torsional stress induces an s1 nuclease-hypersensitive site within
TRANSCRIPT

Proc. Nati. Acad. Sci. USAVol. 82, pp. 4018-4022, June 1985Biochemistry
Torsional stress induces an S1 nuclease-hypersensitive site withinthe promoter of the Xenopus laevis oocyte-type 5S RNA: gene :
(gene expression/transcription factor/DNA-protein interaction)
WANDA F. REYNOLDS AND JOEL M. GOTTESFELDDepartment of Molecular Biology, Research Institute of Scripps Clinic, 10666 North Torrey Pines Road, La Jolla, CA 92037
Communicated by James Bonner, February 22, 1985
ABSTRACT The internal promoter of the Xenopus laevisoocyte-type 5S RNA gene is preferentially cleaved by S1 andBal-31 nucleases in plasmid DNA. S1 nuclease sensitivity islargely dependent on supercoiling; however, Bal-31 cleaveswithin the 5S RNA gene in linear as well as in supercoiled DNA.The S1 nuclease-hypersensitive site is centered at position+48-52 of the gene at the 5' boundary of the promoter. ADNase I-hypersensitive site is induced at this position uponbinding of the transcription factor, TFIIIA, specific for the 5SRNA gene. The somatic-type 5S RNA gene promoter is notpreferentially cleaved by S1 nuclease or Bal-31 nuclease insupercoiled DNA, nor does TFIIIA induce a DNase I site atposition +50. This differential promoter response may berelated to a 4-fold difference in TFIIIA affinity between theoocyte and somatic 5S RNA genes.
The 5' flanking sequences of transcriptionally active genes inchromatin-are often hypersensitive to nucleases. This sensi-tivity extends not only to DNase I, micrococcal nuclease, andrestriction enzymes (1) but also to single-strand-specificreagents such as S1 nuclease and bromoacetaldehyde (2, 3).It has been suggested that hypersensitive regions correspondto regulatory sequences and may be sites of interaction witheffector proteins. Upstream elements that are hypersensitivein active chromatin are in some instances S1 nucleasehypersensitive in supercoiled DNA. The significance of thiscorrelation is made apparent by recent evidence that activechromatin is subject to torsional stress (4, 5).Examples of upstream elements that are hypersensitive as
supercoiled DNA include those of the chicken P-globin gene(2, 6), Drosophila heat shock protein 70 (7) and histone genes(8), sea urchin histone genes (9), adenovirus major late region(10), and the simian virus 40 origin/enhancer region (11).Although these findings are provocative, there is as yet littlecorrelation between hypersensitive sites (HSSs) insupercoiled DNA and known promoter elements. In fact,sequences giving rise to a HSS upstream of the Drosophilaheat shock protein 70 gene can be deleted without signifi-cantly affecting promoter function in vivo (12). Only in thecases of certain (T)ATA homologies (6, 10) and sequenceswithin the simian virus 40 origin/enhancer region (11) do suchHSSs correspond to identifiable promoter elements. To helpclarify the biological significance of S1 nuclease hypersensi-tive structures, a system is needed in which the promotersequences are well defined and specific transcription factorshave been identified. The Xenopus SS RNA genes providesuch a system. Deletion mutant analysis has delineated apromoter element internal to the 120-base-pair (bp) gene (13).A transcription factor, TFIIIA, specific for the 5S RNA genehas been shown to bind this promoter region (14). In thisreport, we present evidence that the oocyte-type 5S RNA
gene promoter adopts an S1 nuclease sensitive conformationin supercoiled DNA. This altered conformation may besimilarly induced or stabilized by TFIIIA in linear DNA.These findings provide a clear correlation between an S1nuclease-sensitive conformation and a promoter element.
METHODSDNAs. For DNase I "footprint" analysis, pXlo 3'A+56
was digested with EcoRI and end-labeled with polynucleotidekinase (Bethesda Research Laboratories) and ['y-32P]ATP.The DNA was secondarily digested with HindIII and the530-bp fragment containing the gene was isolated by poly-acrylamide gel electrophoresis. The fragment was labeled onthe coding strand, 60 bp 3' to the gene. pXlsll was digestedwith Hpa II and end-labeled with the Klenow fragment ofDNA polymerase and [a-32P]dCTP. The DNA was secondar-ily digested with HindIII and the 380-bp fragment containingthe gene was isolated from a polyacrylamide gel. The frag-ment was labeled on the coding strand, 107 bp 5' to the gene.
Digestion of DNA with S1 and Bal-31 Nucleases. DNA (1-2,ug) was digested with up to 1 unit of S1 nuclease (BethesdaResearch Laboratories or P-L Biochemicals) in a reactionvolume of 30 p.1 in a buffer containing 30 mM sodium acetateat pH 4.8, 50 mM NaCl, and 1 mM ZnCl2. After a 30-minincubation at 37°C, the reaction was stopped with EDTA (20mM). The DNA was purified, digested with an appropriaterestriction enzyme, electrophoresed in 1.2% agarose gels in1 x TAE buffer (40 mM Tris acetate, pH 7.8/2 mM EDTA),transferred to nitrocellulose filters, and hybridized with anick-translated probe abutting the restriction site (15).
Plasmids were digested with Bal-31 (Bethesda ResearchLaboratories) (units, as defined by the supplier, indicated infigures) for 1 min at 22°C in a 20-,ul reaction volume in a buffercontaining 20 mM Tris HCl at pH 8.1, 100 mM NaCl, 12 mMCaC12, 12mM MgCl2, and 1 mM EDTA. After extraction withphenol and precipitation with ethanol, the DNA was digestedand analyzed as described above.Mapping of S1 Nuclease Cutting Sites. pXlo8 (5-10 ,ug per
lane) was digested with S1 nuclease at low concentrations(0.1-0.2 unit/30 ,ul) for 30 min at 37°C. This nucleaseconcentration produced less than 5% linear molecules andensured a low level of nicking. The DNA was then digestedwith EcoRI and electrophoresed in a 1.2% agarose gel asabove. Sections of the agarose gel containing DNA fragmentsof interest (see Fig. 3) were excised and the DNA waselectroeluted and digested with HindIII. The DNA wasend-labeled with the Klenow fragment of DNA polymeraseand [a-32P]dATP, followed by electrophoresis in 6%acrylamide nondenaturing gels in TBE buffer (90 mM Trisborate, pH 8.3/2 mM EDTA). Labeled fragments wereexcised from the polyacrylamide gel, crush-eluted and elec-trophoresed in sequencing gels (7.6% acrylamide/0.4%bisacrylamide/8.3 M urea/ix TBE).
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
4018
Abbreviations: bp, base pair(s); HSS, hypersensitive site; TFIIIA,transcription factor specific for 5S RNA gene.
Proc. Natl. Acad. Sci. USA 82 (1985) 4019
DNase I Footprinting. TFIIIA was isolated from immatureoocytes as 7S particles consisting of TFIIIA in associationwith 5S RNA or as purified TFIIIA according to publishedprocedures (16, 17). Identical footprints were obtained with7S particles and purified TFIIIA. Singly end-labeled DNAfragments were incubated with saturating amounts of 7Sparticles in the presence of RNase A (10 gg/ml) for 10 minfollowed by DNase I digestion (2 pg/ml for 1 min at 220C).The DNA fragments were purified and electrophoresed onsequencing gels.
RESULTSS1 Nuclease Cleavage of 5S Plasmid DNA. We examined the
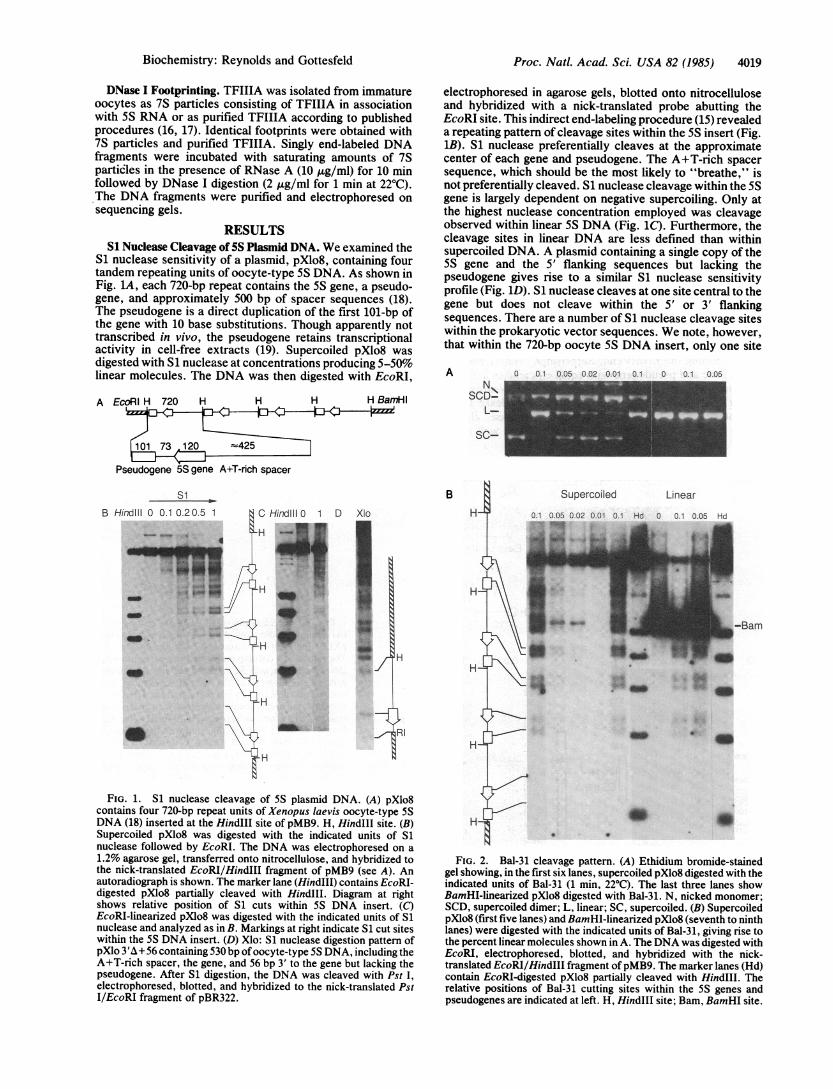
S1 nuclease sensitivity of a plasmid, pXlo8, containing fourtandem repeating units of oocyte-type 5S DNA. As shown inFig. lA, each 720-bp repeat contains the 5S gene, a pseudo-gene, and approximately 500 bp of spacer sequences (18).The pseudogene is a direct duplication of the first 101-bp ofthe gene with 10 base substitutions. Though apparently nottranscribed in vivo, the pseudogene retains transcriptionalactivity in cell-free extracts (19). Supercoiled pXlo8 wasdigested with S1 nuclease at concentrations producing 5-50%linear molecules. The DNA was then digested with EcoRI,
A EcoRI H 720 H H H H BamrHI
11073120 -425
Pseudogene 5S gene A+T-rich spacer
electrophoresed in agarose gels, blotted onto nitrocelluloseand hybridized with a nick-translated probe abutting theEcoRI site. This indirect end-labeling procedure (15) revealeda repeating pattern of cleavage sites within the 5S insert (Fig.1B). S1 nuclease preferentially cleaves at the approximatecenter of each gene and pseudogene. The A+T-rich spacersequence, which should be the most likely to "breathe," isnot preferentially cleaved. S1 nuclease cleavage within the 5Sgene is largely dependent on negative supercoiling. Only atthe highest nuclease concentration employed was cleavageobserved within linear 5S DNA (Fig. 1C). Furthermore, thecleavage sites in linear DNA are less defined than withinsupercoiled DNA. A plasmid containing a single copy of the5S gene and the 5' flanking sequences but lacking thepseudogene gives rise to a similar S1 nuclease sensitivityprofile (Fig. 1D). S1 nuclease cleaves at one site central to thegene but does not cleave within the 5' or 3' flankingsequences. There are a number of S1 nuclease cleavage siteswithin the prokaryotic vector sequences. We note, however,that within the 720-bp oocyte 5S DNA insert, only one site
A 0 0.1 0.05 0.02 0.01 0.1 0 0.1 0.05
NS.DSCD-
SC-
Si
Hindlil 0 1 D XIo
1.i
__=,_
w
H
FIG. 1. S1 nuclease cleavage of 5S plasmid DNA. (A) pXlo8contains four 720-bp repeat units of Xenopus laevis oocyte-type 5SDNA (18) inserted at the HindIII site of pMB9. H, HindIII site. (B)Supercoiled pXlo8 was digested with the indicated units of S1nuclease followed by EcoRI. The DNA was electrophoresed on a1.2% agarose gel, transferred onto nitrocellulose, and hybridized tothe nick-translated EcoRI/HindIII fragment of pMB9 (see A). Anautoradiograph is shown. The marker lane (HindIII) contains EcoRI-digested pXlo8 partially cleaved with HindIlI. Diagram at rightshows relative position of S1 cuts within 5S DNA insert. (C)EcoRI-linearized pXlo8 was digested with the indicated units of S1nuclease and analyzed as in B. Markings at right indicate S1 cut siteswithin the 5S DNA insert. (D) Xlo: S1 nuclease digestion pattern ofpXlo 3'A +56 containing 530 bp ofoocyte-type 5S DNA, including theA+T-rich spacer, the gene, and 56 bp 3' to the gene but lacking thepseudogene. After S1 digestion, the DNA was cleaved with Pst I,
electrophoresed, blotted, and hybridized to the nick-translated PstI/EcoRI fragment of pBR322.
BH 01
Ht
UH
H
H
Supercoiled Linear
-Bam
m
mlso
FIG. 2. Bal-31 cleavage pattern. (A) Ethidium bromide-stainedgel showing, in the first six lanes, supercoiled pXlo8 digested with theindicated units of Bal-31 (1 min, 22°C). The last three lanes showBamHI-linearized pXlo8 digested with Bal-31. N, nicked monomer;SCD, supercoiled dimer; L, linear; SC, supercoiled. (B) SupercoiledpXlo8 (first five lanes) and BamHI-linearized pXlo8 (seventh to ninthlanes) were digested with the indicated units of Bal-31, giving rise tothe percent linear molecules shown in A. The DNA was digested withEcoRI, electrophoresed, blotted, and hybridized with the nick-translated EcoRI/HindIII fragment of pMB9. The marker lanes (Hd)contain EcoRI-digested pXlo8 partially cleaved with HindIII. Therelative positions of Bal-31 cutting sites within the 5S genes andpseudogenes are indicated at left. H, HindIII site; Bam, BamHI site.
B
Biochemistry: Reynolds and Gottesfeld
-
S....fl
4020 Biochemistry: Reynolds and Gottesfeld
(two copies) is recognized and that site is within a regulatoryregion (as shown below).
Bal-31 Cleavage of 5S Plasmid DNA. Bal-31 nuclease hasendonuclease as well as exonuclease activity. Like S1nuclease, Bal-31 preferentially cleaves single-stranded DNA.Unlike S1, Bal-31 is active at physiological pH. SupercoiledpXlo8 was digested with Bal-31 at concentrations resulting in5-70% linear molecules (Fig. 2A), and this digestion wasfollowed by EcoRI digestion, electrophoresis, blotting, andhybridization as described above. As shown in Fig. 2B,Bal-31 preferentially cleaves within the 5S gene and pseudo-gene, producing a pattern similar to that obtained with S1nuclease, although the cleavage sites are broader. Thebreadth of the Bal-31 cleavage site could be due in part to theexonucleolytic activity of this enzyme. We also examined theBal-31 cleavage pattern on linear 5S plasmid DNA. To avoidexonucleolytic digestion at the EcoRI end, pXlo8 was initiallylinearized with BamHI (for restriction map, see Fig. LA),followed by Bal-31. The DNA was then secondarily digestedwith EcoRI, electrophoresed, blotted, and hybridized to theprobe abutting the EcoRI end. Surprisingly, Bal-31 cleavesthe 5S gene and pseudogene with equal efficiencies in linearand supercoiled DNA. Other enzymes such as Neurosporacrassa single-strand nuclease, micrococcal nuclease (datanot shown), and, to a lesser extent, S1 nuclease, cleavewithin the 5S gene and pseudogene in linear as well as in
supercoiled pXlo8. These enzymes, like Bal-31, appear torecognize a broader region than the S1 site in supercoiledDNA. The fact that several enzymes recognize a site withinthe 5S gene in linear DNA suggests a structural feature asopposed to a mere sequence preference.Mapping the S1 Nuclease Cutting Site. To map the S1
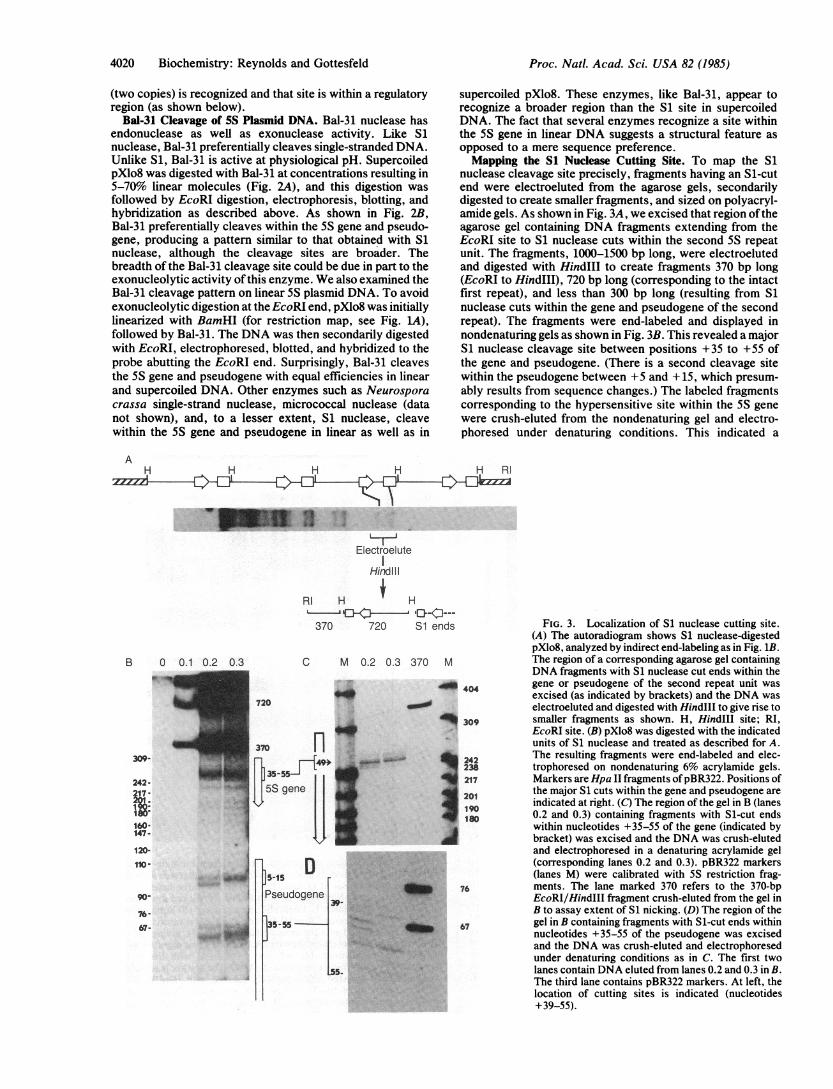
nuclease cleavage site precisely, fragments having an Sl-cutend were electroeluted from the agarose gels, secondarilydigested to create smaller fragments, and sized on polyacryl-amide gels. As shown in Fig. 3A, we excised that region oftheagarose gel containing DNA fragments extending from theEcoRI site to S1 nuclease cuts within the second 5S repeatunit. The fragments, 1000-1500 bp long, were electroelutedand digested with HindIII to create fragments 370 bp long(EcoRI to HindIII), 720 bp long (corresponding to the intactfirst repeat), and less than 300 bp long (resulting from S1nuclease cuts within the gene and pseudogene of the secondrepeat). The fragments were end-labeled and displayed innondenaturing gels as shown in Fig. 3B. This revealed a majorS1 nuclease cleavage site between positions +35 to +55 ofthe gene and pseudogene. (There is a second cleavage sitewithin the pseudogene between +5 and +15, which presum-ably results from sequence changes.) The labeled fragmentscorresponding to the hypersensitive site within the 5S genewere crush-eluted from the nondenaturing gel and electro-phoresed under denaturing conditions. This indicated a
AH H H H H RI
w _ ~~~~X,..IElectroelute
HindI I
RI H i H
370 720 Si ends
B 0 0.1
242-17
160* .147-
120-
110.
90-
76- 167-
oW
370 M
404
_ 309
242
238217
201190ISO8
76
67
FIG. 3. Localization of S1 nuclease cutting site.(A) The autoradiogram shows S1 nuclease-digestedpXlo8, analyzed by indirect end-labeling as in Fig. 1B.The region of a corresponding agarose gel containingDNA fragments with S1 nuclease cut ends within thegene or pseudogene of the second repeat unit wasexcised (as indicated by brackets) and the DNA waselectroeluted and digested with Hind1lI to give rise tosmaller fragments as shown. H, HindIII site; RI,EcoRI site. (B) pXlo8 was digested with the indicatedunits of S1 nuclease and treated as described for A.The resulting fragments were end-labeled and elec-trophoresed on nondenaturing 6% acrylamide gels.Markers are Hpa II fragments ofpBR322. Positions ofthe major S1 cuts within the gene and pseudogene areindicated at right. (C) The region of the gel in B (lanes0.2 and 0.3) containing fragments with Si-cut endswithin nucleotides +35-55 of the gene (indicated bybracket) was excised and the DNA was crush-elutedand electrophoresed in a denaturing acrylamide gel(corresponding lanes 0.2 and 0.3). pBR322 markers(lanes M) were calibrated with 5S restriction frag-ments. The lane marked 370 refers to the 370-bpEcoRI/HindIII fragment crush-eluted from the gel inB to assay extent of S1 nicking. (D) The region of thegel in B containing fragments with Si-cut ends withinnucleotides +35-55 of the pseudogene was excisedand the DNA was crush-eluted and electrophoresedunder denaturing conditions as in C. The first twolanes contain DNA eluted from lanes 0.2 and 0.3 in B.The third lane contains pBR322 markers. At left, thelocation of cutting sites is indicated (nucleotides+39-55).
Proc. Natl. Acad Sci. USA 82 (1985)

Proc. Natl. Acad. Sci. USA 82 (1985) 4021
cluster of minor cutting sites surrounding a distinct site at+48-52 (Fig. 3C) at the 5' boundary of the internal promoter.The sequences surrounding this site exhibit a moderatedegree of purine-pyrimidine asymmetry (Fig. 4B). Thenoncoding strand sequences extending from +47 to +61 arepurine rich (12/15), whereas those 5' of the site are pyrimi-dine rich (11/14). Fragments corresponding to S1 nucleasecleavage at the 5' boundary of the pseudogene promoter weresimilarly crush-eluted and displayed under denaturing condi-tions (Fig. 3D). Cleavage sites extend from +39 to 55,although S1 "nibbling" of breathing ends may create somelength heterogeneity. Also, the pseudogene sequence. differsfrom that of the gene at three positions within this region (Fig.4B) such that the exact pattern of cleavage may differ.
TFIIHA-Induced HSS at +50. As shown by DNase Ifootprinting (Fig. 4), TFIIIA binds the internal promoter from+45 to +95 (14). Within the coding strand footprint, TFIIIAinduces a DNase I-HSS at +49-51 within the oocyte-type 5Sgene (14). This suggests that TFIIIA binding might, likesuperhelical stress, induce an altered conformation at the 5'promoter boundary. We note that the +50 region within thenoncoding strand footprint is not hypersensitive (14). In-stead, enhanced cleavages are located at 60-64 and 74-76.
Nuclease Sensitivity of Somatic 5S DNA. In Xenopus, thereare two major families of 5S RNA genes, the oocyte type,present in 20,000 copies per haploid genome, and the somatictype, present in 400 copies. The oocyte 5S RNA genes are
A M 1 2
160 %147
45122 -15110 W
90 %
76,__ -95
67 ~mm
BTFIIIADNase
40 0w
Xlo G T C TG A T C T C A G A A G C G A T A C A G G G T
Si nucleaseXIS G T C T G A T C T C G G A A G C C A A G C A GG G T
XIo 4/ G T C T G A T C T jG A A G T G A T A C A G G G G
FIG. 4. DNase I-hypersensitive site in presence of TFIIIA. (A)DNase I footprint analysis of TFIIIA bound to oocyte 5S RNA genepromoter. A singly end-labeled (coding strand) fragment containingthe oocyte 5S gene was incubated in the absence (lane 1) or presence(lane 2) of TFIIIA and lightly digested with DNase I. The DNA waspurified and electrophoresed in a sequencing gel. An autoradiographis shown. TFIIIA protects a region extending from nucleotides +45to 95 of the gene with an enhanced DNase I site at nucleotide 49-51.The markers (M) are Hpa II-digested pBR322 fragments. Theposition of the DNase I-hypersensitive site was furthersorroboratedby comparison with a G-only cleavage of the 5S fragment (notshown). (B) Sequences are shown for nucleotides 37 to 62 of theXenopus laevis oocyte-type 5S gene (Xlo) (18), the somatic-type(Xls) (20), and the Xlo pseudogene (Xlo 4O) (18). In Xlo, the arrowabove indicates the position (+2) of the TFIIIA-induced DNase Icleavage site, The arrows below indicate the major S1 nucleasecleavage sites in supercoiled DNA. In Xls and Xlo 41, nucleotides thatdiffer from the Xlo sequence are underlined.
transcribed only during oogenesis and early development(21). The somatic-type genes are not developmentally regu-lated and are transcribed in all active cell types, includingoocytes. We examined the response of somatic 5S DNA tosuperhelical stress. pXlsll contains a single 883-bp repeatunit of X. laevis somatic SS DNA. The repeat contains onecopy of the gene and 760 bp of G+C-rich spacer sequences(20). Supercoiled pXlsll was digested with S1 nuclease orBal-31 and then with Sal I. The DNA was electrophoresed inagarose gels, blotted onto nitrocellulose, and hybridized witha probe abutting the Sal I site. As shown in Fig. SA, major S1nuclease sites are situated at approximately -150 and -25from the 5' end of the gene but not within the gene. Similarly,Bal-31 cleaves at or near the -150 site but does not recognizethe -25 region or the internal promoter. The two types of SSRNA genes have radically different upstream sequences. Theoocyte SS gene is flanked by A+T-rich sequences, whereasthe somatic 5S gene is flanked by G+C-rich sequencescontaining several regions of marked purine/pyrimidineasymmetry (18, 20). It is possible that the 5' flankingsequences could influence the conformation of the 5S pro-moters. For example, the sites upstream of the somatic 5Sgene could act as a sink for torsional stress, thus preventingemergence of an intragenic site. Alternatively, the absence ofa site within the somatic promoter may result from sequencechanges (Fig. 4B). The oocyte and somatic SS promotersdiffer in sequence at four positions (+47, 53, 55, and 56) nearthe 5' boundary. Interestingly, a TFIIIA footprint on thecoding strand of the X. laevis somatic 5S gene does notinclude an enhanced DNase I cleavage at +50 (Fig. 5B).
DISCUSSIONThe 5' boundary of the oocyte-type 5S RNA promoter iscleaved preferentially by S1 nuclease. This site centered at+48-52 is largely dependent on supercoiling. In contrast toS1 nuclease, Bal-31 cleaves within the 5S RNA gene in linearas well as supercoiled DNA. This indicates an unusual DNA
A Bal Si B - -
N wL_
-5
872- ,4, -_
OR~~~\_
603-O.- do-OR_
_. Al.
Lg5
50A5
FIG. 5. Nuclease sensitivity of somatic 5S DNA. (A) Bal-31 andS1 nuclease sensitivity. pXls11 contains a single HindIII repeat unit(883 bp) of X. laevis somatic 5S DNA inserted at the HindIII site ofpBR322 (20). Supercoiled pXls11 was digested with 0.1 unit of Bal-31(1 min 22°C) or 1 unit of S1 nuclease (30 min, 37°C) and then digestedwith Sal I. The DNA was electrophoresed, blotted, and hybridizedwith the nick-translated Sal I/HindIll fragment of pBR322. Markersare end-labeled Hae IlI-digested DNA of phage 4X174. The diagramindicates the relative position of the 5S DNA insert. (B) DNase Ifootprint analysis of TFIIIA bound to somatic 5S DNA. A singlyend-labeled (coding strand) fragment containing the somatic 5S RNAgene was incubated in the absence (-) or presence (+) of TFIIIA as7S particles and lightly digested with DNase I. The DNA fragmentswere isolated and displayed in sequencing gels. An autoradiograph isshown. TFIIIA protection extends from approximately +45 to 95.The +50 region is not hypersensitive.
Biochemistry: Reynolds and Gottesfeld

4022 Biochemistry: Reynolds and Gottesfeld
secondary structure present in linear molecules that is ac-centuated by torsional stress to become accessible to S1nuclease. A TFIIIA-induced DNase I site at +50 in linearDNA suggests that factor binding might trigger or stabilize asimilar conformational change.The question remains as to the nature of the S1 nuclease-
sensitive DNA conformation. The S1 site is dependent, forthe most part, on supercoiling. Since DNA unwinding re-lieves superhelical stress, unwinding probably accompaniessite formation. DNA configurations that result in unwindingand are associated with S1 HSSs include Z-DNA (22),cruciforms (23) and possibly DNA slippage along directrepeats (7). The sequences surrounding the 5S DNA HSS arenot compatible with these structures. The site is, however,situated within a region of purine-pyrimidine asymmetry, acharacteristic commonly associated with S1 nuclease sensi-tivity in or near regulatory DNA. Such asymmetry is thoughtto cause helical distortion, perhaps resulting in non-B-formDNA (11).The +50 HSS is situated within a region shown by deletion
analysis to be crucial for transcription initiation. Deletion ofthe first 50 bp of the Xenopus borealis somatic 5S RNA genestill resulted in 5S-sized transcripts that initiated withinvector sequences. Deletion to +55, however, allowed little tono transcription (13). This 50-55 region, though required fortranscription, is not essential for TFIIIA binding (24). Thosenucleotides that are essential and that make tight contact withthe factor are located at the 3' end of the promoter betweenpositions +70 and 90 (25). Nevertheless, TFIIIA interactionwith the 5' region of the promoter is important for geneactivity. Partial proteolysis of the 40-kDa factor has beenshown to result in three domains (17). A 20-kDa domain thatbinds the 3' promoter region is insufficient for transcriptionin systems reconstituted in vitro. An adjacent 10-kDa domaininteracts with the 5' region of the promoter and permits a lowlevel of transcription. A third domain of 10 kDa apparentlydoes not bind the DNA directly but does give rise to 80% ofthe stimulatory effect of TFIIIA. Interestingly, this latterdomain also enhances DNase I hypersensitivity at the 5'boundary of the footprint and within the 5' half of thepromoter. Thus, the induction of HSSs within this region isassociated with the stimulatory activity of TFIIIA.
Unlike the oocyte-type 5S promoter, the somatic-type doesnot give rise to a HSS in response to superhelical stress.Neither S1 nuclease nor Bal-31 cleaves within the gene. Theoocyte and somatic promoters also respond differently toTFIIIA binding. The TFIIIA footprint within the somatic 5Sgene does not include the +50 HSS characteristic of theoocyte-type gene. Thus, it appears that a supercoil-inducedS1 nuclease HSS at +50 correlates with a factor-inducedDNase I HSS at +50. This correlation suggests that theoocyte 5S promoter undergoes an analogous conformationalchange in response to torsional stress or TFIIIA binding.There is reason to suspect that TFIIIA could locally stress theDNA in that it unwinds oocyte-type 5S DNA to a smalldegree (26) and has single-stranded binding preference (27).The differential response of the oocyte and somatic pro-
moters may be related to their differential expression in vivo.During oogenesis, the oocyte 5S genes are transcribed at highefficiency. Transcription falls during oocyte maturation andis not detectable in somatic cells (21). The somatic 5S genesare transcribed in all cell types, including oocytes. The basisof this differential regulation is unclear, since in vitro bothgenes apparently require the same transcription factors inaddition to RNA polymerase III. The somatic promoter does,however, exhibit a 4-fold greater affinity for TFIIIA incompetition studies in vitro (19). This 4-fold binding prefer-ence, in conjunction with decreasing TFIIIA concentrationsduring embryogenesis, is thought to play a role in thedevelopmental inactivation of the oocyte 5S genes. The
increased TFIIIA affinity of the somatic gene derives fromsequence differences near the 5' promoter boundary (19).These sequence changes (47, 53, 55, and 56) presumablyaccount also for the lack of S1 nuclease and Bal-31 sensitivityat +50 of the somatic 5S promoter. This raises the possibilitythat the nuclease-sensitive conformation within the oocyte 5Spromoter is related to the 4-fold difference in TFIIIA affinity.These findings suggest that the oocyte 5S promoter under-
goes a conformational change in response to torsional stress.The functional relevance of this response is suggested byrecent evidence linking torsional stress and active chromatin.The transcriptionally active fraction of simian virus 40minichromosomes or 5S plasmid DNA assembled in Xenopusoocytes can be relaxed with topoisomerase I or DNase I (4,5). Novobiocin, an inhibitor oftopoisomerase II, has a similarrelaxing effect, suggesting that negative supercoils are ac-tively introduced by this enzyme (5). These findings implythat active nucleosomes do not constrain DNA supercoils butare capable of disassembling, allowing the DNA to uncoil.The energy of negative supercoils would then become avail-able to drive the DNA into alternative conformations. If, likeS1 nuclease, specific DNA binding proteins are able torecognize variations in helical geometry as well as nucleotidesequence, a promoter might exhibit optimal factor affinityonly when associated with active nucleosomes.
We thank D. D. Brown and co-workers for the generous gifts of5Splasmids. We also thank E. Plaza for preparing the manuscript. Thiswork was supported in part by grants from the National Institutes ofHealth (GM26453 and CA06824). This is publication no. 3476-MBfrom the Research Institute of Scripps Clinic.
1. Elgin, S. C. R. (1984) Nature (London) 309, 213-214.2. Larsen, A. & Weintraub, H. (1982) Cell 29, 609-622.3. Kohwi-Shigematsu, T., Gelinas, R. & Weintraub, H. (1983) Proc.
Natl. Acad. Sci. USA 80, 4389-4393.4. Luchnik, A. N., Bakayev, V. V., Zbarsky, I. B. & Georgiev, G. P.
(1982) EMBO J. 1, 1353-1358.5. Ryoji, M. & Worcel, A. (1984) Cell 37, 21-32.6. Schon, E., Evans, T., Welsh, J. & Efstratiadis, A. (1983) Cell 35,
837-848.7. Mace, H. A. F., Pelham, H. R. B. & Travers, A. A. (1983) Nature
(London) 304, 555-557.8. Glikin, G. C., Gargiulo, G., Rena-Descalzi, L. & Worcel, A. (1983)
Nature (London) 303, 770-774.9. Hentschel, C. C. (1982) Nature (London) 295, 714-716.
10. Goding, C. R. & Russell, W. C. (1983) Nucleic Acids Res. 11,21-26.
11. Evans, T., Schon, E., Gora-Maslak, G., Patterson, J. &Efstratiadis, A. (1984) Nucleic Acids Res. 12, 8043-8072.
12. Dudler, R. & Travers, A. A. (1984) Cell 38, 391-398.13. Sakonju, S., Bogenhagen, D. F. & Brown, D. D. (1980) Cell 19,
13-25.14. Engelke, D. R., Ng, S.-Y., Shastry, B. S. & Roeder, R. (1980) Cell
19, 717-728.15. Wu, C. (1980) Nature (London) 286, 854-860.16. Pelham, H. R. B. & Brown, D. D. (1980) Proc. Natl. Acad. Sci.
USA 77, 4170-4174.17. Smith, D. R., Jackson, I. J. & Brown, D. D. (1984) Cell 37,
645-652.18. Miller, J. R., Cartwright, E. M., Brownlee, G. G., Fedoroff, N. V.
& Brown, D. D. (1978) Cell 13, 717-725.19. Wormington, W. M., Bogenhagen, D. F., Jordan, E. & Brown,
D. D. (1981) Cell 24, 809-817.20. Peterson, R. C., Doering, J. L. & Brown, D. D. (1980) Cell 20,
131-141.21. Wormington, W. M. & Brown, D. D. (1983) Dev. Biol. 99, 248-257.22. Singleton, C. K., Klysik, J., Stirdivant, S. M. & Wells, R. D.
(1982) Nature (London) 229, 312-316.23. Panayotatos, N. & Wells, R. D. (1981) Nature (London) 289,
466-470.24. Sakonju, S., Brown, D. D., Engelke, D., Ng, S., Shastry, B. S. &
Roeder, R. G. (1981) Cell 23, 665-669.25. Sakonju, S. & Brown, D. D. (1982) Cell 31, 395-405.26. Reynolds, W. & Gottesfeld, J. (1983) Proc. Natl. Acad. Sci. USA
80, 1862-1866.27. Hanas, J., Bogenhagen, D. & Wu, C. W. (1984) Nucleic Acids Res.
12, 2745-2758.
Proc. Natl. Acad. Sci. USA 82 (1985)