. Electron microscopy of the gonococcal capsule. Journal of Infectious Diseases 1981;143(6):796-802.
[45] Hyams C, Camberlein E, Cohen JM, Bax K, Brown JS. The Streptococcus pneumoniae capsule inhibits complement activity and neutrophil phagocytosis by multiple mechanisms. Infection and Immunity 2010;78(2):704-15.
[46] Springer EL, Roth IL. The ultrastructure of the capsules of Diplococcus pneumoniae and Klebsiella pneumoniae stained with ruthenium red. Journal of General Microbiology 1973;74:21-31.
[47] Waller LN, Fox N, Fox KF, Fox A, Price RL. Ruthenium red staining for ultrastructural visualisation of a glycoprotein layer surrounding the spore of Bacillus anthracis and Bacillus subtilis. Journal of Microbiological Methods 2004;58:23-30.
[48] Shea SM. Lanthanum staining of the surface coat of cells. Its enhancement by the use of fixatives containing alcian blue or cetylpyridinium chloride. The Journal of Cell Biology 1971;51:611-20.
[49] Jacques M, Graham L. Improved preservation of bacterial capsule for electron microscopy. Journal of Electron Microscopy Technique 1989;11:167-9.
[50] Neo Y-L, Li R, Howe J, Hoo R, Pant A Ho S-Y et al. Evidence for an intact polysaccharide capsule in Bordetella pertussis. Microbes and Infection 2010;12:238-45.
[51] Jamieson GA, Pepper DS. Membrane glycoproteins of blood platelets. IN: Johnson SA (ed). The circulating platelet. Academic Press, New York: 1971; p189-207.
[52] Fox AJ, Curry A, Jones DM, Demarco de Hormaeche R, Parsons NJ, Cole JA et al. The surface structure seen on gonococci after treatment with CMP-NANA is due to sialylation of surface lipopolysaccharide previously described as a ‘capsule’. Microbiology and Pathogenesis 1991; 11:199-210.
[53] Findlow H, Vogel U, Mueller JE, Curry A, Njanpop-Lafourcade B-M, Claus H, et al. Three cases of invasive meningococcal disease caused by a capsule null locus strain circulating among healthy carriers in Burkina Faso. Journal of Infectious Diseases 2003;195:1071-7.
[54] Kubler-Kielb J, Coxon B, Schneerson R. Chemical structure, conjugation, and cross-reactivity of Bacillus pumilus Sh18 cell wall polysaccharide. Journal of Bacteriology 2004;186(20):6891-901.
[55] Brown S, Santa Maria Jr JP, Walker S. Wall teichoic acids of Gram-positive bacteria. Annual Review Microbiology 2013;67: doi:10.1146/annurev-micro-092412-155620.
[56] Beveridge TJ. Structures of Gram-negative cell walls and their membrane-derived vesicles. Journal of Bacteriology 1999; 181(16):4725-33.
[57] Genevrois S, Steeghs L, Roholl P, Letesson J-J, van der Ley P. The Omp85 protein of Neisseria meningitidis is required for lipid export to the outer membrane. European Molecular Biology Organization Journal 2003; 22(8):1780-9.
[58] Unkmeir A, Kämmerer U, Stade A, Hübner C, Haller S, Kolb-Mäurer A, et al. Lipooligosaccharide and polysaccharide capsule: virulence factors of Neisseria meningitidis that determine meningococcal interaction with human dendritic cells. Infection and Immunity 2002;70(5):2454-62.
[59] Winstel V, Kühner P, Salomon F, Larsen J, Skov R, Hoffmann W et al. 2015. Wall teichoic acid glycosylation governs Staphylococcus aureus nasal colonization. mBio 6(4):e00632-15. doi:10.1128/mBio.00632-15.
[60] Redfern BG, Wise LD, Spence S. An alternative alcian blue dye variant for the evaluation of fetal cartilage. Birth Defects Research. Part B, Developmental ad Reproductive Toxicology 2007;80(3):171-6.
[61] Du Plessis M, Moodley C, Mothibeli KM, Fali A, Klugman KP, von Gottberg A. Serogroup B meningococcus in South Africa. Journal of Clinical Microbiology 2012: doi:10.1128/JCM.00401-12
[62] Franke WW, Krien S, Brown RM. Simultaneous glutaraldehyde-osmium tetroxide fixation with post osmication. Histochemie 1969;19:162-4.
[63] Hayat MA. Basic techniques for transmission electron microscopy. 1st ed. Florida: Academic Press Inc; 1986. [64] Graham LL, Beveridge TJ. Evaluation of freeze-substitution and conventional embedding protocols for routine electron
microscopic processing of eubacteria. Journal of Bacteriology 1990;172(4):2141-9.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
638
___________________________________________________________________________________________

[65] Romanow A, Haselhorst T, Stummeyer K, Claus H, Bethe A, Mühlenhoff M et al. Biochemical and biophysical characterisation of the sialyl-/hexosyltransferase synthesizing the meningococcal serogroup W135 heteropolysaccharide capsule. Journal of Biological Chemistry 2013;288(17):11718-30.
Microscopy and imaging science: practical approaches to applied research and education (A. Méndez-Vilas, Ed.)
639
___________________________________________________________________________________________