tomato research report for 2008-2009 - university of florida
TRANSCRIPT

Tomato Research Reportfor
2008-2009
Supported by the Florida Tomato Committee

Institute of Food and Agricultural Sciences (IFAS) 1022 McCarty Hall Florida Agricultural Experiment Station PO Box 110200 Office of the Dean for Research Gainesville, FL 32611-0200 Tel: (352) 392-1784 Fax: (352) 392-4965
The Foundation for The Gator Nation An Equal Opportunity Institution
MEMORANDUM
TO: The Florida Tomato Committee
FROM: Mary L. Duryea, Associate Dean for Research
SUBJECT: 2008-2009 Research Report
DATE: September 8, 2009 This report describes research by scientists in UF/IFAS that received support from the Florida Tomato Committee during the past year. The Florida Tomato Committee support combines with State and Federal resources to allow IFAS tomato scientists to have a strong and productive research program focused on the tomato industry of Florida. This is an extremely important partnership between the tomato industry and IFAS scientists. We have worked together to identify the questions and needs for the industry. Together we have prioritized the research to be accomplished. This year’s work included food safety, tomato breeding and whitefly and disease management. Many of the results are immediately applicable by the industry. We hope to continue in our quest for improving production methods and product quality. We are pleased to have the Florida Tomato Committee as a partner in programmatic support for tomato research. On behalf of the scientists involved in tomato research, we thank you and appreciate your support. We hope you find the information in the report useful to all facets of tomato production. MLD:las cc: M. McLellan M. Ferrer

ContentsResearchsupportedbytheFLORIDATOMATOCOMMITTEE
2008‐2009IFASResearchReportsPage Title Investigator(s)
FOODSAFETY
1 TrackingsourcesofmicrobialpathogensinpackinghousestoensurefoodsafetyofFloridatomatoes
J.BonillaG.Toor
9 In‐cartonsanitationoffreshmarkettomatoes J.BartzK.SchneiderS.SargentJ.Brecht
11 Effectoflaserlabelingoncolonization,infiltrationandsurvivalofSalmonellaspp.andErwinaontomatosurfaces
E.EtxeberriaK.SchneiderM.Danyluk
TOMATOBREEDING
17 Marker‐assistedselectionforbreedingandrapiddeploymentofdiseaseresistanceinFloridatomatocultivars
J.EdwardsJ.Scott
23 BreedingtomatoesforFlorida J.ScottJ.Edwards
WHITEFLYANDDISEASEMANAGEMENT
30 EvaluationofTYLCV‐resistanttomatocultivarsundercommercialconditionsinsouthwestFlorida
M.Ozores‐HamptonE.McAvoyE.SimonneP.StanslyS.Sargent
43 Identificationofweedreservoirsoftomatoyellowleafcurlvirus J.PolstonD.Schuster
48 Analysisofrecentsporadicpostharvestdecayevents J.Bartz
OTHERTOPICS
61 ThepotentialimpactsthroughoutthemarketchannelfromfoodsafetypracticesimplementedbyFloridatomatogrowersandtheimpactofrecentfoodsafetyscares
J.VanSickle

Tracking Sources of Microbial Pathogens in Packinghouses to Ensure Food Safety of Florida Grown Tomatoes
Final Project Report
Investigators
Dr. J. A. Bonilla1 Dr. G. S. Toor2 1Department of Infectious Diseases and Pathology, University of Florida, Gainesville, FL. 2Gulf Coast Research and Education Center, University of Florida, Wimauma, FL.
July 20, 2009
Funded by the Florida Tomato Committee
Abstract
A study was undertaken to determine the presence and accumulation of microbes in wash tanks at two tomato packinghouses in Florida over several hours of operation. The wash tank water, pre-processed tomatoes and post-processed tomatoes were assayed for total heterotrophic bacteria (THB) and E. coli by traditional membrane filtration methods, and for Salmonella spp. and E. coli O157:H7 by molecular-based methods. The study demonstrates that negligible concentrations of THB remain viable in the chlorinated wash tank water and no E. coli were detected. Moreover, using highly sensitive DNA-based assays, no Salmonella spp. and E. coli O157:H7 could be detected on the surface of tomato grab samples. The high level of microbial inactivation in the wash tanks suggests possible pathogens introduced into the packinghouse process would be effectively inactivated.
Introduction The United States enjoys one of
the safest food supplies in the world. Nevertheless, the spread of infectious diseases through food and beverages is a common, distressing, and sometimes life-threatening problem for millions of people in the US. Each year, the Center for Disease Control (CDC) estimates that 76 million people suffer from foodborne illnesses in the US, accounting for 325,000 hospitalizations and more than 5,000 deaths. There is also a heavy economical toll from foodborne disease as health experts estimate a yearly cost of 5 to 6 billion dollars in direct medical expenses and lost productivity (CDC, 2009). To this end, food safety programs are important to address important health issues related to food sources, production, and consumption. Recent outbreaks of Salmonellosis in fresh tomatoes have highlighted the importance of ensuring continued food safety of produce (Tauxe, 1997, Crohn & Bianchi, 2008, Heaton & Jones, 2008).
The agriculture community has important economic reasons to be concerned and informed about food safety requirements and issues (Lynch et al., 2009). To be accepted in the marketplace, agricultural products must meet governmental food safety standards, and maintain a safety level that inspires continued consumer confidence. There are more than 250 known foodborne diseases caused by bacteria, viruses or protozoa. Some diseases are caused by toxins from the disease-causing microbe, others by the human body’s reaction to the microbial infection. The sources of food contamination are almost as numerous and varied as the contaminants themselves. Bacteria, and other infectious organisms, are pervasive in
1

the environment and foods may become contaminated at many stages of food production, including in the home. Of the numerous human pathogens transmitted through contaminated food, Salmonella spp. and E. coli bacteria represent two of the most common (and often serious) foodborne infections in the US with approximately 40,000 and 73,000 cases of infection, respectively, reported annually to the CDC (Frenzen et al., 2005, Chang et al., 2009).
Tracking the specific sources of Salmonella and other microbial pathogens in tomatoes is a challenge because of the complexity of potential sources involved in tomato production. Potential sources of contamination include water, soil, wastes, insects, and wildlife that may come in contact with the produce. Once the pathogen is introduced into the system, the spread or multiplication of the pathogens to a significant proportion can occur at one or more of the growing-harvesting-packing-distributing points involved with bringing tomatoes to market. Previous research has reported that pathogens can attach to tomato surfaces in the field and can survive post-contamination treatments and storage (Beuchat & Ryu, 1997, Shi et al., 2007).
Packinghouses present an ideal opportunity to remove the pathogens which may have been carried along from pre-harvest of tomatoes to the packing stage. A variety of treatments is employed in packinghouses’ wash stream to decontaminate the tomatoes by using sanitizing agents. The sanitizing agents may include chlorine, trisodium phosphate, electrolyzed water, or a mixture of lactic acid and hydrogen peroxide. Studies have reported that these treatments have mixed success in
removing Salmonella and other pathogens from tomatoes (Heaton & Jones, 2008). For example, a greater reduction in pathogen concentrations was achieved when Salmonella was attached to the tomato skin while a lower reduction was reported when pathogens were attached to the stem scar and/or penetrated inside the tomatoes. The pH and temperature of wash-tank water (or washwater) has also been reported to influence the survival of pathogens. For example, if the fruit temperature is higher than the washwater temperature, washwater can infiltrate the tomatoes (Buchanan et al., 1999). The implications of this are that if the washwater becomes contaminated with pathogens from previous washing events, it could serve as a source of spreading the pathogen to subsequent batches of tomatoes by infiltrating pathogens into the core of tomatoes via wounds (Janisiewicz et al., 1999) or structures present on the skin called lenticels (Burnett et al., 2000) and stomata (Seo & Frank, 1999). It is much more difficult to decontaminate the tomatoes if pathogens are located inside the tomatoes and secondly it would increase the difficulty of tracing the origin/source of pathogens as it could be assumed that the produce was contaminated before arriving in packinghouses.
The objective of this study was to assess the microbial concentration in the wash stream (specifically wash tanks) at two representative South Florida tomato packinghouses and assay the packinghouses washwater for specific pathogens important to the industry using advance molecular biology techniques. Our first objective was to assay the wash stream water using traditional membrane filtration methods
2

to determine the level of total heterotrophic bacteria (THB) and E. coli in the wash tanks over a period of 4-6 hours of heavy operation in the packinghouses. Secondly, we assayed the water and grab samples of tomatoes (before and after packaging) for Salmonella spp. and E. coli O157:H7 using the polymerase chain reaction (PCR) method, which is a highly sensitive DNA amplification technique for specifically identifying the presence of microorganisms.
Materials and Methods
Ninety one washwater samples were collected over four sampling events and analyzed for total heterotrophic bacteria (THB) and E.coli using standard membrane filtration methods as described in Standard Methods for the Examination of Water and Wastewater (APHA, 2005). Briefly, water samples were collected from wash tanks every 30 minutes and filtered through 47 mm filters with a 0.45 um pore size to entrap the bacteria. The filters were placed onto mHPC agar (Heterotrophic Plate Count medium) for the enumeration of THB and on mTEC agar for the enumeration of thermotolerant E. coli. The agar plates were analyzed at 24 and 48 hours of incubation and the number of colony-forming units (CFU)/100 ml was determined. The concentration of chloride was also determined in every washwater sample collected using a Discrete Analyzer and standard EPA methods.
For the genetic analysis of the washwater, a 25 ml sample was filtered through a 25 mm filter with a 0.45 um pore size. The bacterial population entrapped on the filter was lysed in a lysis solution containing 0.5% sodium
dodecyl (lauryl) sulfate (SDS), Proteinase K (100ug/ml) and 0.7M hexadecyltrimethylammonium bromide (CTAB). The DNA was isolated and purified using phenol chloroform extractions and isopropanol precipitation. Two PCR primer sets for Salmonella spp. (targeting a LuxR family member gene sdiA, and invasion-gene activator hilA), one primer set for E. coli O157:H7 (targeting the virulent rfbE gene), and one primer set for ‘universal’ bacteria (16s rRNA gene) were used as previously described (Guo et al., 2000, Halatsi et al., 2006, Omiccioli et al., 2009).
In an attempt to increase the sensitivity of the PCR detection of Salmonella, selected washwater samples, processed for DNA analysis as described above, were also subjected to an enrichment procedure. Bacteria-containing filters were placed into culture-grade tubes with Rappaport-Vassiliadis R10 broth and Tetrathionate broth and incubated overnight at 37 0C. The broth samples were centrifuged at 10,000 × g for 10 minutes, the pellet was lysed in a SDS/ProK/CTAB lysis solution, and the DNA was extracted and purified.
Grab samples of tomatoes (approximately 500 grams) were collected from the truck bins prior to dumping (pre-processed) and from the final packed boxes (post-processed). The tomatoes were collected in a sterile bag and 100 ml of phosphate-buffered saline (PBS) was added. The bag was shaken and agitated every 5 minutes for 30 minutes to remove and collect bacteria off the surface of the tomatoes. A selected number of tomato samples were analyzed directly by PCR after the enrichment procedure. The PBS wash of
3

the pre-processed and post-processed tomatoes was also analyzed for THB by membrane filtration procedure. Table 1
presents a summary of analysis techniques performed in washwater and tomato samples.
Table 1. Summary of analysis Technique Target mHPC agar Total heterotrophic bacteria (THB) mTEC agar E. coli Tetrathionate Broth Salmonella-enrichment prior to PCR Rappaport-Vassiliadis R10 Broth Salmonella-enrichment prior to PCR PCR Salmonella spp. hilA and sdiA genes PCR E. coli O157:H7 rfbE gene PCR Universal 16s rRNA gene Results and Discussion
Analysis of Total Heterotrophic Bacteria (Thb) and E. Coli Bacteria
The concentrations of THB and E. coli enumerated by the membrane filtration method were very low in all washwater samples analyzed (Figure 1). For THB, less than 10 colony-forming-units (CFU) per 100 ml of washwater were detected in nearly all samples throughout the 5-6 hours of tomato processing in packinghouses. While some samples were positive for presumptive coliform bacteria, upon confirmation tests, E. coli was not detected in any of the washwater samples. An initial concern was the travel time from collection of the sample, to transport and analysis in the laboratory. These samples were analyzed within 8 hours of collection. However, to investigate whether the extended contact time was affecting the microbial concentration in the washwater samples, we decided to process the water sample for membrane filtration onto the agar plates at the packinghouses itself and immediately after collection. The results were comparable to initial collection and
transportation of samples to the laboratory, with very few THB and no E. coli detected in the water. Figure 1 shows the average concentrations of THB and chloride levels in the wash water of packinghouses.
4

Figure 1. Total heterotrophic bacteria (cfu/100ml) and chloride levels (mg/L) in wash tank samples collected over 6 hours at 2 South Florida packinghouses. Each packinghouse was sampled at least twice and the averages are reported.
One of the concerns in the packinghouses is the cost of maintaining a high enough concentration of chloride in the wash stream to effectively inactivate microbes. Our preliminary results suggest that despite the lower concentrations of ~200 mg/L chloride in packinghouse B washwater, the levels of bacteria were comparable to the concentrations observed at packinghouse A, where chloride levels were initially ~300 mg/L and progressively increased to 700-800 mg/L after 6 hours of tomato packing operation. These results suggest that a detailed investigation into the microbiocidal effects of chloride levels in packinghouse washwater may be
warranted and could help reduce the amount of chlorine used without compromising bacterial growth. This could help reduce the cost of chlorine disinfection at packinghouses. The pre-processed and post-processed tomato grab samples had detectable levels of THB. The concentration of THB in the pre-processed tomatoes ranged from 6.1 × 104 to 3.4 × 105 CFU/100ml of the PBS wash used on ~500 grams of tomatoes. The post-processed samples had, on average, an 81% reduction (range 73% to 93%) in THB concentration, suggesting that disinfection in wash stream at packinghouses was effective at killing the bacteria. As a comparison,
5

the EPA “acceptable” level for THB per 100 ml of drinking water is 5.0 × 104 CFU. Therefore, the concentrations detected in this study are relatively low, and the presence of THB does not correlate with the presence of human pathogens in the absence of other indicators of fecal contamination. To this end, there was no E. coli detected in the final tomato samples by membrane filtration or PCR (E. coli O1557:H7). Pcr Analysis of Salmonella Spp. and E. Coli O157:H7 Despite the low levels of bacteria detected in the washwater samples by membrane filtration, the PCR reaction for the universal 16s rRNA gene demonstrated a robust amplification of this gene fragment (Figure 2). The PCR assay is a highly sensitive assay using a DNA amplification procedure to detect the presence of organism-specific genes. The assay, as performed here, is
not quantitative, but the positive reactions observed (and shown in Figure 2) demonstrate that there is not a significant level of PCR inhibition in the reaction. This is a particularly important point as environmental samples can contain numerous inhibitors of PCR such as humic acids and complex polysaccharides that can lead to false-negative results being reported. All of the samples analyzed by PCR were positive for the 16s rRNA gene, as would be expected. However, none of the samples was positive for Salmonella spp. or E.coli O157:H7 by PCR. Samples subjected to the Salmonella-enrichment procedure remained negative for Salmonella spp. by PCR suggesting that a direct analysis of 25 ml of water by PCR is not below a particular detection limit, but that the sample is truly negative for Salmonella spp.
Figure 2. Representative samples of 16s rRNA gene fragments isolated from DNA samples. Lane M, DNA marker. Lanes 1-2, tomato wash samples. Lanes 3-5, sash tank samples. Lane 6, negative control.
�� �
6

The study demonstrates that low-to-undetectable levels of bacteria remain viable in the packinghouse washwater tanks throughout a typical packing day. The packinghouses demonstrated equal levels of microbial removal while one used a high level of chlorine and the other a moderate level of chlorine. On-site sampling was feasible but did not result in higher concentrations detected when compared to transporting the samples to the laboratory. While the tomato grab samples indicated that bacteria could be isolated from post-processed fruit, further studies are needed to determine the source of the bacteria. It is known that harvesting during summer months increases the microbial load on tomatoes and it would be of useful interest to sample Florida packinghouses in winter months for comparison. It was not determined whether there is an adherent population of bacteria that are not washed off the fruit, or whether there are low levels of bacteria re-colonizing the fruit downstream of the wash tank. Importantly, no Salmonella spp, E. coli or E. coli O157:H7 were found on fruit during the entire packinghouse processing, including pre- and post-processing, using traditional culture-based assays and molecular-based assays.
REFERENCES
APHA, (2005) Standard Methods for the Examination of Water and Wastewater, 20th edn. American Public Health Association/American Water Works Association/Water Environment Federation, Washington, D.C.
Beuchat, L. R. & J. H. Ryu, (1997) Produce handling and processing practices. Emerg Infect Dis 3: 459-465.
Buchanan, R. L., S. G. Edelson, R. L. Miller & G. M. Sapers, (1999) Contamination of intact apples after immersion in an aqueous environment containing Escherichia coli O157:H7. J Food Prot 62: 444-450.
Burnett, S. L., J. Chen & L. R. Beuchat, (2000) Attachment of Escherichia coli O157:H7 to the surfaces and internal structures of apples as detected by confocal scanning laser microscopy. Appl Environ Microbiol 66: 4679-4687.
CDC, (2009) Surveillance for foodborne disease outbreaks - United States, 2006. MMWR Morb Mortal Wkly Rep 58: 609-615.
Chang, M., S. L. Groseclose, A. A. Zaidi & C. R. Braden, (2009) An ecological analysis of sociodemographic factors associated with the incidence of salmonellosis, shigellosis, and E. coli O157:H7 infections in US counties. Epidemiol Infect 137: 810-820.
Crohn, D. M. & M. L. Bianchi, (2008) Research priorities for coordinating management of food safety and water quality. J Environ Qual 37: 1411-1418.
Frenzen, P. D., A. Drake & F. J. Angulo, (2005) Economic cost of illness due to Escherichia coli O157 infections in the United States. J Food Prot 68: 2623-2630.
Guo, X., J. Chen, L. R. Beuchat & R. E. Brackett, (2000) PCR detection of Salmonella enterica serotype Montevideo in and on raw tomatoes using primers derived from hilA. Appl Environ Microbiol 66: 5248-5252.
7

Halatsi, K., I. Oikonomou, M. Lambiri, G. Mandilara, A. Vatopoulos & A. Kyriacou, (2006) PCR detection of Salmonella spp. using primers targeting the quorum sensing gene sdiA. FEMS Microbiol Lett 259: 201-207.
Heaton, J. C. & K. Jones, (2008) Microbial contamination of fruit and vegetables and the behaviour of enteropathogens in the phyllosphere: a review. J Appl Microbiol 104: 613-626.
Janisiewicz, W. J., W. S. Conway, M. W. Brown, G. M. Sapers, P. Fratamico & R. L. Buchanan, (1999) Fate of Escherichia coli O157:H7 on fresh-cut apple tissue and its potential for transmission by fruit flies. Appl Environ Microbiol 65: 1-5.
Lynch, M. F., R. V. Tauxe & C. W. Hedberg, (2009) The growing burden of foodborne outbreaks due to contaminated fresh produce: risks and opportunities. Epidemiol Infect 137: 307-315.
Omiccioli, E., G. Amagliani, G. Brandi & M. Magnani, (2009) A new platform for Real-Time PCR detection of Salmonella spp., Listeria monocytogenes and Escherichia coli O157 in milk. Food Microbiol 26: 615-622.
Seo, K. H. & J. F. Frank, (1999) Attachment of Escherichia coli O157:H7 to lettuce leaf surface and bacterial viability in response to chlorine treatment as demonstrated by using confocal scanning laser microscopy. J Food Prot 62: 3-9.
Shi, X., A. Namvar, M. Kostrzynska, R. Hora & K. Warriner, (2007) Persistence and growth of different Salmonella serovars on pre- and postharvest tomatoes. J Food Prot 70: 2725-2731.
Tauxe, R. V., (1997) Emerging foodborne diseases: an evolving public health challenge. Emerg Infect Dis 3: 425-434.
8

Progress Report on “In-Carton Sanitation of Fresh Market Tomatoes.” Jerry A. Bartz, PhD, Plant Pathology Department Keith Schneider, PhD, Food Science & Human Nutrition Department Steven A. Sargent, PhD, Horticultural Sciences Department Jeffrey Brecht, Ph.D., Horticultural Sciences Department University of Florida/IFAS
Chlorine dioxide gas supplied by an aqueous solution was applied
to parts of a standard tomato box in a treatment chamber. The chamber also contained a reactive sink (solution of potassium iodide). This was an attempt to determine if the cardboard provided an unlimited sink or just reacted with a percentage of the gas that was released.
The following two charts illustrate the problem with the interior of a tomato carton or with supplying an external source of chlorine dioxide gas and expecting to get it into the carton and onto the tomatoes. Fig. 1.
Fig. 1. Effect of tomato carton fiberboard on the efficacy of ClO2 gas for control of bacterial soft rot in wound-inoculated tomatoes.
9

Cardboard with a slight waxy surface did not interfere with the action of chlorine dioxide gas (compare with ClO2 (+) for decay incidence at 24 and 48 h). Dry cardboard reduced the control
significantly, whereas torn and moist cardboard was completely ineffective. Additionally, scrape wounds were more easily treated than small puncture wounds. Increasing the dose applied:
Fig. 2. Restored some of the efficacy, which suggested that the effect of the fiberboard could be overcome by applying more ClO2 (Fig. 2). Note that in the absence of the carton material, a 2 mg dose completely stopped bacterial soft rot. In subsequent tests with two sheets of cardboard in a standard treatment chamber, the cardboard reacted with or absorbed over 95% of the chlorine dioxide off-gassed from an aqueous source. The percentage was similar over doses ranging from about 2 to 20 mg, unfortunately less than 5% of the initial dose reached the sink. Consequently, a dose of 20 mg would have to be introduced to achieve 1 mg at the sink. In tests without cardboard, 1 to 2 mg per kg of fruit were required to inactivate soft rot bacteria that were introduced into wounds on the fruit surfaces. In future tests, lightly waxed cardboard (as would be used for romaine lettuce) will be enclosed to determine if the coating will reduce the carton’s reactively with chlorine dioxide gas. Additionally, the sink used (water-solution of potassium iodide) will be compared with a freshly wounded tomato to determine their comparative reactivity to chlorine dioxide gas. Splitting an application up into smaller doses was initially not successful in improving the delivery of the gas to the sink, e.g. the cardboard absorbed and equal or even greater percentage of the split dose than for the single dose.
10

Effect of Laser Labeling on Colonization, Infiltration and Survival of Salmonella spp. and Erwinia on Tomato Surfaces Investigators
Ed Etxeberria, Professor, Department of Horticultural Sciences, CREC, Lake Alfred
Keith R. Schneider, Associate Professor, Food Science and Human Nutrition Department
Michelle Danyluk, Assistant Professor, Food Science and Human Nutrition Department, CREC, Lake Alfred
Abstract
Large outbreaks of salmonellosis have been caused by consumption of contaminated raw tomatoes. Labeling of individual fresh fruits and vegetables has the potential for providing product traceability and thereby strengthening food safety systems against foodborne outbreaks. An alternative to labeling produce with a sticker is to directly imprint an alphanumeric code on the produce surface. The alphanumeric code produced by the laser etching method is permanent, requires no additional adhesive and labeling information can be easily modified. This study evaluated the effects of laser-labeled tomato surfaces, waxed laser-etched surfaces and intact tomato surfaces on the survival of Salmonella spp. during storage at 25, 12 and 4°C. Treatment groups included: (i) unlabelled tomato with Salmonella; (ii) laser-labeled tomato, waxed then inoculated with Salmonella; (iii) laser-labeled tomato, inoculated with
Salmonella then waxed; (iv) labeled tomato, then inoculated with Salmonella; and (v) Salmonella inoculated tomato, then laser etched. No significant differences in Salmonella survival were seen among the treatment groups for tomatoes held at 4 or 25°C over 14 days. Experiments evaluating storage times of up to 28 days, at 12°C and the effects of Erwinia colonization on Salmonella survival are currently being quantified, and are set to be completed by mid-August, 2009.
Introduction
During the past two decades, an increase in consumption of fresh produce has occurred in the U.S. (Tauxe et al., 1997). Large outbreaks of salmonellosis have been caused by consumption of contaminated raw tomatoes (CFSAN-FDA, 2001; Toth et al., 2002; CDC, 2005; Roebuck, 2004; US FDA, 2005). These outbreaks continue to be an indication of the lack of an efficient traceback and biosecurity systems in our fresh produce chain.
Labeling of individual fresh fruits and vegetables has the potential for providing product traceability and thereby strengthening food safety systems against foodborne outbreaks. Currently, each piece of fresh produce is marked with a Price Look Up (PLU) sticker which contains a four digit number developed by the Produce Electronic Identification Board (PEIB) which identifies the variety of fruit or vegetable (PEIB, 1995). The PLU sticker is most commonly adhered to the fruit or vegetable surface at the packing line (Varon and Paddock, 1978). Disadvantages to the PLU sticker include the sticky residue left behind on produce
11

surfaces when the sticker is removed as well as damage to the produce surface during removal, usually by fingernails (Durand-Wayland, 2005). Most importantly, PLU stickers may be detached during postharvest handling, thereby eliminating any traceability the sticker may provide (Etxeberria et al., 2006). An alternative to labeling produce with a sticker is to directly imprint an alphanumeric code on the produce surface. This can be accomplished using a newly designed, soon-to-be-approved, laser-etching device that was designed for labeling produce surfaces using a low energy carbon dioxide laser beam (10,600 nm) (Drouillard and Rowland, 1997). The alphanumeric code produced by the laser etching method is permanent, requires no additional adhesive and labeling information can be easily modified (Etxeberria et al., 2006).
The application of an alphanumeric code directly to the surfaces of produce, such as tomatoes, by the laser etching system disrupts the natural cuticular barrier on fruit surface. It must be determined if this disruption of the natural cuticular barrier facilitates greater survival or infiltration of pathogens, such as Salmonella, over undisrupted produce surfaces, or any other secondary deleterious effect.
Wells and Butterfield (1997) reported a significant correlation between isolation of Salmonella and bacterial soft rot in fruit and vegetables sampled from supermarkets. When Salmonella was co-inoculated with the bacterial soft rot pathogen Erwinia carotovora subsp. carotovora, it not only survived in the tissues of various plants,
including tomato fruit slices (Mahovic and Schneider, unpublished), but multiplied to 10-fold greater numbers (Francis et al., 1999). In combined bacteria studies on wounded tomato surfaces using Salmonella and Erwinia, peak growth of Salmonella occurred under optimum conditions for growth of Erwinia (Joy, 2005). The degree to which colonization of Erwinia is implicated in previous outbreaks of salmonellosis linked to tomatoes is unknown.
Objectives
This study will evaluates the effects of laser-labeled tomato surfaces, waxed laser-etched surfaces and intact tomato surfaces on the survival of Salmonella spp. during storage at 25, 12 and 4°C for up to 28 days. Treatment groups included: (i) unlabelled tomato with Salmonella; (ii) laser-labeled tomato, waxed then inoculated with Salmonella; (iii) laser-labeled tomato, inoculated with Salmonella then waxed; (iv) labeled tomato, then inoculated with Salmonella; and (v) Salmonella inoculated tomato, then laser etched. Additionally, the effects of Erwinia colonization on Salmonella survival will be quantified for intact, and laser-etched tomato surfaces.
Methods
Cultures. Five Salmonella serovars from tomato associated sources will be made resistant to rifampicin by spontaneous mutation will be used in all trials. All microbiological media used in this study will be supplemented with 80 mg/L of rifampicin (rif+) (Fisher Scientific, Fair Lawn, N.J.) to allow selection of inoculated Salmonella and inhibit growth of indigenous microflora from produce
12

surfaces (Yuk et al. 2004). Cultures will be maintained at -80°C in tryptic soy broth rif+ with 20% glycerol. Before each experiment, the 5 serovars will be individually cultivated at 37 °C in tryptic soy broth rif+ The cultures will be centrifuged at 3220 × g for 10 min, and the resulting pellet will be washed 2 times in 0.1% peptone. Finally, all 5 serovars will be resuspended into 0.1% peptone and combined as a cocktail and were approximately equal in number in the final population. Erwinia inoculums preparation will be carried out in the same way, but made resistant to ampicillin rather than rifampicin.
Inoculation and Recovery. Tomatoes were obtained from a local packing house (Ruskin, FL). Five treatment groups were evaluated included: (i) unlabelled tomato with Salmonella; (ii) laser-labeled tomato, waxed then inoculated with Salmonella; (iii) laser-labeled tomato, inoculated with Salmonella then waxed; (iv) labeled tomato, then inoculated with Salmonella; and (v) Salmonella inoculated tomato, then laser etched. Tomatoes were inoculated with the Salmonella cocktail using a spot-inoculation technique. Tomatoes were then stored for up to 28 days at 25, 12 and 4°C. At each sample point, 100 ul
of 0.1% peptone was added to the tomato and a rub-shake-rub technique was used prior to dilutions and enumeration on selective agar.
The effects of Erwinia colonization on Salmonella survival was quantified by the treatment groups: (i) unlabeled tomato with Salmonella; (ii) unlabeled tomato with Erwinia; (iii) laser-labeled tomato with Salmonella; (iv) laser-labeled tomato with Erwinia; (v) laser labeled tomato with a high Erwinia concentration and low Salmonella concentration; (vi) laser labeled tomato with a high Salmonella concentration and low Erwinia concentration; and (vii) equal concentrations of Salmonella and Erwinia. Tomatoes were inoculated with the Salmonella or Erwinia using a spot-inoculation technique. Tomatoes were then stored for up to 14 days at 25, and 12°C. At each sample point, 100 ul of 0.1% peptone was added to the tomato and a rub-shake-rub technique was used prior to dilutions and enumeration on selective agar
Statistics. Statistical analyses will be performed using the Statistical Analysis System (SAS; SAS Institute, Cary, NC). Results that yielded P values of < 0.05 were considered significant in this recovery study.
13

Results
Table 1. Fate of Salmonella on laser etched tomatoes held at 4°C (n=6). Counts expressed in log CFU/tomato. Time Treatment (Days) UL+S LL+W+S LL+S+W LL+S S+LL 0 6.5 + 0.6 7.3 + 0.5 7.3 + 0.4 7.4 + 0.5 6.8 + 0.8 1 6.6 + 0.2 7.2 + 0.5 7.3 + 0.3 7.5 + 0.5 6.8 + 0.1 4 6.8 + 0.2 7.3 + 0.5 7.8 + 0.9 7.3 + 0.5 6.7 + 0.2 7 7.5 + 0.7 7.8 + 1.0 7.6 + 0.9 7.7 + 1.1 6.7 + 0.5 10 7.0 + 0.5 8.0 + 1.1 7.9 + 0.4 8.1 + 0.8 6.8 + 0.4 14 6.5 + 0.2 7.0 + 0.4 7.9 + 0.6 7.4 + 0.5 7.7 + 0.4 UL+S – Unlabeled tomato with Salmonella, control LL+W+S – Labeled tomato, waxed then inoculated with Salmonella LL+S+W – Labeled tomato, inoculated with Salmonella then waxed LL+S – Labeled tomato then inoculated with Salmonella S+LL – Salmonella inoculated tomato, then laser etched Table 2. Fate of Salmonella on laser etched tomatoes held at 25°C (n=6). Counts expressed in log CFU/tomato. Time Treatment (Days) UL+S LL+W+S LL+S+W LL+S S+LL 0 6.9 + 0.3 7.1 + 0.4 7.5 + 0.8 7.4 + 0.5 6.5 + 0.7 1 7.4 + 0.4 7.5 + 0.2 7.6 + 0.7 8.0 + 1.0 7.4 + 0.6 4 7.3 + 0.6 7.8 + 0.7 7.7 + 0.4 7.9 + 0.2 7.3 + 0.5 7 7.7 + 1.3 8.4 + 0.8 8.7 + 0.5 8.3 + 0.8 7.8 + 1.1 10 7.2 + 0.9 8.7 + 0.6 8.6 + 0.7 8.0 + 0.9 7.2 + 0.8 14 8.1 + 0.6 8.2 + 0.6 8.2 + 0.4 8.2 + 0.4 7.2 + 0.5 UL+S – Unlabeled tomato with Salmonella, control LL+W+S – Labeled tomato, waxed then inoculated with Salmonella LL+S+W – Labeled tomato, inoculated with Salmonella then waxed LL+S – Labeled tomato then inoculated with Salmonella S+LL – Salmonella inoculated tomato, then laser etched
No significant differences in Salmonella survival were seen among the treatment groups for tomatoes held at 4 or 25°C over 14 days.
14

Remaining Timetable May – August: During this season, objectives one and two will be completed. The work
performed during these trials will help establish pest practices for handling laser etched fruit as well as the potential impact of this technology
August – September: Gather data and perform statistical analysis. At this stage of the
research any repeat experiments will be performed to finalize the data in preparation of report creation.
October: Prepare final reports for submission to the Florida Tomato Committee.
References Center for Food Safety and Applied Nutrition-U.S. Food and Drug Administration [CFSAN-FDA]. September 30, 2001. Analysis and evaluation of preventative control measures for the control and reduction/elimination of microbial hazards on fresh and fresh-cut produce. http://www.cfsan.fda.gov/~comm. Accessed 2003 September.
Diachun, S., Valleau, W. D. and Johnson, E. M. 1942. Relation of moisture to invasion of tobacco leaves by Bacterium tabacum and Bacterium angulatum. Phytopathology. 32: 379-387. Droulliard, G. and Rowland, R. W. 1997. Method of laser marking of produce. U.S. Patent Office #5,660,747.
Durand-Wayland. 2005. Etched fruit to be featured on “Today Show”. Available at: http://durand-wayland.com/label/docs/DWTodayShowRelease.pdf Accessed 8 May 2006. Etxeberria, E., Miller, W. M. and Achor, D. 2006. Anatomical and morphological characteristics of laser etching depressions for fruit labeling. Submitted to HortTechnology. Francis, G. A., Thomas, C. and O'beirne, D. 1999. The microbiological safety of
minimally processed vegetables. International Journal of Food Science and Technology. 34: 1: 1-22. Johnson, J. 1947. Water-congestion in plants in relation to disease. Univ. Wis. Res. Bul. 160, Madison, WI.
Joy, J. A. 2005. Survival of Salmonella and Shigella on tomatoes in the presence of the soft rot pathogen, Erwinia Carotovora. Master’s Thesis, University of Florida.
PEIB. 1995. Guide to coding fresh produce. Produce Electronic Identification Board. Newark, Del. Roebuck, K. 2004. Cause of salmonellosis outbreak still unknown. Pittsburgh, Pennsylvania. URL: http://pittsburghlive.com/x/search/s_248903.html.
Sargant, S. 1998. Handling Florida Vegetables – Tomato. EDIS Publication, Univ. of Florida. SS-VEC-928. Tauxe, R.V., Kruse, H., Hedburg, C., Potter, M., Madden, J. and Wachsmuth, K. 1997. Microbial hazards and emerging issues associated with produce, a preliminary report to the National Advisory Committee on microbiologic criteria for foods. J. Food Prot. 60(11):1400-1408.
Toth, B., Bodager, D., Hammond, R. M.,
15

Stenzel, S., Adams, J. K., Kass-Hout, T., Hoekstra, R. M., Mead, P. S. and Srikantiah, P. 2002. Outbreak of Salmonella serotype Javiana infections -- Orlando, FL, June 2002. Morbidity and Mortality Weekly Report. 15:683-684. URL: http://www.cdc.gov/mmwr/preview/mmwrhtml/mm5131a2.htm. Date of access: 27 Oct. 2004. U.S. FDA. 2005. California Specialty Produce, Inc. recalls product because of possible health risk. URL: http://www.fda.gov/oc/po/firmrecalls/caspecialty05_05.html. Date of access: 14 June 2005. Yuk, H.-G., Warren, B. R. and Schneider, K. R. 2006. Effects of labeling tomato surfaces using a low energy carbon dioxide laser on infiltration and survival of Salmonella spp. In preparation. Wells, J. M. and Butterfield, J. E. 1997. Salmonella contamination associated with bacterial soft rot of fresh Fruits and vegetables in the marketplace. Plant Disease. 81:8: 867-872.
16

Marker-Assisted Selection for Breeding and Rapid Development of Disease Resistance in Florida Tomato Cultivars.
J.D. Edwards and J. W. Scott Gulf Coast Research & Education Center University of Florida, IFAS 14625 CR 672 Wimauma, FL 33598
Abstract
This project is rapidly producing new tomato varieties combining resistance to multiple diseases affecting Florida tomato production. Six different disease resistance targets are being backcrossed into 14 elite lines using marker assisted selection. At the same time, linkage drag effects are being reduced by molecular screening to identify recombinant individuals where the resistance traits have been separated from linked deleterious effects. Through this project, new molecular marker technologies have been developed for tomato to further speed up the rate of resistance gene integration. Ultimately the resistance genes will be deployed in elite lines that are already proven to perform well as parents of hybrid varieties.
Introduction
Tomato varieties with improved resistance to multiple diseases are urgently needed by the Florida tomato industry. Sources of genetic resistance to many critical diseases affecting tomato production in Florida have been identified within cultivated tomato germplasm or wild relatives, and breeding efforts are required to deploy
the resistance alleles in commercially acceptable varieties.
The integration of multiple resistance traits will be greatly simplified by the marker-directed transfer of each corresponding resistance gene to a common set of commercially acceptable fresh market tomato genetic backgrounds. The resistance genes are generally found in less desirable genetic backgrounds. Traditionally, breeding of new varieties incorporating a disease resistance trait involves crossing susceptible elite lines with a resistance source (donor) and screening of segregating progeny for the resistance. Resistant progeny may then again be crossed (backcrossed) to a susceptible elite line. Each iteration of backcrossing reduces the amount of DNA inherited from the donor (and potential for undesirable traits inherited from the donor) by an average of 50%. Without molecular markers, an additional season is needed for progeny testing in each selection cycle. Breeding for multiple resistance genes decreases the number of useful segregating progeny exponentially. To overcome this limitation, a marker-assisted backcrossing strategy has been implemented to integrate the resistance traits in parallel into the elite genetic backgrounds. A collection of elite lines developed by the UF tomato breeding program are being used as recurrent parents for trait integration. The set of lines consists of parents of recently released hybrid varieties and other promising inbred parent lines. Some of the lines have particular positive attributes such as heat tolerance, bacterial spot tolerance, and enhanced flavor. The lines have been evaluated in
17

multiple seasons and in multiple locations. Their yield and horticultural characteristics are similar to commercially available varieties. The benefit of the parallel breeding strategy is that once resistance genes are individually integrated into elite lines, the resistance genes can quickly be pyramided to form lines with multiple disease resistances in the same uniform genetic background.
A confounding issue often encountered in breeding for disease resistance from wild sources is that the segment of DNA that contains the resistance gene itself may also harbor linked genes with undesirable effects (termed linkage drag). The larger the segment is, the greater the potential for linkage drag effects. Meiotic recombination events that break linkage drag can be detected phenotypically (in cases of severe linkage drag) or through screening with molecular markers. Subtle negative traits associated with the resistance allele may not become apparent until late in large-scale variety testing and hinder the commercial success of the variety. Therefore it is desirable to break linkage drag and to deploy the resistance genes on the smallest segments of wild DNA possible.
A further increase in speed for generating backcrossed lines can be accomplished using molecular marker selection across the entire genome, not just for the gene of interest. Whole genome selection identifies individuals with the least genomic contribution of the donor and speeds the recovery of the recurrent parent genetic background. Using whole genome selection, it is possible to reduce the
number of backcross generations from six to three (Frisch and Melchinger 2005). The limitations to implementing whole genome selection in tomato are cost and availability of polymorphic molecular markers. New technologies may help to alleviate these challenges.
The long-term goal of this project is to separately integrate a large number of disease resistance alleles into sets of inbred parent lines that are known to produce commercially acceptable hybrids. Integrating the resistance alleles separately (in parallel) into the same genetic backgrounds will produce near isogenic lines (NILs) that are genetically identical to the original lines except for the small segment of DNA spanning the integrated allele. This strategy will enable efficient gene pyramiding; Varieties with any combination of disease resistances can be constructed by simply crossing the desired NILs and screening the progeny with molecular markers linked to the resistance genes. The overall objective of this project is to use marker assisted selection to breed near isogenic versions of established and promising UF hybrid tomato parental lines with the addition of alleles for resistance to high priority diseases. There is both a scientific and practical value in deployment of disease resistance alleles as NILs in a consistent set of standard genetic backgrounds. Scientifically, the effects of each resistance allele can be studied separately and in combination with other resistance alleles without other confounding genetic effects. Practically, NILs provide a tomato variety that is upgradable with a wide selection of disease resistances and can be customized for different growing
18

conditions. As additional loci of interest are mapped (e.g., gray-wall resistance, yield traits, etc.), they can also be incorporated into the same genetic backgrounds using marker-assisted backcrossing. The specific objectives of this project were:
1) Marker-assisted backcross breeding of high priority resistance genes into a set of elite breeding lines and parents of hybrid cultivars.
2) Breaking linkage drag surround resistance genes, focusing on problematic TYLCV resistance sources.
3) Developing markers to accelerate backcross breeding using marker-assisted background selection.
Objective 1: Marker-assisted backcross breeding
Methods: An initial cross was made between each donor line carrying a resistant allele with each of the recurrent parents. Seeds from fruit derived from cross-pollinated flowers were collected and sown to obtain the next generation. The resulting F1 plants were crossed back to the recurrent parent. The individuals resulting from this cross (BC1) were screened with molecular markers to identify individuals
that carry the resistant allele, and these individuals were again be crossed back to the recurrent parent. This process of selection and backcrossing has been repeated presently up to the BC3 generation. Once BC6 individuals are obtained, they will be allowed to self-pollinate and progeny homozygous for the allele of interest will be selected.
Results: Multiple generations of backcrossing has been completed for resistance to six different tomato diseases into a panel of 14 elite recurrent parent lines (Table 1). The sources of disease resistance include tomato yellow leaf curl virus (TYLCV) resistance, crown rot resistance, southern blight resistance, and bacterial speck resistance. TYLCV resistance sources include one from the Ty-1 locus and two recombinant sources from the Ty-3 locus. Most lines are at the BC2-BC3 generation. Backcrossing will be complete within 3-4 generations (at the BC6 generation). Molecular markers for marker-assisted selection are in use for Ty-1 (Zamir et al. 1994 and Ty-3 (Ji et al. 2007) loci, and bacterial speck (Martin et al. 1993). Molecular markers for crown rot resistance are under development (M. Stevens personal comm.). Markers for southern blight resistance are being developed through an M.S. student thesis research project.
19

Table 1. Current status of backcrossing disease resistance gene into elite recurrent parents.
TYLCV resistance (Ty-1 gene)
TYLCV resistance (Ty-3 recombinant #1)
TYLCV resistance (Ty-3 recombinant #2)
Crown rot resistance
Southern Blight Resistance
Bacterial speck resistance
7776 BC2 BC3 BC3 BC3 BC2 BC2 7781 BC2 BC2 BC2 N/A BC2 BC2 7804 BC2 BC2 BC3 BC3 BC2 BC2 7907B BC2 BC2 BC2 BC3 BC2 BC2 8000 BC2 BC2 BC2 BC2 BC2 BC2 8059 BC2 BC2 BC2 BC1 BC2 BC2 8083 BC2 BC2 BC2 BC3 BC2 BC2 8111B BC2 BC2 BC2 F1 BC2 BC2 8124C BC2 BC2 BC2 BC3 BC2 BC2 8249 BC2 BC2 BC2 BC3 BC2 BC2 8293 BC2 BC2 BC2 BC3 BC2 BC2 8539C BC2 BC2 BC2 BC3 BC2 BC2 8608 BC2 BC3 BC1 BC3 BC2 BC2 NC2 BC1 F1 F1 F1 BC2 F1
20

Objective 2: Breaking linkage drag
Methods: Efforts to break linkage drag have focused on TYLCV resistance sources derived from the wild tomato relative Solanum chilense. Segregating populations from a parent that is heterozygous at the resistance locus were screened with markers at each end of the introgression. Individuals that have undergone recombination within the introgression were detected by a change in linkage phase of the marker alleles. The recombinant individuals were tested to determine whether or not the smaller introgression still confers the disease resistance phenotype, and they were screened with additional markers to establish the position of the recombination event with greater precision. This process serves to fine map the resistance loci, and to identify resistant lines with reduced linkage drag.
Results: Recombinant screens were done for the TYLCV resistance loci Ty-1 and Ty-3. Approximately two thousand lines segregating at the TYLCV resistance locus Ty-1 were screened for recombinants between a pair of markers on tomato chromosome 6. Severe recombination suppression was encountered for this region. This is consistent with the findings of other research groups. No recombinants were identified from this screen whereas nearly 10% of the lines would be expected to be recombinants based on the previously estimated map length without recombination suppression. However a single recombinant was identified from material derived from a
recombinant screen in an earlier generation. This recombinant reduced the introgression size to roughly half its previous size. This smaller introgression will be targeted for future recombinant screening. If recombination suppression is not overcome in this line, alternative strategies will be explored including recombination with other sources of S. chilense introgressions or introgressions from other wild species nearby the Ty-1 region. At the Ty-3 locus, many recombinants have been obtained and two recombinants were selected as sources to incorporate into the elite lines by backcrossing. These two sources have introgressions with minimal overlap so linked genes with deleterious effects in either source are unlikely to be present in the other. Current backcross generations for the Ty-1 and Ty-3 sources are at the BC2 stage or later.
Objective 3: Markers for background selection
Methods: The first step in implementing whole-genome selection is to identify a set of polymorphic markers distributed throughout the genome. This is particularly difficult to achieve in tomato because polymorphism rates are extremely low. One solution is to focus marker development on highly variable genomic features including various types of repetitive DNA. As a part of this project, a marker system for background selection in tomato is being developed based on transposable elements (TEs) using the transposon display technology (Casa et al. 2000). A set of TEs has been identified using a bioinformatic approach to discover
21

elements with moderate to high copy number and a high level of sequence similarity between the elements indicating recent transposition activity and potential for high rates of polymorphism between closely related lines. One of these elements is the retroelement “Rider” which has been implicated in controlling fruit shape at the sun locus (Jiang et al. 2009).
Results: Eight candidate transposable elements have been screened on a set of tomato varieties and wild species with transposon display and six have patterns of polymorphism that could be useful for background selection. Five of eight transposable elements have revealed moderate rates of polymorphism between cultivated tomato lines and high rates of polymorphism between cultivated tomato and wild species that are common sources of disease resistance including S. pimpinellifolium, S. chilense, and S. habrochaites. In addition to markers from transposon display technology, genome-wide single nucleotide polymorphism (SNP) markers are under development. DNA from elite recurrent parents has been included in a panel of tomato germplasm that will be genotyped as a part of the USDA funded Sol-CAPS project (http://solcap.msu.edu/). This collaboration will serve as an additional source of polymorphic markers that can be used for whole-genome background selection.
Literature Cited
Casa A, Brouwer C, Nagel A, Wang L, Zhang Q, Kresovich S, Wessler S (2000) Inaugural article: the MITE family heartbreaker (Hbr): molecular markers
in maize. Proc Natl Acad Sci U S A 97:10083-10089
Frisch M, Melchinger AE (2005) Selection theory for marker-assisted backcrossing. Genetics 170:909-917.
Ji Y, Schuster DJ, Scott JW (2007) Ty-3, a begomovirus resistance locus near the Tomato yellow leaf curl virus resistance locus Ty-1 on chromosome 6 of tomato. Molecular Breeding 20:285-285.
Jiang N, Gao D, Xiao H, van der Knaap E (2009) Genome organization of the tomato sun locus and characterization of the unusual retrotransposon Rider. Plant J
Martin G, Brommonschenkel S, Chunwongse J, Frary A, Ganal M, Spivey R, Wu T, Earle E, Tanksley S (1993) Map-based cloning of a protein kinase gene conferring disease resistance in tomato. Science 262:1432-1436
Zamir D, Eksteinmichelson I, Zakay Y, Navot N, Zeidan M, Sarfatti M, Eshed Y, Harel E, Pleban T, Vanoss H, Kedar N, Rabinowitch HD, Czosnek H (1994) Mapping and introgression of a Tomato Yellow Leaf Curl Virus Tolerance Gene, Ty-1.Theoretical and Applied Genetics 88:141-146.
22

Breeding Tomatoes For Florida J. W. Scott and J.D. Edwards Gulf Coast Research & Education Center University of Florida, IFAS 14625 CR 672 Wimauma, FL 33598
Abstract
Seed of recent hybrid releases Tasti-Lee and’ Tribeca’ are available from Bejo Seeds and Vilmorin, Inc., respectively. Tasti-Lee will soon be test marketed in supermarkets in three cities. Growers on the peninsula of Florida should try ‘Tribeca’ in their early fall plantings as it is a heat-tolerant variety. It also has tomato spotted wilt resistance for those growing in North Florida or the Southeast. One of the five TYLCV resistant breeding lines representing round, plum, and cherry types released in fall 2007 also carries the newly discovered Ty-4 gene which will allow for marker assisted selection to speed up the breeding process. Fla. 8624B appears to be a promising inbred with TYLCV resistance. Hybrids with bacterial wilt tolerant parent Fla. 8626 did well in a recent trial and will be tested further, some show tolerance to bacterial spot. Fla. 8736 is a heat-tolerant inbred with fusarium crown rot resistance that should make an outstanding hybrid parent. Fla. 8590 is heat tolerant and may also make a good hybrid parent. To further improve flavor of Tasti-Lee, fruity/floral backcross line Fla. 8629 had high flavor scores in all four-taste panels were it was tested, as did heterozygous hybrids from it in the two panels where they were tested. Fla. 8735 is a new inbred with large globe
fruit and outstanding flavor. Jointless, crimson hybrid Fla. 216 yielded similar to ‘Sanibel’ in Dade Co. but had superior interior color. Fla. 8647 is a bacterial spot tolerant inbred that should make a good parent. Bacterial spot tolerant hybrid Fla. 8555 did well in several trials and will continue to be tested. At least two chromosomal regions associated with bacterial spot resistance have been identified by molecular markers and this should advance development of bacterial spot tolerant varieties.
Introduction
Improved varieties are needed to keep the Florida tomato industry competitive in a changing world. Improvements include the areas of yield, pest resistance, and fruit quality. Florida’s geography offers many challenges for tomato improvement, since high temperatures limit fruit set and are conducive to disease and insect problems. Although private companies have tomato-breeding programs that develop varieties for Florida, these companies do not handle many of the long term, high-risk projects that could prove valuable in the future. The University of Florida tomato-breeding program aims at such projects and works in partnership with the private companies to deliver improved varieties of benefit to the Florida tomato industry.
Much of the groundwork for the development of heat-tolerant varieties was done at the University of Florida (Scott et al., 1986). ‘Solar Set’ (Scott et al., 1989) was an important commercial, heat-tolerant variety for 11 years after it was released. The improved heat-
23

tolerance in ‘Solar Fire’, released in 2003, has been evident on some farms, but sensitivity to bacterial spot that has been carried with resistance to fusarium wilt race 3 has limited its use. See results under objective 5 for new advancements in this area. Fusarium wilt race 3 resistant varieties are available because of the introgression of a resistance gene discovered in a wild species by this program in the 1980's (Scott and Jones, 1989; Scott and Jones, 1995). Breeding lines with Fusarium crown and root rot resistance were released in 1999 (Scott and Jones, 2000) and UF breeding lines are in commercial varieties ‘Sebring’, ‘Soraya’, and ‘Crown Jewel’. With the impending loss of methyl bromide, these pathogens could become more widespread. Other diseases such as bacterial wilt and spotted wilt occur in Florida, but cause far more damage in other regions of the world. If these diseases become more prevalent here, resistant varieties will prove beneficial. Tomato yellow leaf curl virus (TYLCV) is a serious threat to Florida tomato production (Polston et al., 1999) and some crops have sustained severe losses in past years. Commercial companies have released TYLCV resistant varieties but none have yet been widely accepted by Florida growers. Our project has been ongoing since 1990 utilizing different resistance genes derived from the wild species Solanum chilense. It appears that four genes have been introgressed, with two genes needed in a variety to provide resistance. These genes must be incorporated into both parents of a hybrid since the resistance is not
dominant. This makes the breeding of finished varieties more difficult, but such hybrids are presently being evaluated. Molecular markers such as Ty-3 (Ji et al., 2007) linked to the resistance genes are being developed that will accelerate future breeding progress and these will be provided to tomato breeders for their use and benefit to the Florida industry.
Tomatoes have met with dissatisfaction in the marketplace. Essentially, this relates to compromises that are made in providing fruit that will ship well. Solutions to this problem are not simple. Research is needed to provide tomatoes that will be more acceptable to consumers. On the bright side, Nugyen and Swartz, (1999) and others have shown that lycopene, the red pigment in tomato, has strong antioxidant properties that reduce several cancers. Work in the breeding program has been ongoing for over 25 years with the crimson gene (ogc) that improves internal tomato color and increases lycopene content. Seven breeding lines with this gene have been released to seed companies over the last 14 years. Crimson varieties may be a boon to the Florida industry in the near future and a crimson hybrid ‘Fla. 8153’ trademarked as Tasti-Lee was released in 2006. Genetic alteration of plant architecture might provide ground tomatoes that reduce labor costs for staking and harvesting. This could also be important for future production in Florida. Objectives of this project were:
24

1) To develop varieties or breeding lines resistant to soil borne pathogens.
2) To develop improved heat-tolerant inbreds and hybrids.
3) To develop commercially acceptable breeding lines and hybrids resistant to TYLCV or tomato spotted wilt virus (TSWV).
4) To improve fruit quality and postharvest characteristics.
5) To develop commercial hybrids for the Florida tomato industry.
OBJECTIVE 1:
Methods. Seedlings of hundreds of lines are inoculated with fusarium wilt races 2 and 3, fusarium crown rot, verticillium wilt, and bacterial wilt to select resistant plants for field evaluation. The lines showing the most promise are tested in yield trials at GCREC and some are tested at NFREC or Dade County. The best of these are crossed to test hybrids for commercial release. The hybrids are evaluated at GCREC (stage 1) and the best ones are advanced to replicated trials at GCREC (stage 2). The best of these hybrids are then tested off station at IFAS trials and on grower farms (stage 3). This report will highlight primarily some results of stage 2 testing that took place in the last year.
Results. Fla. 8626 is a bacterial wilt tolerant inbred with huge fruit size. Five hybrids were selected for stage 2 testing in fall 2008 and yield data were taken in
spring 2009. All performed well with 3 being of particular interest. These will be tested further next year. Several have strong vines with some tolerance to bacterial spot. Fla. 8626 had significantly larger fruit than any other entry in the spring trial, but it was late in maturity. It combines well with early fruiting parents. New crimson inbred Fla. 8736 performed well in spring 2009 both in a yield trial and in the breeding plot. It is heat-tolerant and likely carries resistance to fusarium crown rot. Fla. 8736 has very good flavor and appears to have little to no zippering. Numerous hybrids were made with it in the spring and these will be tested this fall.
OBJECTIVE 2:
Methods. Heat-tolerant (HT) fruit setting ability is being incorporated into all phases of the breeding program. For instance many lines being developed with bacterial spot tolerance have heat tolerance as well. Key selection in this project is done for early fruit setting ability in fall crops when there is high temperature stress. Otherwise procedures are similar as those described under objective 1.
Results. Hybrid Fla. 8455 continued to perform well in trials from Gadsden to Dade counties and is in phase 3 testing. Inbred Fla. 8590 did well in the spring 2009 yield trial. However, only a few hybrids were made with 8590 in the spring and more will be made this fall. As indicated elsewhere the HT trait is now in several lines listed under other projects so is more ubiquitous than indicated here. Heat-tolerant hybrids with the proper horticultural characteristics should
25

provide Florida growers with varieties that set marketable fruit more reliably under stress conditions such as cool temperatures as well as under high temperatures. A heat-tolerant UF parent is in Sakata’s tomato spotted wilt resistant hybrid ‘Bella Rosa’ and growers should try this variety in their early fall plantings as it has done well under heat stress at NFREC.
OBJECTIVE 3:
Methods. Geminivirus resistance. Hundreds of breeding lines from F2 to F10 are inoculated with TYLCV at the seedling stage via viruliferous whiteflies and then rated for disease severity in the field. Once resistance is fixed, generally in the F3 or F4 generation, backcrossing is done to make further horticultural improvements. The best inbreds are also intercrossed and the resultant hybrids then evaluated for possible stage 2 testing. Over the last decade or so we have been working intensively to locate molecular markers linked to resistance genes as this is key to developing horticulturally acceptable parent lines and hybrids with TYLCV resistance. Tomato Committee funding has helped with the marker work, but other funding sources have been critical as well since this research is expensive.
Spotted wilt resistance. Selection for Sw-5 gene resistance is done with a molecular marker that eliminates the need to screen with thrips and potentially spread the virus in the west coast growing region. Our newly identified Sw-7 gene does not yet have a marker, although our cooperator Dr. Mike Stevens at BYU is very close to
developing one. Selections are made without TSWV pressure at GCREC and the resultant lines are assayed at NFREC to identify which carry Sw-7. Several hybrids with each gene have been grown recently to evaluate the incidence of fruit blotch, which has been a problem with Sw-5 hybrids under some conditions. Preliminary data suggests that blotch is reduced or eliminated in Sw-7 varieties but this needs to be verified. Otherwise development procedures as to inbred and hybrid testing are similar to those described above.
Results. Geminivirus resistance. Five breeding lines; 2 with large fruit, 2 plums, and a cherry line, each with the Ty-3 gene and a robust linked molecular marker were released in fall 2007. In 2008 we identified another resistance gene Ty-4 (Ji et al, 2009) and one of the lines also has that gene. Now for the first time breeders can use marker assisted selection to incorporate resistance from our breeding program, which will speed up the process considerably. In 2008 a contract was sent out to seed companies to license the marker and breeding lines. This material should help with future TYLCV resistant variety development for Florida. A few hybrids are showing some commercial possibilities, but more testing is needed and seed availability has often been lacking. Fla. 8624B is a resistant inbred that looked very good in a winter Dade Co. breeding trial and numerous hybrids were made with this line in spring 2009.
Spotted wilt resistance. In fall 2007 ‘Tribeca’ was released, the name
26

was changed from ‘Gulf Stream’ due to some trademark concerns. Vilmorin Inc. is the seed company producing seed. Initial seed production had germination problems, but soon there should be good seed available. For seed contact Caroline Cordier ([email protected] phone 520-940-1539). It is heat-tolerant as well as TSWV resistant and it may do well for fall production on the Florida peninsula. Fruit blotch data is not available at this time from experiments to compare lines with Sw-5 vs. Sw-7 . It appears that hybrids with Sw-7 are not quite good enough for release so crosses were done to develop better inbreds.
OBJECTIVE 4:
Methods. Fruit quality and shelf life are emphasized in all breeding projects. One aspect of fruit quality is the development of high lycopene varieties by using the crimson (ogc) gene. This gene is widely distributed in the various breeding projects. New inbreds and hybrids are continually being made and evaluated. Another aspect is the incorporation of high sugars from a cherry line into large-fruited lines and in developing improved cherry/grape breeding lines. We are backcrossing the desirable fruity/floral note into the parents of Tasti-Lee to further improve the flavor. This is challenging because expression of this note is subject to environmental variation and only a few lines are stable for good expression. The most interesting lines from all flavor work are tested in flavor trials with USDA cooperator Dr. Elizabeth Baldwin. Another project is to develop tomatoes
that do not require staking by use of the brachytic (br) gene that reduces plant height and increases side shoots. These tomatoes are called compact growth habit (CGH) tomatoes. Here we emphasize jointless tomatoes that can be mechanically harvested. Selections take place at GCREC and in Dade County on a grower farm.
Results. Seed of Tasti-Lee is now available from Bejo Seeds; contact Greg Styers ([email protected] phone 805-689-1627). This variety will be test marketed later this year in 3 cities by the Florida Dept. of Agriculture and Consumer Services. In the backcrossing of the fruity/floral note into the parents of Tasti-Lee, line Fla. 8629 was selected. In all four-taste panels it has been in the most preferred flavor group. Moreover two of the panels have tested Fla. 8629 backcrossed again and the resulting heterozygous hybrids have also been in the most preferred flavor group. This indicates that it may be possible to further improve the flavor of Tasti-Lee by incorporating fruity/floral into just one of the parents. This is good news since 8629 needs more size and firmness and selection for good expression of the fruity/floral note is difficult. Fla. 8735 is a new inbred with very large, globe-shaped fruit with outstanding locule formation. It has outstanding tomato flavor and numerous hybrids were made with it in spring 2009 to begin to test its hybrid potential. In the CGH project numerous selections with jointless stems were made (that would allow for mechanical harvest). It appears that another cross from these is needed to obtain
27

horticultural attributes necessary for commercial hybrid production. To do this the best of these will be crossed to elite CGH jointed lines and elite conventional jointless lines in the program.
OBJECTIVE 5.
Methods. A project has been underway for a number of years to develop a superior crimson, good flavored, jointless tomato cultivar or hybrid especially well-suited for Dade County growing conditions. This is a cooperative project with Drs. Jim Strobel and Waldy Klassen. A hybrid trial was conducted on a grower farm in Dade County last winter with the goal to find a hybrid that competes with ‘Sanibel’. Our bacterial spot resistant project was not funded last year but since we did work on bacterial spot it will be reported here. Sam Hutton finished his Ph.D. with part of the work to locate molecular markers linked to race T4 resistance. We also did some experiments in fall 2008 and spring 2009 to confirm the most important markers. Bacterial spot lines are inoculated in the field but natural infections often provide enough disease pressure.
Results. Jointless, crimson hybrids 214 and 216 had yield and fruit size comparable to ‘Sanibel’ in a yield trial conducted on a grower farm in Dade County during the winter. Firmness was similar while interior red color was superior to ‘Sanibel’ and ‘Florida 47’. These hybrids use a parent from our program and a parent from retired UF tomato breeder Jim Strobel. The parent in 216 is Fla. 8653, which has looked
promising. Other jointless inbreds that have been doing well are Fla. 8652B and Fla. 8543B. These are also being test crossed and the hybrids are being evaluated. We have been doing large seed increases on Tasti-Lee and ‘Tribeca’ and moderate seed increases on several of the hybrids mentioned in recent reports. We have had trouble getting seed on several of the TYLCV resistant hybrids for unknown reasons and are trying to correct this with increased efforts.
We have strong evidence for 2 chromosomal regions that are linked to bacterial spot resistance. Moreover, 2 regions were found where alleles resulted in increased susceptibility. One of these was the marker linked to fusarium wilt race 3 resistance. Growers have seen that fusarium wilt race 3 resistant varieties are quite sensitive to bacterial spot. In spring 2009 we identified a line with fusarium race 3 resistance, but with very good bacterial spot resistance and it had the 2 resistant markers in it. This suggests progress can be made to develop fusarium wilt race 3 resistant lines with bacterial spot tolerance in the future. Bacterial spot breeding will be more efficient by selecting for the 2 markers and using modified marker assisted selection in the future. Hybrid Fla. 8555 has good bacterial spot tolerance and has yielded well in 3 of 4 trials. It will move to stage 3 testing. Inbred Fla. 8647 looked good for yield and fruit size in spring 2009 and it is now being crossed to assess its parental potential. Some parental potential has been seen in hybrids with Fla. 8561B and Fla. 8476
28

as parents. These 2 do not have high levels of race T4 tolerance, but they do provide strong vines that are not so affected by the pathogen.
Literature Cited
Ji, Yuanfu, David J. Schuster, and Jay W. Scott. 2007. Ty-3, a begomovirus resistance locus near the Tomato yellow leaf curl virus resistance locus Ty-1 on chromosome 6 of tomato. Molecular Breeding 20:271-284.
Yuanfu Ji, Jay W. Scott, David J. Schuster, and Douglas P. Maxwell. 2009. Molecular Mapping of Ty-4, a New Tomato Yellow Leaf Curl Virus Resistance Locus on Chromosome 3 of Tomato.
J. Amer. Soc. Hort. Sci. 134(2):281–288.
Nguyen, M.L. and S.J. Schwartz. 1999. Lycopene: Chemical and biological properties, Food Technol. 53(2): 38-45.
Polston, J.E., R.J. McGovern, L.G. Brown. 1999. Introduction of Tomato yellow leaf curl virus in Florida and implications for the spread of this and other geminiviruses of tomato. Plant Dis. 8384-988.
Scott, J. W., and J. P. Jones. 1989. Monogenic resistance in tomato to Fusarium oxysporum f. sp. lycopersici race 3. Euphytica 40:49-53.
Scott, J. W., and John Paul Jones. 1995. Fla. 7547 and Fla. 7481 tomato breeding lines resistant to Fusarium oxysporum f. sp. lycopersici races 1, 2, and 3. HortScience 30(3):645-646.
Scott, J. W., and John Paul Jones. 2000.
Fla. 7775 and Fla. 7781: Tomato breeding lines resistant to Fusarium crown and root rot. HortScience 35(6):1183-1184.
Scott, J. W., S. M. Olson, J. J. Bryan, T. K. Howe, P. J. Stoffella, and J. A. Bartz. 1989. Solar Set: A heat tolerant, fresh market tomato hybrid. Fla. Agric. Expt. Sta. Circ. S-359 10p.
Scott, J. W., R. B. Volin, H. H. Bryan, and S. M. Olson. 1986. Use of hybrids to develop heat tolerant tomato cultivars. Proc. Fla. State Hort. Soc. 99:311-314.
29

Evaluation of TYLCV-resistant Tomato Cultivars under Commercial Conditions in Southwest Florida Monica Ozores-Hampton1, Eugene McAvoy2, Eric Simonne3,
Phil Stansly1 and Steve Sargent3.
1University of Florida/IFAS, SWFREC, Immokalee, FL. 2Hendry County Extension Service. 3University of Florida, Horticultural Sciences Department, Gainesville, FL.
Final Report Florida Tomato Committee 20 July 2009
30

Abstract
Tomato yellow leaf curl virus (TYLCV) considered by some to be the worst tomato virus worldwide, is now endemic to Middle East, North Africa, the Caribbean and the USA, including Florida. Management of the whitefly vector and TYLCV rests primarily on insecticides and tomato-free planting periods. However, insecticide resistance is creating an urgent need for alternative management tools such as TYLCV resistant varieties. Therefore, the objective of this study was to evaluate the horticultural characteristics of currently TYLCV-resistant tomato varieties currently in the USA market. The study was conducted in the spring 2009 with twelve TYLCV-resistant varieties plus two grafted varieties. We compared the varieties with two susceptible grower standards in a completely randomized experimental design with three replications in Immokalee, FL. Whitefly populations and TYLCV pressure were low. First harvest of extra-large fruit yields, total first harvest, (all sizes combined), first and second combined extra-large, total first and second harvest (all sizes combined), total extra-large and total harvest [(all sizes and harvest combined) were higher for ‘Tygress’, ‘Tygress on FL47’ grafted plants and ‘Sur 200’ than the rest of the varieties. The lowest total unmarketable yields for the first harvest, first and second harvest combined (all sizes combined) and total harvest (all sizes and harvest combined) were with ‘Tygress’, ‘Tygress on FL47’ grafted plants, ‘FL 47’ on ‘Tygress’ grafted plants, ‘Sur 200’, ‘FL 47’ and ‘Sebring’. The most common
defect types as percentage of the total unmarketable yields harvested per TYLCV-resistant variety were scaring and odd shape, followed by blossom end scar and discoloration. There were no significant differences in firmness, pH, SSC, and TTA among TYLCV varieties. Participant perception under a blind test rated best overall were ‘Tygress’, ‘Sur 200’ and ‘HM 8845’ based in earliness, plant vigor, fruit size, firmness, fruit quality, potential yield and an overall plant rating.
Introduction
Tomato yellow leaf curl virus (TYLCV), considered by some to be the worst tomato virus worldwide, TYLCV is now endemic to Florida. The virus causes stunted growth and flower abortion with early infections resulting in almost no fruit set (Schuster and Stansly, 1996). Tomato growers have focused their attention on whitefly control to reduce losses. Management of whitefly and TYLCV rests primarily on insecticides, particularly the neonicotinoids, and tomato-free planting periods initiated by timely crop destruction after harvest (Schuster and Polston, 1999). However, insufficient control, in part due to insecticide resistance, is creating an urgent need for additional management tools.
The most important of these management tools is to the use of TYLCV-resistant varieties. Companies such as Seminis, Syngenta, De Ruiter, Hazera, Zeraim, Zakata, Harris Moran, and Clause have already developed TYLCV-resistant cultivars adapted to our needs and environment. Some resistant varieties were evaluated 8 years ago
31

(Gilreath et al, 2000) and more recently by Cushman and Stansly (2006). Subsequent trials in spring 2007 included nine round and two plum tomato cultivars resulted in very distinctive performed in the field as well after the post-harvest evaluation under a high virus pressure (Ozores-Hampton et al., 2008). However, information on the virus tolerance and horticultural qualities including post-harvest of many new cultivars is still lacking or insufficient. Therefore, the objective of this study was to document the TYLCV resistant and horticultural characteristics of currently available TYLCV-resistant tomato cultivars under commercial field conditions.
Materials and Methods
The trial was conducted in a commercial farm near Immokalee; FL. Twelve TYLCV-resistant varieties were evaluated and compared to two susceptible grower standards in a completely randomized experimental design with three replications, respectively (Table 1). Two additional treatments included grafted plants: (1) resistant TYLCV (root-stock)/susceptible TYLCV (scion), and (2) susceptible TYLCV (root-stock) /resistant TYLCV (scion).
Seed were planted in flats and grown by Redi Plants, Corp. The field was rototilled, and the pre-plant fertilizer (bottom mix and top “hot” mix) was applied following the modified broadcast method to supply 300-60-462 lb.acre-1 of N-P2O5-K2O. Beds 36-inch wide and 9-inch tall and formed on 6-ft centers (1 acre = 7,260 linear bed feet). Beds were then fumigated with methyl bromide and chloropicrin (50:50, w:w)
at the rate of 200 lb.acre-1. All beds were immediately covered with aluminized VIF polyethylene mulch. On 9 Jan 2009, transplants were established in the field at a within-row spacing of 22 inches, which created a stand of approx. 4,035 plants/acre. Plots were 36-ft long in 2008 (20 plants). On 28 days after transplanting (DAT), each tomato variety was pruned following the seed company’s specifications (Table 1). The field was seepage irrigated and tomato plants staked and tied. Pesticide applications were made as needed in response to regular scouting reports according to UF/IFAS recommendations (Olson et al., 2006). Ten tomatoes plants were harvested three times on 21 April, 6 and 20 May 2009 (102, 117 and 132 DAT).
The whitefly (Bemisia argentifolii) population was monitored using a leaf-turn method and TYLCV symptomatic plants were counted at the third harvest 20 May 2009. The number of plants showing symptoms of fusarium crown rot (caused by Fusarium oxysporum f.sp. radicis-lycopersici) in each plot was also noted at third harvest 20 May 2009. Bacterial spot (caused by Xanthomonas campestris) was rated as percentage foliar cover at the third harvest 20 May 2009. Tomatoes were graded in the field using a potable grading table according to USDA specifications for extra-large (5x6), large (6x6), and medium (6x7) fruit categories (USDA, 1997). Total unmarketable tomato fruit numbers were recorded and categorized into odd shape (OS) blossom end scar (BES), discoloration (DISC), and scaring (SC), as described by Gilreath et al. (2000).
32

After harvest, tomatoes were placed in 25-lb boxes and transported to the Garguilo, Inc. packing house (Immokalee, FL.). After 6 days of ethylene ripening treatment, post-harvest evaluation was done in the first two harvests at the UF Post-harvest Horticulture Laboratory in Gainesville, FL. Tomatoes were held overnight at 68°F, and the four uniform fruits from each treatment were selected the next day. These fruits were stored at 68°F until they reached table-ripe stage defined as “the point at which red-ripe tomatoes became noticeably softer when pressure was applied with thumb and fingertips to the equatorial region of each fruit”. Soluble solids concentration (SSC), total titratable acidity (TTA), and pH were measured at the table-ripe stage. Frozen pulp samples were thawed and homogenized then centrifuged at 15,000 RPM for 20 min at 41 ºF. The supernatant was filtered through cheesecloth, and a 0.013 lb aliquot of filtrate was used to asses SSC (using a Mark Abbe II digital refractometer; Reichert-Jung, Model 10480, Depew, NY), pH (using a Corning 140 pH meter, Medfield, MA), and TTA (by titration with 0.1N NaOH using a Fisher Model 395, Pittsburg, PA). Fruit were stored until red-ripe (3 days) at 20 °C; a subsample of three tomatoes were taken per replicate per variety and individual fruits were measured for firmness. Firmness was tested using a deformation meter (1 kg-force applied for 5 sec (11-mm convex probe) at the fruit equator. Higher values indicate softer fruit.
A field day was conducted at first harvest 21 April 2009. Participants using a 1-to-5 scale (1= very poor; 5=very
good) in a blind test evaluated the varieties in earliness, plant vigor, fruit size, firmness, fruit quality, potential yield and an overall plant rating at first harvest 21 April, 2009. Data yield components, SSC, pH, TTA, firmness, and field evaluation to TYLCV variety responses were analyzed using ANOVA and Duncan’s Multiple Range Test at 5%. Amount of fruit defects by TYLCV, fusarium crown rot, and bacterial spot percentage by variety were transformed by Arcsin distribution before the ANOVA and Duncan’s Multiple Range Test (SAS, 2002).
Results and Discussion
Overall weather conditions during the trials in South Florida recorded by the Florida Automated Weather Network (FAWN) was cool and dry with five (21-23, Jan., 5 Feb. and 3 Mar.) freeze events during the spring of 2009. Rainfall during the season showed accumulations of 3.0 inches.
Whitefly population, TYLCV incidence, bacterial spot, and fusarium crown rot rating: Whitefly pressure was low in spring 2009, with an average whitefly count of 0.5 to 1 ± 1.0 and 2 to 5 ± 1.0 adult per leaf at the beginning and end of the season, respectively. The were no TYLCV symptomatic plants among varieties (Table 1). There were no significant differences in bacterial spot or fusarium crown rot among TYCLV varieties at third harvest (Table 1). The incidence of bacterial spot range from 25 to 37% and fusarium crown rot was 0 to 47% (P ≤ 0.05).
Fruit yields: First harvest extra-large ranges from 336 to 1,733 boxes/acre, total extra-large (all harvest combined) 462 to 2,165 and total harvest (all sizes
33

and harvest combined) 1,549 to 3,210 boxes/acre (Tables 2-4).
First harvest of extra-large fruit yields and total first harvest (all sizes combined) are given in Table 2. First and second combined extra-large, total first and second harvest (all sizes combined) are given in Table 3. Total extra-large and total harvest (all sizes and harvest combined) are given in Table 4. By all measures, yields were greater for ‘Tygress’, ‘Tygress on FL47’ grafted plants and ‘Sur 200’ than the rest of the varieties (P<0.05). However, ‘Tygress’, Tygress on FL47’ grafted plants and ‘Sur 200’ total first harvest (all sizes combined), first and second harvest combined (all sizes combined), total harvest (all sizes and harvest combined) were not significantly different than ‘BHN 765’, HM 8845, FL 47 on Tygress’ grafted plants, FL 47 (control) and Sebring (Control).
Total unmarketable tomato categories for first harvest, first and second harvest combined (all sizes combined) and total harvest (all sizes and harvest combined) were lowest with ‘Tygress’, ‘Tygress on FL47’ grafted plants, ‘FL 47’ on ‘Tygress’ grafted plants, ‘Sur 200’, ‘FL 47’ and ‘Sebring’ and the highest ‘BHN 765’ ‘FLA 8577’ and ‘8578’, ‘HA 3096’, and ‘Sak 5230’ (Table 5). The large number of unmarketable (odd shape, blossom end scar, discoloration, and scar) fruit found with TYLCV-resistant cultivars was consistent with results from spring 2007 and 2008 (Ozores et al., 2008), but in contrast to other studies (Gilreath et al, 2000; Scott, 2004 and Cushman and Stansly, 2006).
The most common defect types as percentage of the total unmarketable yields harvested per TYLCV variety were scaring (SC) and odd shape (OS) followed by blossom end scar (BES) and discoloration (DISC) (Table 5). TYLCV varieties with the highest percentage of odd shaped fruit were ‘Tycoon’, ‘FL 47’, and ‘FL 47’ on ‘Tygress’. Those with the lowest percentage were ‘BNH 765’, ‘HA 3075’ (‘Ofri’), ‘HA 3095’, and ‘Sak 5230’. Highest percentage of blossom end scar were ‘FL 8578’ and ‘Sak 5230’ and lowest with ‘SUR 200’, ‘Tygress’, ‘FL 47’ on ‘Tygress’, ‘FL 47’ and ‘Sebring’. The highest percentage of scaring occurred with ‘SUR 200’, ‘Tygress’ and ‘HM 8845’ and the lowest with ‘FL 8578’.
Post-harvest evaluation: There were no significant differences in firmness, pH, SSC, and TTA among TYLCV varieties (Table 6).
The Extension activities: A field day held at the grower’s cooperator in Immokalee was well-attended (65 attendees). According to 23 responses from participants in the blind test, best overall TYLCV varieties for the spring 2009 were ‘Tygress’, ‘Sur 200’ and ‘HM 8845’ based in earliness, plant vigor, fruit size, firmness, fruit quality, potential yield and an overall plant rating (Table 7).
Previously, no clear advantage or disadvantage has been demonstrated by using TYLCV resistant varieties under conditions of low or no virus pressure during springs TYLCV variety evaluation (Cushman and Stansly, 2006; Ozores et al., 2008). In contrast, this year ‘Tygress’ demonstrated a better performance in the extra-large fruit
34

category over the standard susceptible ‘FL 47’ and ‘Sebring’.
The authors wish like to thanks to Sakata, Hazera, Harris Moran, Seminis, BHN, Redi-Plants, and Six’L Farms for providing monetary or in-king support to this project.
References
Cushman, K and P. A. Stansly. 2006. TYCLV-resistant tomato cultivar trial and whitefly control. Proceedings: Florida Tomato Institute. P. Gilreath [Ed.], Vegetable Crops Special Series, IFAS, U. of Florida, Gainesville, pp. 29-34.
Gilreath, P., K. Shuler, J. Polston, T. Sherwood, G. McAvoy, P. Stansly, and E. Waldo. 2000. Tomato yellow leaf curl virus resistant tomato variety trials. Proc. Fla. State Hort. Soc. 113:190-193.
Olson, S.M., W.M. Stall, M.T. Momol, S.E. Webb, T.G. Taylor, S.A. Smith, E.H.
Simonne, and E. McAvoy. 2006. Tomato production in Florida, pp. 407-426 In: S.M. Olson and E. Simonne (Eds.) 2006-
2007 Vegetable Production Handbook for Florida, Vance Pub., Lenexa, KS.
Ozores-Hampton, M.P., G. McAvoy, E.H. Simonne, and P. Stansly. 2008. Evaluation of TYLC virus-resistant varieties under commercial conditions in Southwest Florida. Fla. Tomato Inst. Proc. PRO525, pp.12-17.
SAS. 2002. SAS/STAT user’s guide, Ver. 9.1, SAS Institute, Cary, NC
Schuster, D. J., P. A. Stansly and Jane E. Polston. 1996. Expressions of plant damage of Bemisia. In: Bemisia 1995: Taxonomy, Biology, Damage Control and Management. Andover, Hants, UKD.D. Gerling and R. T. Mayer {Eds.} PP: 153-165.
Schuster, D. and J. Polston, 1999. Whitefly management guide: Tomato yellow leaf curl virus. Citrus and Vegetable, July, A6-A7.
USDA.1997. United States standards for grades of fresh tomatoes. Agr. Markt. Serv. http://www.ams.usda.gov/standards/tomatfrh.pdf.
35

Table 1. Cultivars and advanced breeder’s varieties evaluation during spring 2009.
Variety Source Number of
Suckers Pruned
Virus incidence y
(%)
Bacterial Spot Rating
y (%)
Fusarium crown rot y
(%)
BHN 765 BHN Seed No 0 31.7 0.0 FLA 8577 UFz 3 0 25.0 0.0 FLA 8578 UF 3 0 30.0 10.0
HA 3075 (Ofri) Hazera 3 0 33.3 0.0 HA 3095 Hazera 3 0 30.0 10.0 HA 3096 Hazera 3 0 31.7 20.0 HM 8845 Harris Moran 2 0 31.7 0.0 Sak 5230 Sakata 3 0 33.3 13.3 Sak 5443 Sakata No 0 36.7 10.0 SUR 200 Seminis 3 0 30.0 6.7 Tycoon Hazera 3 0 35.0 46.7 Tygress Seminis 3 0 28.3 0.0
FL 47 (control) Seminis 3 0 28.3 6.7 Sebring (control) Syngenta 3 0 25.0 0.0
Grafted
Tygress on FL 47 Seminis 3 0 33.3 0.0 FL 47 on Tygress Seminis 3 0 30.0 13.3
P value - - - 0.81 0.10 Sig. - - - ns ns
z University of Florida y Percentage of TYLVC-affected plants, Bacterial spot, Fusarium crown rot at the end of trial, after third harvest. Values are means of three replications of 10 plants. x Within columns, means followed by different letters are significantly different according to Duncan’s Multiple Range Test at 5%. ** Significance at P ≤ 0.01 * Significance at P ≤ 0.05 ns Not significance
36

Table 2. First harvest marketable fruit yield by size categories for selected tomato varieties, Immokalee, FL, grown in Spring 2009. Variety Marketable Yield (Boxes Z /acre)
XLy Ly My Total Unmarketable BHN 765 1,031c x 437 164ab 1,632abcd 908a FLA 8577 575def 445 188ab 1,209efgh 706abc
FLA 8578 336f 351 194ab 881h 417cde HA 3075 (Ofri) 390f 335 192ab 917h 460bcde
HA 3095 457ef 288 214a 959h 531abcd HA 3096 684cdef 316 147ab 1,147fgh 843ab HM 8845 965c 348 138abc 1,452cdefg 358cde Sak 5230 520ef 368 165ab 1,054gh 742abc Sak 5443 812cde 394 182ab 1,387defg 461bcde SUR 200 1,369b 335 121abc 1,824abc 229de Tycoon 922cd 327 129abc 1,378defg 506abcde Tygress 1,733a 225 35c 1,992a 269de
Tygress on FL 47 1,422ab 436 94bc 1,952ab 142de FL 47 on Tygress 963c 401 134abc 1,498cdef 185de FL 47 (control) 985c 456 136abc 1,577bcde 180de
Sebring (control) 407f 395 144ab 946h 93e P value 0.0001 0.29 0.0817 0.0001 0.0004
Sig. ** Ns * ** **
z 25-lb tomatoes/box y XL= Extra-large (5x6 industry grade); L=Large (6x6); M=Medium (6x7). x Within columns, means followed by different letters are significantly different according to Duncan’s Multiple Range Test at 5%. ** Significance at P ≤ 0.01 * Significance at P ≤ 0.05 ns Not significance
37

Table 3. First and second (combined) harvest marketable fruit yield by size categories for selected tomato varieties, Immokalee, FL, grown in Spring 2009.
Variety Marketable Yield (Boxes Z /acre)
XLy Ly My Total Unmarketable BHN 765 1,413bcd x 721b 367bcde 2,501ab 1,746a FLA 8577 858efgh 683bc 432abcd 1,972cd 1,469a FLA 8578 452h 529bcd 454abc 1,435e 1,566a
HA 3075 (Ofri) 715fgh 717b 609a 2,041bcd 1,160ab HA 3095 772efgh 514bcd 551ab 1,837cde 1,194ab HA 3096 1,055defg 516bcd 390bcde 1,962cd 1,612a HM 8845 1,406bcd 733ab 526ab 2,665a 788bc Sak 5230 647gh 534bcd 354bcde 1,534de 1,707a Sak 5443 1,080defg 703b 528ab 2,311abc 1,101abc SUR 200 1,812ab 472cd 218ef 2,502ab 569bc Tycoon 1,222cde 515bcd 300cdef 2,037bcd 1,109abc Tygress 2,112a 364d 118f 2,595a 657bc
Tygress on FL 47 1,863ab 635bc 250def 2,748a 578bc FL 47 on Tygress 1,613bc 731ab 387bcde 2,732a 479c FL 47 (control) 1,496bcd 690bc 321cde 2,507ab 649bc
Sebring (control) 1,178cdef 930a 535ab 2,643a 491c P value 0.0001 0.0003 0.0001 0.0001 0.0001
Sig. ** ** ** ** **
Z 25-lb tomatoes/box y XL= Extra-large (5x6 industry grade); L=Large (6x6); M=Medium (6x7). x Within columns, means followed by different letters are significantly different according to Duncan’s Multiple Range Test at 5%. ** Significance at P ≤ 0.01 * Significance at P ≤ 0.05 ns Not significance
38

Table 4. Total harvest marketable fruit yield by size categories for selected tomato varieties, Immokalee, FL, grown in Spring 2009.
Variety Marketable Yield (Boxes Z /acre)
XLy Ly My Total Unmarketable BHN 765 1,457bcd x 747bcde 464cdef 2,675abc 2,524a FLA 8577 863efg 727bcdef 558cde 2,148cde 2,012abc FLA 8578 462g 543defg 543cdef 1,549e 2,007abc
HA 3075 (Ofri) 757efg 791b 792ab 2,339bcd 1,725abcd HA 3095 801efg 530defg 614bcd 1,945de 1,578bcd HA 3096 1,070def 523efg 429cdef 2,022de 2,222ab HM 8845 1,417bcd 764bc 629bcd 2,810ab 1,301cd Sak 5230 663fg 563cdefg 413def 1,639e 2,130ab Sak 5443 1,106def 748bcd 655bc 2,509bcd 1,695bcd SUR 200 1,876ab 515fg 372ef 2,763abc 928d Tycoon 1,233cde 543defg 368ef 2,144cde 1,634bcd Tygress 2,165a 429g 309f 2,903ab 1,023d
Tygress on FL 47 1,920ab 739bcde 503cdef 3,162a 928d FL 47 on Tygress 1,697abc 815b 661bc 3,173a 1,232cd FL 47 (control) 1,568bcd 781b 598bcde 2,947ab 1,140d
Sebring (control) 1,247cde 1,057a 906a 3,210a 1,098d P value 0.0001 0.0001 0.0001 0.0001 0.0004
Sig. ** ** ** ** **
Z 25-lb tomatoes/box y XL= Extra-large (5x6 industry grade); L=Large (6x6); M=Medium (6x7). x Within columns, means followed by different letters are significantly different according to Duncan’s Multiple Range Test at 5%. ** Significance at P ≤ 0.01 * Significance at P ≤ 0.05 ns Not significance
39
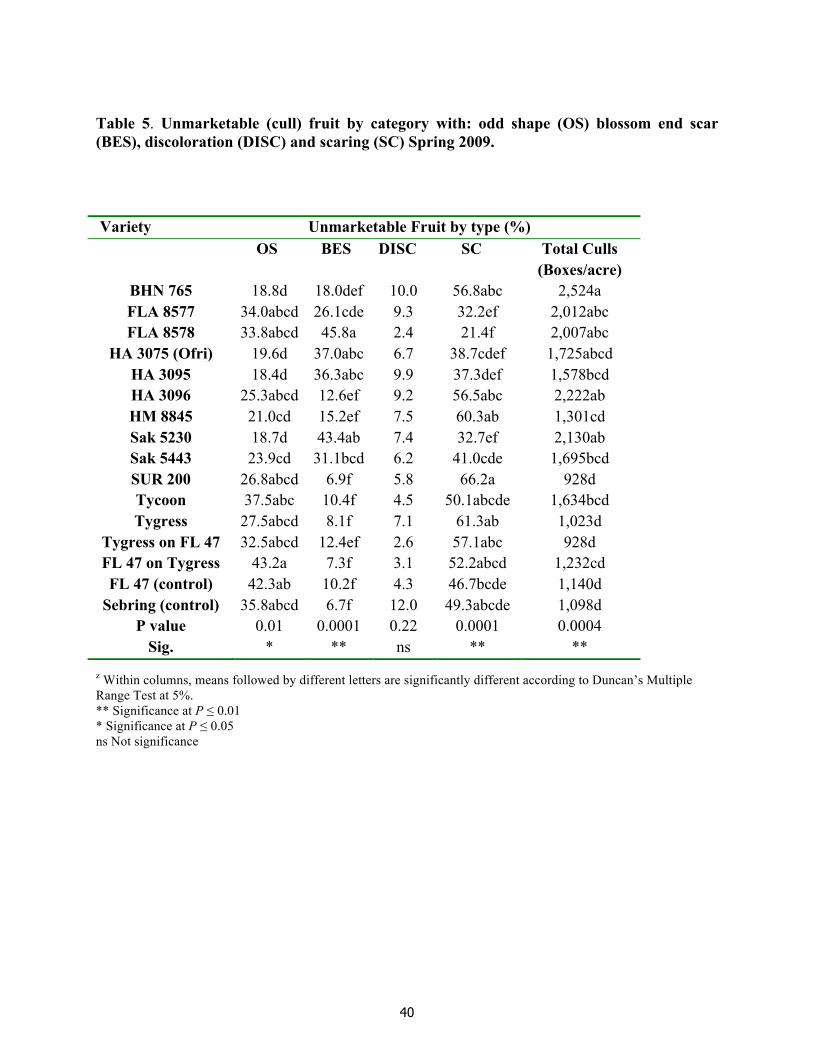
Table 5. Unmarketable (cull) fruit by category with: odd shape (OS) blossom end scar (BES), discoloration (DISC) and scaring (SC) Spring 2009.
z Within columns, means followed by different letters are significantly different according to Duncan’s Multiple Range Test at 5%. ** Significance at P ≤ 0.01 * Significance at P ≤ 0.05 ns Not significance
Variety Unmarketable Fruit by type (%) OS BES DISC SC Total Culls (Boxes/acre)
BHN 765 18.8d 18.0def 10.0 56.8abc 2,524a FLA 8577 34.0abcd 26.1cde 9.3 32.2ef 2,012abc FLA 8578 33.8abcd 45.8a 2.4 21.4f 2,007abc
HA 3075 (Ofri) 19.6d 37.0abc 6.7 38.7cdef 1,725abcd HA 3095 18.4d 36.3abc 9.9 37.3def 1,578bcd HA 3096 25.3abcd 12.6ef 9.2 56.5abc 2,222ab HM 8845 21.0cd 15.2ef 7.5 60.3ab 1,301cd Sak 5230 18.7d 43.4ab 7.4 32.7ef 2,130ab Sak 5443 23.9cd 31.1bcd 6.2 41.0cde 1,695bcd SUR 200 26.8abcd 6.9f 5.8 66.2a 928d Tycoon 37.5abc 10.4f 4.5 50.1abcde 1,634bcd Tygress 27.5abcd 8.1f 7.1 61.3ab 1,023d
Tygress on FL 47 32.5abcd 12.4ef 2.6 57.1abc 928d FL 47 on Tygress 43.2a 7.3f 3.1 52.2abcd 1,232cd FL 47 (control) 42.3ab 10.2f 4.3 46.7bcde 1,140d
Sebring (control) 35.8abcd 6.7f 12.0 49.3abcde 1,098d P value 0.01 0.0001 0.22 0.0001 0.0004
Sig. * ** ns ** **
40

Table 6. Tomato TYLCV post-harvest evaluation at first harvest during spring 2009.
Variety Firmness
(1/100") pH SSC
(oBrix) TTA
(mEq/100g juice) BHN 765 14.00 4.44 4.23 0.38 UF8577 16.33 4.40 4.10 0.42 UF8578 12.33 4.42 3.77 0.40 HA 3075 (Ofri) 11.00 4.44 4.10 0.36 HA 3095 13.33 4.43 2.73 0.31 HA 3096 14.00 4.41 3.40 0.36 HM 8845 14.67 4.50 4.43 0.36 Sak 5230 12.00 4.39 2.97 0.36 Sak 5443 13.67 4.49 3.20 0.31 SUR 200 11.33 4.58 3.33 0.40 Tycoon 13.00 4.37 3.77 0.34 Tygress 10.67 4.54 3.97 0.36 Tygress on FL 47 14.00 4.52 4.30 0.40 FL 47 on Tygress 13.33 4.49 4.47 0.43 FL 47 (Control) 9.67 4.50 3.87 0.37 Sebring (Control) 11.67 4.54 4.00 0.34
P values 0.09 0.56 0.12 0.45 Sig ns ns ns Ns
z Within columns, means followed by different letters are significantly different according to Duncan’s Multiple Range Test at 5%. ** Significance at P ≤ 0.01 * Significance at P ≤ 0.05 ns Not significance
41

Table 15. Field TYLCV variety blind evaluation spring 2009 (tomato plant and fruit evaluation was calculated from the contribution of 23 participants based in rating scale 1-5; 1= very poor and 5 = very good).
Variety Field Tomato Plants and Fruit Evaluation
Earliness type
Plant vigor
Fruit size Firmness Fruit
quality Yield
potential Overall
rate BHN 765 4.0b z 3.6cde 3.6b 3.3b 3.4cde 4.4a 3.6b FLA 8577 3.3cd 3.3def 2.6g 3.0b 2.7fg 3.9b 2.8cd FLA 8578 3.2d 3.0fg 2.7efg 3.0b 2.5gh 2.8de 2.3ef
HA 3075 (Ofri) 3.1de 3.6cde 2.6g 3.1b 2.4gh 2.4e 2.3ef HA 3095 2.8ef 3.1f 2.5g 3.2b 2.8fg 2.6e 2.5def HA 3096 4.5a 2.0i 2.4g 2.4c 2.2h 3.0cd 2.2f HM 8845 3.7bc 3.0fg 3.7b 4.1a 4.0a 3.7b 3.7ab Sak 5230 3.7bc 3.1f 3.1cde 3.2b 3.1def 3.6b 3.0c Sak 5443 3.4cd 3.6cde 3.3bcd 3.8a 2.9f 3.8b 3.1c SVR 200 3.0def 3.2ef 3.4bc 3.9a 3.8abc 4.3a 3.7ab Tycoon 3.9b 2.3h 3.0def 3.0b 2.8fg 3.2c 2.6de Tygress 3.7bc 3.7dc 4.2a 4.0a 3.9ab 3.9b 4.0a
Tygress on FL47 3.0def 2.7gh 2.4g 2.9b 3.0ef 2.5e 2.3ef FL47on Tygress 2.7f 4.1ab 2.7fg 2.9b 3.6bc 2.5e 2.8cd FL 47 (control) 3.0def 3.8bc 2.7efg 3.1b 3.5cd 3.7b 3.1c
Sebring (control) 2.2g 4.4a 2.8efg 3.3b 3.4cde 3.0cd 2.9cd P values 0.0001 0.0001 0.0001 0.0001 0.0001 0.0001 0.0001
Sig ** ** ** ** ** ** ** z Within columns, means followed by different letters are significantly different according to Duncan’s Multiple Range Test at 5%. ** Significance at P ≤ 0.01 * Significance at P ≤ 0.05 ns Not significance.
42

Identification of Weed Reservoirs of Tomato Yellow Leaf Curl Virus in Florida Jane E. Polston Dept. of Plant Pathology, University of Florida Gainesville, FL and David J. Schuster GCREC, University of Florida Wimauma, FL
The recognition of wild plant reservoirs can be a very important component of the management of plant viruses. The role and relative importance of weed reservoirs in virus ecology varies with the virus, the crop and the location. The ecology of Tomato yellow leaf curl virus (TYLCV), a virus, which appeared in Florida in 1997 (Polston et al. 1997), is not well understood. The whitefly vector has a very wide reported range of plants upon which it will feed, more than 500 plant species. While we do know that there are alternative crop hosts for TYLCV, we do not know if wild plant species play any role as reservoirs. This information is important for the improvement of the management of TYLCV. One option that has been proposed is a crop-free period in the summer, between the spring and fall crops. This crop-free will not be effective if there are wild plant species present during the summer that will serve as reservoirs of TYLCV.
We conducted a survey of possible TYLCV reservoirs and collected plants in 2008-2009. This is a challenging project because there are hundreds of wild plant species in the Manatee-Hillsborough area. Many
species are widely dispersed. In addition, we are expecting that only a small percent of the plants of a susceptible species would be infected. To focus our collection and keep within the budget allotted, we did not collect from grasses or plants known not to be hosts of the whitefly vector. Most of the plant species were selected based on their ability to serve as hosts for whiteflies, and/or the presence of virus-like symptoms. We were looking for plants that could serve as sources of virus for young tomato plants. Different species of wild plants are present at different times of the year so we collected samples from plants at the early part of the spring season, and then at the end of the spring season in the Ruskin tomato production region. Five sites in Manatee Co. and one in Hillsborough Co. were sampled. Plants were sampled in and around tomato fields and came from the following types of locations: tomato field, ditch bank, edges of tomato fields, edges of other fields, woody field edge, fallow fields, and fence rows. The sites selected were those where we had observed TYLCV-infected tomatoes early in the season, and therefore where it was likely a weed host might exist.
Results to Date
Samples were collected, identified, and frozen for laboratory analyses for the presence of TYLCV. All samples were assayed for the presence of TYLCV using a nucleic acid spot hybridization assay (NASHA). Briefly, nucleic acid was extracted from frozen samples (Doyle and Doyle et al. year), blotted onto nylon membranes, and hybridized with a radioactively-labeled
43

probe made from the genome of TYLCV. While this assay allows the rapid processing of many samples, it is known to give false positive results, especially on plant samples that have a lot of latex and polysaccharides (present in many tropical wild plants). Therefore, the more specific and sensitive polymerase chair reaction (PCR) using primers, which will amplify TYLCV was conducted on all samples that gave a positive result in the NASHA. DNA was extracted from samples that were positive in the dot spot assay and a PCR was run using appropriate primers. Samples positive by this assay will be retested using a different set of primers to confirm the first results.
Between February 2008 and February 2009 we collected 1,920 plants from 45 known species of wild plants from 15 different plant families (Table 1). We are in the process of identifying the species of approximately 227 of those samples. All of the samples have been tested by NASHA for the presence of TYLCV. Approximately 326 samples gave a positive result in the NASHA. Of those 103 have already been tested by PCR and the rest are in the process of being tested. TYLCV was not detected in any of the plants tested. We conclude that the samples, which were positive by NASHA, were probably the result of non-specific binding of the probe to the sample.
This study will be completed within the next few months. At this point in the study, we only have two weed samples that may be infected with TYLCV; assays to confirm this are in progress. At this time there is no evidence to suggest that there is a wild
plant species that is an important reservoir for TYLCV in the summer months. If our data continues along this trend it would suggest that there are no obvious impediments, in terms of wild plant species, to the success of a tomato-free period during the summer months. This is not to say that there are no wild plant hosts, since our study was not exhaustive, but that we did not find any likely candidates.
Literature Cited
Polston, J.E., R. J. McGovern, and L.G. Brown. 1999. Introduction of Tomato yellow leaf curl virus in Florida and implications for the spread of this and other geminiviruses of tomato. Plant Disease 83:984-988.
44

Table 1. Wild plants sampled from the fields and tested for presence of TYLCV in and around tomato fields in Manatee and Hillsborough Co. Florida.
Family Species Common Name County
No. of
Samples Tested by NASHA
No. Positive by
NASHA
No. Samples Tested by
PCR
No. Positive by
PCR
Amaranthaceae Amaranthus viridis Slender amaranth Manatee 40 10 6 0
Amaranthus. spp. 10 0 Anacardiaceae Schinus terebinthifolius Brazilian pepper Manatee 20 0
Asteraceae Ambrosia artemisiifolia Common ragw eed Manatee 50 0
Bidens pilosa L. Spanish needle Manatee, Hillsborough 170 0
Bidens spp. Manatee 61 0 Heterotheca subaxillaris Camphorw eed Manatee 40 0 Lactuca canadensis Wild lettuce Manatee 20 0 Pseudognaphalium sp. cudw eed Manatee 20 0 Chenopodiaceae Chenopodium album L. Lambs quarters Manatee 19 0
C. ambrosioides L. Mex ican tea Manatee 80 0 C. sp. chenopodium Manatee 40 0
Commelinaceae Commelina diffusa Burm. f.
Spreading day flow er Manatee 20 0
Euphorbaceae Euphorbia hirta L. Garden spurge Manatee 10 0 Euphorbia spp. spurge Manatee 40 0
Poinsettia cyanthophero (Murray) Bartling Wild poinsettia Manatee 30 0
Ricinus communis L. Castorbean Manatee 14 0
45

Table 1 (cont’d). Wild plants sampled from the fields and tested for presence of TYLCV in and around tomato fields in Manatee and Hillsborough Co. Florida.
Family
Species
Common Name
County
No. of
Samples Tested by NASHA
No.
Positive by NASHA
No.
Samples Tested by
PCR
No.
Positive by PCR
Fabaceae Crotalaria spectabilis Roth show y crotalaria Manatee 10 0
Indigofera hirsuta L. Hairy indigo Manatee, Hillsborough 127 71 34 0
Melilotus alba White sw eet clov er Manatee 40 0
Phaseolus sp. (narrow leaf) phasebean 20 0
Phaseolus sp. (broad leaf) phasebean 10 7 4 0
Sesbania sp. Scop Hemp sesbania Manatee 71 40 33 1 Trifolium sp. Clov er sp. Manatee 21 0 Lythraceae Lagerstroemia sp. Crape my rtle Manatee 11 0
Malvaceae Abutilon permolle coastal indian mallow Manatee 7 0
Abutilon sp. Indian mallow Manatee 20 0 Sida spinosa L. Sida Manatee 31 0 S. rhombifolia L. Indian hemp Manatee 20 1
Sida. sp. Sida Manatee, Hillsborough 50 0
Urena lobata Caesar-w eed Manatee 111 23 8 0
46

Table 1 (cont’d). Wild plants sampled from the fields and tested for presence of TYLCV in and around tomato fields in Manatee and Hillsborough Co. Florida.
Family
Species
Common Name
County
No. of
Samples Tested by NASHA
No.
Positive by NASHA
No.
Samples Tested by
PCR
No.
Positive by PCR
Myricaceae Myrica cerifera w ax my rtle Manatee 5 4 2 0
Onagraceae Ludwigia peruviana Primrose w illow Manatee, Hillsborough 70 20
Polygonaceae Rumex crispus curly dock Manatee 20 20 Rubiaceae Richardia brasiliensis Brazilian pusley Manatee 30 0
R. scabra L. Florida pusley Manatee, Hillsborough 65 10 6 0
Solanaceae Physalis spp. Manatee 20 0
Solanum americanum Mill. Nightshade Manatee 42 0
Solanum esculentum L. Cultiv ated tomato Manatee 10 10
S. ptycanthum Dun. Eastern nightshade Manatee 21 0
S. viarum Dunal Tropical soda apple Manatee 10 0
Verbenaceae Lantana sp. Lantana Manatee 28 0 Phyla nodiflora Mat lippia Manatee 10 0 Unknow n Unknow n Unknow n Manatee 233 80 19 1
47

Analysis of Recent Sporadic Postharvest Decay Events.
Jerry A. Bartz Postharvest Pathologist IFAS University of Florida
Situation
This project was begun at the request of several Florida tomato growers in the spring of 2007. They requested help due to an ongoing, but sporadic upsurge in postharvest decay losses in Quincy, FL; South Carolina; and Virginia. Similar decay problems had been observed elsewhere (New Jersey, Ohio, Alabama, coastal North Carolina, and Tennessee). In Florida, the problem was observed in Quincy and West Coast production areas and was primarily associated with the fall harvests (August to November). The problem often starts (excessive decay observed in the ripening rooms) and stops (normal ripening and defect incidence) without apparent reason. It primarily affects standard round green fruit, whereas pink fruit harvested from the same field remain sound and healthy. Roma fruit are usually unaffected except on rare occasions (New Jersey, 2006). Financial losses associated with dumped fruit, repacking costs, adjustment in contracts, etc. endanger farm profitability and, in some situations, viability.
Case study
In the Eastern Shore production area of Virginia, severe postharvest losses due to an
unknown decay appeared early in July and persisted most of the season. The weather in this region had been generally moist. June had been relatively cool, whereas July and August were warm to hot. Three tropical storms affected the region between mid-June and early September. According to Eastern Shore weather records, measureable precipitation occurred on 50% of the days in June and July, whereas August was not as moist (7/31 days) although 9 in. of rainfall fell on Sept. 1. Additionally, according to SkyBit® leaf wetness information, on several days in early to mid-July that did not have measurable precipitation; leaves were wet for several hours each day. On other days the leaves were not wet at all, meaning that dew did not form on plant surfaces. During periods of precipitation, fields would remain persistently wet. Long periods of leaf wetness suggest fogs, which would limit the drying of the crop canopy. Both frequent rainfall and fogs would keep the plants and fruit moist. An absence of dew suggests a cloud cover and absence of nightly cooling. Cloud covers restrict plant transpiration (evaporation of water from leaf surfaces) and water that is absorbed by the roots accumulates in the above ground parts of the plant.
The fruit harvested during the period were called “tender” by growers and significant bruise damage was observed. Fruit quality was poor and packouts at certain houses on the Eastern Shore ranged down to the mid to upper 20% level. Certain lots of fruit were stored
48
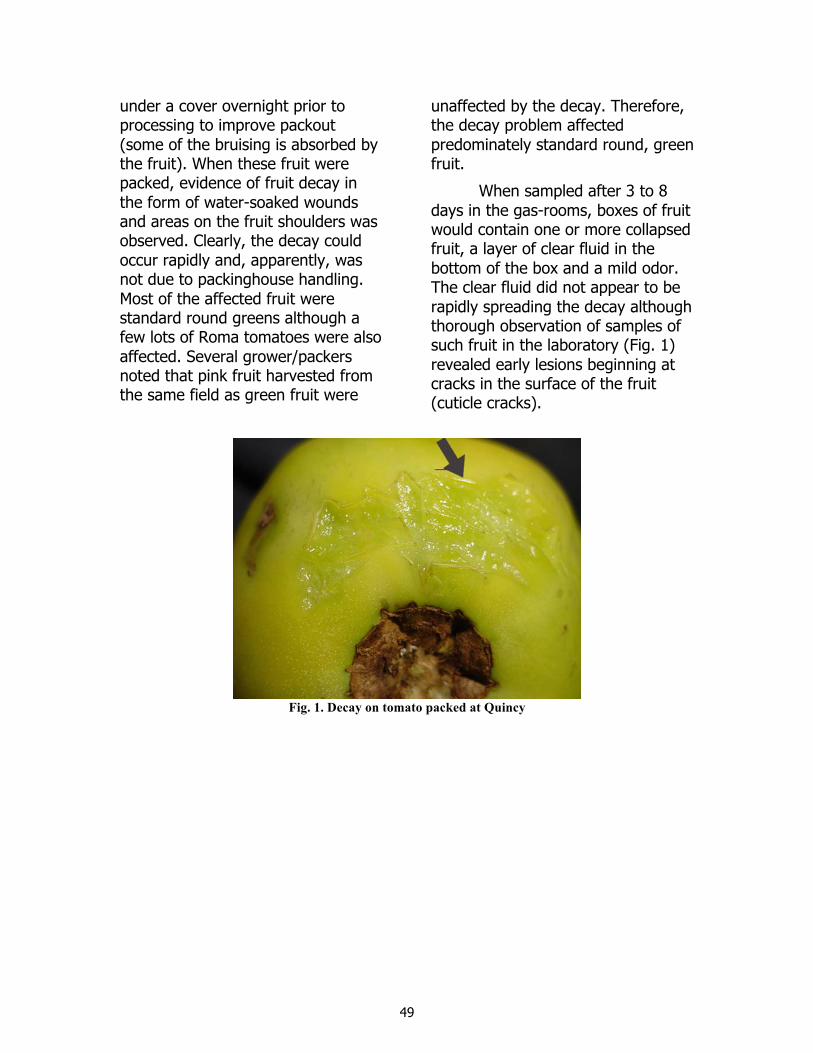
under a cover overnight prior to processing to improve packout (some of the bruising is absorbed by the fruit). When these fruit were packed, evidence of fruit decay in the form of water-soaked wounds and areas on the fruit shoulders was observed. Clearly, the decay could occur rapidly and, apparently, was not due to packinghouse handling. Most of the affected fruit were standard round greens although a few lots of Roma tomatoes were also affected. Several grower/packers noted that pink fruit harvested from the same field as green fruit were
unaffected by the decay. Therefore, the decay problem affected predominately standard round, green fruit.
When sampled after 3 to 8 days in the gas-rooms, boxes of fruit would contain one or more collapsed fruit, a layer of clear fluid in the bottom of the box and a mild odor. The clear fluid did not appear to be rapidly spreading the decay although thorough observation of samples of such fruit in the laboratory (Fig. 1) revealed early lesions beginning at cracks in the surface of the fruit (cuticle cracks).
Fig. 1. Decay on tomato packed at Quincy
49

Fig. 2. Quincy fruit on lab bench
Fruit received by my
laboratory developed a white mold over lesions within a day or two (Fig. 2.).
The growth was identified as Geotrichum candidum, cause of sour rot. The diseased fruit remained relatively intact over a several day period and the lesions enlarged relatively slowly, quite unlike the reports of a development of lesions within 24 h of harvest. Additional decay pathogens isolated from lesions included the soft rot pathogen, Erwinia carotovora subsp. carotovora and bacteria responsible for a lactic acid decay (Lactobacillus and Leuconostoc, spp.). However, G. candidum was consistently isolated from lesions, whereas the others were found in some lesions but not all. When inoculated in wounds of healthy green fruit, none of the isolated pathogens produced symptoms that matched those
observed in the boxes of fruit with decay. Thus, the role each pathogen played in the outbreak is unclear, although circumstantial evidence suggests that the bacterial pathogens were not major influences.
Individual lesions were water soaked, moist, and progressive, with or without a brown discoloration at the margin. When lesions encompassed the locular cavity of fruit, the gel holding the seeds was completely liquefied, whereas the solid tissue surrounding the cavity remained relatively firm and only slowly dissolved. This lesion progression is in contrast to typical bacterial soft rot, which progresses through the walls and locular cavity relatively quickly and uniformly. Moreover, with bacterial soft rot, the fruit completely collapses within 3 to 5 days (Figs. 3).
50

Fig. 3. Bacterial soft rot
The diseased fruit retrieved
from packinghouse ripening rooms remained relatively firm for a week or more. Additionally, the pH of the decaying tissues was typically 4.4 or lower, whereas that for bacterial soft rot is above 5.0 (Bartz, 1980). The disease syndrome (progression of the disease over time) observed in the packinghouse fruit appeared to account for the pool of clear fluids in boxes of fruit. All visual, sensory (odor of diseased fruit), isolation frequency and chemical (pH) evidence implicated sour rot as the major decay. This was consistent with the reports of sour rot being identified as the cause of a postharvest disease of tomato at disease clinics in other Eastern US growing areas.
The epidemic began on the Eastern Shore production area in early July and continued for the rest of the summer, as noted above. All growers in that area experienced major losses. Early harvests at Quincy were affected as was at least
one field in Georgia (Roma’s) but by mid October the problem had disappeared. At least two fields in the West-Coast production area of Florida had similar decay issues. A decay issue also affected Roma types produced in New Jersey during the summer months; reports back from that Eastern US production area indicated sour rot was diagnosed.
The Literature
Initially, attributing the cause of the losses to sour rot caused by G. candidum appeared implausible because this fungus is not considered to be a destructive pathogen of green tomato fruit (Butler, 1960). The disease was first reported in 1922 as being widely distributed in fields in New Jersey (Poole, 1922). The author noted that an unusually high number of fruit on the plants were developing severe cracks, likely due to the weather. If the fruits were red, a white fungus developed in the cracks and rotted
51

the fruit within 2 to 5 days. Cracks on green fruit were not diseased. A year later Pritchard and Porte (1923) published a report about a “watery-rot” of tomatoes that was observed by Federal market inspectors in shipments of “southern grown tomatoes.” They noted that green fruit were readily infected through punctures although their method of surface sanitizing a fruit may have affected its innate resistance to rot. Butler (1960) extensively studied the disease as it affected tomato production in California and claimed that the pathogen would not infect green fruit unless they had been chilled, either in the field or postharvest. By contrast, sour/watery rot was the most common decay cited in USDA inspection reports of California and Florida tomato arrivals in the NYC terminal over the period 1972-1984 (Ceponis, et al. 1986). Since most of these fruit would have been harvested when green and since the most common weather for postharvest decays are during warm moist periods, it seems unlikely that these market losses involved chilled fruit. Bartz (1980) observed the disease in a box of green tomatoes that had been shipped from Quincy to Orlando in early July, which is not a time that green tomatoes would have been chilled. The shipment had been rejected because of progressive decay. Sour rot was one of several decays observed in the box, but did not appear to be the most aggressive one.
With citrus fruit, sour rot incidence and severity are associated
with the water content of the peel. More water equals greater susceptibility. The pathogen is a different species, G. citri aurantifolia, but is a strict wound invader like G. candidum in tomato (Baudoin and Eckert, 1982, 1985). With lemons grown in California, more sour rot was observed when the fruit contain more water such as in coastal as compared with inland grown fruit; fruit harvested after rainfall versus fruit harvested in dry weather; and fruit harvested at 8 AM as compared with 2 PM. Also, the susceptibility decreases with time after harvest as the fruit peels lose moisture. When introduced into wounds on low water content peels, the pathogen grew poorly and caused an “arrested lesion” characterized by an area of slight necrosis around the wound. Flooding the wound surface with water activated arrested lesions if such action was taken on 5 but not 10-day old arrested infections.
The free water content of plant tissues ranges from deficient (wilting) to excessive (free water fills intercellular spaces and restricts air movement). Excessive water was defined by Johnson (1947) as “water congestion,” which is simply an “accumulation of excessive water in the intercellular spaces.” Further, Johnson (1947) wrote that water congested plant tissues “may completely surrender to invasion by a variety of organisms, including saprophytes. . . .” Johnson’s studies involved plant leaves, but tomato fruit can also become congested with water. In greenhouse production, excessive water produces cracks in
52

the surface of the fruit usually beginning on the shoulders and sometimes down toward the equator (Dorais et al., 2004). The resulting condition is called cuticle cracking and if severe, leads to russeting, which destroys the fruits’ marketability. In field production, a similar surface cracking can be caused by excessive rainfall or exposure of fruit to sunlight, particularly during periods of warm air temperatures (Emmons and Scott, 1997 and 1998). At a commercial packinghouse, fruit with large cracks or extensive cuticle cracking (rain checks) would be culled, whereas fruit with mild cuticle cracks would likely be packed. Cuticle cracks, including the most severe form, rain check, could have been prominent in the case study above and may have been the reason for the low packout rates reported. If present and not healed,
cracks in green fruit could be invaded by decay pathogens including G. candidum although it is unclear if the invasion by the latter would have led to an arrested lesion as described for green tomato fruit (Butler, 1960) or a progressive lesion as occurs in water-congested citrus.
Research
Observations from fruit sampled from packinghouses in the case study about were compared with the literature. Watery rot was an apt description of the initial symptoms when boxes were opened (Fig. 4). Cuticle cracks including the severe form, rain check” were apparent (Figs 1, 2, and 4). Moreover, watery infections appeared to begin at cuticle cracks (Figs 1 and 4), whereas lesions exposed to air became covered with a white mold and the fruit only slowly decayed (Fig. 2).
Fig. 4. Watery rot symptom in fruit from Quincy.
53

Efforts in the laboratory included
1. Determine if water congestion affects the susceptibility of tomato fruit to G. candidum.
2. Find if water congestion of green fruit enables a progressive decay
3. Determine if an exposure of freshly wound-inoculated fruit to free water could lead to a progressive sour rot.
Fruit of various stages of ripeness were wounded with a no. 3
cork borer (wound diameter = 6 mm (1/4 in) and depth = about 1/8 in). Alternatively, smaller wounds (1/8 in diameter and 1/8 deep) were made with a pointed probe. Spores suspended in water were added to each wound or the fruit were dipped into a spore suspension. The inoculated fruit were stored under various humidity regimes at 75oF.
With fruit wounded with the cork borer, lesion development was apparent on fully red fruit within 48 h and by 72 h were quite extensive (Fig 5).
Fig. 5. Extensive sour rot-72 hrs
By contrast, green fruit remained healthy and could be fully ripened
without evidence of the decay (Fig. 6).
54

Fig. 6. Inoculated when green, now red
The storage for both groups
of fruit featured high humidity. Next, we attempted to water congest fruit by submerging them in water for 18 to 24 h. One set of fruit was inoculated before being submerged and a second set was inoculated after being submerged. The control fruit were wounded but not submerged or wounded and submerged but not inoculated (clean water added to each wound). The water treatment led to an increase in
weight of the fruit by 1.6 to 6.1% of the original fruit weight. The green fruit absorbed more water than the red fruit. Soaking the fruit in an aqueous suspension of spores led to decay development in essentially the same areas that was infiltrated by the dye. Soaking green fruit in water predisposed them to sour rot, whether the fruit were inoculated before or after the water treatment (Fig 7 & 8).
Fig 7. Soak after inoculation
55

Fig. 8. Soak before inoculation
The symptoms on the green
fruit were similar to those observed in the outbreak tomatoes described in the case study above. The red fruit were still more susceptible than the green fruit, whereas water-congested tissues on the green fruit were susceptible. Use of smaller wounds led to similar results. Preliminary attempts to reverse water congestion by drying the green fruit with a fan after the soaking procedure were
unsuccessful. The moisture loss was not very rapid. The development of sour rot in inoculated green fruit was also promoted by keeping the fruit wet. Fruit with small wounds were wrapped in wet paper towels for up to 48 h. The towels were positioned so that the stem scar was open and normal air exchange could occur. By 48 to 72 h at 75oF, G. candidum was observed growing on the outside of the toweling over the wounds (Fig. 9).
Fig. 9. Partially covered for 48 hr.
Lesions had developed around
the wounds on the fruit partially enclosed by the wet paper towel (Fig 10).
56

Fig. 10. 72 hr after inoculation (covered with wet towel for 24 h).
The level of predisposition by
the wetness increases as the duration of the cover increases. In tests, where fruit were submerged briefly in a spore suspension of G.
candidum, the period that fruit were submerged and the water pressure on the submerged fruit markedly affected the severity of the decay (Fig. 11).
Fig 11. Increase of sour rot symptoms with increased exposure to inoculum/depth of penetration.
57

When fruit were congested by a dye solution, color was noted in the wall tissues around the wound but not directly into the gel (Fig. 12). Dye movement was also observed
into the stem scar. The area colonized within the green tomatoes after they’ve been soaked in water is the same as penetrated by a dye solution (Fig. 13).
Fig 12. Fruit treated with dye solution (soak 24 h or pressure treatment).
Fig 13. Cross section of green fruit that were wounded and inoculated (left column) or soaked in
dilute inoculum (right column).
Harvesting wet tomatoes or allowing fruit to become wet with condensation carries a substantial
decay risk if the moisture is not promptly dried. The risk is quite large if conditions in the field favor
58

water congested fruit and high levels of inocula.
Tests of several fungicides: Keyplex®, Actinovate®, CankerKill®, Proud-3®, Serenade Max®, ProPhyt®, K-Phite®, Omega-Grow®, Phyton B®, Biotune®, Kocide 3000®, Manex®, and Kocide + Manex were compared for direct toxicity (incorporated into an agar culture medium) and for ability to protect red fruit from developing sour rot. The two phosphite chemicals, which stimulate systemic acquired resistance, were applied to green fruit, which were then allowed to ripen before the wounds were made and inoculated. Subsequently, two additions to the chemicals tests included Mentor® (propiconazole) and Graduate® (fludioxonyl). The battery of tests included inoculation of green fruit with Erwinia carotovora subsp. carotovora, cause of bacterial soft rot.
Red fruit wounded, inoculated and then treated 4 or 6 h later with propiconazole did not develop sour rot lesions during up to 6 days of subsequent storage, whereas the control fruit were completely decayed by 6 days. The company, Syngenta Crop Protection, Inc., has funded several projects in different locations (California, Florida, Virginia) and is pursuing labeling the fungicide as a postharvest treatment. The treatment has been used on green fruit that were inoculated and then wrapped in wet towels. Results are still being evaluated although in several tests the propiconazole treated fruit had a similar level of
discoloration at wounds as compared with non-inoculated control fruit (Fig. 9). Currently, the fruit are being ripened to determine if the wounds become decayed as the fruits become red. None of the other materials tested provided more than a slight reduction in the severity of the decay. In contrast, applications of the Serenade Max product or a bacterial culture isolated from it, caused a slowly developing bacterial soft rot in green or red tomato fruit, particularly if the fruit were stored at 75 to 86oF under high humidity.
Summary
A severe outbreak of postharvest decay was observed in several eastern production areas during July through early November, 2006. Growers and packers termed the problem on-going but sporadic and seemingly increasing in severity. The major component of the decay, based on all evidence was found to be G.candidum, cause of sour rot. Although the literature does not indicate that G. candidum is an aggressive pathogen on green tomatoes, evidence was developed here that water-congested tissues or wet fruit surfaces enabled the fungus to infect green tomatoes. Control of the disease is currently problematic other than to avoid conventional high volume harvests of fruit from wet fields. Since water congestion accompanies persistent rainfall, heavy fogs and warm, moist field conditions at a time of reduced evaporation of moisture from plant canopies, avoiding harvests during those periods would be helpful.
59

Additionally, irrigation should be adjusted to plant needs; over irrigation at times of reduced transpiration is potentially hazardous. Fruit harvested when plants are wet require special handling as the longer the fruit remain wet, the greater the decay hazard. Bins of fruit that become wet due to rainfall or condensation also require special handling. If watery, water soaked areas are observed on fruit on the cull belt, the entire lot should be repacked after 48 to 72 h. At this time, boxes with one or more decayed fruit and fluids contaminating nearby fruit likely should be discarded. If a label is obtained for a propiconazole product, application of that product will control sour rot, but not bacterial soft rot.
Literature Cited
Bartz, J. A. 1980. Causes of postharvest losses in a Florida tomato shipment. Plant Dis. 64:934-937.
Baudoin, A. B. A. M. and Eckert, J. W. 1982. Factors influencing the susceptibility of lemons in infection by Geotrichum candidum. Phytopathology 72:1592-1597.
Baudoin, A. B. A. M. and Eckert, J. W. 1985. Development of resistance against Geotrichum candidum in
lemon peel injuries. Phytopathology 75: 174-179.
Butler, E. E. 1960. Pathogenicity and taxonomy of Geotrichum candidum. Phytopathology 50:665-672.
Ceponis, M. J., Cappellini, R. A., and Lightner, G. W. 1986. Disorders in Tomato Shipments to the New York Market, 1972-1984. Plant Dis. 70:261-265.
Dorais, M., Demers, D.-A., Papadopoulos, A. P., Van Ieperen, W. 2004. Greenhouse tomato fruit cuticle cracking. Hort. Rev. 30:163-184.
Emmons, C. L. W. and Scott, J. W. 1997. Environmental and physiological effects on cuticle cracking in tomato. J. Amer. Soc. Hort. Sci. 122:797-801.
Emmons, C. L. W. and Scott, J. W. 1998. Ultrastructural and anatomical factors associated with resistance to cuticle cracking in tomato (Lycopersicon esculentum Mill.). Int. J. Plant Sci. 159:14-22.
Johnson, J. 1947. Water-congestion in plants in relation to disease. Univ. Wis. Res. Bull., 160.
Poole, F. R. 1922. A new fruit rot of tomatoes. Botan. Gaz. 74:210-214.
Pritchard, F. J. and Porte, W. S. 1923. Watery-rot of tomato fruits. J. Agric. Res. 24:895-906.
60

The Potential Impacts Throughout the Market Channel from Food Safety Practices Implemented by Florida Tomato Growers and the Impact of Recent Food Safety Scares
John VanSickle University of Florida, Food & Resource Economics Dept., Gainesville, FL 32611, [email protected] University of Florida/IFAS
It is the responsibility of the food chain to insure that consumers are provided with a safe and abundant supply of fresh fruits and vegetables. Tomato producers have taken important steps to improve the safety of their products including the adoption of “Tomato Good Agricultural Practices” (T-GAP) and Tomato Best Management Practices (T-BMP) at the farm and in packinghouses. T-GAP and T-BMP are important to preventing contamination at the earliest stages of the supply chain, but it does not insure that contamination does not occur once it leaves the packinghouse. Recent food safety scares have caused problems for producers and the supply chain. Fresh tomatoes have been linked to large, multi-state outbreaks of food borne illness at retailer and food service outlets.
The impact of the increased concern for food safety has resulted in additional programs that increase the cost of growing and selling fresh market tomatoes. The impacts of these programs on growers and distributors need to be estimated. In addition, the regulatory framework for reacting to
food borne illnesses often results in unintended consequences to those producers who are not the source of the contamination.
The Cost of Food Safety Programs
The Florida tomato industry has been aggressive in addressing food safety concerns to pre-empt potential problems that could surface for their sector of the industry. There have been multiple food safety scares that have implicated tomatoes at different stages of the discovery process in these cases. The Florida industry has yet to be identified as a source of any identified critical problems in the industry. The Saintpaul Salmonella outbreak initially implicated tomatoes and did impact demand to the industry, in spite of the fact that weeks later it was traced to Serrano peppers grown in Mexico.
The Florida industry has been progressive in addressing food safety concerns. The industry worked with USDA to develop Good Management Practices that would minimize the threat of food safety concerns. These practices began in the field and were carried through the packinghouse operation. Discussion with industry leaders have estimated the cost of these programs as adding $100 per acre in preharvest growing costs and $0.10 per carton in harvest and marketing costs. The added harvest and marketing costs are generally thought to be collected from the buyers with the environmental impact fee that is collected on each carton of tomatoes sold. Assuming this to be the case, then impacts on growers are felt from the added costs at the field level that come from added burden of monitoring field activities and
61

documenting those activities to insure the food safety programs are being followed. Implementation of Good Management Practices for food safety have not impacted the productivity or increased the risk related to yield.
Representative Farm Model
A representative farm model for a grower of fresh market large round tomatoes in southwest Florida was developed to assess the impact of food safety programs implemented in Florida. The model was developed using Simetar© ( Richardson, et al., 2006), an Excel add-in that was developed explicitly for stochastic simulation modeling. Here, Simetar is used to model the representative activities of a Florida tomato grower located in Southwest Florida. The model simulates the farm’s financial results over a ten-year period based on production costs estimated by VanSickle et al. (2009) for a fall tomato crop of 272 acres. This grower is expected to incur variable costs of $6,400.52 for growing tomatoes in 2007/08 and harvest and marketing costs of $5,235 per acre for a yield of 1,500 cartons per acre. Based on annual cash operating and fixed costs and expected prices and yields an annual income statement, an annual cash flow statement, and balance sheets are forecast. Since this is a stochastic simulation, pseudo-random prices and yields are drawn from a multivariate empirical distribution. The random yields and prices are correlated based on historical correlations. Yields are estimated for the simulation based on recorded yields as reported by a grower who produced on 19 different fields in southwest Florida from 1998/99 to
2007/08. Because fields were not always planted and also because of missing data, the total number of observations provided by the grower was 71.
Prices for fresh market tomatoes are modeled from price data for southwest Florida provided by the Florida Tomato Committee (Florida Tomato Committee, Annual Reports 1999 – 2008). A probability distribution function for price was estimated from this data and correlated with yields to produce price and yield estimates for a simulation of the grower’s returns over a 10 year horizon given the state of technology that evolved from 1998/99 to 2007/08.
The simulation provides an estimate of the net worth provided by growing tomatoes given the current state of technology and incidence of bacterial spot. The model is then altered to remove the cost of implementing food safety practices to provide an estimate of net worth should the farmer operate with no risk associated with food safety. The difference between the simulations provide an estimate of the cost of food safety programs implemented at the grower level within this representative farm.
Results
The simulation for the 272 acre farm that grows tomatoes with food safety programs continuing shows that the change in the net present value of net worth is expected to be a loss of $172,520. The model indicates that there is an 86 percent probability that this grower will lose net worth if he continues to grow 272 acres over the
62

next 10 years. If the food safety practices are removed from the production program without any incidence of a food safety scare, then that expected loss in net worth is reduced to $153,417, a gain of $19,103 for this representative grower. The risk of losing net worth is still significant at 83 percent.
The simulation was also used to evaluate the value that this program needs to add in eliminating the threat of a food safety scare in tomatoes to offset the cost of producing tomatoes with food safety measures in place at the field level. The model was run to see how much premium must be added to price to offset the cost of the food safety program, to where the financial result (change in net worth) is the same as with the food safety program removed. The results indicate that a farm must realize a 1.7% premium in the value of their tomatoes as a result of implementing food safety production practices, with the expected outcome on net worth as good or better than it is without any food safety production practices in place. It is assumed this premium is a reflection of the added value that must be realized by growers from these programs.
Conclusions
Food safety is a serious issue to fresh tomato growers in Florida. The objective of this study was to estimate the cost of these programs to a representative grower of 272 acres of tomatoes and to estimate the premium
on returns they would need to realize to offset these costs. A stochastic farm level simulation model was used to model the production and financial activities of a representative grower over a ten-year horizon, using production budgets developed from growers in southwest Florida. The results suggest that a farm could needs to realize a premium of 1.7% on the price of tomatoes to offset the cost of food safety programs that have been implemented since 1996. The results indicate that the threat of losing net worth in the operation is reduced 25 percent by eliminating the threat to this disease. Additional work needs to be completed to assess how food safety risks have changed with these programs and the costs associated with any food safety event that could result. The models used in this research will be a valuable tool for assessing these risks and the potential outcomes to growers.
References
Florida Tomato Committee. 1999-2008. “Florida Tomato Committee Annual Report.” Florida Tomato Committee. Maitland, FL.
Richardson J.W., Schumann, K.D. & Feldman, P.A. 2006. Simetar© Simulation for Applied Risk Management, Version 2006 Texas A&M University, College Station, Texas.
VanSickle, John, Scott Smith and Eugene McAvoy. 2009. “Production Budget for Tomatoes in the Southwest Florida Area, 2007/08.” UF/IFAS EDIS forthcoming. Gainesville, Fl.
63