title page differential regulation of hepatic oct1...
TRANSCRIPT

JPET #205369
1
TITLE PAGE
Differential Regulation of Hepatic Oct1, Oatp1a4, Bsep, and Mrp2 Transporter Expression in
Lymphocyte-deficient Mice Associates with IL-6 Production
Connor E. Bodeman, Anika L. Dzierlenga, Catherine M. Tally, Ryan M. Mulligan, April D. Lake,
Nathan J. Cherrington, and Susan C. McKarns
University of Missouri School of Medicine, Center for Cellular and Molecular Immunology,
Department of Surgery, Columbia, MO 65212 (RMM, CMT, CEB, SCM)
University of Missouri School of Medicine, Department of Microbiology and Immunology,
Columbia, MO 65212 (SCM)
University of Arizona, Department of Pharmacology and Toxicology, Tucson, Arizona 85721
(ALD, ADL, NJC)
JPET Fast Forward. Published on August 8, 2013 as DOI:10.1124/jpet.113.205369
Copyright 2013 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
2
RUNNING TITLE PAGE
Running Title: Lymphocytes on hepatic cytokines and transporters
Address correspondence and reprint requests to Susan C. McKarns, University of Missouri
School of Medicine, Department of Surgery, M616 Medical Sciences Building, Columbia, MO
65212. 573-884-1722 (office), 573-882-4287 (fax) E-mail address:
No. of text pages: 29
No. of tables: 2
No. of figures: 5
No. of references: 54
No. of words in the Abstract: 248
No. of words in the Introduction: 749
No. of words in the Discussion: 1475
Abbreviations: ANOVA, analyses of variance; BA, bile acid; bDNA, branched DNA; bili, bilirubin;
BS, bile salt; Bsep, bile-salt export pump; BW, body weight; CAR, constitutive androstane
receptor; cMoat, canalicular multispecific organic anion transporter; CTL, cytotoxic T
lymphocytes; DAMPs, danger-associated molecular patterns; FoxP3, forkhead box P3; FXR,
farnesoid X receptor; GR, glucocorticoid receptor; GSH, glutathione; HNFα , hepatocyte nuclear
factor alpha; IFN-, interferon gamma; IL, interleukin; KO, knockout; LPS, lipopolysaccharide;
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
3
Mrp, multidrug resistance-associated protein; MU, University of Missouri; Na, sodium; NF-B,
nuclear factor of the B enhancer; NK, natural killer; NKT, natural killer T; Ntcp, Na+-
taurocholate cotransporting polypeptide; OA, organic anion; Oatp, organic anion-transporting
polypeptide; OC, organic cation; Oct, organic cation transporter; PAMPs, pathogen-associated
molecular patterns; PB, phenobarbital; PFIC-2, progressive familial intrahepatic cholestasis 2;
PPARα, peroxisome proliferator-activated receptor alpha; PXR, pregnane X receptor; Rag,
recombination-activating gene; RARα, retinoic acid receptor alpha; RLU, relative light unit; RXR,
retinoid X receptor; SHP, short heterodimer partner; SPGP, sister of P-glycoprotein; TCR, T cell
receptor; Tg, transgenic; TH, T helper; TLR, toll like receptor; TNF-α, tumor necrosis factor-alpha;
Tregs, T regulatory cells; VDR, vitamin D receptor; WT, wild-type
Recommended section assignment: Gastrointestional, Hepatic, Pulmonary, and Renal
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
4
ABSTRACT
Cholestasis results from interrupted bile flow and associates with immune-mediated liver
diseases. It is unclear how inflammation contributes to cholestasis. The aim of this study was
to determine whether T and B cells contribute to hepatic transporter expression under basal
and inflammatory conditions. C57BL/6J wild-type (WT) mice or strains lacking T, B, or both T
and B cells were exposed to lipopolysaccharide (LPS) or saline, and the livers were collected 16
h later. Branched DNA signal amplification was used to assess mRNA levels of organic anion-
transporting polypeptides (Oatp) 1a1, 1a4, and 1b2, organic cation transporter (Oct) 1,
canalicular bile salt export pump (Bsep), multidrug resistance-associated proteins (Mrp) 2 and
3, and the sodium-taurocholate co-transporting polypeptide (Ntcp). Real-time polymerase
chain reaction analyses was used to correlate changes of transporter expression with
interleukin (IL)-1b, IL-6, IL-17A, IL-17F, tumor necrosis factor (TNF)-and interferon (IFN)-
expression in the liver. LPS treatment inhibited Bsep and Oct1 mRNA expression, and this was
abrogated with a loss of T cells but not B cells. In addition, the absence of T cells increased
Mrp2 mRNA expression; whereas, B cell deficiency attenuated Oap1a4 mRNA in LPS treated
mice. Oatp1a1, Oatp1b2, Ntcp, and Mrp3 were largely unaffected by T or B cell deficiency.
Lymphocyte deficiency altered basal and inflammatory IL-6, but not TNF- or IL-1b, mRNA
expression. Taken together, these data implicate lymphocytes as regulators of basal and
inflammatory hepatic transporter expression and suggest that IL-6 signaling may play a critical
role.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
5
INTRODUCTION
The liver is a critical immunological and metabolic organ and is strategically positioned to
receive blood that is rich in bacterial and food-derived antigens from the gut via the portal vein.
Immune-mediated liver diseases are usually progressive and associate with significant
morbidity and mortality. Cholestasis can result from either a functional defect in bile formation
or from interrupted bile flow and is commonly associated with human liver diseases (Hirschfield
and Heathcote, 2009; Hirschfield et al., 2010). Genetic and environmental factors, including
infections and xenobiotics, can contribute to cholestasis. A major consequence of acute
cholestasis is the development of severe liver injury. Hepatocyte and cholangiocyte
proliferation that occurs in response to injury, in turn, leads to periductular fibrosis, biliary
fibrosis, and cirrhosis (Hirschfield et al., 2010). The mechanisms by which inflammation
contribute to the pathophysiology of cholestasis-mediated liver injury remain unclear.
Bile acids are synthesized in the liver from cholesterol, secreted into the bile, and are actively
transported to the gut and then back to the liver via enterohepatic circulation. Enterohepatic
bile flow is essential for the emulsification of dietary fat, fat soluble vitamin absorption, and
elimination of toxic compounds and is regulated through a coordinated hepatobiliary sinusoidal
(basolateral) and canalicular (apical) transport system (Merritt and Donaldson, 2009). Organic
cation transporter (Oct)1 is the major hepatic uptake transporter for small cations
ncreased expression of Mrp) provides and i multidrug resistance-associated protein ( 3
an alternative route for efflux during periods of bile acid and/or xenobiotic overload At the .
hepatic canalicular membrane, the ATP-dependent bile salt export pump (Bsep) and Mrp2
function as the primary hepatic bile acid exporters, reviewed in (Klaassen and Aleksunes, 2010).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
6
In concert, the sodium-taurocholate co-transporting polypeptide (Ntcp) is the predominant
mechanism of bile acid uptake at the sinusoidal membrane. The organic anion (OA)
transporting polypeptides (Oatp) 1a1, 1a4, and 1b2 transport conjugated bile acids, organic
acids, and xenobiotics. Disruption of transporter function results in cholestasis (Ricciardi et al.,
2001).
Lipopolysaccharide (LPS) is a common cause of inflammation–induced cholestasis in humans
and is widely used in experimental animal models to initiate cholestasis, presumably by
increasing synthesis of cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-6,
and IL-1b in response to toll-like receptor (TLR) 4-dependent nuclear factor of the B enhancer
(NF-B) activation in Kupffer cells. This model predicts that pro-inflammatory cytokines, in turn,
signal through their respective membrane receptors to suppress transporter expression and
function (Geier et al., 2006; Lickteig et al., 2007; Mulder et al., 2009). While the underlying
mechanisms remain unclear, LPS-induced alteration of transporter function appears to be
immune-mediated and the direct cause of cholestasis. Thus, LPS-induced cholestasis provides a
valuable experimental tool to study mechanisms of hepatic transporter regulation directly,
rather than secondarily in response to the accumulation of bile constituents resulting from
biliary obstruction.
The liver receives approximately 70% of its blood supply from the intestine through the portal
circulation and is thus exposed to potential gut-derived immune modulators including bile acids
and bacteria. Accordingly, the liver is selectively enriched in a large number of innate and
adaptive immune cells for protection against harmful pathogens and autoreactive self-antigens
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
7
from foodstuffs and commensal l bacteria. Lymphocyte subpopulations in normal human livers
are comprised of approximately 35% T cells, 30% natural killer (NK) T cells, 20% NK cells, and
10% B cells (Dong et al., 2007). TLRs are widely expressed by immune cells, including
conventional CD4+ T helper cells, cytotoxic T lymphocytes, B cells, natural forkhead box P3
(FoxP3)+ regulatory T cells, and NKT cells. This enables responsiveness to microbes through
pathogen-associated molecular patterns (PAMPs) or cell injury via danger-associated molecular
patterns (DAMPs) for cytokine production during LPS-induced cholestasis. Substantial evidence
has recently implicated that TLR4 signaling contributes to the activation of T cells, B cells, and
NKT cells (Kim et al., 2012; Pone et al., 2010; Reynolds et al., 2012). Importantly, while some
cytokines, e.g., TNF- and IL-1b are thought to be pro-inflammatory and detrimental, others
including IL-6 are also anti-inflammatory and suppress liver injury (Sun et al., 2004). The
objective of this study was to test the hypothesis that lymphocytes contribute to the expression
of hepatic transporter expression under basal and acute inflammatory conditions.
METHODS
Animal Models
Male adult C57BL/6J wild type (WT) and homozygous T cell-deficient B6.129S2-Tcratm1Mom
(TCRα KO), B cell-deficient B6.129S2-Ighmtm1Cgn (µMT), and T and B cell-deficient B6.129S7-
Rag1tm1Mom (Rag1 KO) mice (8-10 weeks of age) were purchased from The Jackson Laboratory
(Bar Harbor, ME). B10.A 5C.C7 T cell receptor (TCR) transgenic (Tg) Rag2 KO mice (Seder et al.,
1992) were obtained from the National Institute of Allergy and Infectious Diseases (NIAID)
contract facility (Taconic Farms, Germantown, NY). All mice were acclimated to the housing
conditions for at least 14 days prior to use. Groups for LPS-induced cholestasis studies were
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
8
designated WT (n = 6), TCRα KO (n = 6), µMT (n = 6), and Rag1 KO (n = 6). Groups for sex-
preference expression studies were designated C57BL/6J (n = 3) and B10.A TCR Rag2 KO (n = 3).
All mice were maintained in 12 h light/dark cycles and allowed water and standard chow ad
libitum. All animals were used according to the guidelines of the University of Missouri Animal
Care and Use Committee and in accordance with the U.S. National Institutes of Health and the
American Association for Laboratory Animal Care International.
LPS Model of Cholestasis
Four groups (WT, TCRα KO, µMT, and Rag1 KO) of adult (8-10 weeks old) age-matched male
mice (22–31 g) underwent treatment. LPS (4 mg/kg in a volume of 5 l/g) or sterile saline
vehicle was administered to mice by i.p. injection. Livers were excised 16 h after LPS
administration, snap-frozen in liquid nitrogen, and stored at -80oC until RNA isolation. Mice
were maintained under specific pathogen-free conditions in University of Missouri (MU) animal
facilities that are accredited by the Association for Assessment and Accreditation of Laboratory
Animal Care International. All protocols were approved by the MU Animal Care and Use
Committee. Mice were monitored for signs of distress during the study (abdominal distention,
respiratory difficulty).
Chemicals
LPS (from Escherichia coli serotype 055:B5) and all other chemicals, unless otherwise indicated,
were purchased from Sigma-Aldrich (St. Louis, MO). 1 mg LPS was added to 0.5 ml sterile saline
to create stock solutions and frozen (-20oC) prior to use.
RNA Preparations
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
9
Total RNA was isolated from mouse liver using RNAzol B reagent (Tel-Test Inc., Friendswood,
TX) per the manufacturer's recommendations. RNA concentrations were determined by UV
spectrophotometry, and the integrity of the RNA was confirmed by ethidium bromide staining
after agarose gel electrophoresis.
Branched DNA (bDNA) Assay
Specific oligonucleotide probes for Oatp 1a1, 1a4, 1b2, Oct1, Ntcp, Bsep, Mrp-2 and 3 genes
(Augustine et al., 2005) were diluted in lysis buffer supplied by the Quantigene HV Signal
Amplification Kit (Panomics, Fremont, CA). The substrate solution, lysis buffer, capture
hybridization buffer, amplifier, and label probe buffer used in the analysis were all obtained
from the Quantigene Discovery Kit (Panomics). The assay was performed in 96-well format with
10 μg of total RNA added to the capture hybridization buffer and 50 μl of the diluted probe set.
The total RNA was then allowed to hybridize to the probe set overnight at 53°C. Hybridization
steps were performed per the manufacturer's protocol on the following day. Luminescence of
the samples was measured with a Quantiplex 320 bDNA luminometer interfaced with
Quantiplex Data Management Software (version 5.02).
Real time RT-PCR
Real-time quantification of IL-1b, IFN-, TNF-, IL-6, IL-17A, and IL-17F relative to β-actin mRNA
was performed using SYBR Green PCR master mix (Applied Biosystems) and an ABI PRISM
7900HT sequence detector. Total RNA (5 μg) was reverse-transcribed to cDNA using
SuperScript™ III and random hexamer primers (Invitrogen) in a 20 μL reaction. PCR was
performed using the following primers: IL-1β, 5’-GAAAGCTCTCCACCTCAATG and 5’-
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
10
GCCGTCTTTCATTACACAGG, IFN- primers 5’-CTGCCACGGCACAGTCATTG and 5’-
TGCATCCTTTTTCGCCTTGC, TNF-α primers 5’-GACCCTCACACTCAGATCATCTTCT and 5’-
CCACTTGGTGGTTTGCTACGA, IL-6 primers 5’-TGTCTATACCACTTCACAAGTCGGAG and 5’-
GCACAACTCTTTTCTCATTTCCAC, IL-17A primers 5’-GCTCCAGAAGGCCCTCAGACT and 5’-
CCAGCTTTCCCTCCGCATTGA, IL-17F primers 5’-CTGGAGGATAACACTGTGAGAGT and 5’-
TGCTGAATGGCGACGGAGTTC, and β-actin primers 5’-ATGGTGGGAATGGGTCAGAA and 5’-
CCATGTCGTCCCAGTTGGTAA. PCR reactions incorporated 5 pmol forward and reverse primer
and used the following cycling conditions: 95°C for 10 min, 95°C for 15 s, and 60°C for 60 s
followed by dissociation curve analysis. For the negative controls, the reverse-transcriptase
step was omitted and PCR was performed directly from the RNA. The normalized threshold
cycle (Ct) values were subtracted from the target Ct values of each sample (ΔCt). Relative levels
of target mRNA were calculated as 2−ΔΔCt.
Statistical Analysis
Statistical analysis was performed using InStat 3 (GraphPad, La Jolla, CA) software. Data are
shown as mean + standard error (S.E.). The Mann-Whitney, Wilcoxon matched pairs test or
unpaired two-tailed t-test were applied to compare 2 groups; the one-way ANOVA with
Bonferroni’s multiple comparisons test was used to compare three or more means. Best fit of
correlation was measured by the root mean square. All p values <0.05 were considered
statistically significant. In the figures and tables, p values are displayed according to the
following scheme: **<0.01; *<0.05.
RESULTS
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
11
A genetic loss of function approach was used to determine the regulatory role of lymphocytes
on hepatic transporter steady-state mRNA expression prior to and after LPS treatment. Uptake
and efflux transporters investigated in this study were chosen on the basis that they encompass
sinusoidal and canalicular membrane localization and respond to inflammation. Given that
transporter and cytokine expression are sex-dependent and adult female responses are likely
influenced by estrous cycle (Klaassen and Aleksunes, 2010; Kwekel et al., 2010), male mice were
used. Hepatic transporter expression has also been shown to be age-dependent (Fu et al.,
2012); therefore, despite C57BL/6J Rag1 KO mice being smaller in size relative to WT, TCRα KO,
and µMT mice (p < 0.01), age-matched mice were studied (Fig. 1). Unfasted body weights (BW)
were recorded during the acclimatization period (days -14, -10, and -5), on the day of treatment
(day 1), and at necropsy (16 h post treatment). The administration of LPS resulted in a loss of
BW in WT, TCRα KO, and µMT, but not Rag1 KO, mice, amounting to 12.97 + 1.86 % (p < 0.05),
16.22 + 0.99 % (p < 0.01), 13.36 + 1.05 % (p < 0.01), and 8.44 + 1.18 % (p > 0.05) of saline
controls, respectively (Fig. 1). Overall, these results suggest that mice lacking T and B cells likely
respond differently to acute LPS exposure.
Consistent with previous studies (Klaassen and Aleksunes, 2010), basal steady-state mRNA
expression of sinusoidal Oatp 1a1, 1a4, 1b2, Ntcp, Oct1, and Mrp3 and canalicular Mrp2 and
Bsep transporters were readily detected (Table 1). Oatp1a4 expression was reduced in TCRα KO
mice relative to WT mice (p < 0.05). Although not significant, basal Oct1 expression was
reduced in Rag1 KO mice. In comparison, basal Mrp3, Mrp2, and Bsep expression was lower in
TCRα KO, µMT, and Rag1 KO mice compared to WT mice. Meanwhile, basal Oatp1a1, Oatb1b2,
and Ntcp expression were unaffected by lymphocyte deficiency. Of the transporter genes
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
12
studied, Oatp1a4 mRNA was uniquely down-regulated in the absence of T cells, but not B cells
or the combined deficiency of T and B cells (Table 1) to suggest that B cells, in a T cell-
independent manner, may augment Oatp1a4 expression. In summary, these data demonstrate
that T and B cells contribute to the maintenance of steady-state mRNA expression levels of
selective transporters in the liver of adult male C57BL/6J mice in the absence of inflammation,
infection, or autoimmunity.
We next investigated the effect of T cell and B cell deficiency on transporter mRNA expression
during inflammation. Fig. 2 demonstrates augmented Mrp2 mRNA and a loss of LPS-induced
suppression of Bsep mRNA in TCRα KO mice following acute LPS exposure. We further
investigated the potential for lymphocytes to impact hepatic sinusoidal transporter expression
following LPS treatment. Fig. 3 shows that LPS treatment did not alter Oatp1b2, Ntcp, or Mrp3
mRNA in any of the genotypes. In contrast, Oatp1a1 mRNA was reduced in response to LPS
treatment in all four genotypes. Meanwhile, LPS-induced inhibition of Oct1 and Oatp1a4 was
observed only in WT and B cell-deficient mice, respectively (Fig. 3). Overall, these data
demonstrate that lymphocytes contribute to differential regulation of hepatic transporter
expression in a transporter- and inflammation-dependent manner.
TNF-α, IL-1b, and IL-6 likely contribute to the regulation of hepatic drug transporters during
injury. Given that these cytokines are produced by multiple cell types, including lymphocytes,
we next asked whether T and B cells influence the expression of these soluble mediators in the
liver. For these studies, RT-PCR analyses were performed on liver homogenates from LPS or
saline treated WT, TCR KO, MT, and Rag1 KO mice. We demonstrate decreased basal IL-6
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
13
mRNA in TCR- KO and MT mice and exacerbated LPS-induced IL-6 expression in Rag1 KO
mice (Fig. 4). In contrast, alterations in IL-1b and TNF- mRNA were modest as shown by
tempered up-regulation during inflammation. Noteworthy, selective B cell loss lessened hepatic
TNF-, IL-1b, and IL-6 mRNA levels in LPS treated mice (Fig. 4). IL-6 mRNA levels were increased
in MT and Rag1 KO mice following LPS treatment. Meanwhile, IL-17A and IL-17F mRNA was
only slightly detectable (Ct = 38-42) and these levels did not differ with loss of T and/or B cells
(data not shown). Hepatic IFN-expression was below the level of detection in all samples
tested (data not shown). Overall, these results suggest that T and B cells contribute to basal and
acute inflammatory IL-6 production in the liver.
NKT cells represent a population of unconventional T lymphocytes that can mediate both
innate and adaptive immune responses and are abundant in the liver (Duwaerts and Gregory,
2011; Eberl et al., 1999; Parker and Picut, 2012). In the mouse, NKT cells represent
approximately 0.5% of the total T cell population in the blood, peripheral lymph nodes, and
spleens. In striking contrast, almost 30% of total hepatic T cells have the NKT phenotype (Eberl
et al., 1999) and the majority of these cells, like conventional T cells and NK cells, are localized
in the sinusoids (Geissmann et al., 2005). Therefore, we further investigated the contribution of
NKT cells on the regulation of sex-biased expression of hepatic bile acid transporters (Buist and
Klaassen, 2004; Cheng et al., 2007; Klaassen and Aleksunes, 2010; Maher et al., 2005). For these
experiments, male and female WT C57BL/6J and B10.A TCR Tg Rag2 KO mice were utilized to
determine the influence of combined genetic deficiency of CD8+, B, NKT, and natural CD4+ T
regulatory (Treg) cells on hepatic transporter expression. We first compared WT C57BL/6J and
CD8+, B, NKT, and natural Treg cells deficient (henceforth referred to as NKT-deficient) males
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
14
and females (Table 2). Although hepatic transporter mRNA expression was unaltered in NKT-
deficient males, increased Mrp3 mRNA (p < 0.01) and modestly decreased Oatp1a4 mRNA was
observed in NKT-deficient female mice relative to WT controls (Table 2).
We next determined whether NKT-deficiency influenced the gender predominance of
transporter mRNA expression. First, Oatp1a1 and Oatp1b2 mRNA was male-predominant in WT
mice (p < 0.01) and this sex bias was exacerbated in NKT-deficient mice. Second, Oatp1a4 (p <
0.05), Mrp3 (p > 0.05), Ntcp (p < 0.05), Oct1 (p > 0.05), and Bsep (p > 0.05) expression was
greater in WT females than males, and the magnitude of the female-predominance was either
unchanged or exacerbated with NKT deficiency. Mrp2 mRNA expression in WT mice was
comparable between males and females. Mrp3 mRNA expression was female-predominant in
WT as well as NKT-deficient mice (Table 2). In summary, the above analyses provide evidence
that lymphocytes may further function in a sex-bias manner to modulate hepatic transporters.
DISCUSSION
The possibility that lymphocytes may control hepatic transporter expression has not been
addressed until now. Using a genetic loss-of-function approach, we now provide evidence
suggesting that T and B cells contribute to the regulation of hepatic Mrp2, Bsep, Oct1, and
Oatp1a4 transporter expression. We further associate elevated IL-6 production during
conditions of T cell deficiency with abrogated suppression of Bsep and Oct 1 expression during
acute inflammation. In addition, we provide evidence that lymphocyte-mediated regulation of
hepatic transporter expression is sex-biased. Given that transporters can be significantly
affected by drug-drug interactions or genetic polymorphisms and the immune system has been
linked to sex bias human liver disorders, the contribution of lymphocytes to changes in drug
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
15
transporter activity warrants further attention. These findings contribute novel insights to the
cellular basis of cytokine production in the liver and to the mechanisms of inflammation-
induced liver diseases.
Previous studies demonstrating that altered hepatic transporter expression in response to LPS
have implicated that Kupffer cells directly respond to LPS through TLR4-dependent signaling to
up-regulate TNF-α, IL-6, and IL-1b transcription (Augustine et al., 2005; Cherrington et al., 2004;
Green et al., 1996; Kim et al., 2000; Kubitz et al., 1999; Li et al., 2004; Li and Klaassen, 2005). In
theory, these presumed pro-inflammatory cytokines, in turn, signal through their respective
receptors to repress transcription of genes encoding hepatic transporters causing bile acid
accumulation and cholestatic liver disease. We now demonstrate that T and B cells play
substantial, yet differential, roles in IL-6, but not IL-1b and TNF- production during basal and
acute inflammatory conditions. IL-6 has been implicated in both pro-inflammatory and anti-
inflammatory responses (Scheller et al., 2011) and most studies have investigated pro-
inflammatory contributions by IL-6. However, our data illustrating down- and up-regulated IL-6
in WT and T cell-deficient mice, respectively, are consistent with recent reports suggesting that
B cell-induced production of IL-6 prevents down-regulation of hepatic transporters to reduce
liver injury associated with acute inflammation (Klein et al., 2005; Taub, 2003). The molecular
mechanisms by which IL-6 protects in the liver are not well understood and this is further
confounded by a poor understanding of how T cells propagate hepatic inflammation. None the
less, it is well established that IL-6 binds the membrane-bound IL-6 receptor α (gp80) and
heterodimerizes with gp130 to form a signaling complex and activates downsteam STAT1,
STAT3, and SHP2-RAS-MAPK signaling pathways (Ishihara and Hirano, 2002). A lack of gp130
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
16
expression in hepatocytes has previously been shown to promote liver injury (Streetz et al.,
2003). Thus, it is unclear whether B cell induced IL-6 production directly targets T cells or
hepatocytes to modulate transporter expression. Given that IL-6-gp130-STAT3 in T cells
promotes IL-17 producing CD4 T cells (Camporeale and Poli, 2012), we were surprised by the
low levels of IL-17 expression in LPS treated Rag1 KO mice. However, these results suggest that
hepatocytes may be the critical recipient of the IL-6 signaling during acute TLR4-mediated
inflammation.
Although not well-appreciated, recent evidence has established that T, B, and NKT cells also
express TLR4 (Gururajan et al., 2007; Kim et al., 2012; Meyer-Bahlburg et al., 2009). Thus,
lymphocytes also retain the capacity to rapidly respond to endotoxin and other microbes
including commensal bacteria (Godfrey and Rossjohn, 2011; Reynolds et al., 2012; Tough et al.,
1997). Little is known about TLR4 signaling in lymphocytes, and it remains to be determined
whether TLR4 signaling pathways differ between lymphocytes and macrophages, particularly
with regards to the requirement for CD14, MyD88, and TRIF. Given the abundance of T, B, and
NKT cells in the liver and their capacity to propagate pro-inflammatory, but also anti-
inflammatory, cytokine production, we have utilized a genetic loss of function approach to test
the hypothesis that lymphocytes contribute to the regulation of genes encoding uptake and
efflux transporters in mouse liver. Our results are in agreement with previous studies
demonstrating down-regulation of sinusoidal and canalicular transporters in response to LPS as
previously reviewed (Klaassen and Aleksunes, 2010).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
17
Regarding the role of lymphocytes in regulating transporter expression, our findings do, in fact,
support our hypothesis that lymphocytes contribute to the transcriptional control of hepatic
bile acid transporters. Given that most lymphocytes including T and B, NK, and NKT cells are
thought to provide critical defense against invading pathogens and modulate liver injury by
circulating through hepatic sinusoids (Geissmann et al., 2005), it was somewhat unexpected
that the efflux transporters positioned on the apical membrane were most affected by
lymphocyte deficiency. None the less, it is well accepted that migration through hepatic
sinusoids enables lymphocytes to come in close contact to antigens displayed by endothelial
cells, Kupffer cells, and microbial pathogens undergoing enterohepatic circulation. Since altered
Mrp3 and Bsep mRNA expression in response to lymphocyte deficiency was evident only during
inflammation, we speculate that lymphocytes likely gain access, via direct contact or through
the secretion of soluble mediators, to hepatocytes at times when the vasculature in the liver is
inflamed as the sinusoidal endothelium is highly fenestrated and lacks a basement membrane.
In agreement, the sinusoidal membrane has previously been identified as a unique means by
which liver is able to differentially regulate immune-mediated responses (Crispe, 2012; Parker
and Picut, 2012). This line of reasoning is also consistent with our observation that lymphocytes
retain the capacity to modulate efflux transporters that are located on the apical membrane,
i.e., Mrp3, in the absence of LPS-induced inflammation.
Without doubt, hepatic transporter expression is critical for effective elimination of conjugated
bilirubin and bile acids as well as xenobiotics and their metabolites to maintain metabolic
homeostasis and simultaneous removal of toxic waste. However, it is also appreciated that
transient repression of hepatic efflux transporter function during acute inflammation, such as
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
18
sepsis or gallstones, offers protection to the liver by enhancing the retention of antioxidants
such as glutathione (Kong et al., 2012; Roeb et al., 2003). Thus, it is the sustained disruption of
these efflux pumps, in combination with uptake transporters, which exacerbates pathology in
liver diseases. It is appreciated that administration of LPS in experimental mouse models
reproduces the common clinical features of sepsis including down-regulation of transporter
mRNA. So, is altered hepatic transporter expression in response to lymphocyte deficiency
pathogenic or protective? This is a particularly important question given the liver provides a
unique environment that favors immunological tolerance. Of particular importance, CD4+
FoxP3+ Tregs are mediators of immune tolerance, express toll like receptors including TLR4,
exert enhanced suppressor function in response to LPS treatment, and have previously been
shown to contribute to extrahepatic cholestasis (Caramalho et al., 2003; Katz et al., 2011).
Additional studies to identify which lymphocytes are involved and gain a better understanding
of their mechanism(s) of action during acute and chronic exposure are required to
appropriately address this question. Here, we have clearly determined that hepatic
transporters are sensitive to modulation by lymphocyte activity. Importantly, while we and
others propose that LPS-induced cytokines directly target hepatocytes and ductal epithelial
cells to alter hepatic transporter mRNA, it has also been established that transporters, in
particular ABC transporters, are expressed on lymphocytes. Therefore, given that lymphocytes
also express TLR4, we cannot ignore the possibility that alteration of lymphocyte transporter
expression may contribute to inflammation in the liver, as has been previously suggested
(Giraud et al., 2010; van de Ven et al., 2009; Verbrugge et al., 2012).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
19
As hepatic bile acid transporter expression and immune response in other liver diseases are
both sex-dependent, a second aspect of this study involved elucidating whether lymphocytes
contribute to the sex-biased basal expression of hepatic transporters. Exacerbated basal Mrp3
expression in only female mice suggests that lymphocytes may contribute to sex-biased hepatic
transporter expression and implicate a particular important role for CD4+ T cells. Consistent
with this, conventional CD4+ T cells rapidly produce IL-2 upon activation. IL-2 receptor signaling
is predominantly mediated through STAT5a and STAT5b signaling pathways (Liao et al., 2013;
Lin et al., 2012), and hepatocytes respond to IL-2 signaling (Sunman et al., 2004). A role for
STAT5 signaling in gender-specific hepatic transporter expression has been identified for Ntcp
(Clodfelter et al., 2006; Clodfelter et al., 2007; Zhang et al., 2012). Thus, it is plausible that
cytokine signaling crosstalk with bile acid-induced nuclear receptor activation also contributes
to gender-specific regulation of hepatic transporters. Alternatively, gender discrepancy could
be the result of other divergent sex-dependent immune responses. For example, women have
higher absolute numbers of total CD4+ T cells and produce higher levels of IFN-.
In conclusion, the present study implicates that T and B cells regulate hepatic transporters (Fig.
5). In addition, we further implicate a protective role for IL-6 and identify lymphocytes as
modulators of sex-biased hepatic transporter expression. Given that targeted therapies that
affect lymphocyte function could adversely affect bile flow, it is critical that further work be
done to identify the lymphocytes that regulate transporter function.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
20
AUTHORSHIP CONTRIBUTIONS
Participated in Research design: Cherrington, McKarns
Conducted experiments: Tally, Bodeman, Dzierlenga, Lake, McKarns
Performed data analysis: Tally, Bodeman, Dzierlenga, Cherrington, McKarns
Wrote or contributed to the writing of the manuscript: Mulligan, Tally, Dzierlenga, Cherrington,
McKarns
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
21
REFERENCES
Augustine, L. M., Markelewicz, R. J., Jr., Boekelheide, K., and Cherrington, N. J. (2005).
Xenobiotic and endobiotic transporter mRNA expression in the blood-testis barrier. Drug Metab
Dispos 33, 182-189.
Buist, S. C., and Klaassen, C. D. (2004). Rat and mouse differences in gender-predominant
expression of organic anion transporter (Oat1-3; Slc22a6-8) mRNA levels. Drug Metab Dispos
32, 620-625.
Camporeale, A., and Poli, V. (2012). IL-6, IL-17 and STAT3: a holy trinity in auto-immunity? Front
Biosci 17, 2306-2326.
Caramalho, I., Lopes-Carvalho, T., Ostler, D., Zelenay, S., Haury, M., and Demengeot, J. (2003).
Regulatory T cells selectively express toll-like receptors and are activated by lipopolysaccharide.
J Exp Med 197, 403-411.
Cheng, X., Buckley, D., and Klaassen, C. D. (2007). Regulation of hepatic bile acid transporters
Ntcp and Bsep expression. Biochem Pharmacol 74, 1665-1676.
Cherrington, N. J., Slitt, A. L., Li, N., and Klaassen, C. D. (2004). Lipopolysaccharide-mediated
regulation of hepatic transporter mRNA levels in rats. Drug Metab Dispos 32, 734-741.
Clodfelter, K. H., Holloway, M. G., Hodor, P., Park, S. H., Ray, W. J., and Waxman, D. J. (2006).
Sex-dependent liver gene expression is extensive and largely dependent upon signal transducer
and activator of transcription 5b (STAT5b): STAT5b-dependent activation of male genes and
repression of female genes revealed by microarray analysis. Mol Endocrinol 20, 1333-1351.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
22
Clodfelter, K. H., Miles, G. D., Wauthier, V., Holloway, M. G., Zhang, X., Hodor, P., Ray, W. J., and
Waxman, D. J. (2007). Role of STAT5a in regulation of sex-specific gene expression in female but
not male mouse liver revealed by microarray analysis. Physiol Genomics 31, 63-74.
Crispe, I. N. (2012). Migration of lymphocytes into hepatic sinusoids. J Hepatol 57, 218-220.
Dong, Z., Wei, H., Sun, R., and Tian, Z. (2007). The roles of innate immune cells in liver injury and
regeneration. Cell Mol Immunol 4, 241-252.
Duwaerts, C. C., and Gregory, S. H. (2011). Targeting the diverse immunological functions
expressed by hepatic NKT cells. Expert Opin Ther Targets 15, 973-988.
Eberl, G., Lees, R., Smiley, S. T., Taniguchi, M., Grusby, M. J., and MacDonald, H. R. (1999).
Tissue-specific segregation of CD1d-dependent and CD1d-independent NK T cells. J Immunol
162, 6410-6419.
Fu, Z. D., Csanaky, I. L., and Klaassen, C. D. (2012). Effects of aging on mRNA profiles for drug-
metabolizing enzymes and transporters in livers of male and female mice. Drug Metab Dispos
40, 1216-1225.
Geier, A., Fickert, P., and Trauner, M. (2006). Mechanisms of disease: mechanisms and clinical
implications of cholestasis in sepsis. Nat Clin Pract Gastroenterol Hepatol 3, 574-585.
Geissmann, F., Cameron, T. O., Sidobre, S., Manlongat, N., Kronenberg, M., Briskin, M. J.,
Dustin, M. L., and Littman, D. R. (2005). Intravascular immune surveillance by CXCR6+ NKT cells
patrolling liver sinusoids. PLoS Biol 3, e113.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
23
Giraud, C., Manceau, S., Decleves, X., Goffinet, F., Morini, J. P., Chappuy, H., Batteux, F.,
Chouzenoux, S., Yousif, S., Scherrmann, J. M., et al. (2010). Influence of development, HIV
infection, and antiretroviral therapies on the gene expression profiles of ABC transporters in
human lymphocytes. J Clin Pharmacol 50, 226-230.
Godfrey, D. I., and Rossjohn, J. (2011). New ways to turn on NKT cells. J Exp Med 208, 1121-
1125.
Green, R. M., Beier, D., and Gollan, J. L. (1996). Regulation of hepatocyte bile salt transporters
by endotoxin and inflammatory cytokines in rodents. Gastroenterology 111, 193-198.
Gururajan, M., Jacob, J., and Pulendran, B. (2007). Toll-like receptor expression and
responsiveness of distinct murine splenic and mucosal B-cell subsets. PLoS One 2, e863.
Hirschfield, G. M., and Heathcote, E. J. (2009). Cholestasis and cholestatic syndromes. Curr Opin
Gastroenterol 25, 175-179.
Hirschfield, G. M., Heathcote, E. J., and Gershwin, M. E. (2010). Pathogenesis of cholestatic liver
disease and therapeutic approaches. Gastroenterology 139, 1481-1496.
Ishihara, K., and Hirano, T. (2002). Molecular basis of the cell specificity of cytokine action.
Biochim Biophys Acta 1592, 281-296.
Katz, S. C., Ryan, K., Ahmed, N., Plitas, G., Chaudhry, U. I., Kingham, T. P., Naheed, S., Nguyen,
C., Somasundar, P., Espat, N. J., et al. (2011). Obstructive jaundice expands intrahepatic
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
24
regulatory T cells, which impair liver T lymphocyte function but modulate liver cholestasis and
fibrosis. J Immunol 187, 1150-1156.
Kim, J. H., Kim, H. S., Kim, H. Y., Oh, S. J., and Chung, D. H. (2012). Direct engagement of TLR4 in
invariant NKT cells regulates immune diseases by differential IL-4 and IFN-gamma production in
mice. PLoS One 7, e45348.
Kim, P. K., Chen, J., Andrejko, K. M., and Deutschman, C. S. (2000). Intraabdominal sepsis down-
regulates transcription of sodium taurocholate cotransporter and multidrug resistance-
associated protein in rats. Shock 14, 176-181.
Klaassen, C. D., and Aleksunes, L. M. (2010). Xenobiotic, bile acid, and cholesterol transporters:
function and regulation. Pharmacol Rev 62, 1-96.
Klein, C., Wustefeld, T., Assmus, U., Roskams, T., Rose-John, S., Muller, M., Manns, M. P., Ernst,
M., and Trautwein, C. (2005). The IL-6-gp130-STAT3 pathway in hepatocytes triggers liver
protection in T cell-mediated liver injury. J Clin Invest 115, 860-869.
Kong, B., Csanaky, I. L., Aleksunes, L. M., Patni, M., Chen, Q., Ma, X., Jaeschke, H., Weir, S.,
Broward, M., Klaassen, C. D., et al. (2012). Gender-specific reduction of hepatic Mrp2
expression by high-fat diet protects female mice from ANIT toxicity. Toxicol Appl Pharmacol
261, 189-195.
Kubitz, R., Wettstein, M., Warskulat, U., and Haussinger, D. (1999). Regulation of the multidrug
resistance protein 2 in the rat liver by lipopolysaccharide and dexamethasone.
Gastroenterology 116, 401-410.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
25
Kwekel, J. C., Desai, V. G., Moland, C. L., Branham, W. S., and Fuscoe, J. C. (2010). Age and sex
dependent changes in liver gene expression during the life cycle of the rat. BMC Genomics 11,
675.
Li, N., Choudhuri, S., Cherrington, N. J., and Klaassen, C. D. (2004). Down-regulation of mouse
organic anion-transporting polypeptide 4 (Oatp4; Oatp1b2; Slc21a10) mRNA by
lipopolysaccharide through the toll-like receptor 4 (TLR4). Drug Metab Dispos 32, 1265-1271.
Li, N., and Klaassen, C. D. (2005). Lipopolysaccharide-induced down-regulation of organic anion
transporting polypeptide 4 (Oatp4; Slc21a10) is independent of tumor necrosis factor-alpha,
Interleukin-1beta, interleukin-6, or inducible nitric oxide synthase. Toxicol Sci 83, 197-203.
Liao, W., Lin, J. X., and Leonard, W. J. (2013). Interleukin-2 at the crossroads of effector
responses, tolerance, and immunotherapy. Immunity 38, 13-25.
Lickteig, A. J., Slitt, A. L., Arkan, M. C., Karin, M., and Cherrington, N. J. (2007). Differential
regulation of hepatic transporters in the absence of tumor necrosis factor-alpha, interleukin-
1beta, interleukin-6, and nuclear factor-kappaB in two models of cholestasis. Drug Metab
Dispos 35, 402-409.
Lin, J. X., Li, P., Liu, D., Jin, H. T., He, J., Ata Ur Rasheed, M., Rochman, Y., Wang, L., Cui, K., Liu,
C., et al. (2012). Critical Role of STAT5 transcription factor tetramerization for cytokine
responses and normal immune function. Immunity 36, 586-599.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
26
Maher, J. M., Slitt, A. L., Cherrington, N. J., Cheng, X., and Klaassen, C. D. (2005). Tissue
distribution and hepatic and renal ontogeny of the multidrug resistance-associated protein
(Mrp) family in mice. Drug Metab Dispos 33, 947-955.
Merritt, M. E., and Donaldson, J. R. (2009). Effect of bile salts on the DNA and membrane
integrity of enteric bacteria. J Med Microbiol 58, 1533-1541.
Meyer-Bahlburg, A., Bandaranayake, A. D., Andrews, S. F., and Rawlings, D. J. (2009). Reduced
c-myc expression levels limit follicular mature B cell cycling in response to TLR signals. J
Immunol 182, 4065-4075.
Mulder, J., Karpen, S. J., Tietge, U. J., and Kuipers, F. (2009). Nuclear receptors: mediators and
modifiers of inflammation-induced cholestasis. Front Biosci 14, 2599-2630.
Parker, G. A., and Picut, C. A. (2012). Immune functioning in non lymphoid organs: the liver.
Toxicol Pathol 40, 237-247.
Pone, E. J., Zan, H., Zhang, J., Al-Qahtani, A., Xu, Z., and Casali, P. (2010). Toll-like receptors and
B-cell receptors synergize to induce immunoglobulin class-switch DNA recombination:
relevance to microbial antibody responses. Crit Rev Immunol 30, 1-29.
Reynolds, J. M., Martinez, G. J., Chung, Y., and Dong, C. (2012). Toll-like receptor 4 signaling in T
cells promotes autoimmune inflammation. Proc Natl Acad Sci U S A 109, 13064-13069.
Ricciardi, R., Schaffer, B. K., Shah, S. A., Quarfordt, S. H., Banner, B. F., Wheeler, S. M., Donohue,
S. E., Meyers, W. C., and Chari, R. S. (2001). Bosentan, an endothelin antagonist, augments
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
27
hepatic graft function by reducing graft circulatory impairment following ischemia/reperfusion
injury. J Gastrointest Surg 5, 322-329.
Roeb, E., Purucker, E., Gartung, C., Geier, A., Jansen, B., Winograd, R., and Matern, S. (2003).
Effect of glutathione depletion and hydrophilic bile acids on hepatic acute phase reaction in rats
with extrahepatic cholestasis. Scand J Gastroenterol 38, 878-885.
Scheller, J., Chalaris, A., Schmidt-Arras, D., and Rose-John, S. (2011). The pro- and anti-
inflammatory properties of the cytokine interleukin-6. Biochim Biophys Acta 1813, 878-888.
Schenten, D., and Medzhitov, R. (2011). The control of adaptive immune responses by the
innate immune system. Adv Immunol 109, 87-124.
Seder, R. A., Paul, W. E., Davis, M. M., and Fazekas de St Groth, B. (1992). The presence of
interleukin 4 during in vitro priming determines the lymphokine-producing potential of CD4+ T
cells from T cell receptor transgenic mice. J Exp Med 176, 1091-1098.
Streetz, K. L., Wustefeld, T., Klein, C., Kallen, K. J., Tronche, F., Betz, U. A., Schutz, G., Manns, M.
P., Muller, W., and Trautwein, C. (2003). Lack of gp130 expression in hepatocytes promotes
liver injury. Gastroenterology 125, 532-543.
Sun, R., Tian, Z., Kulkarni, S., and Gao, B. (2004). IL-6 prevents T cell-mediated hepatitis via
inhibition of NKT cells in CD4+ T cell- and STAT3-dependent manners. J Immunol 172, 5648-
5655.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
28
Sunman, J. A., Hawke, R. L., LeCluyse, E. L., and Kashuba, A. D. (2004). Kupffer cell-mediated IL-2
suppression of CYP3A activity in human hepatocytes. Drug Metab Dispos 32, 359-363.
Taub, R. (2003). Hepatoprotection via the IL-6/Stat3 pathway. J Clin Invest 112, 978-980.
Tough, D. F., Sun, S., and Sprent, J. (1997). T cell stimulation in vivo by lipopolysaccharide (LPS).
J Exp Med 185, 2089-2094.
van de Ven, R., Oerlemans, R., van der Heijden, J. W., Scheffer, G. L., de Gruijl, T. D., Jansen, G.,
and Scheper, R. J. (2009). ABC drug transporters and immunity: novel therapeutic targets in
autoimmunity and cancer. J Leukoc Biol 86, 1075-1087.
Verbrugge, S. E., Assaraf, Y. G., Dijkmans, B. A., Scheffer, G. L., Al, M., den Uyl, D., Oerlemans,
R., Chan, E. T., Kirk, C. J., Peters, G. J., et al. (2012). Inactivating PSMB5 mutations and P-
glycoprotein (multidrug resistance-associated protein/ATP-binding cassette B1) mediate
resistance to proteasome inhibitors: ex vivo efficacy of (immuno)proteasome inhibitors in
mononuclear blood cells from patients with rheumatoid arthritis. J Pharmacol Exp Ther 341,
174-182.
Zhang, Y., Laz, E. V., and Waxman, D. J. (2012). Dynamic, sex-differential STAT5 and BCL6
binding to sex-biased, growth hormone-regulated genes in adult mouse liver. Mol Cell Biol 32,
880-896.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
29
FOOTNOTES
This work was supported in part by the National Institutes of Health National Institute of Allergy
and Infectious Diseases Extramural Activities [Grant AI083927]; the National Institutes of Health
Eunice Kennedy Shriver National Institute of Child Health and Human Development [Grant
HD062489] (to N.J.C.) and the National Institute of Environmental Health Science Toxicology
Training [Grant, ES007091] to A. D. L.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
30
LEGENDS FOR FIGURES
Figure 1. The influence of LPS treatment of body weight. The body weights of 16-week old
male WT, TCRα KO, µMT, and Rag1 KO mice (n = 3 mice per experimental group) were obtained
one day prior to and 16 h following LPS administration (4 mg/kg in a volume of 5 l/g; i.p.) or an
equal volume of sterile saline (vehicle control). The BW (g) for individual mice before and after
saline (top) or LPS (bottom) treatment is shown. Open and closed circles represent prior to and
after treatment **<0.01; *<0.05.
Figure 2. Steady-state mRNA levels of hepatic canalicular efflux transporters in response to
LPS. Male WT, TCRα KO, µMT, or Rag1 KO mice (22–31 g) were administered LPS (4 mg/kg in a
volume of 5 µl/g) or sterile saline vehicle by injection (i.p.). Livers were excised 16 h after LPS
administration, collected, snap-frozen in liquid nitrogen, and stored at -80oC until RNA isolation
(n = 3 mice per experimental group). Livers were evaluated for steady-state mRNA levels of
Bsep and Mrp2 as determined by the bDNA assays. Data are expressed as mean relative light
units (RLU)/ 10 g total RNA + S.E **<0.01; *<0.05.
Figure 3. Steady-state mRNA levels of hepatic sinusoidal transporters in response to LPS. Male
WT, TCRα KO, µMT, or Rag1 KO mice (22–31 g) were administered LPS (4 mg/kg in a volume of
5 µl/g) or sterile saline vehicle by injection (i.p.). Livers were excised 16 h after LPS
administration, collected, snap-frozen in liquid nitrogen, and stored at -80oC until RNA isolation
(n = 3 mice per experimental group). Livers were evaluated for steady-state mRNA levels of the
hepatic bile acid transporters Oatp1a1, Oatp1a4, Oatp1b2, Ntcp, Oct1, and Mrp3 as determined
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
31
by branched DNA analyses. Data are expressed as mean RLU/ 10 g total RNA + S.E **<0.01;
*<0.05.
Figure 4. The influence of T and B cell deficiency on basal and LPS-induced hepatic cytokine
gene expression. Male WT, TCRα KO, µMT, or Rag1 KO mice were administered LPS (4 mg/kg in
a volume of 5 µl/g) or sterile saline vehicle by injection (i.p.). Livers were excised 16 h after LPS
administration, collected, snap-frozen in liquid nitrogen, and stored at -80oC until RNA isolation.
Gene expression was quantified by RT- PCR using SYBR green PCR master mix and the ABI Prism
7900HT sequence detection system. Fold change was calculated as 2−ΔΔCt using -actin as
endogenous control. The results shown are normalized to WT vehicle (saline) controls.
Quantification was calculated as vehicle versus LPS-treated cells. Data are mean ± S.E. of three
mice.
Figure 5. Schematic depicting differential regulation of hepatic transporter expression by T
and B cells. Bsep and Mrp2 transporters are expressed on hepatocyte canalicular membranes
and export monovalent tauro- and glyco-conjugated (conj.) hepatic bile acids (BAs) and divalent
sulphated (sulpho-) or glucuronidated (glucurono-) BAs amidated with a taurine or a glycine,
organic anions (OAs), reduced glutathione (GSH), and bilirubin (bili) out of the hepatocyte into
the bile canaliculi. Mrp3, like Mrp2, is an ATP-binding cassette transporter, but unlike Mrp2, is
localized on hepatocyte basolateral surfaces and functions to efflux bile and organic acids
during periods of BA overload, e.g., gallstones or dysfunctional Mrp2 and/or Bsep. Ntcp,
Oatp1a1, Oatp1a4, and Oatp1b2 are positioned at the basolateral membrane and facilitate
enterohepatic cycling of BAs and OAs. Oct1 is also expressed at the basolateral membrane and
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

JPET #205369
32
functions in a similar manner, except it transports conj. BAs and organic cations (OCs) from the
portal vein.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

Table 1. Basal Transporter Gene Expression in Liver Isolated from male WT mice and mice lacking T cells (TCRα KO), B cells (µMT), or both T and B cells (Rag1 KO)a.
Transporter Human Gene Accession
#
Nuclear
Receptorb Ligandc WT TCR KO MT Rag1 KO
Oatp1a1 (Oatp1)
OATP1A2 Slco1a1 NM_013797 FXR, RXR, SHP, HNF4α, GR
BA (↑) 69.56 + 7.78 74.09 + 13.39 63.43 + 8.85 64.26 + 4.16
Oatp1a4 (Oatp2)
OATP1B1 Slco1a4 NM_030687 FXR, SHP, HNF4α
SHP (↓) 5.81 + 0.88 0.90 + 0.41* 6.03 + 0.61 5.03 + 0.92
Oatp1b2 (Oatp4)
OATP2B2 (OATP1B3)
Sclo1b2 NM_020495 FXR BA (↑) 82.27 + 17.96 88.53 + 19.38 63.80 + 3.33 85.07 + 7.10
Ntcp (lx1)
NTCP Slc10a1 NM_011387 FXR, RXR, SHP, HNFα, GR,
RAR
Retinoids (↑)
SHP (↓)
64.04 + 11.80 57.95 + 2.83 76.06 + 8.80 80.73 + 18.80
Oct1 OCT1 Slc22a1 NM_009202 PXR, PPAR, HNF1α
OC (↑) 192.93 + 29.88 124.42 + 20.75 151.48 + 30.08 104.84 + 3.80
Mrp3 MRP3 Abcc3 NM_029600 CAR, PXR, VDR BS, BIli, PB, phytoestrogens (↑)
15.21 + 3.58 11.24 + 4.80 11.89 + 3.13 5.56 + 1.00
Mrp2 MRP2, cMoat
Abcc2 NM_013806 PXR, CAR, FXR BA, Bili, GSH, Xenobiotics(↑)
29.87 + 8.59 20.40 + 1.91 13.13 + 3.27 14.55 + 3.65
Bsep BSEP, SPGP, PFIC-2
Abcb11 NM_021022 FXR, RXR BA (↑) 50.48 + 8.52 21.81 + 4.35 31.27 + 0.38 38.67 + 15.93
aResults represent the mean RLU/10 g total RNA + SE (n = 3). *p < 0.05 relative to WT.
bReviewed in (Beuers et al, 2001; Chandra & Brouwer, 2004; Halilbasic et al, 2013; Hofmann, 2007; Hofmann, 2009; Shu et al, 2008; Tirona & Kim, 2005; Trauner & Boyer, 2003).
cReviewed in (Beuers et al, 2001; Chandra & Brouwer, 2004; Halilbasic et al, 2013; Shu et al, 2008).
BA, bile acid; Bili, bilirubin; BS, bile salts; BSEP, bile salt export pump; CAR, constitutive androstane receptor; cMOAT, canalicular multi-organic anion transporter; FXR, farnesoid X receptor; GSH, glutathione; GR, glucocorticoid receptor; HNFα, hepatocyte nuclear factor alpha; MRP, multidrug resistance associated protein; NTCP, sodium-taurocholate cotransporting polypeptide; OATP, organic anion transporter; OC, organic cation; OCT,
organic cation transporter; PB, phenobarbital; PFIC-2, progressive familial intrahepatic cholestasis; PPAR, peroxisome proliferator-activated
receptor alpha; PXR, pregnane X receptor; RAR retinoid acid receptor alphaRXR, retinoid X receptor; SHP, short heterodimer partner; SPGP, sister of P-glycoprotein; VDR, vitamin D receptor; ↑, increases transporter expression; ↓, decreases transporter expression.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

Table 2. Sex-dependent differences in constitutive hepatic transporter gene expression in WT mice and mice genetically-deficient in NKT, CD8+, B, and natural FoxP3+ regulatory T cellsa
.
Transporter Males Females
WT TCRα Tg Rag2 KO WT TCRα Tg Rag2 KO
Oatp1a1 89.07 + 26.73 100.32 + 17.10 37.87 + 22.75 4.41 + 1.93
Oatp1a4 6.80 + 1.30 5.09 + 1.11 44.53 + 5.91 27.09 + 3.15
Oatp1b2 91.75 + 8.18 93.07 + 3.75 61.29 + 14.74 50.60 + 20.58
Ntcp 64.49 + 12.90 97.89 + 10.96 146.58 + 21.35 163.08 + 4.55
Oct1 128.78 + 26.81 154.74 + 14.71 182.76 + 56.49 254.43 + 49.54
Mrp3 6.31 + 0.53 7.22 + 1.98 9.30 + 0.97 17.46 + 3.07**
Mrp2 28.95 + 3.17 25.71 + 4.91 24.38 + 13.87 41.69 + 5.16
Bsep 74.97 + 12.99 60.70 + 1.32 128.67 + 56.47 102.47 + 9.06
aResults represent the mean RLU/10 g total RNA + SE (n = 3). **, p < 0.01, comparing gender-
specific WT and TCR Tg Rag2 KO mice.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

WT TCR KO MT Rag1 KO
15
25
35
0 h Saline
16 h Saline
WT TCR KO MT Rag1 KO
15
25
35
0 h LPS
16 h LPS
*** **B
ody W
eig
ht (g
)
McKarns_JPET #205369_Figure 1
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

WT TCR KO MT Rag1 KO0
30
60
90 Mrp2
*
Saline
LPS
WT TCR KO MT Rag1 KO0
30
60
90 Bsep
*
**
mR
NA
(R
LU
/ 10 µ
g tota
l R
NA
)
McKarns_JPET #205369_Figure 2
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

0
40
80
120 Oatp1a1
** ***
*
WT TCR KO MT Rag1 KO0
5
10
15Oatp1a4
**
0
75
150
225Oatp1b2
LPS
Saline
WT TCR KO MT Rag1 KO
50
100
150Ntcp
0
100
200
300 Oct1*
WT TCR KO MT Rag1 KO0
8
16
24Mrp3
mR
NA
(R
LU
/ 10 µ
g tota
l R
NA
)
McKarns_JPET #205369_Figure 3
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from

Rela
tive m
RN
A e
xpre
ssio
n 0
2
4
6
8IL-1b
*
Saline
LPS
0.0
0.5
1.0
1.5
2.08
1012
IL-6
*
**
WT TCR KO MT Rag1 KO0
10
20
30TNF-
**
**
McKarns_JPET #205369_Figure 4
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from
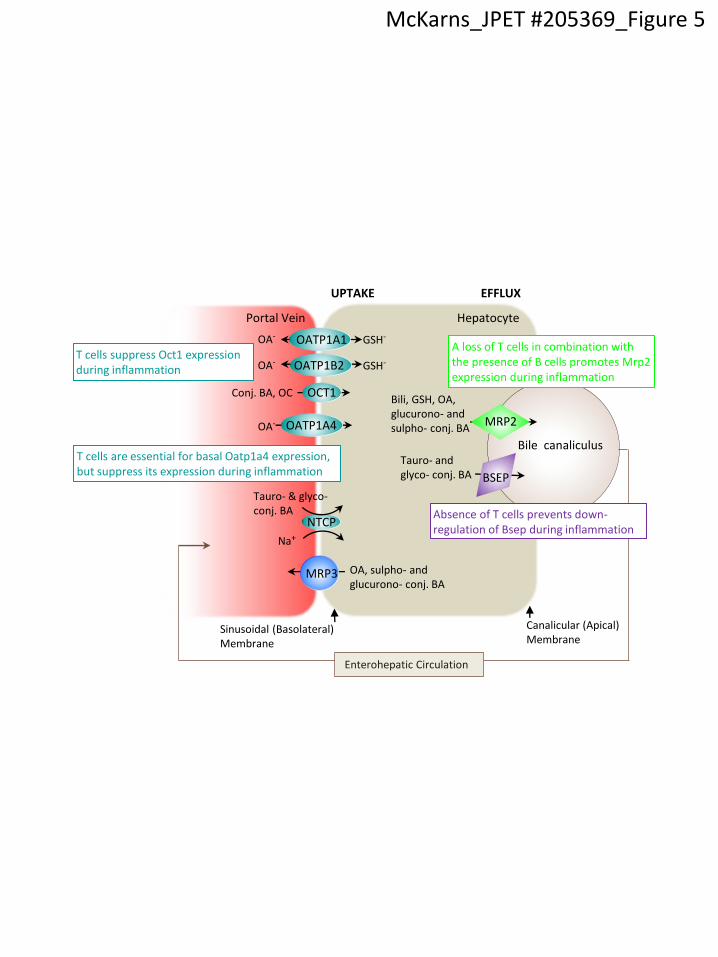
Bile canaliculus
Hepatocyte Portal Vein
GSH-
Tauro- & glyco-conj. BA
OA, sulpho- and glucurono- conj. BA
Sinusoidal (Basolateral) Membrane
Canalicular (Apical) Membrane
BSEP
MRP3
Na+
OATP1A1
OCT1
OATP1B2
NTCP
UPTAKE EFFLUX
MRP2
Conj. BA, OC
Enterohepatic Circulation
GSH-
OA-
OA-
OATP1A4 OA-
Bili, GSH, OA, glucurono- and sulpho- conj. BA
Tauro- and glyco- conj. BA
A loss of T cells in combination with the presence of B cells promotes Mrp2 expression during inflammation
Absence of T cells prevents down-regulation of Bsep during inflammation
T cells suppress Oct1 expression during inflammation
T cells are essential for basal Oatp1a4 expression, but suppress its expression during inflammation
McKarns_JPET #205369_Figure 5
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on August 8, 2013 as DOI: 10.1124/jpet.113.205369
at ASPE
T Journals on June 26, 2018
jpet.aspetjournals.orgD
ownloaded from